
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Endocrinol. , 20 July 2022
Sec. Developmental Endocrinology
Volume 13 - 2022 | https://doi.org/10.3389/fendo.2022.945736
This article is part of the Research Topic Maternal-Fetal Interface: New Insight in Placenta Research View all 18 articles
 María Luján Scalise1
María Luján Scalise1 Nicolás Garimano1
Nicolás Garimano1 Marcelo Sanz2
Marcelo Sanz2 Nora Lia Padola2
Nora Lia Padola2 Patricia Leonino3Adriana Pereyra3
Patricia Leonino3Adriana Pereyra3 Roberto Casale3María Marta Amaral1
Roberto Casale3María Marta Amaral1 Flavia Sacerdoti1†*
Flavia Sacerdoti1†* Cristina Ibarra1†*
Cristina Ibarra1†*The presence of Escherichia coli in the vaginal microbiome has been associated with pregnancy complications. In previous works, we demonstrated that Shiga toxin-producing Escherichia coli (STEC) can produce abortion and premature delivery in rats and that Shiga toxin type 2 (Stx2) can impair human trophoblast cell lines. The hypothesis of this work was that STEC may colonize the lower female reproductive tract and be responsible for adverse pregnancy outcomes. Thus, the aim of this work was to evaluate the presence and prevalence of virulence factor genes from STEC in the endocervix of asymptomatic pregnant women. For that purpose, endocervical swabs were collected from pregnant women during their prenatal examination. Swab samples were enriched in a differential medium to select Enterobacteria. Then, positive samples were analyzed by PCR to detect genes characteristic of Escherichia sp. (such as uidA and yaiO), genes specific for portions of the rfb (O-antigen-encoding) regions of STEC O157 (rfbO157), and STEC virulence factor genes (such as stx1, stx2, eae, lpfAO113, hcpA, iha, sab, subAB). The cytotoxic effects of stx2-positive supernatants from E. coli recovered from the endocervix were evaluated in Vero cells. Our results showed that 11.7% of the endocervical samples were positive for E. coli. Additionally, we found samples positive for stx2 and other virulence factors for STEC. The bacterial supernatant from an isolate identified as E. coli O113:NT, carrying the stx2 gene, exhibited cytotoxic activity in Vero, Swan 71 and Hela cells. Our results open a new perspective regarding the presence of STEC during pregnancy.
Shiga toxin-producing Escherichia coli (STEC), a bacterium that belongs to the family Enterobacteriaceae, can cause severe foodborne diseases. The main reservoir for STEC is the intestinal tract of cattle, and the bacterium can survive for months in soil, water, or organic material (1). The main route of transmission is through contaminated food, mainly minced meat (undercooked meat below 71°C), unpasteurized food, contaminated vegetables and person-to-person contact by the fecal-oral route (2, 3).
STEC comprises strains producing two Shiga toxins: Stx type 1 (Stx1) and Stx type 2 (Stx2) (4) and its variants, Stx1 (a – c) and Stx2 (a – k) (5–7). Not all Stx subtypes have been associated with severe illness (8). In this sense, it has been described that Stx2a and Stx2c are clinically more related with severe cases of hemolytic uremic syndrome (HUS), and that STEC O157 strains carrying stx2a predominate in human infections, causing more severe disease symptoms than those carrying stx2c (9, 10).
Stx is the main virulence factor of STEC and, in combination with other virulence factors, contributes to the pathogenic potential of STEC strains. In this sense, intimin (eae gene) plays an essential role in the intimate attachment and colonization of intestinal cells of STEC O157:H7, although many STEC isolates lacking the eae gene are also able to efficiently colonize the human gut (11). Many other proteins, such as the long polar fimbriae (Lpf) (12), the hemorrhagic coli pilus (Hcp) (13), the IrgA homolog adhesin (IhA) (14), and the STEC autotransporter contributing to biofilm formation (Sab) (15), are also involved in the adherence of O157 and non-O157 STEC strains. Non-O157 STEC strains also produce Subtilase cytotoxin (SubAB), able to cause cytotoxic effects in epithelial cells (16).
It is currently known that E. coli strains can colonize the vagina, usually asymptomatically, although epidemiological studies have shown that the presence of these bacteria in the female reproductive tract may be a risk factor for pregnancy (17). Some studies developed in Argentina and other countries have shown a high prevalence of E. coli in vaginal microbiota of adult women (17, 18). However, to our knowledge, there are no reports about the effects of STEC infection on human pregnancy, although some reports have indicated that STEC transmission from the mother to the child during delivery can cause neonatal HUS (19) or STEC-mediated HUS during pregnancy (20). In Argentina, the most common STEC serotype is O157:H7 (21, 22). Nevertheless, the appearance of non-O157 STEC strains is a clear evidence of the dynamic genome of these E. coli pathogens and their ability to transfer or acquire important virulence factors (23, 24). Since there are no epidemiological studies regarding STEC infections during pregnancy, it is difficult to know their impact on reproductive health.
Previous works in our laboratory have demonstrated that Stx2 injection in pregnant rats in early and late pregnancy causes premature delivery, miscarriage and impairments in placental development (25, 26). Also, we have shown that the immunization of rats against Stx2 can prevent the detrimental effects of the toxin during pregnancy (27). Taking these previous findings into account, we hypothesized that STEC can colonize the endocervix and may be responsible for complications in pregnancy. The main goal of this work was to evaluate the presence and prevalence of virulence factor genes from STEC in the endocervix from asymptomatic pregnant women and to better understand the possible clinical relevance of this pathogen during human pregnancy.
Asymptomatic pregnant women (from 17 to 37 years old) with gestational age from 12 to 34 weeks were enrolled (n=103) during their prenatal examination in the Obstetrics Service of the Maternal and Child Department of the Prof. A. Posadas National Hospital (Buenos Aires, Argentina) between January and March 2019. Endocervical swab samples were collected under direct visualization during a speculum examination. Swabs were transported in sterile tubes containing Cary Blair transport medium (Britania, Argentina) and stored at 4-8°C until used.
Endocervical swabs were enriched in 3 mL of Tryptic Soy Broth (Oxoid, UK) overnight (ON) at 37°C and shaken at 150 rpm. Then, cultures were streaked into Sorbitol MacConkey (SMAC) agar (Oxoid, UK) and incubated ON at 37°C. SMAC agar was used for a preliminary selection of Enterobacteria based on a differential sorbitol and lactose fermentation profile. SMAC agar is recommended as a selective and differential medium for the detection of STEC O157:H7. Enterobacteria capable of fermenting sorbitol form pink colonies on SMAC agar, while non-sorbitol fermenting bacteria, such as STEC O157:H7, form white/colorless colonies.
Total genomic DNA from colonies grown in SMAC agar was obtained to detect the presence of E. coli by PCR. DNA was purified using PURO bacteria Kit (PB-L products, Bio-Logicos, Argentina) according to the manufacturer’s instructions and quantified by Nanodrop One (Thermo Fisher Scientific, USA). Specific genes encoding for the enzyme beta-glucuronidase (uidA gene) (28) and external membrane protein of E. coli (yaiO gene) (29), and genes specific for portions of the rfb (O-antigen-encoding) regions of STEC O157 (rfbO157gene) (11) were detected by PCR using specific primers (Table 1).

Table 1 Primers used.
PCRs were performed in a total volume of 20 µL: 6 µL H2O, 3 µL DNA, 0.5 µL of each forward and reverse primer (10 µM) and 10 µL Master Mix 2X Kit (M024, Inbio Highway, Argentina). The thermocycler (Thermo Fisher Scientific, USA) program used consisted of one cycle at 95°C for 10 min followed by 35 cycles of 94°C for 40 s, 58°C for 30 s, and 72°C for 60 s, ending with one cycle of 72°C for 2 min. A total of 10 μL of each PCR reaction product was loaded into an agarose gel to confirm the presence and size of the corresponding amplicon.
Samples positive for yaiO were analyzed for rfbO157 as well as for the presence of other STEC virulence factor genes, including: stx1, stx2, stx2a, stx2c (4), subAB (30), eae (31), lpfAO113 (12), hcp (13), iha (32) and sab (15) by PCR (Table 2).

Table 2 Evaluation of STEC virulence factors, primer sequences and size of amplicons .
The Vero cell line (Vero), derived from African green monkey kidney, was purchased from the American Type Culture Collection (ATCC-CCL-81, Manassas, VA, USA). The HeLa cell line (ATCC-CCL-2), derived from a human cervix adenocarcinoma, was used as a human endocervical model. The Swan 71 cell line, derived by telomerase-mediated transformation of a 7-week cytotrophoblast isolate, was kindly provided by Dr. Gil Mor, Yale University, New Haven, CT, USA (33).
All cell lines were cultured in Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 (DMEM-F12 medium, Sigma Aldrich, USA) supplemented with 10% Fetal Bovine Serum (FBS, Internegocios S.A., Argentina), 100 U/mL penicillin/streptomycin, and 2 mM L-glutamine (GIBCO, USA), and grown at 37°C in a humidified 5% CO2 incubator. For growth-arrested conditions, the medium was used without FBS.
Endocervical samples that were positive for stx2 by PCR were grown ON in Luria-Bertani Broth (LB) (Sigma Aldrich, USA) at 37°C with shaking at 150 rpm. Then, the cultures were diluted (1:20) in DMEM-F12 medium (Sigma Aldrich, USA), supplemented with 1 mM of HEPES buffer solution (4-(2-Hydroxyethyl)piperazine-1-ethanesulfonic acid, N-(2-Hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid)) (GIBCO, USA) and grown until exponential phase (OD600 = 0.4-0.5) at 37°C with shaking at 150 rpm. Subsequently, bacterial supernatants were collected after centrifugation at 10,000 x g for 5 min and filtered (0.22-μm pore-size filter; Millipore, USA). In some experiments, to induce Stx2 production, mitomycin-C (1 µg/mL) was added at the exponential phase and the bacterial culture was grown for additional 3 h (34).
Cell viability assays were performed as previously described (35). Briefly, 15x 103 cells/well (Vero, Swan 71, and HeLa) were seeded in 96-well plates and grown at 80% of confluence. Then, cells were exposed for 72 h to different concentrations (1x10-6 – 0.1 µg/mL) of purified Stx2 (Phoenix Laboratory, Tufts Medical Center, Boston, USA) or to serial dilutions of bacterial supernatant (10-1-10-6) in growth-arrested conditions. After treatment, cells were incubated with a neutral red solution (50 µg/mL) at 37°C for 1 h in 5% CO2. Then, cells were washed and fixed with 1% CaCl2-1% formaldehyde and finally lysed with 1% acetic acid in 50% ethanol solution to solubilize the neutral red uptake by cells. Absorbance in each well was read at 540 nm in an automated plate spectrophotometer (RT-6000, Rayto Life and Analytical Sciences Co. Ltd., China). Results are expressed as percentage of cell viability, where 100% represents control cells without toxin or supernatant treatment. The 50% cytotoxic dose (CD50) of Stx2 and the bacterial supernatant corresponded to the dilution required to kill 50% of cells.
For the neutralization assay, bacterial supernatant in a dilution required to kill 50% of Vero cells (CD50) was co-incubated with 25 µg/mL of the mouse monoclonal antibody 2E11 against the A-subunit of Stx2 (anti-Stx2 mAb) at 37°C for 1 h (36). The mixture was added to a 96-well culture plate containing Vero cells grown at 80% of confluence and then incubated for 72 h. The cytotoxicity was analyzed by neutral red uptake as previously described. The CD50 of purified Stx2 (1 ng/mL) was used as control.
The enriched bacterial culture from endocervical samples carrying the stx2 gene and showing cytotoxic effects on Vero was streaked onto LB agar plates and incubated ON at 37°C. Grown colonies were picked and added in 96-well plates and grown in LB medium ON at 37°C with agitation at 150 rpm. The next day, wells of each column were pooled and the DNA was purified to test the presence of the stx2 gene. Afterward, individual colonies grown in each well of stx2-positive columns were directly picked and confirmed to be STEC by testing the presence of the stx2 gene by PCR and by Vero cell assays. Isolates were then sub-cultured for further genetic or phenotypic typing.
The serotypes (O and H) of STEC isolates were determined by microagglutination, as described by Blanco etal. (37).
This study had the approval of the Human Research Ethics Committee of the Prof. A. Posadas National Hospital, Buenos Aires, Argentina (Ref # 423 EMnPeSe/20), in accordance with the Argentine Good Clinical Practice Guidelines. All pregnant women were thoroughly informed about the purpose of the study and provided a written informed consent. The exclusion criteria were: women with genital papilloma virus infection (HPV), human immunodeficiency virus (HIV), pelvic inflammatory disease (PID) and sexually transmitted diseases such as chlamydia, gonorrhea, and genital herpes.
Data were plotted and statistically analyzed using Graph Pad Prism 5.0 (San Diego, CA, USA). Cytotoxicity curves were fitted using a four-parameter logarithmic regression. Statistical significance for all experiments was assessed using analysis of variance (ANOVA) with Tukey’s multiple comparison test as a posteriori test. In all cases, statistical significance was set at *p < 0.05.
Sixteen out of the 103 endocervical swab samples developed colonies on SMAC agar. Only non-sorbitol fermenting colonies were observed. Fifteen out of those sixteen samples had the uidA gene and 12 out of the 16 had the yaiO gene (Table 3). Considering that the yaiO gene is more specific than the uidA gene for the identification of E. coli, only those containing the yaiO gene were considered positive for E. coli (29). One sample (S12) was negative for both genes (Table 3). Therefore, 12 out of the 103 endocervical samples recruited for this study (11.7%) were considered positive for E. coli spp. in the endocervical microbiota. The absence of sorbitol-fermenting colonies (characteristic of O157:H7 E. coli) on SMAC agar was confirmed by PCR (none of the samples evaluated were positive for the rfbO157 gene) (Table 3).
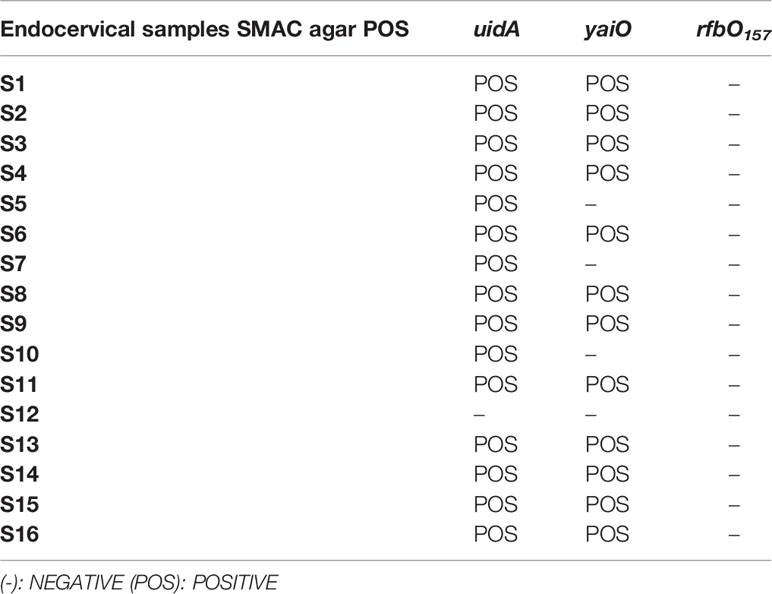
Table 3 Genotypic characterization of E. coli in endocervical samples grown on SMAC agar.
The presence of STEC virulence factors was analyzed in the 12 endocervical samples that developed colonies on SMAC agar and were confirmed positive for E. coli by the presence of the yaiO gene. Seven out of the 12 samples (58.3%) amplified for the stx2 gene, 5 out of the 12 (41.7%) for the lpfAO113 gene, 8 out of the 12 (66.7%) for the hcpA gene and 3 out of the 12 (25%) for the iha gene (Table 4). None of them were positive for the eae, stx1 or sab genes.
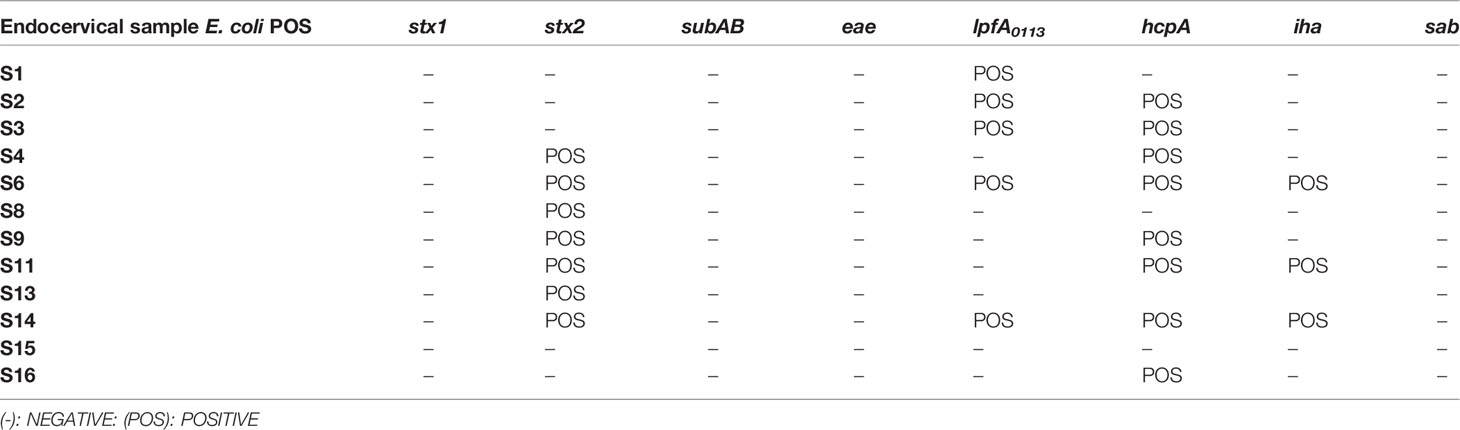
Table 4 Genotypic identification of STEC virulence factors in E. coli-positive samples .
The cytotoxic activity of filter-sterilized bacterial supernatants obtained from endocervical samples positive for E. coli carrying the stx2 gene was evaluated in Vero cells (Figure 1).
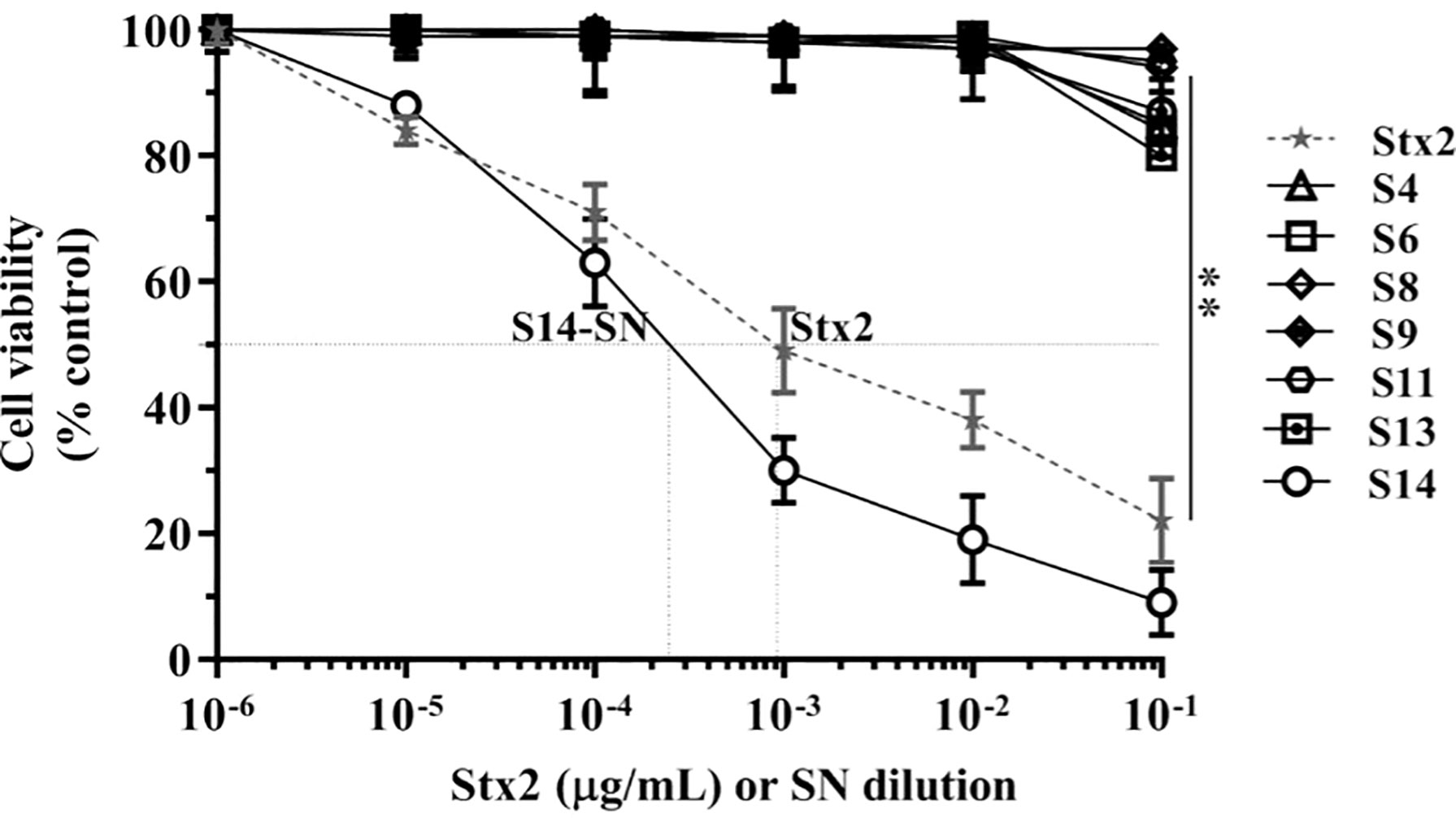
Figure 1 Cytotoxicity of E. coli-positive endocervical samples carrying the stx2 gene on the viability of Vero cells. Vero cells were exposed to serial dilutions of bacterial supernatants (SN) from endocervical samples carrying the stx2 gene or different concentrations of purified Stx2 under growth-arrested conditions. Cell viability was determined by neutral red uptake after 72 h of incubation, and 100% represents cells incubated under identical conditions but without treatment. Data are shown as means ± S.D from at least three independent experiments performed in triplicate. **p < 0.01.
Incubation of Vero cells for 72 h with serial dilutions of the bacterial supernatant corresponding to sample 14 (S14-SN) caused a significant cytotoxicity in a dose-dependent manner, reaching a CD50 at 2x10-4 dilution. The CD50 of Stx2 in S14-SN was equivalent to that elicited by approximately 5 µg/mL of purified Stx2 (Figure 1). In contrast, the other filter-sterilized bacterial supernatants showed no cytotoxic effects on the viability of Vero cells, even if previously grown with mitomycin-C, an inductor of stx2 phages (data not shown).
Neutralization studies were used to confirm that the significant decrease in Vero cell viability caused by S14-SN was due to the cytotoxic action of Stx2. The results showed that S14-SN cytotoxicity was prevented by preincubation of the bacterial S14-SN (1:5000) with the anti-Stx2 mAb (25 µg/mL) at 37°C for 1 h. Neutralization of purified Stx2 was used as control (Figure 2).
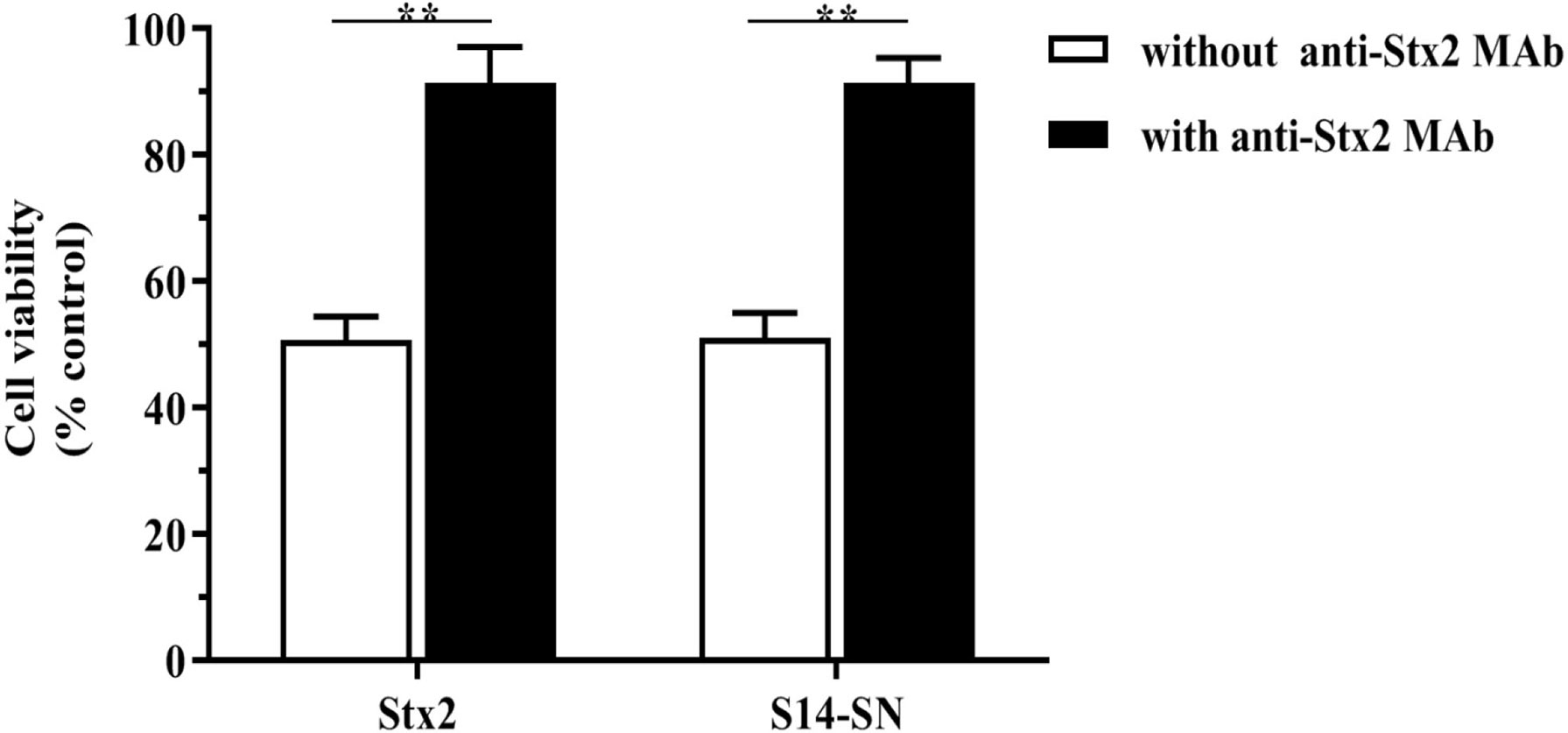
Figure 2 Neutralization of the Stx2 cytotoxicity of bacterial supernatants isolated from endocervical samples on the viability of Vero cells. The bacterial supernatant from the endocervical sample 14 (S14-SN, dilution 1:5000) was preincubated with the mouse monoclonal antibody 2E11 against the A-subunit of Stx2 (anti-Stx2 mAb, 25 µg/mL) at 37°C for 1 h. The mixture was then added to a 96-well culture plate containing Vero cells and incubated for 72 h. The CD50 of purified Stx2 (1 ng/mL) incubated with anti-Stx2 mAb was used as control. Cell viability was analyzed by neutral red uptake. Data are shown as mean ± S.D from at least three independent experiments performed in triplicate. **p < 0.01, n=3.
A significant cytotoxic effect was observed when monolayers of human endocervical (HeLa) cells were exposed to different concentrations of Stx2 or serial dilutions of the filter-sterilized bacterial supernatants or bacteria isolated from endocervical sample S14 (S14-SN) for 72 h. The CD50 was obtained with 10 ng/mL of purified Stx2 and with a 5x10-3 dilution of S14-SN. In agreement with the results described above, cytotoxicity was also observed with purified Stx2 on monolayers of extravillous trophoblasts (Swan 71 cells). The CD50 after 72 h of incubation with purified Stx2 was obtained at 100 ng/mL and with a dilution of approximately 2x10-2 of S14-SN (Figure 3). These results indicate that filter-sterilized bacterial supernatants from the STEC-positive endocervical microbiota can impair endocervical and trophoblast cell viabilities mediated by Stx2.
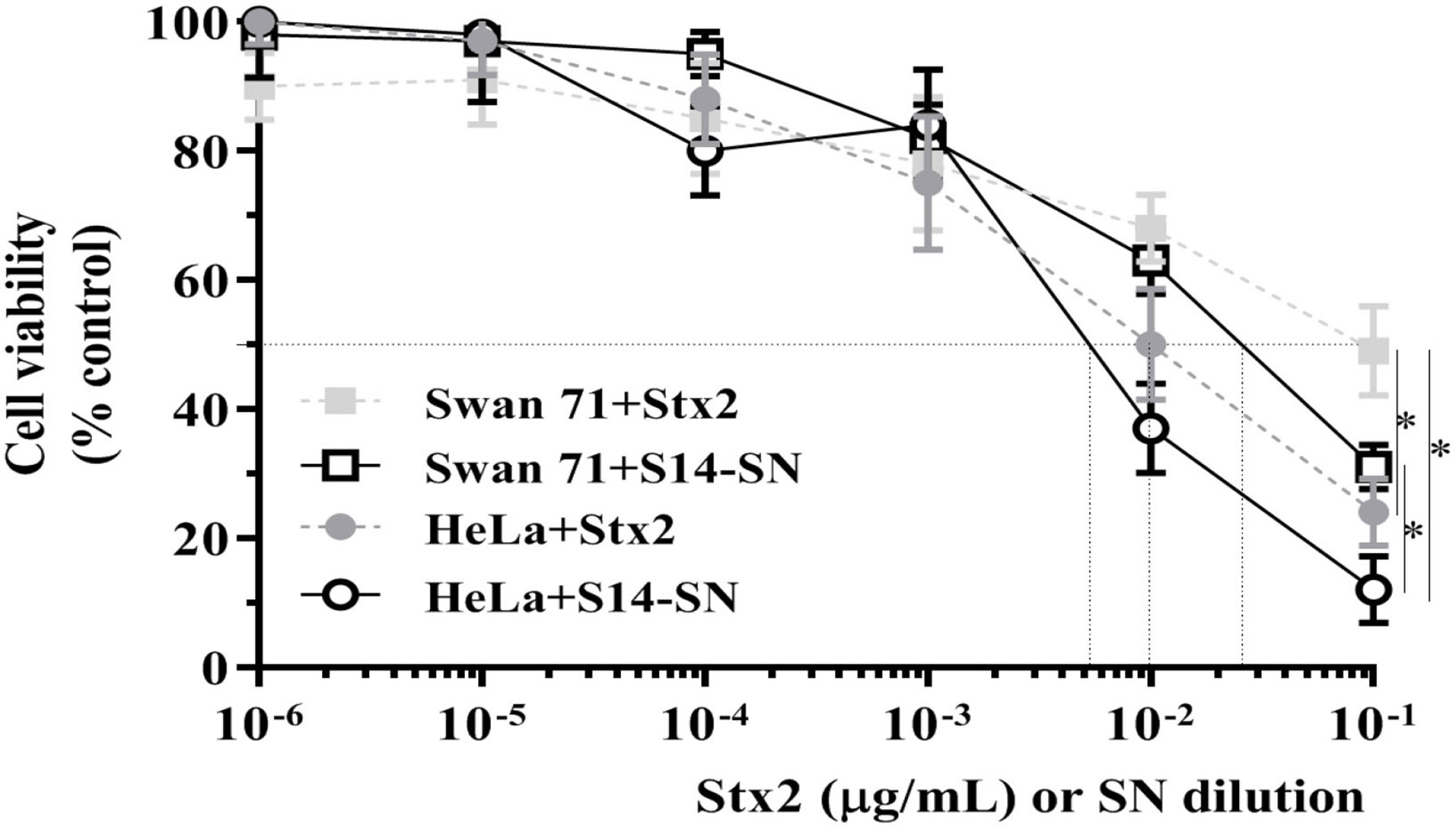
Figure 3 Cytotoxic effects of STEC-positive endocervical samples on the viability of Swan 71 and HeLa cell lines. Cells were exposed to purified Stx2 or serial dilutions of the supernatant from endocervical sample S14 (S14-SN) for 72 h. Cell viability was analyzed by neutral red uptake. Data are shown as mean ± S.D from at least three independent experiments performed in triplicate. *p < 0.05, n=3.
The bacterial isolate from endocervical sample S14, named STEC 123/21, was subtyped for stx2 by PCR, using specific primers for the stx2a and stx2c genes (Table 2). Figure 4 shows a representative gel of the STEC isolate subjected to PCR assay. Results showed PCR products of the expected sizes, consistent with the presence of the stx2a gene (Figure 4) and the absence of the subAB gene (S14 in Table 3).
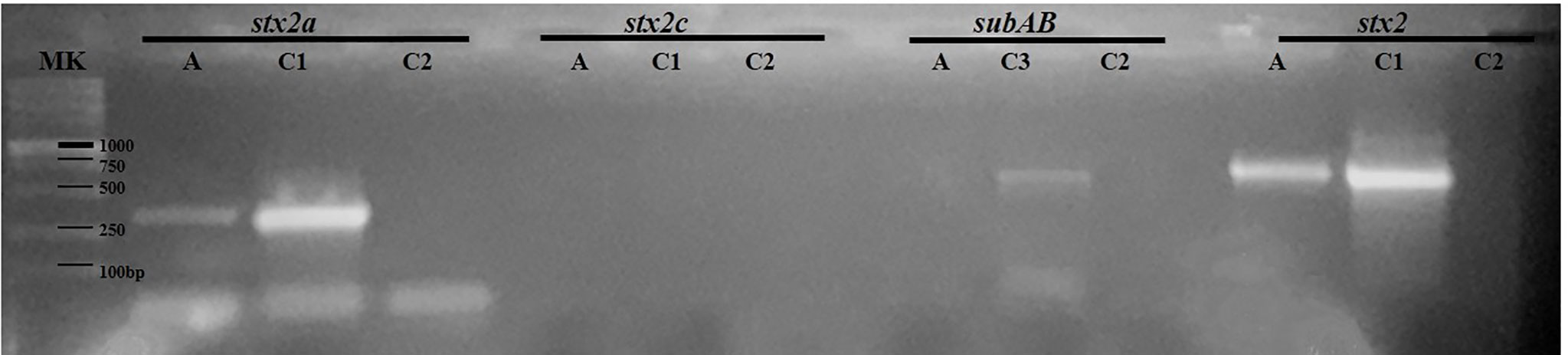
Figure 4 Characterization of a STEC strain isolated from an endocervical sample by PCR assays. The STEC strain named 123/21 was subtyped for the stx2 gene. The amplicons were seeded on 1.5% agarose gel for 30 min at 80 Volts. A: STEC strain 123/21. C1: E. coli O157:H7 strain 125/99 (stx2a+ 349 bp; stx2 627 bp), C2: negative control of PCR; C3: E. coli O113:H21 (subAB; 556 bp); DNA size marker (MK).
STEC strain 123/21 belonged to the O113 type, but the H type could not be determined (nondetermined, NT). The analysis of the supernatant from STEC O113:NT demonstrated that it was able to induce a marked decrease in cell viability. The cytotoxicity of the supernatant of this STEC isolate was similar to that obtained from the initial endocervical sample S14 containing other bacteria (SN-S14) (Figure 5).

Figure 5 Cytotoxic effects of the STEC isolate O113:NT obtained from an endocervical sample on the viability of Vero cells. Cells were exposed to purified Stx2 or serial dilutions of supernatants from STEC O113:NT or sample S14 for 72 h. Cell viability was analyzed by neutral red uptake. Data are shown as mean ± S.D from at least three independent experiments performed in triplicate. *p < 0.05.
The vaginal microbiota is a complex ecosystem consisting of around 200 species of microorganisms, being Lactobacillus crispatus, L. jensenii and L. gasseri the most represented species. Several studies have shown that around 70% of the vaginal microbiota in pregnant and non-pregnant women is composed of Lactobacillus (38–40). The substitution of Lactobacillus by pathogenic or opportunistic microorganisms is associated with the development of bacterial vaginosis and lower urinary tract infections, and the presence of non-Lactobacillus species has been associated with poor reproductive health and/or complications in pregnancy (41–44). In this regard, the presence of E. coli in the female reproductive tract has been found to be a risk factor for the progression of pregnancy (17, 45, 47).
The results of this work demonstrate the prevalence of E. coli in the endocervical microbiota in 11.7% of asymptomatic pregnant women. Previous studies performed in Argentina (18) and other countries of South America, Europe and North America (17) have reported an incidence similar to that found in this work. E. coli are commensal bacteria of the intestine of humans and animals, and some pathogenic strains, such as STEC, can cause moderate to severe gastrointestinal disease in humans. However, the risk of spontaneous abortion or preterm delivery in humans associated with STEC infection or its main virulence factor, Stx2, has not yet been evaluated.
Our results showed that 7 out of the 12 endocervical samples positive for E. coli carried the stx2 gene, although only one of them (1/12, 8.3%) caused significant cytotoxicity in Vero cells due to the presence of Stx2 in the bacterial culture supernatant. The isolated STEC was identified as O113:NT Stx2a-positive. The Stx2 cytotoxicity evaluated by neutral red uptake method in the remaining stx2-positive E. coli samples was undetectable. We hypothesized that the composition of the endocervical gram-negative microbiota may be negatively regulating Stx2 expression and/or STEC growth, as previously described (46, 47). Moreover, the relationship between induction of Stx2-encoding phages and toxin production in STEC (48) may vary considerably in response to the microenvironment (49).
An interesting finding was the detection of lpfA0113 and hcpA in our samples, considering that lpfAO113-positive STEC strains are associated with the appearance of small outbreaks of intestinal and extra-intestinal diseases in humans (50) and that the hcpA gene is associated with the pathogenicity of non-O157 STEC (51).
The fact that the microbiological selection by SMAC was negative for O157:H7, together with the absence of amplification for the eae gene that encodes for Locus of Enterocyte Effacement (LEE), led us to study the expression of the iha gene, which encodes an outer membrane adhesin protein. This adhesin was detected in 25% (3/12) of the endocervical samples also positive for E. coli, which is an interesting finding because previous studies have demonstrated the positivity of the iha gene in non-STEC uropathogenic E. coli (52). LEE-negative STEC strains have been found to be associated with clinical cases of HUS (53, 54). In the present work, we also studied the sab gene, which encodes an adhesin that promotes adherence to human epithelial cells, mediates the formation of biofilm, and is involved in intestinal colonization of LEE-negative STEC strains (15), but found no positive samples.
It is known that non-O157 STEC expressing Stx2 can affect the adult population, as observed in the outbreak that occurred in Germany in 2011, which produced approximately 3,500 cases of infection, 810 cases of HUS, and 39 deaths (55). Detailed studies of this outbreak demonstrated that the stx2-encoding phage can insert into the genome of non-O157 STEC strains and cause serious disease in the adult population, mostly women (56), although details on complications of pregnant vs non-pregnant women were not reported.
In summary, our results show the presence of a STEC O113:NT strain with Stx2a production in the endocervix of an asymptomatic woman during the first trimester of pregnancy. The patient (sample S14) received cephalexin, a broad-spectrum antibiotic, during the second trimester of gestation (20 weeks) to treat uterine inflammation and urinary infection. Probably antibiotic administration is the reason for no longer presence of the STEC strain in subsequent analysis of the endocervical samples during the second and third trimester as well as urine and fecal sample during the third trimester. At birth, the newborn presented a good general condition, and subsequent clinical checks of the mother and baby were normal. Therefore, the contributions of this study encourage us about the importance of the detection of E. coli during prenatal studies to prevent possible complications in pregnancy. Epidemiological studies related to the presence of vaginal STEC may establish its association with possible risks for the reproductive health of women.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by Human Research Ethics Committee of the Prof. A. Posadas National Hospital, Buenos Aires, Argentina (Ref # 423 EMnPeSe/20). The patients/participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
The authors contributions are as follows: Conceived and designed the experiments: SML, GN, CR, MA, SF, IC. Performed the experiments: SML, GN, SM, PN, MA, SF. Analyzed the data: SML, GN, SM, PN, MA, SF, IC. Contributed reagents/materials/analysis tools: SML, LP, PA, CR, MA, SF, IC. Wrote the paper: SML, GN, SF, IC. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We are grateful to Roxane M. F. Piazza from the Laboratorio de Bacteriología, Instituto Butantan, São Paulo, Brazil, for kindly providing mAb 2E11 directed against the A-subunit of Stx2.
1. Sapountzis P, Segura A, Desvaux M, Forano E. An Overview of the Elusive Passenger in the Gastrointestinal Tract of Cattle: The Shiga Toxin Producing. Microorganisms (2020) 8(6):877. doi: 10.3390/microorganisms8060877
2. Rivas M, Chinen I, Miliwebsky E, Masana M. Risk Factors for Shiga Toxin-Producing Escherichia coli-Associated Human Diseases. Microbiol Spectr (2014) 2(5):1–14. doi: 10.1128/microbiolspec.ehec-0002-2013
3. Alconcher LF, Rivas M, Lucarelli LI, Galavotti J, Rizzo M. Shiga Toxin-Producing Escherichia coli in Household Members of Children With Hemolytic Uremic Syndrome. Eur J Clin Microbiol Infect Dis (2020) 39(3):427–32. doi: 10.1007/s10096-019-03738-1
4. Scheutz F, Teel LD, Beutin L, Piérard D, Buvens G, Karch H, et al. Multicenter Evaluation of a Sequence-Based Protocol for Subtyping Shiga Toxins and Standardizing Stx Nomenclature. J Clin Microbiol (2012) 50(9):2951–63. doi: 10.1128/JCM.00860-12
5. Bai X, Fu S, Zhang J, Fan R, Xu Y, Sun H, et al. Identification and Pathogenomic Analysis of an Escherichia coli Strain Producing a Novel Shiga Toxin 2 Subtype. Sci Rep (2018) 8(1):1–11. doi: 10.1038/s41598-018-25233-x
6. Yang X, Bai X, Zhang J, Sun H, Fu S, Fan R, et al. Escherichia coli Strains Producing a Novel Shiga Toxin 2 Subtype Circulate in China. Int J Med Microbiol (2020) 310(1):151377. doi: 10.1016/j.ijmm.2019.151377
7. Hughes AC, Zhang Y, Bai X, Xiong Y, Wang Y, Yang X, et al. Structural and Functional Characterization of Stx2k, a New Subtype of Shiga Toxin 2. Microorganisms (2020) 8(1):1–18. doi: 10.3390/microorganisms8010004
8. Hofer E, Cernela N, Stephan R. Shiga Toxin Subtypes Associated With Shiga Toxin-Producing Escherichia coli Strains Isolated From Red Deer, Roe Deer, Chamois, and Ibex. Foodborne Pathog Dis (2012) 9(9):792–5. doi: 10.1089/fpd.2012.1156
9. Orth D, Grif K, Khan AB, Naim A, Dierich MP, Würzner R. The Shiga Toxin Genotype Rather Than the Amount of Shiga Toxin or the Cytotoxicity of Shiga Toxin In Vitro Correlates With the Appearance of the Hemolytic Uremic Syndrome. Diagn Microbiol Infect Dis (2007) 59(3):235–42. doi: 10.1016/j.diagmicrobio.2007.04.013
10. Pianciola L, D’Astek BA, Mazzeo M, Chinen L, Masana M, Rivas M. Genetic Features of Human and Bovine Escherichia coli O157: H7 Strains Isolated in Argentina. Int J Med Microbiol (2016) 306:123–30. doi: 10.1016/j.ijmm.2016.02.005
11. Paton JC, Paton AW. Pathogenesis and Diagnosis of Shiga Toxin-Producing Escherichia coli Infections. Clin Microbiol Rev (1998) 11(3):450–79. doi: 10.1128/cmr.11.3.450
12. Doughty S, Sloan J, Bennett-Wood V, Robertson M, Robins-Browne RM, Hartland EL. Identification of a Novel Fimbrial Gene Cluster Related to Long Polar Fimbriae in Locus of Enterocyte Effacement-Negative Strains of Enterohemorrhagic Escherichia coli. Infect Immun (2002). doi: 10.1128/IAI.70.12.6761-6769.2002
13. Xicohtencatl-Cortes J, Monteiro-Neto V, Ledesma MA, Jordan DM, Francetic O, Kaper JB, et al. Intestinal Adherence Associated With Type IV Pili of Enterohemorrhagic Escherichia coli O157:H7. J Clin Invest (2007) 117(11):3519–29. doi: 10.1172/JCI30727
14. Tarr PI, Bilge SS, Vary JC, Jelacic S, Habeeb RL, Ward TR, et al. Iha: A Novel Escherichia coli O157:H7 Adherence-Conferring Molecule Encoded on a Recently Acquired Chromosomal Island of Conserved Structure. Infect Immun (2000) 68(3):1400–7. doi: 10.1128/IAI.68.3.1400-1407.2000
15. Herold S, Paton JC, Paton AW. Sab, a Novel Autotransporter of Locus of Enterocyte Effacement-Negative Shiga-Toxigenic Escherichia coli O113:H21, Contributes to Adherence and Biofilm Formation. Infect Immun (2009) 77(8):3234–43. doi: 10.1128/IAI.00031-09
16. Paton A, Srimanote P, Talbot U, Wang H, Paton J. A New Family of Potent AB(5) Cytotoxins Produced by Shiga Toxigenic Escherichia coli. J Exp Med (2004) 200(1):35–46. doi: 10.1084/jem.20040392
17. Cools P. The Role of Escherichia coli in Reproductive Health: State of the Art. Res Microbiol (2017) 168(9-10):892–901. doi: 10.1016/j.resmic.2017.02.002
18. Villar HE, Aubert V, Baserni MN, Jugo MB. Maternal Carriage of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli Isolates in Argentina. J Chemother (2013) 25(6):324–7. doi: 10.1179/1973947813Y.0000000081
19. Ulinski T, Lervat C, Ranchin B, Gillet Y, Floret D, Cochat P. Neonatal Hemolytic Uremic Syndrome After Mother-to-Child Transmission of Escherichia coli O157. Pediatr Nephrol (2005) 20(9):1334–5. doi: 10.1007/s00467-005-1871-3
20. Chart H, Perry NT, Cheasty T, Wright PA. The Kinetics of Antibody Production to Antigens of Escherichia coli O157 in a Pregnant Woman With Haemolytic Uraemic Syndrome. J Med Microbiol (2002) 51(6):522–5. doi: 10.1099/0022-1317-51-6-522
21. Carbonari CC, Fittipaldi N, Teatero S, Athey TBT, Pianciola L, Masana M, et al. Whole-Genome Sequencing Applied to the Molecular Epidemiology of Shiga Toxin-Producing Escherichia coli O157:H7 in Argentina. Genome Announc (2016) 4(6):4–5. doi: 10.1128/genomeA.01341-16
22. Pianciola L, Rivas M. Genotypic Features of Clinical and Bovine Escherichia coli O157 Strains Isolated in Countries With Different Associated-Disease Incidences. Microorganisms (2018) 6(2):36. doi: 10.3390/microorganisms6020036
23. Oderiz S, Leotta GA, Galli L. Detection and Characterization of Shiga Toxin-Producing Escherichia coli in Children Treated at an Inter-Zonal Pediatric Hospital in the City of La Plata. Rev Argent Microbiol (2018) 50(4):341–50. doi: 10.1016/j.ram.2017.08.008
24. Cundon C, Carbonari CC, Zolezzi G, Rivas M, Bentancor A. Putative Virulence Factors and Clonal Relationship of O174 Shiga Toxin-Producing Escherichia coli Isolated From Human, Food and Animal Sources. Vet Microbiol (2018) 215(11):29–34. doi: 10.1016/j.vetmic.2017.12.006
25. Burdet J, Zotta E, Franchi A, Ibarra C. Intraperitoneal Administration of Shiga Toxin Type 2 in Rats in the Late Stage of Pregnancy Produces Premature Delivery of Dead Fetuses. Placenta (2009) 30(6):491–6. doi: 10.1016/j.placenta.2009.03.012
26. Sacerdoti F, Amaral MM, Zotta E, Franchi AM, Ibarra C. Effects of Shiga Toxin Type 2 on Maternal and Fetal Status in Rats in the Early Stage of Pregnancy. BioMed Res Int (2014) 2014:384645. doi: 10.1155/2014/384645
27. Sacerdoti F, Mejías MP, Bruballa AC, Alvarez RS, Amaral MM, Palermo MS, et al. Immunization With BLS-Stx2B Chimera Totally Protects Dams From Early Pregnancy Loss Induced by Shiga Toxin Type 2 (Stx2) and Confers Anti-Stx2 Immunity to the Offspring. Vaccine (2016) 34(39):4732–7. doi: 10.1016/j.vaccine.2016.07.049
28. Tantawiwat S, Tansuphasiri U, Wongwit W, Wongchotigul V, Kitayaporn D. Development of Multiplex PCR for the Detection of Total Coliform Bacteria for Escherichia coli and Clostridium perfringens in Drinking Water. Southeast Asian J Trop Med Public Health (2005) 36(1):162–9.
29. Molina F, López-Acedo E, Tabla R, Roa I, Gómez A, Rebollo JE. Improved Detection of Escherichia coli and Coliform Bacteria by Multiplex PCR. BMC Biotechnol (2015) 15(1):1–9. doi: 10.1186/s12896-015-0168-2
30. Paton A, Paton J. Multiplex PCR for Direct Detection of Shiga Toxigenic Escherichia coli Strains Producing the Novel Subtilase Cytotoxin. J Clin Microbiol (2005) 43(6):2944–7. doi: 10.1128/JCM.43.6.2944-2947.2005
31. Blanco M, Blanco JE, Mora A, Dahbi G, Alonso MP, Gonza EA, et al. Serotypes, Virulence Genes, and Intimin Types of Shiga Toxin (Verotoxin)-Producing Escherichia coli Isolates From Cattle in Spain and Identification 427 of a New Intimin Variant Gene (Eae-Xi). J Clin Microbiol (2004) 42(2):645–51. doi: 10.1128/JCM.42.2.645-651.2004
32. Schmidt H, Beutin L, Karch H. Molecular Analysis of the Plasmid-Encoded Hemolysin of Escherichia coli O157:H7 Strain EDL 933. Infect Immun (1995) 63(3):1055–61. doi: 10.1128/iai.63.3.1055-1061.1995
33. Straszewski-Chavez SL, Abrahams VM, Alvero AB, Aldo PB, Ma Y, Guller S, et al. The Isolation and Characterization of a Novel Telomerase Immortalized First Trimester Trophoblast Cell Line, Swan 71. Placenta (2009). doi: 10.1016/j.placenta.2009.08.007
34. Thuraisamy T, Lodato PB. Influence of RNase E Deficiency on the Production of Stx2-Bearing Phages and Shiga Toxin in an RNase E-Inducible Strain of Enterohaemorrhagic Escherichia coli (EHEC) O157:H7. J Med Microbiol (2018) 67(5):724–32. doi: 10.1099/jmm.0.000728
35. Creydt VP, Silberstein C, Zotta E, Ibarra C. Cytotoxic Effect of Shiga Toxin-2 Holotoxin and Its B Subunit on Human Renal Tubular Epithelial Cells. Microbes Infect (2006) 8(2):410–9. doi: 10.1016/j.micinf.2005.07.005
36. Rocha LB, Luz DE, Moraes CTP, Caravelli A, Fernandes I, Guth BEC, et al. Interaction Between Shiga Toxin and Monoclonal Antibodies: Binding Characteristics and In Vitro Neutralizing Abilities. Toxins (2012) 4(9):729–47. doi: 10.3390/toxins4090729
37. Blanco J, Blanco M, Alonso MP, Blanco JE, Garabal J, González EA. Serogroups of Escherichia coli Strains Producing Cytotoxic Necrotizing Factors CNF1 and CNF2. FEMS Microbiol Lett (1992) 96(2–3):155–9. doi: 10.1016/0378-1097(92)90396-6
38. Romero R, Hassan SS, Gajer P, Tarca AL, Fadrosh DW, Nikita L, et al. The Composition and Stability of the Vaginal Microbiota of Normal Pregnant Women Is Different From That of Non-Pregnant Women. Microbiome (2014) 2(1):1–19. doi: 10.1186/2049-2618-2-4
39. Chen X, Lu Y, Chen T, Li R. The Female Vaginal Microbiome in Health and Bacterial Vaginosis. Front Cell Infect Microbiol (2021) 11:631972. doi: 10.3389/fcimb.2021.631972
40. Juliana NCA, Suiters MJM, Al-Nasiry S, Morré SA, Peters RPH, Ambrosino E. The Association Between Vaginal Microbiota Dysbiosis, Bacterial Vaginosis, and Aerobic Vaginitis, and Adverse Pregnancy Outcomes of Women Living in Sub-Saharan Africa: A Systematic Review. Front Public Health (2020) 8:567885(12). doi: 10.3389/fpubh.2020.567885
41. Moreno I, Codoñer FM, Vilella F, Valbuena D, Martinez-Blanch JF, Jimenez-Almazán J, et al. Evidence That the Endometrial Microbiota has an Effect on Implantation Success or Failure. Am J Obstet Gynecol (2016) 215(6):684–703. doi: 10.1016/j.ajog.2016.09.075
42. Smith SB, Ravel J. The Vaginal Microbiota, Host Defense and Reproductive Physiology. J Physiol (2017) 595(2):451–63. doi: 10.1113/JP271694
43. Han C, Li H, Han L, Wang C, Yan Y, Qi W, et al. Aerobic Vaginitis in Late Pregnancy and Outcomes of Pregnancy. Eur J Clin Microbiol Infect Dis (2019) 38(2):233–9. doi: 10.1007/s10096-018-3416-2
44. Riganelli L, Iebba V, Piccioni M, Illuminati I, Bonfiglio G, Neroni B, et al. Structural Variations of Vaginal and Endometrial Microbiota: Hints on Female Infertility. Front Cell Infect Microbiol (2020) 10:350(7). doi: 10.3389/fcimb.2020.00350
45. Sáez-López E, Guiral E, Fernández-Orth D, Villanueva S, Goncé A, López M, et al. Vaginal Versus Obstetric Infection Escherichia coli Isolates Among Pregnant Women: Antimicrobial Resistance and Genetic Virulence Profile. PloS One (2016) 11(1):1–11. doi: 10.1371/journal.pone.0146531
46. Krüger A, Lucchesi PMA. Shiga Toxins and Stx Phages: Highly Diverse Entities. Microbiol (United Kingdom) (2015) 161(3):1–12. doi: 10.1099/mic.0.000003
47. Liu TH, Wang HP, Cho FN, Wang JL, Hung CH, Chiou YH, et al. Rectovaginal Colonization With Pathogenic Escherichia coli During Pregnancy and Neonatal Outcomes. Infect Drug Resist (2019) 12:3103–12. doi: 10.2147/IDR.S207857
48. Muniesa M, Blanco JE, de Simón M, Serra-Moreno R, Blanch AR, Jofre J. Diversity of Stx2 Converting Bacteriophages Induced From Shiga-Toxin-Producing Escherichia coli Strains Isolated From Cattle. Microbiology (2004) 150(9):2959–71. doi: 10.1099/mic.0.27188-0
49. Berger M, Aijaz I, Berger P, Dobrindt U, Koudelka G. Transcriptional and Translational Inhibitors Block SOS Response and Shiga Toxin Expression in Enterohemorrhagic Escherichia coli. Sci Rep (2019) 9(1):1–9. doi: 10.1038/s41598-019-55332-2
50. Dos Santos LF, Gonçalves EM, Vaz TMI, Irino K, Guth BEC. Distinct Pathotypes of O113 Escherichia coli Strains Isolated From Humans and Animals in Brazil. J Clin Microbiol (2007) 45(6):2028–30. doi: 10.1128/JCM.00340-07
51. Cadona JS, Bustamante AV, Parma AE, Lucchesi PMA, Sanso AM. Distribution of Additional Virulence Factors Related to Adhesion and Toxicity in Shiga Toxin-Producing Escherichia coli Isolated From Raw Products in Argentina. Lett Appl Microbiol (2013) 56(6):449–55. doi: 10.1111/lam.12067
52. Colello R, Krüger A, Velez MV, Del Canto F, Etcheverría AI, Vidal R, et al. Identification and Detection of Iha Subtypes in LEE-Negative Shiga Toxin-Producing Escherichia coli (STEC) Strains Isolated From Humans, Cattle and Food. Heliyon (2019) 5(12):1–6. doi: 10.1016/j.heliyon.2019.e03015
53. Hauser E, Mellmann A, Semmler T, Stoeber H, Wieler LH, Karch H, et al. Phylogenetic and Molecular Analysis of Food-Borne Shiga Toxin-Producing Escherichia coli. Appl Environ Microbiol (2013) 79(8):2731–40. doi: 10.1128/AEM.03552-12
54. Galli L, Miliwebsky E, Irino K, Leotta G, Rivas M. Virulence Profile Comparison Between LEE-Negative Shiga Toxin-Producing Escherichia coli (STEC) Strains Isolated From Cattle and Humans. Vet Microbiol (2010) 143(2–4):307–13. doi: 10.1016/j.vetmic.2009.11.028
55. Frank C, Werber D, Cramer J, Askar M, Faber M, An der Heiden M, et al. Epidemic Profile of Shiga-Toxin–Producing. N Engl J Med (2011) 365(19):1771–80. doi: 10.1056/NEJMoa1106483
Keywords: Endocervical microbiota, pregnancy, STEC, virulence factors, Stx2
Citation: Scalise ML, Garimano N, Sanz M, Padola NL, Leonino P, Pereyra A, Casale R, Amaral MM, Sacerdoti F and Ibarra C (2022) Detection of Shiga Toxin-Producing Escherichia coli (STEC) in the Endocervix of Asymptomatic Pregnant Women. Can STEC Be a Risk Factor for Adverse Pregnancy Outcomes? Front. Endocrinol. 13:945736. doi: 10.3389/fendo.2022.945736
Received: 16 May 2022; Accepted: 14 June 2022;
Published: 20 July 2022.
Edited by:
Mariana Farina, CONICET Centro de Estudios Farmacológicos y Botánicos (CEFYBO), ArgentinaReviewed by:
Mohammad Yousef Alikhani, Hamadan University of Medical Sciences, IranCopyright © 2022 Scalise, Garimano, Sanz, Padola, Leonino, Pereyra, Casale, Amaral, Sacerdoti and Ibarra. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cristina Ibarra, aWJhcnJhQGZtZWQudWJhLmFy; Flavia Sacerdoti, ZnNhY2VyZG90aUBmbWVkLnViYS5hcg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.