 Alison E. King
Alison E. King Genevieve A. Ali
Genevieve A. Ali Adam W. Gillespie
Adam W. Gillespie Claudia Wagner-Riddle
Claudia Wagner-Riddle
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Environ. Sci. , 08 May 2020
Sec. Soil Processes
Volume 8 - 2020 | https://doi.org/10.3389/fenvs.2020.00050
This article is part of the Research Topic Frontiers in Environmental Science – Editor’s Picks 2021 View all 25 articles
The positive effect of soil organic matter (SOM) on crop yield has historically been attributed to the ability of SOM to supply crops with nitrogen and water. Whether management-induced increases in SOM meaningfully supplement water supply has received recent scrutiny, introducing uncertainty to the mechanisms by which SOM benefits crops. Here, we posit that to benefit crops SOM does not need to increase the supply of a growth-limiting resource; it only needs to facilitate root access to extant resource stocks. We highlight evidence for the ability of SOM to alleviate negative impacts of inadequate aeration (mainly waterlogging) and compaction on roots. Waterlogging, even if transient, can permanently downregulate root biosynthesis and call for expensive growth of new roots. Management practices that promote SOM reduce waterlogging by accelerating water infiltration and may promote aeration in non-saturated soils. Compaction as a restriction to root development manifests in drying soils, when mechanical impedance (MI) inflates photosynthate required to extend root tips, leading to short, thick, and shallow roots. SOM reduces MI in dry soils and is associated with root channels to subsoil, granting crops access to deep soil water. Both waterlogging and compaction necessitate additional belowground investment per unit resource uptake. In this framework, crop response to SOM depends on interactions of crop susceptibility to inadequate aeration or compaction, soil moisture, and “baseline” soil aeration and compaction status. By exploring the proposition that SOM catalyzes resource uptake by permitting root development, future research may constrain crop yield improvements expected from SOM management.
Despite comprising a small proportion of the mass of agricultural soils, soil organic matter (SOM) is associated with improved soil structure (Feller and Beare, 1997; Six et al., 2000; Dexter et al., 2008; King et al., 2019). Multiple features of agricultural systems limit the extent by which management can alter SOM levels, but managing for even a modicum of increased SOM offers societal benefits, including climate change mitigation from the storage of carbon (C; Paustian et al., 2016; Minasny et al., 2017), a component of SOM, and reduced erosion (Barthès and Roose, 2002). Another, potential benefit of SOM is increasing crop yield (Pan et al., 2009; Oldfield et al., 2018), however, we have limited ability to explain inconsistent effects of management-induced increases in SOM on crops (Xin et al., 2016; Bradford et al., 2019; Wade et al., 2020) or to constrain potential crop benefits from SOM in future climates, as crop stressors shift (IPCC, 2019). Progress on these fronts relies on a sound understanding of mechanisms through which SOM benefits crops in the first place.
Historically, improved crop yield from SOM has primarily been attributed to SOM’s role in resource supply, either of water or nitrogen (Gregorich et al., 1994; Arshad and Martin, 2002; Lal, 2020). A large effect of SOM on available water capacity (AWC) was established without consideration of limits of management-induced SOM (Hudson, 1994). A recent synthesis, however, concluded that management-induced increases in SOM (10 g C/kg soil) effected on average only 1.16 mm additional AWC in the top 10 cm of soil (Minasny and McBratney, 2018). This is a limited contribution to crop transpiration, which can exceed 450 mm (Kimball et al., 2019, maize). If a 1.16 mm increase in SOM-derived AWC is multiplied by rain events during crop maturation, signifying the number of times AWC is “used,” and including 10–20 cm soil, the augmentation of AWC by SOM is larger, but an effect on crop performance is likely context-dependent. While SOM is linked to AWC and management increases AWC in some situations, crop water supply does not appear well-justified as a universal mechanism linking management-induced SOM increases to crop yield.
Larger SOM pools are linked to higher production rates of plant-available nitrogen (N) via net N mineralization (Schimel, 1986; Booth et al., 2005), leading to the view that SOM benefits crops by increasing N supply. However, to determine that N supply from SOM limits crop growth, rates of plant N uptake should approach rates of N mineralization, indicating a potential for crop N demand that outpaces soil N supply. Results of this comparison depend on method, with net N mineralization deceeding (Brye et al., 2003; Loecke et al., 2012) but gross N mineralization exceeding (Osterholz et al., 2016) crop N demand. If gross N mineralization is indeed a better indicator of plant-available N than net N mineralization (Schimel and Bennett, 2004), or an underestimate given crop uptake of amino acids (Hill et al., 2011), then increasing SOM would simply be increasing an already sufficient N supply.
Here, we posit that SOM benefits crops not necessarily by increasing resource supply but by catalyzing crop resource capture (Figure 1). Roots, the locus of nutrient and water uptake, rely on contact with soil for nutrient (Wang et al., 2006), and water uptake (Javot and Maurel, 2002). Vigorous root development therefore multiplies the surface area of root-soil contact through which resource uptake occurs. We highlight below the consequences of inadequate aeration and compaction on roots and the ability of SOM-enhancing management practices to alleviate these stressors. While we acknowledge that relationships between SOM and soil structure are known (Karlen et al., 2001; Bünemann et al., 2018), we argue that their connections to related crop stressors—poor aeration and compaction—have received inadequate attention as mechanisms that link SOM to crops. We also acknowledge that compaction and inadequate aeration are often co-located, and we separate them to discuss how they manifest differently in the soil environment and as root stressors.
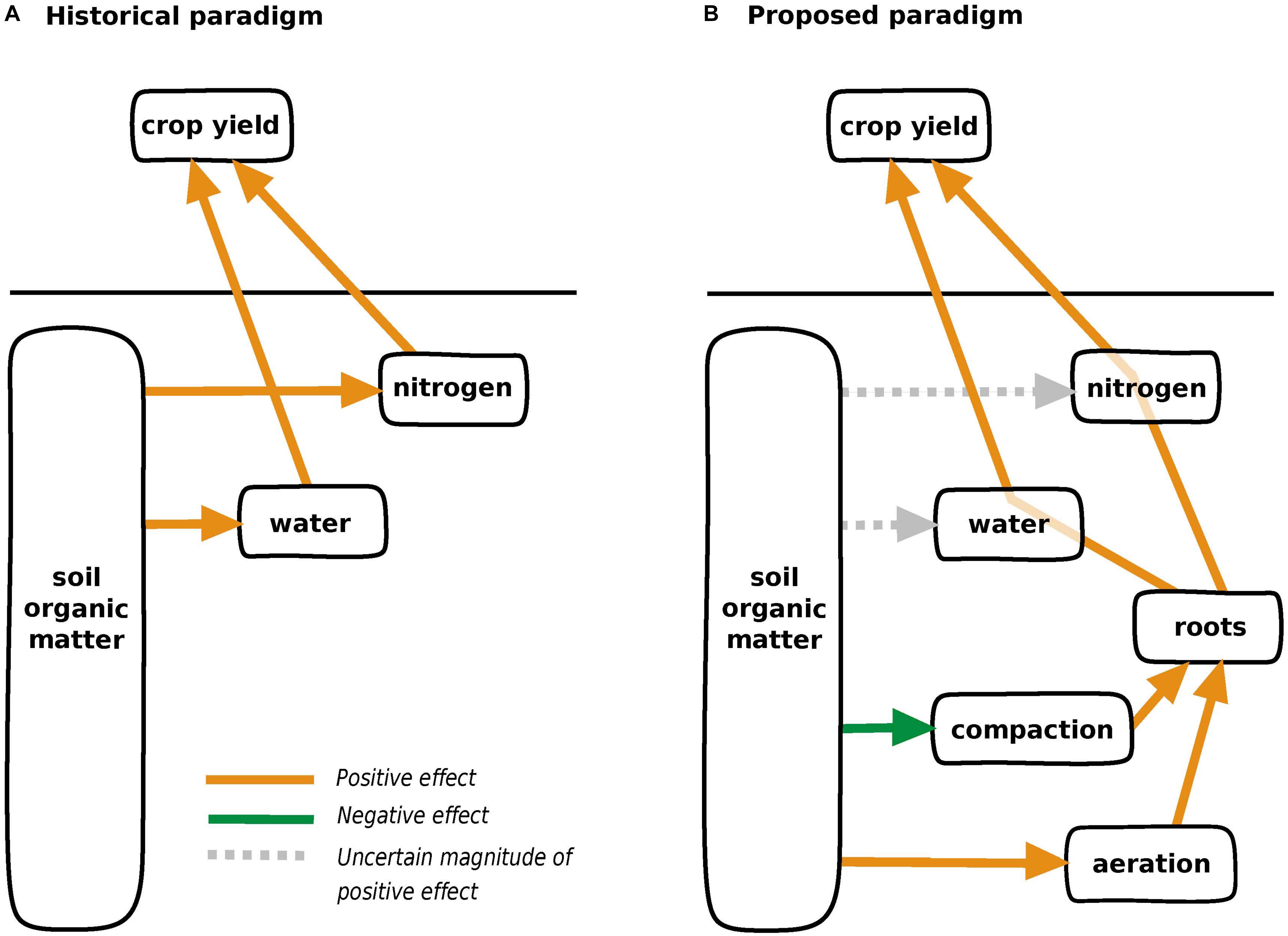
Figure 1. Conceptual models representing (A) historically acknowledged and (B) newly proposed mechanisms linking management-induced increases in soil organic matter with crop yield. We recognize that the effect of SOM in mediating soil structure has been known for decades, but we argue that exploring the response of crops to related stressors (inadequate aeration and compaction) has potential to explain effects of SOM on crop yield.
We refer to differences in SOM induced by agricultural practices, such as cover crops (Poeplau and Don, 2015), perennial forages (King and Blesh, 2018), manure application (Maillard and Angers, 2014), straw retention (Liu et al., 2014), or reduced tillage (in surface soil, Luo et al., 2010), except where noted. Although some of the mechanisms discussed may apply to naturally occurring variability in SOM, we defer their discussion for brevity.
As a substrate for respiration, O2 and its transport potentially affect all functions of crop roots (Grable, 1966). To study the effects of O2 deficiency in the field, researchers commonly impose an extreme constriction of aeration with waterlogging, the saturation of soil pores with water (Hodgson and Chan, 1982; Bange et al., 2004). Our consideration of inadequate aeration focuses accordingly on waterlogging, and we do not review impacts of inadequate aeration in non-saturated soils on crops, as these remain largely unexplored.
Waterlogging damages diverse crops worldwide (Velde and Van Der Tubiello, 2012; Shaw et al., 2013; Zhang et al., 2015; Li et al., 2019), and increases in waterlogging due to climate change are forecasted to exacerbate these damages (Rosenzweig et al., 2002). Waterlogging reduces soil aeration due to a 104 fold slower rate of O2 diffusion through water than through air (Grable, 1966). Within hours, soil O2 can be depleted as root or soil organisms’ respiration demands exceed atmospheric O2 supply, with rate and extent of O2 depletion depending on depth (Malik et al., 2001) and temperature (Trought and Drew, 1982). Over slightly longer time frames, waterlogged soils also accumulate byproducts of root or microbial metabolism, e.g., CO2, Fe2+, and Mn2+, potential plant toxins (Shabala, 2011). Crop responses to waterlogging depend on crop species and cultivar (Huang, 1997; Boru, 2003; Ploschuk et al., 2018), as well as timing (Rhine et al., 2010; de San Celedonio et al., 2014; Ren et al., 2014) and duration (Malik et al., 2002; Rhine et al., 2010; Kuai et al., 2014; Ren et al., 2014; Arduini et al., 2019) of waterlogging, but a well-supported view holds that crop yield reductions from waterlogging are largely attributable to impaired function and inadequate recovery of roots (Malik et al., 2002; Herzog et al., 2016; Arduini et al., 2019).
In crops that are susceptible to waterlogging, stress response can be considered both during and after release from waterlogging. During waterlogging, O2 deficiency induces an energy crisis in the root due to inefficient production of ATP (Gibbs and Greenway, 2003), which curtails energy-dependent nutrient uptake (Trought and Drew, 1980; Morard et al., 2000; Colmer and Greenway, 2011) and root growth (Palta et al., 2010; Arduini et al., 2019). After release from waterlogging, root growth of certain root forms, i.e., seminal roots, can be permanently inhibited (Malik et al., 2002; Palta et al., 2010; Colmer and Greenway, 2011), attributable to cell death in apical meristems (Trought and Drew, 1980; Malik et al., 2002), beyond the reach of plant-transported O2 (Colmer and Greenway, 2011). Crops must then rely on energetically-expensive production of new roots, i.e., adventitious roots (Palta et al., 2010; Steffens and Rasmussen, 2016), and/or increase nutrient uptake per unit root (Arduini et al., 2019). These adaptations do not necessarily allow crops to escape a waterlogging yield penalty, as reductions in root biomass (Grassini et al., 2007; de San Celedonio et al., 2017; Ploschuk et al., 2018) or root length density (Hayashi et al., 2013) are often linked to reductions in shoot biomass or yield.
Uncertainty remains about the threshold duration at which waterlogging damages crops. In some crops and growth stages, waterlogging as short as 3 days reduced yield (Malik et al., 2002; Ren et al., 2014), but the shortest waterlogging we found in field studies was 2 days (Rhine et al., 2010). Due to the risk of crop damage from even transient waterlogging, there is interest in management practices that reduce waterlogging risk (Manik et al., 2019) and improve root aeration across the soil water spectrum (Rabot et al., 2018).
Management practices that promote SOM reduce risk and duration of waterlogging by increasing rate of water infiltration (Boyle et al., 1989; Adekalu et al., 2007; Abid and Lal, 2009; Blanco-Canqui et al., 2011), which increases the time soil can receive rain before ponding occurs (McGarry et al., 2000) and reduces time required to drain from saturation to field capacity (Wuest et al., 2005). Among the many measurable soil water variables, infiltration is the most commonly assessed, and we note the need to better establish relationships between infiltration, time to ponding, and drainage. It is also important to note, as reviewed by Blanco-Canqui and Ruis (2018) for no-till, that management practices that promote SOM can have a neutral effect on water infiltration in some cases, despite positive effects in majority of cases.
Accelerated infiltration associated with SOM is attributable to several soil features. The redistribution of soil mass to larger aggregate size classes associated with SOM (King et al., 2019) helps to explain an increase in total (Pikul and Zuzel, 1994; Yang et al., 2011; Blanco-Canqui and Benjamin, 2013) or macro- porosity (>0.3–0.4 mm, Deurer et al., 2009; Yagüe et al., 2016), although this effect is not detectable in all cases (Ruiz-Colmenero et al., 2013). SOM also stabilizes aggregates (Chenu et al., 2000; Annabi et al., 2011), minimizing their dissolution into smaller, and pore-clogging size fractions that seal the soil surface against water infiltration (Bissonnais and Arrouays, 1997; Lado et al., 2004). The most dramatic effects of SOM on infiltration can likely be traced to earthworms and/or termites and their creation of wide, continuous, vertically-oriented pores through which water flows preferentially (McGarry et al., 2000; Guo and Lin, 2018). More abundant – or more active (Pérès et al., 2010) – soil fauna may be due in part to reduced disturbance associated with some SOM-promoting practices, e.g., no-till. However, close relationships between SOM and earthworm abundance without the confounding effect of disturbance (Fonte et al., 2009; Guo et al., 2016) also indicate a role for SOM as faunal substrate supply.
Few studies have attempted to link SOM-induced reductions in waterlogging with crop yield. Gómez-paccard et al. (2015), however, find crop yields increased from reduced surface soil waterlogging associated with no-till. The ability of SOM-mediated reductions in waterlogging to benefit crops are most likely when (1) crop is sensitive to waterlogging and (2) rainfall intensity can be mediated by SOM on a timescale relevant to waterlogging stress (neither drizzle nor deluge); and (3) soil is otherwise poorly-drained (Rhine et al., 2010).
If crops experience inadequate aeration in non-saturated soils, it is reasonable to expect that SOM would improve gas diffusivity given its effects on related parameters of soil structure (Neira et al., 2015, and below). Few studies investigate the isolated effect of SOM on gas diffusivity, however, Colombi et al. (2019) find a positive relationship between SOM and gas diffusivity at field capacity across a soil texture gradient. Future work should examine net effects of SOM on O2 diffusivity and consumption in soils.
Soil compaction reduces crop yields (Coelho et al., 2000; Ishaq et al., 2001a; Bayhan et al., 2002; Czyz, 2004; Whalley et al., 2008) and is quantified via either bulk density or mechanical impedance (MI; Ehlers et al., 1987; Bengough et al., 2011). MI estimates the force encountered by the elongation of a living root, and is consequential for crops because greater MI inflates the photosynthate required for root elongation (Herrmann and Colombi, 2019). Although MI measurements ignore biopores used preferentially by roots (Stirzaker et al., 1996; White and Kirkegaard, 2010), MI is more descriptive than bulk density because it is sensitive to soil water. Drying soils present increasing MI (Vaz et al., 2011), and to isolate effects of water stress from compaction stress per se on crop development, researchers use experimental compaction.
Compaction studies indicate that reduced crop yield from compaction is due in large part to constraints on root development (Ishaq et al., 2001b; Czyz, 2004; Colombi and Keller, 2019). A root restricted by soil compaction is generally thicker than a root in non-compacted soil (Nadian et al., 1997), likely due to greater axial force needed to overcome compaction (Bengough, 2012). Reductions in total number of roots, rate of root elongation, total root length, or root biomass are also reported (Panayiotopoulos et al., 1994; Chan et al., 2006; Lipiec et al., 2012). Root length is generally more reduced than root dry mass (Panayiotopoulos et al., 1994), indicating the accumulation of belowground photosynthate without commensurate expansion of soil-contacting surface area available for nutrient and water uptake. Compaction is sometimes characterized by a hardpan around 20 cm depth, which leads to restricted root access to subsoil and concentrated root development in the topsoil (Czyz, 2004). This pattern of root development prevents crop access of deep soil water most implicated in crop drought resistance (Uga et al., 2013; Lynch, 2018).
A single threshold MI for crop sensitivity is unlikely to serve universally, and not only because cultivars (Houlbrooke et al., 1997) and crops (Rosolem et al., 2002) differ in MI tolerance. Threshold MI values also likely depend on definition by energy required to extend roots (Herrmann and Colombi, 2019) or by crop yield penalty. The MI required to reduce root growth efficiency is likely less than required to affect yields, and yield penalty due to restricted roots can be counteracted somewhat by fertilization (Robertson et al., 2009). Whatever the threshold, the detrimental effects of compaction on crops has generated attention toward means to reduce it.
Although SOM is often promoted for its ability to alleviate soil compaction and associated increases in MI (Hamza and Anderson, 2005), the generation of data confirming a negative SOM-MI relationship has been hampered by the convention of measuring MI in soils near field capacity (Duiker, 2002). Even in large datasets, no relationship between SOM and MI in soil near field capacity is found (Fine et al., 2017), likely because very wet soils (∼1–10 kPa) offer minimal MI regardless of SOM. It is as MI increases in drying soils (Vaz et al., 2011; Filho et al., 2014) that an effect of SOM becomes apparent (Stock and Downes, 2008; Gao et al., 2012).
We highlight two studies showing the effect of SOM in reducing MI as soils dry. Stock and Downes (2008) and Gao et al. (2012) investigated soils differing only in SOM concentrations. With few exceptions, MI increased as soils approached permanent wilting point, but the increase in MI was not as much in higher SOM soils. In other words, SOM allows soil to become drier before reaching a potentially root-constraining MI. For instance, Stock and Downes (2008) find an MI of 1.5 MPa is reached in the 1% OM soil at ∼-100 kPa, whereas the 3% OM soil reaches the same MI at about ∼-200 kPa. For these soils, the difference in volumetric water content between -100 and -200 kPa is ∼0.02 m–3 H2O m–3 soil, or ∼5 mm of water when considered over the top 25 cm. While Stock and Downes (2008) added organic amendments to glacial till to create fixed SOM percentages, their results resemble those of Gao et al. (2012), who compared fallow to grassland soils. The extent to which management-induced SOM benefits crops via reductions in compaction likely vary with context, particularly those relevant to MI thresholds (see section “Soil compaction constrains root development”).
Although not connected to the physical or biological properties of SOM, management practices that promote SOM may also alleviate the effects of compaction by facilitating crop root access to the subsoil. Deep-rooted cover crops or perennial crops create root channels to subsoil (McCallum et al., 2004), which are used by subsequent cash crops (Rasse and Smucker, 1998; Williams and Weil, 2004). Crops are most likely to benefit from these root channels if subsoil is compacted and if crops experience sufficient water stress for subsoil water stores to be relevant.
In the historical conceptual model linking crop performance to SOM, SOM benefits crops primarily by supplying nitrogen and water. Here we propose SOM as a mediator of resource uptake via root growth. We note caveats to the proposed framework. The extent to which SOM affects nutrient and water supply still merits research, and not all mechanisms discussed are contingent on increased SOM. We focus on MI to characterize compaction, but SOM may alter soil structure in ways relevant to root growth that are not captured by MI. As a simple diagram, Figure 1 does not depict that yield penalty in low SOM soils may be due to cost of constructing new or thicker roots. However, considering SOM as catalyzing resource uptake via root development can help explain recent reports of SOM effects on crops. Wade et al. (2020) report maize yield increases from management-induced SOM across a range of N fertilizer levels, consistent with the concept that root N uptake—as well as soil N supply—can limit crop yield. Recognizing the importance of aeration for roots may also explain a parabolic crop response to a SOM gradient in a pot study (Oldfield et al., 2020), in which synthesized mixtures of minerals and organic horizons higher in SOM may have supported higher O2 consumption by microbes without improved gas diffusivity expected in natural high-SOM soils (Colombi et al., 2019).
We propose the exploration of SOM as a catalyst for resource capture focuses on:
• Characterizing the context-mediated effect of SOM on aeration and compaction by describing the effect of SOM gradients on (a) aeration and risk and duration of waterlogging and (b) MI across a range of soil moisture contents.
• Investigating crop response to SOM as a function of aeration and compaction by identifying the thresholds of hypoxia affecting roots and yields. To relate SOM-mediated MI to root development, describing soil moisture status during crop maturation will be crucial.
• Simulating future crop response to SOM; currently, Basche et al. (2016) and Jarecki et al. (2018) are two of few examples to model the potential for management-induced SOM to stabilize crop yields under future climates.
We hope the lens proposed will help illuminate the effect of SOM on crops.
AK conceptualized and drafted the manuscript. GA, AG, and CW-R provided input to arrive at the final version.
This work was funded by a Natural Sciences and Engineering Research Council of Canada Discovery Grant to CW-R.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Peter Pellitier for feedback on an early version of the manuscript and Rebecca Johnson for comments that improved the figure. Two reviewers identified careful distinctions in the literature review.
Abid, M., and Lal, R. (2009). Tillage and drainage impact on soil quality: II. Tensile strength of aggregates, moisture retention and water infiltration. Soil Tillage Res. 103, 364–372.
Adekalu, K. O., Olorunfemi, I. A., and Osunbitan, J. A. (2007). Grass mulching effect on infiltration, surface runoff and soil loss of three agricultural soils in Nigeria. Bioresour. Technol. 98, 912–917. doi: 10.1016/j.biortech.2006.02.044
Annabi, M., Bissonnais, Y. L., Villio-Poitrenaud, M. L., and Houot, S. (2011). Improvement of soil aggregate stability by repeated applications of organic amendments to a cultivated silty loam soil. Agricu. Ecosyst. Environ. 144, 382–389.
Arduini, I., Baldanzi, M., and Pampana, S. (2019). Reduced growth and nitrogen uptake during waterlogging at tillering permanently affect yield components in late sown oats. Front. Plant Sci. 10:1087. doi: 10.3389/fpls.2019.01087
Arshad, M. A., and Martin, S. (2002). Identifying critical limits for soil quality indicators in agro-ecosystems. Agric. Ecosyst. Environ. 88, 153–160.
Bange, M. P., Milroy, S. P., and Thongbai, P. (2004). Growth and yield of cotton in response to waterlogging. Field Crops Res. 88, 129–142. doi: 10.1093/pcp/pcp163
Barthès, B., and Roose, E. (2002). Aggregate stability as an indicator of soil susceptibility to runoff and erosion; validation at several levels. Catena 47, 133–149.
Basche, A. D., Archontoulis, S. V., Kaspar, T. C., Jaynes, D. B., Parkin, T. B., and Miguez, F. E. (2016). Simulating long-term impacts of cover crops and climate change on crop production and environmental outcomes in the Midwestern United States. Agric. Ecosyst. Environ. 218, 95–106.
Bayhan, Y., Kayisoglu, B., and Gonulol, E. (2002). Effect of soil compaction on sunflower growth. Soil Tillage Res. 68, 31–38.
Bengough, A. G. (2012). Root elongation is restricted by axial but not by radial pressures: so what happens in field soil? Plant Soil 360, 15–18.
Bengough, A. G., Mckenzie, B. M., Hallett, P. D., and Valentine, T. A. (2011). Root elongation, water stress, and mechanical impedance: a review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 62, 59–68. doi: 10.1093/jxb/erq350
Bissonnais, Y. L., and Arrouays, D. (1997). Aggregate stability and assessment of soil crustability and erodibility: II. Application to humic loamy soils with various organic carbon contents. Eur. J. Soil Sci. 48, 39–48.
Blanco-Canqui, H., and Benjamin, J. G. (2013). “Impacts of soil organic carbon on soil physical behavior,” in Quantifying and Modeling Soil Structure Dynamics, eds S. Logsdon, M. Berli, and R. Horn (Madison, WI), 11–40.
Blanco-Canqui, H., Mikha, M. M., Presley, D. R., and Claassen, M. M. (2011). Addition of cover crops enhances no-till potential for improving soil physical properties. Soil Sci. Soc. Am. J. 75, 1471–1482.
Blanco-Canqui, H., and Ruis, S. J. (2018). No-tillage and soil physical environment. Geoderma 326, 164–200.
Booth, M. S., Stark, J. M., and Rastetter, E. (2005). Controls on nitrogen cycling in terrestrial ecosystems: a synthetic analysis of literature data. Ecol. Monogr. 75, 139–157.
Boru, G. (2003). Responses of soybean to oxygen deficiency and elevated root-zone carbon dioxide concentration. Ann. Bot. 91, 447–453. doi: 10.1093/aob/mcg040
Boyle, M., Frankenberger, W. T., and Stolzy, L. H. (1989). Influence of organic matter on soil aggregation and water infiltration. J. Prod. Agric. 2, 290–299.
Bradford, M. A., Carey, C. J., Atwood, L., Bossio, D., Fenichel, E. P., Gennet, S., et al. (2019). Soil carbon science for policy and practice. Nat. Sustainabil. 2, 1070–1072. doi: 10.1007/s13280-016-0773-x
Brye, K. R., Norman, J. M., Gower, S. T., and Bundy, L. G. (2003). Effects of management practices on annual net N-mineralization in a restored prairie and maize agroecosystems. Biogeochemistry 63, 135–160.
Bünemann, E. K., Bongiorno, G., Bai, Z., Creamer, R. E., De Deyn, G., de Goede, R., et al. (2018). Soil quality – A critical review. Soil Biol. Biochem. 120, 105–125.
Chan, K. Y., Oates, A., Swan, A. D., Hayes, R. C., Dear, B. S., and Peoples, M. B. (2006). Agronomic consequences of tractor wheel compaction on a clay soil. Soil Tillage Res. 89, 13–21.
Chenu, C., Le Bissonnais, Y., and Arrouays, D. (2000). Organic matter influence on clay wettability and soil aggregate stability. Soil Sci. Soc. Am.J. 64:1479.
Coelho, M. B., Mateos, L., and Villalobos, F. J. (2000). Influence of a compacted loam subsoil layer on growth and yield of irrigated cotton in Southern Spain. Soil Tillage Res. 57, 129–142.
Colmer, T. D., and Greenway, H. (2011). Ion transport in seminal and adventitious roots of cereals during O2 deficiency. J. Exp.Bot. 62, 39–57. doi: 10.1093/jxb/erq271
Colombi, T., and Keller, T. (2019). Developing strategies to recover crop productivity after soil compaction — A plant eco-physiological perspective. Soil Tillage Res. 191, 156–161.
Colombi, T., Walder, F., Büchi, L., Sommer, M., Liu, K., Six, J., et al. (2019). On-farm study reveals positive relationship between gas transport capacity and organic carbon content in arable soil. Soil 5, 91–105.
Czyz, E. A. (2004). Effects of traffic on soil aeration, bulk density and growth of spring barley. Soil Tillage Res. 79, 153–166.
de San Celedonio, R. P., Abeledo, L. G., Mantese, A. I., and Miralles, D. J. (2014). Identifying the critical period for waterlogging on yield and its components in wheat and barley. Plant Soil 378, 265–277.
de San Celedonio, R. P., Abeledo, L. G., Mantese, A. I., and Miralles, D. J. (2017). Differential root and shoot biomass recovery in wheat and barley with transient waterlogging during preflowering. Plant Soil 417, 481–498.
Deurer, M., Grinev, D., Young, I., Clothier, B. E., and Muller, K. (2009). The impact of soil carbon management on soil macropore structure: a comparison of two apple orchard systems in New Zealand. Eur. J. Soil Sci. 60, 945–955. doi: 10.2134/jeq2007.0508
Dexter, A. R., Richard, G., Arrouays, D., Czyż, E. A., Jolivet, C., and Duval, O. (2008). Complexed organic matter controls soil physical properties. Geoderma 144, 620–627.
Duiker, S. W. (2002). Diagnosing Soil Compaction Using a Penetrometer. University Park, PA: College of Agricultural Sciences.
Ehlers, W., Kopke, U., Hesse, F., and Bohm, W. (1987). Penetration resistance and root growth of oats in tilled and untilled loess soil. Soil Tillage Res. 3, 261–275.
Feller, C., and Beare, M. H. (1997). Physical control of soil organic matter dynamics in the tropics. Geoderma 79, 69–116.
Filho, E. A. M., da Silva, A. P., Figueiredo, G. C., Gimenes, F. H. S., and Vitti, A. C. (2014). Compared performance of penetrometers and effect of soil water content on penetration resistance measurements. Rev. Brasil. Ciên. Solo 38, 744–754.
Fine, A. K., van, E. S. H. M., and Schindelbeck, R. R. (2017). Statistics, scoring functions, and regional analysis of a comprehensive soil health database. Soil Sci. Soc. Am. J. 81, 589–601.
Fonte, S. J., Winsome, T., and Six, J. (2009). Earthworm populations in relation to soil organic matter dynamics and management in California tomato cropping systems. Appl. Soil Ecol. 41, 206–214.
Gao, W., Watts, C. W., Ren, T., and Whalley, W. R. (2012). The effects of compaction and soil drying on penetrometer resistance. Soil Tillage Res. 125, 14–22.
Gibbs, J., and Greenway, H. (2003). Mechanisms of anoxia tolerance in plants. I. Growth, survival and anaerobic catabolism. Func. Plant Biol. 30, 1–47.
Gómez-paccard, C., Hontoria, C., Mariscal-sancho, I., Pérez, J., León, P., González, P., et al. (2015). Soil – water relationships in the upper soil layer in a Mediterranean Palexerult as affected by no-tillage under excess water conditions – Influence on crop yield. Soil Tillage Res. 146, 303–312.
Grassini, P., Indaco, G. V., Hall, A. J., and Tra, N. (2007). Responses to short-term waterlogging during grain filling in sunflower. Field Crops Res. 101, 352–363.
Gregorich, E. G., Carter, M. R., Angers, D. A., Monreal, C. M., and Ellert, B. H. (1994). Towards a minimum data set to assess soil organic matter quality in agricultural soils. Can. J. Soil Sci. 74, 367–385.
Guo, L., Wu, G., Li, Y., Li, C., Liu, W., Meng, J., et al. (2016). Effects of cattle manure compost combined with chemical fertilizer on topsoil organic matter, bulk density and earthworm activity in a wheat-maize rotation system in Eastern China. Soil Tillage Res. 156, 140–147.
Guo, L., and Lin, H. (2018). “Addressing two bottlenecks to advance the understanding of preferential flow in soils,” in Advances in Agronomy, Vol. 147 (Amsterdam: Elsevier Inc), 61–117.
Hamza, M. A., and Anderson, W. K. (2005). Soil compaction in cropping systems: a review of the nature, causes and possible solutions. Soil Tillage Res. 82, 121–145.
Hayashi, T., Yoshida, T., Fujii, K., and Mitsuya, S. (2013). Maintained root length density contributes to the waterlogging tolerance in common wheat (Triticum aestivum L.). Field Crops Res. 152, 27–35.
Herrmann, A. M., and Colombi, T. (2019). Energy use efficiency of root growth – a theoretical bioenergetics framework. Plant Signal. Behav. 14:1685147. doi: 10.1080/15592324.2019.1685147
Herzog, M., Striker, G. G., Colmer, T. D., and Pedersen, O. (2016). Mechanisms of waterlogging tolerance in wheat – a review of root and shoot physiology. Plant Cell Environ. 39, 1068–1086. doi: 10.1111/pce.12676
Hill, P. W., Quilliam, R. S., Deluca, T. H., Farrar, J., Farrell, M., Roberts, P., et al. (2011). Acquisition and assimilation of nitrogen as peptide- bound and d-enantiomers of amino acids by wheat. PLoS One 6:e19220. doi: 10.1371/journal.pone.0019220
Hodgson, A., and Chan, K. (1982). The effect of short-term waterlogging during furrow irrigation of cotton in a cracking grey clay. Austr. J. Agric. Res. 33, 109–116.
Houlbrooke, D. J., Thom, E. R., Chapman, R., Mclay, C. D. A., Thom, E. R., Chapman, R., et al. (1997). A study of the effects of soil bulk density on root and shoot growth of different ryegrass lines. N. Z. J. Crop Hortic. Sci. 40, 429–435.
Huang, B. (1997). Responses to root-zone CO2 enrichment and hypoxia of wheat genotypes differing in waterlogging tolerance. Crop. Sci. 37, 464–468.
Hudson, B. D. (1994). Soil organic matter and available water capacity. J. Soil Water Conserv. 49, 189–194.
Ishaq, M., Hassan, A., Saeed, M., Ibrahim, M., and Lal, R. (2001a). Subsoil compaction effects on crops in Punjab, Pakistan I. Soil physical properties and crop yield. Soil & Tillage Research 59, 57–65.
Ishaq, M., Ibrahim, M., Hassan, A., Saeed, M., and Lal, R. (2001b). Subsoil compaction effects on crops in Punjab, Pakistan: II. Root growth and nutrient uptake of wheat and sorghum. Soil Tillage Res. 60, 153–161.
Jarecki, M., Grant, B., Smith, W., Deen, B., Drury, C., VanderZaag, A., et al. (2018). Long-term trends in corn yields and soil carbon under diversified crop rotations. J. Environ. Q. 7, 635–643. doi: 10.2134/jeq2017.08.0317
Javot, H., and Maurel, C. (2002). The role of aquaporins in root water uptake. Ann. Bot. 90, 301–313. doi: 10.1093/aob/mcf199
Karlen, D. L., Andrews, S. S., and Doran, J. W. (2001). Soil quality: current concepts and applications. Adv. Agron. 74, 1–40.
Kimball, B. A., Boote, K. J., Hatfield, J. L., Ahuja, L. R., Stockle, C., Archontoulis, S., et al. (2019). Simulation of maize evapotranspiration: an inter-comparison among 29 maize models. Agri. For. Meteorol. 271, 264–284.
King, A. E., and Blesh, J. (2018). Crop rotations for increased soil carbon: perenniality as a guiding principle. Ecol. Appl. 28, 249–261. doi: 10.1002/eap.1648
King, A. E., Congreves, K. A., Deen, B., Dunfield, K. E., Voroney, R. P., and Wagner-Riddle, C. (2019). Quantifying the relationships between soil fraction mass, fraction carbon, and total soil carbon to assess mechanisms of physical protection. Soil Biol. Biochem. 135, 95–107.
Kuai, J., Liu, Z., Wang, Y., Meng, Y., Chen, B., Zhao, W., et al. (2014). Waterlogging during flowering and boll forming stages affects sucrose metabolism in the leaves subtending the cotton boll and its relationship with boll weight. Plant Sci. 223, 79–98. doi: 10.1016/j.plantsci.2014.03.010
Lado, M., Paz, A., and Ben-Hur, M. (2004). Organic matter and aggregate-size interactions in saturated hydraulic conductivity. Soil Sci. Soc. Am. J. 68:234.
Li, Y., Guan, K., Schnitkey, G. D., Delucia, E., and Peng, B. (2019). Excessive rainfall leads to maize yield loss of a comparable magnitude to extreme drought in the United States. Global Change Biol. 25, 2325–2337. doi: 10.1111/gcb.14628
Lipiec, J., Horn, R., Pietrusiewicz, J., and Siczek, A. (2012). Effects of soil compaction on root elongation and anatomy of different cereal plant species. Soil Tillage Res. 121, 74–81.
Liu, C., Lu, M., Cui, J., Li, B., and Fang, C. (2014). Effects of straw carbon input on carbon dynamics in agricultural soils: a meta-analysis. Global Change Biol. 20, 1366–1381. doi: 10.1111/gcb.12517
Loecke, T. D., Cambardella, C. A., and Liebman, M. (2012). Synchrony of net nitrogen mineralization and maize nitrogen uptake following applications of composted and fresh swine manure in the Midwest U.S. Nutr. Cycl. in Agroecosyst. 93, 65–74.
Luo, Z., Wang, E., and Sun, O. J. (2010). Can no-tillage stimulate carbon sequestration in agricultural soils? A meta-analysis of paired experiments. Agric., Ecosyst. Environ. 139, 224–231.
Lynch, J. P. (2018). Rightsizing root phenotypes for drought resistance. J. Exp. Bot. 69, 3279–3292. doi: 10.1093/jxb/ery048
Maillard, É, and Angers, D. A. (2014). Animal manure application and soil organic carbon stocks: a meta-analysis. Global Change Biol. 20, 666–679. doi: 10.1111/gcb.12438
Malik, A. I., Colmer, T. D., Lambers, H., and Schortemeyer, M. (2001). Changes in physiological and morphological traits of roots and shoots of wheat in response to different depths of waterlogging. Aust. J. Plant Physiol. 28, 1121–1131.
Malik, A. I., Colmer, T. D., Lambers, H., Setter, T. L., and Schortemeyer, M. (2002). Short-term waterlogging has long-term effects on the growth and physiology of wheat. New Phytol. 4, 225–236.
Manik, S. M. N., Pengilley, G., Dean, G., Field, B., Shabala, S., and Zhou, M. (2019). Soil and crop management practices to minimize the impact of waterlogging on crop productivity. Front. Plant Sci. 10:140. doi: 10.3389/fpls.2019.00140
McCallum, M. H. A., Kirkegaard, J. A. A., Green, T. W. B., Cresswell, H. P. B., Davies, S. L. A., Angus, J. F. A., et al. (2004). Improved subsoil macroporosity following perennial pastures. Austr. J. Exp. Agric. 44, 299–307.
McGarry, D., Bridge, B. J., and Radford, B. J. (2000). Contrasting soil physical properties after zero and traditional tillage of an alluvial soil in the semi-arid subtropics. Soil Tillage Res. 53, 105–115.
Minasny, B., Malone, B. P., McBratney, A. B., Angers, D. A., Arrouays, D., Chambers, A., et al. (2017). Soil carbon 4 per mille. Geoderma 292, 59–86. doi: 10.1016/j.scitotenv.2017.12.263
Minasny, B., and McBratney, A. B. (2018). Limited effect of organic matter on soil available water capacity. Eur. J. Soil Sci. 69, 39–47.
Morard, P., Lacoste, L., Silvestre, J., Morard, P., Lacoste, L., and Silvestre, J. (2000). Effect of oxygen deficiency on uptake of water and mineral nutrients by tomato plants in soilless culture. J. Plant Nutr. 23, 1063–1078.
Nadian, H., Smith, S. E., Alston, A. M., and Murray, R. S. (1997). Effects of soil compaction on plant growth, phosphorus uptake and morphological characteristics of vesicular-arbuscular mycorrhizal colonization of Trifolium suhterraneum. New Phytol. 135, 303–311.
Neira, J., Ortiz, M., Morales, L., and Acevedo, E. (2015). Oxygen diffusion in soils: Understanding the factors and processes needed for modeling. Chil. J. Agric. Res. 75, 35–44.
Oldfield, E. E., Bradford, M. A., and Wood, S. A. (2018). Global meta-analysis of the relationship between soil organic matter and crop yields. Soil 5, 15–32.
Oldfield, E. E., Woods, S. A., and Bradford, M. A. (2020). Direct evidence using a controlled greenhouse study for threshold effects of soil organic matter on crop growth. Ecol. Appl. e02073 (in press). doi: 10.1002/eap.2073
Osterholz, W. R., Rinot, O., Liebman, M., and Castellano, M. J. (2016). Can mineralization of soil organic nitrogen meet maize nitrogen demand? Plant Soil 451, 73–84.
Palta, J. A., Ganjeali, A., Turner, N. C., and Siddique, K. H. M. (2010). Effects of transient subsurface waterlogging on root growth, plant biomass and yield of chickpea. Agric. Water Manag. 97, 1469–1476.
Pan, G., Smith, P., and Pan, W. (2009). The role of soil organic matter in maintaining the productivity and yield stability of cereals in China. Agric. Ecosyst. Environ. 129, 344–348.
Panayiotopoulos, K. P., Papadopoulou, C. P., and Hatjiioannidou, A. (1994). Compaction and penetration resistance of an Alfisol and Entisol and their influence on root growth of maize seedlings. Soil Tillage Res. 31, 323–337.
Paustian, K., Lehmann, J., Ogle, S., Reay, D., Robertson, G. P., and Smith, P. (2016). Climate-smart soils. Nature 532, 49–57. doi: 10.1038/nature17174
Pérès, G., Bellido, A., Curmi, P., Marmonier, P., and Cluzeau, D. (2010). Relationships between earthworm communities and burrow numbers under different land use systems. Pedobiol. Int.J. Soil Biol. 54, 37–44.
Pikul, J. L., and Zuzel, J. F. (1994). Soil crusting and water infiltration affected by long-term tillage and residue management. Soil Sci. Soc. Am. J. 58, 1524–1530.
Ploschuk, R. A., Miralles, D. J., Colmer, T. D., Ploschuk, E. L., and Striker, G. G. (2018). Waterlogging of winter crops at early and late stages: impacts on leaf physiology, growth and yield. Front. Plant Sci. 9:1863. doi: 10.3389/fpls.2018.01863
Poeplau, C., and Don, A. (2015). Carbon sequestration in agricultural soils via cultivation of cover crops: a meta-analysis. Agric. Ecosyst. Environ. 200, 33–41.
Rabot, E., Wiesmeier, M., Schlüter, S., and Vogel, H. J. (2018). Soil structure as an indicator of soil functions: a review. Geoderma 314, 122–137.
Rasse, D. P., and Smucker, A. J. M. (1998). Root recolonization of previous root channels in corn and alfalfa rotations. Plant Soil 204, 203–212.
Ren, B., Zhang, J., Li, X., Fan, X., Dong, S., Liu, P., et al. (2014). Effects of waterlogging on the yield and growth of summer maize under field conditions. Can. J. f Plant Sci. 94, 23–31.
Rhine, D. M., Shannon, G., Wrather, A. E. A., and Sleper, A. E. D. (2010). Yield and nutritional responses to waterlogging of soybean cultivars. Irrig. Sci. 28, 135–142.
Robertson, D., Zhang, H., Palta, J. A., Colmer, T., and Turner, N. C. (2009). Waterlogging affects the growth, development of tillers, and yield of wheat through a severe, but transient, N deficiency. Crop Pasture Sci. 60, 578–586.
Rosenzweig, C., Tubiello, F. N., Goldberg, R., Mills, E., and Bloomfield, J. (2002). Increased crop damage in the US from excess precipitation under climate change. Global Environ. Change 12, 197–202.
Rosolem, C., Foloni, J. S., and Tiritan, C. (2002). Root growth and nutrient accumulation in cover crops as affected by soil compaction. Soil Tillage Res. 65, 109–115.
Ruiz-Colmenero, M., Bienes, R., Eldridge, D. J., and Marques, M. J. (2013). Vegetation cover reduces erosion and enhances soil organic carbon in a vineyard in the central Spain. Catena 104, 153–160.
Schimel, D. S. (1986). Carbon and nitrogen turnover in adjacent grassland and cropland ecosystems. Biogeochemistry 357, 345–357.
Schimel, J. P., and Bennett, J. (2004). Nitrogen mineralization: challenges of a changing paradigm. Ecology 85, 591–602.
Shabala, S. (2011). Physiological and cellular aspects of phytotoxicity tolerance in plants: the role of membrane transporters and implications for crop breeding for waterlogging tolerance. New Phytol. 190, 289–298. doi: 10.1111/j.1469-8137.2010.03575.x
Shaw, A., Ruth, E., Wayne, S., and Stephen, D. (2013). Waterlogging in Australian agricultural landscapes: a review of plant responses and crop models. Crop Pasture Sci. 64, 549–562.
Six, J., Paustian, K., Elliot, E. T., and Combrink, C. (2000). Soil structure and organic matter: I. distribution of aggregate size-classes and aggregate-associated carbon. Soil Sci. Soc. Am. J. 64, 681–689.
Steffens, B., and Rasmussen, A. (2016). The physiology of adventitious roots. Plant Physiol. 170, 603–617. doi: 10.1104/pp.15.01360
Stirzaker, R. J., Passioura, J. B., and Wilms, Y. (1996). Soil structure and plant growth: impact of bulk density and biopores. Plant Soil 185, 151–162.
Stock, O., and Downes, N. K. (2008). Effects of additions of organic matter on the penetration resistance of glacial till for the entire water tension range. Soil Tillage Res. 99, 191–201.
Trought, M. C. T., and Drew, M. C. (1980). The development of waterlogging damage in young wheat plants in anaerobic solution cultures. J. Exp. Bot. 31, 1573–1585.
Trought, M. C. T., and Drew, M. C. (1982). Effects of waterlogging on young wheat plants (Trificum aestivum L.) and on soil solutes at different soil temperatures. Plant Soil 326, 311–326.
Uga, Y., Sugimoto, K., Ogawa, S., Rane, J., Ishitani, M., Hara, N., et al. (2013). Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 45, 1097–1102. doi: 10.1038/ng.2725
Vaz, C. M. P., Manieri, J. M., de Maria, I. C., and Tuller, M. (2011). Modeling and correction of soil penetration resistance for varying soil water content. Geoderma 166, 92–101.
Velde, M., and Van Der Tubiello, F. N. (2012). Impacts of extreme weather on wheat and maize in France: evaluating regional crop simulations against observed data. Clim. Change 113, 751–765.
Wade, J., Culman, S. W., Logan, J. A. R., Po, H., Demyan, M. S., Grove, J. H., et al. (2020). Improved soil biological health increases corn grain yield in N fertilized systems across the Corn Belt. Nat. Sci. Rep. 10, 3917. doi: 10.1038/s41598-020-60987-3
Wang, H., Inukai, Y., Yamauchi, A., and Wang, H. (2006). Root Development and Nutrient Uptake. Crit. Rev. Plant Sci. 25:2689.
Whalley, W. R., Watts, C. W., Gregory, A. S., Mooney, S. J., Clark, L. J., and Whitmore, A. P. (2008). The effect of soil strength on the yield of wheat. Plant Soil 306, 237–247. doi: 10.13287/j.1001-9332.201604.009
White, R. G., and Kirkegaard, J. A. (2010). The distribution and abundance of wheat roots in a dense, structured subsoil - Implications for water uptake. Plant Cell Environ. 33, 133–148. doi: 10.1111/j.1365-3040.2009.02059.x
Williams, S. M., and Weil, R. R. (2004). Crop cover root channels may alleviate soil compaction effects on soybean crop. Soil Sci. Soc. Am. J. 68, 1403.
Wuest, S. B., Caesar-TonThat, T. C., Wright, S. F., and Williams, J. D. (2005). Organic matter addition, N, and residue burning effects on infiltration, biological, and physical properties of an intensively tilled silt-loam soil. Soil Tillage Res. 84, 154–167.
Xin, X., Zhang, J., Zhu, A., and Zhang, C. (2016). Effects of long-term (23 years) mineral fertilizer and compost application on physical properties of fluvo-aquic soil in the North China Plain. Soil Tillage Res. 156, 166–172.
Yagüe, M. R., Domingo-Olivé, F., Bosch-Serra, À, Poch, R. M., and Boixadera, J. (2016). Dairy cattle manure effects on soil quality: porosity, earthworms, aggregates and soil organic carbon fractions. Land Degrad. Dev. 27, 1753–1762.
Yang, X., Li, P., Zhang, S., Sun, B., and Xinping, C. (2011). Long-term-fertilization effects on soil organic carbon, physical properties, and wheat yield of a loess soil. J. Plant Nutr. Soil Sci. 174, 775–784.
Keywords: aeration, available water capacity, compaction, crop yield, mechanical impedance, soil health and quality, soil organic matter, waterlogging
Citation: King AE, Ali GA, Gillespie AW and Wagner-Riddle C (2020) Soil Organic Matter as Catalyst of Crop Resource Capture. Front. Environ. Sci. 8:50. doi: 10.3389/fenvs.2020.00050
Received: 28 February 2020; Accepted: 14 April 2020;
Published: 08 May 2020.
Edited by:
Pascal Boivin, University of Applied Sciences Western Switzerland, SwitzerlandReviewed by:
Jörg Schnecker, University of Vienna, AustriaCopyright © 2020 King, Ali, Gillespie and Wagner-Riddle. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alison E. King, YWtpbmcxNUB1b2d1ZWxwaC5jYQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.