 Jianhui Hao1,2†
Jianhui Hao1,2† Hongkun Han
Hongkun Han Jiahuan Li
Jiahuan Li Jiyun Yang
Jiyun Yang Baihui Ren
Baihui Ren Long Bai
Long Bai- 1College of Horticulture, Shenyang Agricultural University, Shenyang, China
- 2College of Agronomy, Inner Mongolia Minzu University, Tongliao, China
Atmospheric nitrogen(N) deposition increased soil active N, and excessive N content led to the increase of the ratio of nitrogen to phosphorus (N: P), which changed plant growth from N limitation to phosphorus(P) limitation. Potentilla is not only an important native greening material, but also a common diversity component in various grasslands in China. Its population fluctuation in the process of N deposition will affect the species diversity and productivity of grassland ecosystem. Potting experiment was conducted for 2 years with Potentilla tanacetifolia, a common species in the northern warm steppe, as the material. Through the interactive treatment of different N addition (0, 10, 20, 40 kg N ha yr−1) and P addition (4, 6, 8 kg P ha yr−1) gradients, to analysis the feedback effect between leaf N and P content and net photosynthetic rate (Pn). We explored the N: P threshold of N and P limitation from the perspective of Pn. The results showed that: 1) Under low soil N concentration, P addition can promote N absorption of P.tanacetifolia, while the high soil N concentration can reduce the N: P by increasing the leaf P content to weaken the limiting effect caused by nutrient imbalance of plants. 2) In N addition environment, proper P addition increased Pn by increasing stomatal conductance (Gs), while excessive P addition decreased Gs and inhibited Pn. 3) The Pn showed a single peak normal distribution characteristic with the enhancement of the N: P of leaves, and the Pn was at a high level between 14.5–17.0. It was preliminarily believed that the threshold value of N: P in leaves of P.tanacetifolia was 14.5–17.0. Plant photosynthesis is very sensitive to the environment and easy to be affected by the external environment. The results of N and P addition showed that Pn of broad-leaf forbs was easily affected by N and P restriction, and P addition increased Pn of broad-leaf forbs under N restriction. There was a certain relationship between N:P and Pn. It was preliminarily believed that the N: P of P.tanacetifolia leaves is not limited by nitrogen and phosphorus in the range of 14.5-17.0.
Introduction
Atmospheric nitrogen(N) deposition has become the third driving factor for biodiversity loss after land use and global climate change. Since 2005, the Biodiversity Committee of the UNEP has listed N deposition as an important indicator for assessing biodiversity change (Sala et al., 2000; Payne et al., 2017). N deposition increases the content of soil ionic N (NO3-, NH4+), the inherent nutrient balance between soil and plants is changed, which has a serious impact on the growth and reproduction of plants, and ultimately leads to the loss of species diversity (Liu et al., 2013; Stevens et al., 2018). In recent years, the N deposition rate in northern China has reached 40–70 kg ha yr−1 and has been growing continuously, which has become one of the important environmental variables in the warm steppe of northern China (Yin et al., 2017; Tian et al., 2020). Due to the responses of different plant to soil N addition, photosynthetic capacity, growth and reproduction of plants showed different fluctuation trends under a certain range of N addition (Soons et al., 2017; Ibáñez et al., 2018; Stevens et al., 2018). Therefore, in order to further clarify the impact of N deposition on the warm steppe communities in northern China, further studies are still needed on different vegetation combinations in different regions.
According to 1,091 sets of plant observation data from 181 research sites worldwide, N addition could significantly increase the N content of plant leaves and lead to further increase in the ratio of nitrogen to phosphorus (N:P) (You et al., 2018). In N limited ecosystems, moderate N deposition increased the N content of plants, and some plants had stronger growth and reproduction ability due to the release of N limitation (Yu et al., 2022). However, when the increase of N deposition exceeded a certain threshold, due to the phosphorus(P) content in the soil was not supplemented, the high N:P led to the transformation of the ecosystem from N limitation to P limitation. Again inhibiting the growth of the plant (Bai et al., 2010; Yuan et al., 2021). Study on the tundra zone of Changbai Mountain in Northeast China found that N addition increased the leaf N content of the dominant plant Rhododendron aureum, decreased the plasticity of N and P, and decreased the plant viability, while another dominant plant Vaccinium uliginosum showed a batter growth trend (Yuan et al., 2021). Therefore, N deposition will inevitably lead to the replacement of dominant species or species decline. In order to determine the limit range of plant N:P, we need an index as a basis for judgment.
Photosynthesis plays a key role in plant energy acquisition and transformation, which will directly affect the growth and reproductive capacity of plants. The loss of plant diversity in warm steppe due to N enrichment mainly results from changes in plant functional groups, through the rapid accumulation of NH4+-N in the soil after N addition, which first inhibits the net photosynthetic rate of broad-leaved non-root nodule plants, leading to the loss of these N sensitive species in the early stages (≤3 years) of N enrichment, which has become an important mechanism for the reduction of plant diversity due to N deposition in warm alkaline calcareous steppe (Tian et al., 2020). Therefore, changes in net photosynthetic rate is a stronger indicator of the early stages of N addition. Studies in the eastern Tibetan Plateau showed that Pn of Potentilla fruticosa was suppressed when N additions exceeded 15 g N m−1 yr−1 (Guo et al., 2014). N additions significantly increased the Pn of the dominant species Kobresia capillifolia and Elymus nutans in cold grassland, but Pn of Potentilla cranbergii was still inhibited (Xu et al., 2018). Pn of Leymus chinensis was suppressed in temperate grasslands when N additions exceeded 50 kg N · ha−1 · yr−1 (Wang et al., 2019a), while Pn of the dominant species Cryptocarya concinna was suppressed at N additions above 100 kg N · ha−1 · yr−1 in subtropical forests (Mao et al., 2018). These studies show that the relationship between Pn response and N addition of grass plants in different ecological regions has not been uniformly concluded. With most grassland plants changing from N limitation to P limitation, the net photosynthetic rate decreased with the increase of N deposition. N:P in soil of alpine meadow, warm steppe and mountain steppe was positively correlated with N:P in leaves of their grassland vegetation (Stipa baicalensis, Leymus chinensis, etc.) (Prach et al., 2021; Qian et al., 2018; Wang et al., 2019a). This indicated that the net photosynthetic rate was inhibited when N:P exceeded a certain threshold. Therefore, we supposed that whether N:P could be reduced by P addition to alleviate the inhibition of photosynthesis caused by plant nutrient limitation and improve the growth and reproduction ability of plants in N deposition environment.
Studies have shown that P addition can alleviate the P limitation of plants when N is excessive (Liu et al., 2018; Zong et al., 2022). However, many studies have focus on grasses and legumes, and the current judgment of N and P limitation threshold is usually defined by the biomass change of N and P addition experiments (Du et al., 2014; Turner and Wright, 2014). Potentilla tanacetifolia is widely distributed in temperate grasslands in northern China. It has the characteristics of early turning green, strong drought resistance, high ornamental value, and natural material for landscaping (Hao et al., 2020). We propose the following hypotheses. Can changes in net photosynthetic rate reveal the trend of N and P limitation of broad-leaf forbs by N deposition? Does P addition in the context of N deposition improve net photosynthetic rate of broad-leaf forbs? Is there any relationship between N:P and net photosynthetic rate? Therefore, we selected P.tanacetifolia, a common broad-leaved forbs in warm steppe of northern China, as the material. Used pot experiment and the method of adding N and P artificially, mastered the response of plant N and P content and photosynthesis after the addition of N and P. Revealed the relationship between leaf N:P and net photosynthetic rate, and explored the N:P threshold of N and P limitation. This study provides a theoretical basis for predicting the potential impact of N deposition on grassland plants in northern China and coping strategies in the future.
Materials and methods
Study area and experimental design
The seed of P.tanacetifolia were collected from the Aerxiang Teaching and Research Base of Shenyang Agricultural University (42°08′-42°;50′N, 121°;53′-122°;58′E), which is located in the transition zone from the eastern forest to the warm steppe, with an annual precipitation of 550 mm (Figure 1). It is hot in summer and cold in winter, belonging to the temperate continental climate. In September 2018, the collected seeds were planted in experimental pots (with an inner diameter of 21 cm, a height of 27 cm, and a hole with a diameter of 1 cm at the bottom), and then placed in a greenhouse to seedlings cultivation. In April 2019, one healthy seedling was retained in each pot and moved to the awning for test. The pot was filled with 8 kg brown soil (pH 5.74, organic matter 32.11 g·kg−1, total N 1.02 g·kg−1, and total P .42 g·kg−1). The experimental period is from June 2019 to September 2020, and samples will be taken after 2 years of treatment.
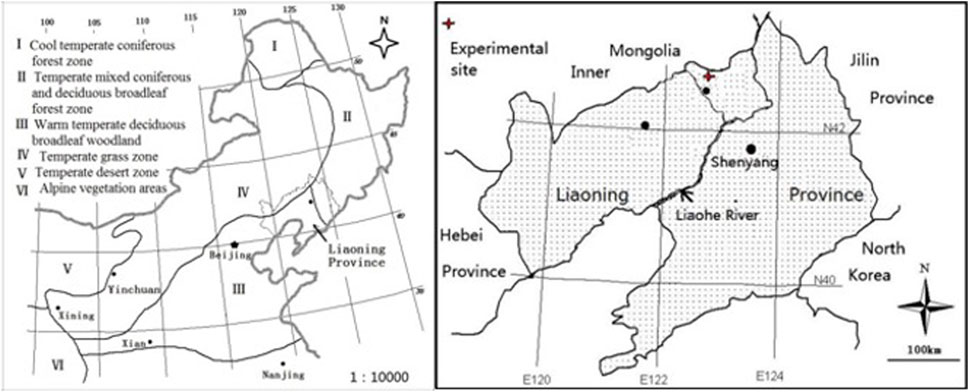
FIGURE 1. Basic information of test site, the red cross in the figure is the location of the test site.
The N treatment agent was urea (CH4N2O) with a N content of 46%, and KH2PO4 was used as a P additive. The amount of N and P added in 1 year were divided into two equal parts, dissolved in water and sprayed on June 10 and August 10 each year. According to the measured data, the total atmospheric dry deposition of N was 24.1 kg N ha yr−1 in 2017 in the grassland near the Dahekou Reservoir (42°13′19.17″N, 116°38′4.00″E) in Inner Mongolia, 200 km from the experimental area (Lu et al., 2021). 4 gradients of N concentration are set. Marked as N0 (N free treatment, 0 kg N ha yr−1), N10 (low N treatment, 10 kg N ha yr−1), N20 (medium N treatment, 20 kg N ha yr−1) and N40 (high N treatment, 40 kg N ha yr−1). According to the data of Hulun Buir Meadow Steep Field Experiment Station of the Chinese Academy of Sciences (Gao et al., 2017), 3 gradients of P concentration were set, which were marked as P4 (low P treatment, 4 kg P ha yr−1), P6 (medium P treatment, 6 kg P ha yr−1) and P8 (high P treatment, 8 kg P ha yr−1), and there were 12 in total after two-factor interaction treatment. Each treatment was repeated 5 per, with a total of 60 pots. To avoid the influence of precipitation, it is calculated according to the annual average precipitation of the test site that artificial water replenishment is carried out every 4 days during the test period, which is equivalent to an increase of 1% in annual precipitation.
Data collection
Photosynthesis was measured from 9:00 a.m. to 12:00 a.m. on 2 September 2020. Before the measurement, the leaves were wrapped in tinfoil to adapt to the darkness at ambient temperature for 1 h, and then the net photosynthetic rate (Pn), stomatal conductance (Gs) and other light and parameters of the leaves of different plants were measured by portable photosynthesis system (Li6800, Li-Cor, United States). The CO2 cylinder was used for the measurement, and the CO2 concentration was set to (400 ± 5) mmol mol−1. The red and blue light source fluorescence chamber (LI6800-02) was used with 90% red light and 10% blue light as light sources, and the light intensity was set to 1,350 μmol m−2 s−1 (Liu et al., 2019). Three leaves in the same position were selected for each treatment, and each leaf was collected five times, and the plant photosynthesis was recorded as the average value of 15 data.
The plant leaves were taken immediately after the determination of photosynthetic parameters, washed with distilled water and dried, and then put into an oven for deactivation of enzymes (105°C, 30 min) and dried at 65°C to constant weight. The dried plant samples were ground into powder for determination, and the total N and P contents of the leaves were determined by using an intermittent chemical analyzer (Smartchem140).
Statistical analysis
We used SPSS (Version 23.0, SPSS Inc., United States) to analyze the collected data, and two-way ANOVA was used to analyze the significance of the two factors of N and P addition, and then LSD multiple comparisons were performed. The significance analysis was performed at the 95% confidence level for the P addition treatments. The analyzed data were plotted using Sigmaplot (Version 12.5, Systat Software Inc., United States). The variation degree of plant N and P nutrients is calculated according to the ratio of the standard deviation of the plant N and P nutrient contents to the average value of the N and P nutrient contents in the same concentration N treatment group and the same concentration P treatment group, i.e.
Results
Effects of N and P addition on N and P absorption in leaves
The leaf total N content of P.tanacetifolia was significantly different only in N0 group. Compared with N0P4, the leaf total N content significantly increased (p < .01) by 17.5% (N0P6) and 52.1% (N0P8), and the leaf total N content increased with P addition (Figure 2A). Leaf P content was different from N content. In N0, N10 and N40 groups, P8 treatment significantly increased leaf total P content (p < .05), while in N20 group, P6 treatment significantly increased (p < .05) (Figure 2B).
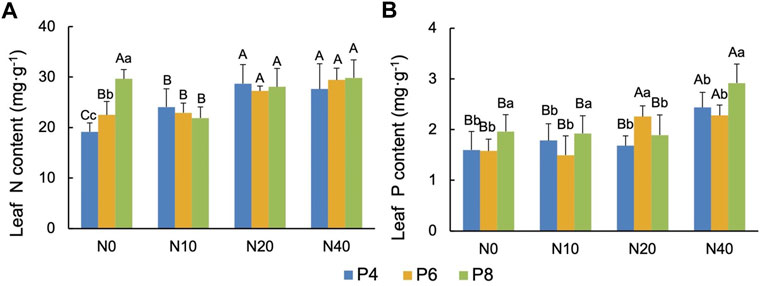
FIGURE 2. Effects of N and P addition on (A) total N content and (B) total P content in leaves of P.tanacetifolia. Different capital letters indicate significant differences (p < .05) between groups of N addition treatments in the same P treatment group, and different small letters indicate significant differences (p < .05) between different P addition treatments in the same N treatment group, similarly for the following figure.
Under N addition treatment, there was no significant difference in the variation coefficient of P.tanacetifolia N nutrient, which was maintained at a low level, while the variation coefficient of P nutrient was significantly increased in the N0 and N10 groups (p < .05) (Figure 3A). Under P addition, the variation coefficient of N increased significantly within P4 group (p < .05), while there was no significant change in the coefficient of variation of P nutrient (Figure 3B). Plant P nutrients were more sensitive under low N (N0, N10), and plant N was also more sensitive under low P conditions (P4).
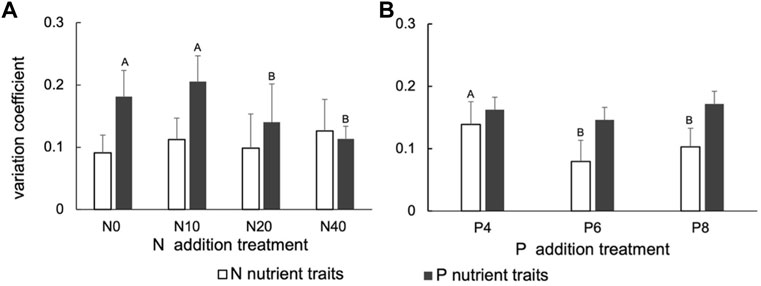
FIGURE 3. Coefficient of variation of N and P contents in P.tanacetifolia under (A) N addition treatment and (B) P addition treatment. Different capital letters indicate significant differences (p < .05) between groups of CV of N and CV of P.
Effects of N and P addition on leaf photosynthetic parameters
In the N0 and N10 treatment groups, only the Pn of P4 was significantly increased (p < .01), and in the N20 treatment group, the Pn of P4 and P6 was significantly increased (p < .01), while in the N40 treatment group, only the net photosynthesis rate of P6 was significantly increased (Figure 4A). The change of Gs was basically consistent with the change of Pn. The Gs of P4 in N0, N10 and N20 treatments was significantly increased (p < .01), while the Gs of P6 in N40 treatment was significantly increased (p < .01) (Figure 4B).
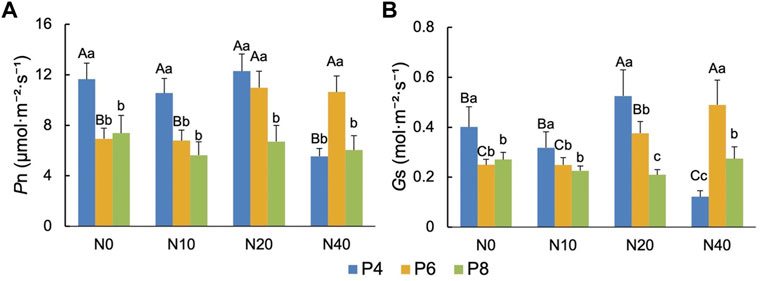
FIGURE 4. Effects of N and P addition on (A) net photosynthetic rate (Pn) and (B) stomatal conductance (Gs) of P.tanacetifolia.
In order to demonstrate the effects of N and P addition on Pn and Gs of P.tanacetifolia, we took the highest Pn in N0 group as the boundary for comparison. In the N0 and N10 treatments, only part of the Pn of P4 treatment could exceed this limit, and the Gs of N10P4 treatment was lower when it exceeded this limit (Figures 5A, B). In the N20 treatment group, the Pn of most P4 treatments were higher than this limit, and the Pn of most P6 treatments also fluctuated at this limit, and the overall increase in Pn was promoted by higher Gs (Figure 5C). However, the N40 treatment was different from other N addition groups. The photosynthesis of P4 treatment was inhibited, while P6 treatment could still maintain the Pn above and below the limit through higher Gs (Figure 5D). The P8 treatment exhibited lower photosynthetic parameters at all N addition levels and did not show significant changes (Figure 5).
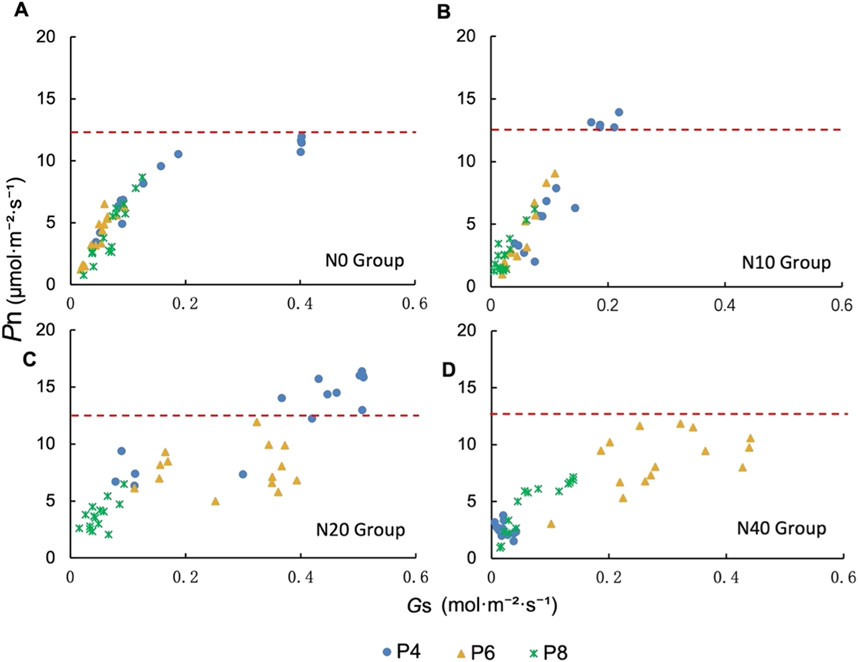
FIGURE 5. Relationship between net photosynthetic rate (Pn) and stomatal conductance (Gs) under different N and P addition treatments. (A) N0 group, (B) N10 group, (C) N20 group and (D) N40 group.
Correlation between photosynthetic parameters and N and P in leaves
Leaf N and P status is one of the important factors affecting Pn. We found that with the increase of leaf N content, the Pn of plants decreased, and there was a negative correlation between leaf N content and Pn (Figure 6A). There was also a negative correlation between leaf P content and Pn, and the Pn of the plant decreased with the increase of leaf P content (Figure 6B).
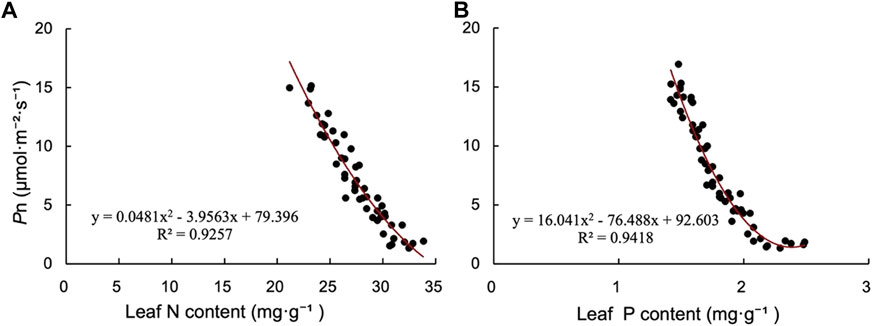
FIGURE 6. Relationship between photosynthetic rate and (A) total N and (B) total P in P.tanacetifolia leaves.
We analyzed the leaf N:P and Pn under different treatments. The Pn of P4 treatment was higher in the N0 and N10 treatment groups (Figures 7A, B). The Pn of P4 and P6 treatments were higher in the N20 treatment group (Figure 7C). In the N40 treatment group, the Pn was higher in the P6 treatment (Figure 7D). We found that the Pn and leaf N:P of P.tanacetifolia leaves were normally distributed. The peak value of Pn appeared in the range of 14.5–17 of leaf N:P, and the Pn decreased when the leaf N:P was less than 14.5 or greater than 17 (Figure 7).
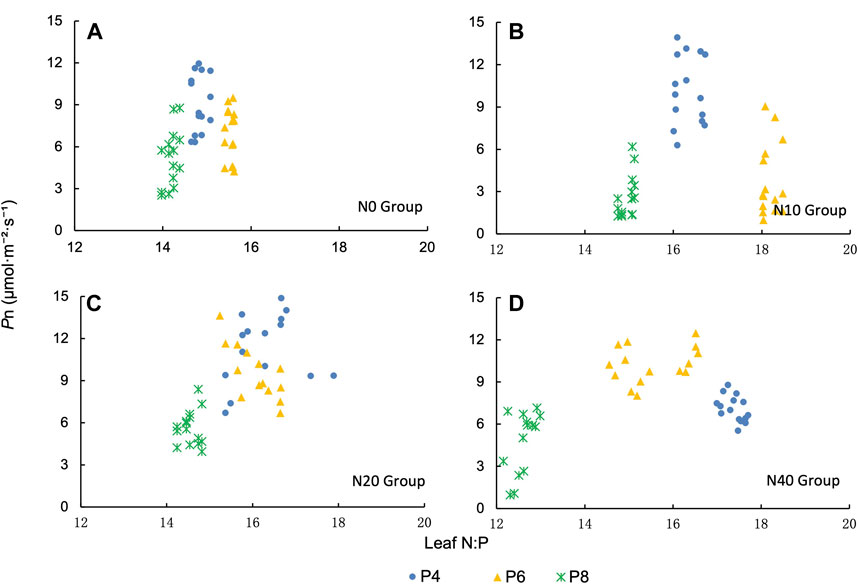
FIGURE 7. Relationship between N/P ratio and net photosynthetic rate (Pn) in P.tanacetifolia leaves. (A) N0 group, (B) N10 group, (C) N20 group and (D) N40 group.
Discussion
Plants can adopt different strategies to adapt to environmental changes, and different life forms have different responses to N and P environment. Their nutrient distribution patterns will show different ecological stoichiometry characteristics, under the environment of N addition the N content of plants will fluctuate accordingly (Soons et al., 2017; He et al., 2021). In our study, higher N concentrations (N20, N40) increased the leaf N content of P.tanacetifolia, but the N40 treatment did not show higher leaf N concentration than the N20 treatment as the N concentration continued to increase. This indicates that leaf N content will stabilize after a certain amount of uptake and will not increase with the increase of N addition. Leaf P content was elevated at higher N concentrations (N20, N40) compared to lower N concentrations (N10, N20) treatments (Figure 2). Similar results were found in Pennisetum centrasiaticum, where N addition increased leaf N and P content of P.centrasiaticum (Huang et al., 2018). Moreover, it can be seen from the coefficient of variation of leaf N and P nutrients in P.tanacetifolia that plant N nutrients are less sensitive to N addition, but more sensitive to P addition, and a small amount of P addition in low N (N0, N10) environment will increase leaf N (Figure 3). The results showed that P.tanacetifolia had unique characteristics of N and P nutrient adaptation. In the environment with low soil N concentration, N absorption can be promoted by P addition, while in the environment with high soil N concentration, the N:P could be reduced by increasing the P content in leaves, to weaken the limitation of plant nutrient imbalance.
Photosynthesis is an important way of plant energy acquisition and transformation, will produce different degrees of changes in different environments, which can be used as an important indicator of plant vitality in different environments (Atkin et al., 2015). It was found that the net photosynthetic rate of P.tanacetifolia shrubs in the eastern Qinghai-Tibet Plateau was significantly increased by 10 g N·m−1·yr−1 N addition, but inhibited when the concentration reached 15 g N·m−1·yr−1 (Xu et al., 2018). N addition significantly increased the net photosynthetic rate of Salix viminalis and Elymus nutans, but inhibited the net photosynthetic rate of Potentilla chinensis (Wang et al., 2019b). Therefore, the response of N addition to the net photosynthetic rate of Potentilla in different environments has not been unified. We found that the net photosynthetic rate decreased with P addition under no N (N0) and low N (N10) treatments, indicating that N was the main limiting factor for P.tanacetifolia photosynthesis within this range, and that photosynthesis reached the highest under medium N (N20) treatment, and P addition began to show a mitigating effect; Photosynthesis was not significantly improved when N was applied at high concentration (N40), but P addition still had a mitigating effect on photosynthesis (Figure 4A). These results confirm our first hypothesis that changes in net photosynthetic rate can reveal trends in N and P limitation by nitrogen deposition in broad-leaf forbs. However, high P addition did not increase net photosynthetic rate under P limitation. We further explored the changes of stomatal conductance under different treatments. Stomatal conductance is one of the important limiting factors of net photosynthetic rate. When stomatal conductance decreases, the amount of CO2 entering the stomata decreases, which cannot meet the requirements of photosynthesis, and it is called stomatal limitation of photosynthesis (Iturrate-Garcia et al., 2020). Studies have shown that the stomatal conductance of grassland plants will change with the addition of N, which has a serious impact on the net photosynthetic rate through the change of CO2 concentration (Cabrera et al., 2021; Zangani et al., 2021). The addition of N and P helps to improve the stomatal conductance and net photosynthetic rate, but the environment with higher P concentration will inhibit the CO2 assimilation rate, resulting in the decrease of stomatal conductance and net photosynthetic rate (Ben et al., 2019; Cabrera et al., 2021; Zangani et al., 2021). In our study, the response trend of stomatal conductance to N and P addition was basically consistent with that of net photosynthetic rate, and showed a positive correlation. Moderate N and P addition promoted net photosynthetic rate by increasing stomatal conductance, while excessive P addition inhibited net photosynthetic rate by reducing stomatal conductance (Figure 4B, Figure 5). This also explained the reason why the high concentration of P addition treatment cannot improve the net photosynthetic rate when P limitation occurred. However, the specific reasons for the inhibition of P.tanacetifolia photosynthesis by excessive P concentration need further experimental explanation. It can be concluded that P.tanacetifolia photosynthesis is mainly N-limited when the N application rate is less than 20 kg N·ha·yr−1, while it is mainly P-limited when the N application rate is greater than this value (Figure 5). This supports our second hypothesis that P addition in the context of N deposition improves the net photosynthetic rate of broad-leaf forbs. However, this improvement is limited. When P is added in excess, the resulting P limitation also inhibits net photosynthetic rate.
When the environmental N and P change, the content of N and P in plants will change accordingly, which makes the plat N:P fluctuate. Study have pointed out that the N:P in plants can be used as a basis for judging whether plants are limited by N or P in the environment (Güsewell, 2004). In the study of Sphagnum swamp plants in northern and southern Sweden, it was proposed that plants are limited by N when the N:P is less than 10, and limited by P when the N:P is greater than 14; In the study of 74 plants in Dutch grassland suggested that when the N:P is less than 10, it was limited by N, and when the N:P is greater than 20, it was limited by P; In the study of L.chinensis and Carex korshinskyi in the typical steppe of Inner Mongolia, it is proposed that when the N:P is less than 21, it is limited by N, and when the N:P is more than 23, it is limited by P (Güsewell, 2004; Zhang et al., 2004; Zhao et al., 2019). Therefore, the threshold of N/P ratio proposed by different plant communities in different regions is not fixed. In our study, the net photosynthetic rate of P.tanacetifolia had a linear relationship with the content of N and P in leaves, and the correlation between the N:P and net photosynthetic rate was normal distribution. When the N:P is 14.5–17.0, the net photosynthetic rate was at a higher level. The net photosynthetic rate of P.tanacetifolia decreased when it was below or above this range (Figures 6, 7). This supports our third hypothesis that there is a relationship between N:P and net photosynthetic rate of broad-leaf forbs. We preliminarily considered that the P.tanacetifolia is limited by N when the N:P is less than 14.5, and limited by P when the N:P is more than 17.0. The results of our experiment are not the same as those of the above experiments, indicating that there are certain differences between the stoichiometric ratio of N and P and the limitation of N and P in different plants, and more research accumulation is needed to verify the threshold of N or P limitation in main species.
Conclusion
N deposition caused nutrient imbalance of P.tanacetifolia by changing the proportion of soil nutrients, which had a serious impact on plant growth and reproduction. Therefore, appropriate P supplement should be carried out to avoid or reduce the impact of N deposition on community ecology when using P.tanacetifolia to build orchards or cultivate and expand.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
YL, JL, JY, and BR contributed to conceptualization and design of the study. JH performed the lab work. JH and HH analyzed the data. JH wrote the first draft of the manuscript. JH, HH, and LB contributed to writing, review and editing. LB contributed to funding acquisition. All authors have read and approved to the published version of the manuscript. JH and HH contributed equally to this work and share first authorship.
Funding
This research was financially supported by Liaoning Provincial Science and Technology Plan Projects (2020JH1/10300006) in China.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Atkin, O. K., Bloomfield, K. J., Reich, P. B., Tjoelker, M. G., Asner, G. P., Bonal, D., et al. (2015). Global variability in leaf respiration in relation to climate, plant functional types and leaf traits. New Phytol. 206 (2), 614–636. doi:10.1111/nph.13253
Bai, Y. F., Wu, J. G., Clark, C. M., Naeemz, S., Pan, Q. M., Huang, J. H., et al. (2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia grasslands. Glob. Change Biol. 16 (1), 358–372. doi:10.1111/j.1365-2486.2009.01950.x
Ben, H. S., Lefi, E., and Chaieb, M. (2019). Effect of phosphorus concentration on the photochemical stability of PSII and CO2 assimilation in Pistacia vera L. and Pistacia atlantica Desf. Plant Physiol. biochem. 142, 283–291. doi:10.1016/j.plaphy.2019.07.016
Cabrera, J. C. B., Hirl, R. T., Schaufele, R., Macdonald, A., and Schnyder, H. (2021). Stomatal conductance limited the CO2 response of grassland in the last century. BMC Biol. 19 (1), 50–14. doi:10.1186/s12915-021-00988-4
Du, E. Z., Liu, X. Y., and Fang, J. Y. (2014). Effects of nitrogen additions on biomass, stoichiometry and nutrient pools of moss Rhytidium rugosum in a boreal forest in Northeast China. Environ. Pollut. 188, 166–171. doi:10.1016/j.envpol.2014.02.011
Gao, Z. B., Wang, H. Y., Lu, X. T., and Wang, Z. W. (2017). Effects of nitrogen and phosphorus additionon C: N: P stoichiometry in roots and leaves of four dominant plant species in a meadow steppe of hulunbuir. Chin. J. Ecol. 36 (01), 80–88. doi:10.13292/j.1000-4890.201701.015
Guo, S. Q., Li, W. J., Zhang, R. Y., and Wang, G. (2014). Effects of N and P additions on foliar stoichiometry and photosynthetic characteristics of Potenitilla fruiticosa. Guihaia 34 (5), 629–634. doi:10.3969/j.isn.1000-3142.2014.05.009
Güsewell, S. (2004). N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 165 (2), 243–266. doi:10.1111/j.1469-8137.2004.01192.x
Hao, J. H., Chen, X., Qi, F. L., Li, Y. F., Liu, Y., Liu, J., et al. (2020). Comprehensive evaluation of native plant resources and their landscape application in sandy grassland of northwest Liaoning [J]. Acta Agric. Sin. 28 (03), 852–856. doi:10.11733/j.issn.1007-0435.2020.03.033
He, Y. E., Hong, M., Liang, Z., Tu, N. R., Wu, Z. D., Wang, L. Q., et al. (2021). Effects of precipitation and nitrogen deposition on litter decomposition of two perennial grasses in a desert steppe. Acta ecolo. Sin. 42 (7), 2910–2920. doi:10.5846/stxb202103230769
Huang, J. Y., Yu, H. L., Liu, J. L., Luo, C. K., Sun, Z. J., Ma, K. B., et al. (2018). Phosphorus addition changes belowground biomass and C:N:P stoichiometry of two desert steppe plants under simulated N deposition. Sci. Rep. 8 (1), 3400. doi:10.1038/s41598-018-21565-w
Ibáñez, I., Zak, D. R., Burton, A. J., and Pregitzer, K. S. (2018). Anthropogenic nitrogen deposition ameliorates the decline in tree growth caused by a drier climate. Ecology 99 (2), 411–420. doi:10.1002/ecy.2095
Iturrate-Garcia, M., Heijmans-Monique, M. P. D., Cornelissen, J. H. C., Schweingruber, F. H., Niklaus, P. A., and Schaepman-Strub, G. (2020). Plant trait response of tundra shrubs to permafrost thaw and nutrient addition. Biogeosciences 17 (20), 4981–4998. doi:10.5194/bg-17-4981-2020
Liu, C. C., Liu, Y. G., Guo, K., Qiao, X. G., Zhao, H. W., Wang, S. J., et al. (2018). Effects of nitrogen, phosphorus and potassium addition on the productivity of a karst grassland: Plant functional group and community perspectives. Ecol. Eng. 117, 84–95. doi:10.1016/j.ecoleng.2018.04.008
Liu, J. J., Wang, X. P., Gao, Y. F., Rong, Z. L., Zhang, G. D., Wang, W. B., et al. (2019). Leaf gas exchange and photosynthesis curves of Elymus nutans and Potentilla anserina under fencing and grazing conditions in the Qilian Mountains, Northwest China. J. Arid. Land. 11 (3), 431–445. doi:10.1007/s40333-019-0009-6
Liu, X. J., Zhang, Y., Han, W. X., Tang, A. H., Shen, J. L., Cui, Z. L., et al. (2013). Enhanced nitrogen deposition over China. Nature 494 (7438), 459–462. doi:10.1038/nature11917
Lu, J. P., Zhang, X. J., Liu, T. X., Zhang, W. R., and Liu, Y. (2021). Variation characteristics and source analysis of atmospheric nitrogen deposition flux on a reservoir in sand source areas of Beijing-Inner Mongolia. China Environ. Sci. 41 (3), 1034∼1044. doi:10.19674/j.cnki.issn1000-6923.2021.0115
Mao, Q. G., Lu, S. K., Mo, H., Gundersen, P., and Mo, J. M. (2018). Effects of simulated N deposition on foliar nutrient status, N metabolism and photosynthetic capacity of three dominant understory plant species in a mature tropical forest. Sci. Total Environ. 610, 555–562. doi:10.1016/j.scitotenv.2017.08.087
Payne, R. J., Dise, N. B., Field, C. D., Dore, A. J., Capron, S. J. M., and Stevens, C. J. (2017). Nitrogen deposition and plant biodiversity: Past, present, and future. Limnol. Oceanogr. Meth. 15 (8), 431–436. doi:10.1002/fee.1528
Prach, K., Vitovcova, K., Rehounkova, K., and Kralovec, J. (2021). Three decades of vegetation changes in a submontane grassland after the cessation of intensive fertilization. Preslia 93 (2), 169–179. doi:10.23855/preslia.2021.169
Qian, S. Y., Wang, Q., Liu, N., Quan, L., Lei, Z. F., Song, X. Z., et al. (2018). Effects of nitrogen deposition and phosphorus addition on photosynthesis and chlorophyll fluorescence characteristics of Chinese fir. Ecol. Sci. 37 (5), 113–121. doi:10.14108/j.cnki.1008-8873.2018.05.015
Sala, O. E., ChapinIII, F. S., Armesto, J. J., Berlow, E., Bloomfield, J., Dirzo, R., et al. (2000). Global biodiversity scenarios for the year 2100. Science 287 (5459), 1770–1774. doi:10.1126/science.287.5459.1770
Soons, M. B., Hefting, M. M., Dorland, E., Lamers, L. M. P., Versteeg, C., and Bobbink, R. (2017). Nitrogen effects on plant species richness in herbaceous communities are more widespread and stronger than those of phosphorus. Biol. Conserv. 212, 390–397. doi:10.1016/j.biocon.2016.12.006
Stevens, C. J., David, T. I., and Storkey, J. (2018). Atmospheric nitrogen deposition in terrestrial ecosystems: Its impact on plant communities and consequences across trophic levels. Funct. Ecol. 32 (7), 1757–1769. doi:10.1111/1365-2435.13063
Tian, Q. Y., Yang, L. Y., Ma, P. F., Zhou, H., Liu, N. N., et, al., et al. (2020). Below-ground-mediated and phase-dependent processes drive nitrogen-evoked community changes in grasslands. J. Ecol. 108 (5), 1874–1887. doi:10.1111/1365-2745.13415
Turner, B. L., and Wright, S. J. (2014). The response of microbial biomass and hydrolytic enzymes to a decade of nitrogen, phosphorus, and potassium addition in a lowland tropical rain forest. Biogeochemistry 117 (1), 115–130. doi:10.1007/s10533-013-9848-y
Wang, B., Gong, J. R., Zhang, Z. H., Yang, B., Liu, M., and Zhu, C. C. (2019a). Nitrogen addition alters photosynthetic carbon fixation, allocation of photoassimilates, and carbon partitioning of Leymus chinensis in a temperate grassland of Inner Mongolia. Agric. For. Meteorol. 279, 107743. doi:10.1016/j.agrformet.2019.107743
Wang, X. M., Liu, Q., Yan, B. C., Yang, B., Zhao, C., and Liu, G. C. (2019b). Responses of photosynthetic characteristics of Dodonaea visosa to nitrogen and phosphorus addition in a dry-hot valley. Acta Ecol. Sin. 39 (22), 8615–8629. doi:10.14067/j.cnki.1673-923x.2018.05.016
Xu, D. H., Gao, X. G., Gao, T. P., Mou, J., Li, J. H., Bu, H. Y., et al. (2018). Interactive effects of nitrogen and silicon addition on growth of five common plant species and structure of plant community in alpine meadow. Catena 169, 80–89. doi:10.1016/j.catena.2018.05.017
Yin, X., Zhang, L. J., Liu, X. J., Xu, W., Ni, Y. X., and Liu, X. Y. (2017). Nitrogen deposition in suburban croplands of Hebei Plain. Sci. Agric. Sin. 50 (4), 698–710. doi:10.3864/j.issn.0578-1752.2017.04.010
You, C. M., Wu, F. Z., Yang, W. Q., Xu, Z. F., Tan, B., Yue, K., et al. (2018). Nutrient-limited conditions determine the responses of foliar nitrogen and phosphorus stoichiometry to nitrogen addition: A global meta-analysis. Environ. Pollut. 241, 740–749. doi:10.1016/j.envpol.2018.06.018
Yu, Y., Cheng, H., Wu, B., and Wang, C. (2022). Combined effects of drought stress and different forms of nitrogen deposition as response mechanism to environmental change. Int. J. Environ. Sci. Technol. 2022, 1–16. doi:10.1007/s13762-022-04408-0
Yuan, Q., Zhang, Y., Chen, H., Zhou, L., Yu, D. P., Wang, Q. W., et al. (2021). Plasticity of N and P content in typical plants responses to short-term nitrogen input in tundra zone of the Changbai Mountain, China. Chin. J. Ecol. 40 (11), 3493–3500. doi:10.13292/j.1000-4890.202111.014
Zangani, E., Afsahi, K., Shekari, F., Mac, S. E., and Mastinu, A. (2021). Nitrogen and phosphorus addition to soil improves seed yield, foliar stomatal conductance, and the photosynthetic response of rapeseed (Brassica napus L.). Agriculture-Basel. 11 (6), 483. doi:10.3390/agriculture11060483
Zhang, L. X., Bai, Y. F., and Han, X. G. (2004). Different responses of N:P stoichiometry of leymus chinensis and carex korshinskyi to n additions in a steppe ecosystem in Nei Mongo. Acta Bot. Sin. 46 (3), 259–270. doi:10.3321/j.issn:1672-9072.2004.03.002
Zhao, Y. A., Yang, B., Li, M. X., Xiao, R. Q., Rao, K. Y., Wang, J. Q., et al. (2019). Community composition, structure and productivity in response to nitrogen and phosphorus additions in a temperate meadow. Sci. Total Environ. 654, 863–871. doi:10.1016/j.scitotenv.2018.11.155
Keywords: nitrogen deposition, phosphorus addition, Potentilla tanacetifolia, nitrogen and phosphorus content, photosynthetic rate, stomatal conductance, N:P ratio
Citation: Hao J, Han H, Liu Y, Li J, Yang J, Ren B and Bai L (2023) Phosphorus addition alleviates the inhibition of nitrogen deposition on photosynthesis of Potentilla tanacetifolia. Front. Environ. Sci. 11:1099203. doi: 10.3389/fenvs.2023.1099203
Received: 21 November 2022; Accepted: 02 January 2023;
Published: 13 January 2023.
Edited by:
Junhong Bai, Beijing Normal University, ChinaReviewed by:
Fude Liu, Tianjin University of Technology, ChinaPetr Holub, Czech Academy of Sciences, Czechia
Copyright © 2023 Hao, Han, Liu, Li, Yang, Ren and Bai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Long Bai, YmFpbG9uZzIwMThAc3lhdS5lZHUuY24=
†These authors have contributed equally to this work and share first authorship