 Michael Niederwanger1*
Michael Niederwanger1* Josef Wieser2Melanie Thaler2Agnes Felber1Katja Schmölz3Wolfgang Mark1
Josef Wieser2Melanie Thaler2Agnes Felber1Katja Schmölz3Wolfgang Mark1 Erich Tasser3Rüdiger Kaufmann4
Erich Tasser3Rüdiger Kaufmann4 Bernd Pelster1
Bernd Pelster1- 1Institute of Zoology and Center of Molecular Biosciences Innsbruck, University of Innsbruck, Innsbruck, Austria
- 2Agentur Landesdomäne der Autonomen Provinz Bozen, Aquatic Species Conservation Centre, Bozen, Italy
- 3Eurac Research, Institute of Alpine Environment, Bozen, Italy
- 4Institute of Ecology, University of Innsbruck, Innsbruck, Austria
The increasing efflux on a large scale of organic contaminants holding endocrine disrupting activity from sewage treatment plants produces detrimental biological effects to various fish species. However, the impact of small-scale sewage treatment plant-efflux in small river streams and narrow creeks is largely unknown. Extensive pesticide output especially in vineyards and orchards also causes adverse effects on the endocrine system of wildlife fish species inhabiting nearby rivers. To elaborate whether fish species and populations in the areas of interest were at risk of experiencing endocrine disruption, we identified different biomarkers related to endocrine disruption in Cottus gobio and Salmo trutta fario and applied this approach to selected Austrian freshwater streams pre and post sewage treatment plants and permanent cultures with extensive pesticide output in South Tyrol. Overall, mRNA expression levels of vitellogenin, estrogen receptor α and zona pellucida genes in wildlife fish, compared to a control population reared under constant conditions in the laboratory were significantly increased. Sewage-treatment plant efflux did not significantly affect the mRNA expression levels while extensive use of pesticides altered mRNA expression significantly in C. gobio. C. gobio and S. trutta fario display different levels of mRNA expression. Cadmium and copper concentrations in liver tissues varied but did not indicate significant levels of contamination. Our results demonstrate the presence of endocrine disrupting chemicals in the tested freshwater streams. We anticipate our study to be a starting point for further studies focusing on the effects of endocrine disrupting chemicals on individuals and populations. Especially the fact that the two selected species reveal highly different levels of mRNA expression levels is of interest when applying biomarker approaches which can be a useful tool for monitoring projects and risk-assessment associated studies.
1 Introduction
The increasing input of endocrine-disrupting compounds (EDCs) such as pharmaceuticals, pesticides and industrial chemicals in aquatic ecosystems has become an issue of growing concern regarding potential threats to the animal kingdom (Vieira et al., 2021). Amongst others, scientific attention has focused on “xenoestrogens” with the potential to alter hormonal function and physiological status of wildlife species (Schwaiger and Negele, 1998; Arukwe, 2001; Goksøyr, 2006). In particular, the increasing number of reproductive abnormalities, mainly of feminizing nature such as retardation of gonadal development in combination with reduced fertility are known to be a result of endocrine disruption (Hutchinson et al., 2006; Yamamoto et al., 2017) and may affect the integrity of exposed fish populations (Jobling and Tyler, 2003). The feminization of male individuals in freshwaters affected by discharges from large EDC burdened sewage treatment plants has been described previously (Rodgers-Gray et al., 2001; Jobling et al., 2002; Harris et al., 2011). EDCs like natural and synthetic estrogens and/or estrogen mimicking chemicals enter the aquatic ecosystem via domestic and industrial sewage effluents and by deployment of pesticides (Bereswill et al., 2012; Herrero-Hernández et al., 2013; Yamamoto et al., 2017). Biomarkers are widely used to screen for the presence of EDCs, and to assess the sub-lethal changes that an organism encounters when exposed to environmental stressors, such as xenobiotic contaminants or toxic metals like Cadmium (Cd), which also can act as an endocrine disruptor (Depledge et al., 1995; Gómez González et al., 2017). Vitellogenin (VTG) mRNA expression has been used in different species to demonstrate exposure to EDCs (García-Reyero et al., 2004; Bjerregaard et al., 2008; Körner et al., 2008). VTGs are precursors of vitelline, which is crucial for embryonic development by providing required energy reserves (Matozzo et al., 2008). In the liver of oviparous vertebrates, they are present in high concentrations upon stimulation of the estrogen receptor by endogenous 17β-estradiol (Soverchia et al., 2005). Estrogens and xenoestrogens share a likewise structure (Sonnenschein and Soto, 1998) and xenoestrogens bind and activate the same estrogen receptor (Safe, 1995; Yamaguchi et al., 2005). In addition to VTG mRNA estrogen receptor α- and β (ERα, ERβ) (Körner et al., 2008) and zona pellucida (ZOP) mRNA have been used as biomarkers (Fossi et al., 2002; Baker et al., 2013). ERα and ERβ are nuclear transcription factors involved in the regulation of various physiological processes such as development, maturation and function of reproductive, central nervous, skeletal and cardiovascular systems (Bondesson et al., 2015) and are expressed in various tissues including the liver, ovary and gonads (Paterni et al., 2014). Zona radiata proteins like ZOP, synthesized by teleost fish hepatocytes in response to 17β-estradiol (Oppen-Berntsen et al., 1992) protect the developing embryo from environmental stress by enclosing it with an extracellular envelope (Tesoriero, 1978). Induction of different envelope proteins has been used as a marker for the presence of estrogenic endocrine disruptors (Oppen-Berntsen et al., 1999). Due to industrial pollution and agricultural usage, concentrations of essential metals like copper (Cu), zinc (Zn) and nonessential metals like Cd have been severely elevated in various marine and terrestrial ecosystems, and these compounds are known to be toxic to organisms above certain concentrations (Spurgeon et al., 1994; Jelassi et al., 2019). Cu and Zn in particular, used as an effective fungicide, has been applied extensively for more than 150 years (Genova et al., 2022). Although its use as a fungicide is strictly regulated nowadays, residual pollution and ongoing application causes increased Cu concentrations in the soil. By stealthy seeping into ground water, such compounds enter the freshwater streams and affect cellular metabolism (Pelgrom et al., 1995). Heavy metals have been shown to alter VTG expression in different ways (Zheng et al., 2010; Huang et al., 2014). Therefore, metallothioneins (MTs) may also be valuable biomarkers not only for metal contamination but also for the presence of EDCs (Rhee et al., 2009; Huang et al., 2014; Moncaleano-Niño et al., 2017). MTs are widely spread, low-molecular-mass, cysteine-rich, metal binding proteins. In most animal species, MTs are involved in the homeostatic regulation and detoxification of metallic trace elements (Klaassen et al., 1999; Egli et al., 2006) and participate in the cellular protection from oxidative stress (Ghoshal et al., 1998; Baird et al., 2006). MTs and also VTG are linked due to their function as metal ligands for Cd, Cu and Zn (Olsson et al., 1995; Werner et al., 2003).
The objective of the present study was to clarify whether fish species and populations in the areas of interest were at risk of experiencing endocrine disruption. To this end, we identified and measured potential biomarkers related to endocrine disruption and metal pollution in fish of the Alpine river system via quantitative real-time detection PCR (RT-qPCR) and assess Cd and Cu concentrations in the tissues. To screen for possible effects of sewage treatment effluents and pesticides we sampled European bullhead (Cottus gobio; C. gobio) and brown trout (Salmo trutta fario; S. trutta fario), two non-migratory species with therefore increased susceptibility to local environmental factors. C. gobio is a benthic freshwater fish inhabiting mainly river systems and estuaries of small and middle-sized rivers. The salmonid fish S. trutta fario, lives in streams with higher water flow and less pollution compared to typical habitats of C. gobio and was sampled alternatively when C. gobio was absent. Sampling sites were chosen upstream and downstream of sewage treatment plants in river streams of North and South Tyrol (Austria and Italy, respectively), with the former acting as a “wildlife control” and the latter being affected by sewage treatment plant effluent. We selected the sampling sites far apart from each other to minimize the risk of migration due to the absence of physical barriers. Additionally, we sampled fish in rivers passing permanent cultures (vineyards and orchards) in South Tyrol that are of significance due to the application of pesticides in high quantities.
The results of this study contribute to the current knowledge of endocrine disruption by describing in depth mRNA expression levels of different populations in varying habitats revealing species and site dependent differences of the two selected fish species.
2 Materials and methods
2.1 Sampling sites and experimental animals
The sampling sites are located in four rivers in the federal state Tyrol (Austria, Europe) and five rivers in the Autonomous province of South Tyrol (Italy, Europe) covering six sewage treatment plants and five areas with permanent cultures (Figure 1). Additional information (coordinates of sampling sites, treatment type, number and sex of sampled individuals) is listed in the supplementary material (Supplementary Table S1).
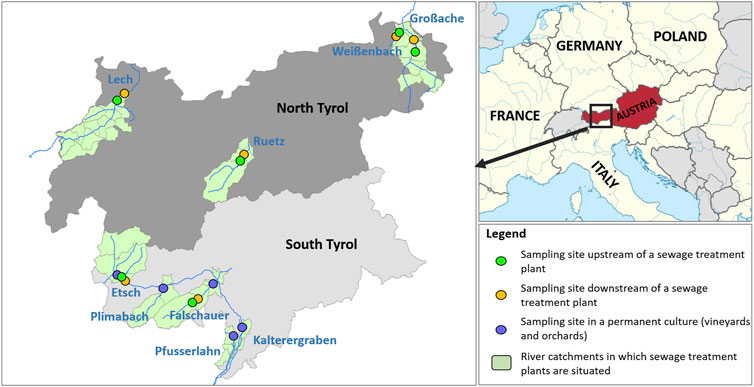
FIGURE 1. Map of Europe and augmented Map of North- and South Tyrol displaying the location and type of the sampling sites. Names of rivers are labeled in blue.
Sexually mature individuals of C. gobio and S. trutta fario were sampled via electrical fishing following Austrian guidelines for fish sampling (Haunschmid et al., 2006; Wagner et al., 2010), according to Zippin (1956) and Moran (2008). Fish were sacrificed using a physical approach directly upon withdrawal and dissected on site on an RNase free ice-cooled stainless-steel plate. Sample aliquots of liver and gonadal tissues were stored in RNA Later solution (Ambion Life Technologies, Carlsbad, California, United States) for subsequent total RNA isolation. Weight of the remaining tissues was determined before oven drying and prepared for metal analysis as described below. The work described in this article is complying with the ARRIVE (Animal Research: Reporting of in Vivo Experiments) guidelines and was carried out in accordance with the EU Directive 2010/63/EU for animal experiments. In order to evaluate expression levels of populations from different habitats, we sampled C. gobio from our laboratory fish culture at the Institute of Zoology in Innsbruck. Despite the fact that differences in basal expression levels between individuals from the wild and individuals from laboratory cultures can occur, it is vital for understanding species and population related differences of mRNA expression levels as a whole. Individuals of C. gobio were kept in freshwater tanks at 9°C and were used as “non-wildlife” controls. We purchased “non-wildlife” control individuals of S. trutta fario from a commercial dealer located in Italy (Fish farm Schiefer KG, San Leonardo, South Tyrol, Italy). We used these controls to compare the expression levels of fish, mostly unaffected from outside influences and wildlife fish. We sacrificed and dissected the controls from each species using the same procedure than for the fish sample in the wild.
2.2 Physical and chemical water parameters
At each sampling site, the following physical and chemical water parameters were assessed with a Multisonde (HI 9829-00042, HANNA Instruments, Vöhringen, Germany) on site: temperature, pH, dissolved oxygen, conductivity and turbidity. Additional water samples were taken, directly cooled to 4°C and analyzed within 24 h using a DR1900 spectrophotometer (HACH Lange GmbH, Colorado, United States). Corresponding Kits (HACH Lange GmbH, Colorado, United States) were used to measure Nitrate (NO3), Nitrite (NO2), Ammonium (NH4+), Phosphate (PO43−), water hardness, Nickel (Ni), Barium (Ba), Iron (Fe), Copper (Cu), Aluminum (Al) and Lead (Pb) concentrations in the water. Cd concentrations in the waters were measured with an atomic absorption spectrophotometer (model Z-8200, Hitachi, Tokyo, Japan) following the same protocol described above.
2.3 Pesticides
At four sampling sites characterized by intensive agricultural use and orcharding as well as viniculture, the Biological State Laboratory of the Autonomous Province of Bolzano (Supplementary Table S2) measured 180 pesticides in the river water in June, October and November. The pesticides were classified into seven groups: Herbicides, Fungicides, Insecticides, Molluscicides, Nematicides, Acaricides, and Rodenticides. The method was APAT NCR IRSA 5060 man 29 2003 (APAT—Agenzia per la protezione dell’ambiente e per i servizi tecnic, 2003) via LC-MS/MS (liquid chromatography-tandem mass spectrometry) and GC-MS/MS (Gas chromatography-mass spectrometry) (Payá et al., 2007; Melo et al., 2020), as well as the method DIN 38407—36:2014 through LC-MS/MS (Núñez et al., 2005). To condense the information from the 180 pesticides a principal component analysis (PCA) was carried out. As criteria, an eigenvalue larger than one and a satisfactory variance was chosen. As a result, two components with different pesticide mix resulted [for details see (Schmölz et al., 2022)].
2.4 Transcriptome generation and biomarker identification
To identify relevant biomarkers, isolated RNA from a liver tissue sample of one female individual of C. gobio was sent to the Duke Center for Genomic and Computational Biology (GBC, Duke University, Durham, NC, United States) for transcriptome generation. The RNA was subjected to HiSeq 4000 150 bp paired-end Illumina sequencing and one library was generated. The raw data is available within the NCBI Bio Project database (PRJNA579253) and was assembled at the Institute of Zoology (University of Innsbruck, Tyrol, Austria) using Trinity Version v2.8.8 (GitHub Inc., San Francisco, United States) (Grabherr et al., 2011) and provided for analysis on a local TBlast page. Known nucleotide sequences of close relatives coding for diverse biomarkers related to endocrine disruption were blasted against the transcriptome data set of C. gobio using TBlast of the Sequence Server software (Priyam et al., 2015). Nucleotide sequences were analyzed with CLC Main workbench v8.0.1 (Qiagen, Aarhus, Denmark) and aligned with the online tool Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/) in order to identify the respective reading frames. Gene specific primers were designed from identified sequences derived from transcriptomic data (see above) in order to confirm the sequences via PCR in three biological replicates for each examined biomarker. A 50-µl approach was set up using the Titanium Taq PCR system (Takara Clontech, Shimogyo-ku, Kyoto, Japan). PCR product separation occurred on a 1.5% agarose gel (Biozym, Hessisch Oldendorf, Germany) and gene specific bands were excised. DNA was purified using the QIAquick™ kit (Qiagen, Hilden, Germany), and pure samples were sent to Microsynth AG (Balgach, Switzerland) for Sanger sequencing. When necessary, DNA was subjected to subsequent cloning with the TOPO® TA Cloning® Kit for sequencing (Invitrogen, Thermo Fisher Scientific, Waltham, MA, United States). Inserts containing accurate plasmids were purified using the QIAprep Spin Miniprep Kit (Qiagen, Hilden, Germany) and sent to Microsynth AG (Balgach, Switzerland) for Sanger sequencing. Primer design and sequence analysis were performed applying CLC Main Workbench v8.0.1 (Quiagen, Aarhus, Denmark). Proven sequences of the following biomarkers from C. gobio were submitted to NCBI GenBank: VTG (MN577631), ERα (MN577627), ERβ (MN577628), ZOP (MN577629), and MT (MN577630).
2.5 mRNA isolation and cDNA synthesis
Total RNA isolation from ∼10 mg of homogenized (Precellys, Bertin Instruments, Montigny-le-Bretonneux, France) liver and gonadal tissues was achieved with Tri Reagent (Lab consulting, Vienna, Austria) and application of DNase 1 digestion (Invitrogen, Thermo Fisher Scientific, Waltham, MA, United States). RNA integrity screen was performed visually on an agarose gel stained with GelRed (Biotium Inc., Bay Area, CA, United States) and quantified with the RiboGreen®RNA Quantification Kit for Molecular Probes (Invitrogen, Thermo Fisher Scientific, Waltham, MA, United States) on a VICTOR™X4 2030 Multilabel Reader (PerkinElmer, Waltham, MA, United States). To remove RNases within the used water, it was treated with DMPC (Merck KGaA, Darmstadt, Germany). First strand cDNA was synthesized from 450 ng of total RNA with Reverse Transcriptase (Life technologies, Thermo Fisher Scientific, Waltham, MA, United States) in a 50-µl approach for RT-qPCR.
2.6 RT qPCR
RT-qPCR using Power SYBR Green (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, United States) was performed on a Quant Studio 3 (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, United States). The transcripts were amplified using the primers and concentrations shown in Table 1, following a protocol of 40 cycles: denaturation at 95°C for 15 s, annealing and extension combined at 60°C for 60 s. The 10 µl PCR reaction contained 1 µl of cDNA, 5 µl Power SYBR Green PCR master mix, 1 μl U-BSA, 1 µl water (Direct-Q®3 UV, Merck Millipore SAS, Molsheim, France) and 1 µl of each forward and reverse primer. The primers were designed with the Primer Express 3.0 software (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, United States). A primer matrix with implemented dissociation curves was created to determine ideal primer concentrations and quality. Calibration curves were generated by using sequence proven amplicon plasmids from three biological replicates for each mRNA sequence assessed. Cycle quantification (CT) values were estimated as followed: C. gobio: VTG: y = −3.4657x + 36.974; ERα: y = −3.4251x + 36.535; ERβ: y = −3.3539x + 34,574; ZOP: y = −3.2088x + 34.15; MT: y = −3.7347x + 38.867; S. trutta fario: VTG: y = −3.3104x + 34.777; ERα: y = −3.3438x + 34.459; ERβ: y = −3.3335x + 33.463 (PCR efficiencies shown in Table 1). Data evaluation was performed by using the Thermo Fisher Cloud Software, Version 1.0 (Life Technologies Corporation, Carlsbad, California, United States). The calculated amounts of mRNA are displayed in mRNA copy numbers per 10 ng total RNA.

TABLE 1. Used quantitative real time PCR primers and PCR efficiencies for Cottus gobio and Salmo trutta fario. qRT PCR primers for vitellogenin, estrogen receptor α- and β from S. trutta fario were adopted from Körner et al. (2008).
2.7 Metal analysis
Liver and gonadal tissue aliquots were dried in an oven at 65°C. Subsequently, the samples were pressure digested in 2 ml flat bottom tubes (Eppendorf, Hamburg, Germany) with a mixture of nitric acid (65%) (Suprapure, Merck, Darmstadt, Germany) and deionized water (1:1), in an aluminum oven covered with a heated lid at 69°C. Digested samples, were diluted with deionized water to 2 ml and Cd and Cu concentrations were measured with an atomic absorption spectrophotometer (model Z-8200, Hitachi, Tokyo, Japan). System calibration was performed with Cd and Cu standard solutions containing 1% nitric acid. The accuracy of metal measurements was verified using certified standard reference material TORT-2 (Lobster Hepatopancreas Reference Material for Trace Metals; National Research Council Canada) (n = 5) and DOLT-1 (Dogfish Muscle and Liver Reference Material for trace metals; National Research Council Canada) (n = 5).
2.8 Statistical method
Mixed effect models (site as random factor) were computed in Statistica 13 (TIBCO Software Inc., Palo Alto, 1984-2017). Boxplots with median, quartiles and range were created in R (R Development Core Team, 2008) using the package ggplot 2 (Wickham, 2016). Details of the statistical analyses are given as Supplementary Tables S3–S5.
The effects of water quality on vitellogenin, estrogen receptor α (ERα) and zona pellucida were assessed by General Linear Model procedure (GLM) in which factors (type of tissue, gender, species) and covariates (physical and chemical water parameters, pesticide mix), as well as their interactions are used as predictors. Collinearities among the covariates were assessed with the Variance Inflation Factor (VIF) measures. Of the chemical predictors, the following remained based on a level of VIF > 10 in the analysis: Nitrite (NO2), Phosphate (PO43−), water hardness (Ca, Mg), Barium (Ba), Iron (Fe), Copper (Cu), Aluminum (Al), Lead (Pb), N and P total as well as the pesticide mix 1. Of the water physical parameters, temperature, conductivity and turbidity were used. Data of vitellogenin and zona pellucida expression were transformed [Box-Cox transformation (Hemmerich, 2016)] to achieve the assumptions of multivariate normality and the homogeneity of variance-covariance matrices (Levene’s Test of Equality of Error Variances, White Test for Heteroscedasticity). The analyses were performed with IBM SPSS Statistics for Windows, Version 27.0.Armonk, NY: IBM Corp.
3 Results
3.1 Cottus gobio
3.1.1 Basal expression levels of vitellogenin, estrogen receptor α and zona pellucida
Basal wildlife VTG levels were significantly elevated in male liver tissues reaching up to 10,000 copies per 10 ng total RNA compared to male samples from a laboratory control population, where levels reached 500 copies per 10 ng total RNA or were even below detection limits (Figure 2A; all statistical examinations are shown in the Supplementary Table S3. Compared to males, VTG levels were up to 500-fold elevated in female liver tissues. The average levels from sampling sites ranged from 700 to 10 million copies. VTG levels in all male samples from the wild were significantly elevated when compared to laboratory controls but significantly lower when compared to female individuals. VTG levels measured in the liver of females from the different sampling sites were all higher than the levels detected in female laboratory controls. Particularly high VTG expression levels were measured in the rivers Lech, Weissenbach, Großache in North Tyrol, and the River Etsch in South Tyrol. Compared to females, expression levels of ERα were about 10-fold decreased in liver tissues of males (Figure 2B). In male wildlife samples, ERα expression levels in the liver tissues were more than 3-fold elevated compared to laboratory controls. In the Großache they were almost elevated to 10-fold, reaching the level of females. We detected the highest expression levels in females of the rivers Lech and Großache. In the liver of females, ZOP mRNA expression levels displayed a 10-fold increase compared to male liver samples (Figure 2C). As observed for VTG and ERα expression, ZOP expression levels in the liver of wildlife male individuals was higher than the values recorded for laboratory control males. Likewise, in females ZOP mRNA levels were significantly elevated when compared to laboratory control females, except for the Falschauer. In the rivers Lech, Weissenbach, Großache and Etsch elevated levels up to 100-fold were recorded. Associated data for sewage treatment plants, pesticide loaded permanent cultures and controls for the respective biomarkers are displayed in Figures 2D–F, indicating significant differences between female and male individuals.

FIGURE 2. Log transformed expression levels (mRNA copy numbers per 10 ng total RNA) of (A) vitellogenin (VTG) (ANOVA; p = 0.0004), (B) estrogen receptor α (ERα) (ANOVA; p = 0.0006), (C) zona pellucida (ZOP) (ANOVA; p = 0.0002), in liver tissues of male (turquoise) and female (red) individuals of C. gobio (C. gobio) at the respective sampling rivers. Combined data of expression levels of (D) VTG, (E) ERα, (F) ZOP in liver tissues of male (blue) and female (red) individuals of C. gobio. Significant differences of mRNA expression between males and females were confirmed by ANOVA (p ≤ 0.001) and are indicated with stars.
3.1.2 Sewage treatment plants and permanent cultures
mRNA expression levels were measured in livers of male C. gobio collected upstream (control) and downstream of a sewage treatment plant at three different rivers in North Tyrol and one river in South Tyrol (Figures 3A–C; Supplementary Table S4). VTG expression was increased significantly in the river Lech at both sampling sites compared to the other rivers. Apart from that, the mRNA expression levels of VTG, ERα, and ZOP were not altered significantly at the sampling sites due to sewage treatment plant-efflux. Expression levels of the three genes VTG, ERα, and ZOP from samples collected in rivers passing through permanent cultures in South Tyrol were significantly elevated compared to the basal expression levels recorded for laboratory control fish (Figures 3D–F; Supplementary Table S4). Especially samples collected from the Leiferer Graben displayed very high mRNA expression levels for all tested biomarkers. In addition to ERα we also tested the expression of ERβ, but neither in liver samples taken from fish collected in the different rivers nor in samples collected from fish from rivers passing through permanent cultures in South Tyrol any significant difference between laboratory controls and wildlife samples were detected (data not shown). We also applied all measurements described for liver tissues to gonadal tissues alike. Basal mRNA expression levels in gonadal tissues were significantly lower than in liver tissues, sometimes even close to or below the detection limits. The results did not show significant differences for any of the tested biomarkers between males and females, pre- and post-sewage treatment plants or at pesticide loaded locations (data not shown).
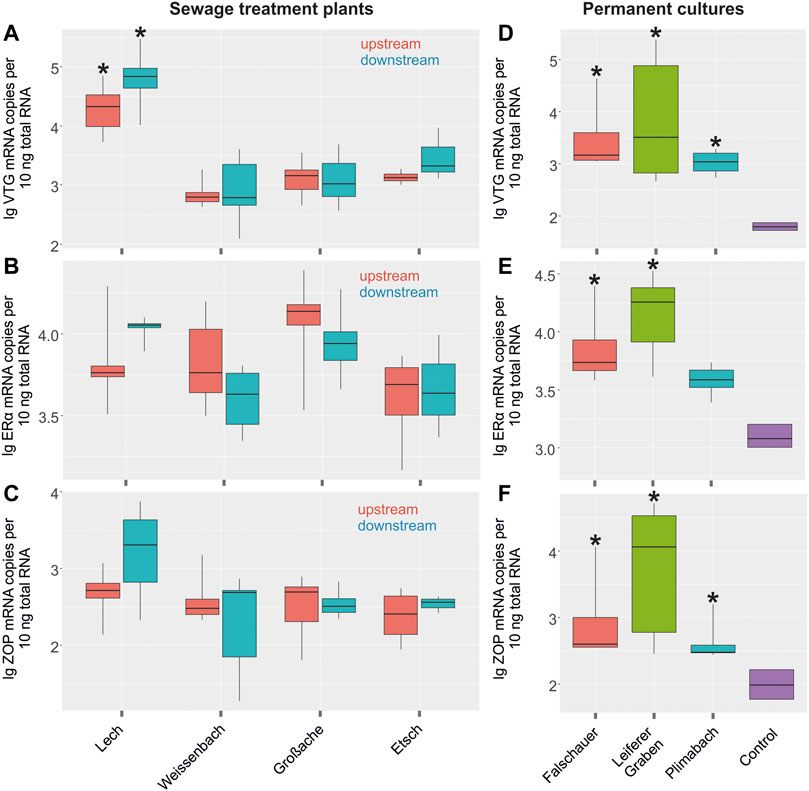
FIGURE 3. Log transformed expression levels (mRNA copy numbers per 10 ng total RNA) of (A) vitellogenin (VTG), (B) estrogen receptor α (ERα), (C) zona pellucida (ZOP) in liver tissues of control individuals upstream of a sewage treatment plant (red) and individuals downstream the sewage treatment plant (turquoise) in individuals of C. gobio. Asterisks indicate a significant difference to other sampling sites of the study (significance level <0.05). Log transformed expression levels (mRNA copy numbers per 10 ng total RNA) of (D) VTG, (E) ERα, (F) ZOP in liver tissues of control individuals from a laboratory culture (lilac) and individuals from permanent cultures (red, green and turquoise). Asterisks indicate a significant difference to the controls (significance level <0.05).
3.1.3 Metallothionein expression
MT expression levels were tested sex dependent in fish collected upstream and downstream of the sewage treatment plants since it has been shown that sex could have an impact on MT expression in some vertebrates (Blazka and Shaikh, 1991; Adeogun et al., 2020). However, no sex related significant differences in MT expression were observed. Therefore, we displayed MT mRNA expression levels graphically sex-independent (Figure 4). MT mRNA levels measured in control liver samples reached on average 330,000 copies per 10 ng total RNA. Fish collected upstream in the river Etsch in South Tyrol had slightly higher MT expression levels than the fish collected upstream in North Tyrol, but this difference was not significant. MT mRNA levels detected in liver samples of fish collected downstream of the sewage plants ranged from 50,000 to 600,000 copies, and they were not significantly different from the levels detected in fish collected upstream. In permanent cultures, no effect on MT expression levels was detected. MT expression levels recorded in gonadal tissues of these fishes were significantly lower than the levels measured in liver samples. No differences were detected between the MT expression levels measured in laboratory control samples and the samples collected in the rivers (data not shown). Furthermore, no coherence between VTG and MT mRNA expression was discovered.
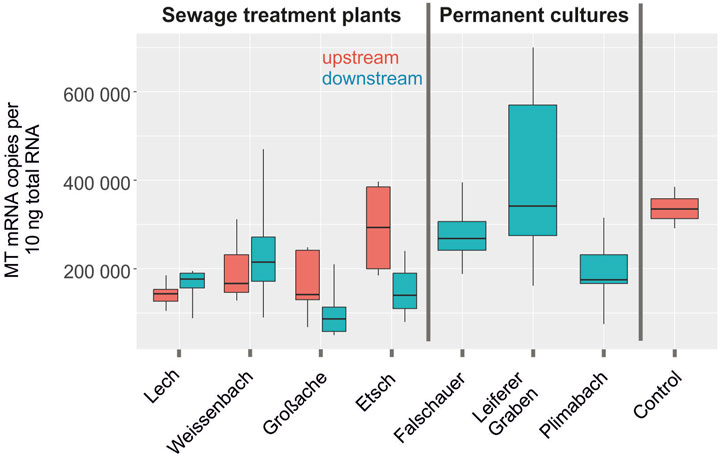
FIGURE 4. Metallothionein expression levels (mRNA copy numbers per 10 ng total RNA) in liver tissues of C. gobio in river streams of control individuals upstream of a sewage treatment plant (red) and individuals downstream the sewage treatment plant (turquoise) (all labeled sewage treatment plants). Rivers passing vineyards and orchards are labeled permanent cultures.
3.2 Salmo trutta fario
3.2.1 Basal expression levels of vitellogenin and estrogen receptor α
We sampled S. trutta fario at three sampling sites due to the absence of C. gobio. Similar to C. gobio, sex-dependent basal expression levels of biomarkers in the livers of S. trutta fario were determined by comparing male and female individuals (Figures 5A,B; Supplementary Table S5). VTG expression in the liver of S. trutta fario females collected in the rivers Ruetz and Falschauer in South Tyrol were significantly higher than the levels recorded in male liver samples. Similarly, liver samples from females collected in the Punibach showed much higher VTG expression levels than samples taken from male fish. ERα expression levels measured in fish from these rivers revealed no difference between males and females except for the Punibach, where the expression levels in males were lower when compared to females (Figure 5B).
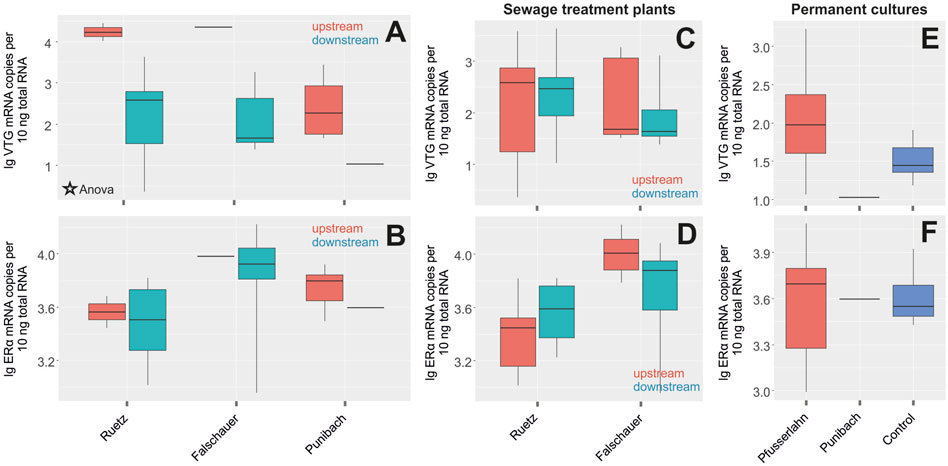
FIGURE 5. Log transformed expression levels (mRNA copy numbers per 10 ng total RNA) of (A) vitellogenin (VTG) (ANOVA; p = 0.0009), (B) estrogen receptor α (ERα) in liver tissues of male (turquoise) and female (red) individuals of S. trutta fario at the respective sampling sites. Significant differences of mRNA expression between males and females were confirmed by ANOVA (p ≤ 0.001). Log transformed expression levels (mRNA copy numbers per 10 ng total RNA) of (C) VTG, (D) ERα in liver tissues of control individuals upstream of a sewage treatment plant (red) and individuals downstream the sewage treatment plant (turquoise) of S. trutta fario. Log transformed expression levels of (E) VTG, (F) ERα in liver tissues of control individuals from a laboratory culture (blue) and individuals from permanent cultures of S. trutta fario.
3.2.2 Sewage treatment plants and permanent cultures
VTG and ERα expression tested in S. trutta fario males collected upstream and downstream of the sewage treatment plant in the rivers Ruetz and Falschauer revealed no significant differences (Figures 5C,D; Supplementary Table S5). Wildlife samples from freshwater streams passing permanent cultures did not reveal enhanced mRNA expression levels compared to control individuals from an aquaculture (Figures 5E,F; Supplementary Table S5). Additionally, ERβ expression was analyzed, but no differences were detected between controls and the samples collected in the different rivers nor between controls and the samples collected from fish from rivers passing through permanent cultures in South Tyrol (data not shown). No significant differences in the expression levels between males and females, pre- and post-sewage treatment plants or controls and pesticide-loaded locations (data not shown) were detected in gonadal tissue.
3.3 Cd and Cu accumulation in Cottus gobio and Salmo trutta fario
Measurements of Cd accumulation in the liver of C. gobio (Figure 6A) collected pre and post sewage treatment plants revealed no significant differences. Noteworthy, however, were the elevated levels of Cd in liver samples from the rivers Lech and Etsch, compared to laboratory controls and the rivers Weissenbach and Großache. Compared to laboratory controls, liver samples collected from the Leiferer Graben and Plimabach, rivers passing through permanent cultures, also showed significantly elevated levels of Cd. Cd concentrations in gonadal tissues were significantly lower than in liver tissue, and no difference was detected between the different sampling sites (data not shown). Cu concentrations measured in the liver of C. gobio collected were not different from laboratory control values and no difference was detected between sampling sites (Figure 6B). Only samples collected from the Großache showed a remarkable variation in the Cu concentration.
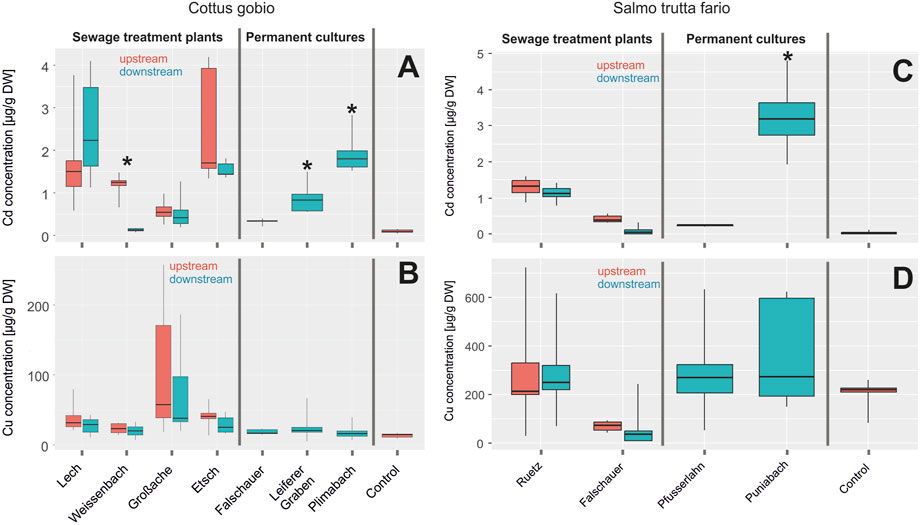
FIGURE 6. (A) Cd and (B) Cu accumulation (in µg per g dry weight) in liver tissues of C. gobio in river streams of control individuals upstream of a sewage treatment plant (red) and individuals downstream the sewage treatment plant labeled impact (turquoise) (all labeled sewage treatment plants). Rivers passing vineyards and orchards are labeled with permanent cultures. Asterisks indicate a significant difference to the controls (significance level 0.05). (C) Cd and (D) Cu accumulation (in µg per g dry weight) in liver tissues of S. trutta fario in river streams of control individuals upstream of a sewage treatment plant (red) and individuals downstream the sewage treatment plant labeled impact (turquoise) (all labeled sewage treatment plants). Rivers passing vineyards and orchards are labeled permanent cultures. Asterisks indicate a significant difference to the controls (significance level 0.05).
Cd accumulation in the liver of S. trutta fario was low and there was no difference between samples collected pre- and post-sewage treatment plants, except for the Punibach, where values were in the range of 3–4 µg per g dry weight (Figure 6C). Interestingly, the Cu accumulation was significantly higher in S. trutta fario than in C. gobio (Figure 6D). Compared to liver tissue, gonadal tissue of S. trutta fario displayed significantly lower values for Cd and Cu, and there were no significant differences between sampling sites (data not shown).
3.4 Physical and chemical water parameters
Temperature measured at the different sampling sites in North and South Tyrol before and behind the sewage treatment plants varied between 8°C and 13°C; dissolved oxygen concentrations were with values of about 10 mg L−1 in the upper range, close to or even slightly above full oxygen saturation at atmospheric oxygen partial pressures (Supplementary Tables S6, S7). None of the sampling sites showed elevated concentrations of ammonia, nitrite or nitrate, and overall conductivity was low with values between 150 and 300 µs cm−1, except for Weissenbach in North Tyrol and Etsch in South Tyrol, where conductivity was near 500 µs cm−1 (Supplementary Table S6). Similarly, concentration of metals measured in these water samples were low, sometimes below the detection threshold. Supplementary Table S7 summarizes physical and chemical water parameters determined in South Tyrolean Rivers passing through permanent cultures. Temperature varied between 13 and 15°C and dissolved oxygen concentration was near 10 mg L−1.
3.5 Effects of water quality on biomarkers
Type of tissue and the sex of the individuals correlated significantly with the expression levels of all biomarkers (Table 2). As shown in the previous results, livers have significantly higher levels of VTG and ZOP, whereas the gonads have higher levels of ERα. Not surprisingly, females also contained higher numbers of mRNA copies of VTG and ZOP overall. Using the General Linear Model procedure (GLM) we tried to identify a possible correlation between these expression data and physical and chemical water parameters. Especially in the case of VTG, our data show that some water chemical and physical parameters were correlated with it. Thus, there was a positive correlation between the concentration of total nitrogen, iron and copper in water with VTG expression. On the other hand, calcium hardness was correlated negatively with the number of VTG mRNA copies. Furthermore, the VTG expression also correlated positively with at higher temperatures and a higher conductivity. Finally, the VTG expression was positively correlated with a higher concentration of pesticides in the water and, proximity to sewage treatment plants. ERα expression was positively correlated only with temperature, whereas turbidity, nitrogen content and iron content were negatively correlated with this biomarker. In the case of ZOP, in addition to tissue and sex, only aluminum content affected expression (negative correlation).
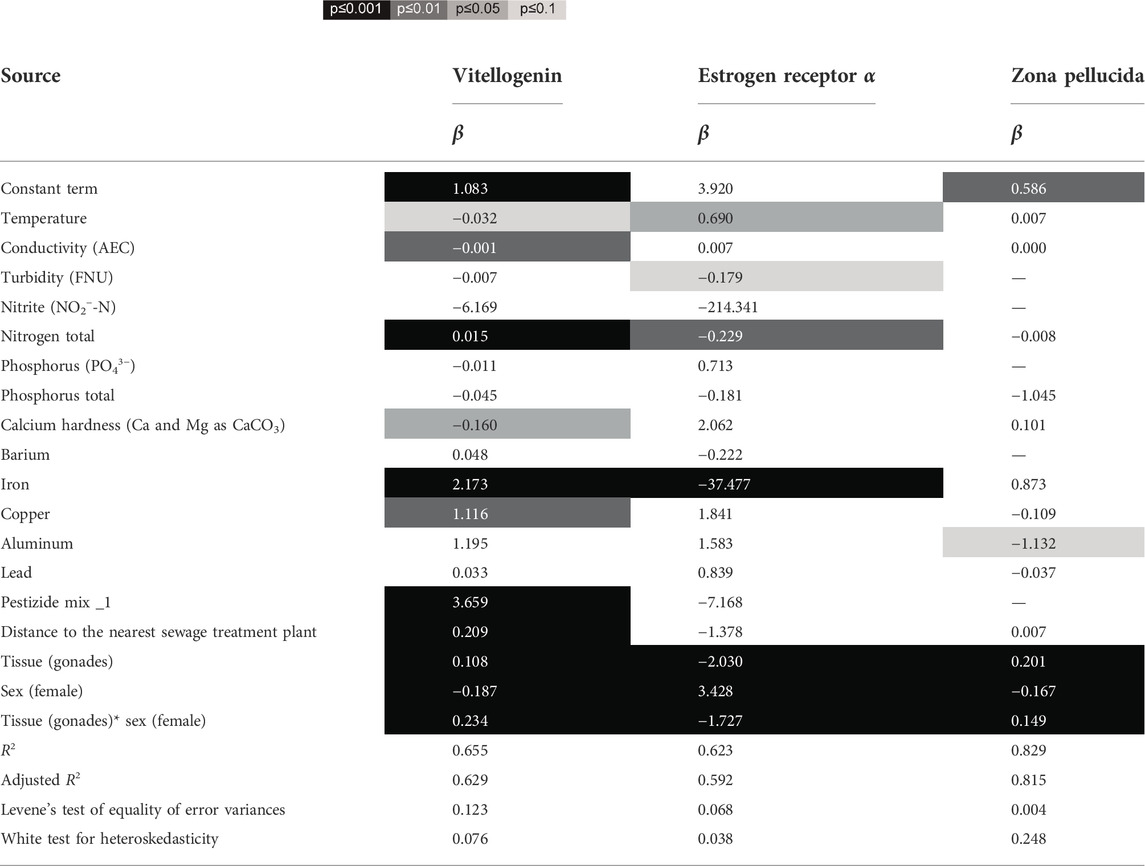
TABLE 2. Statistical results of GLM models to the effects of tissue and sex (factors) and chemical and physical water parameters, as well as pesticide mix (as covariates) on the expression levels of vitellogenin, estrogen receptor α and zona pellucida. Significant correlations and interactions based on type III Wald statistics are highlighted ( ), for all predictors the regression coefficients (β) are given. R2, adjusted R2, Levene’s Test of Equality of Error Variances and White Test for Heteroskedasticity are provided for each model.
), for all predictors the regression coefficients (β) are given. R2, adjusted R2, Levene’s Test of Equality of Error Variances and White Test for Heteroskedasticity are provided for each model.
4 Discussion
Of the tested genes, the expression levels of VTG, ERα, and ZOP in the liver of wildlife control fish were elevated compared to laboratory controls and revealed significant differences between male and female individuals of C. gobio. and of S. trutta fario. Our data suggested three distinct levels of mRNA expression, where the lowest levels of expression were found in male controls from a laboratory- or aquaculture, the intermediate level were detected in male wildlife fish and the highest expression levels in female wildlife fish. VTG and ZOP expression have repeatedly been used as a biomarker (Celius et al., 2000; Fossi et al., 2002; García-Reyero et al., 2004; Barucca et al., 2006; Bjerregaard et al., 2008; Baker et al., 2013). Körner et al. (2008) reported that ethinylestradiol exposure stimulated ERα but not ERß expression in juvenile brown trout. Our results suggest that this holds true for adult individuals alike. While ERα expression was significantly higher in females of C. gobio and S. trutta fario, expression levels of ERß in liver tissues revealed no differences between control males and females. In addition to liver tissue, expression of these genes was assessed in gonadal tissues. In both species, expression levels of VTG, ERα, and ZOP in gonadal tissues were significantly lower than in liver tissues, and there was no significant difference between male and female expression levels. These results support the assumption that the liver is the main organ for mRNA expression of the tested biomarkers and therefore should be used for possible screening studies. Expression levels of VTG, ERα, and ZOP in liver samples of wildlife fish were consistently higher than the expression levels measured in the laboratory control population, suggesting that in the wild fish may have been exposed to EDC’s. However, during dissection, no obvious organ modifications of feminizing nature were observed. Furthermore, no hints of decreased fertility or unstable population density in the rivers were detected. Therefore, it remains unclear whether these significantly increased basal expression levels of VTG, ERα, and ZOP with respect to laboratory controls in male wildlife individuals was sufficient to cause detrimental biological effects and therefore led to a decrease in reproductive success. The different levels of mRNA expression between the two species where ERα and ZOP mRNA expression levels of S. trutta fario were below RT-qPCR detection limits is of particular interest, since this proposes disparities in the response mechanisms of the two species.
The sewage treatment plant-efflux in the tested rivers did not significantly alter mRNA expression levels of the assessed biomarkers in three rivers, except for liver tissue from C. gobio collected downstream of the sewage treatment plant at the river Lech, which showed elevated expression levels especially of VTG mRNA. This could be due to the observation that sewage treatment plants may not eliminate synthetic estrogen, some heavy metals and pesticides from wastewaters effectively enough (Larsson et al., 1999; Parkkonen et al., 2000). Another possible explanation could be the high immigration of new residents in the catchment area of this river within the last few years and an increasing summer- and winter tourism into this region, leading to seasonal peaks of increased wastewater efflux. Especially during winter season, the number of guest nights increased by a factor of five compared to the summer months in recent years (https://gemeinde.lech.eu/servicecenter/gaestelaenderstatistik/). The efflux of an additional sewage treatment plant, located upstream our sampling sites in a likewise touristic region, may have contributed to the already increased mRNA levels detected at the site upstream the sewage treatment plant. Therefore, the present sewage treatment plant may be overstrained due to the altered concentration of EDCs upstream.
Using the General Linear Model procedure, we identified correlations between individual physical and chemical water parameters and the mRNA expression, especially in the liver tissue of males. In liver tissue, VTG mRNA expression increased with increasing total nitrogen, iron and Cu concentration in the water, and it decreased with increasing Ca2+ hardness. The latter effect is in concordance with previous results by Yeo and Mugiya (1997), who detected a decrease of VTG synthesis with increasing extracellular calcium concentrations in rainbow trout. Water hardness is one of the most important factors that affect fish physiology and metal toxicity (Saglam et al., 2013), because their gills are constantly submersed in water containing metal ions. In general, Cd toxicity is increased upon reduced water hardness (Pilehvar et al., 2020) and it has been shown to be more toxic in freshwater compared to saltwater since it associates with saltwater chlorides to form a less available molecule from solution (Bradl, 2005). Through passive diffusion over the gills or ingested and absorbed via endocytosis, metal ions enter chloride cells in the gills through calcium channels (Olsson et al., 1998) and interact with different cytoplasmatic components such as MTs (Perera et al., 2015). In fish, naturally occurring waterborne cations and trace metals interact with gill surfaces and other exchange structures resulting in competitive/non-competitive inhibition of the uptake and accumulation processes, hence, modifying metal toxicity (Paquin et al., 2002). In female fathead minnow Cu exposure had no effect on the expression of ERß and VTG; while ERα expression was slightly decreased (Driessnack et al., 2017). Cd exposure significantly decreased VTG mRNA expression, but increased ERß expression (Driessnack et al., 2016). Exposure of the fish to water containing a mixture of copper and nickel resulted in a decrease in hepatic VTG expression in females (Driessnack et al., 2017). In females of the mosquitofish, Zn, Cd, and Pb have been shown to induce VTG and MT- mRNA expression (Huang et al., 2014). In fathead minnow, Cu exposure also enhanced MT expression (Driessnack et al., 2016; Driessnack et al., 2017). Significant metabolic changes such as altered VTG gene expression, lipid transport activity, defense response, innate immune response and metal ion binding activity support the hypothesis that exposure to Cd induces endocrine disruption in aquatic animals (Kim et al., 2016). Taken together these results support the notion that heavy metals may affect not only MT expression, but also VTG expression or ER expression. It is of note that Yang and Sun showed, that Cd not only affects gene expression but can also decrease VTG accumulation in the hepatopancreas of the benthic freshwater crab Sinopotamon henanense due to elevated energy consumption and an activated defense system (Yang et al., 2017). The energy metabolism is enhanced to balance increased ATP consumption due to an increased protein synthesis which is upregulated due to external stress such as metals (Cherkasov et al., 2006), leading to a downregulated VTG accumulation (Yang et al., 2017). In general, the metabolic utilization of energy to support basal functions is of higher priority than reproduction, growth or nutrient storage (Sokolova et al., 2012). Maternal exposure to Cu and also to Cd has been shown to decrease brood size in live-bearing western mosquito fish Gambusia affinis (Cazan and Klerks, 2015). While many studies focused on female fish, our study revealed that in males VTG and ER expression may also be affected, and we even found stronger effects in males. Our results showed a temperature dependent expression of VTG and ERα mRNA, as reported in a previous study (Körner et al., 2008).
Pesticides may pollute South Tyrolean Rivers passing permanent cultures, and our results confirm this observation. Compared to laboratory controls, expression levels of VTG, ERα, and ZOP expression was significantly elevated in liver samples from C. gobio and S. trutta fario males. However, the expressions levels detected in males were significantly lower than the expression values recorded in females. This suggested that rivers passing through permanent cultures indeed show an increased level of pollution. MT mRNA expression is often used as a biomarker for metal pollution, but it is known that EDCs such as 17ß estradiol can alter MT expression in different ways (Olsson et al., 1995). High concentrations of 17ß estradiol and of Cd significantly altered the MT expression levels (Martin et al., 2003; Zheng et al., 2010; Huang et al., 2014). Elevated MT expression has also been reported for fathead minnow exposed to Cu, or to a mixture of Cu and either Cd or Ni (Driessnack et al., 2016; Driessnack et al., 2017). Additionally, it has been demonstrated that sex can have an impact on MT expression in vertebrates and arthropods (Blazka and Shaikh, 1991; Legras et al., 2000; Adeogun et al., 2020). Statistical analysis of our data, however, did not identify significant sex-related differences. Our expression data for MT mRNA in female and male individuals did not reveal any significant differences between laboratory controls and the various sampling sites, and there was no correlation between VTG and MT mRNA expression levels. The liver is known to be the main storage organ for Cd and Cu in fish (Kamunde et al., 2005; Driessnack et al., 2016), and our results confirmed that MT’s are mainly produced in liver tissue. Metals may induce MTs, and our data therefore indicate that in none of our sampling sites the concentration of metals like Cd and Cu, which are bound by MTs, was high enough to enhance MT expression. Indeed, measurements of Cd and Cu concentrations in the livers of our collected fish did not display signs of severe Cd or Cu accumulation, and the concentrations were even lower in gonadal tissue. In female fathead minnow, exposure to waterborne Cu also did not result in an accumulation of Cu in ovary tissue (Driessnack et al., 2016; Driessnack et al., 2017).
5 Conclusion
Our results suggest that VTG, ERα, and ZOP mRNA expression levels, measured in the livers of C. gobio and VTG in the livers of S. trutta fario serve as valuable biomarkers for EDC related monitoring projects and the two fish species can be used as bio indicators for metal pollution. The constantly increased expression levels in wildlife fish do suggest pollution with EDCs and pesticides to some extend in small rivers that are interspersed with partially overstrained small-scale sewage treatment plants or flow through permanent cultures. The two species also significantly differ regarding overall mRNA expression levels. It is of note, that ERα and ZOP mRNA expression levels in S. trutta fario constantly were below detection limits, which indicates significant differences in the response mechanisms of the two species.
However, further studies are necessary to evaluate the impact of the documented elevated expression levels on the physiology and reproductive biology of C. gobio and S. trutta fario and to investigate in depth whether slightly increased expression levels lead to ascertainable detrimental effects of feminizing nature.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: NCBI GenBank: VTG (MN577631), ERα (MN577627), ERβ (MN577628), ZOP (MN577629), and MT (MN577630).
Ethics statement
Ethical review and approval was not required for the animal study because the work described in this article is complying with the ARRIVE (Animal Research: Reporting of in Vivo Experiments) guidelines and was carried out in accordance with the EU Directive 2010/63/EU for animal experiments.
Author contributions
MN: Conceptualization, methodology, validation, investigation, data curation, formal analysis, writing—original draft, writing—review and editing, visualization JW: Methodology, investigation MT: Methodology, investigation, formal analysis AF: Methodology, investigation, formal analysis KS: Methodology, investigation, formal analysis WM: Conceptualization, methodology, investigation, resources, ET: Conceptualization, methodology, funding acquisition, project administration RK: Software, formal analysis BP: Conceptualization, methodology, funding acquisition, supervision.
Funding
The research leading to these results has received funding from the INTERREG Italia—Österreich European Regional Development Fund, under Grant Agreement ITAT1041, ALFFA Holistic (multiscale) analysis of the factors and their effect on the fish fauna in inner-Alpine space and from the University of Innsbruck.
Acknowledgments
We are grateful to the Department of Hunting and Fishery, Department of Water Conservation, Biological State Laboratory and Agency for Civil Protection of the Autonomous Province of Bolzano as well as the Department of Water Management, Environmental Protection and TIRIS (Tyrolean Geographic Frontiers in Information System) of the Federal State Tyrol for providing data and expert input. We further thank all associated partners in the research project, including fisheries association in Tyrol and South Tyrol, Umbrella Organization for Nature Conservation and Environmental Organizations South Tyrol, FUST‐Tyrol e.V. and Laimburg Research Centre. In addition, our thanks and appreciation to the anglers in South Tyrol and Tyrol for their cooperation. The authors declare that this study received contribution from the TIWAG-Tiroler Wasserkraft AG. The company was not involved in the study design, collection, analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2022.1027062/full#supplementary-material
References
Adeogun, A. O., Ibor, O. R., Omiwole, R., Chukwuka, A. V., Adewale, A. H., Kumuyi, O., et al. (2020). Sex-differences in physiological and oxidative stress responses and heavy metals burden in the black jaw tilapia, Sarotherodon melanotheron from a tropical freshwater dam (Nigeria). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 229, 108676. doi:10.1016/j.cbpc.2019.108676
Arukwe, A. (2001). Cellular and molecular responses to endocrine-modulators and the impact on fish reproduction. Mar. Pollut. Bull. 42 (8), 643–655. doi:10.1016/s0025-326x(01)00062-5
Baird, S. K., Kurz, T., and Brunk, U. T. (2006). Metallothionein protects against oxidative stress-induced lysosomal destabilization. Biochem. J. 394 (1), 275–283. doi:10.1042/bj20051143
Baker, M. E., Vidal-Dorsch, D. E., Ribecco, C., Sprague, L. J., Angert, M., Lekmine, N., et al. (2013). Molecular analysis of endocrine disruption in Hornyhead Turbot at wastewater outfalls in southern California using a second generation multi-species microarray. PLoS One 8 (9), e75553. doi:10.1371/journal.pone.0075553
Barucca, M., Canapa, A., Olmo, E., and Regoli, F. (2006). Analysis of vitellogenin gene induction as a valuable biomarker of estrogenic exposure in various Mediterranean fish species. Environ. Res. 101 (1), 68–73. doi:10.1016/j.envres.2005.07.002
Bereswill, R., Golla, B., Streloke, M., and Schulz, R. (2012). Entry and toxicity of organic pesticides and copper in vineyard streams: Erosion rills jeopardise the efficiency of riparian buffer strips. Agric. Ecosyst. Environ. 146 (1), 81–92. doi:10.1016/j.agee.2011.10.010
Bjerregaard, P., Hansen, P. R., Larsen, K. J., Erratico, C., Korsgaard, B., and Holbech, H. (2008). Vitellogenin as a biomarker for estrogenic effects in Brown trout, Salmo trutta: Laboratory and field investigations. Environ. Toxicol. Chem. 27 (11), 2387–2396. doi:10.1897/08-148.1
Blazka, M. E., and Shaikh, Z. A. (1991). Sex differences in hepatic and renal cadmium accumulation and metallothionein induction. Biochem. Pharmacol. 41 (5), 775–780. doi:10.1016/0006-2952(91)90080-o
Bondesson, M., Hao, R., Lin, C. Y., Williams, C., and Gustafsson, J. Å. (2015). Estrogen receptor signaling during vertebrate development. Biochim. Biophy. Acta. 1849 (2), 142–151. doi:10.1016/j.bbagrm.2014.06.005
Bradl, H. (2005). Heavy metals in the environment: Origin, interaction and remediation. 1st Edn. New York, USA: Academic Press, 282.
Cazan, A. M., and Klerks, P. L. (2015). Effects from a short-term exposure to copper or cadmium in gravid females of the livebearer fish (Gambusia affinis). Ecotoxicol. Environ. Saf. 118, 199–203. doi:10.1016/j.ecoenv.2015.04.039
Celius, T., Matthews, J. B., Giesy, J. P., and Zacharewski, T. R. (2000). Quantification of rainbow trout (Oncorhynchus mykiss) zona radiata and vitellogenin mRNA levels using real-time PCR after in vivo treatment with estradiol-17β or α-zearalenol. J. Steroid Biochem. Mol. Biol. 75 (2–3), 109–119. doi:10.1016/s0960-0760(00)00165-5
Cherkasov, A. S., Biswas, P. K., Ridings, D. M., Ringwood, A. H., and Sokolova, I. M. (2006). Effects of acclimation temperature and cadmium exposure on cellular energy budgets in the marine mollusk Crassostrea virginica: Linking cellular and mitochondrial responses. J. Exp. Biol. 209 (7), 1274–1284. doi:10.1242/jeb.02093
Depledge, M. H., Aagaard, A., and Györkös, P. (1995). Assessment of trace metal toxicity using molecular, physiological and behavioural biomarkers. Mar. Pollut. Bull. 31 (1–3), 19–27. doi:10.1016/0025-326x(95)00006-9
Driessnack, M. K., Jamwal, A., and Niyogi, S. (2017). Effects of chronic exposure to waterborne copper and nickel in binary mixture on tissue-specific metal accumulation and reproduction in fathead minnow (Pimephales promelas). Chemosphere 185, 964–974. [Internet]. doi:10.1016/j.chemosphere.2017.07.100
Driessnack, M. K., Matthews, A. L., Raine, J. C., and Niyogi, S. (2016). Interactive effects of chronic waterborne copper and cadmium exposure on tissue-specific metal accumulation and reproduction in fathead minnow (Pimephales promelas). Comp. biochem. Physiol. C Toxicol. Pharmacol. 179, 165–173. doi:10.1016/j.cbpc.2015.10.009
Egli, D., Domènech, J., Selvaraj, A., Balamurugan, K., Hua, H., Capdevila, M., et al. (2006). The four members of the Drosophila metallothionein family exhibit distinct yet overlapping roles in heavy metal homeostasis and detoxification. Genes Cells 11 (6), 647–658. doi:10.1111/j.1365-2443.2006.00971.x
Fossi, M. C., Casini, S., Marsili, L., Neri, G., Mori, G., Ancora, S., et al. (2002). Biomarkers for endocrine disruptors in three species of Mediterranean large pelagic fish. Mar. Environ. Res. 54 (3–5), 667–671. doi:10.1016/s0141-1136(02)00149-6
García-Reyero, N., Raldúa, D., Quirós, L., Llaveria, G., Cerdà, J., Barceló, D., et al. (2004). Use of vitellogenin mRNA as a biomarker for endocrine disruption in feral and cultured fish. Anal. Bioanal. Chem. 378 (3), 670–675. doi:10.1007/s00216-003-2295-1
Genova, G., Della Chiesa, S., Mimmo, T., Borruso, L., Cesco, S., Tasser, E., et al. (2022). Copper and zinc as a window to past agricultural land-use. J. Hazard. Mat. 424, 126631. doi:10.1016/j.jhazmat.2021.126631
Ghoshal, K., Wang, Y., Sheridan, J. F., and Jacob, S. T. (1998). Metallothionein induction in response to restraint stress. J. Biol. Chem. 273 (43), 27904–27910. doi:10.1074/jbc.273.43.27904
Goksøyr, A. (2006). Endocrine disruptors in the marine environment: Mechanisms of toxicity and their influence on reproductive processes in fish. J. Toxicol. Environ. Health A 69 (1–2), 175–184. doi:10.1080/15287390500259483
Gómez González, N. E., Cabas, I., Rodenas, M. C., Arizcun, M., Mulero, V., and García Ayala, A. (2017). 17α-Ethynylestradiol alters the peritoneal immune response of gilthead seabream. Dev. Comp. Immunol. 76, 143–149. doi:10.1016/j.dci.2017.06.002
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., Amit, I., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 29 (7), 644–652. doi:10.1038/nbt.1883
Harris, C. A., Hamilton, P. B., Runnalls, T. J., Vinciotti, V., Henshaw, A., Hodgson, D., et al. (2011). The consequences of feminization in Breeding groups of wild fish. Environ. Health Perspect. 119 (3), 306–311. doi:10.1289/ehp.1002555
Haunschmid, R., Honsig-Erlenburg, W., Petz-Glechner, R., Schmutz, S., Schotzko, N., Spindler, T., et al. (2006). “Fischbestandsaufnahmen in fließgewässern,” in Methodikhandbuch, Bundesamt für Wasserwirtschaft. Vienna: Bundesministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft Sektion VIIA—1012 Wien.
Hemmerich, W. A. (2016). StatistikGuru: Box-Cox powertransformation berechnen. [Internet]. Available from: https://statistikguru.de/rechner/box-cox.html.
Herrero-Hernández, E., Andrades, M. S., Álvarez-Martín, A., Pose-Juan, E., Rodríguez-Cruz, M. S., and Sánchez-Martín, M. J. (2013). Occurrence of pesticides and some of their degradation products in waters in a Spanish wine region. J. Hydrol. X. 486, 234–245. doi:10.1016/j.jhydrol.2013.01.025
Huang, G. Y., Ying, G. G., Liang, Y. Q., Liu, S. S., and Liu, Y. S. (2014). Expression patterns of metallothionein, cytochrome P450 1A and vitellogenin genes in Western mosquitofish (Gambusia affinis) in response to heavy metals. Ecotoxicol. Environ. Saf. 105 (1), 97–102. doi:10.1016/j.ecoenv.2014.04.012
Hutchinson, T. H., Ankley, G. T., Segner, H., and Tyler, C. R. (2006). Screening and testing for endocrine disruption in fish-biomarkers as “signposts, ” not “traffic lights, ” in risk assessment. Environ. Health Perspect. 114 (Suppl. 1), 106–114. doi:10.1289/ehp.8062
Jelassi, R., Ghemari, C., Khemaissia, H., Raimond, M., Souty-Grosset, C., and Nasri-Ammar, K. (2019). An assessment of copper, zinc and cadmium contamination and their ecotoxicological effects in O. mediterranea Costa, 1853 (Amphipoda, Talitridae). Chem. Ecol. 35 (4), 361–378. doi:10.1080/02757540.2018.1554062
Jobling, S., Beresford, N., Nolan, M., Rodgers-Gray, T., Brighty, G. C., Sumpter, J. P., et al. (2002). Altered sexual maturation and gamete production in wild roach (Rutilus rutilus) living in rivers that receive treated sewage Effluents1. Biol. Reprod. 66 (2), 272–281. doi:10.1095/biolreprod66.2.272
Jobling, S., and Tyler, C. R. (2003). Endocrine disruption in wild freshwater fish. Pure Appl. Chem. 75 (11–12), 2219–2234. doi:10.1351/pac200375112219
Kamunde, C. N., Niyogi, S., and Wood, C. M. (2005). Interaction of dietary sodium chloride and waterborne copper in rainbow trout (Oncorhynchus mykiss): Copper toxicity and sodium and chloride homeostasis. Can. J. Fish. Aquat. Sci. 62 (2), 390–399. doi:10.1139/f04-169
Kim, Y. J., Lee, N., Woo, S., Ryu, J. C., and Yum, S. (2016). Transcriptomic change as evidence for cadmium-induced endocrine disruption in marine fish model of medaka, Oryzias javanicus. Mol. Cell. Toxicol. 12 (4), 409–420. doi:10.1007/s13273-016-0045-7
Klaassen, C. D., Liu, J., and Choudhuri, S. (1999). Metallothionein: An intracellular protein to protect against cadmium toxicity. Annu. Rev. Pharmacol. Toxicol. 39 (1), 267–294. Available from: http://www.annualreviews.org/doi/10.1146/annurev.pharmtox.39.1.267.
Körner, O., Kohno, S., Schönenberger, R., Suter, M. J. F., Knauer, K., Guillette, L. J., et al. (2008). Water temperature and concomitant waterborne ethinylestradiol exposure affects the vitellogenin expression in juvenile Brown trout (Salmo trutta). Aquat. Toxicol. 90 (3), 188–196. doi:10.1016/j.aquatox.2008.08.012
Larsson, D. G. J., Adolfsson-Erici, M., Parkkonen, J., Pettersson, M., Berg, A. H., Olsson, P. E., et al. (1999). Ethinyloestradiol - an undesired fish contraceptive? Aquat. Toxicol. 45 (2–3), 91–97. doi:10.1016/s0166-445x(98)00112-x
Legras, S., Mouneyrac, C., Amiard, J. C., Amiard-Triquet, C., and Rainbow, P. S. (2000). Changes in metallothionein concentrations in response to variation in natural factors (salinity, sex, weight) and metal contamination in crabs from a metal-rich estuary. J. Exp. Mar. Biol. Ecol. 246 (2), 259–279. doi:10.1016/s0022-0981(99)00187-2
Martin, M. B., Reiter, R., Pham, T., Avellanet, Y. R., Camara, J., Lahm, M., et al. (2003). Estrogen-like activity of metals in MCF-7 breast cancer cells. Endocrinology 144 (6), 2425–2436. doi:10.1210/en.2002-221054
Matozzo, V., Gagné, F., Marin, M. G., Ricciardi, F., and Blaise, C. (2008). Vitellogenin as a biomarker of exposure to estrogenic compounds in aquatic invertebrates: A review. Environ. Int. 34 (4), 531–545. doi:10.1016/j.envint.2007.09.008
Melo, M. G., Carqueijo, A., Freitas, A., Barbosa, J., and Silva, A. S. (2020). Modified QuEChERS extraction and HPLC-MS/MS for simultaneous determination of 155 pesticide residues in rice (Oryza sativa L.). Foods 9 (1), 18. doi:10.3390/foods9010018
Moncaleano-Niño, A. M., Barrios-Latorre, S. A., Poloche-Hernández, J. F., Becquet, V., Huet, V., Villamil, L., et al. (2017). Alterations of tissue metallothionein and vitellogenin concentrations in tropical cup oysters (Saccostrea sp.) following short-term (96 h) exposure to cadmium. Aquat. Toxicol. 185, 160–170. doi:10.1016/j.aquatox.2017.02.011
Moran, A. P. (2008). A mathematical theory of animal trapping. Biometrika 38 (3), 307–311. Published by : Biometrika Trust Stable URL: http://www.jstor.org/stable/2332576. doi:10.2307/2332576
Núñez, O., Moyano, E., and Galceran, M. T. (2005). LC-MS/MS analysis of organic toxics in food. TrAC Trends Anal. Chem. 24 (7), 683–703. doi:10.1016/j.trac.2005.04.012
Olsson, P. E., Kling, P., and Hogstrand, C. (1998). “Mechanisms of heavy metal accumulation and toxicity in fish,” in Metal metabolism in aquatic environments. Editors W. J. Langston, and M. J. Bebianno (Boston, MA: Springer), 321–350.
Olsson, P. E., Kling, P., Petterson, C., and Silversand, C. (1995). Interaction of cadmium and oestradiol-17β on metallothionein and vitellogenin synthesis in rainbow trout (Oncorhynchus mykiss). Biochem. J. 307 (1), 197–203. doi:10.1042/bj3070197
Oppen-Berntsen, D. O., Arukwe, A., Yadetie, F., Lorens, J. B., and Male, R. (1999). Salmon eggshell protein expression: A marker for environmental estrogens. Mar. Biotechnol. (NY). 1 (3), 252–260. doi:10.1007/pl00011774
Oppen-Berntsen, D. O., Gram-Jensen, E., and Walther, B. T. (1992). Zona radiata proteins are synthesized by rainbow trout (Oncorhynchus mykiss) hepatocytes in response to oestradiol-17β. J. Endocrinol. 135 (2), 293–302. doi:10.1677/joe.0.1350293
Paquin, P. R., Gorsuch, J. W., Apte, S., Batley, G. E., Bowles, K. C., Campbell, P. G. C., et al. (2002). The biotic ligand model: A historical overview. Comp. Biochem. Physiology Part C Toxicol. Pharmacol. 133 (1–2), 3–35. doi:10.1016/s1532-0456(02)00112-6
Parkkonen, J., Larsson, D. G. J., Adolfsson-Erici, M., Pettersson, M., Berg, A. H., Olsson, P. E., et al. (2000). Contraceptive pill residues in sewage effluent are estrogenic to fish. Mar. Environ. Res. 50 (1–5), 198. doi:10.1016/s0141-1136(00)00181-1
Paterni, I., Granchi, C., Katzenellenbogen, J. A., and Minutolo, F. (2014). Estrogen receptors alpha (ERα) and beta (ERβ): Subtype-selective ligands and clinical potential. Steroids 90, 13–29. doi:10.1016/j.steroids.2014.06.012
Payá, P., Anastassiades, M., MacK, D., Sigalova, I., Tasdelen, B., Oliva, J., et al. (2007). Analysis of pesticide residues using the Quick Easy Cheap Effective Rugged and Safe (QuEChERS) pesticide multiresidue method in combination with gas and liquid chromatography and tandem mass spectrometric detection. Anal. Bioanal. Chem. 389 (6), 1697–1714. doi:10.1007/s00216-007-1610-7
Pelgrom, S. M. G. J., Lock, R. A. C., Balm, P. H. M., and Bonga, S. E. W. (1995). Integrated physiological response of tilapia, Oreochromis mossambicus, to sublethal copper exposure. Aquat. Toxicol. 32 (4), 303–320. doi:10.1016/0166-445x(95)00004-n
Perera, P. A. C. T., Kodithu, S. P., Sundarabarathy, T. V., and Edirisingh, U. (2015). Bioaccumulation of cadmium in freshwater fish: An environmental perspective. Insight Ecol. 4 (1), 1–12. doi:10.5567/ECOLOGY-IK.2015.1.12
Pilehvar, A., Cordery, K. I., Town, R. M., and Blust, R. (2020). The synergistic toxicity of Cd(II) and Cu(II) to zebrafish (Danio rerio): Effect of water hardness. Chemosphere 247, 125942. doi:10.1016/j.chemosphere.2020.125942
Priyam, A., Woodcroft, B. J., Rai, V., Moghul, I., Munagala, A., Ter, F., et al. (2015). Sequenceserver: A modern graphical user interface for custom BLAST databases. Mol. Biol. Evol. 36 (12), 2922–2924. doi:10.1093/molbev/msz185
R Development Core Team (2008). A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. ISBN 3-900051-07-0.
Rhee, J. S., Raisuddin, S., Hwang, D. S., Lee, K. W., Kim, I. C., and Lee, J. S. (2009). Differential expression of metallothionein (MT) gene by trace metals and endocrine-disrupting chemicals in the hermaphroditic mangrove killifish, Kryptolebias marmoratus. Ecotoxicol. Environ. Saf. 72 (1), 206–212. doi:10.1016/j.ecoenv.2008.06.001
Rodgers-Gray, T. P., Jobling, S., Kelly, C., Morris, S., Brighty, G., Waldock, M. J., et al. (2001). Exposure of juvenile roach (Rutilus rutilus) to treated sewage effluent induces dose-dependent and persistent disruption in gonadal duct development. Environ. Sci. Technol. 35 (3), 462–470. doi:10.1021/es001225c
Safe, S. H. (1995). Environmental and dietary estrogens and human health: Is there a problem? Environ. Health Perspect. 103 (4), 346–351. doi:10.1289/ehp.95103346
Saglam, D., Atli, G., and Canli, M. (2013). Investigations on the osmoregulation of freshwater fish (Oreochromis niloticus) following exposures to metals (Cd, Cu) in differing hardness. Ecotoxicol. Environ. Saf. 92, 79–86. doi:10.1016/j.ecoenv.2013.02.020
Schmölz, K., Bottarin, R., Felber, A., Lassacher, F., Lehne, F., Mark, W., et al. (2022). A first attempt at a holistic analysis of various influencing factors on the fish fauna in the Eastern European Alps. Sci. Total Environ. 808, 151886. doi:10.1016/j.scitotenv.2021.151886
Schwaiger, J., and Negele, R. D. (1998). Plasma vitellogenin - a blood parameter to evaluate exposure of fish to xenoestrogens. Acta Vet. Brno 67 (4), 257–264. doi:10.2754/avb199867040257
Sokolova, I. M., Frederich, M., Bagwe, R., Lannig, G., and Sukhotin, A. A. (2012). Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 79, 1–15. [Internet]. doi:10.1016/j.marenvres.2012.04.003
Sonnenschein, C., and Soto, A. M. (1998). An updated review of environmental estrogen and androgen mimics and antagonists. J. Steroid Biochem. Mol. Biol. 65 (1–6), 143–150. doi:10.1016/s0960-0760(98)00027-2
Soverchia, L., Ruggeri, B., Palermo, F., Mosconi, G., Cardinaletti, G., Scortichini, G., et al. (2005). Modulation of vitellogenin synthesis through estrogen receptor beta-1 in goldfish juveniles exposed to 17-β estradiol and nonylphenol. Toxicol. Appl. Pharmacol. 209 (3), 236–243. doi:10.1016/j.taap.2005.04.013
Spurgeon, D. J., Hopkin, S. P., and Jones, D. T. (1994). Effects of cadmium, copper, lead and zinc on growth, reproduction and survival of the earthworm Eisenia fetida (Savigny): Assessing the environmental impact of point-source metal contamination in terrestrial ecosystems. Environ. Pollut. 84 (2), 123–130. doi:10.1016/0269-7491(94)90094-9
Tesoriero, J. V. (1978). Formation of the chorion (zona pellucida) in the teleost, Oryzias latipes. J. Ultrastruct. Res. 64 (3), 315–326. doi:10.1016/s0022-5320(78)90040-0
Vieira, W. T., De Farias, M. B., Spaolonzi, M. P., Da Silva, M. G. C., and Vieira, M. G. A. (2021). Endocrine-disrupting compounds: Occurrence, detection methods, effects and promising treatment pathways - a critical review. J. Environ. Chem. Eng. 9, 104558. doi:10.1016/j.jece.2020.104558
Wagner, F., Mauthner-Weber, R., and Ofenböck, G. (2010). Leitfaden zur Erhebung der biologischen Qualitätselemente. Stubenring, Wien: Bundesministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft Abt. IV/3, Nationale und internationale Wasserwirtschaft, 44.
Werner, J., Wautier, K., Evans, R. E., Baron, C. L., Kidd, K., and Palace, V. (2003). Waterborne ethynylestradiol induces vitellogenin and alters metallothionein expression in lake trout (Salvelinus namaycush). Aquat. Toxicol. 62 (4), 321–328. doi:10.1016/s0166-445x(02)00104-2
Yamaguchi, A., Ishibashi, H., Kohra, S., Arizono, K., and Tominaga, N. (2005). Short-term effects of endocrine-disrupting chemicals on the expression of estrogen-responsive genes in male medaka (Oryzias latipes). Aquat. Toxicol. 72 (3), 239–249. doi:10.1016/j.aquatox.2004.12.011
Yamamoto, F. Y., Garcia, J. R. E., Kupsco, A., and Oliveira Ribeiro, C. A. (2017). Vitellogenin levels and others biomarkers show evidences of endocrine disruption in fish species from Iguaçu River - southern Brazil. Chemosphere 186, 88–99. doi:10.1016/j.chemosphere.2017.07.111
Yang, J., Sun, H., Qian, Y., and Yang, J. (2017). Impairments of cadmium on vitellogenin accumulation in the hepatopancreas of freshwater crab Sinopotamon henanense. Environ. Sci. Pollut. Res. 24 (22), 18160–18167. doi:10.1007/s11356-017-9491-6
Yeo, I. K., and Mugiya, Y. (1997). Effects of extracellular calcium concentrations and calcium antagonists on vitellogenin induction by estradiol-17β in primary hepatocyte culture in the rainbow trout Oncorhynchus mykiss. Gen. Comp. Endocrinol. 105 (3), 294–301. doi:10.1006/gcen.1996.6830
Zheng, S., Chen, B., Wang, Z., Qiu, X., Yu, X., Freestone, D., et al. (2010). Reproductive toxic effects of sublethal cadmium on the marine polychaete Perinereis nuntia. Ecotoxicol. Environ. Saf. 73 (6), 1196–1201. doi:10.1016/j.ecoenv.2010.05.022
Zippin, C. (1956). An evaluation of the removal method of estimating animal populations. Biometrics 12 (2), 163–189. Author ( s ): Calvin Zippin Published by : International Biometric Society Stable URL: http://www.jstor.org/stable/3001759. doi:10.2307/3001759
Keywords: vitellogenin, estrogen receptor, zona pellucida, metallothionein, cadmium, copper
Citation: Niederwanger M, Wieser J, Thaler M, Felber A, Schmölz K, Mark W, Tasser E, Kaufmann R and Pelster B (2022) Expression of biomarkers connected to endocrine disruption in Cottus gobio and Salmo trutta fario in relation to sewage treatment plant-efflux and pesticides. Front. Environ. Sci. 10:1027062. doi: 10.3389/fenvs.2022.1027062
Received: 24 August 2022; Accepted: 27 September 2022;
Published: 11 October 2022.
Edited by:
André Alberto Weber, University of California, San Diego, United StatesReviewed by:
Ralph Thomé, Universidade Federal de São João del-Rei, BrazilThiago Parente, Oswaldo Cruz Foundation (Fiocruz), Brazil
Copyright © 2022 Niederwanger, Wieser, Thaler, Felber, Schmölz, Mark, Tasser, Kaufmann and Pelster. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael Niederwanger, bWljaGFlbC5uaWVkZXJ3YW5nZXJAdWliay5hYy5hdA==