 Tabitha C. Y. Hui
Tabitha C. Y. Hui Eleanor M. Slade
Eleanor M. Slade Ju Lian Chong
Ju Lian Chong
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Sci., 22 October 2021
Sec. Conservation and Restoration Ecology
Volume 9 - 2021 | https://doi.org/10.3389/fenvs.2021.637462
This article is part of the Research TopicProceedings of the 29th International Congress for Conservation Biology (ICCB 2019)View all 9 articles
As Malaysia’s economy continues to grow, an ever-expanding road network is being built to support the new developments. Whilst roads increase accessibility to resources and connect human communities, they also bring with them a host of environmental problems such as destruction and fragmentation of habitat, poaching, pollution, and roadkills. Of these, roadkills are the most direct and visible impact of building roads, yet few dedicated studies have been conducted on the extent of roadkills and how to minimize them in Malaysia. In this study, we examined the species richness and abundance of vertebrate roadkills along eight one-km stretches (two in each habitat) of highways in Terengganu and Pahang across four habitat types: rainforest, oil palm plantation, subsistence agricultural land, and village. Each stretch of highway was surveyed by walking it six times between May to October 2017 for the Terengganu segments and May to October 2018 for the Pahang segments. A total of 305 roadkilled animals were recorded, consisting of at least 24 species. Three of these are protected by Malaysian law. The most common roadkilled animals were frogs (102 individuals) followed by birds (70 individuals), mammals (67 individuals) and reptiles (66 individuals). The highest number of roadkills were recorded in roads going through villages (112 individuals), followed by oil palm plantations (81 individuals), forests (65 individuals) and subsistence agricultural land (47 individuals). For every roadkill we also recorded the location’s specific habitat and landscape characteristics such as distance to the nearest fruit tree, tree, shrub, water body, human structure, and overhead cable. Most roadkills were found close to vegetation cover and water bodies. However, we found that roadkills decreased to near zero between 30 and 80 m from the nearest shrub or tree. Roadkills decreased over larger distances from the nearest water body, but in oil palm and village habitats, reducing water bodies within 20–50 m from a road would almost half the number of roadkills. We suggest that making changes to the structure and composition of roadside vegetation and other landscape features, such as thinning vegetation to discourage wildlife from gathering near roads, could significantly reduce the number of roadkills.
Road traffic kills hundreds of millions of animals throughout the world every year (Spellerberg, 1998; Beckmann and Hilty, 2010), posing a significant threat to many species. In addition, roads promote the establishment of human settlements with subsequent habitat fragmentation and destruction, and changes in the dynamics of ecosystems (Trombulak and Frissell, 2000). Road construction tends to reduce landscape permeability and impact animal movements, causing populations to become more isolated, smaller and prone to local extinctions (Bissonette, 2002). Roads may also increase the spread of invasive species (Gelbard and Belnap, 2003), disturbance effects such as traffic noise and light (Reijnen et al., 1995; Reijnen and Foppen, 1995), and greenhouse gas emissions (Fuglestvedt et al., 2008).
Some authors suggest that roadkill rates can exceed those of natural mortality (Ferreras et al., 1992) and this will affect population density if there is no natural compensation mechanism of increased breeding (Fahrig et al., 1995; Huijser and Bergers, 2000). Similarly, imbalances can occur in the sex structure of populations (Gibbs and Steen, 2005), and for many species highway related mortality can serve as a limiting factor for foraging and dispersion. Although the impact of road related mortality is not as notable in populations with high abundances, it may have a considerable effect on threatened or endangered species, which are often characterized by their low abundances (Forman and Alexander, 1998; Glista et al., 2008). Roads and their traffic could, in fact, be compared to a new predator for which the animals have not developed any defense mechanisms or behavioral adaptations that might increase their survival. As a result, some populations may become unstable due to the additional mortality of adult and juvenile individuals (Andrews et al., 2008).
As animal casualties are among the more generalized and conspicuous effects of roads (Spellerberg, 1998; Beckmann and Hilty, 2010), the ecological effects of roads have been widely studied in temperate ecosystems in North America and Europe (e.g. Forman and Alexander, 1998; Spellerberg, 1998; Coffin, 2007; Beckmann and Hilty, 2010). However, there are fewer studies in tropical ecosystems (e.g. Goosem, 2001; Goosem, 2002; Pohlman et al., 2007; Kaiser and Hammers, 2009). This geographic bias of studies regarding roadkills is important because the severity of ecological effects varies among places due to differences in topography, variations in ecosystems, animal and human communities and population dynamics, and the life traits of the animal species (Forman and Alexander, 1998; Seiler, 2001; Forman et al., 2003).
Given the extent of wildlife killed on roads, there is a need to minimize the number of fauna-vehicle collisions. To take effective measures to mitigate wildlife road casualties, it is necessary to understand which factors determine their incidence and to identify ways of reducing the negative impacts of these factors (Clevenger et al., 2003). Because it is not financially feasible for most governments to create roadkill management strategies along the total length of all roads, it is necessary to choose locations where application of these measures will be most effective and efficient; in other words, to optimize the number of lives saved at as low a cost as possible.
Planned systematic studies of roadkills in Malaysia are few, e.g. Jamhuri et al. (2020) and Kasmuri et al. (2020), but there are numerous opportunistic reports from the public about mammals, birds and reptiles, especially endangered species such as the Malayan tiger Panthera tigris and Malayan tapir Acrocodia indica, being hit (Kamil et al., 2011; Kasmuri et al., 2020). This so-called “citizen science” can produce data that are very different from those generated by scientists in terms of the proportion of affected groups (Monge-Nájera, 2018), and tend to be skewed toward the larger, more visible and charismatic species. The lack of accurate and representative information about roadkills is of grave concern, especially in the tropics where land use expansion for food and energy production is a priority for economic development, leading to the expansion of road networks and the widening of older roads. Most studies on the subject only provide checklists of vertebrates killed on roads with no information on the relationships between abundance and landscape variables (Baker, 2019). Thus, roadkill assessment for Malaysian roads is important because it will allow us to document the significance of the impacts of roadkills on wildlife populations and improve the way new roads are built and existing roads are upgraded.
Our study areas were located just beyond the northern and western edges of Taman Negara, Malaysia’s oldest national park, on land gazetted as forest reserves which can be logged under permit. These areas are prone to illegal logging, conversion to plantations and poaching as they do not have the same protection status as a national park. The terrain is hilly and consists mostly of lowland dipterocarp forests with oil palm plantations, subsistence agriculture (forests interplanted with various types of fruit trees, herbs and vegetables for local consumption) and villages common along roads. Rainfall averages 3,000 mm year with a pronounced wet season from November to March. Flooding is common during this period, and roads often become inundated and impassable to traffic. We chose to focus our study on roads which run through four habitats which are becoming more common throughout Malaysia: forest reserves, subsistence agriculture, oil palm plantations, and villages. Even though these are not pristine habitats, higher than expected numbers of native wildlife, especially medium-sized mammals (2–15 kg), have been recorded as roadkills in these areas (Jamhuri et al., 2020). The findings from this study can therefore provide a representative baseline roadkill rate for roads found outside national park areas. Our two main aims were to: 1) compare the number of species and abundance of vertebrate roadkills in the four different habitat types; 2) identify adjacent landscape characteristics that influence the likelihood of road casualties along these roads. The information obtained allows us to make recommendations for mitigating road impacts on wildlife, particularly for smaller species such as herpetofauna, birds, civets, and rodents, which are usually underestimated.
We conducted this study on eight 1 km road segments in Peninsular Malaysia: four located on Terengganu state route T151, two on Federal Route 185, near Tasik Kenyir, Terengganu (Figure 1A), and two on Federal Route 8, near Merapoh, Pahang (Figure 1B). All the road segments were two-lane single carriageways with a lane width of 3–4 m and shoulder width of 2–3 m. There were no street lamps on these sections of road. As far as possible, we chose straight and flat sections of road to enable comparisons to be made among them. Road segments were at least 1 km apart from each other. The speed limit on T151 and 185 was 90 km/h and 60 km/h on route 8. The road segments on T151 ran through subsistence agricultural land and oil palm plantations (two road segments in each habitat type). Federal Route 185 ran through state forest reserves. The segments on Federal Route 8 had a lower speed limit because they ran through villages (Figure 1B). Average (± s.e.) vehicular traffic (counted at least two different points on each road segment between 0700 and 2,300 h for at least six separate days) was 44 ± 13 cars, 50 ± 17 motorcycles and 6 ± 2 heavy vehicles (e.g. trucks, lorries) per hour on Terengganu state route T151; 23 ± 5 cars, 16 ± 5 motorcycles and 10 ± 4 heavy vehicles per hour on Federal Route 185; and 7 ± 1 cars, 8 ± 1 motorcycles and 6 ± 1 heavy vehicles per hour on Federal Route 8.

FIGURE 1. Map of the surveyed road segments within Terengganu (A) and Pahang (B), Peninsular Malaysia.
We walked along both sides of each road segment, looking for vertebrate carcasses. Each road segment was surveyed six times during the dry season, once a month (one road segment per day of sampling, with approximately 4 weeks between surveys for each road segment). Surveys were conducted between 1,000 and 1,300 h so we could record both nocturnal as well as diurnal animals. We observed that the skin, bones and feathers of carcasses often stayed intact over several days, so removal by scavengers was deemed to be a minor factor affecting the number of roadkills we observed. However, deterioration of the carcasses caused by the often hot and humid weather conditions, and an increase in traffic every morning after about 0700 h, did make it difficult to identify roadkills to species level in many cases. Terengganu state route T151 and Federal Route 185 were surveyed from May to October 2017. Federal Route 8 was surveyed from May to October 2018. After identifying the roadkills to the lowest taxonomic group possible and recording the locations using a Global Positioning System (GPS), we moved the carcasses to the side of the road to avoid recounting. All carcasses were assigned to one of four vertebrate groups: birds, mammals, amphibians and reptiles. Records of domestic animals such as domestic chickens, cats and dogs, and individuals that were unidentifiable even to taxonomic group were excluded from the analyses. We assumed that all specimens found dead on the road were killed by vehicular traffic.
Separate chi square tests were conducted to determine whether there were statistically significant differences in the number of roadkills within and among 1) taxonomic groups (birds, mammals, amphibians and reptiles), and 2) habitats (subsistence agriculture, oil palm, rainforest and village). To determine how the immediate surrounding habitat factors influenced the incidence of roadkills, we performed a logistic regression to assess the relationship between the occurrence of roadkills and a set of six habitat variables. For this analysis, we measured the distance, in increments of 5 m (using a measuring tape if the roadkill was up to 30 m away from the target feature, or using the measuring tool in ArcMap 10.6.1 to measure the distances between the GPS locations of the roadkills and the physical features in Google Earth images if the roadkill was >30 m away) of each roadkill to the nearest fruit tree (not measured in oil palm habitat as this was the only tree species present and due to the regular spacing all roadkills were equidistant from an oil palm tree), non-fruit tree (not measured in oil palm habitat for the same reason described above), shrub, water body, human dwelling, and overhead cable (only measured in village habitat as cables were only present in village road segments). To avoid unbalanced models, a number of locations where no roadkills were recorded, corresponding to the number of locations where roadkills were recorded, were randomly selected to represent absences in logistic regression models (Gomes et al., 2009). The same six habitat characteristics were measured from each absence point. Logistic regression models were generated using presence/absence of roadkills as dependent variables and habitat characteristics as independent variables, and a binomial error structure. Data from the six surveys for each road segment, and both road segments within a habitat were combined for analysis.
We used the Information Theoretic (IT) approach (Anderson and Burnham, 2004; Richards, 2005) to compare sets of biologically plausible logistic regression models. The IT approach finds the best model from a suite of models, with the fewest parameters necessary (Johnson and Omland, 2004; Boyce et al., 2007). We followed a three-step approach (Merckx and Berwaerts, 2010). First, candidate models were compared for a dataset including all roadkills recorded across all taxa and habitats except oil palm, as only three variables were measured here out of six. We also excluded the variable cable as this variable was only assessed for village road segments and not for any other habitats. Second, a candidate set of models was generated for each habitat separately, including all variables that were found in each habitat. Third, a candidate set of models was generated for each of the taxa, dropping oil palm habitat and the variable cable for the reasons stated above. We checked for multicollinearity through variance inflation factors (VIF) using the vif function in the car package (Fox et al., 2012). No more than two variables in a model had high VIF values (>3), thus we retained all variables in all models. Akaike’s Information Criterion (AIC; Akaike, 1974) corrected for small sample sizes (AICc) was used to determine Akaike’s weights of evidence for each model given the other models considered. Using the mod.sel function in the MuMIn package (Barton, 2020), the models were sorted from highest to lowest AICc.
For each of the three steps, we defined a confidence set of models which have similar levels of substantial support from the data, and which have more support than all other models tested. The best models (up to a maximum of 10) where the Akaike’s difference with the top-ranked model was less than 5.0 were retained in this confidence set (Anderson and Burnham, 2004; Richards, 2005). These models were used to assess, and order, the variables in their ability to explain roadkill occurrence. Inference was based upon the simplest models in the confidence set, whereas factors present in the more complex models (but absent from the simpler models) in the confidence set only receive marginal support. To specifically assess the direction and strength of effects, we ran the best-ranked logistic regression models in each of the three steps to assess the variables they contained. All statistical analyses were conducted in R version 4.0.2 (R Core Team, 2020). Significance was considered at the α = 0.05 level.
We found a total of 305 carcasses: 70 birds, 67 mammals, 102 amphibians and 66 reptiles. 206 carcasses could be identified to species level (Table 1). At least 24 species were found. Of the carcasses that could be identified to species level, the species which exhibited the greatest mortality by far was the rice field frog Fejervarya limnocharis with 64 carcasses, more than half of which were found in the oil palm habitat, followed by the Malaysian wood rat Rattus tiomanicus (this is the most common Rattus species found in the area based on an unpublished study by TH on the impacts of roads on small mammal community structure; R. tiomanicus is distinguished from R. rattus by its creamy white underparts as opposed to brownish or greyish underparts) with 22 carcasses, almost half of which were also found in the oil palm habitat (Table 1). There was one carcass of a Malayan porcupine Hystrix brachyura, found along the subsistence agriculture habitat, one of a buffy fish owl Ketupa ketupu, found along the village habitat, and two leopard cats Prionailurus bengalensis, found on subsistence and oil palm road segments. All three species are protected in Malaysia (Table 1). The vertebrate roadkill rate in our study was 6.4 individuals/km/day of sampling. The highest rate was in village habitat where 112 carcasses, consisting mostly of rice field frogs and unidentified reptiles (usually snakes) were recorded (Table 1). The lowest number of roadkills was found in subsistence agriculture habitat (Table 1).
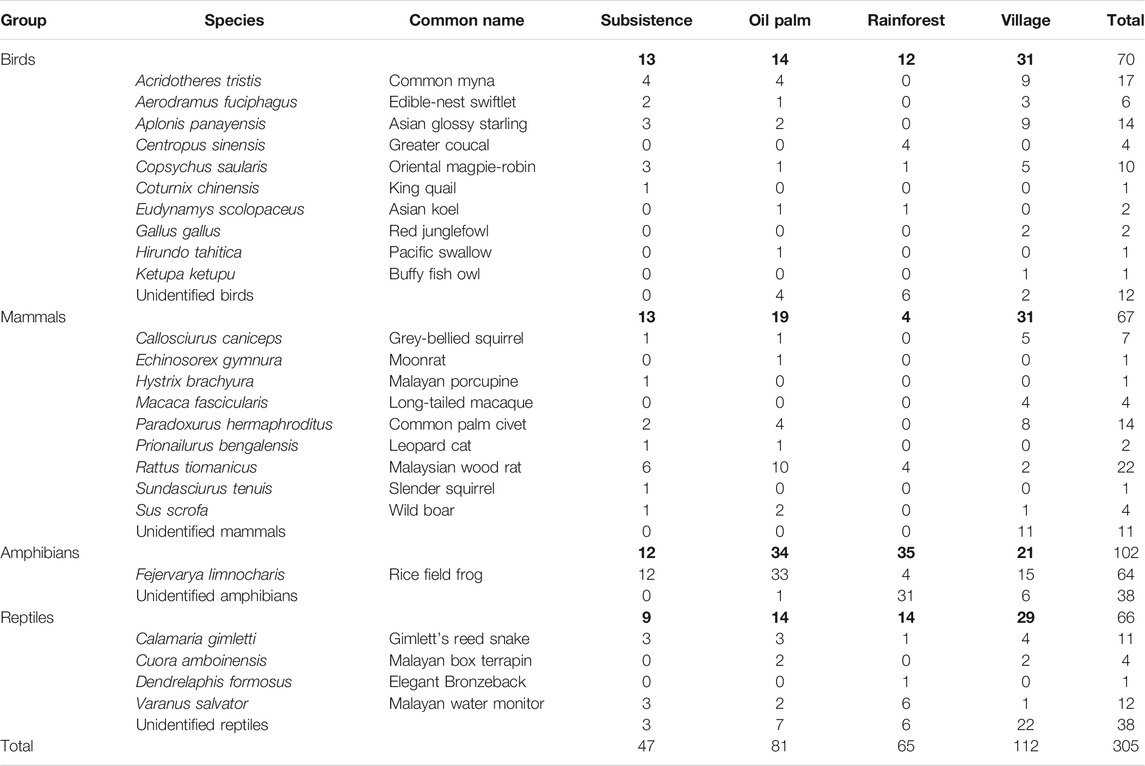
TABLE 1. Vertebrate roadkills found in the road segments listed by number of individuals per taxonomic group, species, and habitat type. A number of roadkills were unidentifiable because carcasses were too degraded. Bold values signifies “Total values”.
Overall, there were significant differences in the frequency of carcasses (i.e. frequency of mortality) among taxa in different habitats (χ2 1,9 = 32.832, p < 0.001). Between habitats, there were significant differences in the number of birds (χ2 1,3 = 14.000, p = 0.003), mammals (χ2 1,3 = 22.970, p < 0.001) and reptiles (χ2 1,3 = 13.636, p = 0.003) killed, with the highest number of casualties on village road segments. Amphibians were most commonly killed on rainforest road segments followed closely by oil palm road segments (χ2 1,3 = 14.314, p = 0.003).
Significantly more amphibians than other taxa were killed on oil palm (χ2 1,3 = 13.272, p = 0.004) and rainforest (χ2 1,3 = 32.292, p < 0.001) road segments. There were no significant differences between taxa in the number of animals killed in subsistence agriculture (χ2 1,3 = 0.915, p = 0.822) and village (χ2 1,3 = 2.429, p = 0.488) habitats.
All habitats and taxa: A total of five models were retained in the confidence set of models for all habitats and taxa combined. Four variables – distance to the nearest human structure (χ2 1,437 = 4.588, p = 0.032), shrub (χ2 1,437 = 6.441, p = 0.011), tree (χ2 1,437 = 42.472, p < 0.001) and water body (χ2 1,437 = 24.750, p < 0.001) – were part of the two best models. The more parsimonious model, containing these four variables only, was the best-ranked model overall (Table 2). The probability of roadkills occurring decreased sharply beyond 40 m from the nearest non-fruit tree and was almost zero beyond 80 m (Figure 2A). Distance from water and shrubs had a more gradual effect on roadkills, with distances of several hundred meters needed to reduce roadkills near water bodies (Figures 2B,C). Roadkills increased gradually the farther away the road was from human structures (Figure 2D).
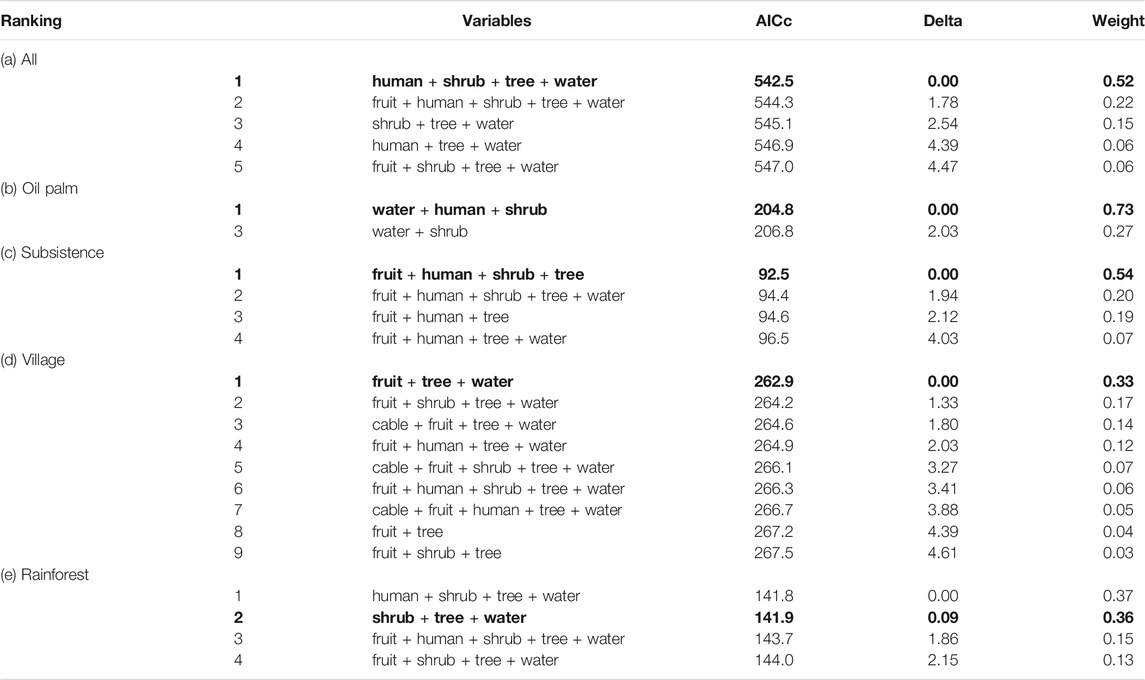
TABLE 2. Ranking of the best statistical models which were retained in each confidence set, where the Akaike's difference (Delta) with the top-ranked model was less than 5.0, following the Information Theoretic approach, for (a) all habitats and taxa combined, (b) oil palm, (c) subsistence agriculture, (d) village, and (e) rainforest habitats. The most parsimonious models are in bold. Weight refers to the cumulative weight of the models starting from the top ranked model. Bold values signifies “The most parsimonious models”.

FIGURE 2. Probability that a roadkill will occur with increasing distance from the nearest water body (A), tree (B), shrub (C) or human dwelling (D) in all habitats and for all taxa combined; water body (E), shrub (F) or human dwelling (G) in oil palm habitat; tree (H), shrub (I), fruit tree (J) or human dwelling (K) in subsistence agriculture habitat; water body (L), tree (M) or fruit tree (N) in village habitat; and water body (O), tree (P) or shrub (Q) in rainforest habitat.
Oil palm: For oil palm habitats, only two models were retained in the confidence set, and all three variables measured in this habitat were important in explaining the incidence of roadkills – distance to the nearest human structure (χ2 1,158 = 4.138, p = 0.042), water body (χ2 1,158 = 8.158, p = 0.004) and shrub (χ2 1,158 = 18.895, p < 0.001) (Table 2). The occurrence of roadkills in oil palm habitats decreased with increasing distance from the nearest water body over just 20 m (Figure 2E). Roadkills dropped most sharply with distance from the nearest shrub; and were nearly zero by 50 m distance (Figure 2F). The occurrence of roadkills in oil palm increased the farther away the road was from human dwellings (Figure 2G).
Subsistence agriculture: Four models were retained in the confidence set for subsistence agriculture habitats (Table 2). Distance to the nearest fruit tree (χ2 1,82 = 18.088, p < 0.001), non-fruit tree (χ2 1,82 = 8.380, p = 0.004), shrub (χ2 1,82 = 4.377, p = 0.036) and human structure (χ2 1,82 = 9.585, p = 0.002) were part of the best two models explaining the incidence of roadkills in subsistence agriculture habitats. Road casualties increased with decreasing distance from the nearest non-fruit tree (Figure 2H), shrub (Figure 2I), and fruit tree (Figure 2J). There were almost no roadkills beyond 30 m from the nearest fruit tree (Figure 2J). The occurrence of roadkills increased gradually with distance from human dwellings (Figure 2K).
Village: A total of nine models were retained in the confidence set for village road segments (Table 2). Three variables – distance to the nearest fruit tree (χ2 1,219 = 8.942, p = 0.003), non-fruit tree (χ2 1,219 = 18.917, p < 0.001) and water body (χ2 1,219 = 6.460, p = 0.011) – were part of seven out of nine models. The most parsimonious model containing these three variables was the best-ranked model overall (Table 2). The probability of roadkills occurring decreased with increasing distance from the nearest water body (Figure 2L), tree (Figure 2M) and fruit tree (Figure 2N), dropping to zero by around 70 m, 80 m, and 200 m respectively. There was no significant effect of overhead cables on the incidence of roadkills in villages (Table 2).
Rainforest: A total of four models were retained in the confidence set for rainforest road segments (Table 2). All of these models contained the variables distance to the nearest shrub (χ2 1,128 = 11.802, p = 0.001), tree (χ2 1,128 = 16.649, p < 0.001) and water body (χ2 1,128 = 12.469, p < 0.001). The most parsimonious model, containing these three variables only, was the second best-ranked model overall (Table 2). The occurrence of roadkills in rainforest habitats increased with decreasing distance from the nearest water body (Figure 2O), non-fruit tree (Figure 2P) and shrub (Figure 2Q). Probability of roadkills occurring in rainforest habitats decreased to zero beyond 30 m from a tree (Figure 2P). Roadkills dropped most sharply with distance from the nearest shrub in rainforest habitats, and by about 25 m, there was hardly any chance of a roadkill occurring due to proximity to a shrub (Figure 2Q). However, the decline in roadkills with distance to a water body was more gradual and occurred over several hundred meters.
Birds and Mammals: For birds, there were more than ten models where the Akaike’s difference with the top-ranked model was less than 5.0, but only one variable appeared in all of them and that was distance to the nearest tree (Table 3). The occurrence of roadkilled birds (χ2 1,110 = 13.096, p < 0.001) increased closer to tree cover and diminished to zero beyond 60 m (Figure 3A). Likewise with mammals, there were more than ten models where the Akaike’s difference with the top-ranked model was less than 5.0, but only one variable appeared in all of them and that was distance to the nearest shrub (Table 3). Occurrence of mammal roadkills (χ2 1,134 = 6.034, p = 0.014) decreased more gradually with increasing distance from the nearest shrub, dropping significantly beyond 40 m (Figure 3B).
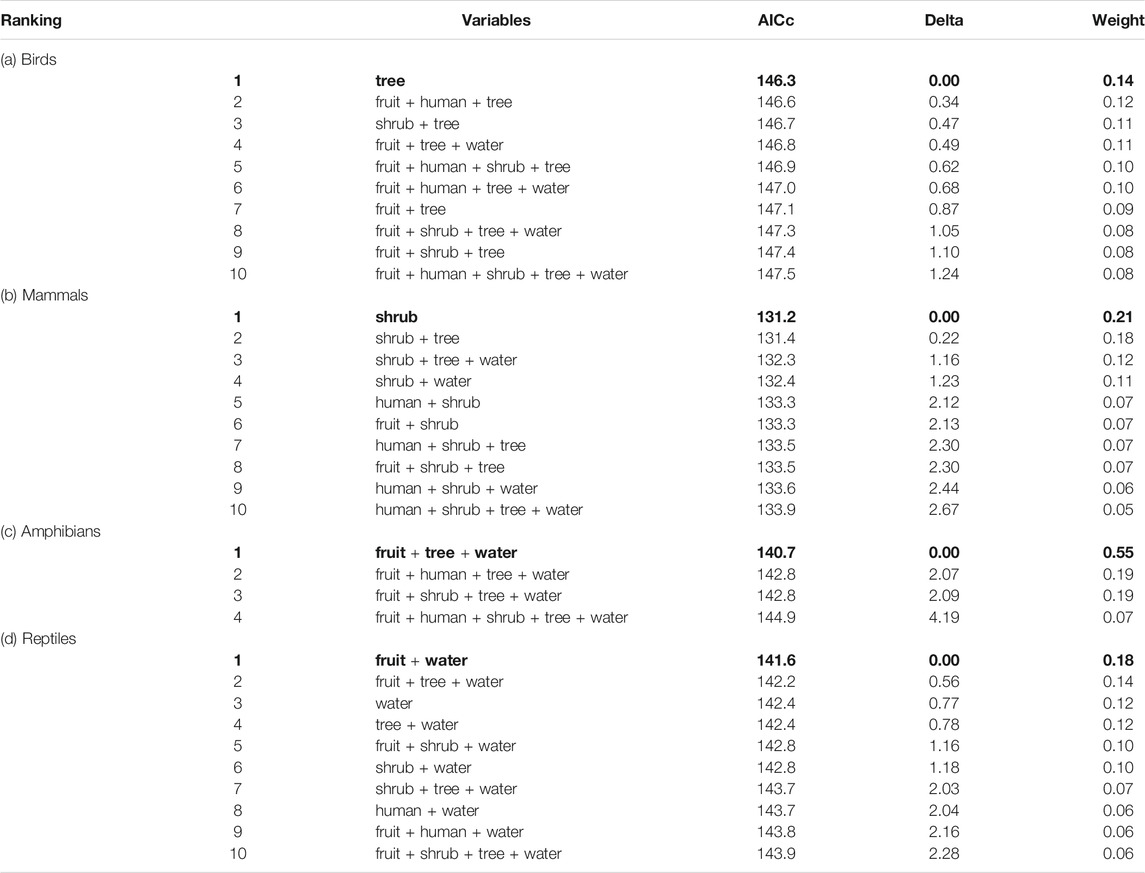
TABLE 3. Ranking of the best statistical models which were retained in each confidence set, where the Akaike's difference (Delta) with the top-ranked model was less than 5.0, following the Information Theoretic approach, for (a) birds, (b) mammals, (c) amphibians, and (d) reptiles. The most parsimonious models are in bold. Weight refers to the cumulative weight of the models starting from the top ranked model. Bold values signifies “The most parsimonious models”.
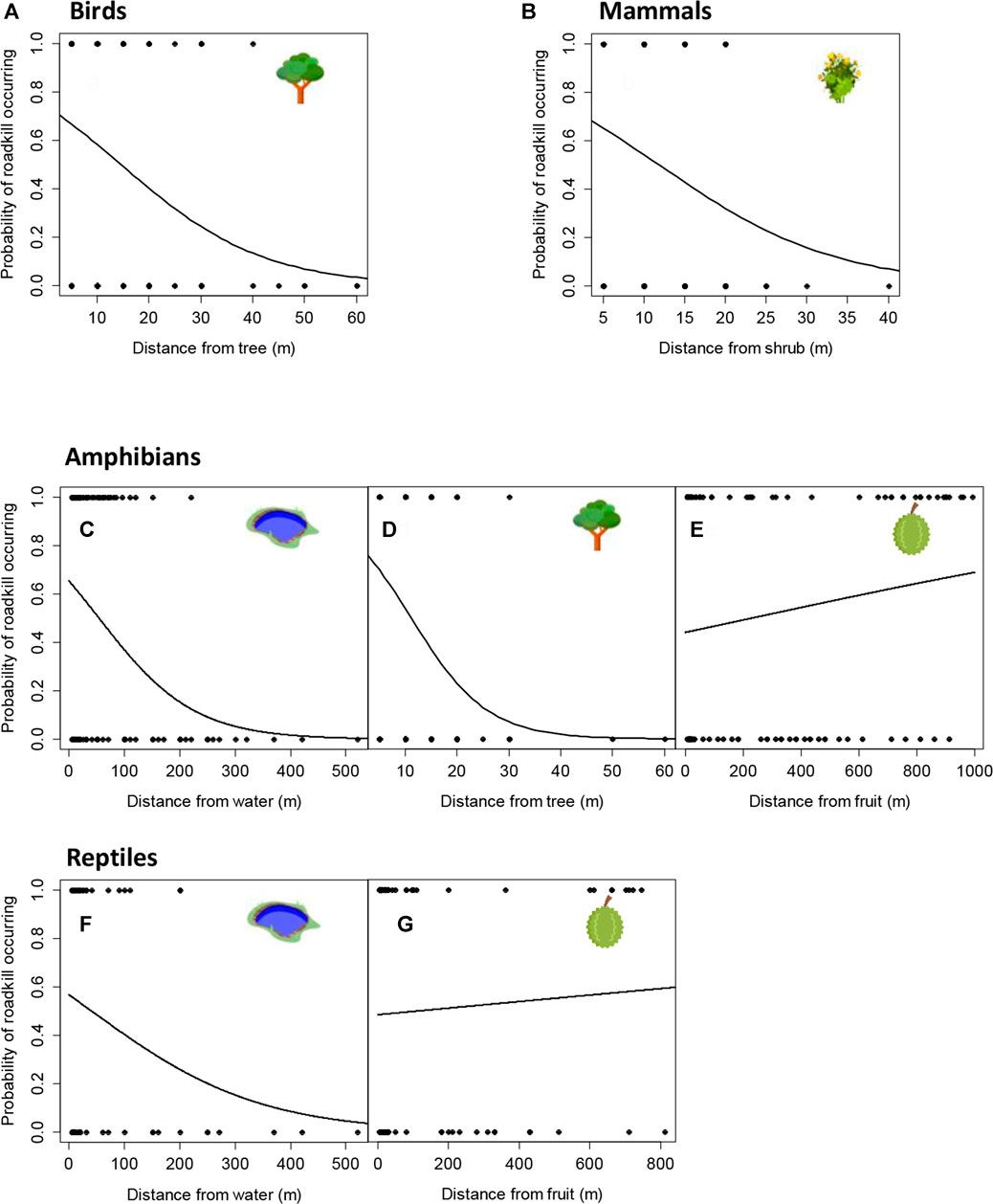
FIGURE 3. Probability that a bird roadkill will occur with increasing distance from the nearest tree (A); that a mammal roadkill will occur with increasing distance from the nearest shrub (B); that an amphibian roadkill will occur with increasing distance from the nearest water body (C), tree (D) or fruit tree (E); and that a reptile roadkill will occur with increasing distance from the nearest water body (F) or fruit tree (G).
Amphibians: There were only four models retained in the confidence set for amphibians and these contained the three variables - distance to the nearest fruit tree (χ2 1,132 = 12.334, p < 0.001), tree (χ2 1,132 = 15.807, p < 0.001) and water body (χ2 1,132 = 37.060, p < 0.001). The best model was thus deemed as the one containing only these three variables (Table 3). The occurrence of roadkilled amphibians increased closer to water bodies (Figure 3C) and tree cover (Figure 3D). The probability of roadkills occurring due to proximity to a tree diminished to zero beyond 60 m (Figure 3D). The impacts of distance from the nearest water body on amphibian roadkills did not dissipate until beyond 300 m (Figure 3C). In contrast, amphibian roadkills significantly increased with increasing distance from fruit trees (Figure 3E), but this effect occurred over hundreds of meters.
Reptiles: There were more than ten models where the Akaike’s difference with the top-ranked model was less than 5.0 for reptiles. Most of these contained the variables distance to the nearest fruit tree and water body. The best-ranked model contained both these variables (Table 3). Reptile roadkills decreased with increasing distance from water (χ2 1,101 = 8.444, p = 0.004) over several hundred meters (Figure 3F), but similar to amphibians, increased with increasing distance from the nearest fruit tree (Figure 3G). However, the increase in roadkills with increasing distance from the nearest fruit tree was not significant at the α = 0.05 level for reptiles (χ2 1,101 = 2.893, p = 0.089) (Figure 3G).
The impact of roads on wildlife has been understudied and sometimes even ignored; however, this may represent a major factor in the conservation of biodiversity, especially that of small vertebrates. The number of vertebrates we observed as roadkill was high (305 animals) over a relatively short distance (8 km of transects), or about 38 animals/km over 6 days of sampling. In comparison, other studies ranged from 1.25 to 8.65/km/year for all vertebrates (Clevenger et al., 2003; Taylor and Goldingay, 2004; Dornas, 2012). Laurance et al. (2009) raised the possibility of wildlife from tropical regions being more vulnerable to road impacts when compared to temperate regions due to their greater diversity and different ecological demands.
Trends in roadkills varied according to taxonomic group and habitat variables. Of the total number of roadkilled animals, amphibians were the most numerous, followed by birds, mammals and reptiles. Just as in natural communities, within each group the abundance of the roadkilled species was dominated by a few taxa. Fejervarya limnocharis was the most common amphibian killed except along rainforest roads. Similar to other studies (e.g. Arroyo-Cabrales et al., 2003; Orlowski and Nowak, 2006), we found that rodents, specifically Rattus tiomanicus, formed a substantial proportion of the mammal roadkills (46.3% of all individuals).
Overall, amphibian (frog) mortality in the study areas was higher than the roadkill of any other vertebrates. Most of these casualties were found on oil palm and rainforest road segments. Mammals and birds were the most frequently hit vertebrate groups in subsistence and village habitats, though the number of casualties on oil palm road segments was also high. Reptile casualties in oil palm and rainforest habitats were higher than in subsistence habitat, but the highest number of casualties was in villages.
Other studies have found that mammals were the most common vertebrates found as roadkill. For instance, censuses during 18 months on a 50 km road segment on the Atlantic coast of Colombia documented roadkills of 216 vertebrates of 41 species: 41.7% mammals, 26.8% birds, 26.4% reptiles and 5.1% amphibians (Argotte and Monsalvo, 2002). However, we found that amphibians made up most of the roadkills. The differences can mainly be explained by our survey approach. Detectability of the carcasses was maximized by doing the surveys by walking instead of driving, taking about 1 h per km. This made it possible to search for roadkills exhaustively, and to detect even small vertebrate carcasses such as songbirds and frogs, which are often underestimated.
The probability of birds, mammals and frogs being struck by vehicles increased nearer to trees and shrubs, which offer habitat and cover for moving across their ranges. The presence of vegetation at the roadside makes a good sheltered foraging area, especially during the fruiting season (June to August), suggesting that individuals may be attracted there by the availability of food (Meunier et al., 2000; Orłowski, 2008). This may increase the frequency of animal movement along and across the road, thereby increasing the likelihood of roadkills. Surprisingly, mortality of amphibians and reptiles increased farther away from fruit trees. This could be due to an inverse relationship between the presence of non-fruit trees and shrubs, and fruit trees.
Amphibian and reptile mortality was higher closer to water bodies and drains. However, unlike for trees, mortality decreased much more gradually over distance, showing the extensive pull of water bodies as a factor influencing roadkills for these two taxa. For frogs, this was an obvious indication of roadside drains being used as habitat. Additionally, the scarcity of water, particularly during the dry season, can also explain why fatalities were concentrated near water bodies. The draw of water for reptiles could be due to the abundance of frog prey near these habitats, in addition to their own water needs as many of the snake species killed were tropical rainforest species. Reptiles may be drawn to open areas with higher temperatures, such as roads, for thermoregulation, especially when traffic is not too frequent. They may also be attracted to roads because of the presence of roadkill carcasses, in turn becoming roadkills themselves.
Species vulnerability to being killed on roads also depends on their behavior (Clevenger et al., 2003). For instance, many species avoid humans and that could explain why fewer roadkills of wildlife were found nearer houses and other manmade structures. The majority of bird species found as roadkill, such as the Oriental magpie-robin Copsychus saularis and greater coucal Centropus sinensis, are low flying or spend time on the ground. This suggests that the vulnerability of birds may relate to their flying behaviour. Although small mammals tend to avoid roads (McGregor et al., 2008) and their carcasses tend to degrade rapidly or be removed in whole by predators, affecting their detectability compared to larger mammals (Slater, 2002), we found many Rattus tiomanicus carcasses, especially on roads passing through oil palm plantations. This could indicate their relatively higher abundance in such habitats, as rats are regarded as pests which feed on oil palm fruits and shoots in such areas (Puan et al., 2011). Other native species which were often seen and killed on other stretches of roads passing through oil palm include one of the Malaysian wood rat’s main predators, the leopard cat Prionailurus bengalensis, which is also a threatened and protected species in Malaysia.
The highest number of roadkills recorded in our study came from village road segments, making up more than a third of the total number of carcasses found. This may be due to the abundance of fruit trees such as rambutan, durian, jackfruit, banana and langsat planted by the villagers along the roadside. During the fruiting season for most trees, many birds and mammals such as bats and civets are attracted to these food sources and become vulnerable as they move back and forth across the road from one fruit or flower to the next. Our results showed that reducing fruit and non-fruit trees planted within 100 m from the road would significantly reduce roadkills.
It is worrisome that three threatened species were recorded as roadkills. Throughout its distribution, the Malayan porcupine Hystrix brachyura is protected under legislation. In Peninsular Malaysia, the species is protected under the Wildlife Conservation Act (2010). Overhunting of Hystrix brachyura for its meat and its bezoars (used in traditional medicine) is the greatest threat to their populations (Borschberg, 2006). Habitat loss, fragmentation (e.g. by roads) and landscape modification also contribute to the decline in Hystrix brachyura numbers (Corlett, 2003). Another threatened species carcass we found was that of a buffy fish owl Ketupa ketupu. Owls are susceptible to being killed on roads due to their nocturnal behaviour. At night, it is more likely that car lights cause temporary blindness in individuals that attempt to cross roads, further inducing collisions (Hernandez, 1988).
In general, the species most frequently killed on roads are the most abundant and with broad distributions (Dornas, 2012; Cáceres et al., 2011; Rosa and Bager, 2012). This is probably why larger mammals such as tapirs and leopards were not recorded as roadkills in this study, as they are already rare in Malaysia and confined mostly to undisturbed rainforests (Jamhuri et al., 2020). Larger mammals are also more visible from a farther distance to drivers who would then have time to take evasive action to avoid a collision. Most of the roadkilled species found in our study are common and typical of habitats modified or inhabited by humans, but may still indicate roadkill rates possible for threatened species. The lack of population studies on and data on the size of the populations of the species we recorded in this study meant that we could not assess the true impact of the observed deaths. However, it is known that roadkills randomly eliminate healthy individuals of a population, which can exacerbate declines in populations which are already fragmented by roads (Bujoczek et al., 2011). For a comprehensive assessment of road impacts on wildlife in Malaysia, studies on population size and long-term effects of fragmentation on population dynamics are needed. Knowing how many individuals per species are being roadkilled along with their abundance adjacent to the roads would give us a clearer idea on where we should concentrate conservation efforts. We often think of roadkills from a wildlife protection perspective, but they can have negative impacts on humans too. Collisions with large animals such as wild boars Sus scrofa could cause serious accidents, including loss of human lives or substantial material damages for the driver (Forman et al., 2003; Huijser et al., 2009). It is therefore important to come up with appropriate mitigation measures for managing roadkills.
Little has been done to maintain the connectivity of the natural elements of landscapes intersected by roads or diminish roadkill, despite the fact that vehicle traffic is one of the most serious causes of mortality for many endangered species of mammals around the world (Blaustein and Wake, 1990; Fahrig et al., 1995; Seiler, 2001). Measures aimed at reducing traffic volume, speed or altering the timing of traffic seasonally or daily will not fully mitigate the effects of roads on animal movement. Additional measures such as the construction of suitable wildlife crossings (Clevenger et al., 2001; Mcdonald and St. Clair, 2004) and in extreme cases (e.g. mitigation for endangered species), removal of the road itself are also needed. Conservation of large roadless areas should be a priority (Crist et al., 2005) to maximize population connectivity, and reduce fragmentation effects and roadkills.
However, given that it is projected that another 25 million km of paved roads will be built by 2050 (Alamgir et al., 2017), primarily in the tropics, our study suggests some simple local efforts that can be done to reduce roadkills. Most roadkills occurred near shrubs and trees, efforts could therefore focus on the management of roadside vegetation, reducing vegetation cover, especially fruit trees within a minimum of 30 m from roads bordering subsistence agriculture, and 100 m away from roads in village areas, to reduce their attractiveness to wildlife. Our study showed that roadkills start declining even within relatively short distances from vegetation and decrease markedly beyond 30 m in most habitats and for most taxa. The maintenance of suitable substitute habitats in areas greater than 100 m from roads should then be considered. Similarly, draining ponds and drains within 500 m from roads could discourage frogs from gathering near roads, but replacement water bodies away from the roads should then be provided. We understand that these recommendations can be controversial (Forman, 2012), especially when roadside vegetation is often seen as an important refuge for wildlife (Carthew et al., 2013; Gardiner et al., 2018), wildlife corridor (Sodhi et al., 1999; Bolger et al., 2001), barrier against noise, pollution and dust (Bałazy, 2002), or has even been shown to provide ecosystem services for humans (Säumel et al., 2016). It seems there will be a tradeoff between the number of roadkills and the connectivity of habitats across roads. We propose that vegetation be planted extensively along road sections where it is safer for animals to cross, such as at a wildlife crossing, or where there is less traffic, and thinned out at sections which are deemed unsafe. In any case, it must be stressed that the direction of road planning requires further investigation into roadkill-habitat relationships as well as finding ways of reducing negative traffic impacts on the environment. These should be subsequently implemented both in planning applications for new road schemes and in the management rules of existing roadside hedgerows and tree belts with the aim to reduce losses of wildlife in rural landscapes.
Lastly, road users should also play a role in reducing roadkills. Wildlife roadkills could be reduced if drivers are educated in road awareness and can anticipate the behavior of wildlife near roads and understand what they can do to avoid collisions (Fudge et al., 2007). This is especially important in areas where there are endangered species.
The impact roads have on wildlife through vehicle collisions spans all taxonomic groups, although a few taxa such as frogs dominate the roadkills. In Malaysia, the effectiveness of mitigation measures such as viaducts in reducing roadkills have not been assessed. We found significant effects of trees, shrubs and water bodies adjacent to the road on the number of roadkills, indicating that simpler and likely cheaper methods such as roadside vegetation and drainage management may decrease roadkills significantly. However, there may be tradeoffs between reducing roadkills and maintaining connectivity and habitats for wildlife. More road ecology studies are required within Southeast Asian landscapes if we want to understand effective mitigation measures and implement more environmentally friendly road development.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by Department of Wildlife and National Parks Peninsular Malaysia.
TH designed, carried out the study, conducted the analyses and wrote the manuscript. ES made suggestions on analysis methods and edited the manuscript. JC helped with the design and location of the study, analyses and edited the manuscript.
Rufford Foundation (21698-1) and Animal Behavior Society funded fieldwork. Skyrail Rainforest Foundation Research Grant and IDEA WILD grant were used to purchase equipment and supplies.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Sohud bin Yaacob for assistance with searching for roadkills. Tan Poai Ean helped with identification of roadkills. We would like to thank the Economic Planning Unit (Permit numbers 3823 and 3949) of the Prime Minister’s Department and the Department of Wildlife and National Parks (HQ, Terengganu and Pahang) for granting permission to undertake this project, as well as the Department of Forestry (Terengganu and Pahang) for granting permission to enter the forest reserves in Kenyir and Sungai Yu. Our research was financially supported by The Rufford Foundation Grant for Nature Conservation, an IDEA WILD Equipment Grant, Skyrail Rainforest Foundation Research Grant and Animal Behavior Society Student Research Grant.
Akaike, H. (1974). A New Look at the Statistical Model Identification. IEEE Trans. Automat. Contr. 19, 716–723. doi:10.1109/tac.1974.1100705
Alamgir, M., Campbell, M. J., Sloan, S., Goosem, M., Clements, G. R., Mahmoud, M. I., et al. (2017). Economic, Socio-Political and Environmental Risks of Road Development in the Tropics. Curr. Biol. 27, R1130–R1140. doi:10.1016/j.cub.2017.08.067
Anderson, D., and Burnham, K. (2004). Model Selection and Multi-Model Inference. Second, 63. NY: Springer-Verlag, 10.
Andrews, K. M., Gibbons, J. W., Jochimsen, D. M., and Mitchell, J. (2008). Ecological Effects of Roads on Amphibians and Reptiles: a Literature Review. Herpetological Conservation 3, 121–143.
Argotte, S. D., and Monsalvo, M. (2002). Incidencia de la carretera Barranquilla-Ciénaga sobre la mortalidad de vertebrados y su relación con el medio ecológico en la vía Parque Isla de Salamanca, Magdalena, Colombia. Barranquilla, Colombia: Universidad del Atlántico.
Arroyo-Cabrales, J., Polaco, O. J., Johnson, E., and Guzmán, A. (2003). The Distribution of the Genus Mammuthus in Mexico. Deinsea 9, 27–40.
Baker, N. (2019). Some Snake Records from Gunung Arong Forest Reserve, Johor, Peninsular Malaysia. Southeast Asia Vertebr. Rec. 2019, 001–006.
Bałazy, S. (2002). “Ecological Guidelines for the Management of Afforestations in Rural Areas,” in Landscape Ecology in Agroecosystems Management. Boca Raton: CRC Press.
Barton, K. 2020. Mu-MIn: Multi-Model Inference. R Package Version 0.12. 2/r18; 2009, Available at: http://R-Forge.R-project.org/projects/mumin/,
Beckmann, J. P., and Hilty, J. A. (2010). Connecting Wildlife Populations in Fractured landscapesSafe Passages: Highways, Wildlife, and Habitat Connectivity, 3–16.
Bissonette, J. A. (2002). Scaling Roads and Wildlife: the Cinderella Principle. Z. für Jagdwissenschaft 48, 208–214. doi:10.1007/bf02192410
Blaustein, A. R., and Wake, D. B. (1990). Declining Amphibian Populations: a Global Phenomenon. Trends Ecol. Evol. 5 (7), 203–204. doi:10.1016/0169-5347(90)90129-2
Bolger, D. T., Scott, T. A., and Rotenberry, J. T. (2001). Use of Corridor-like Landscape Structures by Bird and Small Mammal Species. Biol. conservation 102, 213–224. doi:10.1016/s0006-3207(01)00028-3
Borschberg, P. (2006). The Trade, Forgery and Medicinal Use of porcupine Bezoars in the Early Modern Period (C. 1500–1750). Oriente 14, 60–78.
Boyce, M., Rushton, S., and Lynam, T. (2007). Does Modelling Have a Role in Conservation. Key Top. Conservation Biol. 2007, 134–144.
Bujoczek, M., Ciach, M., and Yosef, R. (2011). Road-kills Affect Avian Population Quality. Biol. Conservation 144, 1036–1039. doi:10.1016/j.biocon.2010.12.022
Cáceres, M. D., San Román, J. A. R., and Brändle, G. (2011). El uso de la televisión en un contexto multipantallas: viejas prácticas en nuevos medios. Anàlisi: quaderns de comunicació i cultura, 21–44.
Carthew, S. M., Garrett, L. A., and Ruykys, L. (2013). Roadside Vegetation Can Provide Valuable Habitat for Small, Terrestrial Fauna in South Australia. Biodivers Conserv 22, 737–754. doi:10.1007/s10531-013-0445-0
Clevenger, A. P., Chruszcz, B., and Gunson, K. E. (2001). Highway Mitigation Fencing Reduces Wildlife-Vehicle Collisions. Wildl. Soc. Bull. 2001, 646–653.
Clevenger, A. P., Chruszcz, B., and Gunson, K. E. (2003). Spatial Patterns and Factors Influencing Small Vertebrate Fauna Road-Kill Aggregations. Biol. conservation 109, 15–26. doi:10.1016/s0006-3207(02)00127-1
Coffin, A. W. (2007). From Roadkill to Road Ecology: a Review of the Ecological Effects of Roads. J. transport Geogr. 15, 396–406. doi:10.1016/j.jtrangeo.2006.11.006
Crist, M. R., Wilmer, B., and Aplet, G. H. (2005). Assessing the Value of Roadless Areas in a Conservation reserve Strategy: Biodiversity and Landscape Connectivity in the Northern Rockies. J. Appl. Ecol. 42, 181–191. doi:10.1111/j.1365-2664.2005.00996.x
Dornas, R. A. P., Kindel, A., Bager, A., and Freitas, S. R. (2012). “Avaliação da mortalidade de vertebrados em rodovias no Brasil,” in Ecologia de Estradas: tendências e pesquisas. Editor A. BAGER (Lavras: UFLA).
Fahrig, L., Pedlar, J. H., Pope, S. E., Taylor, P. D., and Wegner, J. F. (1995). Effect of Road Traffic on Amphibian Density. Biol. conservation 73, 177–182. doi:10.1016/0006-3207(94)00102-v
Ferreras, P., Aldama, J. J., Beltrán, J. F., and Delibes, M. (1992). Rates and Causes of M Ortality in a Fragment Population of Iberian lynx, Felis Pardina (Temminck). Biol. Conservation. 61 (3), 197–202. doi:10.1016/0006-3207(92)91116-a
Forman, R. T. (2012). Safe Passages: Highways, Wildlife, and Habitat Connectivity. Washington D.C.: Island Press.
Forman, R. T., Sperling, D., Bissonette, J. A., Clevenger, A. P., Cutshall, C. D., Dale, V. H., et al. (2003). Road Ecology: Science and Solutions. Washington D.C.: Island press.
Forman, R. T. T., and Alexander, L. E. (1998). Roads and Their Major Ecological Effects. Annu. Rev. Ecol. Syst. 29, 207–231. doi:10.1146/annurev.ecolsys.29.1.207
Fox, J., Weisberg, S., Adler, D., Bates, D., Baud-Bovy, G., Ellison, S., et al. (2012). Package ‘car’. Vienna: R Foundation for Statistical Computing.
Fudge, D., Freedman, B., Crowell, M., Nette, T., and Power, V. (2007). Road-kill of Mammals in Nova Scotia. Can. Field Nat. 121, 265–273. doi:10.22621/cfn.v121i3.474
Fuglestvedt, J., Berntsen, T., Myhre, G., Rypdal, K., and Skeie, R. B. (2008). Climate Forcing from the Transport Sectors. Proc. Natl. Acad. Sci. 105, 454–458. doi:10.1073/pnas.0702958104
Gardiner, M. M., Riley, C. B., Bommarco, R., and Öckinger, E. (2018). Rights‐of‐way: a Potential Conservation Resource. Front. Ecol. Environ. 16, 149–158. doi:10.1002/fee.1778
Gelbard, J. L., and Belnap, J. (2003). Roads as Conduits for Exotic Plant Invasions in a Semiarid Landscape. Conservation Biol. 17, 420–432. doi:10.1046/j.1523-1739.2003.01408.x
Gibbs, J. P., and Steen, D. A. (2005). Trends in Sex Ratios of Turtles in the United States: Implications of Road Mortality. Conservation Biol. 19, 552–556. doi:10.1111/j.1523-1739.2005.000155.x
Glista, D. J., Devault, T. L., and Dewoody, J. A. (2008). Vertebrate Road Mortality Predominantly Impacts Amphibians. Herpetological conservation Biol. 3, 77–87.
Gomes, L., Grilo, C., Silva, C., and Mira, A. (2009). Identification Methods and Deterministic Factors of Owl Roadkill Hotspot Locations in Mediterranean Landscapes. Ecol. Res. 24, 355–370. doi:10.1007/s11284-008-0515-z
Goosem, M. (2002). Effects of Tropical Rainforest Roads on Small Mammals: Fragmentation, Edge Effects and Traffic Disturbance. Wildl. Res. 29, 277–289. doi:10.1071/wr01058
Goosem, M. (2001). Effects of Tropical Rainforest Roads on Small Mammals: Inhibition of Crossing Movements. Wildl. Res. 28, 351–364. doi:10.1071/wr99093
Huijser, M. P., and Bergers, P. J. M. (2000). The Effect of Roads and Traffic on Hedgehog (Erinaceus europaeus) Populations. Biol. conservation 95, 111–116. doi:10.1016/s0006-3207(00)00006-9
Huijser, M. P., Duffield, J. W., Clevenger, A. P., Ament, R. J., and Mcgowen, P. T. (2009). Cost–benefit Analyses of Mitigation Measures Aimed at Reducing Collisions with Large Ungulates in the United States and Canada: a Decision Support Tool. Ecol. Soc. 14. doi:10.5751/es-03000-140215
Jamhuri, J., Edinoor, M. A., Kamarudin, N., Lechner, A. M., Ashton‐Butt, A., and Azhar, B. (2020). Higher Mortality Rates for Large‐ and Medium‐sized Mammals on Plantation Roads Compared to Highways in Peninsular Malaysia. Ecol. Evol. 10, 12049–12058. doi:10.1002/ece3.6827
Johnson, J. B., and Omland, K. S. (2004). Model Selection in Ecology and Evolution. Trends Ecol. Evol. 19, 101–108. doi:10.1016/j.tree.2003.10.013
Kaiser, K., and Hammers, J. (2009). The Effect of Anthropogenic Noise on Male Advertisement Call Rate in the Neotropical Treefrog, Dendropsophus Triangulum. Behav 146, 1053–1069. doi:10.1163/156853909x404457
Kamil, S., Zainuddin, Z. Z., and Abidin, F. A. Z. (2011). Roadkill of a Flat-Headed Cat in Pahang. Peninsular Malaysia: Catnews.
Kasmuri, N., Nazar, N., and Mohd Yazid, A. Z. (2020). Human and Animals Conflicts: A Case Study of Wildlife Roadkill in Malaysia. E-BPJ 5, 315–322. doi:10.21834/e-bpj.v5i13.2093
Laurance, W. F., Goosem, M., and Laurance, S. G. W. (2009). Impacts of Roads and Linear Clearings on Tropical Forests. Trends Ecol. Evol. 24, 659–669. doi:10.1016/j.tree.2009.06.009
Mcdonald, W., and St. Clair, C. C. (2004). Elements that Promote Highway Crossing Structure Use by Small Mammals in Banff National Park. J. Appl. Ecol. 41, 82–93. doi:10.1111/j.1365-2664.2004.00877.x
Mcgregor, R. L., Bender, D. J., and Fahrig, L. (2008). Do small Mammals Avoid Roads Because of the Traffic? J. Appl. Ecol. 45, 117–123. doi:10.1111/j.1365-2664.2007.01403.x
Merckx, T., and Berwaerts, K. (2010). What Type of Hedgerows Do Brown Hairstreak (Thecla Betulae L.) Butterflies Prefer? Implications for European Agricultural Landscape Conservation. Insect Conservation Divers. 3, 194–204. doi:10.1111/j.1752-4598.2010.00088.x
Meunier, F. D., Verheyden, C., and Jouventin, P. (2000). Use of Roadsides by Diurnal Raptors in Agricultural Landscapes. Biol. Conservation 92, 291–298. doi:10.1016/s0006-3207(99)00094-4
Monge-Nájera, J. (2018). Road Kills in Tropical Ecosystems: a Review with Recommendations for Mitigation and for New Research. Rbt 66, 722–738. doi:10.15517/rbt.v66i2.33404
Orlowski, G., and Nowak, L. (2006). Factors Influencing Mammal Roadkills in the Agricultural Landscape of South-Western Poland. Polish J. Ecol. 54, 283–294.
Orłowski, G. (2008). Roadside Hedgerows and Trees as Factors Increasing Road Mortality of Birds: Implications for Management of Roadside Vegetation in Rural Landscapes. Landscape Urban Plann. 86, 153–161. doi:10.1016/j.landurbplan.2008.02.003
Pohlman, C. L., Turton, S. M., and Goosem, M. (2007). Edge Effects of Linear Canopy Openings on Tropical Rain forest Understory Microclimate. Biotropica 39, 62–71. doi:10.1111/j.1744-7429.2006.00238.x
Puan, C. L., Goldizen, A. W., Zakaria, M., Hafidzi, M. N., and Baxter, G. S. (2011). Relationships Among Rat Numbers, Abundance of Oil palm Fruit and Damage Levels to Fruit in an Oil palm Plantation. Integr. Zoolog. 6, 130–139. doi:10.1111/j.1749-4877.2010.00231.x
Reijnen, R., Foppen, R., Braak, C. T., and Thissen, J. (1995). The Effects of Car Traffic on Breeding Bird Populations in woodland. III. Reduction of Density in Relation to the Proximity of Main Roads. J. Appl. Ecol. 32, 187–202. doi:10.2307/2404428
Reijnen, R., and Foppen, R. (1995). The Effects of Car Traffic on Breeding Bird Populations in woodland. IV. Influence of Population Size on the Reduction of Density Close to a Highway. J. Appl. Ecol. 32, 481–491. doi:10.2307/2404646
Richards, S. A. (2005). Testing Ecological Theory Using the Information-Theoretic Approach: Examples and Cautionary Results. Ecology 86, 2805–2814. doi:10.1890/05-0074
Rosa, C. A. d., and Bager, A. (2012). Seasonality and Habitat Types Affect Roadkill of Neotropical Birds. J. Environ. Manage. 97, 1–5. doi:10.1016/j.jenvman.2011.11.004
Säumel, I., Weber, F., and Kowarik, I. (2016). Toward Livable and Healthy Urban Streets: Roadside Vegetation Provides Ecosystem Services where People Live and Move. Environ. Sci. Pol. 62, 24–33. doi:10.1016/j.envsci.2015.11.012
Seiler, A. (2001). Ecological Effects of Roads: A Review. Uppsala: Swedish University of Agricultural Sciences.
Slater, F. M. (2002). An Assessment of Wildlife Road Casulties - the Potential Discrepancy between Numbers Counted and Numbers Killed. Web Ecol. 3, 33–42. doi:10.5194/we-3-33-2002
Sodhi, N. S., Briffett, C., Kong, L., and Yuen, B. (1999). Bird Use of Linear Areas of a Tropical City: Implications for Park Connector Design and Management. Landscape Urban Plann. 45, 123–130. doi:10.1016/s0169-2046(99)00028-6
Spellerberg, I. (1998). Ecological Effects of Roads and Traffic: a Literature Review. Glob. Ecol. Biogeogr. Lett. 7, 317–333. doi:10.1046/j.1466-822x.1998.00308.x
Taylor, B. D., and Goldingay, R. L. (2004). Wildlife Road-Kills on Three Major Roads in north-eastern New South Wales. Wildl. Res. 31, 83–91. doi:10.1071/wr01110
Team, R. C. (2020). R: A Language and Environment for Statistical Computing. Vienna: The R Foundation.
Keywords: infrastructure impacts, threatened species, vertebrate, rural habitats, agriculture, oil palm, forest, land use planning
Citation: Hui TCY, Slade EM and Chong JL (2021) Roadkills in Northern Peninsular Malaysia. Front. Environ. Sci. 9:637462. doi: 10.3389/fenvs.2021.637462
Received: 03 December 2020; Accepted: 08 October 2021;
Published: 22 October 2021.
Edited by:
John A. Cigliano, Cedar Crest College, United StatesReviewed by:
Chong Leong Puan, Putra Malaysia University, MalaysiaCopyright © 2021 Hui, Slade and Chong. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tabitha C. Y. Hui, dGFiaXRoYS5odWlAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.