 Min Li
Min Li Huijie Xiao
Huijie Xiao Zhiming Xin2
Zhiming Xin2- 1School of Soil and Water Conservation, Beijing Forestry University, Beijing, China
- 2Inner Mongolia Dengkou Desert Ecosystem National Observation Research Station, Experimental Center of Desert Forestry, Chinese Academy of Forestry, Dengkou, China
- 3Department of Geosciences, The University of Tulsa, Tulsa, OK, United States
- 4Department of Watershed and Range Management, Faculty of Water and Soil, University of Zabol, Zabol, Iran
- 5Shandong Key Laboratory of Eco-Environmental Science for the Yellow River Delta, Binzhou University, Binzhou, China
Understanding soil seed banks (SSBs) of Nitraria tangutorum nebkhas is critical for vegetation restoration and ecological management in desert–oasis ecotones. In this study, we conducted a survey in the field and a seed germination experiment in the laboratory to investigate the characteristics of SSB in different sizes and parts of nebkhas and to examine their relationship with aboveground vegetation. The results showed that there were 17 species of plants in six families in the SSB of Nitraria tangutorum nebkhas in a desert–oasis ecotone, dominated by Chenopodiaceae. The life forms were mainly annual herbs, accounting for 58.82%–71.43%. The number of species in the SSB were in the order of large nebkha > medium nebkha > small nebkha, and there was no significant difference in SSB density. However, SSB densities in different parts of each nebkha were in the order bottom > middle > top. The densities were significantly different and showed a decreasing trend as soil depth increased. Most of the plant seeds were distributed in the 0–5 cm soil layer. The similarity coefficient between the SSB of each nebkha and the aboveground vegetation was high. From the bottom to the top, the number of common species and similarity coefficient between the aboveground vegetation and the SSB decreased, and the number of species in the SSB was greater than that of the aboveground vegetation. Vegetation plays a decisive role in the initial development of a nebkha. When the nebkha develops to a certain extent, it reacts to the vegetation, thereby intensifying the differences in the composition of vegetation species in different parts of the nebkha and further affecting the SSB.
1 Introduction
The term soil seed bank (SSB) refers to the sum of all viable seeds present on and in the soil (Roberts, 1981). As a potential population, it provides provenance for plant species renewal, community succession, and dispersal processes (Bakker et al., 1996; Zhang et al., 2021b). SSBs are fundamental to the process of plant population settlement, survival, reproduction, and diffusion. They play an important role in reshaping and maintaining biodiversity and can indicate the vegetation on the ground (Bakker et al., 1996; Peterson and Baldwin, 2004; Schwienbacher et al., 2010; He et al., 2018). For this reason, analyzing the regularity and succession of vegetation growth and development in SSBs is a critical component of vegetation reconstruction and restoration studies (Yang et al., 2010).
Nebkhas are aeolian landforms produced by sand deposited near to and obstructed by shrubs, and their development is mainly affected by plants, aeolian sand deposition, and hydrogeological conditions (Li and Ravi, 2018). In China, nebkhas are widely distributed in desert–oasis ecotones, desert steppe marginal zones, interlaced agricultural–pastoral zones, and other regions (Wang et al., 2006). Natural factors and human activities interfere with their formation and development to a large extent, and their appearance has a great impact on the ecological environment (Tengberg and Chen, 1998; Langford, 2000; Li and Ravi, 2018). A large number of Nitraria tangutorum thickets grow in the transition zone between the desert oases in the northeastern part of the Ulan Buh Desert, and nebkhas of different sizes are formed by the annual accumulation of sand. These form a natural “sand prevention wall” around the desert oasis, effectively preventing quicksand from invading the ecological environment of the oasis and playing a positive role in the stability of its environment (Dang et al., 2019). As important sand-fixing vegetation in the transition zone of desert oases, N. tangutorum shrub communities, however, have gradually declined, and some parts have even been activated and disintegrated (Xu et al., 2008a). This has greatly weakened the ecological function of the community, increased the vulnerability of the oasis, and jeopardized associated industrial and agricultural production of the oasis (Jia et al., 2002).
There has been considerable research into the development and evolution of nebkhas: morphological changes, soil physicochemical properties, vegetation biomass predictions, reasons for vegetation degradation, restoration techniques, conservation measures, and wind erosion (Zhang et al., 2021a; Abdurimithi and Yusuf, 2016; Du et al., 2010; Li et al., 2010). However, a majority of these studies have ignored the ecological function of nebkhas. Additionally, studies of species composition and size, species diversity characteristics, spatial distribution characteristics, and plant succession of the underground SSB of potential plant provenance on the horizontal scale of N. tangutorum nebkhas are also limited. Here, we examined a suite of N. tangutorum nebkhas with different sizes in the field and conducted seed germination experiments in the laboratory to analyze the characteristics of SSB and their relationship with aboveground vegetation. The goal of this study is to reveal the relationship between SSBs and the establishment, evolution, and degradation of nebkhas in a desert–oasis ecotone, which is critical for vegetation restoration and land degradation prevention in desert areas.
2 Materials and Methods
2.1 Study Area
The study area was located in the northeastern part of the Ulan Buh Desert (40°18′–40°30′, 106°34′–106°58′ E), Inner Mongolia, northern China, which is a transition zone from steppe desert to desert steppe. It has a temperate continental monsoon climate, with an annual average temperature of 7.6°C and annual precipitation of 100–145 mm. About 80% of the annual precipitation occurs from July to September. Strong northwestern winds are common in spring and winter seasons. In the study area, brown calcium and gray desert soils with quicksand are common. Native vegetation in the study area is dominated by communities of N. tangutorum, which exist in the form of nebkhas. The height of nebkhas ranges from approximately 1 to 3 m. The nebkhas are moderately dynamic and advance from west to east at a speed of 3–5 m every year. The nebkhas are characterized by mixed species with three to eight species per 1 m2. The plant communities have a height of 34–74 cm and cover 25–80% of the dune surface. A total of 22 species of plants were identified in the study area, belonging to six families and 18 genera. More information about vegetation species composition in the study area is listed in Table 1.
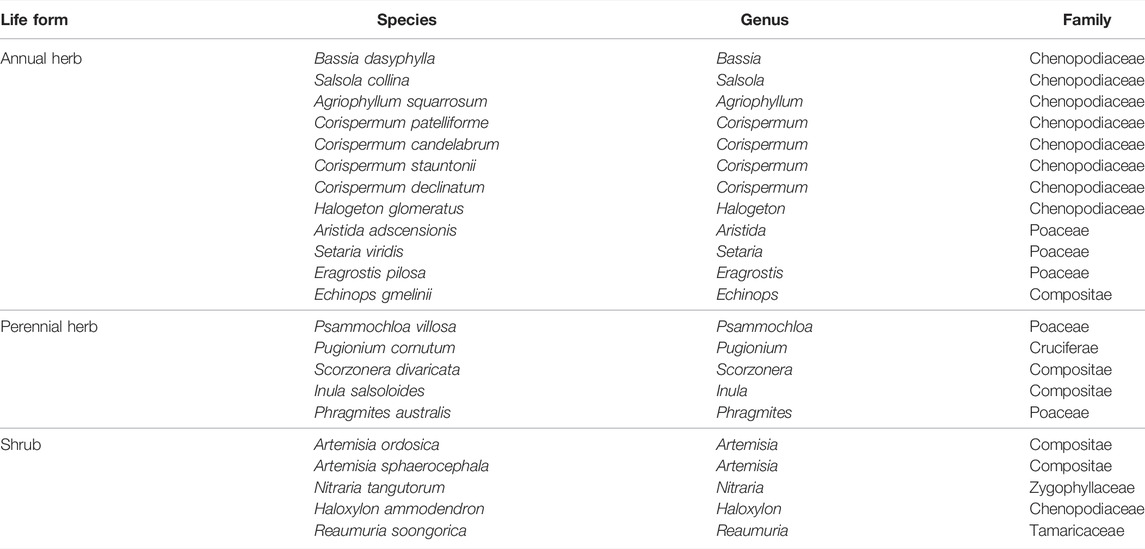
TABLE 1. Vegetation species composition in the study area.
2.2 Methods
2.2.1 Experimental Setup and Vegetation Survey
In April 2021, a continuous distribution of N. tangutorum in the northeastern part of the Ulan Buh Desert, covering an area of 1 km2, was selected as the experimental quadrat. We randomly surveyed 100 N. tangutorum nebkhas in the quadrat. The nebkhas selected were classified based on the height of the nebkhas into large (height >1.5 m), medium (1 m < height <1.5 m), and small (height <1 m). We also selected nine similar typical nebkhas from each of the three types for a vegetation survey and SSB sampling, which yielded a total of 27 nebkhas surveyed (Table 2).
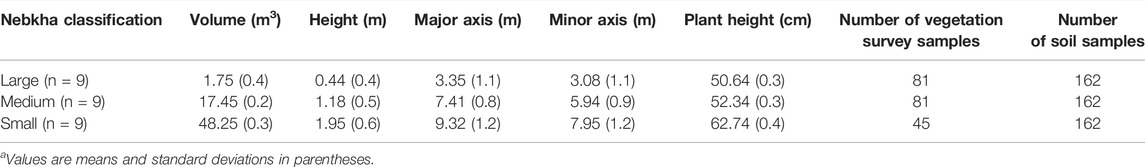
TABLE 2. Characteristics of the nebkhas measured in this studya.
The height, volume, long axis, short axis, slope, vegetation coverage, and vegetation height of these nebkhas were measured. Nine 50 cm × 50 cm sampling points were set up at the bottom, middle, and top of each nebkha in the four directions (east, west, south, and north), and the composition and quantity of aboveground plant species were investigated. Among them, vegetation survey was not carried out in the middle of the small nebkha (Figure 1; Table 2).
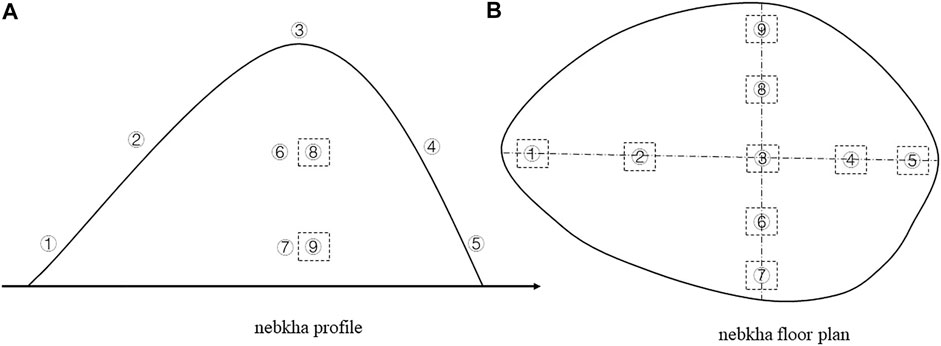
FIGURE 1. Schematic diagram of sampling sites in Nitraria tangutorum nebkhas. ① West bottom, ② West middle, ③ Top, ④ East middle, ⑤ East bottom, ⑥ South middle, ⑦ South bottom, ⑧ North middle, ⑨ North bottom.
2.2.2 Soil Sampling and Greenhouse Experiment
A metal soil sampler (8 cm in diameter) was used to collect soil samples at depths of 0–10 cm (divided into two layers: 0–5 cm, 5–10 cm) in the center of the bottom, middle, and top of each nebkha. A total of 486 soil samples were collected and brought to the laboratory for further analysis.
The seed bank density and species composition were determined by the indoor seed germination method. The germination test was carried out at the Inner Mongolia Dengkou Desert Ecosystem National Observation Research Station in May 2021. Soil samples for the seed bank determination were placed on 30 cm × 20 cm sample trays at room temperature. Non-soil components were removed (e.g., residual roots and animal and plant debris), and sufficient water was added to keep the soil moisture. The types and number of seeds germinating in each soil sample were recorded daily. After the species were identified, we moved the germinated but unidentified seedlings to pots and placed them at a certain depth until they were identified and removed, as their root systems were too long to fit in the sample tray. This avoided distortion of the accuracy of the experimental results compared with the seedlings of desert vegetation. The germination culture continued for 2 months (Table 2).
2.3 Data Analysis
In this study, the density of an SSB was expressed by the number of viable seeds per unit area of soil, obtained by converting the seeds in the sampling area into the number of 1 m2. The counted plants were divided into four types: annual herbs, perennial herbs, shrubs, and trees, and the percentage of each type of species in the total number of species was calculated separately. The Margalef richness index, Simpson diversity index, Shannon–Wiener diversity index, and Pielou evenness index were used to describe the characteristics of species diversity (Jia et al., 2017), and the Jaccard similarity coefficient was used to characterize the similarity of species (Jaccard, 1908) as follows:
Margalef richness index:
Simpson diversity index:
Shannon–Wiener diversity index:
Pielou evenness index:
Jaccard similarity coefficient:
Here, S is the total number of species in the aboveground vegetation or SSB; N is the number of plants or SSBs of the total species; Pi is the ratio of the number of SSBs or plants of the ith species to the total number of all species; a is the total number of species in SSB; b is the total number of species in aboveground vegetation; and j is the number of species common to SSB and aboveground vegetation.
Soil seed bank density at different locations of the nebkhas was analyzed using the one-way analysis of variance (ANOVA) with a significant level at 0.05. Finally, correlation analyses were performed to reveal the relationship between the characteristics of nebkha, plant species, and SSB.
3 Results
3.1 Species Composition and Floristic Compositions of Soil Seed Banks
A total of 17 plant species in six families were identified through the seed germination test. Among them, the family of Chenopodiaceae comprised the largest proportion, with seven species accounting for 41.18% of the total species. There were five species in the family of Compositae, accounting for 29.14% of the total species. Species in the families of Poaceae and Cruciferae account for 11.77% and 5.88% of the total species, respectively. It should be noted that the study area is not suitable for trees. Ulmus pumila seeds are likely carried by the wind in a farm shelter 3 km away from the study area.
There were 17, 16, and 12 species of plants at the bottom, middle, and top of the large nebkhas, respectively. There were 16, 16, and 13 species of plants at the bottom, middle, and top of the medium nebkhas, respectively. For small nebkhas, we found 14, 14, and 15 species of plants at the bottom, middle, and top, respectively. It was clear that the larger the nebkha, the fewer the SSB species at the top. The SSB density of companion species such as Bassia dasyphylla, Salsola collina, and Agriophyllum squarrosum was relatively large. In addition, the seeds of Halogeton glomeratus, Pugionium cornutum, Psammochloa villosa, and Scorzonera divaricata were only present at the bottom and in the middle of the nebkhas.
We did not find notable difference of life forms in the SSBs of different nebkhas (Figure 2). Irrespective of the nebkha size, the proportion of annual herbs was the highest (58.82%–71.43%), followed by shrubs (17.65%–23.08%), perennial herbs (0%–17.65%), and trees (0%–5.88%). Similar to nebkhas with different sizes, SSBs in nebkhas with the same size category but different location are also dominated by annual herbs, and the proportion generally increased from the bottom to the top of the dunes, except for the small nebkhas.
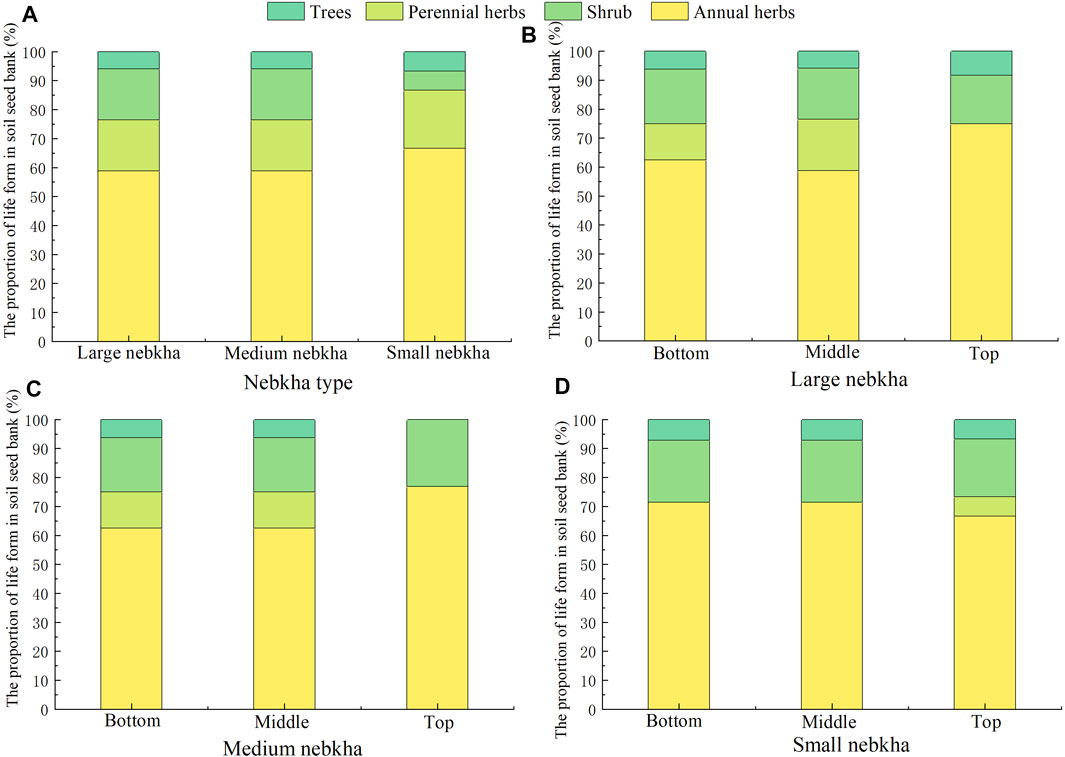
FIGURE 2. Life forms of soil seed banks in Nitraria tangutorum nebkhas. (A) Comparison of life forms of soil seed banks in large, medium and small nebkha. (B–D) Comparison of the life forms of soil seed banks at the top, middle and bottom of the large, medium and small nebkha.
3.2 Spatial Distribution of Soil Seed Bank
3.2.1 Horizontal Distribution Characteristics
The SSB density at the bottom of large and medium nebkha sites was higher than that at the top (p < 0.05). The SSB density between the middle and top locations of the nebkhas was not statistically significant, regardless the size of the nebkhas. We did not observe consistent patterns of SSB density for the same relative locations of the nebkhas with various sizes (Figure 3).
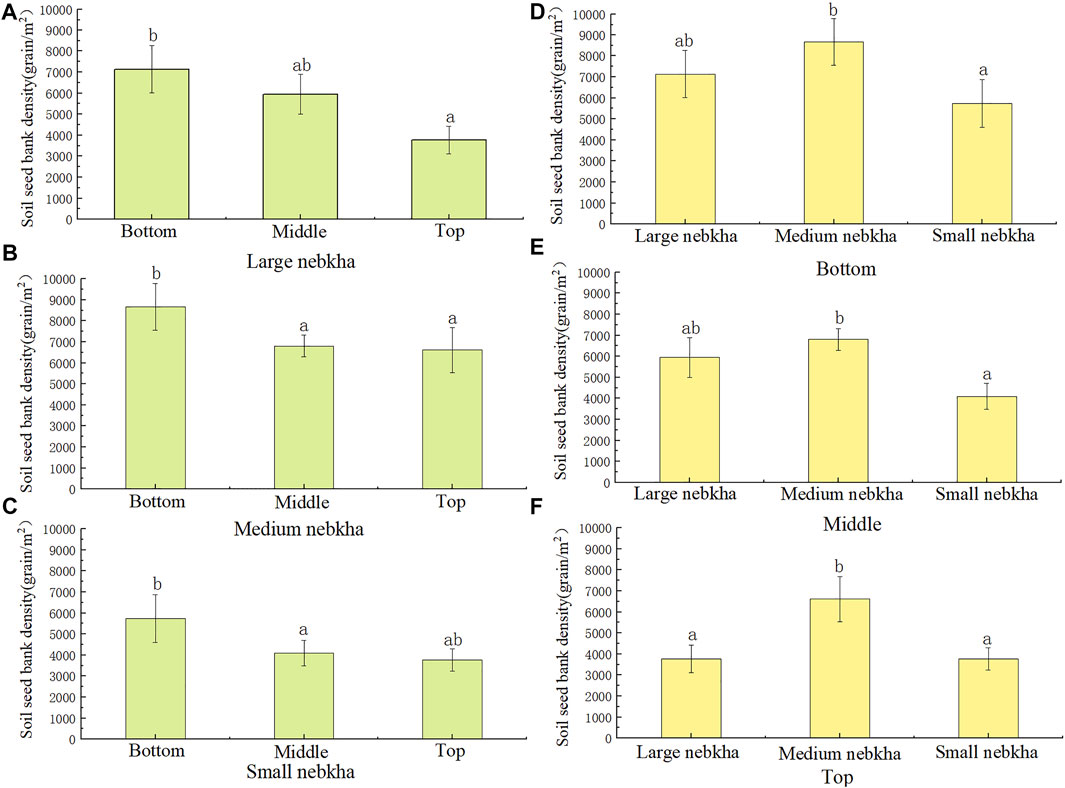
FIGURE 3. Soil seed bank density (grain/m2) in Nitraria tangutorum nebkhas. The letters in the figure indicate significant differences (p < 0.05). (A–C) Comparison of soil seed bank density in different parts of the same size nebkha. (D–F) Comparison of soil seed bank density of different nebkhas in the same place.
3.2.2 Vertical Distribution Characteristics
The number of species in the top 5 cm of the soil profile fell in the range of 6–14, with a general trend that bottom parts of the nebkhas have a higher number of species than those of the top ones (Table 3). A similar pattern was found for the SSB at a deeper soil profile (5–10 cm), with the largest number of species of 16 found at the bottom of the large nebkhas and the smallest number of species of seven found at the top of both large and small nebkhas.
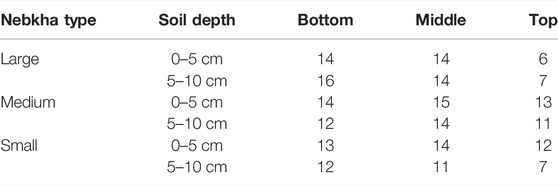
TABLE 3. Vertical distribution of soil seed banks (number of species) in Nitraria tangutorum nebkhas.
Most plant seeds were distributed in the 0–5 cm soil layer (Figure 4). The average SSB density in the 0–5 cm and 5–10 cm soil layers was medium nebkha > large nebkha > small nebkha, and the difference between the three types of nebkha was not significant (p > 0.05). In the range of 0–10 cm soil layer, the SSB density at the bottom of large, medium, and small nebkha sites was higher than that at the top (p < 0.05). We did not detect a consistent pattern of SSB density between the top 5 cm and 5–10 cm of the soil profiles, regardless of the size of the nebkhas.
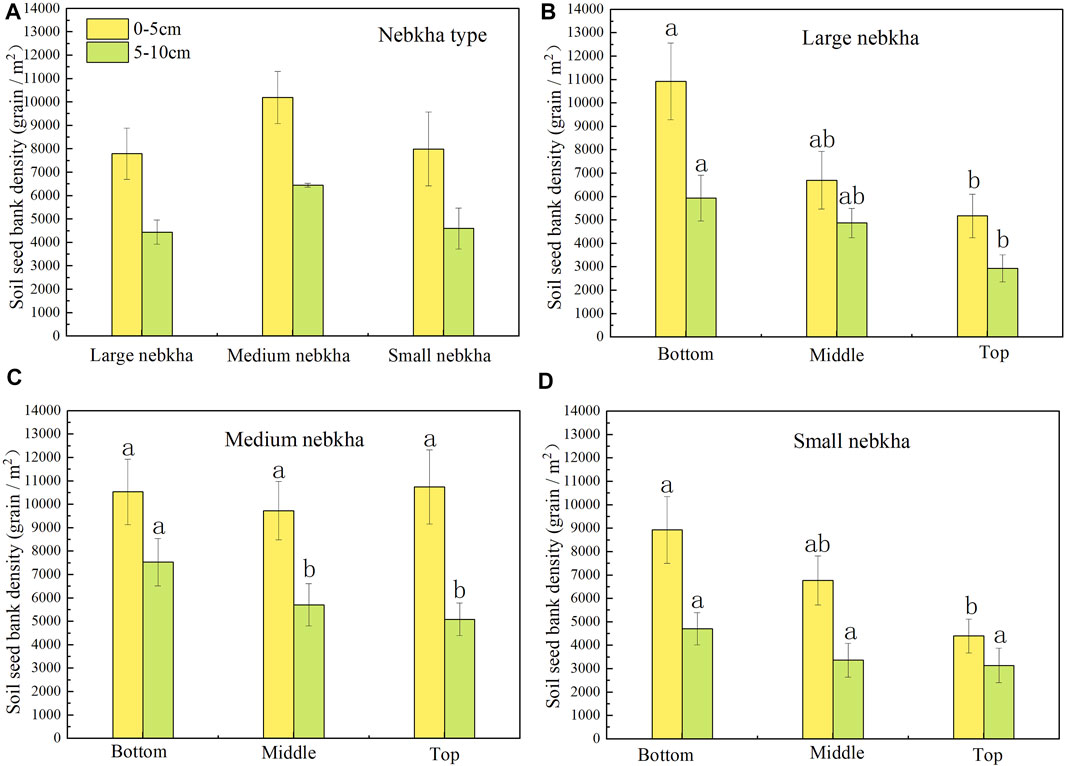
FIGURE 4. Soil seed bank density at different soil depths. The letters in the figure indicate significant differences (p < 0.05). (A) Comparison of soil seed bank density in large, medium and small nebkha. (B–D) Comparison of soil seed bank density at the top, middle and bottom of the large, medium and small nebkha at different soil depths.
3.3 Relationship Between Soil Seed Bank and Aboveground Vegetation
The survey found 14 species of aboveground vegetation in N. tangutorum nebkhas, belonging to 5 families and 12 genera. Among them, Chenopodiaceae accounted for the largest proportion with six species, accounting for 42.86% of the total species. In addition, Compositae (four species), Poaceae (two species), Cruciferae (one species), and Tribulus (one species) accounted for 28.57%, 14.29%, 7.14%, and 7.14% of the total species, respectively. In general, SSBs in nebkhas are similar to the number of aboveground vegetation species (Table 4).
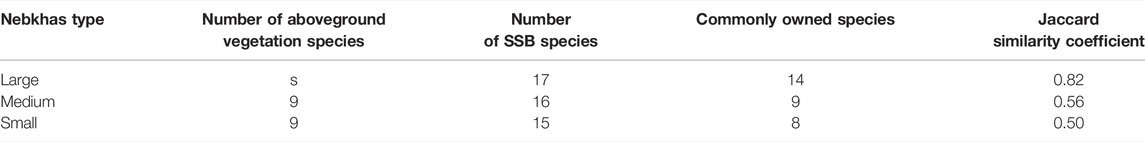
TABLE 4. Similarity of soil seed bank (SSB) and aboveground vegetation in Nitraria tangutorum nebkhas.
The vegetation survey also showed that at the bottom of the nebkhas, the number of aboveground vegetation species is the highest, followed by middle and then top, regardless of the size of the nebkhas (Table 5). A similar pattern was found for the commonly owned species at different locations of the nebkhas. Overall, we found some strong similarities for the bottom of the nebkhas for all the nebkhas we measured.
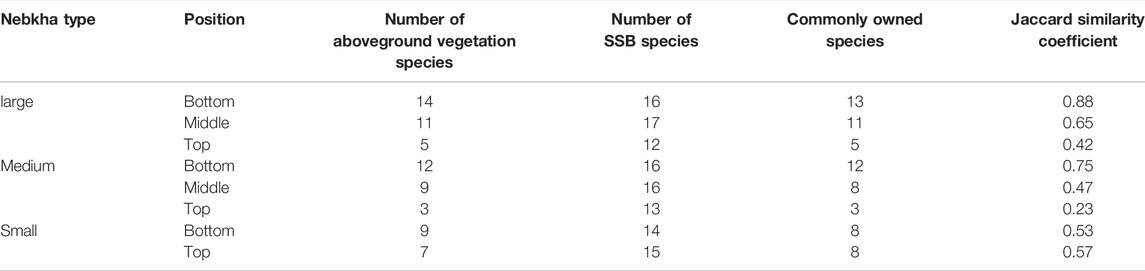
TABLE 5. Similarity between soil seed bank (SSB) and aboveground vegetation in different parts of nebkhas.
In all parts of the nebkhas, the number of species in the SSB was more than that of the aboveground vegetation. N. tangutorum, Bassia dasyphylla, and Salsola collina appeared both on the ground and in the soil. Some species appeared only in the SSB and did not appear in the aboveground vegetation, such as Corispermum patelliforme, C. candelabrum, C. stauntonii, Aristida adscensionis, Pugionium cornutum, Psammochloa villosa, Scorzonera divaricate, and Artemisia ordosica (Table 5). The Jaccard similarity coefficients between the SSB and the aboveground vegetation species of different types of nebkha were 0.82 (large nebkha) > 0.56 (medium nebkha) > 0.50 (small nebkha) (Table 4). In addition, the difference of the Jaccard similarity coefficient between different parts of large nebkha and medium nebkha was obvious (range: 0.23–0.52), while the difference between the top and bottom of small nebkha was only 0.038. From the bottom to the top of the nebkhas, the number of common species and similarity between the aboveground vegetation and the SSB showed a decreasing trend (Table 5).
3.4 Influence of Morphological Characteristics of N. tangutorum Nebkhas on Vegetation and Soil Seed Bank
This study used the Pearson method to analyze the morphological parameters, vegetation, and SSB parameters of 27 N. tangutorum nebkhas (Table 6). There were significant differences in the volume, height, long axis, and short axis of different nebkhas (p < 0.05). There were different degrees of correlation between the morphological parameters of N. tangutorum nebkhas and vegetation parameters. The species number in the SSB at the top of the nebkhas was significantly negatively correlated with the volume, height, long axis, short axis, and slope of the nebkhas (p < 0.01). There was a significant positive correlation between the vegetation coverage of a nebkha and its volume, long-axis slope, and short-axis slope (p < 0.05). The number of vegetation species of a nebkha had a strong and significant positive correlation with the nebkha volume, height, long axis, short axis, long-axis slope, and plant height (p < 0.05). It was shown that the morphological parameters of N. tangutorum nebkhas affected the plant height, the number of species in the nebkha vegetation, and the number of species in the SSB at the top of a nebkha. Vegetation plays a decisive role in the initial development of a nebkha. When the nebkha develops to a certain extent, it will react to the vegetation, thereby intensifying the differences in the composition of vegetation species in its different parts and further affecting the SSB.
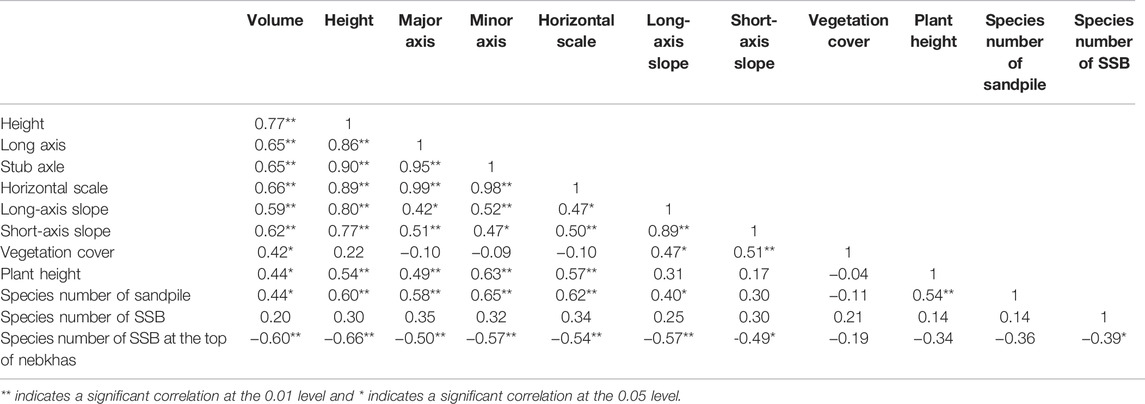
TABLE 6. Correlation analysis of nebkha, vegetation, and soil seed bank (SSB) parameters in Nitraria tangutorum nebkhas.
4 Discussion
We found that annual herbaceous plants, primarily Bassia dasyphylla, Salsola collina, Agriophyllum squarrosum, and Aristida adscensionis, accounted for the largest proportion of species on nebkhas in a desert–oasis ecotone in northwestern China. This finding was consistent with the studies of SSBs in the Alxa Desert region (Zeng et al., 2003), the central desert of the Hexi Corridor (Lu et al., 2019), and the Gurbantunggut Desert (Zhang and Li, 2018), which also reported that the species life forms were mainly annual herbs. This is not surprising as annuals have strong drought tolerance and larger niche breadth (Jia et al., 2017). Additionally, the rainfall in our study area was low and mostly concentrated in July–September, allowing most annual herbs to complete their entire life cycle during this period and to produce a large number of seeds.
In this study, the SSB density is relatively high and is consistent with studies conducted in a South American warm desert (Luis et al., 1998) and a desert–oasis marginal zone in middle reaches of Heihe River, China (Li and Fang, 2008). The density of SSB plants is different from the SSB density observed in other arid and extremely arid deserts (Yu et al., 2015; Zhang and Li, 2018; Hadinezhad, et al., 2021). This difference may be caused by the fact that nebkhas, with various shrub species, increase buried seeds in soil by directly trapping seeds or by indirect mechanisms through an intermediary animal or plant species (Giladi et al., 2013). More importantly, abiotic factors such as wind regime, landform, and soil condition also affected SSB density (Chambers and MacMahon, 1994). There were also significant differences in the spatial distribution patterns of SSB in different nebkha sites because of the strong habitat heterogeneity in the study area. Previous studies have shown that at the bottom of nebkhas, the light intensity is relatively weak, the soil water storage capacity is relatively strong, and thus soil moisture and nutrient conditions are better, which favor seed germination, survival, and establishment of plants (Xu et al., 2008b; Liu et al., 2011; Wang, 2021). However, compared with the middle and bottom of nebkhas, the light intensity is higher at the top, and the plants cannot obtain the soil moisture and nutrients needed for their growth, so there are relatively few species. Likewise, in the southern margin of the Gurbantunggut Desert, the density of each dune slope was found to decrease in sequence at the bottom, lower middle, upper middle, and top (Jia et al., 2017). Plant seeds generally fall on the soil surface or enter the surface layer after they mature, and only a small proportion enter deeper soil layers as a result of wind erosion and sand burial (Yang et al., 2016; Liu et al., 2021).
The study area is located in a windy desert area, and a majority of the windy conditions occur in spring (Cui et al., 2010; Gao et al., 2014). When the wind passes through the nebkhas, areas of low wind speed form in the lower-middle parts of the nebkhas because of the influence of the shrubs on the wind–sand flow. When the wind speed decreases, the ability of the wind–sand flow to carry seeds decreases, so that more seeds will accumulate in the lower-middle parts (Qian et al., 2016; Ma et al., 2021). Consequently, at the bottom of nebkhas, the SSB density in the surface layer is relatively high. Due to the fluctuation of nebkha height, soil moisture, light radiation, soil nutrients, and sand buried by wind erosion would change slightly in small local areas (Jia et al., 2021); at the bottom of nebkhas, wind erosion was not too severe. More plant litter accumulates, which reduces the loss of soil water and nutrients (Shao et al., 2001; Haas, 2004), and the increase in soil fertility improves vegetation growth, resulting in an increase in the number of species in aboveground vegetation (Zhang et al., 2006; Hadinezhad et al., 2021).
Previous studies have shown that in desert regions, more than one-half of all species were found in SSB as well as in the aboveground vegetation (Ma, et al., 2015; Teng et al., 2020; Lv et al., 2021). In this study, 14 species appeared both in the aboveground vegetation and in the SSB, three species appeared only in the SSB, and no species appeared only in the aboveground vegetation. These observations were similar to those reported in a previous study (Teng et al., 2020). The main reason for this was that the study area was located in the transition zone of desert and oasis and there were few plant species, which increased the probability of the same species appearing both in SSB and in aboveground vegetation in the different parts of the nebkhas. Bassia dasyphylla, Salsola collina, and N. tangutorum appeared in the aboveground vegetation and SSB. Thus, B. dasyphylla and S. collina could be the dominant species on nebkhas because their reproductive ability was very strong. In addition to seed germination, the main propagation method of N. tangutorum, as a constructive species in this area, is to bury its own branches by intercepting sand by itself and carrying out clonal propagation with branches or root tillers under suitable water and heat conditions.
The succession process of N. tangutorum nebkhas is strongly related to the process of plant growth, development, and change. With sufficient sand supply under the same wind conditions, vegetation with different ecological characteristics creates different wind speed and wind flow fields, thus developing different forms of nebkha (Yin et al., 2017). Plant morphology, quantity, structure, and litter affect the development of nebkhas through their impact on the environment and ability to block sandstorms (Miao et al., 2020; Han et al., 2021). Some studies have found that the changes in nebkha morphology are determined by the coverage, distribution, abundance, and height of plants (Hesp and McLachlan, 2000; Hesp, 2002). A study of nebkha morphology in Ulan Buh Desert by Gao et al. (2015) found that crown size and plant height affected the development of nebkhas. The nebkha slope may also affect the distribution of nebkha SSB. Seeds on a steep slope are more likely to slide to the bottom of the slope under the action of wind and rainfall (Pa et al., 2016; Li et al., 2018). Our results showed that the morphological characteristics of N. tangutorum nebkhas can affect the number of aboveground vegetation and SSB species and the vegetation height. During the development of N. tangutorum nebkhas, the growth and development of vegetation can inversely affect nebkha morphology. This reflects the decisive role that vegetation plays in the initial development of nebkhas. When they develop to a certain extent they react to the vegetation, thus intensifying the difference in the vegetation species composition in different parts of the nebkhas and further affecting the SSB.
5 Conclusion
We recorded 17 species of plants in six families in the SSB of N. tangutorum nebkhas in a desert–oasis ecotone, dominated by Chenopodiaceae. The life forms were mainly annual herbs, accounting for 58.82%–71.43%. The number of species in the SSB followed the sequence large nebkha > medium nebkha > small nebkha, and there was no significant difference in SSB density. However, the SSB densities in different parts of each nebkha were in the sequence bottom > middle > top. The SSB densities were significantly different and showed a decreasing trend with the increase in soil depth. In the range of 0–10 cm soil layer, most of the plant seeds were distributed in the 0–5 cm layer, which decreased with the increase of soil depth. The similarity coefficient between the SSB of each nebkha and the aboveground vegetation was high. From the bottom to the top, the number of common species and similarity coefficient between the aboveground vegetation and the SSB decreased, and the number of species in the SSB was more than that of the aboveground vegetation. Vegetation plays a decisive role in the initial development of a nebkha. When a nebkha develops to a certain extent, it reacts to the vegetation, thereby intensifying the differences in the composition of vegetation species in its different parts and further affecting the SSB.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material; further inquiries can be directed to the corresponding author.
Author Contributions
ML, HX, and ZX contributed to conception and design of the study. ML wrote the original draft. HX wrote sections of the manuscript. ML, HX, JL, AM, XL, and QC contributed to the revision of the manuscript. All authors contributed to manuscript revision and read and approved the submitted version.
Funding
This study was funded by the Project of Intergovernmental International Cooperation in Science and Technology Innovation (No. 2019YFE0116500).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful for the support from the Inner Mongolia Dengkou Desert Ecosystem National Observation Research Station, Dengkou, China. We also thank the journal editors and reviewers for their comments and efforts on this article.
References
Abdurimithi, A., and Yusuf, B. (2016). Morphological Characteristics of White Thorn Scrub Sandpiles in Different Habitats in the Yanqi Basin. Chin. Agric. Sci. Bull. 32 (32), 117–123. doi:10.11924/j.issn.1000-6850.casb16030228
Bakker, J. P., Bakker, E. S., Rosén, E., Verweij, G. L., and Bekker, R. M. (1996). Soil Seed Bank Composition along a Gradient from Dry Alvar Grassland toJuniperusshrubland. J. Veg. Sci. 7, 165–176. doi:10.2307/3236316
Chambers, J. C., and MacMahon, J. A. (1994). A Day in the Life of a Seed: Movements and Fates of Seeds and Their Implications for Natural and Managed Systems. Annu. Rev. Ecol. Syst. 25 (1), 263–292. doi:10.1146/annurev.es.25.110194.001403
Cui, Y., Wang, X., Pan, Y. X., Wang, Z., and Qi, P. (2010). Differences in Soil Seed Bank in Different Microhabitats of Natural Fixed Sandy Areas. Acta Ecol. Sin. 30 (08), 1981–1989.
Dang, X., Xia, P., Yong, G., Yang, L., Zhenyi, W., and Zhongju, M. (2019). Spatial Heterogeneity of Wind-Eroded Soil Particles Around Nitraria Tangutorum Nebkhas in the Ulan Buh Desert. Écoscience 26 (4), 347–358. doi:10.1080/11956860.2019.1646064
Du, J., Yan, P., and Dong, Y. (2010). Current Status and Perspectives of Research on Scrub Sandpiles in Arid Regions. Acta Geogr. Sin. 65 (03), 339–350. doi:10.11821/xb201003009
Gao, J., Xin, Z., Liu, F., Yuan, W. J., Feng, W., Xu, J., et al. (2014). Analysis of Atmospheric Dust Fall Characteristics and Influencing Factors in Northeastern Ulaanbaatar Desert. J. Arid. Land Res. Environ. 28 (08), 145–150. doi:10.13448/j.cnki.jalre.2014.08.014
Gao, Y., Dang, X., Yu, Y., Wang, J., Wang, S., Yuan, W., et al. (2015). Nabkha Morphological Characteristics and Sand Fixing Capacity of Artemisia sphaerocphala in the Southeastern Edge of the Ulan Buh Desert. J. Desert Res. 35 (01), 1–7. doi:10.7522/j.issn.1000-694X.2014.00080
Giladi, I., Segoli, M., and Ungar, E. D. (2013). Shrubs and Herbaceous Seed Flow in a Semi-arid Landscape: Dual Functioning of Shrubs as Trap and Barrier. J. Ecol. 101 (1), 97–106. doi:10.1111/1365-2745.12019
Ha, S. (2004). Preliminary Study on the Structural Variation of Wind and Sand Flow on the Surface of Sand Dunes at the Southeast Edge of Tengger Desert. Chin. Sci. Bull. 49 (11), 1099–1104. doi:10.3321/j.issn:0023-074X.2004.11.015
Hadinezhad, M., Erfanzadeh, R., and Ghelichnia, H. (2021). Soil Seed Bank Characteristics in Relation to Different Shrub Species in Semiarid Regions. Land Degrad. Dev. 32 (5), 2025–2036. doi:10.1002/ldr.3856
Han, D., Zhang, W., Nurmati, Y., and Yang, Y. (2021). Complementary Constraints on the Renewal of Plant Populations. Chin. J. Plant Ecol. 45 (01), 1–12. doi:10.17521/cjpe.2020.0246
He, F. L., Guo, C., Ma, J. M., Wu, H., and Jin, H. X. (2018). Dynamic Changes in Soil Seed Banks and Their Relationships with Aboveground Vegetation during the Decaying of Haloxylon Ammodendron Plantations at the Edge of the Minqin Oasis. Acta Ecol. Sin. 38 (13), 4657–4667. doi:10.5846/stxb201706141084
Hesp, P. (2002). Foredunes and Blowouts: Initiation, Geomorphology and Dynamics. Geomorphology 48, 245–268. doi:10.1016/s0169-555x(02)00184-8
Hesp, P., and McLachlan, A. (2000). Morphology, Dynamics, Ecology and Fauna of Arctotheca Populifolia and Gazania Rigens Nabkha Dunes. J. Arid Environ. 44, 155–172. doi:10.1006/jare.1999.0590
Jaccard, P. (1908). Nouvelles recherches sur la distribution fiorale. Bull. Soc. Vaud. Sci. Nat. 44, 223–270. doi:10.5169/seals-268384
Jia, B., Ci, L., Cai, T., Gao, Z., and Ding, F. (2002). Preliminary Study on the Characteristics of Soil Moisture Changes in the Oasis-Desert Interlacing Zone. Chin. J. Plant Ecol. 26 (02), 203–208. doi:10.3321/j.issn:1005-264X.2002.02.011
Jia, F., Tiyip, T., Wu, N., Tian, C., and Zhang, Y. (2017). Characteristics of Soil Seed Banks at Different Geomorphic Positions within the Longitudinal Sand Dunes of the Gurbantunggut Desert, China. J. Arid. Land 9 (3), 355–367. doi:10.1007/s40333-017-0055-x
Jia, F., Bai, Y., and Ren, J. (2021). A Review of Studies on the Seed Bank of Sand Dune Soils and its Relationship with Above-Ground Vegetation. J. Ili Agric. Univ. Nat. Sci. 15 (01), 37–44. doi:10.3969/j.issn.1673-999X.2021.01.008
Langford, R. P. (2000). Nabkha (Coppice Dune) Fields of South-Central New Mexico, U.S.A. J. Arid Environ. 46 (9), 25–41. doi:10.1006/jare.2000.0650
Li, H. R., Wang, J., Jiang, Z., and Ji, Y. F. (2010). Piercing White Sandbags Development Process of Soil Water and the Relationship Between Root Biomass. J. Gansu Agri. Univ. 45 (6), 133–138. doi:10.13432/j.cnki.jgsau.2010.06.002
Li, J., and Ravi, S. (2018). Interactions Among Hydrological-Aeolian Processes and Vegetation Determine Grain-Size Distribution of Sediments in a Semi-arid Coppice Dune (Nebkha) System. J. Arid Environ. 154, 24–33. doi:10.1016/j.jaridenv.2018.03.011
Li, Q., and Fang, H., Y. (2008). Study on Soil Seed Bank of Nitraria Sphaerocarpa Coppice Dune in a Desert Oasis Marginal Zone. Arid. Zone Res. 4, 502–506. doi:10.13866/j.azr.2008.04.019
Li, L., Fang, Z., Aiqbayr, M., Zhou, L., and Lu, B. (2018). Study on Soil Seed Bank and Seedling Regeneration of Different Populations of Phellodendron Chinense in Wild Fruit Forests in the Western Tianshan Mountains. Plant Sci. J. 36 (04), 534–540. doi:10.11913/PSJ.2095-0837.2018.40534
Liu, H.-L., Shi, X., Wang, J.-C., Yin, L.-K., Huang, Z.-Y., and Zhang, D.-Y. (2011). Effects of Sand Burial, Soil Water Content and Distribution Pattern of Seeds in Sand on Seed Germination and Seedling Survival of Eremosparton Songoricum (Fabaceae), a Rare Species Inhabiting the Moving Sand Dunes of the Gurbantunggut Desert of China. Plant Soil 345 (1-2), 69–87. doi:10.1007/s11104-011-0761-7
Liu, S. Q., Du, X., Li, Q., Zhang, Z., Wei, Y. C., and Ai, F. (2021). Characterization of the Seed Bank of Salix Community in the Southeastern Margin of Mu Us Sandy Land. J. Northwest Fore. Univ. 36 (06), 140–144. doi:10.3969/j.issn.1001-7461.2021.06.20
Lu, Y. F., Ma, L., Zhan, Y. F., Qian, W. J., Zhen, W. L., and Teng, Y. F. (2019). A Review of the Soil Seed Bank of Artificial Desert Vegetation in the Hexi Corridor. Fore. Sci. Technol. 60 (11), 64–66. doi:10.13456/j.cnki.lykt.2018.08.08.0001
Luis, M., Bertilde, E. R., and Manuel, E. H. (1998). Timing and Spatial Patterning of Seed Dispersal and Redistribution in a South American Warm Desert. Plant Ecol. 137 (2), 143–150. doi:10.2307/20050657
Lv, Y., Shen, M., Meng, B., Zhang, H., Sun, Y., Zhang, J., et al. (2021). The Similarity between Species Composition of Vegetation and Soil Seed Bank of Grasslands in Inner Mongolia, China: Implications for the Asymmetric Response to Precipitation. Plants 10 (9), 1890. doi:10.3390/PLANTS10091890
Ma, Q., Lu, Q., Wei, L., and Jin, H. (2015). Varying Characteristics of Soil Seed Banks during the Succession Process ofNitraria Tangutorumvegetation in an Arid Desert Area. Acta Eco Sin. 35 (7), 326. doi:10.5846/stxb201306041326
Ma, R., Zhao, J. M., Ma, Y., and Lu, J. (2021). Morphological Characteristics and Spatial Distribution of Nitraria Tangutorum Nebkhas Near Shifting Sand of Southern Margin of Badain Jaran Desert. J. Soil Water Conserv. 35 (04), 217–221+236. doi:10.13870/j.cnki.stbcxb.2021.04.030
Miao, B. L., Pei, H., Jiang, Y. F., Jia, C. Z., and Wu, H. (2020). Classification of Sand Cover Surface Morphology and Morphological Characteristics and Developmental Processes. Meteor. J. Inn. Mong. 29 (05), 37–42. doi:10.14174/j.cnki.nmqx.2020.05.008
Pa, R., Dong, X. L., Zhang, Y. L., and Mou, C. (2016). Characteristics of Soil Seed Bank and its Effect on Seed Germination in Staphylinia Variegata in Xinjiang. J. Xinjiang Agric. Univ. 39 (06), 458–462. doi:10.3969/j.issn.1007-8614.2016.06.006
Peterson, J. E., and Baldwin, A. H. (2004). Seedling Emergence from Seed Banks of Tidal Freshwater Wetlands: Response to Inundation and Sedimentation. Aquat. Bot. 78 (3), 243–254. doi:10.1016/j.aquabot.2003.10.005
Qian, J., Liu, Z., Hatier, J.-H. B., and Liu, B. (2016). The Vertical Distribution of Soil Seed Bank and its Restoration Implication in an Active Sand Dune of Northeastern Inner Mongolia, China. Land Degrad. Dev. 27 (2), 305–315. doi:10.1002/ldr.2428
Schwienbacher, E., Marcante, S., and Erschbamer, B. (2010). Alpine Species Seed Longevity in the Soil in Relation to Seed Size and Shape - A 5-year Burial Experiment in the Central Alps. Flora - Morphol. Distribution, Funct. Ecol. Plants 205 (1), 19–25. doi:10.1016/j.flora.2008.10.007
Shao, Y. Q., Zhao, J., and Bao, Q. H. (2001). Study on the Vertical Distribution of Microbial Biomass in Soil of Kubuqi Fixed Dune. J. Desert Res. 21 (01), 91–95. doi:10.3321/j.issn:1000-694X.2001.01.018
Teng, Y. F., Zhan, Y. F., Ma, L., Lu, Y. F., Qian, W. J., and Tian, X. P. (2020). Characteristics of Soil Seed Bank of Different Artificial Vegetation Types in Jinta Desert. Bull. Soil Water Conserv. 40 (03), 163–169. doi:10.13961/j.cnki.stbctb.2020.03.023
Tengberg, A., and Chen, D. (1998). A Comparative Analysis of Nebkhas in Central Tunisia and Northern Burkina Faso. Geomorphology 22 (2), 181–192. doi:10.1016/s0169-555x(97)00068-8
Wang, W. (2021). F. Effects of Sand Dune Microtopography on Soil, Vegetation and Arthropod Community Distribution in a Desert Sand-Fixing Vegetation Zone. Ningxia Univershaity. doi:10.27257/d.cnki.gnxhc.2021.001172
Wang, X., Wang, T., Dong, Z., Liu, X., and Qian, G. (2006). Nebkha Development and its Significance to Wind Erosion and Land Degradation in Semi-arid Northern China. J. Arid Environ. 65, 129–141. doi:10.1016/j.jaridenv.2005.06.030
Xu, H., Ye, M., Li, J. M., and Zhang, Q. Q. (2008a). Effect of Different Water Availability on Seed Germination in the Seed Bank of the Lower Tarim River Soil. Arid. Land Geogr. 31 (05), 650–658. doi:10.13826/j.cnki.cn65-1103/x.2008.05.003
Xu, L. H., Wang, J. H., Li, Y., Ma, Q. L., Zhang, D. K., Liu, Y. J., et al. (2008b). Characteristics of Soil Physical Property Changes during the Reversal of Desertification in the Southern Edge of Tengger Desert. J. Desert Res. 28 (04), 690–695.
Yang, L., Wang, Y. R., and Yu, J. D. (2010). Progress of Research on Soil Seed Bank in Arid Desert Areas. Acta pratac. Sin. 19 (02), 227–234. doi:10.3724/SP.J.1077.2010.01263
Yang, C. B., Li, Y., Shan, L. S., and Duan, Y. N. (2016). Assessment of the Seed Bank of Red Sand Scrub Soils and Their Natural Regeneration Potential in Loess Hilly Gully Areas. Bull. Soil Water Conserv. 36 (02), 105–109+114. doi:10.13961/j.cnki.stbctb.2016.02.020
Yin, R. P., Guo, J. Y., Dong, Z., He, J. L., Li, J. R., Tian, S. M., et al. (2017). Wind and Sand Characteristics of Typical Sand Dunes of Different Heights along the Ulaanbaatar Desert Section of the Yellow River. Res. Soil Water Conserv. 24 (05), 157–161. doi:10.13869/j.cnki.rswc.2017.05.024
Yu, J., Gao, L., Yan, Z., and Wang, Z. (2015). Characteristics of Soil Seed Bank of Sand Dune in Different Succession Stages in the Eastern Kubuqi Desert. Chin. J. Grass. 37 (04), 80–85. doi:10.3969/j.issn.1673-5021.2015.04.013
Zeng, Y., Wang, Y., and Nan, Z. (2003). Study on the Soil Seed Bank of Different Vegetation Types in the Alashan Arid Desert Region. Chin. J. Appl. Ecol. 14 (9), 1457–1463. doi:10.3321/j.issn:1001-9332.2003.09.011
Zhang, Y., and Li, J. (2018). Spatial Pattern of Seed Bank and Seedlings in Gurbantunggut Desert Soils the "protection Effect" of Scrub. Arid. Zone Res. 35 (05), 1138–1145. doi:10.13866/j.azr.2018.05.17
Zhang, T., Tian, C. G., Sun, Y., and Feng, G. (2006). Study on the Soil Seed Bank of Short-Lived Plants in the Gurbantunggut Desert Region. Arid. Land Geogr. 33 (05), 675–681. doi:10.13826/j.cnki.cn65-1103/x.2006.05.010
Zhang, Y. N., Li, C. L., Zhao, P., Duan, X. F., and Qiu, X. N. (2021a). Progress of Research on Soil Seed Bank in Arid Desert Areas. Seed Sci. Te. 39 (17), 9–11. doi:10.19904/j.cnki.cn14-1160/s.2021.17.004
Keywords: Nitraria tangutorum nebkhas, soil seed bank, spatial distribution, aboveground vegetation, desert–oasis ecotone
Citation: Li M, Xiao H, Xin Z, Li X, Li J, Miri A and Cao Q (2022) Soil Seed Bank Characteristics of Nitraria tangutorum Nebkhas in a Desert–Oasis Ecotone. Front. Environ. Sci. 10:937257. doi: 10.3389/fenvs.2022.937257
Received: 06 May 2022; Accepted: 13 June 2022;
Published: 04 August 2022.
Edited by:
Shengbo Xie, Northwest Institute of Eco-Environment and Resources (CAS), ChinaReviewed by:
Jiabing Wu, University of Chinese Academy of Sciences, ChinaZhongjie Shi, Chinese Academy of Forestry, China
Copyright © 2022 Li, Xiao, Xin, Li, Li, Miri and Cao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Huijie Xiao, c29pbGFuZHdhdGVyMjAwNkBob3RtYWlsLmNvbQ==