
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Endocrinol. , 19 April 2023
Sec. Reproduction
Volume 14 - 2023 | https://doi.org/10.3389/fendo.2023.1120988
This article is part of the Research Topic Uterine Factors Associated with Fertility Impairment View all 16 articles
 Nerea M. Molina1,2*†
Nerea M. Molina1,2*† Lucas Jurado-Fasoli3,4†
Lucas Jurado-Fasoli3,4† Alberto Sola-Leyva1,2
Alberto Sola-Leyva1,2 Raquel Sevilla-Lorente5
Raquel Sevilla-Lorente5 Analuce Canha-Gouveia1,6,7Susana Ruiz-Durán2,8Juan Fontes2,8
Analuce Canha-Gouveia1,6,7Susana Ruiz-Durán2,8Juan Fontes2,8 Concepción M. Aguilera2,9,10,11
Concepción M. Aguilera2,9,10,11 Signe Altmäe1,2,12*
Signe Altmäe1,2,12*Introduction: Several metabolite classes have been identified in human endometrium, including lipids, nucleotides, amino acids, organic acids, and sugars. The first studies suggest the importance of metabolites in endometrial functions, as imbalance in uterine metabolites has been associated with low implantation rate and endometriosis. Nevertheless, most of studies have put emphasis on specific metabolite classes, and we lack the knowledge of the whole metabolome composition in human uterus. Further, a healthy dietary pattern has been shown to potentially protect against different endometrial dysfunctions and is a potential modulator of metabolomic composition and, consequently, the intrauterine microenvironment. The Mediterranean Diet (MD), characterized by a high intake of fruits, vegetables, cereals, nuts, legumes, fish, and olive oil, and a low consumption of meat, dairy products, and processed foods, has been associated with a wide range of benefits for health. Indeed, the MD pattern has displayed a beneficial role in endometriosis management and fertility; however, the relationship between the MD and the endometrial metabolome is still unknown. In our study, we set out to analyze receptive-phase endometrial metabolome profiles among women with infertility and their associations with MD.
Methods: The study included women with male factor infertility (n=8), unexplained infertility (n=10), recurrent implantation failure (n=14), and endometriosis (n=13). The endometrial metabolome was analyzed with ultrahigh-performance liquid chromatography-tandem mass spectroscopy (UPLC–MS/MS). The MD adherence of the participants was assessed using the 14-point MEDAS questionnaire of adherence to the MD.
Results: We provide the whole metabolome profile of the endometrium, where 925 different metabolites were identified. Among these metabolites, lipids comprised the largest part, where polyunsaturated fatty acids (PUFAs) prevailed. Women with endometriosis and recurrent implantation failure were found to have lower levels of PUFAs compared to women with male factor and unexplained infertility (i.e., no clear endometrial alterations), identifying a metabolome profile associated with infertility diagnoses where altered endometrial functions are suspected. Moreover, MD adherence seemed to be associated with the endometrial metabolomic profile in a manner dependent on the health status of the uterus.
Conclusion: The study findings provide insight into the molecular background of female infertility and lead to identification of potential molecular biomarkers and possibilities for modulating the endometrial microenvironment and, thereby, endometrial functions involved in embryo implantation and infertility.
The human endometrium is a dynamic tissue that undergoes continuous cycles of shed, repair, regeneration, and remodeling to prepare for the embryo implantation (1). Due to the complexity of the involved mechanisms and the relative lack of information, embryo implantation process in humans remains the “black box” (2). Unfavorable endometrial microenvironment could lead to endometrial dysfunction promoting different disorders including embryo implantation failure and infertility, abnormal uterine bleeding, endometriosis, and endometrial cancer among others (1, 3). In this sense, the novel “omics” techniques based on the massive molecular characterization of DNA, RNA, proteins, and other small molecules synthesized have allowed to define the molecular phenotype of the receptive-phase endometrium at the mid-secretory phase (so-called “window of implantation”) and the genomic, epigenomic, proteomic, and metabolomic methods have helped to gain insight into the molecular markers of the mid-secretory endometrial functions in health and disease (3–8).
Metabolomics has emerged as a powerful tool to identify small molecules with a molecular mass below 1200Da that participate as intermediates and final downstream products of cellular processes (i.e., metabolites), which contribute to a close real-time understanding of the functional phenotype (9). Despite the growing number of metabolomic studies, the use of the whole metabolite profile analysis related to uterine health is still scarce. The first endometrial metabolome studies have analyzed women with endometriosis and have suggested several potential biomarkers of the disease (10–12). The whole metabolome profiling in the uterus of fertile and infertile women undergoing assisted reproductive techniques showed that specific metabolomic signatures contribute to beneficial endometrial microenvironment to achieve pregnancy (13–15). These are the first findings indicating that metabolomics is a powerful tool in unravelling uterine health and understanding receptive-phase endometrial functions to achieve successful embryo implantation.
Factors that may influence uterine microenvironment include nutrition together with other lifestyle factors. It has been previously shown that a healthy dietary pattern might protect against different endometrial dysfunctions such as infertility, endometriosis, polycystic ovary syndrome, dysmenorrhea, or endometrial cancer (16, 17). In fact, food groups, such as fruits and vegetables, could promote fertility; while other dietary elements, including trans-fatty acids, alcohol, and caffeine, have been associated with adverse effects on fertility (18). The Mediterranean Diet (MD), characterized by a high consumption of fruits, vegetables, cereals, nuts, legumes, fish, and olive oil, and a low consumption of meat, dairy products, and processed foods, has been associated with a wide range of benefits for health (19). Indeed, the MD has been shown to have a beneficial role in endometriosis management (20), fertility (21, 22), and polycystic ovary syndrome (23). However, although lifestyle interventions have shown to influence the uterine proteome (24), the relationship between the MD and the endometrial metabolome is still unknown. In the current study, we set out to analyze receptive-phase endometrial metabolome profiles among women with infertility and their associations with the MD.
This cross-sectional study was approved by the Ethics Committee of the Junta de Andalucía (CEIM/CEI 0463-M1-18r). All procedures carried out in the present study were in accordance with the last revised ethical guidelines of the Declaration of Helsinki and the legally enforced Spanish regulation, which regulates the clinical investigation of human beings (RD 223/04). Written informed consent was obtained from all subjects prior to inclusion.
Forty-five women of infertile couples were recruited at the Reproductive Unit (Virgen de las Nieves University Hospital, Granada) between March 2019 and April 2021. All participants attended the Reproductive Unit after at least 1 year of unprotected sex and not being able to conceive. The inclusion criteria involved women entering an infertility treatment. Infertility causes comprised endometriosis, recurrent implantation failure (RIF), unexplained infertility, or male factor infertility. Endometriosis was diagnosed by laparotomy or laparoscopic surgery and histological confirmation, and RIF was defined as repeated implantation failure after the transfer of 3 good-quality embryos. Unexplained infertility was considered when the medical examination yielded no information about reproductive complications in the couple. Lastly, male factor was defined considering the reference values of the World Health Organization manual for semen analysis (25). Patients with age ≥43 years old, with hormone therapy, gynecological tumors, systemic diseases, pelvic inflammatory disease, or other pelvic pathological conditions were excluded from the study.
Participants were invited to complete an extensive lifestyle questionnaire to collect information including demographic characteristics. Body mass index (BMI) was calculated from the measured weight and height data. Ovulation day was estimated using a digital ovulation test (ClearBlue, Swiss Precision Diagnostics GmbH, Geneva, Switzerland) to precisely schedule the endometrial biopsy collection 7-9 days after the luteinizing hormone (LH) peak (day LH+7-9, window of implantation). After insertion of the speculum, the endometrial tissue sample was collected using the endometrial curette device (Gynétics, Medical Products, Hamont-Achel, Belgium). All biopsies were placed in cryovials, were snap-frozen in the gas phase of liquid nitrogen and stored at -80°C until analysis.
The untargeted metabolomics of endometrial samples were analyzed at Metabolon Inc. (Morrisville, NC, USA) on a platform consisting of four independent ultrahigh-performance liquid chromatography-tandem mass spectroscopy (UPLC–MS/MS) instruments as previously described (26). Briefly, samples were prepared using the automated MicroLab STAR® system from the Hamilton Company. Several recovery standards were added prior to the first step in the extraction process for quality-control (QC) purposes. To recover chemically diverse metabolites, proteins and other macromolecules were precipitated with methanol under vigorous shaking for 2 min (Glen Mills GenoGrinder 2000) followed by centrifugation. The resulting supernatant was divided into 4 aliquots, then placed briefly in a TurboVap (Zymark, Hopkinton, MA, USA) to remove the organic solvent and stored overnight under nitrogen before preparation for analysis. The sample extract was then reconstituted in solvents compatible with each of the four methods. Thus, each aliquot was run using a different method: 1) acidic positive-ion conditions chromatographically optimized for more hydrophilic compounds (reverse phase [RP]/UPLC-MS/MS Pos Early); 2) acidic positive-ion conditions chromatographically optimized for more hydrophobic compounds (RP/UPLC-MS/MS Pos Late); 3) basic negative-ion-optimized conditions (RP/UPLC-MS/MS Neg). These methods used separate acid- and base-dedicated C18 columns (Waters UPLC BEH C18, 2.1×100 mm, 1.7 μm). The fourth aliquot was analyzed via negative ionization after elution from a hydrophilic interaction liquid chromatographic (HILIC) column (Waters UPLC BEH Amide, 2.1x150 mm, 1.7 µm) (HILIC/UPLC-MS/MS Polar). The details of the used solvents and chromatography are described before (27). All UPLC-MS/MS methods used a Waters ACQUITY UPLC (Waters, Milford, MA, USA) coupled to a Thermo Scientific Q-Exactive high resolution/accurate mass spectrometer (Thermo Fischer, Waltham, MA, USA) equipped with a heated electrospray ionization (HESI-II) source and Orbitrap mass analyzer. The MS analysis alternated between MS and data-dependent MSn scans using dynamic exclusion. The MS scan range varied slightly between methods but covered 70-1000 m/z and operated at 35,000 mass resolution.
Three types of controls were analyzed along with the experimental samples: 1) a pooled sample generated from a small volume of each experimental sample (i.e., technical replicate); 2) extracted water (i.e., blank); and 3) a cocktail of QC standards spiked into every analyzed sample for instrument performance monitoring. Instrument variability was determined by calculating the median relative standard deviation (RSD) for the internal standards that were added to each sample prior to injection into the mass spectrometers (median RSDs = 3%). Overall process variability was determined by calculating the median RSD for all endogenous metabolites (i.e., non-instrument standards) present in 100% of the pooled matrix samples, which were technical replicates of pooled samples (median RSD = 7%). Experimental samples and controls were randomized across the platform run.
Raw data, peak-identified, and QC was extracted and processed using Metabolon’s hardware and software as described previously (28). Endometrial metabolites were identified by comparison of the ion features in the experimental samples to reference libraries of authenticated standards with known chromatographic retention index (RI), mass to charge ratios (m/z), and MS/MS spectral fragmentation signatures. Further, biochemical identification was based on 3 criteria: RI within a narrow RI window (150 RI units [≈10s]), accurate mass match ( ± 10ppm), and quality of MS/MS fragmentation match. While there may be similarities between these molecules based on one of these factors, the use of all three data points allows to distinguish and differentiate biochemicals. To ensure high quality of the dataset, QC and curation processes were subsequently used to confirm accurate and consistent chemical assignment and remove system artefacts and background noise. Peaks were quantified through area-under-the-curve analysis and these peak area values allowed the determination of relative quantification among samples (29).
Some compounds were detected by multiple methods (i.e., RP/UPLC-MS/MS Pos Early, RP/UPLC-MS/MS Pos Late, RP/UPLC-MS/MS Neg, or HILIC/UPLC-MS/MS Polar). The UPLC–MS/MS method used to evaluate each compound was pre-selected based on sensitivity and reproducibility criteria (“Platform” column of Supplementary Table S1).
Adherence to the MD was assessed using the 14-point MEDAS questionnaire of adherence to the MD as previously reported and validated (30). The MEDAS questionnaire includes 12 questions related to frequency intake of key foods, and 2 questions related to specific dietary habits of the MD (Supplementary Table S2). Each question scores 0 or 1 point. The global score ranges from 0 to 14, being 0 points null adherence and 14 points complete adherence to the MD. Then, participants were categorized in high adherence (≥9 points) or low adherence (<9 points) to the MD, following previous categorization (30). The consumption of folic acid supplement was also registered among the participants with an additional question together with the MEDAS questionnaire (Supplementary Table S2).
The normal distribution assumption was tested using the Shapiro-Wilk test, visual histograms, and Q-Q plots. Non-normally distributed variables (i.e., endometrial metabolomic signature) were log10-transformed before further analysis. The baseline characteristics and outcomes of the study participants were expressed as mean ± standard deviation (unless otherwise stated). Differences in demographic characteristics between the subjects of our study were analyzed using independent sample t-test or chi-square tests where appropriate.
Multivariate statistical analyses (i.e., partial least squares discriminant analysis [PLS-DA]) were performed on the entire metabolomics dataset using the MetaboAnalyst 5.0 (https://www.metaboanalyst.ca) to investigate potential difference in the metabolomic signature between women with low and high adherence to MD, different infertility diagnoses, and between women supplemented with folic acid and women with no supplementation. To increase the importance of low abundance ions without significant amplification of noise, raw data were normalized by Pareto scaling (31).
To find further differences in metabolite composition between the different groups, one-way analysis of covariance (ANCOVA) controlling for age and BMI was applied. Besides, we conducted Pearson partial correlation analyses to examine the relationship between the adherence to the MD and the endometrial metabolomic signature adjusting for age and BMI. The correlation analyses were performed pooling all participants together and splitting the cohort based on the results of the uterine examination (see results section).
Normality and descriptive analyses were performed in Statistical Package for the Social Sciences (SPSS 28.0, IBM Corp, Armonk, NY, USA). All statistical and correlation analyses were performed using the R software (V.3.6.0; R Foundation for Statistical Computing). Whisker’s plot and volcano plots were built using GraphPad Prism software v.9 (GraphPad Software, San Diego, CA, USA). All p-values were corrected by controlling for the multiple testing, False Discovery Rate (FDR) (32). Statistical significance was set p-value<0.05 after FDR correction.
The study cohort included a total of 45 women with infertility. The characteristics of the participants are presented in Table 1. Participants were primarily Caucasian, with just one Hispanic woman.

Table 1 Patient’s demographics based on the adherence to Mediterranean Diet (MD).
Based on the MEDAS score, the participants were categorized as high adherence (n=25) or low adherence (n=20) to the MD. We also studied the differences in metabolome profiles depending on the infertility diagnosis to relate the uterine microenvironment with endometrial dysregulation. To increase the statistical power of our analysis and based on the thorough clinical examination, 27 patients with endometriosis and RIF were grouped as endometrial-factor infertile (EFI) and 18 women with male factor infertility and unexplained infertility were grouped together as no endometrial-factor infertility (NEFI) was detected. This grouping was further supported by the metabolome signatures detected (Figure 1), where women with RIF or endometriosis demonstrated similar metabolite profiles while women with male factor infertility and unexplained infertility grouped together. Statistical tests revealed that the different groups were age- and BMI-matched (Table 1).
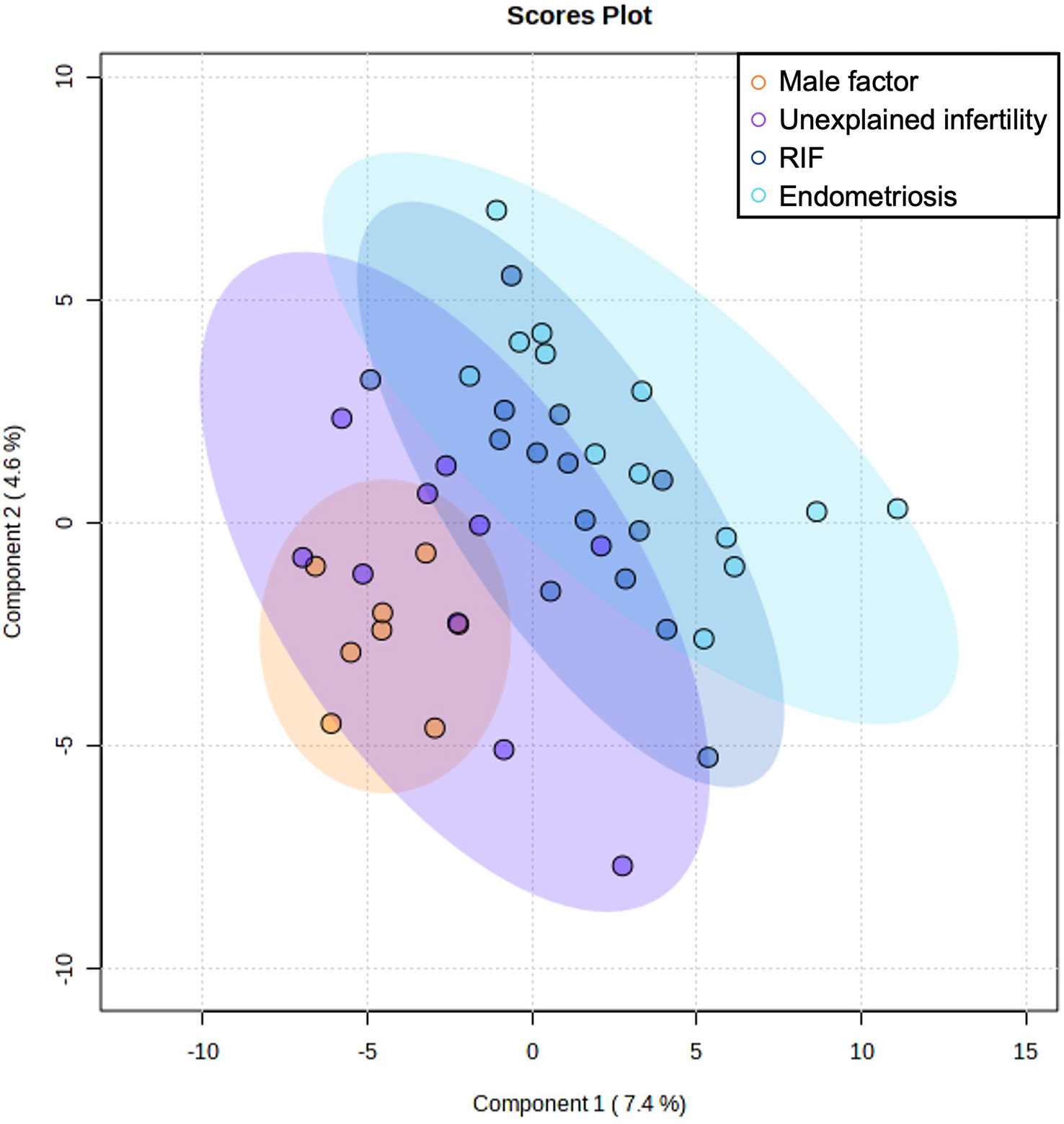
Figure 1 Multivariate Partial Least Squares Discriminant Analysis scores plot from women with male factor infertility, unexplained infertility, endometriosis, and recurrent implantation failure (RIF).
Nine hundred twenty-five metabolites from diverse chemical classes were identified from endometrial tissue samples using an untargeted metabolomic approach. These 925 metabolites included amino acids, lipids, nucleotides, carbohydrates, and xenobiotics among others. The list of identified compounds is provided in Supplementary Table S1. The most abundant metabolites in the entire cohort were N1, N12-diacetylspermine, adenosine 5’-monophosphate (AMP), 4-ethylphenylsulfate, guanosine 5’-monophosphate (5’-GMP), 2-hydroxy-3-methylvalerate, and UDP- glucuronate.
When we compared metabolome based on adherence to diet, the PLS-DA scores plot did not show any different metabolomic signature between the study groups with high and low adherence to the MD (Figure 2A) and no significant differences were detected after correction between the metabolites and the different groups (Supplementary Table S3).
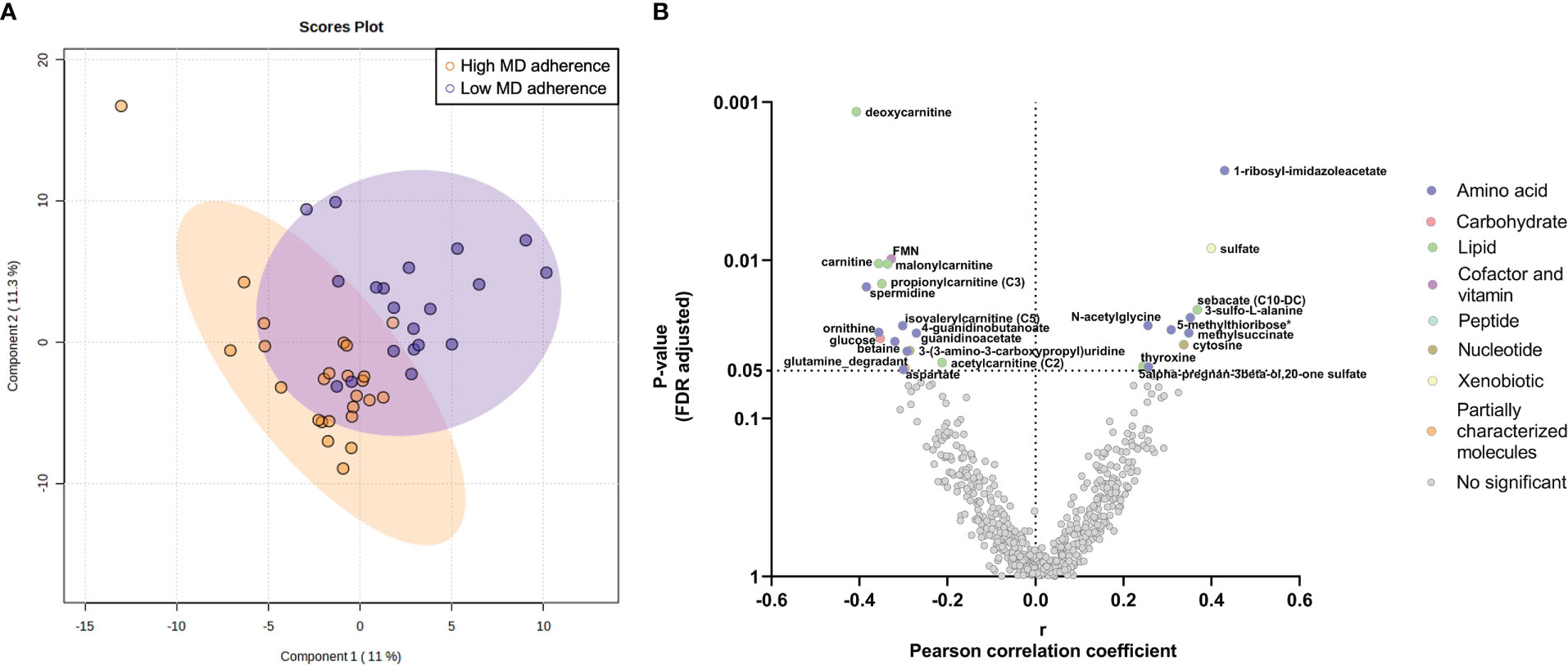
Figure 2 Endometrial metabolomic signature analysis in women with high and low adherence to Mediterranean Diet (MD). (A) Multivariate Partial Least Squares Discriminant Analysis scores plot from women with high and low MD adherence. (B) Partial correlation analyses between the MD global score and the endometrial metabolome. The X-axis represents Pearson partial correlations coefficients, whereas the Y-axis represents the FDR-adjusted p-values of the correlations. Grey dots represent non-significant correlations, whereas colored dots represent statistically significant correlations (p-value<0.05 after FDR correction).
When focusing on metabolite classes, the adherence to the MD showed statistically significant correlations with amino acids and lipids levels (all p-values<0.05 after FDR correction; Figure 2B). Specifically, the adherence to the MD was positively correlated with amino acids related to the histidine metabolism (1-methyl-5-imidazoleacetate and 1-ribosyl-imidazoleacetate), to the methionine, cysteine, SAM, and taurine metabolism (N-acetylmethionine sulfoxide, 5-methylthioribose, and 3-sulfo-L-alanine), and the thyroxine amino acid (Figure 2B, Supplementary Table S4). In contrast, we observed that the adherence to the MD was negatively correlated with different amino acid metabolites (argininate, ornithine, betaine, spermidine, and aspartate) and with carnitines (e.g., deoxycarnitine, carnitine, malonylcarnitine, and 2-methylhexanoylcarnitine) (Figure 2B, Supplementary Table S4).
Comparing groups based on the uterine examination, the PLS-DA scores plot did not discriminate the metabolomic signature between patients with EFI and second group with NEFI (Figure 3A). However, dihomolinolenate (20:3 n-3 or n-6), linolenate (18:3 n-3 or n-6), and linoleate (18:2 n-6) metabolites related with the polyunsaturated fatty acids (PUFAs) pathway were significantly increased in NEFI women compared to EFI women (all p-values<0.001 after FDR correction; Figure 3B, Supplementary Table S5).
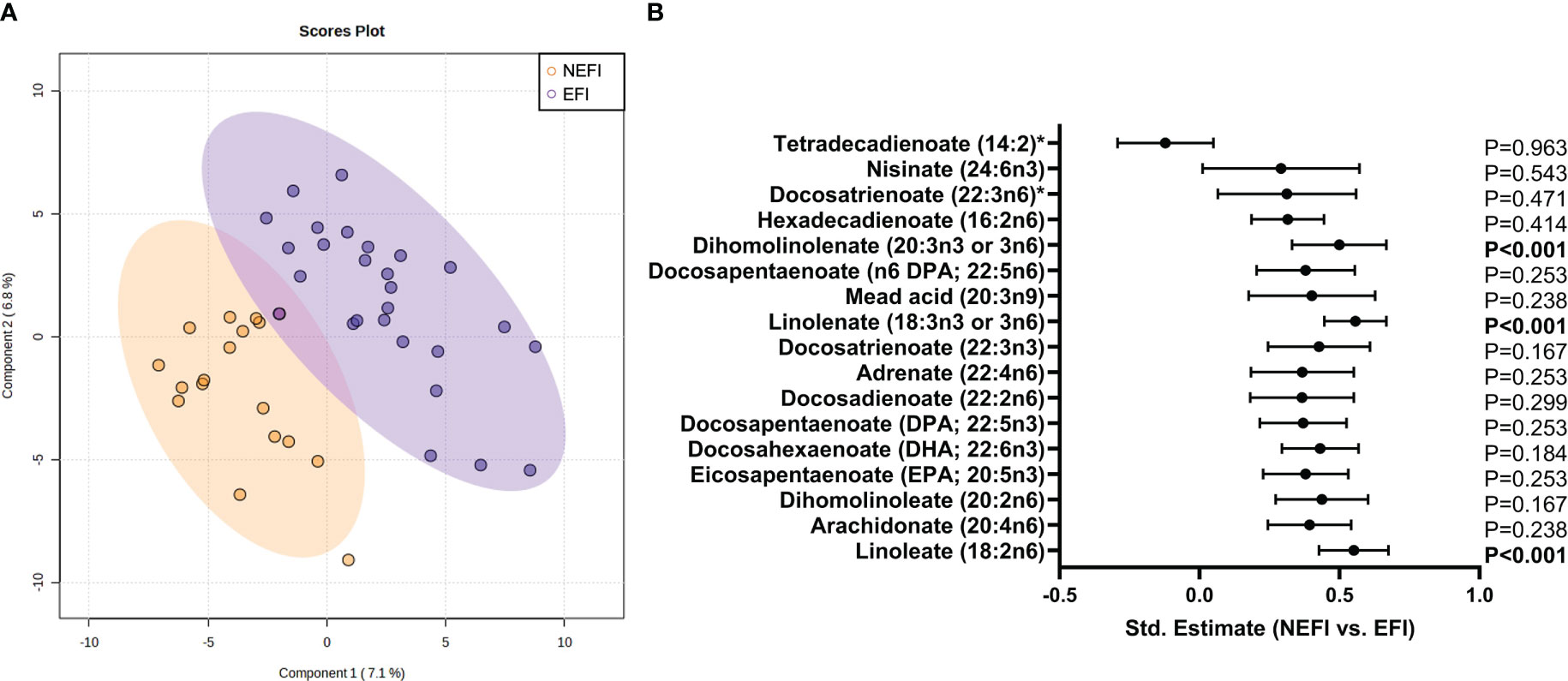
Figure 3 Endometrial metabolomic signature analysis in women with no detectable endometrial dysfunction in comparison to patient group with endometriosis and recurrent implantation failure. (A) Multivariate Partial Least Squares Discriminant Analysis scores plot from women with no detectable endometrial dysfunction and women with endometrial-factor infertility. (B) Statistical differences in long chain polyunsaturated fatty acid metabolism between women with no detectable endometrial dysfunction and women with endometrial-factor infertility. Statistical significance was set p-value<0.05 after FDR correction. NEFI, no endometrial-factor infertility; EFI, endometrial-factor infertility.
Next, we performed the correlation analyses in parallel among patient group of women with EFI and among group of women with NEFI and we observed a higher number of significant correlations between the adherence to the MD and metabolites in women with EFI than in women with no detectable endometrial dysfunction (Figure 4). In the NEFI group, up to 31 metabolites correlated to MD. On one hand, the adherence to the MD was positively correlated to different fatty acids (2S,3R-dihydroxybutyrate, 2-hydroxystearate, and 2-hydroxypalmitate), to progestin steroids (pregnanediol-3-glucuronide, and 5alpha-pregnan-3beta,20beta-diol monosulfate) while was negatively correlated to metabolites involved in the xanthine metabolism (1,3,7-trimethylurate, 5-acetylamino-6-amino-3-methyluracil, caffeine, 1,7-dimethylurate, and 1-methylurate) (Figure 4A, Supplementary Table S4). Contrary, in patients with EFI, a total of 73 metabolites correlated to MD. We observed a positive correlation between the adherence to the MD and metabolites involved in the xanthine metabolism (i.e., 5-acetylamino-6-amino-3-methyluracil, 1,3,7-trimethylurate, 1-methylxanthine, and paraxanthine) and in the metabolism of ceramides (i.e., dihydroceramides, hexosylceramides, and lactosylceramides) (Figure 4B, Supplementary Table S4). We also found in women with EFI, a negative correlation between the adherence to the MD and carnitine metabolites (e.g., carnitine, deoxycarnitine, or acyl-carnitines), primary bile acid metabolites (i.e., cholate, glycochenodeoxycholate 3-sulfate, glycochenodeoxycholate, and chenodeoxycholate), and different amino acid metabolites (e.g., aspartate, argininate, ornithine, orotidie, spermidine, betaine, or homocysteine) (Figure 4B, Supplementary Table S4).
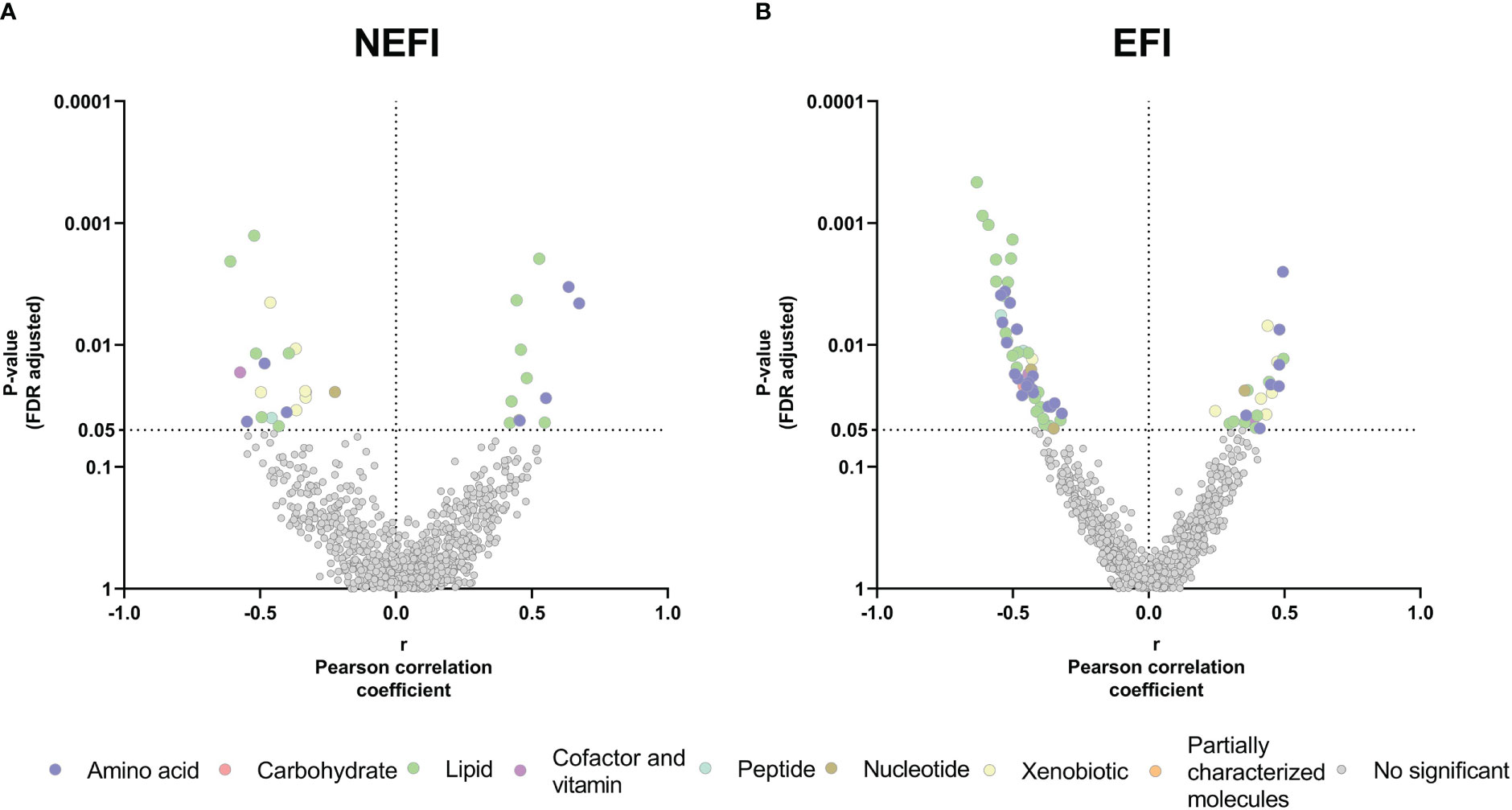
Figure 4 Relationship between the Mediterranean Diet adherence and the endometrial metabolome in (A) women with no detectable endometrial dysfunction and (B) patient group with endometriosis and recurrent implantation failure. The X-axis represents Pearson partial correlations coefficients, whereas the Y-axis represents the FDR-adjusted p-values of the correlations. Grey dots represent non-significant correlations, whereas colored dots represent statistically significant correlations (p-value<0.05 after FDR correction). NEFI, no endometrial-factor infertility; EFI, endometrial-factor infertility.
Additionally, we combined the 2 classification features (i.e., MD adherence and endometrial-factor infertility) for comparisons and 4 groups were established: high MD adherence and NEFI (n=11), high MD adherence and EFI (n=14), low MD adherence and NEFI (n=7), and low MD adherence and EFI (n=13) (Table 1). When we compared metabolome profiles among these groups, linolenate metabolite was significantly more present among women with high MD adherence and NEFI compared to women with low MD adherence and EFI (p-value=0.014 after FDR correction, Supplementary Figure S1).
Regarding the folic acid supplementation, the PLS-DA scores plot did not show any different clusters (Figure 5) nor were significant differences observed in the metabolomic profile when comparing the groups of women who were supplemented with folic acid or not (Supplementary Table S6).
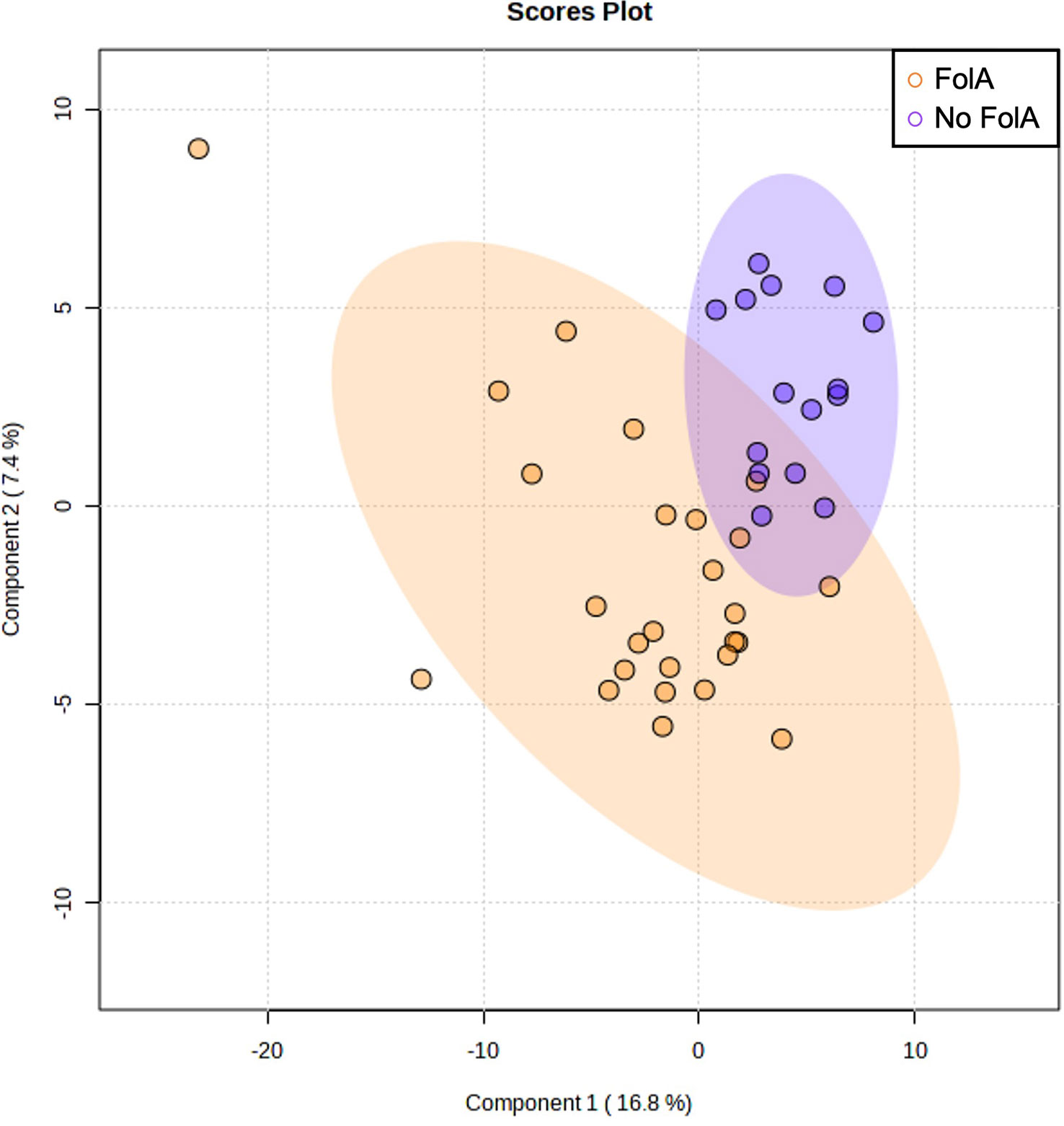
Figure 5 Multivariate Partial Least Squares Discriminant Analysis scores plot from women supplemented with folic acid (FolA) and women with no supplementation.
In this cross-sectional study, we used an untargeted metabolomics approach to profile the endometrial metabolome and to assess possible differences in the metabolite composition between women with low and high adherence to MD, different infertility diagnoses (i.e., male factor infertility, unexplained infertility, RIF, and endometriosis), and taking also into account folic acid supplementation. We provide the whole metabolome profile of the endometrium, where 925 different metabolites were identified. Among these metabolites, lipids comprised the largest percentage (40%), where PUFAs such as mead acid, dihomo-linolenate, and docosatrienoate prevailed. Our main results indorse those women with endometrium-related infertility (i.e., endometriosis or RIF) have lower levels of PUFAs in the endometrium compared to women with no clear endometrial factor infertility (i.e., male factor and unexplained infertility). Further, supporting our grouping based on infertility diagnosis, male factor and unexplained infertility demonstrated closer metabolome patterns while RIF and endometriosis were more similar. Moreover, adherence to MD seemed to be associated with the endometrial metabolomic profile, differing between women with distinct infertility diagnoses. In this way, adherence to MD could be related to the levels of different fatty acids, progestin steroids, xanthine metabolites, carnitines, amino acids, and bile acids in a manner dependent on the health status of the uterus.
In our study, PUFAs prevailed in all the women studied. When comparing the study groups based on the uterine examination, dihomolinolenate, linoleate, and linolenate were detected at lower levels in the endometrium of women with RIF or endometriosis. PUFAs are fatty acids with multiple double bonds in their structures which cannot be synthesized by the human body, being considered essential fatty acids (33). Linoleate and linolenate are the conjugated acids of linoleic acid (LA) and α-linolenic acid (ALA), respectively, which are the primary PUFAs in Western diets (34). PUFAs are shown to play a substantial role in the regulation of body homeostasis and are considered crucial for reproductive health (35). These lipids participate in female fertility at different reproductive phases, including oocyte maturation and quality, and embryo implantation (36–39). Our study results indicate that microenvironment in the uteri of women with RIF or endometriosis could be lacking in PUFAs, which could negatively impact the endometrial functions.
We detected low levels of linolenate in women with EFI compared to women with NEFI. Interestingly, when we combined MD adherence and endometrial-factor infertility features, the results did support that linolenate was less present in the endometrium of women with low MD adherence and EFI compared to women with high MD adherence and NEFI. Previous studies show contradicting results, as negative correlations between ALA levels in different biological fluids (i.e., serum, follicular fluid) of women undergoing infertility treatment and pregnancy rates (40) and number of metaphase II oocytes (41) have been found. Similarly, serum ALA levels have shown positive association with the presence of endometriosis (40). These discrepancies could be due to the different analyzed specimens (serum versus endometrial biopsy). Some evidence also manifests that LA has negative effects on oocyte maturation (33), which could be mediated by the role of lipids in the physical properties and biological functions of membranes (13). Other authors measured the LA : ALA ratio (n-6 to n-3) and they found that women with higher LA : ALA ratios had greater embryo implantation and pregnancy rates compared to women with reduced LA : ALA ratios (42, 43), suggesting a potential role for increased LA to enhance endometrial inflammation and receptivity (43) and for elevated ALA to disturb embryo implantation (35). It is worth mentioning that LA is a precursor to docosapentaenoic acid (n-6). Thus, our data seem to support the previous results as we found that the docosapentaenoate (i.e., conjugated acid of docosapentaenoic acid) is less present in the endometrium of women with EFI. Nevertheless, another study revealed that docosapentaenoic acid was higher in women with no implantation compared to women with successful implantation (44). In this way, further investigation is required to unravel the role of n-6 and n-3 PUFAs in endometrial receptivity as this ratio imbalance could lead to aberrant endometrial functions.
Likewise, we observed a positive correlation between adherence to MD and metabolites involved in the metabolism of ceramides in women with EFI. In line, we noticed that ceramide metabolites were higher in women with no endometrial dysfunction. Although with no statistical significance, these results agree with a previous study where ceramides were detected at lower amount in the endometrial fluid of women with endometriosis compared to women without endometriosis (10).
Another family of metabolites that we found dysregulated in women with endometrium-related infertility were progestin steroids (i.e., intermediates of progesterone metabolism) which are shown to contribute to fertility and the maintenance of pregnancy (45). We revealed that lower adherence to MD was related to lower endometrial levels of these hormonal intermediates in women with endometrium-related infertility. A recent work demonstrated that premature rise in the serum progesterone level in the late follicular phase led to a disrupted endometrial lipid profile in the endometrium during the peri-implantation period (44), suggesting that the altered lipid pattern could desynchronize endometrial receptivity and early embryo implantation. There seems to be a link between progesterone metabolism and lipid profiles, which needs further investigation.
Additional interesting result of our study is that a higher adherence to MD correlated with lower levels of carnitine metabolites within the whole cohort. Accordingly, the patient group of women with EFI exhibited a negative correlation between MD adherence and acylcarnitines. It could be that MD confers protection against endometriosis since increased carnitine levels were observed in women with endometriosis (10) and correlated with the cytokine and cellular profile of endometriosis (46). Moreover, high levels of acylcarnitines have also been associated with beta-oxidation dysfunction, participating in inflammation processes (47). Further, we detected a negative correlation of MD adherence with bile acid metabolites in women with EFI, which corroborates that high levels of bile acids are a negative factor for health (48). All these results related to lipid metabolism endorse that lipidome is an essential aspect of the complex process of endometrium receptivity and embryo implantation (35) and alterations in lipid metabolism could lead to endometrial dysfunctions.
We also found a negative correlation between the MD adherence and xanthine metabolites (i.e., products of the purine and xenobiotics degradation) in women with NEFI and a positive correlation in women with EFI. A healthy diet in women without endometrial abnormal functioning has been associated with lower levels of these metabolites, which were found to be elevated in previous studies in women with endometrial disorders (e.g., caffeine (49), xanthosine (11)). However, studies involving xanthines in female reproduction have been mainly focused on their use in oocyte in vitro maturation (49) and other reproductive outcomes need to be considered.
Some aspects of the current study should be considered as limitations. Firstly, the cross-sectional design does not allow the establishment of causality. Secondly, our results should be interpreted as preliminary due to the limited sample size, which might have underpowered the detection of statistical differences in metabolite composition between the studied groups. Further studies investigating a MD intervention on the endometrial metabolomic signature are thus required to confirm our results.
Despite these shortcomings, the strength of our study is the focus on a very narrow time-frame of the cycle, the mid-secretory phase, and all the samples were collected at the day LH+7, minimizing thereby the heterogeneity and it enabled us to study the receptive-phase uterine microenvironment. Further, existing metabolomic studies of female reproductive health have mainly been performed in serum samples and assessed a preestablished groups of metabolites, while in our study we analyzed for the first time the endometrial whole metabolic signature with an untargeted method.
This study presents the endometrial whole metabolome profile, demonstrating that PUFAs are prevailing in the human uterus. In conclusion, we showed that women with RIF or endometriosis displayed lower levels of PUFAs in the endometrium compared to women with no clear endometrial alterations, identifying a metabolomic profile associated with infertility diagnoses where altered endometrial functions are suspected (i.e., endometriosis and RIF). In addition, adherence to the MD was associated with the endometrial metabolic profile in a manner dependent on the health status of the uterus. Our study findings help to understand the molecular background of female infertility, which could allow to identify potential molecular biomarkers and take personalized medicine into the reproductive clinical practices. With a healthy diet, endometrial microenvironment could be changed from unfavorable to favorable, which could have an influence on endometrial functions.
The original contributions presented in the study are included in the article/Supplementary Materials. Further inquiries can be directed to the corresponding authors.
The studies involving human participants were reviewed and approved by Ethics Committee of the Junta de Andalucía (CEIM/CEI 0463-M1-18r). The patients/participants provided their written informed consent to participate in this study.
NM, LJ-F, AS-L, RS-L, and SA conceived and designed the study. NM, AS-L, JF, and SR-D recruited the participants and obtained the samples. LJ-F and RS-L processed the diet data. NM and LJ-F analyzed the data and drafted the manuscript. AS-L, RS-L, AC-G, JF, SR-D, CA, and SA revised the manuscript. All authors contributed to the article and approved the submitted version.
This work is supported by Grants Endo-Map PID2021-12728OB-100, ENDORE SAF2017-87526-R, and PRE2018-085440 funded by MCIN/AEI/10.13039/501100011033 and ERFD A way of making Europe; Grants RYC-2016-21199, FPU19/01638, FPU19/01609, and FPU19/03745 funded by MCIN/AEI/10.13039/501100011033 and by ESF Investing in your future; FEDER/Junta de Andalucía-Consejería de Economía y Conocimiento: ROBIN A-CTS-614-UGR20 and IRENE P20_00158; Grant PPJIB2021-02 funded by the University of Granada Plan Propio de Investigación; Plan de Recuperación, Transformación y resiliencia, Ayudas para la recualificación del sistema universitario español, Ayudas Margarita Salas.
The authors would like to thank all the participants of this study for their time and the biological material. We also acknowledge the research support by Copan Italia S.p.A Inc., and Clearblue, SPD Swiss Precision Diagnostics GmbH. This study was performed as part of a Ph.D. Thesis conducted within the Official Doctoral Program in Biomedicine of the University of Granada, Spain.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2023.1120988/full#supplementary-material
Supplementary Table 1 | Dataset comprising the average peak intensity and detection platform of each identified metabolite in women with high and low adherence to the Mediterranean Diet.
Supplementary Table 2 | Quantitative score (14-item) of adherence to the Mediterranean Diet. The participants were asked to choose the best option for each question. ‘<’ means less than. The option marked with grey color is the one used as criteria for 1 point. The participants also self-reported their folic acid intake.
Supplementary Table 3 | Statistical differences in metabolites between groups based on Mediterranean Diet (MD) adherence (i.e., high and low MD adherence).
Supplementary Table 4 | Correlation analyses between the Mediterranean Diet global score and the endometrial metabolome.
Supplementary Table 5 | Statistical differences in metabolites between groups based on uterine examination (i.e., women with no endometrial-factor infertility and patients with endometrial-factor infertility).
Supplementary Table 6 | Statistical differences in metabolites between groups based on folic acid supplementation.
Supplementary Figure 1 | Statistical difference in linolenate metabolite between women with high Mediterranean Diet (MD) adherence and no endometrial-factor infertility and women with low MD adherence and endometrial-factor infertility. Statistical significance was set p-value<0.05 after FDR correction. EFI, endometrial-factor infertility; NEFI, no endometrial-factor infertility.
1. Evans J, Salamonsen LA, Winship A, Menkhorst E, Nie G, Gargett CE, et al. Fertile ground: human endometrial programming and lessons in health and disease. Nat Rev Endocrinol (2016) 12:654–67. doi: 10.1038/nrendo.2016.116
2. Ban Z, Knöspel F, Schneider MR. Shedding light into the black box: advances in in vitro systems for studying implantation. Dev Biol (2020) 463:1–10. doi: 10.1016/j.ydbio.2020.04.003
3. Garcia-Alonso L, Handfield L-F, Roberts K, Nikolakopoulou K, Fernando RC, Gardner L, et al. Mapping the temporal and spatial dynamics of the human endometrium in vivo and in vitro. Nat Genet (2021) 53:1698–711. doi: 10.1038/s41588-021-00972-2
4. Tokarz J, Adamski J, Rižner TL. Metabolomics for diagnosis and prognosis of uterine diseases? a systematic review. J Pers Med (2020) 10:294. doi: 10.3390/jpm10040294
5. Suhorutshenko M, Kukushkina V, Velthut-Meikas A, Altmäe S, Peters M, Mägi R, et al. Endometrial receptivity revisited: endometrial transcriptome adjusted for tissue cellular heterogeneity. Hum Reprod (2018) 33:2074–86. doi: 10.1093/humrep/dey301
6. Sola-Leyva A, Andrés-León E, Molina NM, Terron-Camero LC, Plaza-Díaz J, Sáez-Lara MJ, et al. Mapping the entire functionally active endometrial microbiota. Hum Reprod (2021) 36:1021–31. doi: 10.1093/humrep/deaa372
7. Altmäe S, Koel M, Võsa U, Adler P, Suhorutšenko M, Laisk-Podar T, et al. Meta-signature of human endometrial receptivity: a meta-analysis and validation study of transcriptomic biomarkers. Sci Rep (2017) 7:10077. doi: 10.1038/s41598-017-10098-3
8. Altmäe S, Esteban FJ, Stavreus-Evers A, Simón C, Giudice L, Lessey BA, et al. Guidelines for the design, analysis and interpretation of “omics” data: focus on human endometrium. Hum Reprod Update (2014) 20:12–28. doi: 10.1093/humupd/dmt048
9. Wishart DS. Emerging applications of metabolomics in drug discovery and precision medicine. Nat Rev Drug Discovery (2016) 15:473–84. doi: 10.1038/nrd.2016.32
10. Domínguez F, Ferrando M, Díaz-Gimeno P, Quintana F, Fernández G, Castells I, et al. Lipidomic profiling of endometrial fluid in women with ovarian endometriosis†. Biol Reprod (2017) 96:772–9. doi: 10.1093/biolre/iox014
11. Li J, Guan L, Zhang H, Gao Y, Sun J, Gong X, et al. Endometrium metabolomic profiling reveals potential biomarkers for diagnosis of endometriosis at minimal-mild stages. Reprod Biol Endocrinol (2018) 16:42. doi: 10.1186/s12958-018-0360-z
12. Li J, Gao Y, Guan L, Zhang H, Sun J, Gong X, et al. Discovery of phosphatidic acid, phosphatidylcholine, and phosphatidylserine as biomarkers for early diagnosis of endometriosis. Front Physiol (2018) 9:14. doi: 10.3389/fphys.2018.00014
13. Matorras R, Martinez-Arranz I, Arretxe E, Iruarrizaga-Lejarreta M, Corral B, Ibañez-Perez J, et al. The lipidome of endometrial fluid differs between implantative and non-implantative IVF cycles. J Assist Reprod Genet (2020) 37:385–94. doi: 10.1007/s10815-019-01670-z
14. Hong L, Zhu YC, Liu S, Wu T, Li Y, Ye L, et al. Multi-omics reveals a relationship between endometrial amino acid metabolism and autophagy in women with recurrent miscarriage. Biol Reprod (2021) 105:393–402. doi: 10.1093/biolre/ioab101
15. Yurci A, Dokuzeylul Gungor N, Gurbuz T. Spectroscopy analysis of endometrial metabolites is a powerful predictor of success of embryo transfer in women with implantation failure: a preliminary study. Gynecol Endocrinol (2021) 37:415–21. doi: 10.1080/09513590.2021.1883584
16. Ciebiera M, Esfandyari S, Siblini H, Prince L, Elkafas H, Wojtyła C, et al. Nutrition in gynecological diseases: current perspectives. Nutrients (2021) 13:1178. doi: 10.3390/nu13041178
17. Gaskins AJ, Nassan FL, Chiu Y-H, Arvizu M, Williams PL, Keller MG, et al. Dietary patterns and outcomes of assisted reproduction. Am J Obstet Gynecol (2019) 220:567.e1–567.e18. doi: 10.1016/j.ajog.2019.02.004
18. Hsiao PY, Fung JL, Mitchell DC, Hartman TJ, Goldman MB. Dietary quality, as measured by the alternative healthy eating index for pregnancy (AHEI-p), in couples planning their first pregnancy. Public Health Nutr (2019) 22:3385–94. doi: 10.1017/S1368980019001290
19. Guasch-Ferré M, Willett WC. The Mediterranean diet and health: a comprehensive overview. J Intern Med (2021) 290:549–66. doi: 10.1111/joim.13333
20. Ott J, Nouri K, Hrebacka D, Gutschelhofer S, Huber JC, Wenzl R. Endometriosis and nutrition-recommending a Mediterranean diet decreases endometriosis-associated pain: an experimental observational study. J Aging Res Clin Pract (2012) 1:162–6.
21. Skoracka K, Ratajczak AE, Rychter AM, Dobrowolska A, Krela-Kaźmierczak I. Female fertility and the nutritional approach: the most essential aspects. Adv Nutr (2021) 12:2372–86. doi: 10.1093/advances/nmab068
22. Muffone ARMC, de Oliveira Lübke PDP, Rabito EI. Mediterranean Diet and infertility: a systematic review with meta-analysis of cohort studies. Nutr Rev (2022). doi: 10.1093/nutrit/nuac087
23. Mei S, Ding J, Wang K, Ni Z, Yu J. Mediterranean Diet combined with a low-carbohydrate dietary pattern in the treatment of overweight polycystic ovary syndrome patients. Front Nutr (2022) 9:876620. doi: 10.3389/fnut.2022.876620
24. Abdulkhalikova D, Sustarsic A, Vrtačnik Bokal E, Jancar N, Jensterle M, Burnik Papler T. The lifestyle modifications and endometrial proteome changes of women with polycystic ovary syndrome and obesity. Front Endocrinol (Lausanne) (2022) 13:888460. doi: 10.3389/fendo.2022.888460
25. World Health Organization. WHO laboratory manual for the examination and processing of human semen, 6th ed. Geneva: World Health Organization (2021).
26. Evans AM, Bridgewater B, Qiang L, Mitchell M, Robinson R, Dai H, et al. High resolution mass spectrometry improves data quality and quality as compared to unit mass resolution mass spectrometry in high throughput profiling metabolomics. Metabolomics (2014) 4:132. doi: 10.4172/2153-0769.1000132
27. Visconti A, Le Roy CI, Rosa F, Rossi N, Martin TC, Mohney RP, et al. Interplay between the human gut microbiome and host metabolism. Nat Commun (2019) 10:4505. doi: 10.1038/s41467-019-12476-z
28. DeHaven CD, Evans AM, Dai H, Lawton KA. Organization of GC/MS and LC/MS metabolomics data into chemical libraries. J Cheminform (2010) 2:9. doi: 10.1186/1758-2946-2-9
29. Evans AM, DeHaven CD, Barrett T, Mitchell M, Milgram E. Integrated, nontargeted ultrahigh performance liquid chromatography/electrospray ionization tandem mass spectrometry platform for the identification and relative quantification of the small-molecule complement of biological systems. Anal Chem (2009) 81:6656–67. doi: 10.1021/ac901536h
30. Schröder H, Fitó M, Estruch R, Martínez-González MA, Corella D, Salas-Salvadó J, et al. A short screener is valid for assessing Mediterranean diet adherence among older Spanish men and women. J Nutr (2011) 141:1140–5. doi: 10.3945/jn.110.135566
31. Eriksson L, Kettaneh-Wold N, Wold S. Scaling: introduction to multi- and megavariate data analysis using projection methods (PCA & PLS). Umetrics (1999), 213–25.
32. Benjamini Y, Krieger AM, Yekutieli D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika (2006) 93:491–507. doi: 10.1093/biomet/93.3.491
33. Wang R, Feng Y, Chen J, Chen Y, Ma F. Association between polyunsaturated fatty acid intake and infertility among American women aged 20–44 years. Front Public Health (2022) 10:938343. doi: 10.3389/fpubh.2022.938343
34. Simopoulos AP. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed Pharmacother (2002) 56:365–79. doi: 10.1016/S0753-3322(02)00253-6
35. Yang T, Zhao J, Liu F, Li Y. Lipid metabolism and endometrial receptivity. Hum Reprod Update (2022) 28:858–89. doi: 10.1093/humupd/dmac026
36. Sturmey R, Reis A, Leese H, McEvoy T. Role of fatty acids in energy provision during oocyte maturation and early embryo development. Reprod Domest Anim (2009) 44:50–8. doi: 10.1111/j.1439-0531.2009.01402.x
37. Norwitz ER, Schust DJ, Fisher SJ. Implantation and the survival of early pregnancy. New Engl J Med (2001) 345:1400–8. doi: 10.1056/NEJMra000763
38. Nehra D, Le HD, Fallon EM, Carlson SJ, Woods D, White YA, et al. Prolonging the female reproductive lifespan and improving egg quality with dietary omega-3 fatty acids. Aging Cell (2012) 11:1046–54. doi: 10.1111/acel.12006
39. Hammiche F, Vujkovic M, Wijburg W, de Vries JHM, Macklon NS, Laven JSE, et al. Increased preconception omega-3 polyunsaturated fatty acid intake improves embryo morphology. Fertil Steril (2011) 95:1820–3. doi: 10.1016/j.fertnstert.2010.11.021
40. Jungheim ES, Macones GA, Odem RR, Patterson BW, Moley KH. Elevated serum alpha-linolenic acid levels are associated with decreased chance of pregnancy after in vitro fertilization. Fertil Steril (2011) 96:880–3. doi: 10.1016/j.fertnstert.2011.07.1115
41. Mirabi P, Chaichi MJ, Esmaeilzadeh S, Ali Jorsaraei SG, Bijani A, Ehsani M, et al. The role of fatty acids on ICSI outcomes: a prospective cohort study. Lipids Health Dis (2017) 16:18. doi: 10.1186/s12944-016-0396-z
42. Jungheim ES, Frolova AI, Jiang H, Riley JK. Relationship between serum polyunsaturated fatty acids and pregnancy in women undergoing In Vitro fertilization. J Clin Endocrinol Metab (2013) 98:E1364–8. doi: 10.1210/jc.2012-4115
43. Moran L, Tsagareli V, Noakes M, Norman R. Altered preconception fatty acid intake is associated with improved pregnancy rates in overweight and obese women undertaking in vitro fertilisation. Nutrients (2016) 8:10. doi: 10.3390/nu8010010
44. Li J, Gao Y, Guan L, Zhang H, Chen P, Gong X, et al. Lipid profiling of peri-implantation endometrium in patients with premature progesterone rise in the late follicular phase. J Clin Endocrinol Metab (2019) 104:5555–65. doi: 10.1210/jc.2019-00793
45. di Renzo GC, Mattei A, Gojnic M, Gerli S. Progesterone and pregnancy. Curr Opin Obstet Gynecol (2005) 17:598–600. doi: 10.1097/01.gco.0000191899.84567.4d
46. Dionyssopoulou E, Vassiliadis S, Evangeliou A, Koumantakis EE, Athanassakis I. Constitutive or induced elevated levels of l-carnitine correlate with the cytokine and cellular profile of endometriosis. J Reprod Immunol (2005) 65:159–70. doi: 10.1016/j.jri.2004.12.002
47. Vouk K, Hevir N, Ribič-Pucelj M, Haarpaintner G, Scherb H, Osredkar J, et al. Discovery of phosphatidylcholines and sphingomyelins as biomarkers for ovarian endometriosis. Hum Reprod (2012) 27:2955–65. doi: 10.1093/humrep/des152
48. Osuna-Prieto FJ, Rubio-Lopez J, Di X, Yang W, Kohler I, Rensen PCN, et al. Plasma levels of bile acids are related to cardiometabolic risk factors in young adults. J Clin Endocrinol Metab (2022) 107:715–23. doi: 10.1210/clinem/dgab773
Keywords: endometriosis, endometrium, folic acid, lifestyle, metabolomics, Mediterranean diet, recurrent implantation failure, unexplained infertility
Citation: Molina NM, Jurado-Fasoli L, Sola-Leyva A, Sevilla-Lorente R, Canha-Gouveia A, Ruiz-Durán S, Fontes J, Aguilera CM and Altmäe S (2023) Endometrial whole metabolome profile at the receptive phase: influence of Mediterranean Diet and infertility. Front. Endocrinol. 14:1120988. doi: 10.3389/fendo.2023.1120988
Received: 10 December 2022; Accepted: 05 April 2023;
Published: 19 April 2023.
Edited by:
Bassem Refaat, Umm Al-Qura University, Saudi ArabiaReviewed by:
Bei Shi, China Medical University, ChinaCopyright © 2023 Molina, Jurado-Fasoli, Sola-Leyva, Sevilla-Lorente, Canha-Gouveia, Ruiz-Durán, Fontes, Aguilera and Altmäe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nerea M. Molina, bW9saW5hbmVyZWFAdWdyLmVz; Signe Altmäe, c2lnbmVhbHRtYWVAdWdyLmVz
†These authors have contributed equally and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.