 Ivan Raniero Hernández-Salmerón*
Ivan Raniero Hernández-Salmerón* Milena Holmgren*
Milena Holmgren*- Wildlife Ecology and Conservation Group, Department of Environmental Sciences, Wageningen University and Research, Wageningen, Netherlands
Agricultural and ecological droughts, extreme heat and aridity have high impacts on livestock and pasture systems worldwide. Finding ways to adapt production systems and increase biomass under these new conditions is urgently needed. The availability of tree shade in these pastures could potentially ameliorate the impacts of warm weather. Yet, the effects of tree cover on the productivity of livestock rangelands are hotly debated. We performed a global meta-analysis to evaluate the effects of tree cover on grass biomass during contrasting seasons within the same system and along environmental gradients in tropical and temperate productive systems. We also assessed the levels of canopy density at which tree cover effects are observable. We observed that trees facilitate grass biomass during dry seasons, especially in the tropics and dry regions. These positive effects are more likely to occur at intermediate levels of evapotranspiration and irradiance. Our findings suggest that integrating trees in pastures might increase resilience of current livestock production systems to drier and warmer conditions.
Introduction
Livestock rangelands expand through tropical and temperate regions covering approximately 25% of our planet (Steinfeld et al., 2006). They differ in structure and composition from practically treeless pastures to landscapes with scattered trees or woody plant patches within a matrix of grasses. How trees and grasses interact has fascinated rangers, ecologists, and conservationists alike because the outcome determines the resources available to livestock and wild species (Scholes & Archer, 1997; Treydte et al., 2007; Harvey et al., 2011; Costa et al., 2016) and therefore food security (Vira et al., 2015), ecosystem services (Millennium Ecosystem Assessment, 2005) and nature conservation (Harvey et al., 2008). Tree-grass interactions have enormous implications for the management of rangelands under climate change and the conservation of biodiversity. Agricultural and ecological droughts, extreme heat and aridity have high impacts on livestock and pasture systems (IPCC, 2021). Increasing tree cover in rangelands could contribute to ameliorate the impacts of higher temperatures, and erratic rains as climate change progresses (Murgueitio et al., 2011; Altieri et al., 2015; Solorio et al., 2017). This climate change adaptation strategy in productive systems could also have positive effects on biodiversity conservation. By increasing tree cover, wild species movement across landscapes is facilitated, fostering genetic diversity and migration that may contribute to maintaining ecological networks. However, the management of multifunctional landscapes aiming to promote win-win solutions that combine sustainable production with nature conservation and climate change mitigation remains challenging (Bustamante et al., 2014; Mbow et al., 2014).
Early work on tree-grass interactions emphasized competitive effects and promoted a view on production systems where grasses would benefit from treeless landscapes (Walter & Burnett, 1971; Walker & Noy-Meir, 1982; Walker & Salt, 2012). Young trees and grasses usually compete for water and soil resources, but as trees become taller and reach deeper soil layers, competition for soil resources becomes less relevant for trees and light competition more relevant for grasses (Bazzaz, 1979; Smith & Huston, 1990). This emphasis on negative interactions was softened in the nineties by the appreciation of positive effects of trees on grass productivity under abiotic stress conditions, such as droughts and heatwaves or poor soil fertility (Belsky, 1994; Holmgren et al., 1997; Scholes & Archer, 1997; Valladares et al., 2016).
There is a vast literature on the effects of trees on grasses in natural ecosystems such as savannas (Blaser et al., 2013; Dohn et al., 2013) and woodlands (Jackson & Ash, 1998; Barbier et al., 2008). Yet, the debate on the overall effects of trees on grasses in productive systems persists. Different studies have reported contrasting results about the levels of tree canopy density and the environmental conditions under which trees may have positive, negative or neutral effects (Treydte et al., 2007; Moustakas et al., 2013; Bernardi et al., 2016; Ansley et al., 2019). The most recent syntheses of the published literature have found tree facilitation on grasses to be stronger under N2-fixing trees and in dry environments (Rivest et al., 2013; Mazía et al., 2016). We build upon these last studies to systematically evaluate whether tree cover increases grass biomass 1) during contrasting seasons, 2) along expanded environmental gradients in tropical and temperate pastures, and 3) at different levels of canopy density. We report, for the first time, the contrasting seasonal effects of trees on productive systems around the globe and show that trees have stronger positive effects on grass yields under intermediate levels of abiotic stress. These findings contribute to promote win-win solutions in current livestock production systems with focus on climate change mitigation, food security, and sustainability.
Methods
Paper selection
We searched for studies that evaluated grass performance (i.e., dry matter) under two contrasting seasons and two or more levels of tree cover. Searches were conducted in Web of Science (1945–2020) using the following keywords: “shade” OR “light” OR “irradiance” OR “shelter” OR “tree*” OR “canop*” OR “crown” OR “Sun*” OR “arbol” OR “cobertura arborea” OR “luz” OR “irradiaci?n” OR “protecci?n” OR “sombra” OR “dosel arbo*”; “drought” OR “water” OR “precipitation” OR “wet” OR “humid” OR “dissecat*” OR “arid*” OR “irrigat*” OR “dry” OR “rain*” OR “microclimat*” OR “temperat*” OR “sequia” OR “desecaci?n” OR “agua” OR “precipitaci?n” OR “humed*” OR “irrigaci?n” OR “seco” OR “microclima” OR “temperature”; “grass” OR “forage” OR “fodder” OR “grass” OR “pasture” OR “animal producti*” OR “producti*” OR “herbs” OR “herbace*” OR “forraje” OR “pasto” OR “hierba” OR “pastizal*” OR “leche” OR “carne” OR “pastura”; “pasture” OR “S?lvo-pastor*” OR “Agro-s?lvo-pastor*” OR “Agros?lvopastor*” OR “livestock” OR “wood* pasture*” OR “wood* grassland” OR “wood* rangeland” OR “ranching land” OR “pastureland” OR “ganado” OR “ganader*” OR “vaca” OR “cattle” OR “s?lvopastor*.” We chose dry matter as it was the most commonly used indicator of forage biomass. We registered the levels of tree cover and expressed it in a percentage scale of irradiance where 100% would represent the irradiance that reaches and open microsite (i.e., 0% of tree cover). This relative scale is adequate to compare the response of species that naturally occur under different ranges of light availability (Holmgren et al., 2012). We decided not to include studies that reported grass performance at only one season or one level of tree cover because we wanted to explicitly assess season-tree cover interactions. We screened publications for studies that included grass responses within the same calendar year or continuous year, specific location or coordinates where the study was carried out, and period in time when the measures were taken. We only considered studies conducted in the field and with natural tree shade.
The search from 5,135 papers yielded 174 studies from 33 suitable publications. When publications involved several grass species or irradiance levels, each species and each irradiance level was treated as a separate study. We decided to include several studies from the same paper because, although it tends to reduce the overall heterogeneity in effect sizes, excluding multiple results from a paper can underestimate effect sizes (Gurevitch & Hedges, 1999; Karst et al., 2008).
Data collection
Mean values of grass dry matter were collected from text and tables in the main publication and/or supplementary information. We used WebPlotDigitizer (Rohatgi, 2020) to extract mean and standard error values from figures when raw data was not provided. If not provided, standard deviations were back calculated from standard errors and sample sizes (SD = SE × √n). When there were multiple studies within the same publication, we calculated several means (i.e., one per study), pairing the different levels of tree cover with the one with the lowest tree cover (highest irradiance). Some publications reported multiple responses under the same levels of tree cover within the same season, thus we averaged those responses to one value per tree cover level and season.
For each record in our dataset, we converted the grass dry matter mean to kg/ha/d and classified the grass species as C3 or C4. We registered the location, country and biome where the study took place. We obtained evapotranspiration (mm/day), monthly precipitation (mm) and maximum daily temperature (°C) based on the period of time of the records and coordinates, and distinguished between dry and wet seasons. Environmental data was obtained using the R package climatrends (de Sousa et al., 2020). The literature search workflow is presented in Figure 1.
FIGURE 1. Flow diagram for the selection of manuscripts.
Effect size calculations
Following identification, means (X), standard deviations (SDs), and sample sizes (n) were extracted from the published studies. If not reported, these statistics were derived from other metrics. We conducted a meta-analysis to assess the effects of tree cover and season on grass biomass following (Koricheva et al., 2013). For each study, we calculated the effect size using the natural log of the response ratio [ln (RR)] and its associated variance
We used the escalc and rma. mv functions in the metafor package in R 4.0.3 (R Core Team, 2020) to calculate the ln (RR) and to perform the statistical analysis. To address the question of how tree cover influences the response of grasses under contrasting seasons, we performed several multivariate meta-analysis models with random effects. We first analyzed whether the effect of tree cover differs between seasons and C3/C4 species across the different biomes. We used the ln (RR) for tree cover as the response variable, and the Seasons, biome and C3/C4 species as predictors (with interactions: Seasons *C3/C4). We assessed the effects of tree cover along environmental gradients of evapotranspiration, monthly precipitation, maximum, daily temperature and irradiance. We grouped biomes into two major regions: Temperate (i.e., Temperate, Mediterranean and Desert biomes) and Tropical (i.e., Tropical and subtropical biomes). Mediterranean and desert biomes were grouped in the Temperate region based on their mean annual precipitation and mean annual temperature. We fitted meta-regression models with evapotranspiration, major region and C3/C4 species as predictors (with interactions Evapotranspiration*C3/C4 and major region* C3/C4). Both linear and quadratic regressions were fit and the best model chosen using the Akaike Information Criterion corrected for small sample sizes (AICc, Supplementary Table S1). The same models were fitted replacing evapotranspiration for maximum daily temperature and monthly precipitation separately. Lastly, we analyzed the type of response of the effect of tree cover to irradiance and tree type (i.e., functional group) with Irradiance, Tree type, Seasons and C3/C4 species as predictors (with interactions Irradiance* Seasons, Irradiance * Tree type and C3/4* Seasons). In all the models we included the Study nested within the Paper as random factors.
Results
Tree cover effects vary between seasons and biomes
We identified 174 studies that assessed grass biomass under contrasting seasons and levels of tree cover across seven biomes following PRISMA guidelines (Moher et al., 2009, Supplementary Figure S1). About 61% were conducted in the tropics and 39% in temperate regions (Figure 2).

FIGURE 2. (A) Temperate pastures in Durazno, Uruguay during the dry season. (B) Dry tropical pastures in Yucatan, Mexico during the dry season. (C) Worldwide locations of studies. Studies from the same site are represented by a single dot.
The effect of tree cover on grass biomass varied between biomes, seasons and C3/C4 grass species (p < 0.0001, Figure 3; Supplementary Figure S2; Supplementary Table S2). We found the largest tree facilitative effects in the Desert and Xeric shrublands, especially during the dry season. In tropical and subtropical biomes, we found mostly tree facilitative effects on C4 grasses during the dry season. Also in the Mediterranean, tree facilitative effects, for the dominant C3 grasses, were higher during the dry season than during the wet season. In contrast, in temperate regions the effects varied per biome; in temperate grasslands, savannas and shrublands, tree facilitative effects on C4 grass species occurred only during the wet season; while in broadleaf forests we found neutral effects on C3 grasses during both seasons and higher facilitative effects on C4 grass species during the dry season.

FIGURE 3. Tree effects on grass biomass in the dry and wet seasons across biomes. Facilitative effects are stronger in Desert and Xeric biomes and Tropical and subtropical biomes where C4 grasses occur, especially during the dry season. Tree effects vary in the temperate biomes: facilitation occurs in grasslands where C4 grasses dominate; in temperate forests mostly neutral to negative effects are observed. Dotted lines indicate zero effect sizes. 174 studies were identified of which 61% were conducted in the tropics and 39% in temperate regions. Point size indicates the sample size.
Tree cover effects depend on rainfall, temperature and irradiance
The positive effects of trees on grass biomass for C3 and C4 grasses peak at intermediate levels of daily evapotranspiration (4 and 5 mm for C3 and C4 grasses, respectively) and become negative at both ends of the evapotranspiration gradient (Figure 4A). We grouped the biomes in two major regions: tropical and temperate, and did not find differences between them.
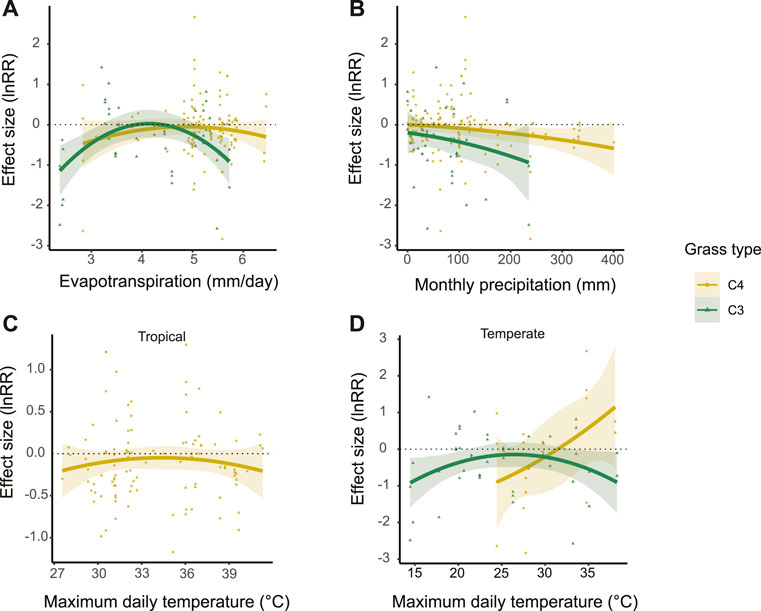
FIGURE 4. Tree effects on C3 and C4 grass biomass along gradients of (A) evapotranspiration, (B) monthly precipitation and maximum daily temperature in tropical (C) and temperate regions (D). Dotted lines indicate zero effect sizes.
When analyzing the contribution of rainfall and temperature separately, we found contrasting patterns on the role of trees on grass biomass along these climatic gradients. The effects of trees on both C3 and C4 grasses became increasingly negative as seasonal rainfall increases (p < 0.005, Figure 4B), especially for C3 species (p < 0.0001), in both tropical and temperate regions. While we found only negative effects on C3 grass species along the whole gradient of precipitation, we observed neutral to slightly positive effects on C4 grasses when monthly precipitation drops below ∼50 mm.
The nonlinear patterns of how trees and grasses interact are mostly explained by temperature. In the tropics, the effects of trees on C4 grass biomass are mostly neutral at intermediate maximum daily temperatures (∼35°C) and become negative at both colder and hotter ends of the temperature gradient (Figure 4C). Also in temperate regions, trees have neutral effects on C3 grasses at intermediate temperature (∼27°C) that become negative at both ends of the gradient of maximum daily temperature. In temperate regions, trees have positive effects on C4 grasses as conditions become warmer than ∼32°C (Figure 4D).
Tree cover density determines the levels of irradiance received by grasses. Overall, we found that the positive effect of trees on grass biomass peaks at roughly 60%–80% of irradiance and becomes negative at lower or higher irradiance levels (Supplementary Figure S2). This facilitative effect tends to be higher under N2 fixing trees than under Non-N2 fixing trees although it is not significantly different (p = 0.5). The response of grasses to the irradiance levels are strongly dependent on the rainfall season and the grass type. C4 grasses respond negatively to lower irradiance levels especially during the rainy season. In contrast, C3 grasses are facilitated by intermediate levels of irradiance both in the dry and wet seasons (Figure 5).
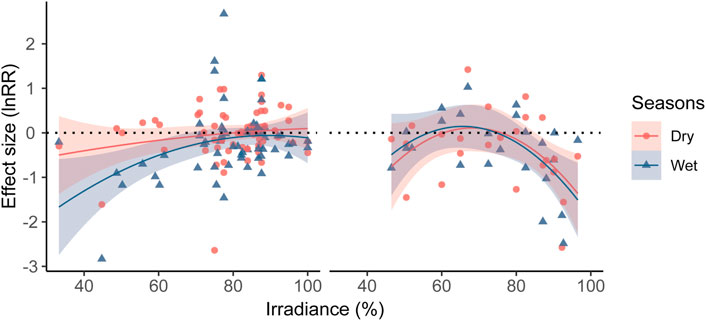
FIGURE 5. Tree effects on C3 and C4 grass biomass along the irradiance gradient in the dry and wet seasons. Dotted lines indicate zero effect sizes.
Discussion
Here we report for the first time the contrasting seasonal effects of trees on productive systems around the globe. Our meta-analysis shows that the effects of trees on grass biomass have been studied more often in tropical and subtropical pastures than in temperate pastures. Overall, our results indicate that positive effects of trees on grasses: 1) are higher during seasonal droughts, especially on C4 grasses in tropical pastures and 2) peak at intermediate levels of evapotranspiration, temperature and irradiance, and at low levels of precipitation. These findings show that water, temperature and light, are key resources for grass growth and their interactions can shape the nature of tree-grass interactions in a wide range of productive ecosystems.
Facilitation is higher during seasonal droughts in the tropics
This meta-analysis shows that trees increase grass biomass in pastures during seasonal droughts and that these effects are stronger in tropical pastures where C4 grasses dominate, than in temperate pastures where C3 grasses are more common. The higher tree facilitative effects during droughts on C4 grasses can result from a combination of mechanisms that ameliorate abiotic stress. Although C4 grasses have been widely promoted across tropical productive systems (Edwards et al., 2010) for being highly tolerant to water stress and high temperatures (Chaves et al., 2003), they still benefit strongly from the shade of trees. During seasonal droughts in the dry tropics, when temperature and evapotranspiration increase sharply, the canopy of trees ameliorates environmental stress by reducing temperature and mediating irradiance levels even for highly tolerant C4 grasses.
In temperate regions, we found mostly, but not exclusively, neutral to negative effects of tree cover on grass biomass. C3 grasses occur more often in temperate regions and have a higher tolerance and photosynthetic capacity in colder temperatures (Saborsky & Mitsui, 1982; Gardner et al., 2017). Interestingly, the only positive effect of trees we found in temperate regions was described for open temperate savanna, specifically on C4 grasses during the wet season. This is very contrasting to the facilitative effects for C4 grasses in the tropical pastures which occur mostly in the dry season. Temperate savannas are open biomes with high levels of irradiance which in combination with high water availability are less stressful for grasses. However, the wet season often corresponds with the coldest period of the year. In such open biomes, trees might ameliorate temperature in an opposite direction than in the tropics. While in the tropics abiotic amelioration implies a reduction in high temperature levels, in the temperate pastures, amelioration may be related to an increase in temperature under the tree canopy (Zellweger et al., 2019), creating a warmer microclimate that would benefit C4 grasses during wet and cold seasons.
Facilitation tends to peak at intermediate abiotic stress
We found a shift from neutral effects to negative effects of trees on grasses as monthly precipitation increases. Previous meta-analyses on the effect of trees on grass biomass in natural savannas also found stronger facilitation at drier places (Moustakas et al., 2013) or with decreasing annual precipitation (Dohn et al., 2013).
Our meta-analysis also revealed non-linear patterns of tree facilitative effects on grasses. These effects peak at intermediate levels of evapotranspiration, temperature and irradiance and can be lost at very low or high levels. Tree cover plays a key role in the interplay between positive and negative effects. In plant communities, the shade of neighboring plants is expected to increasingly ameliorate drought stress as conditions become drier (Bertness & Callaway, 1994), although these facilitative effects may be lost under extremely stressful conditions (Holmgren & Scheffer, 2010; Soliveres et al., 2015; Zhang et al., 2018). These non-linear effects of shade and drought have been found in meta-analyses of plant performance in field and experimental conditions (Holmgren et al., 2012) as well as in agroforests (Blaser et al., 2018). We now found comparable results for productive livestock pastures.
Although there was a tendency for N2-fixing trees to have slightly higher facilitative effects than Non-N2-fixing trees on grass biomass, we did not find significant differences between these two tree functional groups. N2-fixing trees have been reported previously to increase pasture yields as drought pressure increases abiotic stress on livestock grazing systems (Rivest et al., 2013). Palm trees are highly abundant in tropical livestock pastures where farmers preserve them for their multipurpose value (Martínez-Ballesté et al., 2008; Macía et al., 2011), however, their effect on grasses is still poorly known compared to other groups of trees that have been studied more in detail. We found only one publication where the effects of palm trees on livestock pastures have been reported (Esquivel, 2007). Field experiments and long-term observational studies are needed to understand the effects of palm trees on grasses and determine whether they can contribute to increasing resilience to drought in livestock pastures.
We compared studies from seven different biomes that varied in time, period of the year of sampling, duration, and research methodology. Despite this variation, we identified higher facilitative effects of trees on grasses during seasonal droughts, especially in the tropical biomes where C4 grasses dominate and to a lesser degree in the temperate pastures. We showed that these effects are not only strongly related to the available levels of precipitation but also to temperature and irradiance. Understanding the effect of trees is crucial for managing and transforming current livestock production systems into multifunctional landscapes with increased resilience to seasonal droughts around the globe. We encourage the integration of trees with intermediate canopy density (i.e. ∼20%–40%) in drier and warmer pastures, especially in the tropics where trees have positive and neutral effects on grass biomass. Positive and neutral effects of trees on grass biomass imply that trees can be used in productive pastures to ameliorate abiotic weather conditions favoring, or at least not compromising grass yields, while also contributing to other ecosystem services such as biodiversity conservation, carbon sequestration, habitat provision, construction materials, etc.
Challenges ahead
Our understanding of how trees influence the functioning of productive pasture systems is still fragmented. The existing literature has focused on identifying the effects of different types of trees according to their functional traits such as N2-fixation or deciduousness (Rivest et al., 2013; Mazía et al., 2016) but we lack holistic assessments of the direct and indirect effects of tree diversity on livestock productivity. One major challenge is to widely assess how tree diversity affects both grass and animal production and how these effects can be translated into economic benefits for farmers while considering multiple ecosystem services at meaningful temporal scales.
Most of the studies we identified were carried out in the neotropics, especially in Latin America, which may reflect different cultural traditions in either how farmers perceive the separation between productive versus natural landscapes or how scientists study them in different regions around the world. We encourage researchers to bridge across scientific disciplines to expand the knowledge of tree-grass interactions in agroecosystems and multifunctional landscapes across environmental gradients and cultures. Merging the existing traditional knowledge from farmers with the ecological theory developed in natural and productive systems could facilitate generating effective strategies for sustainable productive systems that can contribute to biodiversity conservation and be better adapted to changing climate conditions.
Data availability statement
The raw data supporting the conclusion of this article can be found in the repository https://10.5281/zenodo.6824373 (Hernández-Salmerón and Holmgren 2022).
Author contributions
IH-S and MH conceived the ideas, designed the study, analysed and interpreted the results, and wrote the manuscript. IH-S screened all potential papers, extracted the data and prepared the figures.
Funding
IH-S was funded by a PhD grant from Consejo Nacional de Ciencia y Tecnología, México, CVU 610547. Funds for open access publishing were granted by Wageningen University and Research.
Acknowledgments
The authors acknowledge the Tropical Agricultural Research and Higher Education Center (CATIE) for facilitating access to their library.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2022.949185/full#supplementary-material
References
Altieri, M. A., Nicholls, C. I., Henao, A., and Lana, M. A. (2015). Agroecology and the design of climate change-resilient farming systems. Agron. Sustain. Dev. 35, 869–890. doi:10.1007/s13593-015-0285-2
Ansley, R. J., Cooper, C., and Zhang, T. (2019). Invasion promotes invasion: Facilitation of C3 perennial grass dominance in mixed C3/C4 grassland by an invasive C3 woody sprouter (prosopis glandulosa). Ecol. Evol. 9 (23), 13438–13449. doi:10.1002/ece3.5800
Barbier, S., Gosselin, F., and Balandier, P. (2008). Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manage. 254, 1–15. doi:10.1016/j.foreco.2007.09.038
Bazzaz, F. (1979). The physiological ecology of plant succession. Annu. Rev. Ecol. Syst. 10, 351–371. doi:10.1146/annurev.es.10.110179.002031
Belsky, A. J. (1994). Influences of trees on savanna productivity: Tests of shade, nutrients, and tree‐grass competition. Ecology 75, 922–932. doi:10.2307/1939416
Bernardi, R. E., de Jonge, I. K., and Holmgren, M. (2016). Trees improve forage quality and abundance in South American subtropical grasslands. Agric. Ecosyst. Environ. 232, 227–231. doi:10.1016/j.agee.2016.08.003
Bertness, M. D., and Callaway, R. (1994). Positive interactions in communities. Trends Ecol. Evol. 9, 191–193. doi:10.1016/0169-5347(94)90088-4
Blaser, W. J., Sitters, J., Hart, S. P., Edwards, P. J., and Olde Venterink, H. (2013). Facilitative or competitive effects of woody plants on understorey vegetation depend on N‐fixation, canopy shape and rainfall. J. Ecol. 101, 1598–1603. doi:10.1111/1365-2745.12142
Blaser, W. J., Oppong, J., Hart, S. P., Landolt, J., Yeboah, E., and Six, J. (2018). Climate-smart sustainable agriculture in low-to-intermediate shade agroforests. Nat. Sustain. 1, 234–239. doi:10.1038/s41893-018-0062-8
Bustamante, M., Robledo‐Abad, C., Harper, R., Mbow, C., Ravindranat, N. H., Sperling, F., et al. (2014). Co‐benefits, trade‐offs, barriers and policies for greenhouse gas mitigation in the agriculture, forestry and other land use (AFOLU) sector. Glob. Change Biol. 20, 3270–3290. doi:10.1111/gcb.12591
Chaves, M. M., Maroco, J. P., and Pereira, J. S. (2003). Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 30, 239–264. doi:10.1071/fp02076
Costa, S. B. d. M., de Mello, A. C., Dubeux, J. C., dos Santos, M. V., Lira, M. d. A., Oliveira, J. T., et al. (2016). Livestock performance in warm‐climate silvopastures using tree legumes. Agron. J. 108, 2026–2035. doi:10.2134/AGRONJ20-16.03.0180
Dohn, J., Dembélé, F., Karembé, M., Moustakas, A., Amévor, K. A., and Hanan, N. P. (2013). Tree effects on grass growth in savannas: Competition, facilitation and the stress‐gradient hypothesis. J. Ecol. 101, 202–209. doi:10.1111/1365-2745.12010
Edwards, E. J., Osborne, C. P., Stromberg, C. A., Smith, S. A., Consortium, C. G., Bond, W. J., et al. (2010). The origins of C4 grasslands: Integrating evolutionary and ecosystem science. Science 328, 587–591. doi:10.1126/science.1177216
Esquivel, H. (2007). Tree resources in traditional silvopastoral systems and their impact on productivity and nutritive value of pastures in the dry tropics of Costa Rica. San José: CATIE.
Gardner, F. P., Pearce, R. B., and Mitchell, R. L. (2017). Physiology of crop plants. Iowa: Scientific publishers.
Gurevitch, J., and Hedges, L. V. (1999). Statistical issues in ecological meta‐analyses. Ecol 80, 1142–1149. doi:10.1890/0012-9658(1999)080[1142:SIIEMA]2.0.CO;2
Harvey, C. A., Komar, O., Chazdon, R., Ferguson, B. G., Finegan, B., Griffith, D. M., et al. Working Group on, B. & Conservation Value of Agricultural Landscapes of, M. (2008). Integrating agricultural landscapes with biodiversity conservation in the Mesoamerican hotspot. Conserv. Biol. 22, 8–15. doi:10.1111/j.1523-1739.2007.00863.x
Harvey, C. A., Villanueva, C., Esquivel, H., Gómez, R., Ibrahim, M., Lopez, M., et al. (2011). Conservation value of dispersed tree cover threatened by pasture management. For. Ecol. Manage. 261, 1664–1674. doi:10.10-16/j.foreco.2010.11.004
Hedges, L. V., Gurevitch, J., and Curtis, P. S. (1999). The meta‐analysis of response ratios in experimental ecology. Ecology 80, 1150–1156. doi:10.1890/0012-9658(1999)080[1150:tmaorr]2.0.co;2
Hernández-Salmerón, I. R., and Milena, H. (2022). Data from: Native palms and trees mediate drought impacts on dry neotropical pastures, doi:10.5281/zenodo.6824373
Holmgren, M., Gomez-Aparicio, L., Quero, J. L., and Valladares, F. (2012). Non-linear effects of drought under shade: Reconciling physiological and ecological models in plant communities. Oecologia 169, 293–305. doi:10.1007/s00442-011-2196-5
Holmgren, M., Scheffer, M., and Huston, M. A. (1997). The interplay of facilitation and competition in plant communities. Ecology 78, 1966–1975. doi:10.1890/0012-9658(1997)078[1966:TIOFAC]2.0.CO;2
Holmgren, M., and Scheffer, M. (2010). Strong facilitation in mild environments: The stress gradient hypothesis revisited. J. Ecol. 98, 1269–1275. doi:10.1111/j.13652745.2010.01-709.x
IPCC (2021). Climate change 2021: The physical science basis. Contribution of working group I to the sixth assessment report of the intergovernmental panel on climate. United Kingdom and New York: Cambridge University Press. In press. doi:10.1017/97810091-57896
Jackson, J., and Ash, A. (1998). Tree-grass relationships in open eucalypt woodlands of northeastern Australia: Influence of trees on pasture productivity, forage quality and species distribution. Agrofor. Syst. 40, 159–176. doi:10.1023/A:1006067110870
Karst, J., Marczak, L., Jones, M. D., and Turkington, R. (2008). The mutualism–parasitism continuum in ectomycorrhizas: A quantitative assessment using meta‐analysis. Ecology 89, 1032–1042. doi:10.1890/07-0823.1
Koricheva, J., Gurevitch, J., and Mengersen, K. (2013). Handbook of meta-analysis in ecology and evolution. Princeton University Press.
Macía, M. J., Armesilla, P. J., Cámara-Leret, R., Paniagua-Zambrana, N., Villalba, S., Balslev, H., et al. (2011). Palm uses in northwestern south America: A quantitative review. Bot. Rev. 77, 462–570. doi:10.1007/s12229-011-9086-8
Martínez-Ballesté, A., Martorell, C., and Caballero, J. (2008). The effect of Maya traditional harvesting on the leaf production, and demographic parameters of Sabal palm in the Yucatan Peninsula. Mexico. For. Ecol. Manag. 256, 1320–1324. doi:10.1016/j.foreco.2008.06.029
Mazía, N., Moyano, J., Perez, L., Aguiar, S., Garibaldi, L. A., and Schlichter, T. (2016). The sign and magnitude of tree–grass interaction along a global environmental gradient. Glob. Ecol. Biogeogr. 25, 1510–1519. doi:10.1111/geb.12518
Mbow, C., Smith, P., Skole, D., Duguma, L., and Bustamante, M. (2014). Achieving mitigation and adaptation to climate change through sustainable agroforestry practices in Africa. Curr. Opin. Environ. Sustain. 6, 8–14. doi:10.1016/j.cosust.2013.09.002
Millennium Ecosystem Assessment (2005). Ecosystems and human well-being: Synthesis. Washington, DC: Island Press.
Moher, D., Liberati, A., Tetzlaff, J., Altman, D. G., and Group, P. (2009). Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 6, e1000097. doi:10.1371/journal.pmed.1000097
Moustakas, A., Kunin, W. E., Cameron, T. C., and Sankaran, M. (2013). Facilitation or competition? Tree effects on grass biomass across a precipitation gradient. PLoS ONE 8, e57025. doi:10.1371/journal.pone.0057025
Murgueitio, E., Calle, Z., Uribe, F., Calle, A., and Solorio, B. (2011). Native trees and shrubs for the productive rehabilitation of tropical cattle ranching lands. For. Ecol. Manage. 261, 1654–1663. doi:10.1016/j.foreco.2010.09.027
R Core Team (2020). R: A language and environment for statistical computing. Vienna, Austria. Available at: https://www.R-project.org/.
Rivest, D., Paquette, A., Moreno, G., and Messier, C. (2013). A meta-analysis reveals mostly neutral influence of scattered trees on pasture yield along with some contrasted effects depending on functional groups and rainfall conditions. Agric. Ecosyst. Environ. 165, 74–79. doi:10.1016/j.agee.2012.12.010
Rohatgi, A. (2020). WebPlotDigitizer. Pacifica, CA, USA. Available at: https://automeris.io/WebPlotDigitizer.
Saborsky, O., and Mitsui, A. (1982). “Response of terrestrial plants to light quality, light intensity, temperature, CO2, and O2,” in Handbook of biosolar resources (Boca Raton: CRC Press).
Scholes, R. J., and Archer, S. R. (1997). Tree-grass interactions in savannas. Annu. Rev. Ecol. Syst. 28, 517–544. doi:10.1146/annurev.ecolsys.28.1.517
Smith, T., and Huston, M. (1990). “A theory of the spatial and temporal dynamics of plant communities,” in Progress in theoretical vegetation science. Editors G. Grabherr, L. Mucina, M. B. Dale, and C. J. F. Braak (Springer), 49–69.
Soliveres, S., Smit, C., and Maestre, F. T. (2015). Moving forward on facilitation research: Response to changing environments and effects on the diversity, functioning and evolution of plant communities. Biol. Rev. 90, 297–313. doi:10.1111/brv.12110
Solorio, S., Wright, J., Franco, M., Basu, S., Sarabia, S., Ramírez, L., et al. (2017). “Silvopastoral systems: Best agroecological practice for resilient production systems under dryland and drought conditions,” in Quantification of climate variability, adaptation and mitigation for agricultural sustainability. Editors M. Ahmed, and C. O. Stockle Mukhtar (Springer International Publishing), 233–250.
Steinfeld, H., Gerber, P., Wassenaar, T., Castel, V., Rosales, M., Rosales, M., et al. (2006). Livestock's long shadow: Environmental issues and options. Rome: Food & Agriculture Org.
Treydte, A. C., Heitkönig, I. M., Prins, H. H., and Ludwig, F. (2007). Trees improve grass quality for herbivores in African savannas. Perspect. Plant Ecol. Evol. Syst. 8, 197–205. doi:10.1016/j.ppees.2007.03.001
Valladares, F., Laanisto, L., Niinemets, Ü., and Zavala, M. A. (2016). Shedding light on shade: Ecological perspectives of understorey plant life. Plant Ecol. divers. 9, 237–251. doi:10.1080/17550874.2016.1210262
Vira, B., Agarwal, B., Mansourian, S., and Wildburger, C. (2015). Introduction: Forests, trees and landscapes for food security and nutrition. Vienna: International Union of Forest Research Organizations.
Walker, B. H., and Noy-Meir, I. (1982). “Aspects of the stability and resilience of savanna ecosystems,” in Ecology of tropical savannas in Ecology of tropical savannas. Editors B. J. Huntley, and B. H. Walker (Springer Berlin, Heidelberg), 556–590.
Walker, B., and Salt, D. (2012). Resilience thinking: Sustaining ecosystems and people in a changing world. Washington: Island press.
Walter, H., and Burnett, J. H. (1971). Ecology of tropical and subtropical vegetation. Edinburgh: Oliver & Boyd.
Zellweger, F., Coomes, D., Lenoir, J., Depauw, L., Maes, S. L., Wulf, M., et al. (2019). Seasonal drivers of understorey temperature buffering in temperate deciduous forests across Europe. Glob. Ecol. Biogeogr. 28, 1774–1786. doi:10.1111/geb.12991
Keywords: facilitation, livestock, rangeland, silvo pastoral system, tree-grass interaction, tropics, temperate, tree shade
Citation: Hernández-Salmerón IR and Holmgren M (2022) Global meta-analysis: Sparse tree cover increases grass biomass in dry pastures. Front. Environ. Sci. 10:949185. doi: 10.3389/fenvs.2022.949185
Received: 20 May 2022; Accepted: 18 July 2022;
Published: 17 August 2022.
Edited by:
Karsten Wesche, Senckenberg Museum of Natural History Görlitz, GermanyReviewed by:
Juying Huang, Ningxia University, ChinaLilia L. Roa-Fuentes, Pontifical Javeriana University, Colombia
Copyright © 2022 Hernández-Salmerón and Holmgren. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ivan Raniero Hernández-Salmerón, aXZhbnJhbmllcm8uaHNAZ21haWwuY29t; Milena Holmgren, bWlsZW5hLmhvbG1ncmVuQHd1ci5ubA==