
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Sci., 20 September 2021
Sec. Conservation and Restoration Ecology
Volume 9 - 2021 | https://doi.org/10.3389/fenvs.2021.754286
This article is part of the Research TopicFisheries Ecological Environment in South China SeaView all 18 articles
 Xiangbao Shan1
Xiangbao Shan1 Yulu Shi1Longxiang Fang2,3Yuan Gui1Luchang Xing1Liping Qiu2,3Gengdong Hu1,2,3
Yulu Shi1Longxiang Fang2,3Yuan Gui1Luchang Xing1Liping Qiu2,3Gengdong Hu1,2,3 Jiazhang Chen1,2,3,4*
Jiazhang Chen1,2,3,4*The antibiotics sulfamethoxazole (SMZ) and enrofloxacin (ENF) are commonly used in aquaculture in China, but their effects on the primary productivity of phytoplankton are unclear. On the basis of the fishery drug use standards (SMZ, 100 mg kg−1 d−1; ENF, 15 mg kg−1 d−1), these antibiotics were put into culture ponds to observe their effects on the phytoplankton community and primary productivity. The results showed that the changes in phytoplankton species at SMZ and ENF sites were different. At the ENF sites, Bacillariophyta species increased from 7 to 9, euglenophyta from 8 to 12 and cyanophyta from 11 to 9. Chlorophyta species at the SMZ sites increased from 10 to 15 and euglenophyta from 7 to 4. SMZ and ENF significantly promoted the number of cyanophyta (p < 0.05). The promoting effect of SMZ was more obvious than that of ENF. SMZ was more effective than ENF in promoting cyanophyta. The maximum number of cyanophyta at SMZ sites was 52.39 million L−1, and the concentration of SMZ was positively correlated with the number of cyanophyta. The maximum number of cyanophyta at ENF sites was 33.13 million L−1, and the promoting effect of low concentrations was more significant than that of high concentrations. Both SMZ and ENF promoted increased phytoplankton biomass, consistent with the residual time of the antibiotics in the aquaculture environment. The greatest biomass at the SMZ sites was 129.31 g m−³, and that at the ENF sites was 117.85 g m−³. The changes in the α diversity index showed that both SMZ and ENF led to a decrease in phytoplankton biodiversity, and that SMZ was more harmful to it. There were significant differences in the Shannon–Wiener, Pielou and Simpson indexes of the SMZ sites within groups (p < 0.05). The β diversity index showed that both antibiotics could change the phytoplankton habitat, but the effect of ENF on the habitat was recoverable, while that of SMZ prevented its restoration. These data will be valuable in protecting the ecological environment of fisheries and ensuring the safety and stability of fishery aquatic ecosystems.
In recent years, antibiotics have been widely used in the prevention and treatment of a variety of bacterial diseases (Bialk-Bielinska et al., 2011; Johnson et al., 2015). However, antibiotics are usually not completely metabolized (Zheng et al., 2017). Some of them are excreted into the culture environment through the urine and feces of aquatic animals in the form of crude drugs or metabolites (Aks et al., 2006; Gao et al., 2018). The antibiotics sulfamethoxazole (SMZ) and enrofloxacin (ENF) have been found most often (Xu et al., 2006; Yan et al., 2020), and their residues in the greatest amounts in water (Zhang et al., 2019). The release of residual antibiotics into the environment may affect the phytoplankton community and endanger the safety and stability of the ecosystem.
Algae are an important part of aquatic ecosystems as primary producers, providing oxygen and organic matter to other aquatic organisms through photosynthesis (Laurens et al., 2017; Farooqui et al., 2021). Changes in the number of algae may cause changes in the dominant species and biomass of the community, affect the stability and complexity of the community structure, lead to habitat changes and even harm the structure and function of the whole ecosystem (Rakowski and Cardinale, 2016). Algae are also an important indicator in an environmental quality assessment of an aquatic ecosystem, and so helpful in a comprehensive and effective evaluation of the comprehensive effects of pollutants on phytoplankton and the whole aquatic ecosystem (Li et al., 2008; Khalil et al., 2021). The diversity and complexity of aquatic ecosystems can be effectively understood by observing the species composition and quantitative changes in algae.
α and β diversity indexes are commonly used to describe the diversity of a community. The α diversity index is mainly concerned with the number of species in local and uniform habitats, so describes within-habitat diversity. The β diversity index refers to the differences in species composition or the replacement rate of species along the environmental gradient between different habitat communities, and so describes between-habitat diversity, and emphasizes the change in species diversity within the community (Baczkowski et al., 1998). The diversity index is positively correlated with the number of species. If a region is rich in species, the value of the diversity index is high. If the region is disturbed by external forces, resulting in a reduction in the number of organisms and in the ecological positions of the species remaining, it is easier for other species to replace them and so to maintain the stability and balance of the ecosystem (Peng et al., 2020).
The algae growth inhibition test is one of the most common aquatic ecological toxicity tests. Significant differences in the effects of different antibiotics have been found in the physiological and biochemical indexes of different algae (Aderemi et al., 2018; Fu et al., 2017). Barsha Roy (Br et al., 2020) reported that different concentrations of tetracycline inhibited the growth of Scenedesmus obliquus, and (González-Pleiter et al., 2019a) found that Microcystis was highly sensitive to quinolones (ciprofloxacin). The characteristics of the algae community in the culture environment of the shallow lakes (Peng et al., 2021), the marine (Chong et al., 2020) and the river (Tian et al., 2021) have been reported in the literature, but its structure and the potential risk of SMZ and ENF in the culture environment are not clear. There is an urgent need for research into the risk these antibiotics pose to product and environmental quality. We studied the effects of SMZ and ENF on primary productivity of phytoplankton in the culture environment, to obtain basic experimental data and a theoretical basis to promote the sustainable development of fisheries and to protect the ecological environment.
Seven tilapia cultivation ponds with the same conditions were selected (Table 1). One blank control site (A), three ENF sites (B, C, D) and three SMZ sites (E, F, G) were set up. According to the “Guidelines for the use of fishery drugs” (Ministry of Agriculture of the People’s Republic of China, NY5071-2002) and the “Code for the use of Enrofloxacin in aquaculture” (Ministry of Agriculture of the People’s Republic of China, SC/T1083-2007), SMZ (100 mg kg−1 d−1) and ENF (15 mg kg−1 d−1) were fed for 5 consecutive days, once a day for SMZ and twice a day for ENF (Mix the material feeding). According to the national drug use standard, the SMZ withdrawal period was 30 days and the ENF withdrawal period was 10 days. Tests at the SMZ sites lasted for 40 days and at the ENF sites they lasted for 20 days. Eight samples of aquaculture water, sediment, tilapia muscle, liver and intestinal contents were collected from each pond at 1, 6, 10, 15, 20, 25, 32 and 40 days. The first sample was the blank, and the second sampling time was the second day after continuous feeding for 5 days. At the same time, to ensure the samples representative, samples of culture water, sediment and tilapia in each pond were collected by the five-point method. The muscle, liver and intestinal contents were separated and packed into plastic food bags and transported to the laboratory where they were frozen at −20°C.
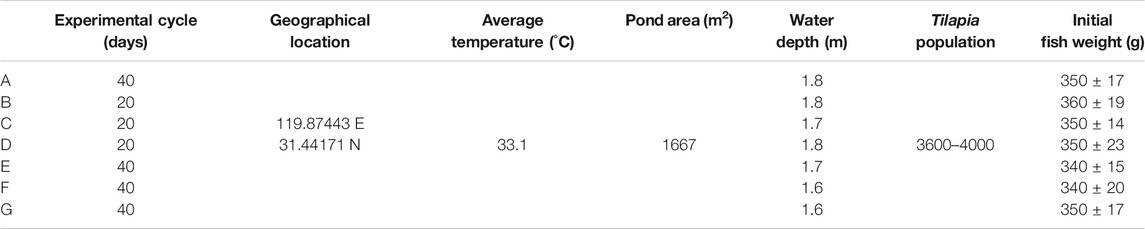
TABLE 1. Basic information of Tilapia pond.
The qualitative samples were collected with a No. 25 plankton net and were taken back to the laboratory for observation and classification under a 10 × 40 optical microscope (BM2000, Jiangnan, China). A quantitative sample was collected in a 1000 ml plexiglass water collector; After standing for 24 h, 30 ml was taken for microscopic examination. The taxonomy of the phytoplankton was recorded according to the Freshwater biota of China (ISBN:7-5027-3729-4).
The phytoplankton counting method was obtained according to the formula as follows:
In the formula: M is the number of phytoplankton in 1 L of water; C is the area of the counter box, the unit is mm2; FS is the area per field of view, the unit is mm2; FN is the number of fields passed per slice count; PN is the number of phytoplankton actually counted by counting each sheet.
The content of Chlorophyll a was determined by the 90% acetone method to calculate the cumulative biomass of phytoplankton. 200 ml of the water sample was filtered and were refrigerated (−4°C) for more than 12 h. We added 3–4 ml 90% acetone solution, used a homogenate machinea (Pro200, United States ) to break the filter paper, then centrifuged (2–16P, Sartorius, Germany) the tube for 10 min at 2460×g, Continued to volume with 90% acetone to 10 ml. The absorbance values were measured at 630, 645, 663 and 750 wavelengths by spectrophotometer (MCL8, Shangai).
The biomass was recorded and calculated, the formula as follows:
In the formula: B is the amount of chlorophyll A (mg/m3); V is water volume (ml); D is the absorbance value at different wavelengths; V1 is constant volume (ml); δ is the cuvette thickness.
Aquaculture water sample: 200 ml samples were taken for extraction and concentration. The HLB solid phase extraction column (ENF:60 mg/3 ml, Anpel, Shanghai; SMZ:500 mg/6ml, Anpel, Shanghai) was selected and activated with 5 ml methanol and 5 ml ultra pure water successively. Then the water sample was added. After the water sample was drained, 5 ml methanol was used for two times (3 ml + 2 ml) for elution, and 10 ml centrifuge tube was used to collect the elution. The eluent was diluted with methanol to a certain scale. After shaking well, part of the eluent was taken through a 0.22 μm aperture filter membrane and transferred to the sample bottle for testing.
Sediment samples: 2 ± 0.01 g sample was weighed in a 50 ml centrifuge tube, 10 ml 0.1% formic acid acetonitrile was added and mixed by shaking. The sample was rotated at 2000 r min−1 for 10 min (Henry Troemner, United States ), and centrifuged at 10680×g for 5 min (GL-22MS, Bioridge, Shanghai). The supernatant of 5 ml after centrifugation was added to the enhanced lipid removal purification tube activated by 5 ml ultra pure water, and then was rotated at 2000 r min−1 for 5 min and centrifuged at 2460×g for 5 min. After centrifugation, the supernatant was transferred to the Bond Elu EMR-Lipid tube (Agilent) containing 1.7 g MgSO4, which was vibrated at 2000 r min−1 for 5 min, and centrifuged at 2460×g for 5 min. The upper organic phase was passed through a 0.22 μm aperture filter membrane and transferred to a sample bottle for testing.
An ultra performance liquid chromatography tandem mass spectrometry (LC-MS/MS, Agilent 6420, United States ) was used to analyze the two selected antibiotics. Standard curves were plotted with the gradient concentrations of 1, 5, 10, 20 and 50 μg L−1. The regression coefficients of SMZ and ENF were all greater than 0.998 and 0.9996, respectively. The recovery rates of SMZ and ENF were 78.3–98.4% and 81–112% respectively. The limits of detection and quantification were less than 0.01 μg L−1 for sulfamethoxazole and 0.2 μg L−1 for enrofloxacin.
The biodiversity index is used to express the relationship between the number and species of mixed biological communities composed of many kinds of organisms, and reflects the complexity of biological communities or habitats (Diaz et al., 2020). The effects of SMZ and ENF on the species and quantity of phytoplankton were analyzed by α and β diversity indexes, and the ecological characteristics of phytoplankton in the culture environment were evaluated comprehensively (Ers et al., 2020).
The α diversity index, including Shannon–Wiener index (H), Pielou index (E), Simpson’s diversity index (C) and Margalef richness index (D). The β diversity index included the Jaccard index (CJ). The calculation formulas and evaluation criteria of the above indexes are shown in Table 2.

TABLE 2. Formula principle and evaluation criteria of α diversity index and β diversity index.
The data were analyzed by Microsoft Excel 2013 (United States ) and SPSS 25 software (Chicago, United States ), and plotted by Origin 2021 (Northampton, United States ) function and Heml Heatmap drawing software (United States ).
We found 109 species in the eight phyla Chlorophyta, Cyanophyta, Bacillariophyta, Euglenophyta, Cryptophyta, Pyrrophyta, Xanthophyta and Chrysophyta. The species composition of the phytoplankton is shown in Table 3. Among them were 38 Chlorophyta species, accounting for 34.9% of the total phytoplankton species; Euglenophyta were second with 26 species, accounting for 23.9% of the total phytoplankton species. The numbers of species from other phyla were 18, 17, 4, 4, 1 and 1 in Cyanophyta, Bacillariophyta, Cryptophyta, Dinoflagellata, Chrysophyta and Xanthophyta, respectively. The number of algae species at the SMZ sites was similar to that at the ENF sites. We found 101 species of algae at the SMZ sites, including 36 species of Chlorophyta, 23 species of Euglenophyta, 17 species of Bacillariophyta and 15 species of Cyanophyta. The ENF sites held 98 species of algae, including 34 species of Chlorophyta, 24 species of Euglenophyta, 15 species of Bacillariophyta and 15 species of Cyanophyta, and the species number of cryptomophyta, dinoflagellata, chrysophyta and xanthophyta remained the same at both sites. Four phytoplankton species related to the two antibiotic sites were 4, 4, 1 and 1, respectively.
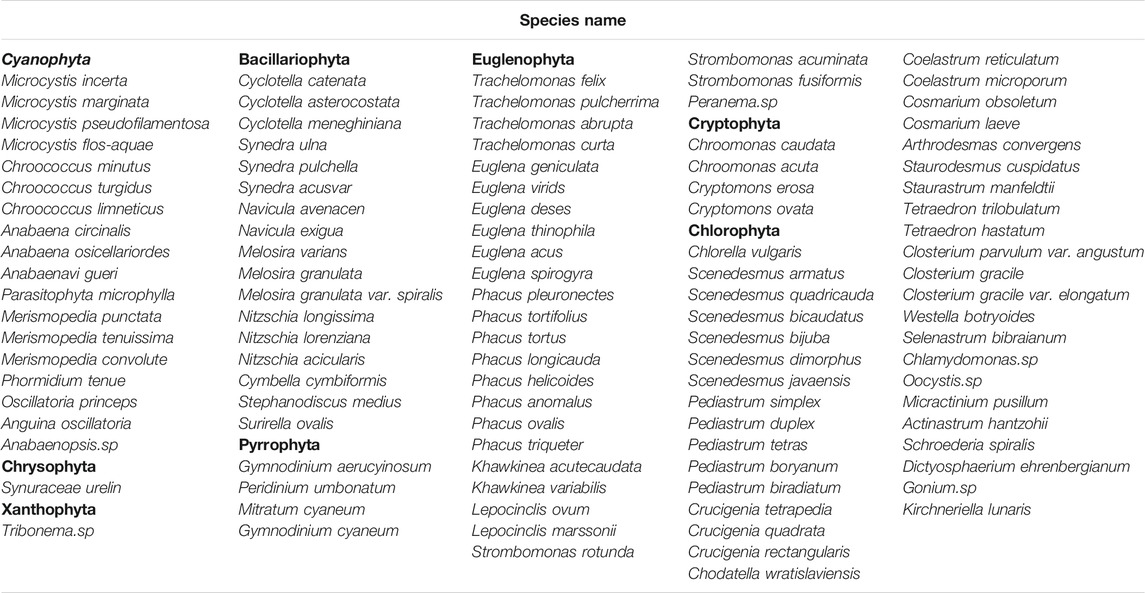
TABLE 3. Phytoplankton species composition in a typical tilapia pond.
After SMZ and ENF entered the aquaculture environment, there were significant differences in the number of phytoplankton species of Cyanophyta, Chlorophyta, Bacillariophyta and Euglenophyta (Figure 1). After adding ENF for 5 days, the Bacillariophyta and Euglenophyta species increased on the 6th day, the number of Bacillariophyta species increased from 7 to 9 and returned to their initial levels on the 10th day, while the number of Euglenophyta species increased from 8 to 12 on the 15th day. This indicated that ENF promoted the growth of Bacillariophyta and Euglenophyta and was positively correlated with them. In the early stage of ENF dosage there was no significant effect on Cyanophyta species, but the number of Cyanophyta species decreased from 11 to 9 on the 10th day as antibiotic concentrations fell. ENF had no significant effect on Chlorophyta species. The effect of SMZ was opposite to that of ENF. We found that Cyanophyta and Bacillariophyta species remained at 10 and 7, respectively, with no significant change. The number of Chlorophyta species increased from 10 species on day 1–15 species on day 25, then declined. SMZ inhibited the growth of Euglenophyta, and the number of Euglenophyta decreased from seven species on day 1–4 species on day 10.
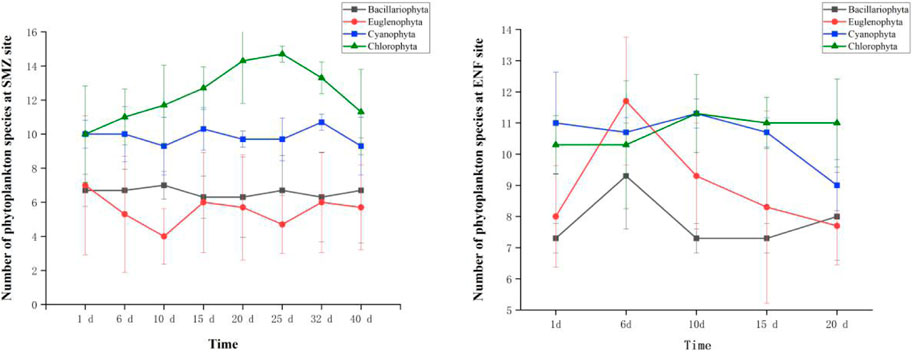
FIGURE 1. Effects of sulfamethoxazole and enrofloxacin on phytoplankton species.
There were significant differences between the two antibiotics on the number of Cyanophyta compared with Bacillariophyta, Chlorophyta and Euglenophyta. The main results were: both SMZ and ENF greatly increased the number of Cyanophyta. The promoting effect of SMZ was more obvious than that of ENF, and the promoting effect of antibiotics on Cyanophyta was related to its concentration in the culture environment. Taking into account the difference in the initial habitat of each pond, pond A was selected as a blank group without any treatment. The results showed that the numbers of four species of algae in the blank site remained stable throughout the experimental period, and the number of Cyanophyta was stable at 7–11 million L−1 (Figure 2A). The number of Cyanophyta at the SMZ sites almost doubled and was 25.54 million L−1 on the first day. The number began to decrease after increasing to 52.39 million L−1 on the sixth day, and fell to its lowest value on the 20th day. At this time, the number of Cyanophyta stabilized at about 12 million L−1, which was half of the initial level. The number of Cyanophyta at the ENF sites almost doubled, from 16.31 million L−1 on the first day to a peak of 33.13 million L−1 on the tenth day, then began to decrease slowly and returned to the initial level on the 20th day.
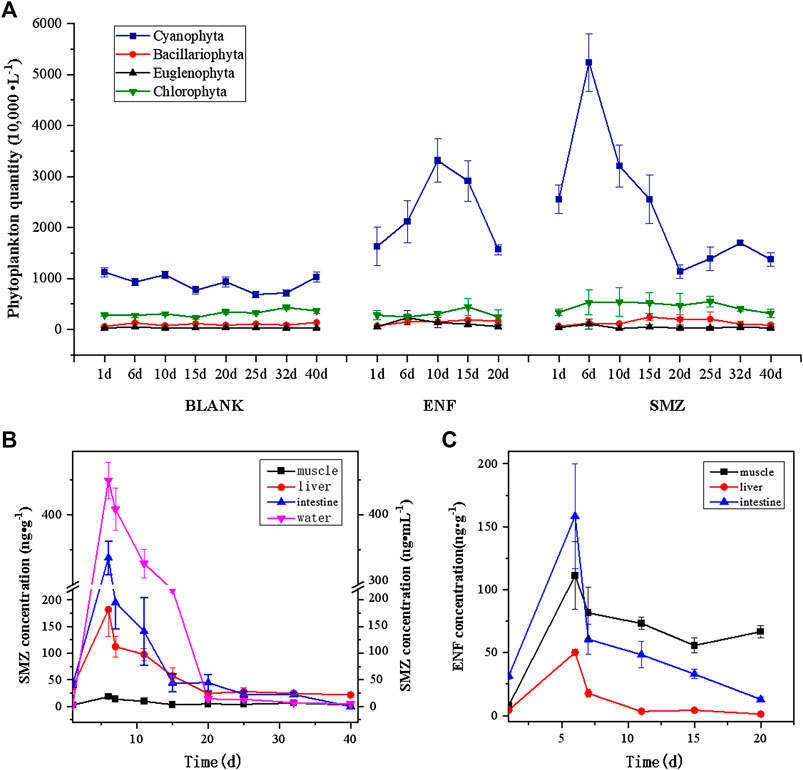
FIGURE 2. Effects of sulfamethoxazole and enrofloxacinon on phytoplankton population changes.
The concentrations of the two antibiotics in different media in the aquaculture environment varied with time (Figures 2B,C). The concentration of antibiotics was its highest on the sixth day after feeding. SMZ was completely degraded in tilapia after about 20 days, and ENF was completely degraded in tilapia after about 10–12 days. Analysis of specific algae species revealed that the sites tested with the two antibiotics were mainly characterized by the outbreak and growth of Anabaena circinalis; Chroococcus turgidus; Chroococcus limneticus; Merismopedia punctata; Merismopedia tenuissima and Merismopedia convolute, but there were differences. The number of Cyanophyta at the SMZ sites was at its highest on the sixth day, and the concentration of SMZ was the highest at this time; while the number of Cyanophyta at the ENF sites was highest on the tenth day, and the withdrawal period of ENF was 10 days. The results showed that there was a positive correlation between the concentration of SMZ and the number of Cyanophyta; that is, the higher the concentration of SMZ, the better its promoting effect on Cyanophyta. Although ENF was also able to increase the number of Cyanophyta, the effect of low concentrations was more significant than that of high concentrations.
The biomass of phytoplankton changed with the alterations in species and quantity. Compared with the blank site, the phytoplankton biomass at the SMZ and ENF sites increased significantly, and the cumulative biomass at the SMZ sites was its highest on the 20th day, with the highest value of 129.31 g/m³ (Figure 3). The cumulative biomass at the ENF sites reached its highest value of 117.85 g/m³ on the tenth day, and then began to decrease. The phytoplankton biomass on the 20th day at the ENF sites was 63.77 g/m³, which was close to the initial level (62.49 g/m³), but the phytoplankton biomass at the 40th day at the SMZ sites was 91.91 g/m³, which was different from the initial level (63.28 g/m³). The change in phytoplankton biomass was consistent with the residual time of antibiotics in the culture environment, and the cumulative biomass increased because antibiotics greatly promoted the growth of Cyanophyta in the culture environment. When the promoting effect of antibiotics on Cyanophyta disappeared after their complete degradation, the cumulative biomass of phytoplankton decreased.

FIGURE 3. Effects of sulfamethoxazole and enrofloxacinon on phytoplankton biomass.
After SMZ and ENF were fed into the culture environment, the α diversity index of the phytoplankton decreased (Table 4). The higher the diversity index, the richer the biodiversity of the species, and vice versa. The results showed that both SMZ and ENF were able to reduce the biodiversity of phytoplankton. At the same time, we found that the values of the Shannon–Wiener, Margalef richness and Pielou indexes at the SMZ sites were lower than those at the ENF sites, which indicated that SMZ was more harmful to phytoplankton biodiversity. Moreover, the values of the three indexes at the SMZ sites were their lowest on the sixth day; the lowest values were 2.32 ± 0.43, 3.93 ± 0.92 and 0.65 ± 0.09, respectively. The lowest values at the ENF sites, however, were 2.67 ± 0.14, 5.03 ± 0.98 and 0.71 ± 0.04, respectively, on the 10th and 15th days, which was consistent with the maximum number of Cyanophyta. Analysis of α diversity index showed that the Shannon–Wiener, Pielou and Simpson diversity indexes at the SMZ sites were significant different within groups (p < 0.05), but there was no significant difference in each index at the ENF sites. The difference between SMZ groups verified the conclusion that the antibiotics aggravated the degree of water pollution by changing the structure of the algae community. According to the evaluation criteria of the α diversity index, SMZ will exacerbate the degree of water pollution of culture ponds, and many indexes show that the water quality of culture ponds changes from mild pollution to moderate pollution after SMZ enters the culture environment. ENF does not intensify the degree of water pollution of culture ponds, and ENF sites always maintain mild levels of pollution. When the Simpson diversity index of the SMZ sites exceeded 0.1 on the 6th and 11th days, the Simpson diversity index of the ENF sites exceeded 0.1 on the 10th day, which was consistent with the time when the number of Cyanophyta reached their peak at both the SMZ and the ENF sites. At this time, Cyanophyta was absolutely dominant in phytoplankton, other species with narrow nutritional niches had died out, the community structure tended to be simple, and both stability and biodiversity decreased.
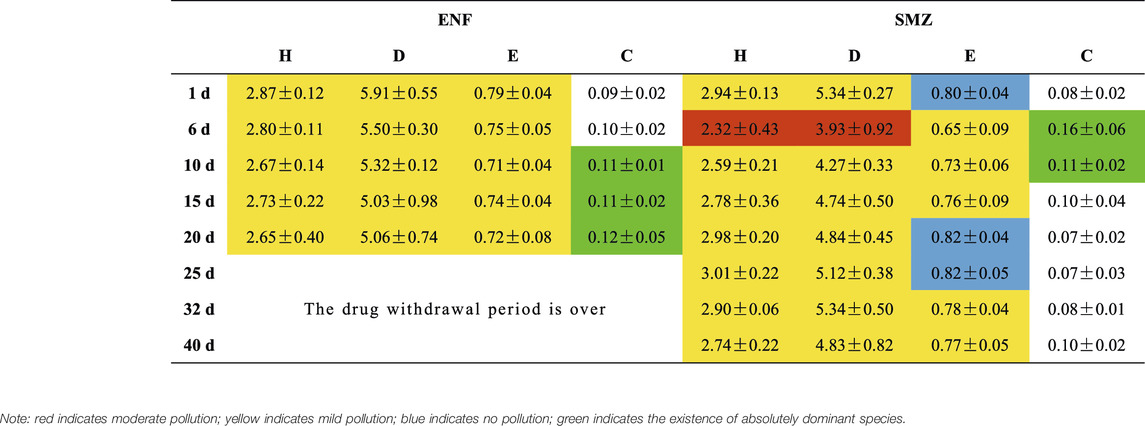
TABLE 4. α diversity index of culture water at SMZ site and ENF site.
The habitats of phytoplankton communities at different times at the sites tested with the two antibiotics are shown in Figure 4. Compared with the blank site, the comprehensive Jaccard index of the SMZ and ENF sites on the sixth day was 0.30–0.46 (mild similarity); that of the ENF sites increased to 0.60–0.75 with the passage of time (moderate similarity), while that of the SMZ sites was still 0.30–0.50. The results showed that both antibiotics could affect the community structure of phytoplankton and change the habitat, but that the community could recover from the effects of ENF. The Jaccard index value increased gradually with the passage of time, remaining unchanged until the end of the recommended withdrawal period. The changes in habitat and biomass of phytoplankton at the SMZ and ENF sites were consistent. At the ENF sites they returned to their initial levels. Those at the SMZ sites, however, could not be completely restored and the phytoplankton community structure changed, so the cumulative biomass was different from the initial level.
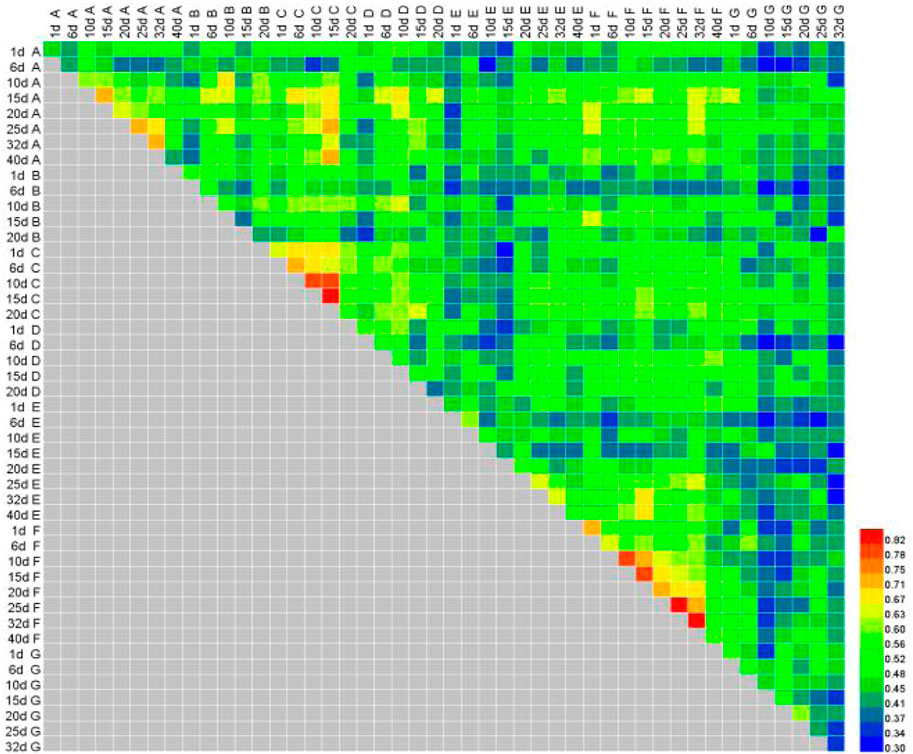
FIGURE 4. Effects of sulfamethoxazole and enrofloxacinon on habitat of culture ponds.
The aquaculture industry has long been plagued by a variety of bacterial diseases. The emergence of antibiotics to avoid the impact of such diseases on aquaculture species can effectively improve the economies of fisheries (Ming et al., 2020). Their widespread use, however, has also caused many environmental problems. These include the phenomenon of antibiotic residues in the culture environment, which is becoming increasingly serious (Zhao et al., 2019). Attention is often only paid to the food health and safety problems caused by the residues of antibiotics in the cultured species, and their harmful effects on the culture environment is ignored (Ana et al.,1987). Phytoplankton are the cornerstones of aquatic ecosystems but, at present, research into the effects of antibiotics on phytoplankton primary productivity in culture environments has not yielded clear results.
We found that both SMZ and ENF can affect the primary productivity of phytoplankton to different degrees. Change in primary productivity was mainly due to the more obvious increase in the number of Cyanophyta than of other phytoplankton. The reason for this phenomenon may be that neither SMZ nor ENF can induce the formation of reactive oxygen species (ROS) (Ling et al., 2015). Chenshan (Shan et al.) found that ROS is an intracellular by-product of aerobic metabolism, and that the toxicity of antibiotics is related to the increase in ROS formation at an early stage. The formation of ROS and the imbalance in cellular antioxidant defense mechanisms will lead to oxidative stress and to protein, lipid and DNA damage, so SMZ and ENF have no inhibitory effect on cyanobacterial cells (Gomes et al., 2016; Mullineaux et al., 2018). EC50 (the median effective toxicity concentration with a 50% effect in the blank group) is a standardized parameter for evaluating different compounds to clarify their toxicity (González-Pleiter et al., 2019b) found that the EC50 value of ENF in Microcystis aeruginosa cells was lower than 1 mg L−1, indicating that, at the current environmental level, the antibiotic may lead to its ecological risk being classified as “toxic to aquatic organisms”. The EC50 values of SMZ and other antibiotics are 0.985–630 mg L−1, which are higher than those of quinolones, macrolides and tetracyclines (Rajaniemi et al., 2005). Therefore, it is an antibiotic with less cytotoxicity to cyanobacteria. Our results showed that the number of cyanobacteria at the SMZ sites was higher than at ENF sites, which was consistent with the experimental results.
After SMZ and ENF entered the aquaculture water body, the dominant species of the algae community changed. This affected the community structure, caused the habitat to change and finally led to a change in the nutritional status of the water body. There is a certain correlation between the dominant population and the nutritional status of the water body (Wyta et al.): for example, the appearance of a large number of Chrysophyta often reflects that the water body is Oligotrophic; and Cyanophyta, Chlorophyta and Chlorophyta indicate a eutrophic water body (Dokulil et al., 2003; Parus and Karbowska, 2020). The results showed that cyanobacteria comprised the dominant community at SMZ and ENF sites, and we inferred that aquaculture water using SMZ and ENF was strongly eutrophic. At the same time, and by evaluating the water bodies by indicator species, the emergence of a large number of Anabaena (Rajaniemi et al., 2005) species at the initial stage of SMZ and ENF entry led to a deterioration in water quality. Water quality improved with the natural degradation of the antibiotics and the self-purification capacity of indicator species such as Scenedesmus (Rosas et al., 1993) and Spirulina (Liu et al., 2018). The α and β diversity indexes showed that the biodiversity of the phytoplankton community decreased and the water quality deteriorated at the sites tested with the two antibiotics, which was consistent with the evaluation results of indicator species.
SMZ and ENF have significant–and different–effects on the primary productivity of phytoplankton in the culture environment. These two antibiotics cause water pollution by affecting the species and quantity of phytoplankton and changing the dominant species and community structure. The degree of water pollution caused by SMZ was higher than that of ENF, and the habitat change caused by ENF was not restored during the drug withdrawal period. This endangers the living environments of aquatic organisms and increases the dietary risk from fishery products. Our test results and actual production management needs indicate that we should pay more attention to, and better manage, these two antibiotics in the fishery culture environment. This will help in reducing the environmental risk caused by drug residues, and protect the fishery ecological environment, as well as ensuring the quality and safety of aquatic products.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethical review and approval was not required for the animal study because Tilapia is a cultured species.
XS designed and completed experiments, wrote articles, etc. Others offered help and participated in the experiments.
The authors acknowledge the financial support provided by National Key R&D Program of China (2017YFC1600704), National Natural Science Foundation of China (31802271) and Young Talents Project of Jiangsu Association for Science and Technology.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aderemi, A. O., Novais, S. C., Lemos, M. F. L., Alves, L. M., Hunter, C., and Pahl, O. (2018). Oxidative Stress Responses and Cellular Energy Allocation Changes in Microalgae Following Exposure to Widely Used Human Antibiotics. Aquat. Toxicol. 203, 130–139. doi:10.1016/j.aquatox.2018.08.008
Aks, A., Mtm, B., and Ab, C. (2006). A Global Perspective on the Use, Sales, Exposure Pathways, Occurrence, Fate and Effects of Veterinary Antibiotics (VAs) in the Environment - ScienceDirect. Chemosphere 65 (5), 725–759. doi:10.1016/j.chemosphere.2006.03.026
Ana, K., Madriaga, J., and Espino, M. P. (1987). β-Lactam Antibiotics and Antibiotic Resistance in Asian Lakes and Rivers: An Overview of Contamination, Sources and Detection Methods. Environ. Pollut. (Barking, Essex 275, 116624.
Baczkowski, A. J., Joanes, D. N., and Shamia, G. M. (1998). Range of Validity of α and β for a Generalized Diversity index H(α,β) Due to Good. Math. Biosciences 148 (2), 115–128. doi:10.1016/s0025-5564(97)10013-x
Bialk-Bielinska, A., Stolte, S., Aming, J., Ue Be Rs, U., Boschen, A., Stepnowski, P., and Matzke, M. (2011). Ecotoxicity Evaluation of Selected Sulfonamides. Chemosphere 85 (6), 928–933. doi:10.1016/j.chemosphere.2011.06.058
Br, A., Pks, A., Nc, B., and Am, B. (2020). Antibiotic Tetracycline Enhanced the Toxic Potential of Photo Catalytically Active P25 Titanium Dioxide Nanoparticles towards Freshwater Algae Scenedesmus Obliquus. Chemosphere.
Chong, B. W. K., Leong, S. C. Y., Kuwahara, V. S., and Yoshida, T. (2020). Monsoonal Variation of the marine Phytoplankton Community in Kota Kinabalu, Sabah. Reg. Stud. Mar. Sci. 37, 101326. doi:10.1016/j.rsma.2020.101326
Díaz, M., Concepción, E. D., Oviedo, J. L., Caparrós, A., Farizo, B. Á., and Campos, P. (2020). A Comprehensive index for Threatened Biodiversity Valuation. Ecol. Indicators 108 (Jan.), 105696–105691. doi:10.1016/j.ecolind.2019.105696
Dokulil, M. T., Markert, B. A., Breure, A. M., and Zechmeister, H. G. (2003). Algae As Ecological Bio-Indicators: Algae as Ecological Bio-Indicators. Amsterdam, Netherlands: Elsevier.
Ers, T., Czeglédi, I., Tóth, R., and Schmera, D. (2020). Multiple Stressor Effects on Alpha, Beta and Zeta Diversity of Riverine Fish. Sci. Total Environ. 748, 141407. doi:10.1016/s0048-9697(20)36593-1
Farooqui, A., Tripathi, G., Moheet, K., Dubey, P., and Mahfooz, S. (2021). Algal Biomass: Potential Renewable Feedstock For Bioenergy Production: Bioenergy Research: Integrative Solution for Existing Roadblock.
Fu, L., Huang, T., Wang, S., Wang, X., Su, L., Li, C., et al. (2017). Toxicity of 13 Different Antibiotics Towards Freshwater Green Algae Pseudokirchneriella Subcapitata and Their Modes of Action - ScienceDirect. Chemosphere 168, 217–222.
Ling, F., Hamzeh, M., Dodard, S., Zhao, Y. H., and Sunahara, G. I. (2015). Effects of TiO2 Nanoparticles on ROS Production and Growth Inhibition Using Freshwater green Algae Pre-exposed to UV Irradiation. Environ. Toxicol. Pharmacol. 39 (3), 1074–1080. doi:10.1016/j.etap.2015.03.015
Ling, F., Haung, T., Wang, S., Wang, X., Su, L., and Li, C. (2017). Toxicity of 13 Different Antibiotics towards Freshwater green Algae Pseudokirchneriella Subcapitata and Their Modes of Action - ScienceDirect. Chemosphere 168, 217–222.
Gao, N., Liu, C. X., Xu, Q. M., Cheng, J. S., and Yuan, Y. J. (2018). Simultaneous Removal of Ciprofloxacin, Norfloxacin, Sulfamethoxazole by Co-producing Oxidative Enzymes System of Phanerochaete Chrysosporium and Pycnoporus Sanguineus. Amsterdam, Netherlands: CHEMOSPHERE -OXFORD-.
Gomes, T., Xie, L., Brede, D., Lind, O. C., Solhaug, K. A., Salbu, B., and Tollefsen, K. E. (2016). Sensitivity of the green Algae Chlamydomonas Reinhardtii to Gamma Radiation: Photosynthetic Performance and ROS Formation. Aquat. Toxicol. 183, 1–10. doi:10.1016/j.aquatox.2016.12.001
González-Pleiter, M., a, S. C., Hurtado-Gallego, J., Leganés, F., Fernández-Pias, F., and Velázquez, D. (2019b). Ecotoxicological Assessment of Antibiotics in Freshwater Using Cyanobacteria - ScienceDirect. Amsterdam, Netherlands: Elsevier
González-Pleiter, M., Cirés, S., Hurtado-Gallego, J., Leganés, F., Fernández-Piñas, F., and Velázquez, D. (2019a). Ecotoxicological Assessment of Antibiotics in Freshwater Using Cyanobacteria. Cyanobacteria, 399–417. doi:10.1016/b978-0-12-814667-5.00020-9
Johnson, A. C., Keller, V., Dumont, E., and Sumpter, J. P. (2015). Assessing the Concentrations and Risks of Toxicity from the Antibiotics Ciprofloxacin, Sulfamethoxazole, Trimethoprim and Erythromycin in European Rivers. Sci. Total Environ. 511 (apr.1), 747–755. doi:10.1016/j.scitotenv.2014.12.055
Khalil, S., Mahnashi, M. H., Hussain, M., Zafar, N., and Irfan, M. (2021). Exploration and Determination of Algal Role as Bioindicator to Evaluate Water Quality – Probing Fresh Water Algae. Saudi J. Biol. Sci. 1.
Laurens, L. M. L., Chen-Glasser, M., and Mcmillan, J. D. (2017). A Perspective on Renewable Bioenergy from Photosynthetic Algae as Feedstock for Biofuels and Bioproducts. Algal Res. 24, 261–264. doi:10.1016/j.algal.2017.04.002
Li, Y., Horsman, M., Wang, B., Wu, N., and Lan, C. Q. (2008). Effects of Nitrogen Sources on Cell Growth and Lipid Accumulation of green Alga Neochloris Oleoabundans. Appl. Microbiol. Biotechnol. 81 (4), 629–636. doi:10.1007/s00253-008-1681-1
Liu, L., Champagne, P., and Hall, G. (2018). “The Role of Algae in the Removal and Inactivation of Pathogenic Indicator Organisms in Wastewater Stabilization Pond Systems.” in Paper presented at the IWA SWWS2018 15th Conference on Small Water & Wastewater Systems and 7th Conference on Resources Oriented Sanitation, Haifa, Israel, October 2018.
Ming, Z. A., Zc, A., Gz, A., Yin, Z. A., Nxb, C., and Dz, A. (2020). Effectively Reducing Antibiotic Contamination and Resistance in Fishery by Efficient Gastrointestine-Blood Delivering Dietary Millispheres. J. Hazard. Mat. 409, 1–11. doi:10.1016/j.jhazmat.2020.125012
Mullineaux, P. M., Exposito-Rodriguez, M., Laissue, P. P., and Smirnoff, N. (2018). ROS-dependent Signalling Pathways in Plants and Algae Exposed to High Light: Comparisons with Other Eukaryotes. Free Rad Biol. Med. doi:10.1016/j.freeradbiomed.2018.01.033
Parus, A., and Karbowska, B. (2020). Marine Algae as Natural Indicator of Environmental Cleanliness. Water Air Soil Pollut. 231 (3), 1–8. doi:10.1007/s11270-020-4434-0
Peng, G., Zhou, X., Xie, B., Huang, C., Uddin, M. M., Chen, X., and Huang, L. (2020). Ecosystem Stability and Water Quality Improvement in a Eutrophic Shallow lake via Long-Term Integrated Biomanipulation in Southeast China - ScienceDirect. Ecol. Eng.
Peng, X., Zhang, L., Li, Y., Lin, Q., He, C., Huang, S., Li, H., Zhang, X., Liu, B., Ge, F., Zhou, Q., Zhang, Y., and Wu, Z. (2021). The Changing Characteristics of Phytoplankton Community and Biomass in Subtropical Shallow Lakes: Coupling Effects of Land Use Patterns and lake Morphology. Water Res. 200 (1), 117235. doi:10.1016/j.watres.2021.117235
Rajaniemi, P., Hrouzek, P., Kaštovská, K., Willame, R., Rantala, A., Hoffmann, L., Komárek, J., and Sivonen, K. (2005). Phylogenetic and Morphological Evaluation of the Genera Anabaena, Aphanizomenon, Trichormus and Nostoc (Nostocales, Cyanobacteria). Int. J. Syst. Evol. Microbiol. 55 (Pt 1), 11–26. doi:10.1099/ijs.0.63276-0
Rakowski, C., and Cardinale, B. J. (2016). Herbivores Control Effects of Algal Species Richness on Community Biomass and Stability in a Laboratory Microcosm experiment. Oikos 125, 1627–1635. doi:10.1111/oik.03105
Rosas, I., Velasco, A., Belmont, R., Báez, A., and Martínez, A. (1993). The Algal Community as an Indicator of the Trophic Status of Lake Patzcuaro, Mexico. Environ. Pollut. 80 (3), 255–264. doi:10.1016/0269-7491(93)90046-q
Shan, C. A., Wei, Z. A., Jl, A., My, A., Jz, A., Fan, X. A., and Lw, A. Ecotoxicological Effects of Sulfonamides and Fluoroquinolones and Their Removal by a green Alga (Chlorella Vulgaris) and a Cyanobacterium (Chrysosporum Ovalisporum). Environmental Pollution.
Tian, Y., Jiang, Y., Liu, Q., Xu, D., Liu, Y., and Song, J. (2021). The Impacts of Local and Regional Factors on the Phytoplankton Community Dynamics in a Temperate River, Northern China. Ecol. Indicators 123, 107352. doi:10.1016/j.ecolind.2021.107352
Wyta, C., Wlhb, C., and Fs, B. Identifying Diatom Indicator Species of Nutrient Enrichment: An In Situ Nutrient Enrichment experiment in Subtropical upland Streams. Ecol. Indicators 119, 1–10. doi:10.1016/j.ecolind.2020.106744
Xu, W., Zhu, X., Wang, X., Deng, L., and Zhang, G. (2006). Residues of Enrofloxacin, Furazolidone and Their Metabolites in Nile tilapia (Oreochromis niloticus). Aquaculture 254 (1-4), 1–8. doi:10.1016/j.aquaculture.2005.10.030
Yan, W., Bai, R., Wang, S., Tian, X., Li, Y., Wang, S., Yang, F., Xiao, Y., Lu, X., and Zhao, F. (2020). Antibiotic Resistance Genes Are Increased by Combined Exposure to Sulfamethoxazole and Naproxen but Relieved by Low-Salinity. Environ. Int. 139, 105742. doi:10.1016/j.envint.2020.105742
Zhang, Y., Wang, L., Zhuang, H., Li, X., Gao, X., An, Z., . . ., Liu, X., Yang, H., Wei, W., and Zhang, X. (2019). Excessive Use of Enrofloxacin Leads to Growth Inhibition of Juvenile Giant Freshwater Prawn Macrobrachium Rosenbergii. Ecotoxicology Environ. Saf. 169, 344–352. doi:10.1016/j.ecoenv.2018.11.042
Zhao, F., Yang, L., Chen, L., Xiang, Q., Li, S., Sun, L., . . ., Yu, X., and Fang, L. (2019). Soil Contamination with Antibiotics in a Typical Peri-Urban Area in Eastern China: Seasonal Variation, Risk Assessment, and Microbial Responses. J. Environ. Sci. (China) 79 (5), 200–212. doi:10.1016/j.jes.2018.11.024
Keywords: sulfamethoxazole, enrofloxacin, phytoplankton, primary productivity, biodiversity
Citation: Shan X, Shi Y, Fang L, Gui Y, Xing L, Qiu L, Hu G and Chen J (2021) Sulfamethoxazole and Enrofloxacin Antibiotics Affect Primary Productivity of Phytoplankton in Fishery Environment. Front. Environ. Sci. 9:754286. doi: 10.3389/fenvs.2021.754286
Received: 06 August 2021; Accepted: 07 September 2021;
Published: 20 September 2021.
Edited by:
Wei Wu, University of Southern Mississippi, United StatesReviewed by:
Liqiang Zhong, Freshwater Fisheries Research Institute of Jiangsu Province, ChinaCopyright © 2021 Shan, Shi, Fang, Gui, Xing, Qiu, Hu and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiazhang Chen, ZmZyY2NoZW5AaG90bWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.