
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Endocrinol., 15 March 2022
Sec. Cancer Endocrinology
Volume 13 - 2022 | https://doi.org/10.3389/fendo.2022.852382
This article is part of the Research TopicDouble-Edged Swords: Important Factors Connecting Metabolic Disorders and Cancer Development - From Basic Research to Translational ApplicationsView all 11 articles
 Makoto Matsushita1
Makoto Matsushita1 Kazutoshi Fujita1,2*
Kazutoshi Fujita1,2* Koji Hatano1
Koji Hatano1 Marco A. De Velasco2,3
Marco A. De Velasco2,3 Hirotsugu Uemura2
Hirotsugu Uemura2 Norio Nonomura1
Norio Nonomura1Prostate cancer (PCa) is the most common malignancy in men worldwide, thus developing effective prevention strategies remain a critical challenge. Insulin-like growth factor 1 (IGF-1) is produced mainly in the liver by growth hormone signaling and is necessary for normal physical growth. However, several studies have shown an association between increased levels of circulating IGF-1 and the risk of developing solid malignancies, including PCa. Because the IGF-1 receptor is overexpressed in PCa, IGF-1 can accelerate PCa growth by activating phosphoinositide 3-kinase and mitogen-activated protein kinase, or increasing sex hormone sensitivity. Short-chain fatty acids (SCFAs) are beneficial gut microbial metabolites, mainly because of their anti-inflammatory effects. However, we have demonstrated that gut microbiota-derived SCFAs increase the production of IGF-1 in the liver and prostate. This promotes the progression of PCa by the activation of IGF-1 receptor downstream signaling. In addition, the relative abundance of SCFA-producing bacteria, such as Alistipes, are increased in gut microbiomes of patients with high-grade PCa. IGF-1 production is therefore affected by the gut microbiome, dietary habits, and genetic background, and may play a central role in prostate carcinogenesis. The pro-tumor effects of bacteria and diet-derived metabolites might be potentially countered through dietary regimens and supplements. The specific diets or supplements that are effective are unclear. Further research into the “Gut–IGF-1–Prostate Axis” may help discover optimal diets and nutritional supplements that could be implemented for prevention of PCa.
Prostate cancer (PCa) is the most common malignancy in men worldwide and the fifth most common cause of cancer-related death with as many as 360,000 men dying of PCa annually (1). PCa morbidity varies somewhat by region and race, and has consistently been increasing in recent years (2, 3). Although androgen deprivation therapy is very effective for PCa, high-grade PCa becomes androgen resistant, which makes subsequent treatment challenging. Therefore, it is important to find new targets for the prevention and treatment of high-risk PCa.
The age-adjusted prevalence of latent PCa at autopsy in Japanese migrants in Hawaii >50 years old was higher than that of Japanese men living in Japan (25.6% vs. 20.5%), suggesting that PCa risk is not only altered by genetic factors but also by various environmental factors, such as diet (4). Many studies have reported that excessive intake of animal fat, carbohydrate, and dairy products increases PCa risk. However, different cohorts have yielded different results, and no consensus has been reached (5). This is because diet affects PCa development and progression through multiple mechanisms (6). The relationship between the diet and PCa is complex and not fully understood and as a result has hindered PCa prevention and treatment strategies via dietary interventions.
We recently identified a novel mechanism by which specific intestinal bacteria promote PCa through insulin-like growth factor I (IGF-1) signaling (7). Various studies have shown that diet can disrupt gut microbial composition resulting in dysbiosis and loss of homeostasis, affecting local intestinal disease as well as diseases and disorders in distant organs, such as the liver and brain (8–13). These relationships have been referred to as gut–liver axis and gut–brain axis, respectively. In cancer biology, the relationships between gut microbiota and various types of cancer, such as colorectal, hepatocellular, and breast cancer, have been well studied. In contrast, little is known about the influence of the gut microbiota on PCa (14). IGF-1 is implicated in the pathogenesis of PCa and may be the key player that links diet to prostate carcinogenesis and progression that is mediated by gut microbes. This review summarizes the present knowledge of the functions of IGF-1 in PCa progression, especially its relationship to diet and gut microbiota.
IGF-1 is a growth factor that plays a crucial role in cell proliferation and physical growth. IGF-1 signal transduction is mediated through the IGF-1 receptor (IGF1R) and insulin receptor (INSR). The structures of IGF1R and INSR are highly homologous (15). These receptors stimulated by IGF-1 activate tyrosine kinase activity directed at the β subunit, resulting in substrate phosphorylation, such as insulin receptor substrate (IRS) 1, IRS2, and Src homology collagen. The phosphorylated residues are recognized by the signaling molecules p85 and Grb2, which stimulate the phosphoinositide 3-kinase (PI3K) and mitogen-activated protein kinase (MAPK) signaling cascades. These signaling cascades mediate crucial biological functions of IGF-1 (15).
IGF-1 production in the healthy liver is mainly positively regulated by growth hormone (GH) signaling through the GH receptor pathway (16, 17). IGF-1 is released into circulation. IGF-1 is produced in other organs by various other cells that express IGF1R and is activated via autocrine signaling, although hepatocytes do not express enough IGF1R (18). Therefore, in liver-specific IGF-1-deficient mouse models, circulating IGF-1 levels are reduced to <20% of control mice and GH levels are elevated, but physical growth is normal (19). The bioavailability of circulating IGF-1 is regulated by the IGF-binding protein (IGFBP) family that blocks access to its receptor (20). In particular, IGFBP3 produced by Kupffer cells in the liver is important in IGF-1 homeostasis (21).
IGF-1 has tissue-specific roles through IGF1R and INSR. Muscle-specific IGF1R knockout mice display disrupted muscle fiber formation and reduced muscle weight early in development (22). However, muscle weight of differentiated muscles is not affected in mice with IGF1R knockout (23). IGF-1 has a significant effect on physical growth, but not on adult physiology. IGF-1 is also critical in bone growth and maintenance during postnatal life. IGF-1 can directly affect chondrocytes and osteoblasts and increase ephrin ligand-receptor signaling, leading to the differentiation of each cell. IGF-1 signaling also inhibits the formation of differentiated osteoclasts, contributing to bone growth (24). In the pancreas, IGF-1 signaling retains normal β-cell function, which is necessary to maintain glucose tolerance in vivo (25, 26). In vitro, IGF-1 stimulates expression of cellular communication network factor 5 (CCN5) and promotes β-cell proliferation (27). The phenotype of diabetes by blocking IGF-1 signaling is more obvious in mouse models lacking both IGF1R and INSR in β-cells (28). IGF-1 signaling is required for adipocyte development and function in adipose tissue, which is a major nutrient storage site. Mice lacking IGF1R and INSR in adipocytes contain almost no adipose tissue and develop significant diabetes, dyslipidemia, and fatty liver (29). IGF-1 is involved in myeloid cell function. IGF-1 activates M2 macrophages. Secretion of IGF-1 by the macrophages in turn leads to insulin resistance in mice fed a high-fat diet (30). A very important role of IGF-1 is its effect on the endocrine system. IGF-1 can directly support thyroid hormone production, and organ-specific IGF-1 signal loss reduces thyroid hormone and significantly increases thyroid stimulating hormone (TSH) levels (31). IGF1R and INSR knockout inhibit the development of the adrenal cortex and testes, and reduce testosterone levels. How IGF-1 signaling affects adrenal and testes function remains unknown (32). The role of IGF-1 signaling in prostate development and normal prostate physiology has not been established in vivo, however, silencing IGF-1 in not only the WPMY-1 prostate stroma cell line, and but also BPH-1, a prostate epithelium cell line, decreased cell proliferation and increased apoptosis rate in vitro (33). In human, IGF1R is located on the long arm of chromosome 15, and 36 different probable mutations have been reported (34). Most patients are heterozygous carriers, and all show pre- and postnatal growth retardation and dysmorphic features, such as a triangular face. The collective findings reveal that IGF-1 is an essential hormone for normal growth and maintain homeostasis.
IGF-1 is involved in several diseases. It is clear that diabetes is influenced by IGF-1 because of its effect on pancreatic β-cell function. IGF-1 increases nutrient-stimulated insulin release. The increased level of insulin increases IGF-1 production by stimulating GH signaling (35). Impaired insulin secretion due to type 1 diabetes lowers serum IGF-1 levels, and improves glycemic control in patients with type 2 diabetes leads to increased IGF-1 levels (36–38). Obese individuals have lower serum IGF-1 levels than normal-weight individuals, although over nourishment is associated with high insulin and IGF-1 levels (39). Several large studies found that serum IGF-1 levels are highest in both men and women with a body mass index of 24–27 kg/m2 (40, 41). A possible reason why serum IGF-1 levels are decreased in obese patients is that increased free IGF-1 fraction by reduction in IGFBP production enhances negative feedback on GH secretion by the pituitary gland (42). Therefore, IGF-1 bioactivity may not be decreased, even in obese patients. In a study of 27 samples of benign prostatic hyperplasia (BPH) patients, there was no significant relationship between serum IGF-1 levels and prostate volume (P = 0.91). However, the gene expression of IGF-1 in prostate tissue was significantly increased (P = 0.001) and the expression of IGFBP3 was significantly decreased (P = 0.003) in patients with larger prostate size (>30 mL) (43). Local IGF-1 was reportedly upregulated in hyperplastic prostate tissues (33). Patients with acromegaly characterized by GH hypersecretion display high IGF-1 levels, and acromegaly patients are highly susceptible to IGF-1 related diseases including diabetes mellitus and BPH, suggesting IGF-1 regulation of various diseases. Acromegaly patients <40 years of age were found to have significantly larger prostate than healthy men (18.2 vs. 28.5 mL, P < 0.001), and suppression of GH and IGF-1 using octreotide caused prostate shrinkage (44).
IGF-1 has been associated with the development and progression of some cancer types due to its function in activating the MAPK and PI3K signaling pathways (15). A positive association was observed between serum IGF-1 level and overall cancer risk in men in the United Kingdom (hazard ratio [HR] = 1.03 per 5-nmol/L increment in IGF-1) and specific cancer risk, such as prostate, melanoma, kidney, and thyroid (HR = 1.09, 1.08, 1.10, and 1.22, respectively) (45). In these cancer types, basic studies have also shown an association with IGF-1 signaling (46–48). Although melanoma cells do not produce IGF-1, activation of the MAPK and PI3K signaling pathway by paracrine stimulation of IGF-1 from stromal fibroblasts enhanced survival, migration, and growth of melanoma cells only from biologically early tumors (46). A cell line derived from metastatic clear cell renal cell carcinoma highly expresses IGFBP3 and IGF-1 compared to normal proximal tubule cell, and the autocrine actions of IGF-1 and IGFBP3 promote and inhibit cell proliferation, respectively (47). IGF-1 secreted by M2-like tumor−associated macrophages promote the invasion and stemness of C643 cells, an anaplastic thyroid carcinoma cell line, by activating PI3K signaling (48). Furthermore, IGF-1 is involved in bone metastasis biology, such as in homing, dormancy, colonization, and expansion (49). In an in vivo study, the presence of high IGF-1 levels in the primary tumor environment tended to induce cancer cells to metastasize to bone, and cancer cell lines that highly expressed IGF1R were prone to display enlarged bone mass (50, 51). IGF1R is highly expressed in PCa cells. Therefore, PCa may be susceptible to IGF-1 signaling (52). The relationship between IGF-1 and PCa is detailed in the next section.
IGF-1 promotes the proliferation of 22RV1 and DU145 PCa cell lines in vitro (7). In these cell lines, protein kinase B (AKT) in the PI3K pathway and extracellular signal-regulated kinase (ERK) in the MAPK pathway were phosphorylated in an IGF-1 dose dependent manner, suggesting that IGF-1 directly influences PCa proliferation (7). In vivo, IGF-1 expression was reportedly reduced in xenografts of Los Angeles PCa-4 (LAPC-4) in mice fed a low-fat diet, and tumor volume was suppressed (53). IGF-1 decreased miR-143 expression and increased IGF1R expression in PC-3 and DU145 cells, and made these cell lines more resistant to docetaxel treatment, suggesting that IGF-1 levels are also involved in resistance to treatment in PCa (54). IGF-1 is also implicated in castration-resistant PCa and has been shown to activate androgen receptor (AR) signaling in prostate cancer cells via the IGF-1R-forkhead box protein O1 (FOXO1) signaling axis (Figure 1) (55–57).
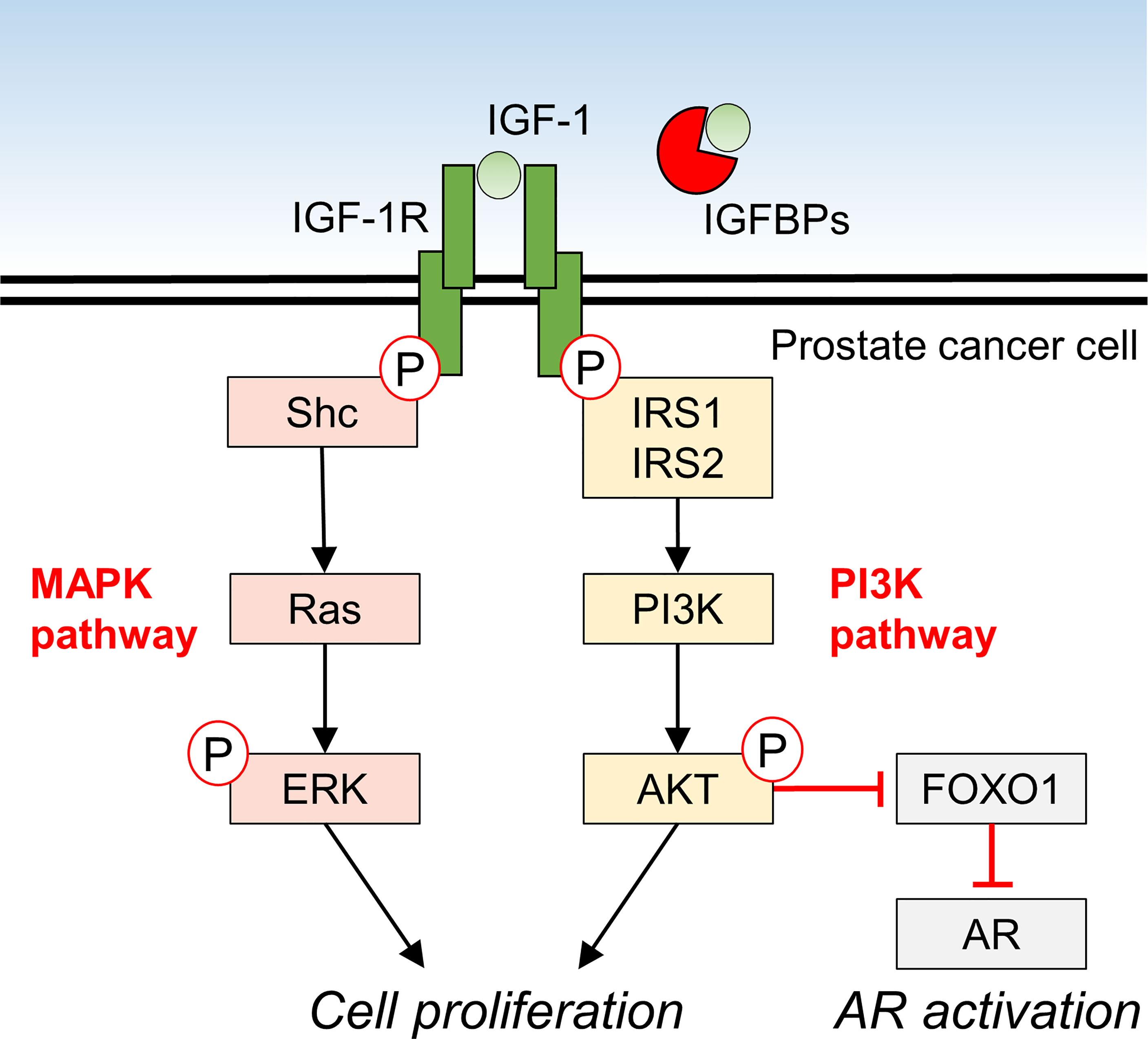
Figure 1 Molecular mechanism of IGF-1 signaling and downstream effects in prostate cancer cells.
Elevated blood IGF-1 levels increase the future risk of PCa in healthy men (45). Acromegaly patients with systemically high GH and IGF-1 levels also have significantly higher incidence of PCa and risk of PCa-related mortality (HR = 1.33 and 1.44, respectively), suggesting that IGF-1 has a positive effect on PCa development and progression, even in humans (58). Several studies reported that blood IGF-1 levels in elderly men with suspected PCa on screening tests are not associated with cancer positivity (59, 60). Serum IGF-1 levels in 94 men who required prostate biopsy showed no significant difference between positive and negative cancer (26.4 vs. 23.7 nmol/L; P = 0.08) (59). This discrepancy suggests that prostate epithelial cells may be at an increased risk of cancer development or progression only after prolonged exposure to high concentrations of IGF-1. Suppression of IGF-1 signaling is a potential therapeutic approach, because the IGF1R inhibitor in combination with castration inhibited PCa growth in rodent models of bone metastasis and subcutaneous xenografts (61, 62). However, in a phase 2 study, limsitinib, the most extensively evaluated IGF1R inhibitor, failed to significantly improve levels of prostate-specific antigen after 12 weeks of treatment and did not improve overall survival in men with metastatic castrate-resistant PCa (63). In the future, as a more potent treatment strategy, a combination of novel IGF1R inhibitors and existing prostate cancer therapies is expected to be effective.
In recent years, studies investigating the interactions between gut microbiota and its host has focused on recognizing an essential factor that influences homeostasis. One of the mechanisms by which intestinal bacteria affect humans is through bacterial structural components and their metabolites. Short-chain fatty acids (SCFAs) are major bacterial metabolites that play an important role in physiology. SCFAs include fatty acids with six or fewer carbon atoms. Of these, acetate (C2), propionate (C3), and butyrate (C4) are mainly produced by fermentation of dietary fiber by intestinal bacteria (64). Bacterial-derived SCFAs affect not only locally the gut but also distant organs in various ways. The anti-inflammatory effect is one of the major characteristics of SCFAs. Bacterial-derived butyrate promotes the differentiation of colonic regulatory T cells, suppresses inappropriate mucosal immunity, and improves local colitis and distant arthritis (65, 66). Propionic acids that reach the liver via the portal circulation increase glycogen synthesis and storage, improve insulin sensitivity, and repress lipogenesis in hepatocytes, resulting in the maintenance of energy homeostasis (67, 68). In the central nervous system (CNS), bacterial-derived SCFAs contribute to normal maturation of microglia via the free fatty acid receptor 2 (FFAR2) as a SCFA receptor (69). SCFAs also have some effects on several brain functions involved in circadian rhythm and appetite control (70).
SCFAs are also involved in the development and progression of cancer (71). In colorectal cancer, SCFAs, especially butyrate, function as anti-inflammatory metabolites and histone deacetylate (HDAC) inhibitors, resulting in the suppression of cancer progression (72). HDAC is a very important enzyme that modulates the expression of genes involved in signaling pathways, such as MAPK and Wnt (72). Therefore, accumulation of butyrate in some types of cancer cells that abundantly express a variety of HDACs, could inhibit their proliferation and promote apoptosis. In PCa, HDAC1, 2, and 3 are highly expressed (73). In vitro, sodium butyrate can directly decrease androgen receptor gene expression in LNCaP and C4-2 PCa cells, and can decrease the viability of these cells at concentrations >2.5 mM (74). However, it was demonstrated that high concentrations of butyrate were needed to inhibit colorectal cancer growth as an HDAC inhibitor, while <5 mM butyrate promoted cancer growth (75). SCFAs metabolized by intestinal bacteria are absorbed into the portal circulation and reach the liver, where most of the SCFAs are consumed (76). Therefore, only a small amount of bacteria-derived SCFAs can reach the prostate via systemic circulation. In vivo, bacteria-derived butyrate is not likely to work as an HDAC inhibitor in PCa due to this low concentration.
In young mice, gut microbiota-derived SCFAs are likely to induce IGF-1 production, suggesting that SCFAs modulate the bone and physical growth (77). The authors also described that germ-free mice and mice orally administered antibiotics showed lower cecal SCFA concentrations and IGF-1 production, resulting in decreased bone growth (77). SCFAs play a positive role in bone formation via an IGF-1-mediated mechanism. Unfortunately, the pathway by which SCFAs result in the elevation of IGF-1 is still not well understood.
We have reported that SCFAs metabolized by intestinal bacteria contribute to PCa growth by increasing systemic and prostate local IGF-1 productions, and revealed the “gut–prostate axis” involving bacterial metabolites (7). Prostate-specific phosphatase and tensin homolog (PTEN)-knockout mice [Pb-Cre+; Ptenfl/fl] were used as a PCa model. In these mice, a western-style high-fat diet (HFD) containing mainly lard accelerated PCa growth (78). This diet-induced PCa growth was inhibited by oral administration of metformin or celecoxib, as well as by an antibiotic mixture (ampicillin, vancomycin, neomycin, and metronidazole) (7, 78, 79). Antibiotics cause substantial changes in the composition of the gut microbiota of HFD-fed mice. Fecal SCFAs in the mice were reportedly reduced by 75%, resulting in decreased production of IGF-1 in the liver and prostate. In addition, phosphorylation of IGF-1R, ERK, and AKT was reduced in PCa cells of mice fed a HFD who received antibiotic, suggesting that decreased IGF-1 might suppress the activity of MAPK and PI3K signaling cascades. Oral supplementation of SCFAs to mice fed a HFD who received antibiotic resulted in increased serum IGF-1 levels and promoted prostate cancer growth. These results suggest that SCFAs derived from intestinal bacteria promote PCa growth through IGF-1 signaling, although butyrate in SCFAs may inhibit cancer cell proliferation as an HDAC inhibitor.
The examination of mice treated with antibiotics has revealed the absence of members of the family Rikenellaceae, order Clostridiales in the gut microbiota. Examination of the gut microbiota of men with a high-risk of PCa has revealed the increased abundance of genus Alistipes belonging to Rikenellaceae and the genus Lachnospira compared to men at low risk of PCa and those who are PCa-free (80). These bacterial taxa are associated with SCFA content in the stool and are SCFA-producing bacteria (81–86). These results suggest that SCFAs and their producing bacteria in the gut may be risk factors for PCa in humans and mice. Consumption of milk and other dairy products increase the dietary intake of SCFAs, thus resulting in the increase of serum IGF-1 levels and that is because these are the among the few foods that contain butyrate (87, 88). Many epidemiological studies have indicated that consumption of milk and dairy products increases the risk of PCa (5). This increased risk may be due to the butyrate contained in these foods (88). While dairy products are essential for nutrition and may a preventive effect in various diseases, including colorectal cancer (89), the roles are complex and most likely context-dependent. For example, low-fat milk containing no SCFAs does not increase the risk of PCa, unlike whole milk (90). Additional studies have reinforced this notion. In the NIH-AACR Diet and Health Study, during 7 years of follow-up, the highest quintile of dairy food intake had a significantly lower risk of colorectal cancer (relative risk [RR] = 0.85, P = 0.01) and a higher risk of prostate cancer (RR = 1.06, P = 0.01) compared to the lowest quintile (91). We hypothesize that the regulation of IGF-1 signaling contributing to prostate cancer risk in a real-world setting is increased by intestinal factors.
There are still some questions that need to be clarified regarding the gut–prostate axis involving SCFAs and IGF-1 signaling. It has been also reported that butyrate and propionate may have inhibitory effects on prostate cancer (74, 92). In our animal study, we found that a mixture of SCFAs (acetate, propionate, and butyrate) promoted prostate cancer growth (7), but we have not been able to determine which types of SCFAs are responsible for this promotive effect and at what concentration. Perhaps these may act cooperatively. Furthermore, although several G protein-coupled receptors, such as GPR41 and GPR43, are known as SCFA receptors, neither the receptor nor the signaling pathway(s) involved in the regulation of IGF-1 by SCFAs have been established (93, 94). Finally, the impact of interventions on the gut–prostate axis in human using fecal microbiota transplantation (FMT) or pro/prebiotics has not been studied. There are however several basic studies that have reported that FMT derived from prostate cancer individuals altered prostate cancer progression in mouse (95, 96), and we think that this axis may be a promising therapeutic target.
IGF-1 is an essential hormone for physical growth and has various effects in several diseases, especially prostate cancer, where it functions as an exacerbating factor. In vivo, local and systemic IGF-1 production might be regulated by SCFAs, which is in turn influenced by gut factors, such as gut microbiota and diet. The data thus far indicate that the gut–IGF-1–prostate axis is connected by SCFAs (Figure 2). This axis could provide a new direction for effective PCa treatment and prevention strategies. However, there are factors that remain unclear such as detailed mechanisms of IGF-1 regulation by SCFAs and the continuous control of SCFA levels in humans. Further study of the gut–IGF-1–prostate axis is needed to provide additional answers.
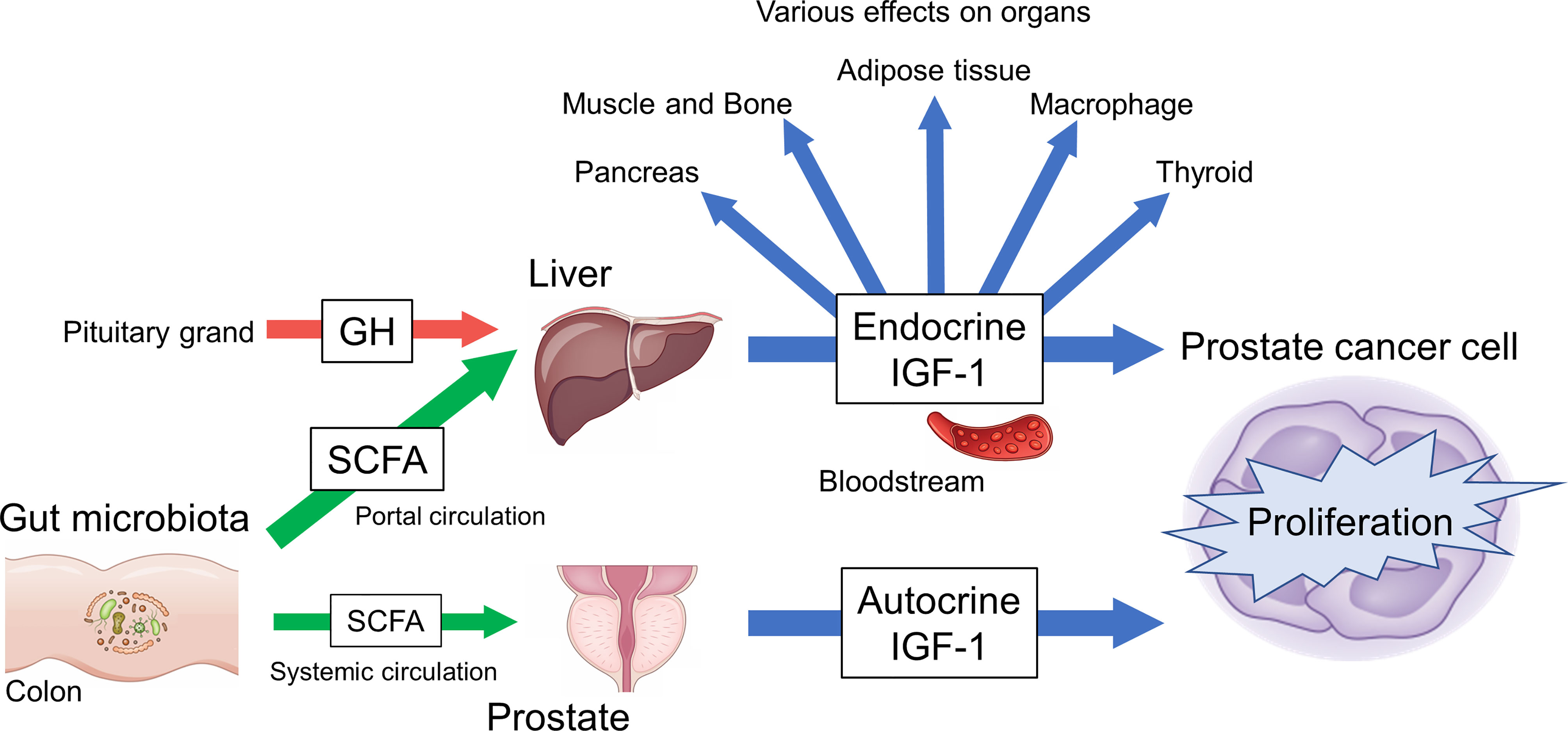
Figure 2 Overview of gut–IGF-1–prostate axis mediated by gut microbiota-derived SCFAs.
MM: writing—original draft preparation. KF, KH, and MV: conceptualization and writing—review and editing. HU and NN: supervision. All authors have read and agreed to the published version of the manuscript.
This work was supported by JSPS KAKENHI Grant Number JP21K09421.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin (2018) 68:394–424. doi: 10.3322/caac.21492
2. Marima R, Hull R, Mathabe K, Setlai B, Batra J, Sartor O, et al. Prostate Cancer Racial, Socioeconomic, Geographic Disparities: Targeting the Genomic Landscape and Splicing Events in Search for Diagnostic, Prognostic and Therapeutic Targets. Am J Cancer Res (2021) 11:1012–30.
3. Lloyd T, Hounsome L, Mehay A, Mee S, Verne J, Cooper A. Lifetime Risk of Being Diagnosed With, or Dying From, Prostate Cancer by Major Ethnic Group in England 2008-2010. BMC Med (2015) 13:171. doi: 10.1186/s12916-015-0405-5
4. Yatani R, Chigusa I, Akazaki K, Stemmermann GN, Welsh RA, Correa P. Geographic Pathology of Latent Prostatic Carcinoma. Int J cancer (1982) 29:611–6. doi: 10.1002/ijc.2910290602
5. Matsushita M, Fujita K, Nonomura N. Influence of Diet and Nutrition on Prostate Cancer. Int J Mol Sci (2020) 21:1447. doi: 10.3390/ijms21041447
6. Narita S, Nara T, Sato H, Koizumi A, Huang M, Inoue T. Research Evidence on High-Fat Diet-Induced Prostate Cancer Development and Progression. J Clin Med (2019) 8:597. doi: 10.3390/jcm8050597
7. Matsushita M, Fujita K, Hayashi T, Kayama H, Motooka D, Hase H, et al. Gut Microbiota-Derived Short-Chain Fatty Acids Promote Prostate Cancer Growth via IGF1 Signaling. Cancer Res (2021) 81:4014–26. doi: 10.1158/0008-5472.CAN-20-4090
8. Stange EF, Schroeder BO. Microbiota and Mucosal Defense in IBD: An Update. Expert Rev Gastroenterol Hepatol (2019) 13:963–76. doi: 10.1080/17474124.2019.1671822
9. Nishida A, Inoue R, Inatomi O, Bamba S, Naito Y, Andoh A. Gut Microbiota in the Pathogenesis of Inflammatory Bowel Disease. Clin J Gastroenterol (2018) 11:1–10. doi: 10.1007/s12328-017-0813-5
10. Yu LX, Schwabe RF. The Gut Microbiome and Liver Cancer: Mechanisms and Clinical Translation. Nat Rev Gastroenterol Hepatol (2017) 14:527–39. doi: 10.1038/nrgastro.2017.72
11. Usami M, Miyoshi M, Yamashita H. Gut Microbiota and Host Metabolism in Liver Cirrhosis. World J Gastroenterol (2015) 21:11597–608. doi: 10.3748/wjg.v21.i41.11597
12. Doifode T, Giridharan VV, Generoso JS, Bhatti G, Collodel A, Schulz PE. The Impact of the Microbiota-Gut-Brain Axis on Alzheimer’s Disease Pathophysiology. Pharmacol Res (2021) 164:105314. doi: 10.1016/j.phrs.2020.105314
13. Rutsch A, Kantsjö JB, Ronchi F. The Gut-Brain Axis: How Microbiota and Host Inflammasome Influence Brain Physiology and Pathology. Front Immunol (2020) 11:604179. doi: 10.3389/fimmu.2020.604179
14. Dzutsev A, Badger JH, Perez-Chanona E, Roy S, Salcedo R, Smith CK, et al. Microbes and Cancer. Annu Rev Immunol (2017) 35:199–228. doi: 10.1146/annurev-immunol-051116-052133
15. Hakuno F, Takahashi SI. IGF1 Receptor Signaling Pathways. J Mol Endocrinol (2018) 61:T69–86. doi: 10.1530/JME-17-0311
16. Feigerlova E, Hwa V, Derr MA, Rosenfeld RG. Current Issues on Molecular Diagnosis of GH Signaling Defects. Endocr Dev (2013) 24:118–27. doi: 10.1159/000342586
17. Rotwein P. Mapping the Growth Hormone–Stat5b–IGF-I Transcriptional Circuit. Trends Endocrinol Metab (2012) 23:186–93. doi: 10.1016/j.tem.2012.01.001
18. Kineman RD, del Rio-Moreno M, Sarmento-Cabral A. 40 YEARS of IGF1: Understanding the Tissue-Specific Roles of IGF1/IGF1R in Regulating Metabolism Using the Cre/Loxp System. J Mol Endocrinol (2018) 61:T187–98. doi: 10.1530/JME-18-0076
19. LeRoith D. Clinical Relevance of Systemic and Local IGF-I: Lessons From Animal Models. Pediatr Endocrinol Rev (2008) 5:739–43.
20. Clemmons DR. Role of IGF-Binding Proteins in Regulating IGF Responses to Changes in Metabolism. J Mol Endocrinol (2018) 61:T139–69. doi: 10.1530/JME-18-0016
21. Novosyadlyy R, Dargel R, Scharf JG. Expression of Insulin-Like Growth Factor-I and Insulin-Like Growth Factor Binding Proteins During Thioacetamide-Induced Liver Cirrhosis in Rats. Growth Horm IGF Res (2005) 15:313–23. doi: 10.1016/j.ghir.2005.06.015
22. O’Neill BT, Lauritzen HP, Hirshman MF, Smyth G, Goodyear LJ, Kahn CR. Differential Role of Insulin/IGF-1 Receptor Signaling in Muscle Growth and Glucose Homeostasis. Cell Rep (2015) 11:1220–35. doi: 10.1016/j.celrep.2015.04.037
23. Mavalli MD, DiGirolamo DJ, Fan Y, Riddle RC, Campbell KS, Van Groen T, et al. Distinct Growth Hormone Receptor Signaling Modes Regulate Skeletal Muscle Development and Insulin Sensitivity in Mice. J Clin Invest (2010) 120:4007–20. doi: 10.1172/JCI42447
24. Lindsey RC, Rundle CH, Mohan S. Role of IGF1 and EFN-EPH Signaling in Skeletal Metabolism. J Mol Endocrinol (2018) 61:T87–T102. doi: 10.1530/JME-17-0284
25. Xuan S, Kitamura T, Nakae J, Politi K, Kido Y, Fisher PE, et al. Defective Insulin Secretion in Pancreatic Beta Cells Lacking Type 1 IGF Receptor. J Clin Invest (2002) 110:1011–9. doi: 10.1172/JCI15276
26. Kulkarni RN, Holzenberger M, Shih DQ, Ozcan U, Stoffel M, Magnuson MA, et al. Beta-Cell-Specific Deletion of the Igf1 Receptor Leads to Hyperinsulinemia and Glucose Intolerance But Does Not Alter Beta-Cell Mass. Nat Genet (2002) 31:111–5. doi: 10.1038/ng872
27. Chowdhury S, Wang X, Srikant CB, Li Q, Fu M, Gong YJ, et al. IGF-I Stimulates CCN5/WISP2 Gene Expression in Pancreatic β-Cells, Which Promotes Cell Proliferation and Survival Against Streptozotocin. Endocrinology (2014) 155:1629–42. doi: 10.1210/en.2013-1735
28. Ueki K, Okada T, Hu J, Chong WL, Assmann A, Dahlgren GM, et al. Total Insulin and IGF-I Resistance in Pancreatic Beta Cells Causes Overt Diabetes. Nat Genet (2006) 38:583–8. doi: 10.1038/ng1787
29. Boucher J, Softic S, El Ouaamari A, Krumpoch MT, Kleinridders A, Kulkarni RN, et al. Differential Roles of Insulin and IGF-1 Receptors in Adipose Tissue Development and Function. Diabetes (2016) 65:2201–13. doi: 10.2337/db16-0212
30. Spadaro O, Camell CD, Bosurgi L, Nguyen KY, Youm YH, Rothlin CV, et al. IGF1 Shapes Macrophage Activation in Response to Immunometabolic Challenge. Cell Rep (2017) 19:225–34. doi: 10.1016/j.celrep.2017.03.046
31. Müller K, Führer D, Mittag J, Klöting N, Blüher M, Weiss RE, et al. TSH Compensates Thyroid-Specific IGF-I Receptor Knockout and Causes Papillary Thyroid Hyperplasia. Mol Endocrinol (2011) 25:1867–79. doi: 10.1210/me.2011-0065
32. Neirijnck Y, Calvel P, Kilcoyne KR, Kühne F, Stévant I, Griffeth RJ, et al. Insulin and IGF1 Receptors Are Essential for the Development and Steroidogenic Function of Adult Leydig Cells. FASEB J (2018) 32:3321–35. doi: 10.1096/fj.201700769RR
33. Qian Q, He W, Liu D, Yin J, Ye L, Chen P, et al. M2a Macrophage can Rescue Proliferation and Gene Expression of Benign Prostate Hyperplasia Epithelial and Stroma Cells From Insulin-Like Growth Factor 1 Knockdown. Prostate (2021) 81:530–42. doi: 10.1002/pros.24131
34. Walenkamp MJE, Robers JML, Wit JM, Zandwijken GRJ, van Duyvenvoorde HA, Oostdijk W, et al. Phenotypic Features and Response to GH Treatment of Patients With a Molecular Defect of the IGF-1 Receptor. J Clin Endocrinol Metab (2019) 104:3157–71. doi: 10.1210/JC.2018-02065
35. Lewitt M, Dent M, Hall K. The Insulin-Like Growth Factor System in Obesity, Insulin Resistance and Type 2 Diabetes Mellitus. J Clin Med (2014) 3:1561–74. doi: 10.3390/jcm3041561
36. Suda K, Matsumoto R, Fukuoka H, Iguchi G, Hirota Y, Nishizawa H, et al. The Influence of Type 2 Diabetes on Serum GH and IGF-I Levels in Hospitalized Japanese Patients. Growth Horm IGF Res (2016) 29:4–10. doi: 10.1016/j.ghir.2016.03.002
37. Kanazawa I, Yamaguchi T, Sugimoto T. Effects of Intensive Glycemic Control on Serum Levels of Insulin-Like Growth Factor-I and Dehydroepiandrosterone Sulfate in Type 2 Diabetes Mellitus. J Endocrinol Invest (2012) 35:469–72. doi: 10.3275/8033
38. Hata S, Mori H, Yasuda T, Irie Y, Yamamoto T, Umayahara Y, et al. A Low Serum IGF-1 Is Correlated With Sarcopenia in Subjects With Type 1 Diabetes Mellitus: Findings From a Post-Hoc Analysis of the Idiamond Study. Diabetes Res Clin Pract (2021) 179:108998. doi: 10.1016/j.diabres.2021.108998
39. Kaaks R, Lukanova A. Energy Balance and Cancer: The Role of Insulin and Insulin-Like Growth Factor-I. Proc Nutr Soc (2001) 60:91–106. doi: 10.1079/pns200070
40. Holmes MD, Pollak MN, Hankinson SE. Lifestyle Correlates of Plasma Insulin-Like Growth Factor I and Insulin-Like Growth Factor Binding Protein 3 Concentrations. Cancer Epidemiol Biomarkers Prev (2002) 11:862–7.
41. Allen NE, Appleby PN, Kaaks R, Rinaldi S, Davey GK, Key TJ. Lifestyle Determinants of Serum Insulin-Like Growth-Factor-I (IGF-I), C-Peptide and Hormone Binding Protein Levels in British Women. Cancer Causes Control (2003) 14:65–74. doi: 10.1023/a:1022518321634
42. Calle EE, Kaaks R. Overweight, Obesity and Cancer: Epidemiological Evidence and Proposed Mechanisms. Nat Rev Cancer (2004) 4:579–91. doi: 10.1038/nrc1408
43. Sreenivasulu K, Nandeesha H, Dorairajan LN, Rajappa M, Vinayagam V, Cherupanakkal C. Gene Expression of Insulin Receptor, Insulin-Like Growth Factor Increases and Insulin-Like Growth Factor-Binding Protein-3 Reduces With Increase in Prostate Size in Benign Prostatic Hyperplasia. Aging Male (2018) 21:138–44. doi: 10.1080/13685538.2017.1401994
44. Colao A, Marzullo P, Ferone D, Spiezia S, Cerbone G, Marinò V, et al. Prostatic Hyperplasia: An Unknown Feature of Acromegaly. J Clin Endocrinol Metab (1998) 83:775–9. doi: 10.1210/jcem.83.3.4645
45. Qian F, Huo D. Circulating Insulin-Like Growth Factor-1 and Risk of Total and 19 Site-Specific Cancers: Cohort Study Analyses From the UK Biobank. Cancer Epidemiol Biomarkers Prev (2020) 29:2332–42. doi: 10.1158/1055-9965.EPI-20-0743
46. Satyamoorthy K, Li G, Vaidya B, Patel D, Herlyn M. Insulin-Like Growth Factor-1 Induces Survival and Growth of Biologically Early Melanoma Cells Through Both the Mitogen-Activated Protein Kinase and Beta-Catenin Pathways. Cancer Res (2001) 61:7318–24.
47. Cheung CW, Vesey DA, Nicol DL, Johnson DW. The Roles of IGF-I and IGFBP-3 in the Regulation of Proximal Tubule, and Renal Cell Carcinoma Cell Proliferation. Kidney Int (2004) 65:1272–9. doi: 10.1111/J.1523-1755.2004.00535.X
48. Lv J, Liu C, Chen FK, Feng ZP, Jia L, Liu PJ, et al. M2-Like Tumour-Associated Macrophage-Secreted IGF Promotes Thyroid Cancer Stemness and Metastasis by Activating the PI3K/AKT/Mtor Pathway. Mol Med Rep (2021) 24:604. doi: 10.3892/MMR.2021.12249
49. Rieunier G, Wu X, Macaulay VM, Lee AV, Weyer-Czernilofsky U, Bogenrieder T. Bad to the Bone: The Role of the Insulin-Like Growth Factor Axis in Osseous Metastasis. Clin Cancer Res (2019) 25:3479–85. doi: 10.1158/1078-0432.CCR-18-2697
50. Zhang XH, Jin X, Malladi S, Zou Y, Wen YH, Brogi E, et al. Selection of Bone Metastasis Seeds by Mesenchymal Signals in the Primary Tumor Stroma. Cell (2013) 154:1060–73. doi: 10.1016/j.cell.2013.07.036
51. Van Golen CM, Schwab TS, Kim B, Soules ME, Su Oh S, Fung K, et al. Insulin-Like Growth Factor-I Receptor Expression Regulates Neuroblastoma Metastasis to Bone. Cancer Res (2006) 66:6570–8. doi: 10.1158/0008-5472.CAN-05-1448
52. Heidegger I, Massoner P, Sampson N, Klocker H. The Insulin-Like Growth Factor (IGF) Axis as an Anticancer Target in Prostate Cancer. Cancer Lett (2015) 367:113–21. doi: 10.1016/j.canlet.2015.07.026
53. Ngo TH, Barnard RJ, Cohen P, Freedland S, Tran C, DeGregorio F, et al. Effect of Isocaloric Low-Fat Diet on Human LAPC-4 Prostate Cancer Xenografts in Severe Combined Immunodeficient Mice and the Insulin-Like Growth Factor Axis. Clin Cancer Res (2003) 9:2734–43.
54. Niu XB, Fu GB, Wang L, Ge X, Liu WT, Wen YY, et al. Insulin-Like Growth Factor-I Induces Chemoresistence to Docetaxel by Inhibiting Mir-143 in Human Prostate Cancer. Oncotarget (2017) 8:107157–66. doi: 10.18632/oncotarget.22362
55. Weyer-Czernilofsky U, Hofmann MH, Friedbichler K, Baumgartinger R, Adam PJ, Solca F, et al. Antitumor Activity of the IGF-1/IGF-2-Neutralizing Antibody Xentuzumab (BI 836845) in Combination With Enzalutamide in Prostate Cancer Models. Mol Cancer Ther (2020) 19:1059–69. doi: 10.1158/1535-7163.MCT-19-0378
56. Plymate SR, Haugk K, Coleman I, Woodke L, Vessella R, Nelson P, et al. An Antibody Targeting the Type I Insulin-Like Growth Factor Receptor Enhances the Castration-Induced Response in Androgen-Dependent Prostate Cancer. Clin Cancer Res (2007) 13:6429–39. doi: 10.1158/1078-0432.CCR-07-0648
57. Fan WQ, Yanase T, Morinaga H, Okabe T, Nomura M, Daitoku H, et al. Insulin-Like Growth Factor 1/Insulin Signaling Activates Androgen Signaling Through Direct Interactions of Foxo1 With Androgen Receptor. J Biol Chem (2007) 282:7329–38. doi: 10.1074/jbc.M610447200
58. Watts EL, Goldacre R, Key TJ, Allen NE, Travis RC, Perez-Cornago A. Hormone-Related Diseases and Prostate Cancer: An English National Record Linkage Study. Int J cancer (2020) 147:803–10. doi: 10.1002/ijc.32808
59. Cutting CW, Hunt C, Nisbet JA, Bland JM, Dalgleish AG, Kirby RS. Serum Insulin-Like Growth Factor-1 Is Not a Useful Marker of Prostate Cancer. BJU Int (1999) 83:996–9. doi: 10.1046/j.1464-410x.1999.00088.x
60. Finne P, Auvinen A, Koistinen H, Zhang WM, Määttänen L, Rannikko S, et al. Insulin-Like Growth Factor I Is Not a Useful Marker of Prostate Cancer in Men With Elevated Levels of Prostate-Specific Antigen. J Clin Endocrinol Metab (2000) 85:2744–7. doi: 10.1210/jcem.85.8.6725
61. Nordstrand A, Bergström SH, Thysell E, Bovinder-Ylitalo E, Lerner UH, Widmark A, et al. Inhibition of the Insulin-Like Growth Factor-1 Receptor Potentiates Acute Effects of Castration in a Rat Model for Prostate Cancer Growth in Bone. Clin Exp Metastasis (2017) 34:261–71. doi: 10.1007/s10585-017-9848-8
62. Fahrenholtz CD, Beltran PJ, Burnstein KL. Targeting IGF-IR With Ganitumab Inhibits Tumorigenesis and Increases Durability of Response to Androgen-Deprivation Therapy in Vcap Prostate Cancer Xenografts. Mol Cancer Ther (2013) 12:394–404. doi: 10.1158/1535-7163.MCT-12-0648
63. Barata P, Cooney M, Tyler A, Wright J, Dreicer R, Garcia JA. A Phase 2 Study of OSI-906 (Linsitinib, an Insulin-Like Growth Factor Receptor-1 Inhibitor) in Patients With Asymptomatic or Mildly Symptomatic (Non-Opioid Requiring) Metastatic Castrate Resistant Prostate Cancer (CRPC). Invest New Drugs (2018) 36:451–7. doi: 10.1007/s10637-018-0574-0
64. Cook SI, Sellin JH. Review Article: Short Chain Fatty Acids in Health and Disease. Aliment Pharmacol Ther (1998) 12:499–507. doi: 10.1046/j.1365-2036.1998.00337.x
65. Takahashi D, Hoshina N, Kabumoto Y, Maeda Y, Suzuki A, Tanabe H, et al. Microbiota-Derived Butyrate Limits the Autoimmune Response by Promoting the Differentiation of Follicular Regulatory T Cells. EBioMedicine (2020) 58:102913. doi: 10.1016/j.ebiom.2020.102913
66. Furusawa Y, Obata Y, Fukuda S, Endo TA, Nakato G, Takahashi D, et al. Commensal Microbe-Derived Butyrate Induces the Differentiation of Colonic Regulatory T Cells. Nature (2013) 504:446–50. doi: 10.1038/nature12721
67. Weitkunat K, Schumann S, Nickel D, Kappo KA, Petzke KJ, Kipp AP, et al. Importance of Propionate for the Repression of Hepatic Lipogenesis and Improvement of Insulin Sensitivity in High-Fat Diet-Induced Obesity. Mol Nutr Food Res (2016) 60:2611–21. doi: 10.1002/mnfr.201600305
68. El Hage R, Hernandez-Sanabria E, Arroyo MC, van de Wiele T. Supplementation of a Propionate-Producing Consortium Improves Markers of Insulin Resistance in an In Vitro Model of Gut-Liver Axis. Am J Physiol Endocrinol Metab (2020) 318:E742–9. doi: 10.1152/ajpendo.00523.2019
69. Erny D, Hrabě de Angelis AL, Jaitin D, Wieghofer P, Staszewski O, David E, et al. Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat Neurosci (2015) 18:965–77. doi: 10.1038/nn.4030
70. Silva YP, Bernardi A, Frozza RL. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front Endocrinol (Lausanne) (2020) 11:25. doi: 10.3389/fendo.2020.00025
71. Mirzaei R, Afaghi A, Babakhani S, Sohrabi MR, Hosseini-Fard SR, Babolhavaeji K, et al. Role of Microbiota-Derived Short-Chain Fatty Acids in Cancer Development and Prevention. BioMed Pharmacother (2021) 139:111619. doi: 10.1016/j.biopha.2021.111619
72. Wu X, Wu Y, He L, Wu L, Wang X, Liu Z. Effects of the Intestinal Microbial Metabolite Butyrate on the Development of Colorectal Cancer. J Cancer (2018) 9:2510–7. doi: 10.7150/jca.25324
73. Li Y, Seto E. Hdacs and HDAC Inhibitors in Cancer Development and Therapy. Cold Spring Harb Perspect Med (2016) 6:a026831. doi: 10.1101/cshperspect.a026831
74. Paskova L, Smesny Trtkova K, Fialova B, Benedikova A, Langova K, Kolar Z. Different Effect of Sodium Butyrate on Cancer and Normal Prostate Cells. Toxicol In Vitro (2013) 27:1489–95. doi: 10.1016/j.tiv.2013.03.002
75. Donohoe DR, Collins LB, Wali A, Bigler R, Sun W, Bultman SJ. The Warburg Effect Dictates the Mechanism of Butyrate-Mediated Histone Acetylation and Cell Proliferation. Mol Cell (2012) 48:612–26. doi: 10.1016/j.molcel.2012.08.033
76. Morrison DJ, Preston T. Formation of Short Chain Fatty Acids by the Gut Microbiota and Their Impact on Human Metabolism. Gut Microbes (2016) 7:189–200. doi: 10.1080/19490976.2015.1134082
77. Yan J, Herzog JW, Tsang K, Brennan CA, Bower MA, Garrett WS, et al. Gut Microbiota Induce IGF-1 and Promote Bone Formation and Growth. Proc Natl Acad Sci USA. (2016) 113:E7554–63. doi: 10.1073/pnas.1607235113
78. Hayashi T, Fujita K, Nojima S, Hayashi Y, Nakano K, Ishizuya Y, et al. High-Fat Diet-Induced Inflammation Accelerates Prostate Cancer Growth via IL6 Signaling. Clin Cancer Res (2018) 24:4309–18. doi: 10.1158/1078-0432.CCR-18-0106
79. Hayashi T, Fujita K, Matsushita M, Hayashi Y, Uemura M, Nonomura N. Metformin Inhibits Prostate Cancer Growth Induced by a High-Fat Diet in Pten-Deficient Model Mice. Int J Urol (2019) 26:307–9. doi: 10.1111/iju.13847
80. Matsushita M, Fujita K, Motooka D, Hatano K, Fukae S, Kawamura N, et al. The Gut Microbiota Associated With High-Gleason Prostate Cancer. Cancer Sci (2021) 112:3125–35. doi: 10.1111/cas.14998
81. Wang B, Kong Q, Li X, Zhao J, Zhang H, Chen W, et al. A High-Fat Diet Increases Gut Microbiota Biodiversity and Energy Expenditure Due to Nutrient Difference. Nutrients (2020) 12:1–20. doi: 10.3390/nu12103197
82. Shi H, Chang Y, Gao Y, Wang X, Chen X, Wang Y, et al. Dietary Fucoidan of Acaudina Molpadioides Alters Gut Microbiota and Mitigates Intestinal Mucosal Injury Induced by Cyclophosphamide. Food Funct (2017) 8:3383–93. doi: 10.1039/c7fo00932a
83. Meehan CJ, Beiko RG. A Phylogenomic View of Ecological Specialization in the Lachnospiraceae, a Family of Digestive Tract-Associated Bacteria. Genome Biol Evol (2014) 6:703–13. doi: 10.1093/gbe/evu050
84. Xu Y, Yu Y, Shen Y, Li Q, Lan J, Wu Y, et al. Effects of Bacillus Subtilis and Bacillus Licheniformis on Growth Performance, Immunity, Short Chain Fatty Acid Production, Antioxidant Capacity, and Cecal Microflora in Broilers. Poult Sci (2021) 100:101358. doi: 10.1016/j.psj.2021.101358
85. Li B, Chen H, Cao L, Hu Y, Chen D, Yin Y. Escherichia Coli Exopolysaccharides Induced by Ceftriaxone Regulated Human Gut Microbiota In Vitro. Front Microbiol (2021) 12:634204. doi: 10.3389/fmicb.2021.634204
86. Allen JM, Mailing LJ, Niemiro GM, Moore R, Cook MD, White BA, et al. Exercise Alters Gut Microbiota Composition and Function in Lean and Obese Humans. Med Sci Sports Exerc (2018) 50:747–57. doi: 10.1249/MSS.0000000000001495
87. Beasley JM, Gunter MJ, LaCroix AZ, Prentice RL, Neuhouser ML, Tinker LF, et al. Associations of Serum Insulin-Like Growth Factor-I and Insulin-Like Growth Factor-Binding Protein 3 Levels With Biomarker-Calibrated Protein, Dairy Product and Milk Intake in the Women’s Health Initiative. Br J Nutr (2014) 111:847–53. doi: 10.1017/S000711451300319X
88. Harrison S, Lennon R, Holly J, Higgins JPT, Gardner M, Perks C, et al. Does Milk Intake Promote Prostate Cancer Initiation or Progression via Effects on Insulin-Like Growth Factors (Igfs)? A Systematic Review and Meta-Analysis. Cancer Causes Control (2017) 28:497–528. doi: 10.1007/S10552-017-0883-1
89. Pereira PC. Milk Nutritional Composition and Its Role in Human Health. Nutrition (2014) 30:619–27. doi: 10.1016/j.nut.2013.10.011
90. Downer MK, Batista JL, Mucci LA, Stampfer MJ, Epstein MM, Håkansson N, et al. Dairy Intake in Relation to Prostate Cancer Survival. Int J cancer (2017) 140:2060–9. doi: 10.1002/ijc.30642
91. Park Y, Leitzmann MF, Subar AF, Hollenbeck A, Schatzkin A. Dairy Food, Calcium, and Risk of Cancer in the NIH-AARP Diet and Health Study. Arch Intern Med (2009) 169:391–401. doi: 10.1001/archintermed.2008.578
92. Mahmud SM, Franco EL, Turner D, Platt RW, Beck P, Skarsgard D, et al. Use of Non-Steroidal Anti-Inflammatory Drugs and Prostate Cancer Risk: A Population-Based Nested Case-Control Study. PloS One (2011) 6:e16412. doi: 10.1371/journal.pone.0016412
93. Kimura I, Ozawa K, Inoue D, Imamura T, Kimura K, Maeda T, et al. The Gut Microbiota Suppresses Insulin-Mediated Fat Accumulation via the Short-Chain Fatty Acid Receptor GPR43. Nat Commun (2013) 4:1829. doi: 10.1038/ncomms2852
94. Kimura I, Inoue D, Maeda T, Hara T, Ichimura A, Miyauchi S, et al. Short-Chain Fatty Acids and Ketones Directly Regulate Sympathetic Nervous System via G Protein-Coupled Receptor 41 (GPR41). Proc Natl Acad Sci U S A. (2011) 108:8030–5. doi: 10.1073/pnas.1016088108
95. Pernigoni N, Zagato E, Calcinotto A, Troiani M, Mestre RP, Calì B, et al. Commensal Bacteria Promote Endocrine Resistance in Prostate Cancer Through Androgen Biosynthesis. Science (2021) 374:216–24. doi: 10.1126/science.abf8403
Keywords: prostate cancer, IGF-1, short-chain fatty acids, gut microbiome, bacteria
Citation: Matsushita M, Fujita K, Hatano K, De Velasco MA, Uemura H and Nonomura N (2022) Connecting the Dots Between the Gut–IGF-1–Prostate Axis: A Role of IGF-1 in Prostate Carcinogenesis. Front. Endocrinol. 13:852382. doi: 10.3389/fendo.2022.852382
Received: 11 January 2022; Accepted: 21 February 2022;
Published: 15 March 2022.
Edited by:
Conghui Yao, Harvard Medical School, United StatesReviewed by:
Yahui Wang, Washington University in St. Louis, United StatesCopyright © 2022 Matsushita, Fujita, Hatano, De Velasco, Uemura and Nonomura. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kazutoshi Fujita, a2F6dWZ1aml0YTJAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.