
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Endocrinol. , 15 March 2022
Sec. Reproduction
Volume 13 - 2022 | https://doi.org/10.3389/fendo.2022.826920
 Diego Crespo1*
Diego Crespo1* Kai Ove Skaftnesmo1
Kai Ove Skaftnesmo1 Erik Kjærner-Semb1
Erik Kjærner-Semb1 Ozlem Yilmaz2
Ozlem Yilmaz2 Birgitta Norberg2Sara Olausson2Petra Vogelsang1
Birgitta Norberg2Sara Olausson2Petra Vogelsang1 Jan Bogerd3Lene Kleppe1Rolf B. Edvardsen1
Jan Bogerd3Lene Kleppe1Rolf B. Edvardsen1 Eva Andersson1
Eva Andersson1 Anna Wargelius1Tom J. Hansen4
Anna Wargelius1Tom J. Hansen4 Per Gunnar Fjelldal4
Per Gunnar Fjelldal4 Rüdiger W. Schulz1,3
Rüdiger W. Schulz1,3Precocious male maturation causes reduced welfare and increased production costs in Atlantic salmon (Salmo salar) aquaculture. The pituitary produces and releases follicle-stimulating hormone (Fsh), the gonadotropin triggering puberty in male salmonids. However, little is known about how Fsh production is regulated in Atlantic salmon. We examined, in vivo and ex vivo, transcriptional changes of gonadotropin-related genes accompanying the initial steps of testis maturation, in pituitaries of males exposed to photoperiod and temperature conditions promoting maturation (constant light and 16°C). Pituitary fshb, lhb and gnrhr2bba transcripts increased in vivo in maturing males (gonado-somatic index > 0.1%). RNA sequencing (RNAseq) analysis using pituitaries from genetically similar males carrying the same genetic predisposition to mature, but differing by responding or not responding to stimulatory environmental conditions, revealed 144 differentially expressed genes, ~2/3rds being up-regulated in responders, including fshb and other pituitary hormones, steroid-related and other puberty-associated transcripts. Functional enrichment analyses confirmed gene involvement in hormone/steroid production and gonad development. In ex vivo studies, whole pituitaries were exposed to a selection of hormones and growth factors. Gonadotropin-releasing hormone (Gnrh), 17β-estradiol (E2) and 11-ketotestosterone (11-KT) up-regulated gnrhr2bba and lhb, while fshb was up-regulated by Gnrh but down-regulated by 11-KT in pituitaries from immature males. Also pituitaries from maturing males responded to Gnrh and sex steroids by increased gnrhr2bba and lhb transcript levels, but fshb expression remained unchanged. Growth factors (inhibin A, activin A and insulin-like growth factor 1) did not change gnrhr2bba, lhb or fshb transcript levels in pituitaries either from immature or maturing males. Additional pituitary ex vivo studies on candidates identified by RNAseq showed that these transcripts were preferentially regulated by Gnrh and sex steroids, but not by growth factors, and that Gnrh/sex steroids were less effective when incubating pituitaries from maturing males. Our results suggest that a yet to be characterized mechanism up-regulating fshb expression in the salmon pituitary is activated in response to stimulatory environmental conditions prior to morphological signs of testis maturation, and that the transcriptional program associated with this mechanism becomes unresponsive or less responsive to most stimulators ex vivo once males had entered pubertal developmental in vivo.
The pituitary gland is a key organ integrating both intrinsic and extrinsic stimuli to regulate vertebrate physiology, including for example reproduction, growth or stress (1, 2). The anterior pituitary (pars distalis of the adenohypophysis) contains various cell types, together producing a range of peptide hormones. Among those cell types, gonadotropes produce the key regulators of reproduction, the gonadotropins follicle-stimulating hormone (FSH) and luteinizing hormone (LH) (3), which are produced by two distinct gonadotrope cell types in teleost fish (4). Gonadotropins are released into the bloodstream and regulate gonadal hormone and germ cell production (4). Extensive research carried out in numerous fish species has shown the complexity (5) but also conserved aspects of some of the main signaling systems regulating expression, synthesis and secretion of pituitary gonadotropins in teleosts, such as hypothalamic gonadotropin releasing-hormone (Gnrh) (6–9), sex steroids (7, 10, 11) and the activin/inhibin system (7, 12). Notably, androgenic and estrogenic sex steroids exert complex feedback effects on the teleost pituitary, depending among others on gender, stage of maturation, or species examined [as reviewed by Fontaine et al. (13)]. In both Atlantic salmon (Salmo salar) (14) and coho salmon (Oncorhynchus kisutch) (11), in vivo treatment with testosterone (T) −and with 17β-estradiol (E2) in the latter species− stimulated the pituitary expression of lhb but not of fshb. Similarly, in coho salmon, pituitary protein levels of Fsh remained unaffected, whereas Lh levels increased in response to T and E2 (11). Further studies using gonad-intact and gonadectomized Atlantic salmon parr showed that plasma Fsh levels decreased after implanting aromatizable and non-aromatizable androgens (15), demonstrating an important role of androgens in mediating negative feedback on Fsh release.
In Atlantic salmon, pituitary fshb expression and plasma 11-ketotestosterone (11-KT) levels both increase when males enter puberty, which is accompanied by an increase in single germ and Sertoli cell proliferation activity in the testis (14, 16). Fsh and androgen plasma levels remain elevated throughout the testicular growth period in spring/early summer (17, 18), probably reflecting the potent steroidogenic activity of Fsh (19–23). After all, Lh cannot be detected in plasma samples in salmonids until close to the spawning season in the fall, when spermatogenesis has been completed and the spermatogenic tubules are filled with mature sperm. This scenario argues for a pivotal role for Fsh regarding the regulation of entering puberty and sustaining spermatogenesis during the prolonged testicular growth period in salmonid fish. Yet, we are far from having a clear picture of the regulation of Fsh production and release in teleosts, particularly when considering the question how concomitantly increasing/elevated Fsh and androgen levels are possible in the light of a negative androgen feedback on Fsh in salmonids (10, 11).
Farmed Atlantic salmon males exposed to growth-promoting light and temperature regimes can enter precocious maturation, which negatively impacts fish welfare, growth rates and their immune status in turn causing important production losses (24). In order to gain knowledge on how to avoid entrance into precocious puberty, we investigated transcriptomic changes in the pituitary entering puberty. As a first step, we confirmed a prominent pituitary expression of fshb over lhb already prior to and even clearer after entering puberty. We also found that exposing fish to stimulatory photoperiod conditions increased the proportion of males entering puberty, which always was associated with elevated fshb mRNA and circulating androgen levels. Secondly, with two types of experiments we elucidated changes in the pituitary transcriptome accompanying the initiation of puberty in males, namely by (i) an RNAseq study comparing pituitaries from males about to enter puberty with immature counterparts with the aim to identify puberty-associated signals; and (ii) primary tissue culture studies using pituitaries obtained from immature or maturing males that were exposed ex vivo to known regulators of pituitary gene expression, such as Gnrh, sex steroids or growth factors, followed by quantifying a selection of candidate gene transcripts. The latter experiments also allowed to address the question, how a concomitant increase in Fsh and androgen plasma levels may be possible.
Atlantic salmon postsmolts used in this study were reared under standard conditions and sampled at Matre Aquaculture Research Station (Matredal, Norway). All experiments herein have been approved by the Norwegian Animal Research Authority (NARA, permit number 5741); use of the experimental animals was in accordance with the Norwegian Animal Welfare Act of 19th of June 2009. Four groups of fish were studied:
Group 1. After smoltification, immature males (14-15 months old) were exposed to 12 hours dark/12 hours light and 16°C (referred to as non-stimulatory conditions) for a period of 16 days. Feeding was done with standard commercial diets. Prior to sampling for body weight, length and gonad weight, all fish were anesthetized with 2 mL/L Finquel vet and sacrificed by cutting into the medulla oblongata. The condition factor (K) was calculated as K = body weight × body length−3 × 100. Pituitaries from the sampled fish were cultured ex vivo (see Pituitary Tissue Incubations sections) and collected after 9 days of incubation in RNAlater (Thermo Fisher Scientific) for RNA extraction. Additional pituitaries were fixed in 4% glutaraldehyde overnight at 4°C and embedded in plastic (Technovit 7100; Kulzer) for subsequent histological analysis, as previously described (25, 26).
Group 2. Immature postsmolt males (14-15 months old) were exposed to continuous light and 16°C (referred to as stimulatory conditions) for 16 days, as previously described by Fjelldal et al. (27). In response to this regime, part of the fish had started pubertal development, as indicated by gonado-somatic index (GSI) levels above 0.1% and the presence of type B spermatogonia, respectively [the germ cells/cysts were identified according to previously published morphological criteria (28)]. Feeding and sampling were performed as mentioned above for Group 1 with minor changes. Briefly, pituitaries from immature and maturing postsmolts were collected before or after ex vivo incubation for 9 days and stored in RNAlater until RNA extraction. Testis tissue was fixed in 4% glutaraldehyde overnight at 4°C and embedded in Technovit 7100 for histological evaluation or collected in RNAlater for RNA extraction. Moreover, plasma samples were collected for 11-KT quantification.
Group 3. An additional batch of (14-15 months old) immature postsmolt males was exposed to stimulatory conditions for 7 weeks. Feeding, sampling and pituitary incubations were performed as described above for Group 2. No testis tissue samples were collected in this experiment.
Group 4. All-male siblings, heterozygous for the puberty-associated vgll3 locus (29, 30), were produced by crossing a double haploid XX female with a YY supermale (31). At the age of 14-15 months, the fish were exposed to stimulatory conditions for 6 months. At 7 and 11 days after starting the maturation regime, the fish were sampled for body weight, length, gonad weight and plasma. Pituitaries were collected and flash frozen in liquid nitrogen for high-throughput RNA sequencing analyses. A testis tissue sample was fixed in 4% glutaraldehyde, embedded in Technovit 7100 and sectioned at 3 µm thickness, and analysis of the stage of maturation showed that all males in Group 4 exhibited testes containing type A spermatogonia as the furthest developed germ cell type, i.e. were classified as immature regarding germ cell development (28). Males were selected for sequencing analysis based on 11-KT plasma levels and on fshb transcript levels in the pituitary.
A previously established primary tissue culture system developed for zebrafish (Danio rerio) (32) was used, which is also similar to the system developed for Japanese eel (Anguilla japonica) pituitaries (33), except that we used an agar cylinder instead of elder pith as support for pituitary tissue. For salmon pituitary incubations, the temperature was set at 14-16°C. First, in order to study the morphological and functional integrity of pituitary tissue after ex vivo culture, whole pituitaries collected from immature postsmolt males were incubated in L15-based medium for 3 and 9 days and subsequently fixed in 4% glutaraldehyde for histological analysis. Additional pituitaries were incubated in the absence or presence of E2 and collected in RNAlater for gene expression analyses.
To study the effects of potential regulators on gene expression ex vivo, pituitaries from immature and maturing postsmolts were incubated for 9 days in the absence or presence of different compounds (Gnrh, E2, 11-KT, inhibin A, activin A and Igf1). After the incubation period, pituitaries were collected for gene expression analyses. E2 and 11-KT were purchased from Sigma-Aldrich and used at a final concentration of 100 ng/mL (33) and 100 nM (34), respectively. Gnrh salmon analog [1 µM (8)] was purchased from Syndel. Recombinant human inhibin A and activin A proteins were purchased from Thermo Fisher Scientific and used at the same final concentration of 50 ng/mL (35, 36); the respective salmon proteins were not available commercially. Also salmon Igf1 was not available, so that we used recombinant gilthead sea bream Igf1 (37) (50 ng/mL) from ProSpec.
Two types of single-chain recombinant hormones were produced in CHO cells after transfection of the respective coding sequences in pcDNA3.1(+) vector (Invitrogen) by Rara-Avis (Valencia, Spain): (i) single-chain recombinant salmon Fsh (referred to as rFsh-A) consisting of Atlantic salmon Fsh beta (Fshb) subunit (acc. no. XM_014126338), a (Gly-Ser) spacer sequence and rabbit common alpha (CGA) subunit (acc. no. AF318299), and (ii) single-chain recombinant salmon Fsh (referred to as rFsh-B) consisting of Atlantic salmon Fshb subunit (acc. no. XM_014126338), a (Gly-Ser) spacer sequence and Atlantic salmon common alpha (Cga) subunit (acc. no. BT056856). The percentage identity between amino acid sequences of rabbit CGA and Atlantic salmon Cga is 70%. Yields of rFsh-A and rFsh-B, after IMAC purification, were 1.37 mg per 8x108 CHO cells and 1.89 mg protein per 4x108 CHO cells, respectively. Both recombinant hormones contained a TEV protease cleavage site followed by a 6X His-Tag (used for affinity purification) at the C-terminus of the protein. Antiserum was raised in rabbits against the rFsh-A protein (Agrisera AB, Vännäs, Sweden). The specificity of the Fsh antiserum was confirmed by Western blotting and immunohistochemistry (see below).
Samples containing 0.25 µg of the single-chain rFsh-B were diluted in lithium dodecyl sulfate sample buffer (NuPAGE LDS Sample Buffer, Invitrogen) with 100 mM DTT (reducing conditions). Pituitaries from 3 maturing male salmon (GSI = 0.78 ± 0.05%) were mixed and homogenized in lysis buffer (10 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.05% Tween 20) containing 1:100 protease inhibitor cocktail (Merck) with 104 mM AEBSF, 80 μM aprotinin, 4 mM bestatin, 1.4 mM E-64, 2 mM leupeptin and 1.5 mM pepstatin. The homogenate was incubated on ice for 10 min before being centrifuged at 14000 g for 10 min at 4°C. All samples were denatured for 10 min at 70°C and separated on Any kD Mini-PROTEAN TGX gels (Bio-Rad) at 120 V for 1 hour. A gel replicate for Coomassie blue staining was fixed in 40% ethanol/10% acetic acid for 15 min, stained overnight at room temperature with QC Colloidal Coomassie Stain (Bio-Rad) and de-stained in deionized water for 3 hours before signal detection. rFsh-B (0.25 µg) and 10 µg pituitary homogenate were separated on gels and then transferred to Trans-Blot Turbo Mini nitrocellulose membranes (Bio-Rad) at 25 V for 7 min on a Trans-Blot Turbo system (Bio-Rad). Blots containing transferred protein were blocked in 1X casein solution (Vector laboratories, Bionordika) for 10 min at room temperature and were then incubated for 1 hour at room temperature with the anti-Fsh serum, anti-Fsh serum preabsorbed with rFsh-B (at 1:28 molar ratio overnight at 4°C) or pre-immune serum, all diluted 1:5000 in PBST with 1% BSA. After washing in PBST, the membranes were incubated with biotinylated goat anti-rabbit IgG (H+L) secondary antibody (1:1000; Life Technologies) at room temperature for 2 hours and signal was developed using VECTASTAIN ABC-AmP reagent (Vector laboratories) and detected using iBright FL1500 imaging system (Invitrogen).
Pituitaries of immature and maturing male salmon (collected from a subset of fish as presented in Figures 1C–F) were fixed in 4% paraformaldehyde, embedded in paraffin, and serial 4 μm thick sagittal sections were prepared for fluorescent immunohistochemistry. Briefly, slides were heated using a 2100 Retriever (Electron Microscopy Sciences) for 20 min in sodium citrate buffer pH 6.0 for antigen retrieval and subsequently cooled down at room temperature for 2 hours. After washing in PBS and blocking with 5% BSA (in PBS), consecutive slides were incubated at 4°C overnight with polyclonal rabbit anti-(Atlantic salmon)Fsh (Agrisera; this study) and rabbit anti-(coho salmon)Lh antibodies (kindly provided by Dr. Penny Swanson), diluted 1:5000 in 1% BSA/PBS. Then, slides were incubated with the secondary antibody (mouse anti-rabbit IgG Alexa Fluor 488; Life Technologies) at room temperature for 90 min. Propidium iodide (Sigma-Aldrich) was used as nuclear counterstain and slides were mounted with SlowFade Diamond Antifade Mountant (Thermo Fisher Scientific). Pituitary sections were analyzed using a standard fluorescence microscope (Nikon Eclipse 80i).
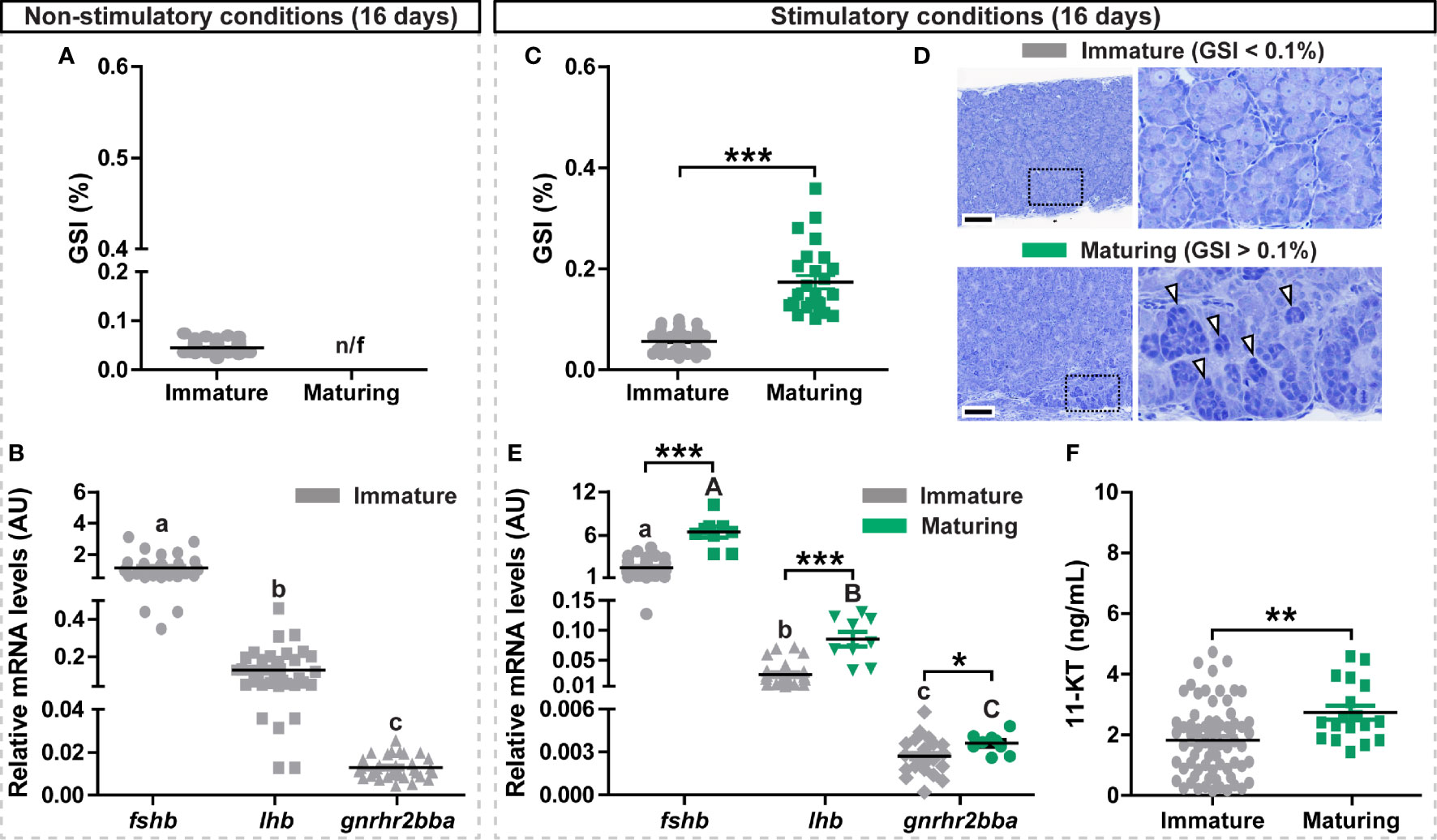
Figure 1 Evaluation of gonadal and pituitary responses to non-stimulatory and stimulatory conditions. (A, B) Gonado-somatic index (GSI; A) and in vivo expression levels of selected pituitary genes (fshb, lhb, gnrhr2bba; B) in immature postsmolt males exposed to non-stimulatory conditions (12 hours dark/12 hours light, and 16°C) for 16 days. (C–F) GSI (C), representative testis histology (D), fshb, lhb, gnrhr2bba expression levels (E) and 11-ketotestosterone (11-KT) plasma levels (F) in immature and maturing males after exposure to stimulatory conditions (constant light and 16°C) for 16 days. In (A, C, F), data are shown as mean ± SEM (N = 18-96; **p < 0.01; ***p < 0.001) and, in (B, E), shown as mean ± SEM (N = 9-35; *p < 0.05; ***p < 0.001; different letters denote significant differences between groups) and expressed relative to the fshb mRNA abundance. n/f, not found. AU, arbitrary units. In (D), boxes identify the testis tissue areas shown at higher magnification, white arrowheads indicate representative groups of type B spermatogonia. Scale bar, 100 µm.
Digital images from fluorescent stained sections were subjected to a quantification pipeline. First, a pixel classification of the nuclei was performed with Ilastik (38) to separate nuclei from background. Using the probability maps produced in Ilastik, nuclei were then segmented in CellProfiler (39–41) and propagation outward from the nuclei was used to locate the cell boundaries. Cell cytoplasm was located as the cell boundary minus the nuclei boundary, and any positive signal inside the cytoplasm was measured by the ‘‘MeasureObjectIntensity’’ and ‘‘MeasureGranularity’’ modules in CellProfiler. Positive cells were then scored by filtering on median intensity and granularity of the staining. Four non-overlapping fields of the proximal pars distalis per individual and antibody (anti-Fsh or anti-Lh) were photographed at 400X magnification in serial pituitary sections, and the percentage of Fsh- and Lh-positive cells quantified. Therefore, the total area examined for fluorescent signal was the same for both groups.
Additional pituitary sections were examined by chromogenic staining of Fsh protein using EnVision+ Dual Link System-HRP (DAKO), according to the manufacturer’s protocol. Briefly, endogenous peroxidase activity was blocked by incubation with 0.3% H2O2 in PBS for 30 min. After incubation with the anti-Fsh serum and washing in PBST, slides were incubated with the polymer-HRP secondary antibody at room temperature for 30 min, after which slides were incubated with DAB substrate kit (VECTASTAIN, Vector laboratories) for 5 min. Nuclei were counterstained with hematoxylin and slides mounted with Histokitt (Chemi-Teknik) after dehydration.
Plasma concentrations of 11-KT were analyzed by ELISA (42) on extracted plasma samples, as previously described (43). Acetylcholine esterase-labeled tracers and microplates pre-coated with monoclonal mouse anti-rabbit IgG were supplied by Cayman Chemicals. 11-KT standard was purchased from Sigma-Aldrich. Anti-11-KT was a kind gift from David E. Kime (Sheffield University, UK).
Each pituitary was collected on RNAlater for preservation and stored at 4°C until use. Individual pituitaries were homogenized in 400 µL of homogenization buffer and processed according to the Maxwell HT-simplyRNA kit instructions (Promega) on a BioMek 4000 instrument (Beckton Dickinson), and RNA was DNase treated as part of the RNA extraction procedure. The quantity and purity of RNA samples were assessed by spectrophotometry on a Nanodrop ND-1000 instrument (Thermo Fisher Scientific). cDNA was prepared by reverse transcription of 200 ng RNA using the SuperScript IV VILO Master Mix with ezDNase Enzyme (Thermo Fisher Scientific) according to the manufacturer’s recommendations.
Except for qPCR assays previously published, primers specific for the genes of interest were designed using the BatchPrimer3 online tool (https://probes.pw.usda.gov/batchprimer3/). Primers are listed with their respective sequence and citation in Supplementary Table 1. A qPCR reaction was prepared according to the manufacturer descriptions to contain 800 nM of each forward and reverse primer in a 6 µL reaction containing a 1x concentration of the PowerUp SYBR Green Master Mix, or TaqMan Fast Advanced Master Mix where a TaqMan probe was required (Thermo Fisher Scientific). 2 µL of a 1/20 dilution of cDNA was added to the reaction and all qPCR assays were analyzed using a QuantStudio 5 Real-Time PCR system (Thermo Fisher Scientific). The relative gene expression level was calculated using the comparative Ct (2−ΔΔCt) method (44). For each gene, all values were normalized to ef1a expression.
We used immature sibling salmon males for the RNAseq study, which showed a low genetic variation since they were produced by crossing a double haploid XX female with an inbreed YY supermale, created by self-fertilization from a hermaphrodite (31). These immature fish, heterozygous for the puberty-associated vgll3 locus, were exposed to constant light and 16°C. Part of the population did (responders), another part did not (non-responders), react to these stimulatory environmental conditions by showing signs for entering puberty (see Results section below). Pituitaries from responders and from non-responders were used to examine transcriptomic differences. Total RNA was isolated from each pituitary using the Maxwell HT-simplyRNA kit (Promega) and quantified, as described above. RNA integrity was checked with an Agilent Bio-analyzer 2100 total RNA Nano series II chip (Agilent). Only samples with an RNA integrity number > 8 were used for library preparation. 200 ng RNA of each pituitary sample were reverse transcribed and the synthesized cDNAs were then used for fshb screening by qPCR, as previously described. Illumina RNAseq libraries were prepared from the remaining 1-2 µg total RNA using the Illumina TruSeq Stranded mRNA Library Prep Kit (Illumina, Inc.) according to the manufacturer’s instructions. The resulting RNAseq libraries were sequenced on an Illumina HiSeq 4000 sequencer (Illumina, Inc.) as 150 bp pair-end reads. Base calling was done by the Illumina pipeline. Quality control of the obtained reads was performed using FastQC suite (v0.11.7; default parameters). RNAseq derived reads were mapped to the salmon genome (ICSASG_v2) using Bowtie2 v2.3.5.1 (45). The resulting files were filtered with SAMtools v1.10 (46), and the read counts for each transcript were extracted using SAMtools idxstats and summed for each unique GeneID. Data analysis was performed with the R/Bioconductor package DESeq (47) (p-adjusted < 0.001). Genes were discarded if all samples had normalized read counts less than 50. The raw RNAseq data of the 10 samples sequenced (5 biological replicates per condition) have been deposited in the NCBI BioProject database with accession number PRJNA778619.
Functional enrichment analysis was carried out using the WebGestalt online tool (48) (http://www.webgestalt.org/), which calculates over-representation (FDR < 0.05) of Gene Ontology (GO) categories (49). The g:Orth option of the g:Profiler tool (50) (http://biit.cs.ut.ee/gprofiler/gost) was used to retrieve mouse (Mus musculus) orthologs from the list of differentially expressed genes (DEGs) and exported as the input for functional enrichment analysis (Biological Process GO category). Regulated KEGG pathways were determined using the KEGG Mapper tool (51). KEGG pathways represented by at least 3 DEGs and by the ratios of regulated genes (up-/down-, and vice versa) higher than 2.5 were considered for the analysis.
GraphPad Prism 9.0.0 package (GraphPad Software, Inc.) was used for statistical analysis. Significant differences between groups were identified using Student’s t test or one-way ANOVA followed by Tukey’s test for multiple group comparisons, as appropriate (*, p < 0.05, **, p < 0.01, ***, p < 0.001; ns, no significant changes observed). For datasets with no normal distribution, the non-parametric two-sided Mann–Whitney test was applied. The correlations between GSI values and fshb, lhb and gnrhr2bba expression levels, and 11-KT plasma concentration, were analyzed using Spearman’s rank test (data were Log2-transformed to meet homogeneity of variances). Results are represented as mean ± SEM.
To investigate the transcriptional changes of selected gonadotropin-related genes (fshb, lhb and gnrhr2bba) accompanying initial steps of testis maturation, we exposed immature postsmolt males to both non-stimulatory and stimulatory conditions. While exposure to non-stimulatory conditions (12 hours dark/12 hours light and 16°C, for a period of 16 days) did not trigger testis growth (GSI < 0.1%; Figure 1A), an increasing proportion of the fish entered pubertal development from day 10 when kept under stimulatory conditions (24 hours light and 16°C, for a period of 16 days) (Supplementary Figure 1E), as indicated by GSI levels above 0.1% and the presence of type B spermatogonia (Figures 1C, D). Condition factor and body weight, but not body length, were also increased in maturing males exposed to stimulatory conditions (Supplementary Figures 1C, D). Pituitary fshb expression levels were at least 10-fold higher than lhb, and at least 100-fold higher than gnrhr2bba levels in immature fish of both photoperiod regimes (Figures 1B, E). fshb, lhb and gnrhr2bba transcripts all increased in the maturing fish exposed to stimulatory conditions (Figure 1E), and all transcripts showed significant correlations with the GSI values (Supplementary Figures 2B, E, H). Remarkably, despite already showing high expression levels, the amplitude of the response to the stimulatory conditions was highest for fshb (4.25-fold), compared to lhb (2.56-fold) and gnrhr2bba (1.32-fold). A significant increase in plasma 11-KT was also observed in maturing fish (Figure 1F), and its levels correlated to GSI and fshb expression values (Supplementary Figure 3).
Considering the prominent response of fshb transcript levels upon entering puberty, we aimed at investigating the cellular expression of Fshb protein in Atlantic salmon pituitaries. For that purpose, a chimeric single-chain Fsh peptide (rFsh-A; containing the mature Atlantic salmon Fshb subunit, a spacer sequence and rabbit CGA common alpha subunit) was produced, and an Fsh antiserum was then raised in rabbits against the rFsh-A protein.
The Fsh antiserum was characterized by Western blotting, using pituitary extracts of maturing male salmon (GSI = 0.78 ± 0.05%) and the single-chain rFsh-B protein (Atlantic salmon Fshb subunit, a Gly-Ser spacer sequence and Atlantic salmon Cga common alpha subunit) as samples for electrophoresis. Coomassie blue protein staining (Figure 2A) and immunoblotting analysis (Figure 2B) indicated that the Fsh antiserum recognized the rFsh-B protein with molecular mass of approximately ~35 kDa. This corresponds well with the molecular weight determined by detecting His-Tags in the rFsh-B protein (Supplementary Figure 4A). Minor bands of approximately 18, 70, 90 and 125 kDa were also visible (left panel in Figure 2B). Additional bands on the Western blot could represent strongly glycosylated proteins, or degradation of the protein. None of the bands were observed using pre-immune serum (right panel in Figure 2B). A clear band of approximately 18 kDa was detected in a pituitary extract of maturing males, corresponding to the molecular weight of Atlantic salmon Fshb subunit (Figure 2C, left panel). This band disappeared when preabsorbing the Fsh antiserum with rFsh-B (Figure 2C, right panel). Two higher molecular weight bands (~75 kDa and ~150 kDa) were found (Figure 2C) when using Fsh antiserum, but also with pre-immune serum and another, unrelated antiserum raised in rabbit (data not shown), suggesting they represent non-specific staining not related to Fsh.
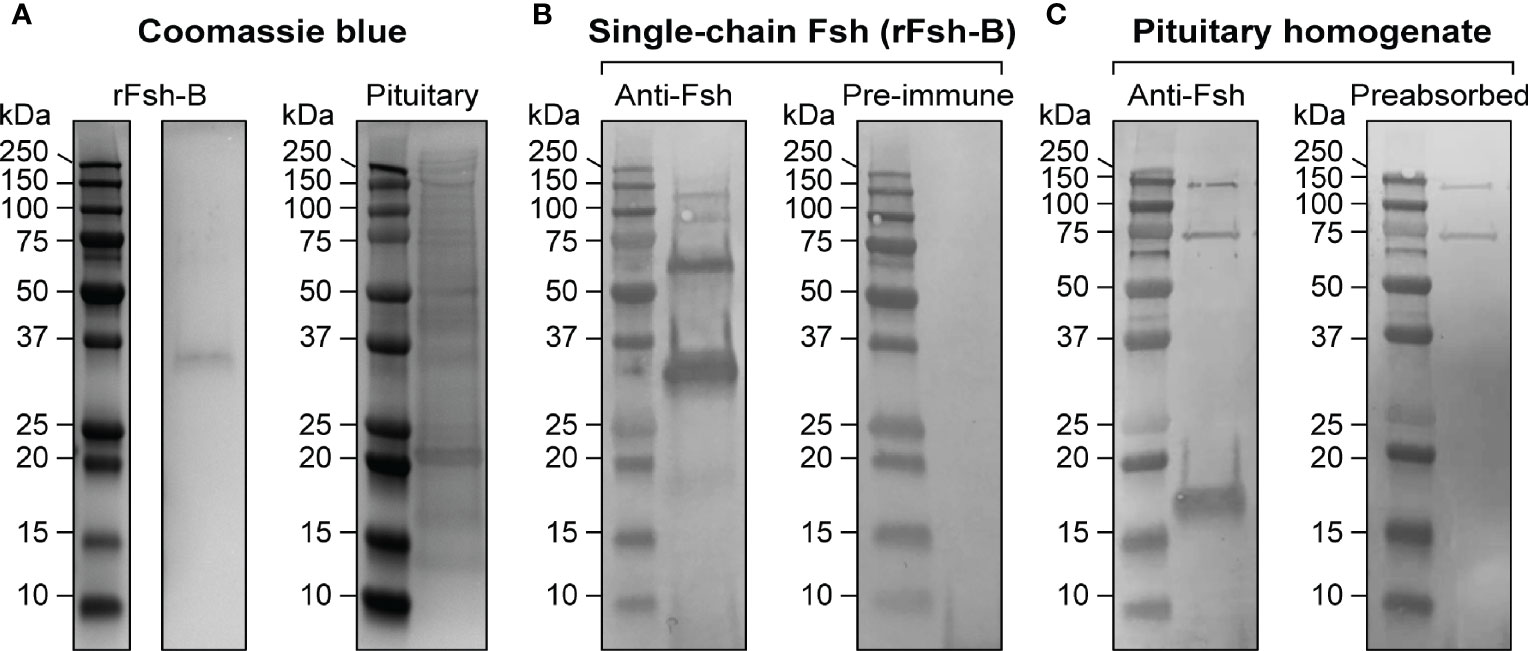
Figure 2 Characterization of a specific antiserum for Atlantic salmon Fsh. (A) Coomassie blue protein staining of the single-chain Fsh protein (rFsh-B) and of a pituitary homogenate. (B) Representative immunoblots of rFsh-B using Fsh antiserum (left panel) and pre-immune serum (as negative control; right panel). (C) Representative immunoblots of a pituitary homogenate using anti-Fsh serum (left panel) and anti-Fsh serum preabsorbed with rFsh-B (as negative control; right panel). In all blots, molecular mass markers (kDa) are shown on the left.
The Fsh antiserum was then used for immunohistochemistry on serial sections from pituitaries of immature and maturing male salmon after 16 days exposure to stimulatory conditions (collected from a subset of fish as presented in Figures 1C–F). In immature pituitaries, the antiserum labeled some but not many Fsh-containing pituitary cells located in the proximal pars distalis (PPD; Figure 3A). However, the number of Fsh-positive cells and the signal intensity was remarkably higher in pituitaries of maturing males, where clearly Fsh-positive cells were widely distributed in the PPD region (magnified areas from the marked yellow and red dashed lines; Figure 3C). No signal was observed in control sections incubated with pre-immune serum (right lower insets in Figures 3A, C; see also Supplementary Figure 4B), indicating that the non-specific staining detected by Western blotting did not result in non-specific staining patterns on tissue sections. Moreover, using an anti-(coho salmon)Lh antibody previously generated and validated (18), very few or no (in 3 out of 5 males) Lh-positive cells were identified in immature Atlantic salmon pituitaries (Figure 3A). Despite observing an increase in their incidence during maturation, the number (and signal intensity) of Lh cells was clearly lower compared to those observed for Fsh cells, in both immature and maturing pituitaries (Figures 3A, C). Quantification of the percentage of Fsh- and Lh-positive cells supported the microscopical evaluation of pituitary sections, since the incidence of Fsh cells was significantly higher, irrespective of the stage of maturation (Figures 3E, F). Furthermore, the percentage of Fsh cells, in contrast to Lh cells, increased during maturation (Figure 3G). Considering that the Fsh and Lh antisera were both raised in rabbits, we were not able to carry out co-localization studies for Fsh/Lh protein expression on a single section. While single immuno-detection of Fsh and Lh on consecutive sections did not provide evidence for co-localization, it also does not allow excluding cellular co-localization with certainty (Figures 3B, D).
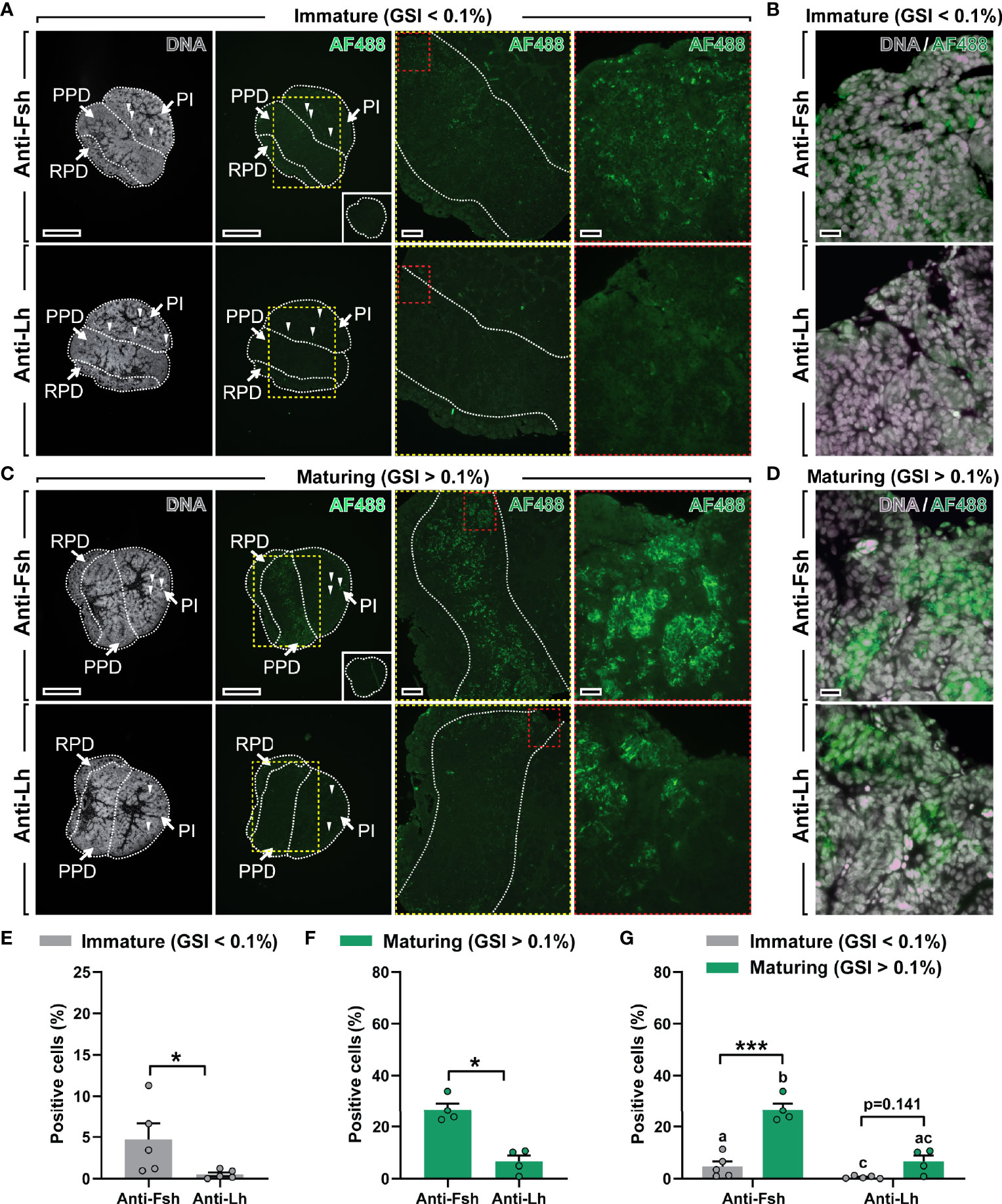
Figure 3 Localization and quantification of Fsh and Lh cells in immature and maturing male salmon pituitaries. (A–D) Immunolocalization of Fsh and Lh proteins in serial sections (sagittally oriented; anterior to the left) of immature and maturing salmon pituitaries using the Atlantic salmon Fsh antiserum generated in this study and an anti-(coho salmon)Lh antibody previously validated (18). Right panels in (A) and C show pituitary tissue magnified from the marked areas (yellow and red dashed lines; scale bar = 200 µm and 50 µm, respectively). Scale bar in low magnification pictures = 500 µm. Negative control (pre-immune serum) for Fsh immunostaining showed no specific staining (insets in right lower panels in (A, C). Arrowheads indicate red blood autofluorescence. Further magnified areas of pituitary tissue of immature (B) and maturing (D) male salmon do not seem to show clear Fsh/Lh co-localization in serial sections (scale bar = 20 µm). Propidium iodide (in grey) was used as nuclear counterstain. GSI, gonado-somatic index; AF488, Alexa Fluor 488; PPD, proximal pars distalis; PI, pars intermedia; RPD, rostral pars distalis. (E–G) Quantification of the percentage of Fsh- and Lh-positive cells in immature (E) and maturing (F) pituitaries, and combined (G). Data are shown as mean ± SEM (N = 4-5; *p < 0.05; ***p < 0.001), and expressed relative to the total number of cells. In (G), different letters indicate significant differences between groups.
Pituitaries from sibling salmon males with low genetic variation (31) and sharing the same genetic predisposition to mature (29, 30), were analyzed by RNAseq to investigate global gene expression of pituitary tissue upon entering puberty. To this end, juvenile males were exposed to stimulatory photoperiod and water temperature conditions (constant light and 16°C, as described above) for 11 days and samples were collected after 7 and 11 days. The pituitaries were assigned to two groups, high or low, depending on the fshb transcript levels (Figure 4A), which also correlated with high or low 11-KT plasma levels (Figure 4B). We refer to these two groups as responders and non-responders, respectively. It is important to note that neither GSI (Figure 4C) nor testicular histology (spermatogonia type A were the furthest developed germ cell type in both groups; data not shown) differed between the two groups.
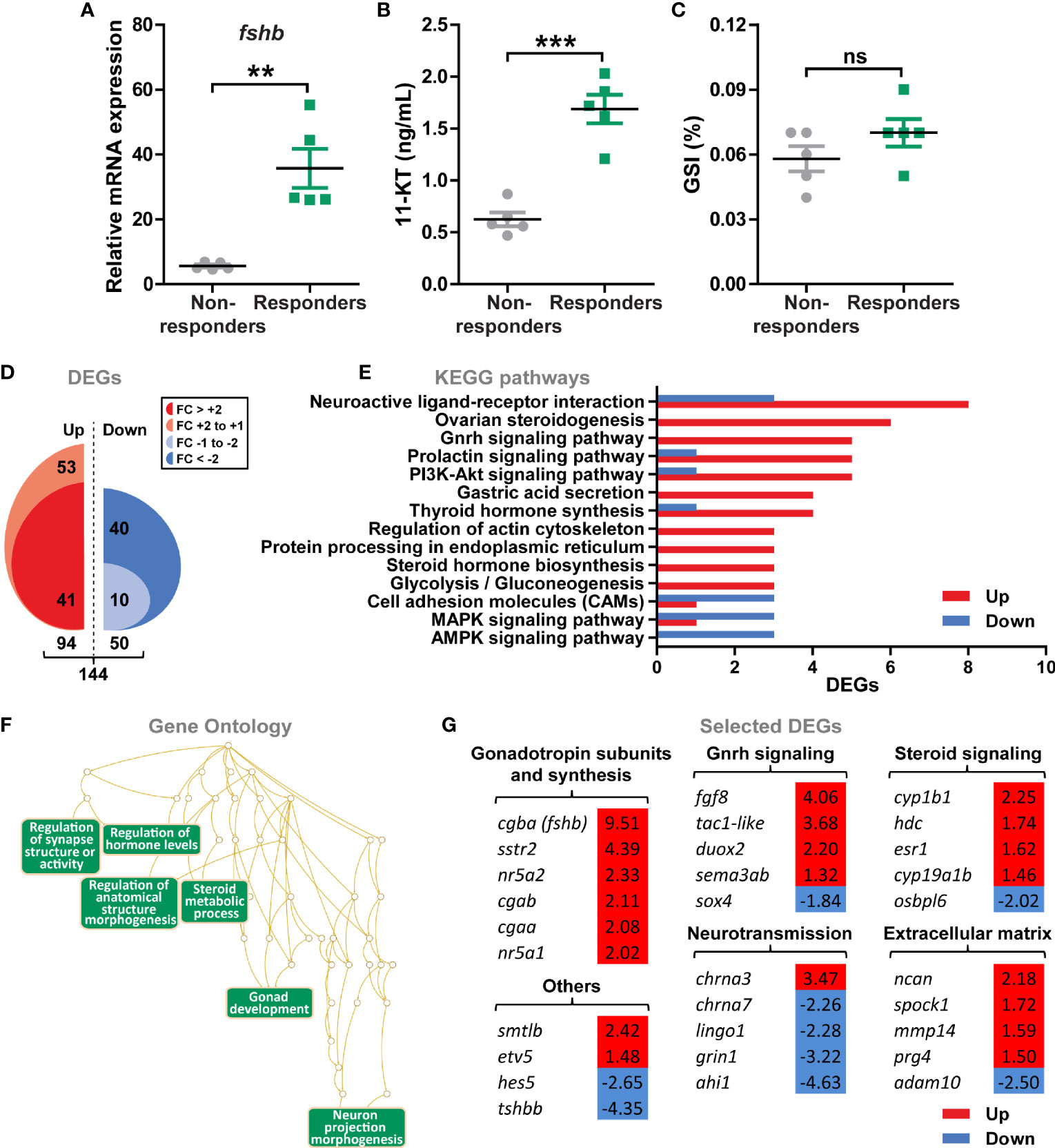
Figure 4 Gene expression profiling of pituitaries from male postsmolts about to enter puberty. (A–C) To select pituitary samples for RNAseq, sibling salmon males with low genetical variance were sampled at day 7 and 11 after exposure to stimulatory conditions (constant light and 16°C) and different puberty-associated parameters evaluated. Analysis of fshb expression (A) and 11-ketotestosterone [11-KT; (B)] levels revealed two groups of fish showing low (non-responders) or high (responders) levels for both parameters, while gonado-somatic indices (GSI; C) were unaffected. Data are shown as mean ± SEM (N = 5; **p < 0.01; ***p < 0.001). ns, no significant differences between groups. (D) Total numbers of up- and down-regulated genes (DEGs) identified by RNAseq (N = 5; p-adjusted < 0.001). (E, F) Regulated KEGG pathways (E) and Gene Ontology (F) terms in pituitaries of males responding to stimulatory conditions. KEGG pathways represented by at least 3 DEGs and ratio of regulated genes higher than 2.5 were considered for the analysis. (G) Selected DEGs identified by KEGG and GO analyses grouped by their function. Fold change values are shown with a red or blue background indicating up- or down-regulation, respectively.
144 genes were differentially expressed (DEGs) in pituitaries from responders versus non-responders (Figure 4D). Two thirds of the DEGs increased (94 or 65%), one third decreased (50 or 35%) in pituitary tissue from responding males, while the proportion of DEGs that reached a more than 2-fold change in expression was higher for the down- versus the up-regulated genes (80% versus 44%; Figure 4D and Supplementary Data 1). The majority of KEGG terms significantly enriched in pituitaries from responders contained genes that were up-regulated (Figure 4E), including pathways related to Gnrh, prolactin and PI3K-Akt signaling, steroid and thyroid hormone biosynthesis, as well as others involved in metabolic processes (e.g. glycolysis/gluconeogenesis) and neurotransmission (e.g. neuroactive ligand-receptor interaction). To further investigate characteristics of the pituitary transcriptome of males responding to stimulatory conditions, Gene Ontology terms were retrieved. Functionally related gene groups emerging from this analysis included factors involved in hormone (steroid) metabolism, gonad development and neuron morphogenesis (Figure 4F). Among the candidates identified by functional analyses, we observed that transcript levels of all gonadotropin subunits −cgba (fshb), cgaa, cgab− and genes involved in the regulation of gonadotropin synthesis (nr5a1, nr5a2 and sstr2) were increased in pituitaries from responder males (Figure 4G). Also, Gnrh and steroid signaling, as well as extracellular matrix genes were preferentially up-regulated, in particular regarding estrogen signaling (esr1, cyp19a1b and cyp1b1; Figure 4G). On the contrary, a higher number of down-regulated genes was identified in the neurotransmission category (Figure 4G). In addition, the gene set “others” included factors previously associated to puberty and the Notch signaling pathway (etv5 and hes5, respectively), as well as other pituitary hormones (smtlb and tshbb; Figure 4G).
First, we established that pituitary tissue from immature fish showed normal morphology following whole-organ tissue culture for up to 9 days (Supplementary Figures 5A–H). We then examined transcriptional changes of selected gonadotropin-related genes in response to E2 and found that gnrhr2bba and lhb, but not fshb, mRNA levels increased (Supplementary Figure 5I). These results indicate that the tissue culture system preserves structural and functional characteristics of the salmon pituitary, including gonadotroph cells.
By exposing immature fish to stimulatory environmental conditions, we generated sibling groups with individuals that did, or did not, enter puberty, and then used primary pituitary tissue cultures to investigate the effect of Gnrh and sex steroids −the most enriched signaling pathways identified by RNAseq during puberty− on gonadotropin gene expression in pituitaries collected from immature and maturing sibling males. We observed an up-regulation of gnrhr2bba and lhb in response to Gnrh, E2 and 11-KT in pituitaries of immature fish exposed to both non-stimulatory and stimulatory conditions (Figures 5A, B). While Gnrh also increased fshb transcript levels in immature pituitaries ex vivo, they were decreased by 11-KT (Figures 5A, B). As reported for immature fish exposed to non-stimulatory photoperiod conditions (Figure 5A), as well as for fish that remained immature despite having been kept under stimulatory conditions (Figure 5B; grey bars), sex steroids consistently increased lhb and gnrhr2bba transcript levels also when incubating pituitaries from fish that had started puberty (Figure 5B; green bars). In contrast, a Gnrh-dependent modulation in the expression of the three selected pituitary genes was not observed, and fshb transcript levels remained unchanged in response to all compounds tested when incubating pituitaries from maturing males (Figure 5B; green bars).
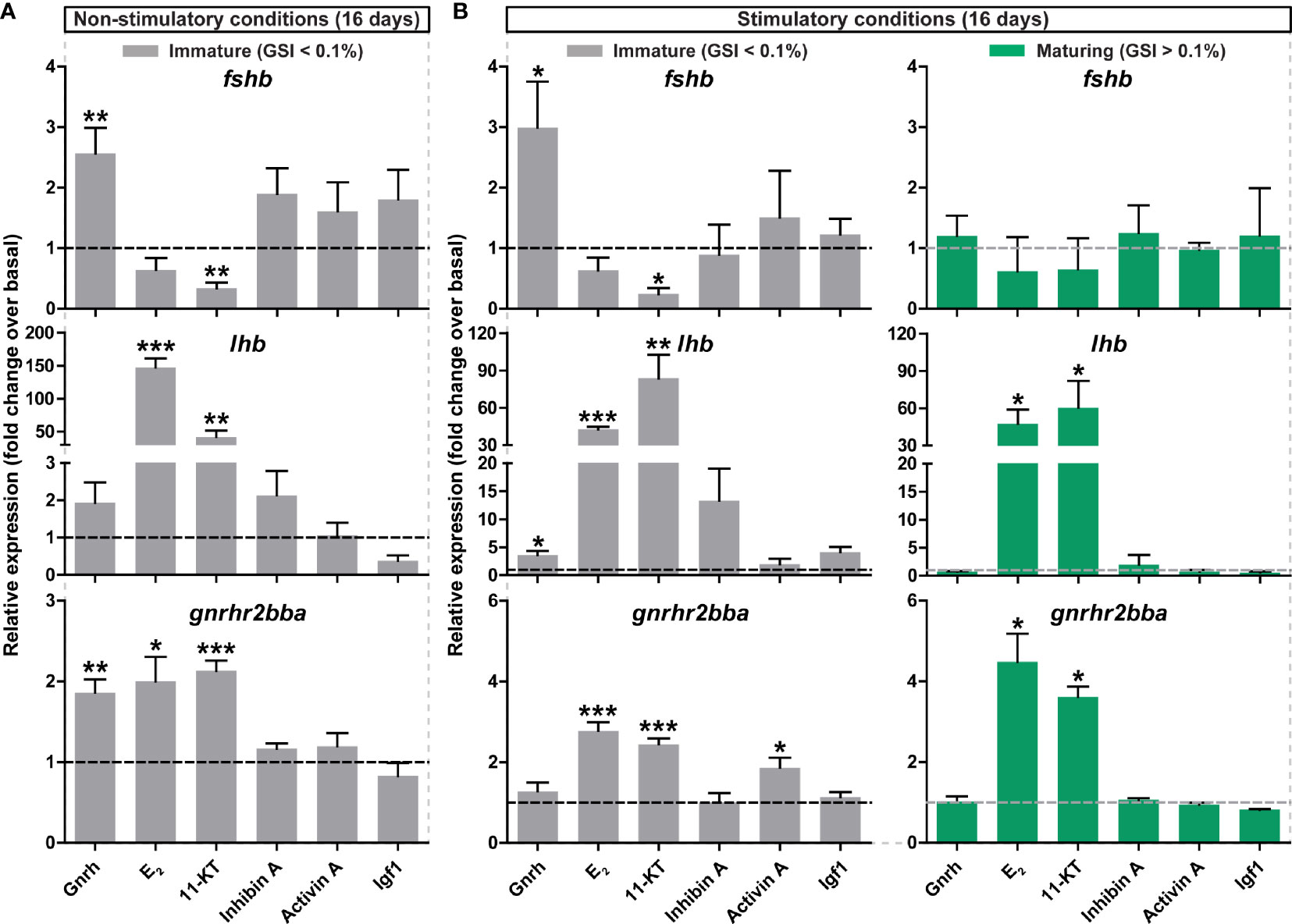
Figure 5 Ex vivo effects of potential regulators on selected pituitary genes. (A, B) Expression levels of fshb, lhb and gnrhr2bba in pituitaries collected from immature postsmolt males exposed to non-stimulatory conditions [12 hours dark/12 hours light, and 16°C; (A)] and immature and maturing postsmolt males exposed to stimulatory conditions [constant light and 16°C; (B)] for 16 days, and subsequently incubated ex vivo for 9 days in the presence of various potential regulators of pituitary gene expression. Results are shown as mean fold change ± SEM (N = 3-6; *p < 0.05; **p < 0.01; ***p < 0.001) and expressed relative to the control basal condition, which is set at 1 (dashed line).
In addition to Gnrh and sex steroids, other potential modulators of gonadotropin gene expression such as inhibin A, activin A and Igf1 were tested ex vivo. However, statistical significance was only reached for gnrhr2bba transcript levels in response to activin A in immature pituitaries of fish exposed to stimulatory conditions (Figure 5B; grey bars).
We then investigated the transcriptional response of candidates identified by pituitary RNAseq upon entry into puberty as differentially expressed, examining the same array of ligands previously tested to evaluate gonadotropin gene expression ex vivo. Selected candidates include: (i) somatolactin (smtlb) and thyrotropin subunit beta (tshbb) pituitary hormones; (ii) genes involved in estrogen signaling (brain aromatase, cyp19a1b; estrogen receptor 1, esr1); and (iii) protachykinin-like (tac1-like). This selection was based on previous studies describing the importance of these factors in the regulation of sexual maturation (see Discussion). In immature fish kept under standard photoperiod conditions, Gnrh (in most cases) and steroids clearly modulated the expression of all candidate genes studied (Figures 6A–D) except of tshbb (data not shown). While transcript levels of cyp19a1b, esr1, tac1-like and smtlb were increased by both E2 and 11-KT, Gnrh up-regulated tac1-like and smtlb and down-regulated cyp19a1b (Figures 6A, D). Interestingly, cyp19a1b was the only candidate gene examined for which Gnrh and steroids induced an opposite effect in expression (Figure 6A). In addition, Igf1 increased the expression of esr1 (Figure 6B).
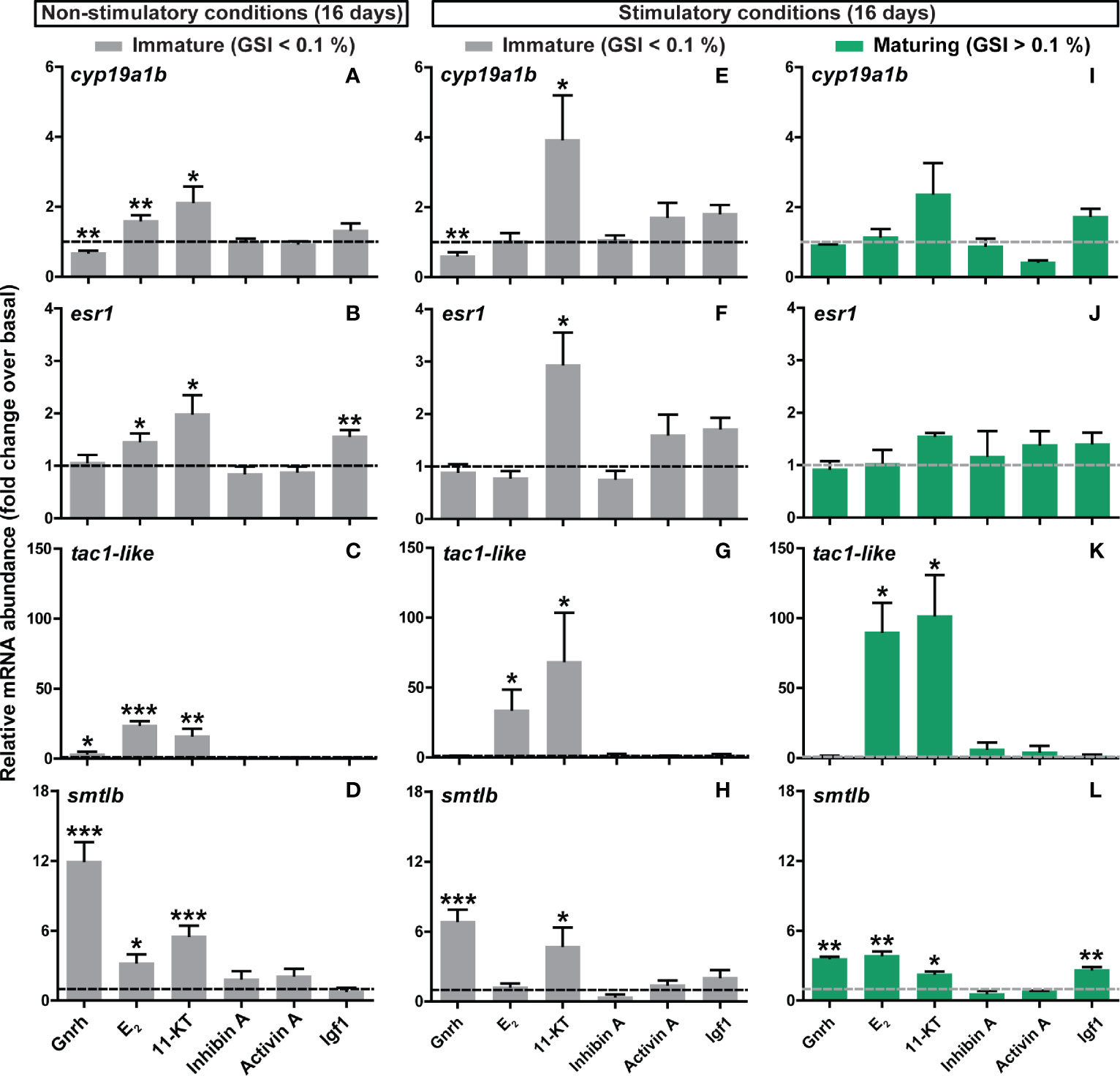
Figure 6 Ex vivo effects of different ligands on selected pituitary genes identified by RNAseq. (A–D) Expression levels of steroid (cyp19a1b, esr1), neurotransmission (tac1-like) and pituitary hormone (smtlb) genes in pituitaries collected from immature postsmolts exposed to standard photoperiod conditions and 16°C for 16 days, and subsequently incubated ex vivo for 9 days in the presence of various ligands. (E–L) The same set of candidate genes was analyzed in immature (E–H) and maturing (I–L) pituitary tissue after exposure to stimulatory conditions (constant light and 16°C) for 16 days, and subsequently incubated ex vivo for 9 days in the presence of the same ligands. Data are shown as mean fold change ± SEM (N = 4-11; *p < 0.05; **p < 0.01; ***p < 0.001) and expressed relative to the basal control group, which is set at 1 (dashed line).
Similar results with respect to Gnrh and steroids were found in pituitaries from males that remained immature after exposure to stimulatory conditions, however, exposure to E2 did not reach statistical significance, except for tac1-like (Figures 6E–H). In fish that did enter pubertal development in vivo in response to the stimulatory regime, only 2 of the genes investigated were modulated ex vivo (tac1-like and smtlb; Figures 6I-L). Transcript levels of tac1-like again showed the strong, and smtlb more modest, response to E2 and 11-KT (Figure 6K); steroids and Gnrh and now also Igf1 significantly up-regulated smtlb expression (Figure 6L).
These results suggest that, as observed for fshb, ex vivo changes in the pituitary transcriptome are more sensitive to regulatory signals in immature salmon males and that treatment with Gnrh and/or steroids consistently modulated pituitary gene expression.
Additional functional studies were carried out to investigate changes in pituitary gene expression in response to long-term maturation trials. For that purpose, immature males were exposed to stimulatory conditions (constant light and 16°C) for 7 weeks and the incidence of maturation was scored weekly from week 4 of treatment. As expected, a higher proportion of maturing males was found compared to ~2 weeks (16 days) of exposure to stimulatory conditions (Supplementary Figures 1E and 6A). Also GSI values of maturing males were clearly higher (Figures 1C and Supplementary Figure 6B) and significant increases in both body weight and length, and condition factor, were found in long-term treated fish (Supplementary Figures 6C-E). In vivo pituitary gnrhr2bba, more clearly lhb, and very clearly fshb transcripts all increased in maturing males kept under long-term stimulatory conditions (Supplementary Figure 6E). Although significant, correlations between GSI and gonadotropin gene expression were weaker in fish exposed to the long-term (Supplementary Figures 2C, F, I) compared to the medium-term maturation regime (Supplementary Figures 2B, E, H). Remarkably, pituitaries from males that remained immature despite 7 weeks of stimulatory conditions, did not respond to Gnrh or 11-KT ex vivo by increased fshb transcript levels (Supplementary Figure 7A), in contrast to the immature fish in the medium-term experiments (Figure 5). The 11-KT-mediated increase in lhb and gnrhr2bba transcript levels, on the other hand, was still observable, albeit less prominently than after ~2 weeks of stimulatory conditions (Figures 5 and Supplementary Figure 7).
The pituitary gland produces a variety of hormones regulating the development and functioning of tissues/organs involved in all major life processes in vertebrates, including the master regulators of gametogenesis and sexual maturation, the gonadotropins FSH and LH (1, 2). In fish species with long, annual reproductive cycles, analysis of plasma levels during the cycle suggest that Fsh release increases when spermatogenesis is initiated and stays elevated during the testicular growth phase while increasing/peak Lh levels appear (much) closer to the spawning season, when the spermatogenic process is already far progressed or completed. This suggests that Lh effects in males, probably mediated to a great part by very high androgen levels, are more related to spawning-related secondary sexual characters/behaviour, while Fsh drives spermatogenesis (52, 53). This scenario seems to also apply to the Atlantic salmon reproductive cycle, where fshb pituitary transcript levels increased during initial steps of pubertal development in males (16, 28). A homologous Fsh immunoassay is not available for Atlantic salmon to date, but Fsh plasma levels peak during sexual maturation in other salmonids (17, 18). When measuring Fsh plasma levels in Atlantic salmon using a radioimmunoassay for coho salmon Fsh, GSI and Fsh values were correlated during the main testicular growth phase in spring and summer (10). Therefore, the Atlantic salmon is an excellently suited model to specifically focus on the role of Fsh at the beginning of puberty/spermatogenesis.
Previous studies on gonadotropins in Atlantic salmon parr (54) or grilse (55), coho salmon (18) or rainbow trout (Oncorhynchus mykiss) (17, 56) collected samples during natural reproductive cycles. Analyzing gonadotropin blood levels, Lh is very low or undetectable while Fsh is clearly present in the circulation for the several months that most of testicular growth takes place. Furthermore, examining Fsh effects on testicular steroidogenesis (22) or gene expression (57) suggested that Fsh can induce testis maturation in salmonids. However, it is not clear what changes are taking place in the pituitary when males enter puberty. Here, we applied an experimental approach and subjected a sibling group of immature fish to environmental conditions known to promote entry into puberty, which split the treatment group into responders and non-responders. Subsequently, we focused on analyzing puberty-associated changes in pituitary gene expression.
Our main findings are: 1) when sampled one week after exposure to stimulatory conditions, changes in the pituitary were already clearly established in responders, while no morphologically discernible changes had occurred in testis tissue yet; 2) analysis of the pituitary transcriptome revealed a transcriptional network during the onset of puberty that was characterized by the regulation of different pituitary hormones and the activation of Gnrh and steroid signaling systems; 3) when sampled two or more weeks after exposure to stimulatory conditions, there was a strong correlation between the up-regulation of fshb in the pituitary and activation of pubertal spermatogenesis and maturing males showed increased growth; 4) the transition into puberty in vivo in response to stimulatory conditions blunts both, the stimulatory effect of Gnrh and the inhibitory effect of 11-KT, on fshb transcript levels otherwise seen as response in pituitaries collected from immature males; 5) the fshb transcript levels in pituitaries of males not responding to long-term exposure to stimulatory conditions in vivo also did not respond ex vivo to 11-KT or Gnrh.
Considering the pituitary hormone of central relevance for puberty, fshb levels largely (~10 times) exceeded those of lhb already in immature postsmolt males kept under standard, non-stimulatory photoperiod conditions (16, 55), and a stronger fshb/GSI than lhb/GSI correlation was found upon maturation. Here, we moreover added to the previous observations the response to stimulatory conditions (27), namely a clear up-regulation of fshb (4.25-fold), while lhb also significantly increased (2.56-fold) but showed a smaller amplitude. Both, the higher baseline expression values and the stronger up-regulatory response, support the view that Fsh is the relevant gonadotropin at the start of puberty in salmon. However, the physiological significance of gonadotropins ultimately relies on their actual plasma concentrations, so that homologous assays for plasma gonadotropins are required to further investigate their involvement in gonad development in Atlantic salmon.
In addition to studies on mRNA abundance in pituitary tissue, parallel analyses were carried out to investigate gonadotropin expression and their cellular localization at the protein level. The specific antiserum for Atlantic salmon Fsh protein generated in the current study and a previously validated anti-(coho salmon)Lh antibody (18) served to show that, akin to the respective mRNA levels, Fsh is abundant in contrast to Lh in immature salmon pituitaries. Several Fsh-positive cells were found in the PPD, while Lh-positive cells were scarce. The number of both gonadotroph cell types increased in maturing fish, though reaching statistical significance only in the case of Fsh cells in the samples analyzed here. Overall, the observed changes in protein and transcript levels agreed well with each other. The background of the strong (~5-fold) increase in the number of Fsh cells requires further studies. Previous work in juvenile African catfish (Clarias gariepinus) showed that an increase in Lh cell number following estrogen treatment was not associated with cell proliferation, but rather with a recruitment of pre-existing quiescent into hormone producing cells (58). In medaka (Oryzias latipes), on the other hand both, recruitment and proliferation was reported during ontogenesis and following estrogen exposure (59).
Previous work suggests that, different from mammals, gonadotropic hormones are produced and secreted by distinct endocrine cells in the teleost pituitary (60, 61). Studies using transgenic zebrafish and tilapia (Oreochromis niloticus) expressing fluorescence-tagged gonadotropes (62) and single-cell RNAseq of tilapia pituitaries (63) confirmed that Fsh and Lh are synthesized and secreted from distinct cells. However, a study in medaka that combined multi-color in situ hybridization (ISH) and single-cell RNAseq analyses revealed the existence of a small fraction (4%) among the gonadotrophs co-expressing fshb and lhb (64). Furthermore, in primary cell culture, medaka Fsh cells can produce Lhb (65). Considering the plasticity of gonadotropes (4), the possible existence of multi-hormonal cells cannot be ruled out for Atlantic salmon, although both transcript and protein data suggest that this is not relevant quantitatively during the initiation of pubertal spermatogenesis.
The regulation of pituitary gonadotropin production and release is based on a network of neuroendocrine factors from the hypothalamus that also integrates signal molecules produced by peripheral organs (4, 66). However, knowledge on genes/pathways operating in the teleost pituitary during the initiation of puberty and their potential involvement in the regulation of gonadotropin production and release is still limited. A few studies have examined global changes on pituitary gene expression, comparing juvenile and adult males in model fish species (64, 67). However, no information is available on transcriptional changes in the pituitary during the narrow time window of initiating male puberty. Previous work of our group has described changes in the testis transcriptome when entering puberty in Atlantic salmon, a period characterized by increased fshb pituitary transcript and 11-KT plasma levels (16). Accordingly, these two parameters have been used here for assigning pituitary samples to the two groups (responders and non-responders). Our RNAseq results showed that the genes differentially expressed in the pituitary of maturing males included factors involved in Gnrh and steroid signaling, neurotransmission and extracellular matrix remodeling.
Regarding pituitary hormones, the transcript levels of fshb, cgaa, cgab, smtlb were all enhanced at the onset of maturation (fshb showing the highest amplitude of 9.51-fold increase), while tshbb expression was strongly inhibited. This pattern is in agreement with what has been described previously in zebrafish (67). Reduced tshb expression in maturing fish is very interesting in context with observations in zebrafish, showing that thyroid hormone potentiated Fsh-induced androgen production (68). Recent genetic evidence supported this thyroid-testis connection, considering that tshba mutant males neither showed secondary sex characters nor reproductive behaviour when tested with wild-type females (69). Assuming the existence of a similar thyroid-testis connection in salmon, it seems possible for example that androgens exert a negative feedback on pituitary thyrotropes. In contrast to tshbb, smtlb expression levels were enhanced in vivo in maturing fish in the present study. Two different forms of somatolactin (alpha and beta) were previously cloned in Atlantic salmon, and both forms increased in the pituitary during vitellogenesis and spawning in females (70). In coho salmon males, plasma somatolactin increased during the rapid testicular growth phase, was highly correlated to the 11-KT plasma concentration, and peaked during final maturation and spawning (71). Here, we found a consistent 11-KT stimulation of smtlb expression in pituitary cultures, irrespective of the maturational status and of the environmental conditions. While this positive feedback of 11-KT on somatolactin production in Atlantic salmon fits well to the observed changes in coho salmon androgen and somatolactin plasma levels, little is known so far regarding potential reproductive functions of somatolactin.
In both zebrafish (67) and medaka (64), but not in salmon (this RNAseq study analyzing samples collected within 1.5 weeks after starting the exposure to stimulatory conditions), lhb expression levels were also increased after the start of puberty. This discrepancy probably reflects the fact that those studies compared juvenile −immature− with adult −mature− stages while here, we compare two early stages of development, representing fish before and about to enter puberty, but prior to histologically visible signs of testis maturation. However, as mentioned above, lhb transcript abundance is elevated also in salmon when exposure to stimulatory conditions lasted longer and developmental changes have become obvious also in testis tissue, viz. the presence of type B spermatogonia and increased GSI values. This suggests that initiating puberty primarily involved activating fshb/Fsh expression and release, while effects on lhb gene expression seem secondary in nature (see below section on sex steroid effects). Next to effects on lhb transcript levels based on Fsh-triggered sex steroid production (see below), recent work in tilapia revealed that Gnrh-stimulated Fsh-producing cells can promote hormone synthesis in Lh-producing cells in a paracrine manner (72). However, as mentioned earlier, Lh plasma levels do not increase until much later in the salmonid reproductive cycle.
In addition to pituitary hormones, the expression levels of relevant signaling molecules involved in the regulation of gonadotropin expression and/or gonadotrope function in mammals, nr5a1 (73, 74), nr5a2 (75, 76) and sstr2 (77), were also significantly elevated in the pituitary gland of maturing salmon. As one of the neuropeptides regulating gonadotropin expression and synthesis in vertebrates (78), enrichment of pituitary-expressed genes related to Gnrh signaling was expected. Among those, tac1-like, fgf8, duox2, sema3ab and sox4 are considered as regulators of GnRH gene expression and/or GnRH secretion in mammals (79–83). Indeed, gene expression profiling of hypothalamic-pituitary-gonadal axis tissues identified Sema3a and Fgf8 as puberty-associated factors in mice (84), and loss-of-function mutations in both genes are associated to pubertal disorders in humans (80, 85). However, in mammals, these factors mainly function in the hypothalamus while we found differential expression in the pituitary. Interestingly, in tilapia, neurokinin B (Tac3) and its receptor (Tac3r) are both expressed by Lh- and Fsh-producing cells (86), and Tac3 analogues increased pituitary fshb mRNA and brain gnrh mRNA levels (72). In mammals, the TAC3/TAC3R genes are critical for the functioning of the Kiss1-neurons in the hypothalamic arcuate nucleus, responsible for both, the negative steroid feedback as well as the pulsatile release of Gnrh (87). However, also TAC1/Tac1, encoding substance P and neurokinin A, have been implicated in the regulation of reproductive functions (88–91). Tac1 mutant male and female mice showed delayed puberty onset (79, 92) and, conversely, exposure to TAC1R agonists triggered sexual maturation of pre-pubertal females (92). Recently, tac1 transcript expression has been reported in the zebrafish brain but, unlike mammals, Tac1 action on Gnrh3 neurons seems independent of kisspeptin (93). In the present study, transcripts encoding different Tac and Tacr variants were detected in pituitary tissue but were not differentially expressed (data not shown), except for tac1-like. Future work will have to show if signaling via peptides derived from the tac1-like transcript in the pituitary is relevant for regulating gonadotroph functions in salmon. For the present study, we included the tac1-like paralog for further qPCR studies, since it was the only differentially expressed one among the 4 paralogs (see below).
A total of six Gnrh receptor paralogs have been identified in the Atlantic salmon genome, gnrhr2bba being the only paralog stimulated during precocious male parr maturation (94). The same study reported that gnrhr2bba transcript was solely expressed in lhb-expressing cells, however, a correlation between fshb and gnrhr2bba transcript levels (both increased) in earlier stages of maturation was also observed (94). Results obtained in other fish species support Gnrhr expression by Fsh-cells: (i) a recent single-cell RNAseq study in tilapia found cell-specific enrichment of gnrhr1 in Fsh cells; (ii) in zebrafish, a close association (direct contact) was observed between Fsh cells and Gnrh3 axons (95). Until confirmation by other techniques than ISH, and studies at other life stages than parr, one should not yet dismiss the possibility of direct Gnrh action on Fsh-cells in salmon. In any case, the published work in different fish species suggest several options for stimulating Fsh-cells, such as direct effects, indirect effects via Gnrhr on Lh-cells, but also via other (non Gnrh) peptides, such as the Tac family (96). Irrespective of the mechanism(s) used by Gnrh, earlier studies in pre-pubertal male coho salmon showed that Gnrh increased fshb transcript levels as well as Fsh release from primary pituitary cell cultures (6). We confirmed the stimulatory effect on fshb transcript levels in the present study when culturing pituitary tissue from immature Atlantic salmon males, and moreover found that Gnrh treatment also increased gnrhr2bba transcript levels. Thus, in immature males, Gnrh seems to prime Fsh-cells to respond to subsequent Gnrh stimuli, thereby triggering in an auto-stimulatory manner increased Fsh production at the onset of puberty. Since positive feedback systems require inhibitory loops to avoid overstimulation, it was interesting to note that when exposure to stimulatory conditions had activated spermatogenesis to proceed to producing type B spermatogonia, the fshb response to Gnrh in culture was blunted. This can be understood by assuming that Gnrh-mediated signaling was already fully stimulated in the more progressed fish, so that their pituitaries were unable to respond to additional stimuli in culture. Alternatively, the more progressed testicular state of development may have blunted the Gnrh effect specifically regarding Fsh. We favour the latter hypothesis, since Gnrh kept stimulating somatolactin (smtlb) transcript levels in pituitary tissue from fish showing type B spermatogonia. Finally, Gnrh only increased gnrhr2bba transcript in immature fish exposed to non-stimulatory conditions, and exposure to stimulatory conditions that did not result yet in producing type B spermatogonia already blunted the Gnrh-induced gnrhr2bba response, which potentially contributes to preventing overstimulation of Fsh-cells. Taken together, it appears that Gnrh signaling plays a self-stimulatory role during the initiation of puberty to kick-start Fsh-production, but that this role may be limited in time, in view of the blunting of the effect on gnrhr2bba or fshb transcript levels once males have experienced stimulatory environmental conditions or commenced the production of type B spermatogonia, respectively. Hence, we assume that the exposure to stimulatory environmental conditions triggers central-nervous processes upstream of Gnrh, resulting in increased hypophysiotropic Gnrh release.
Gonadal sex steroids are key molecules controling gonadotropin production and release in vertebrates. Due to the limited availability of Fsh assays for fish species, information on the regulation of Fsh release is relatively scarce, compared to Lh. Sex steroid effects on lhb gene expression can be mediated by estrogen receptors and their use of estrogen response elements in the lhb promoter/enhancer region (34, 97, 98), but probably also by androgen receptors (15, 99, 100). Despite the essential role of androgens, such as T and 11-KT in male reproductive physiology also in fish (101–103), not a single gene annotated as being androgen-specific, was significantly regulated in our RNAseq study. Literature describing cell type-specific androgen receptor expression at the pituitary level is not available in salmonids to date, but in tilapia three different androgen receptor paralogs were enriched specifically in Fsh cells (63). Evidence for a direct effect of androgens comes from studies, in which non-aromatizable, 11-oxygenated androgens increased lhb transcript levels in three teleost species [Atlantic salmon parr (15), African catfish (99) or stickleback (100)]. Our present experiments confirmed this stimulatory effect of 11-KT on lhb transcript levels and moreover revealed, that this effect is (i) a direct one on the pituitary level, and (ii) an invariable response not depending on the maturational state or the environmental conditions. Regarding the initiation of puberty, on the other hand, this response of lhb transcript levels seems to be of little acute physiological relevance, considering the previously discussed, non-detectable or very low Lh plasma levels in salmonid fish during the testicular growth phase. However, 11-KT is known to exert negative feedback effects on Fsh release in coho (11, 104) and Atlantic salmon (15). We also found a negative feedback effect of 11-KT quantifying fshb transcript levels in our studies, and moreover saw that this inhibitory effect was characteristic of immature males but disappeared after the start of spermatogenesis, i.e. this inhibitory effect is restricted to immature fish. Importantly, this may explain the apparently paradoxical observation of concomitantly increasing Fsh and 11-KT plasma levels, despite the negative feedback effect of 11-KT on Fsh release previously reported by others in salmonids (see above). Moreover, once Fsh release has induced the start of spermatogenesis via modulating Sertoli growth factor (105) and Leydig cell androgen production, including the main teleost androgen 11-KT (22), the elevated 11-KT levels invariably increase pituitary gnrhr2bba transcript levels. We assume that the consistent up-regulation of Gnrh receptor expression in combination with the disappearing inhibition of 11-KT-mediated repression of Fsh production jointly allows for the concomitant rise of the two main stimulators of spermatogenesis: Fsh and androgens, assuming that Gnrh release is not a limiting factor.
Androgen-mediated up-regulation of gonadotropin gene expression has additional components, one of them involving estrogen signaling. We have found elevated expression of estrogen signaling genes (esr1, cyp19a1b and cyp1b1) in pituitaries of maturing male salmon. Pituitary expression of the estrogen receptor 1 (esr1) transcript has been identified in numerous fishes (13) and, in some species, was localized in gonadotropes (34, 59, 106, 107). Similarly, the pituitary of several teleost species, including Atlantic salmon (108), showed brain aromatase gene expression (cyp19a1b) or activity. The use of aromatase inhibitors (34, 109) and of aromatizable and non-aromatizable androgens (59, 65) demonstrated the importance of aromatase-mediated conversion of T to E2. Since gonadotropes in certain fishes express estrogen receptors and cyp19a1b, E2 (or T after intracellular conversion to E2) could regulate this cell type’s functionality, as hypothesized for medaka Lh-producing cells (59).
Our data on the KEGG pathway enrichment analysis also indicated that, similar to zebrafish (67) and medaka (64), genes related to cell adhesion and extracellular matrix (ECM) are modulated in the pituitary during the start of puberty. This is in accordance with previous studies in mammals, in which components of the ECM were shown to modulate pituitary cell proliferation and hormone secretion (110). As reviewed by Fontaine et al. (4), gonadotrope cells display an extraordinary plasticity at cellular, population and structural level in both fish and mammals, and some physiological processes (including pubertal development) require a remarkable reorganization of the anterior pituitary structure. Matrix remodeling in pituitary cells, through members of the metalloproteinase (MMP) family (i.e. MMP2 and MMP9), was described in human cell lines, an effect that was enhanced after GnRH treatment (111). In this study, we found the modulated expression of the Mmp members mmp14 and adam10, and of spock1, a factor known to inhibit MMP2 activity (112). While MMP14 and ADAM10 regulate cell migration and invasion in pituitary adenomas (113, 114), no clear function for SPOCK1 has been described so far in pituitary tissue. However, a genome-wide study has identified SPOCK1 as a key gene underlying age at menarche in Caucasian woman (112). Therefore, it is tempting to speculate that, also in fish, ECM remodeling may play a role in changes of pituitary cell homeostasis and hormone production accompanying puberty in Atlantic salmon.
Our study also investigated potential effects of different growth factors on pituitary gene expression. The selection of candidates was based on literature available in other fish species in which modulatory effects were reported (8, 115, 116). However, none of the candidates tested provided consistent data, under the experimental conditions investigated, preventing to draw conclusions as to their involvement in regulating gonadotropin gene expression. In particular the lack of effect of activin/inhibin was unexpected. In the European eel (Anguilla anguilla) and in goldfish (Carassius auratus), activin stimulated pituitary fshb mRNA levels (12, 115). In adult mammals, activin of pituitary origin and inhibin of gonadal origin stimulate and inhibit pituitary FSH release, respectively. The stimulatory effect of activin reflects changes in the Gnrh pulse frequency-dependent availability of the activin binding protein follistatin rather than changes in the activin amount (117). While no information is available in fish in this regard, our RNAseq data in salmon show that the expression levels of activin, inhibin and their receptors were low or very low and none of them differentially expressed; the same applies to follistatin (data not shown). Therefore, it appears that activin/inhibin/follistatin signaling is of limited relevance in salmon before and at the beginning of puberty. Considering inhibin, finally, recent genetic experiments in zebrafish suggest that this competitive inhibitor of activin is not of critical relevance for male puberty or adult spermatogenesis (118).
We have summarized our main findings and integrated them with literature data in Figure 7. We assume that exposing immature males to stimulatory environmental conditions can trigger the release of Gnrh. The pituitary responds by releasing the at this time dominating gonadotropin Fsh, resulting in an activation of Leydig cell steroid production, including 11-KT and T. Gnrh also increases pituitary Gnrh receptor expression, but this response is restricted to immature fish that had not been exposed to stimulatory environmental conditions yet, and therefore may represent a short-lived positive feedback loop to initiate the maturational response. Gnrh furthermore increased pituitary fshb transcript levels, a response also seen after exposure to stimulatory conditions, but that disappeared with the appearance of type B spermatogonia in the testis, i.e. may also be restricted to the initial phase of puberty. This developmental transition also appeared relevant for the inhibitory effect of 11-KT on fshb transcript levels, considering it was only apparent as long as the testes did not contain type B spermatogonia yet. Since 11-KT always (i.e. irrespective of the maturational status and the environmental conditions) increased pituitary Gnrh receptor expression, this setting may result initially in a somewhat limited, but then soon in a more important increase in Fsh and hence androgen production, jointly further supporting the progress of pubertal spermatogenesis. Both, 11-KT and T via the androgen receptor and T after conversion to E2 via the estrogen receptor, increase Lh production that, however, will not be released until approaching the actual spawning season. Finally, both androgen receptor and estrogen receptor ligands always strongly increased pituitary tac1-like gene expression in a selective manner, considering that other members of the Tac signaling system were not identified as DEGs by RNAseq. Tac-related neuropeptides potentially stimulate gonadotropin production, and this may apply to peptides derived from the tac1-like gene as well. Overall, it appears that in our experimental model the regulatory mechanisms sensitive to photoperiod and temperature mainly use Gnrh signaling to trigger a network of downstream processes. However, genetic evidence suggests that even in the complete absence of Gnrh signaling, zebrafish can still reproduce, demonstrating that the pathways highlighted by our studies are not of the ‘must be taken’ but rather of the ‘can be taken’ category, as has been stressed recently (5, 96) in the light of alternative signaling routes.

Figure 7 Schematic illustration summarizing the regulation of pituitary Fsh/fshb expression and production at the, experimentally induced, onset of puberty in male Atlantic salmon. Described effects are indicated by solid lines, while dashed lines denote no experimental evidence reported here but previously demonstrated in other studies. Yellow lines highlight effects that are potentially limited in time: in vivo response to stimulatory conditions blunts both, the stimulatory effect of Gnrh on fshb and gnrhr2bba, and the inhibitory effect of 11-KT on fshb transcript levels. Gnrh, gonadotropin-releasing hormone; Fsh, follicle-stimulating hormone; Lh, luteinizing hormone; 11-KT, 11-ketotestosterone; T, testosterone; E2, 17β-estradiol; Tac1-like, protachykinin-like; gnrhr2bba, gonadotropin-releasing hormone receptor 2bba; cyp19a1b, brain aromatase; esr1, estrogen receptor 1.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
The animal study was reviewed and approved by the Norwegian Animal Research Authority (NARA, permit number 5741) and the use of these experimental animals was in accordance with the Norwegian Animal Welfare Act.
DC, KS, OY, SO, PV, and JB performed the experiments. DC, KS, EK-S, OY, BN, SO, PV, JB, LK, EA, AW, PF, and RS analyzed and contributed to the interpretation of the results. RE, JB, EA, AW, TH, PF, and RS conceived the project, secured funding, and provided the supervision. DC, AW, PF, and RS wrote the manuscript. All authors contributed to the article and approved the submitted version.
This research was financed with resources from The Research Council of Norway (POSTSMOLTMAT project, No. 254870), the EU Seventh Framework Programme via the AQUAEXCEL project (No. 262336), and the European Union’s Horizon 2020 research and innovation programme under grant agreement No 652831.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors thank Dr Penny Swanson (Northwest Fisheries Science Center, NOAA, USA) for kind donation of anti-Lh antibody, and Tomasz Furmanek (Institute of Marine Research, Bergen, Norway) and Karen Anita Kvestad (Institute of Marine Research, Matre Research Station, Matredal, Norway) for technical support.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2022.826920/full#supplementary-material
1. Edwards W, Raetzman LT. Complex Integration of Intrinsic and Peripheral Signaling Is Required for Pituitary Gland Development. Biol Reprod (2018) 99(3):504–13. doi: 10.1093/biolre/ioy081
2. Perez-Castro C, Renner U, Haedo MR, Stalla GK, Arzt E. Cellular and Molecular Specificity of Pituitary Gland Physiology. Physiol Rev (2012) 92(1):1–38. doi: 10.1152/physrev.00003.2011
3. Coss D. Regulation of Reproduction via Tight Control of Gonadotropin Hormone Levels. Mol Cell Endocrinol (2018) 463:116–30. doi: 10.1016/j.mce.2017.03.022
4. Fontaine R, Ciani E, Haug TM, Hodne K, Ager-Wick E, Baker DM, et al. Gonadotrope Plasticity at Cellular, Population and Structural Levels: A Comparison Between Fishes and Mammals. Gen Comp Endocrinol (2020) 287:113344. doi: 10.1016/j.ygcen.2019.113344
5. Trudeau VL. Neuroendocrine Control of Reproduction in Teleost Fish: Concepts and Controversies. Annu Rev Anim Biosci (2022) 10:107–30. doi: 10.1146/annurev-animal-020420-042015
6. Dickey JT, Swanson P. Effects of Salmon Gonadotropin-Releasing Hormone on Follicle Stimulating Hormone Secretion and Subunit Gene Expression in Coho Salmon (Oncorhynchus Kisutch). Gen Comp Endocrinol (2000) 118(3):436–49. doi: 10.1006/gcen.2000.7482
7. Lin SW, Ge W. Differential Regulation of Gonadotropins (FSH and LH) and Growth Hormone (GH) by Neuroendocrine, Endocrine, and Paracrine Factors in the Zebrafish–An In Vitro Approach. Gen Comp Endocrinol (2009) 160(2):183–93. doi: 10.1016/j.ygcen.2008.11.020
8. Luckenbach JA, Dickey JT, Swanson P. Regulation of Pituitary GnRH Receptor and Gonadotropin Subunits by IGF1 and GnRH in Prepubertal Male Coho Salmon. Gen Comp Endocrinol (2010) 167(3):387–96. doi: 10.1016/j.ygcen.2009.09.010
9. Rebers FE, Bosma PT, van Dijk W, Goos HJ, Schulz RW. GnRH Stimulates LH Release Directly via Inositol Phosphate and Indirectly via cAMP in African Catfish. Am J Physiol Regul Integr Comp Physiol (2000) 278(6):R1572–8. doi: 10.1152/ajpregu.2000.278.6.R1572
10. Antonopoulou E, Swanson P, Borg B. Effects of Aromatase Inhibitors and Different Doses of Testosterone on Gonadotropins in One Year Old Male Atlantic Salmon (Salmo Salar). Comp Biochem Physiol A Mol Integr Physiol (2009) 153(4):408–16. doi: 10.1016/j.cbpa.2009.03.019
11. Dickey JT, Swanson P. Effects of Sex Steroids on Gonadotropin (FSH and LH) Regulation in Coho Salmon (Oncorhynchus Kisutch). J Mol Endocrinol (1998) 21(3):291–306. doi: 10.1677/jme.0.0210291
12. Yam KM, Yoshiura Y, Kobayashi M, Ge W. Recombinant Goldfish Activin B Stimulates Gonadotropin-Ibeta But Inhibits Gonadotropin-IIbeta Expression in the Goldfish, Carassius Auratus. Gen Comp Endocrinol (1999) 116(1):81–9. doi: 10.1006/gcen.1999.7339
13. Fontaine R, Royan MR, von Krogh K, Weltzien FA, Baker DM. Direct and Indirect Effects of Sex Steroids on Gonadotrope Cell Plasticity in the Teleost Fish Pituitary. Front Endocrinol (Lausanne) (2020) 11:605068. doi: 10.3389/fendo.2020.605068
14. Melo MC, van Dijk P, Andersson E, Nilsen TO, Fjelldal PG, Male R, et al. Androgens Directly Stimulate Spermatogonial Differentiation in Juvenile Atlantic Salmon (Salmo Salar). Gen Comp Endocrinol (2015) 211:52–61. doi: 10.1016/j.ygcen.2014.11.015
15. Antonopoulou E, Swanson P, Mayer I, Borg B. Feedback Control of Gonadotropins in Atlantic Salmon, Salmo Salar, Male Parr.II. Aromatase Inhibitor and Androgen Effects. Gen Comp Endocrinol (1999) 114(1):142–50. doi: 10.1006/gcen.1998.7248
16. Crespo D, Bogerd J, Sambroni E, LeGac F, Andersson E, Edvardsen RB, et al. The Initiation of Puberty in Atlantic Salmon Brings About Large Changes in Testicular Gene Expression That Are Modulated by the Energy Status. BMC Genomics (2019) 20(1):475. doi: 10.1186/s12864-019-5869-9
17. Prat F, Sumpter JP, Tyler CR. Validation of Radioimmunoassays for Two Salmon Gonadotropins (GTH I and GTH II) and Their Plasma Concentrations Throughout the Reproductivecycle in Male and Female Rainbow Trout (Oncorhynchus Mykiss). Biol Reprod (1996) 54(6):1375–82. doi: 10.1095/biolreprod54.6.1375
18. Swanson P, Bernard M, Nozaki M, Suzuki K, Kawauchi H, Dickhoff WW. Gonadotropins I and II in Juvenile Coho Salmon. Fish Physiol Biochem (1989) 7(1-6):169–76. doi: 10.1007/BF00004704
19. Garcia-Lopez A, Bogerd J, Granneman JC, van Dijk W, Trant JM, Taranger GL, et al. Leydig Cells Express Follicle-Stimulating Hormone Receptors in African Catfish. Endocrinology (2009) 150(1):357–65. doi: 10.1210/en.2008-0447
20. Garcia-Lopez A, de Jonge H, Nobrega RH, de Waal PP, van Dijk W, Hemrika W, et al. Studies in Zebrafish Reveal Unusual Cellular Expression Patterns of Gonadotropin Receptor Messenger Ribonucleic Acids in the Testis and Unexpected Functional Differentiation of the Gonadotropins. Endocrinology (2010) 151(5):2349–60. doi: 10.1210/en.2009-1227
21. Mazon MJ, Gomez A, Yilmaz O, Carrillo M, Zanuy S. Administration of Follicle-Stimulating Hormone In Vivo Triggers Testicular Recrudescence of Juvenile European Sea Bass (Dicentrarchus Labrax). Biol Reprod (2014) 90(1):6. doi: 10.1095/biolreprod.113.110569
22. Planas JV, Swanson P. Maturation-Associated Changes in the Response of the Salmon Testis to the Steroidogenic Actions of Gonadotropins (GTH I and GTH II) In Vitro. Biol Reprod (1995) 52(3):697–704. doi: 10.1095/biolreprod52.3.697
23. Van der Kraak G, Suzuki K, Peter RE, Itoh H, Kawauchi H. Properties of Common Carp Gonadotropin I and Gonadotropin II. Gen Comp Endocrinol (1992) 85(2):217–29. doi: 10.1016/0016-6480(92)90005-5
24. Taranger GL, Carrillo M, Schulz RW, Fontaine P, Zanuy S, Felip A, et al. Control of Puberty in Farmed Fish. Gen Comp Endocrinol (2010) 165(3):483–515. doi: 10.1016/j.ygcen.2009.05.004
25. Wargelius A, Leininger S, Skaftnesmo KO, Kleppe L, Andersson E, Taranger GL, et al. Dnd Knockout Ablates Germ Cells and Demonstrates Germ Cell Independent Sex Differentiation in Atlantic Salmon. Sci Rep (2016) 6:21284. doi: 10.1038/srep21284
26. Kleppe L, Andersson E, Skaftnesmo KO, Edvardsen RB, Fjelldal PG, Norberg B, et al. Sex Steroid Production Associated With Puberty Is Absent in Germ Cell-Free Salmon. Sci Rep (2017) 7(1):12584. doi: 10.1038/s41598-017-12936-w
27. Fjelldal PG, Hansen T, Huang T. Continuous Light and Elevated Temperature can Trigger Maturation Both During and Immediately After Smoltification in Male Atlantic Salmon (Salmo Salar). Aquaculture (2011) 321:93–100. doi: 10.1016/j.aquaculture.2011.08.017
28. Melo MC, Andersson E, Fjelldal PG, Bogerd J, Franca LR, Taranger GL, et al. Salinity and Photoperiod Modulate Pubertal Development in Atlantic Salmon (Salmo Salar). J Endocrinol (2014) 220(3):319–32. doi: 10.1530/JOE-13-0240
29. Ayllon F, Kjaerner-Semb E, Furmanek T, Wennevik V, Solberg MF, Dahle G, et al. The Vgll3 Locus Controls Age at Maturity in Wild and Domesticated Atlantic Salmon (Salmo Salar L.) Males. PloS Genet (2015) 11(11):e1005628. doi: 10.1371/journal.pgen.1005628
30. Barson NJ, Aykanat T, Hindar K, Baranski M, Bolstad GH, Fiske P, et al. Sex-Dependent Dominance at a Single Locus Maintains Variation in Age at Maturity in Salmon. Nature (2015) 528(7582):405–8. doi: 10.1038/nature16062
31. Fjelldal PG, Hansen TJ, Wargelius A, Ayllon F, Glover KA, Schulz RW, et al. Development of Supermale and All-Male Atlantic Salmon to Research the Vgll3 Allele - Puberty Link. BMC Genet (2020) 21(1):123. doi: 10.1186/s12863-020-00927-2
32. Leal MC, de Waal PP, Garcia-Lopez A, Chen SX, Bogerd J, Schulz RW. Zebrafish Primary Testis Tissue Culture: An Approach to Study Testis Function Ex Vivo. Gen Comp Endocrinol (2009) 162(2):134–8. doi: 10.1016/j.ygcen.2009.03.003
33. Saito K, Chida Y, Adachi S, Yamauchi K. In Vitro Induction of Luteinizing Hormone Synthesis by Estrogen in Organ-Cultured Pituitary Glands of the Japanese Eel, Anguilla Japonica. Zoolog Sci (2003) 20(1):69–73. doi: 10.2108/zsj.20.69
34. Zhang S, Zhang Y, Chen W, Wu Y, Ge W, Zhang L, et al. Aromatase (Cyp19a1b) in the Pituitary Is Dynamically Involved in the Upregulation of Lhb But Not Fshb in the Vitellogenic Female Ricefield Eel Monopterus Albus. Endocrinology (2014) 155(11):4531–41. doi: 10.1210/en.2014-1069
35. Yaron Z, Gur G, Melamed P, Rosenfeld H, Levavi-Sivan B, Elizur A. Regulation of Gonadotropin Subunit Genes in Tilapia. Comp Biochem Physiol B Biochem Mol Biol (2001) 129(2-3):489–502. doi: 10.1016/S1096-4959(01)00345-1
36. Fung RSK, Bai J, Yuen KWY, Wong AOL. Activin/follistatin System in Grass Carp Pituitary Cells: - Regulation by Local Release of Growth Hormone and Luteinizing Hormone and Its Functional Role in Growth Hormone Synthesis and Secretion. PloS One (2017) 12(6):e0179789. doi: 10.1371/journal.pone.0179789
37. Fine M, Amuly R, Sandowski Y, Marchant TA, Chan SJ, Gertler A, et al. Recombinant Gilthead Seabream (Sparus Aurata) Insulin-Like Growth Factor-I: Subcloning, Expression in Escherichia Coli, Purification and Characterization. J Endocrinol (1997) 153(1):139–50. doi: 10.1677/joe.0.1530139
38. Berg S, Kutra D, Kroeger T, Straehle CN, Kausler BX, Haubold C, et al. Ilastik: Interactive Machine Learning for (Bio)Image Analysis. Nat Methods (2019) 16(12):1226–32. doi: 10.1038/s41592-019-0582-9
39. Carpenter AE, Jones TR, Lamprecht MR, Clarke C, Kang IH, Friman O, et al. CellProfiler: Image Analysis Software for Identifying and Quantifying Cell Phenotypes. Genome Biol (2006) 7(10):R100. doi: 10.1186/gb-2006-7-10-r100
40. McQuin C, Goodman A, Chernyshev V, Kamentsky L, Cimini BA, Karhohs KW, et al. CellProfiler 3.0: Next-Generation Image Processing for Biology. PloS Biol (2018) 16(7):e2005970. doi: 10.1371/journal.pbio.2005970
41. Stirling DR, Swain-Bowden MJ, Lucas AM, Carpenter AE, Cimini BA, Goodman A. CellProfiler 4: Improvements in Speed, Utility and Usability. BMC Bioinf (2021) 22(1):433. doi: 10.1186/s12859-021-04344-9
42. Cuisset B, Pradelles P, Kime DE, Kühn ER, Babin P, Davail S, et al. Enzyme Immunoassay for 11-Ketotestosterone Using Acetylcholinesterase as Laberl: Application to the Measurement of 11-Ketotestosterone in Plasma of Siberian Sturgeon. Comp Biochem Physiol Part C: Pharmacol Toxicol Endocrinol (1994) 108(2):229–41. doi: 10.1016/1367-8280(94)90035-3
43. Andersson E, Schulz RW, Male R, Bogerd J, Patina D, Benedet S, et al. Pituitary Gonadotropin and Ovarian Gonadotropin Receptor Transcript Levels: Seasonal and Photoperiod-Induced Changes in the Reproductive Physiology of Female Atlantic Salmon (Salmo Salar). Gen Comp Endocrinol (2013) 191:247–58. doi: 10.1016/j.ygcen.2013.07.001
44. Livak KJ, Schmittgen TD. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (2001) 25(4):402–8. doi: 10.1006/meth.2001.1262
45. Langmead B, Salzberg SL. Fast Gapped-Read Alignment With Bowtie 2. Nat Methods (2012) 9(4):357–9. doi: 10.1038/nmeth.1923
46. Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, et al. The Sequence Alignment/Map Format and SAMtools. Bioinformatics (2009) 25(16):2078–9. doi: 10.1093/bioinformatics/btp352
47. Anders S, Huber W. Differential Expression Analysis for Sequence Count Data. Genome Biol (2010) 11(10):R106. doi: 10.1186/gb-2010-11-10-r106
48. Liao Y, Wang J, Jaehnig EJ, Shi Z, Zhang B. WebGestalt 2019: Gene Set Analysis Toolkit With Revamped UIs and APIs. Nucleic Acids Res (2019) 47(W1):W199–205. doi: 10.1093/nar/gkz401
49. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al. Gene Ontology: Tool for the Unification of Biology. Gene Ontol Consortium Nat Genet (2000) 25(1):25–9. doi: 10.1038/75556
50. Raudvere U, Kolberg L, Kuzmin I, Arak T, Adler P, Peterson H, et al. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res (2019) 47(W1):W191–W8. doi: 10.1093/nar/gkz369
51. Wang S, Furmanek T, Kryvi H, Krossoy C, Totland GK, Grotmol S, et al. Transcriptome Sequencing of Atlantic Salmon (Salmo Salar L.) Notochord Prior to Development of the Vertebrae Provides Clues to Regulation of Positional Fate, Chordoblast Lineage and Mineralisation. BMC Genomics (2014) 15:141. doi: 10.1186/1471-2164-15-141
52. Levavi-Sivan B, Bogerd J, Mananos EL, Gomez A, Lareyre JJ. Perspectives on Fish Gonadotropins and Their Receptors. Gen Comp Endocrinol (2010) 165(3):412–37. doi: 10.1016/j.ygcen.2009.07.019
53. Schulz RW, de Franca LR, Lareyre JJ, Le Gac F, Chiarini-Garcia H, Nobrega RH, et al. Spermatogenesis in Fish. Gen Comp Endocrinol (2010) 165(3):390–411. doi: 10.1016/j.ygcen.2009.02.013
54. Maugars G, Schmitz M. Expression of Gonadotropin and Gonadotropin Receptor Genes During Early Sexual Maturation in Male Atlantic Salmon Parr. Mol Reprod Dev (2008) 75(2):403–13. doi: 10.1002/mrd.20767
55. Schulz RW, Taranger GL, Bogerd J, Nijenhuis W, Norberg B, Male R, et al. Entry Into Puberty is Reflected in Changes in Hormone Production But Not in Testicular Receptor Expression in Atlantic Salmon (Salmo Salar). Reprod Biol Endocrinol (2019) 17(1):48. doi: 10.1186/s12958-019-0493-8
56. Gomez JM, Weil C, Ollitrault M, Le Bail PY, Breton B, Le Gac F. Growth Hormone (GH) and Gonadotropin Subunit Gene Expression and Pituitary and Plasma Changes During Spermatogenesis and Oogenesis in Rainbow Trout (Oncorhynchus Mykiss). Gen Comp Endocrinol (1999) 113(3):413–28. doi: 10.1006/gcen.1998.7222
57. Sambroni E, Lareyre JJ, Le Gac F. Fsh Controls Gene Expression in Fish Both Independently of and Through Steroid Mediation. PloS One (2013) 8(10):e76684. doi: 10.1371/journal.pone.0076684
58. Cavaco JE, van Baal J, van Dijk W, Hassing GA, Goos HJ, Schulz RW. Steroid Hormones Stimulate Gonadotrophs in Juvenile Male African Catfish (Clarias Gariepinus). Biol Reprod (2001) 64(5):1358–65. doi: 10.1095/biolreprod64.5.1358
59. Fontaine R, Ager-Wick E, Hodne K, Weltzien FA. Plasticity of Lh Cells Caused by Cell Proliferation and Recruitment of Existing Cells. J Endocrinol (2019) 240(2):361–77. doi: 10.1530/JOE-18-0412
60. Nozaki M, Naito N, Swanson P, Miyata K, Nakai Y, Oota Y, et al. Salmonid Pituitary Gonadotrophs. I. Distinct Cellular Distributions of Two Gonadotropins, GTH I and GTH II. Gen Comp Endocrinol (1990) 77(3):348–57. doi: 10.1016/0016-6480(90)90224-A
61. Weltzien FA, Hildahl J, Hodne K, Okubo K, Haug TM. Embryonic Development of Gonadotrope Cells and Gonadotropic Hormones–Lessons From Model Fish. Mol Cell Endocrinol (2014) 385(1-2):18–27. doi: 10.1016/j.mce.2013.10.016
62. Golan M, Martin AO, Mollard P, Levavi-Sivan B. Anatomical and Functional Gonadotrope Networks in the Teleost Pituitary. Sci Rep (2016) 6:23777. doi: 10.1038/srep23777
63. Hollander-Cohen L, Golan M, Levavi-Sivan B. Differential Regulation of Gonadotropins as Revealed by Transcriptomes of Distinct LH and FSH Cells of Fish Pituitary. Int J Mol Sci (2021) 22(12):6478. doi: 10.3390/ijms22126478
64. Royan MR, Siddique K, Csucs G, Puchades MA, Nourizadeh-Lillabadi R, Bjaalie JG, et al. 3D Atlas of the Pituitary Gland of the Model Fish Medaka (Oryzias Latipes). Front Endocrinol (Lausanne) (2021) 12:719843. doi: 10.3389/fendo.2021.719843
65. Fontaine R, Ager-Wick E, Hodne K, Weltzien FA. Plasticity in Medaka Gonadotropes via Cell Proliferation and Phenotypic Conversion. J Endocrinol (2020) 245(1):21–37. doi: 10.1530/JOE-19-0405
66. Zohar Y, Munoz-Cueto JA, Elizur A, Kah O. Neuroendocrinology of Reproduction in Teleost Fish. Gen Comp Endocrinol (2010) 165(3):438–55. doi: 10.1016/j.ygcen.2009.04.017
67. He W, Dai X, Chen X, He J, Yin Z. Zebrafish Pituitary Gene Expression Before and After Sexual Maturation. J Endocrinol (2014) 221(3):429–40. doi: 10.1530/JOE-13-0488
68. Morais RD, Nobrega RH, Gomez-Gonzalez NE, Schmidt R, Bogerd J, Franca LR, et al. Thyroid Hormone Stimulates the Proliferation of Sertoli Cells and Single Type A Spermatogonia in Adult Zebrafish (Danio Rerio) Testis. Endocrinology (2013) 154(11):4365–76. doi: 10.1210/en.2013-1308
69. Song J, Lu Y, Cheng X, Shi C, Lou Q, Jin X, et al. Functions of the Thyroid-Stimulating Hormone on Key Developmental Features Revealed in a Series of Zebrafish Dyshormonogenesis Models. Cells (2021) 10(8):1984. doi: 10.3390/cells10081984
70. Benedet S, Bjornsson BT, Taranger GL, Andersson E. Cloning of Somatolactin Alpha, Beta Forms and the Somatolactin Receptor in Atlantic Salmon: Seasonal Expression Profile in Pituitary and Ovary of Maturing Female Broodstock. Reprod Biol Endocrinol (2008) 6:42. doi: 10.1186/1477-7827-6-42
71. Rand-Weaver M, Swanson P. Plasma Somatolactin Levels in Coho Salmon (Oncorhynchus Kisutch) During Smoltification and Sexual Maturation. Fish Physiol Biochem (1993) 11(1-6):175–82. doi: 10.1007/BF00004564
72. Mizrahi N, Gilon C, Atre I, Ogawa S, Parhar IS, Levavi-Sivan B. Deciphering Direct and Indirect Effects of Neurokinin B and GnRH in the Brain-Pituitary Axis of Tilapia. Front Endocrinol (Lausanne) (2019) 10:469. doi: 10.3389/fendo.2019.00469
73. Zhao L, Bakke M, Krimkevich Y, Cushman LJ, Parlow AF, Camper SA, et al. Steroidogenic Factor 1 (SF1) Is Essential for Pituitary Gonadotrope Function. Development (2001) 128(2):147–54. doi: 10.1242/dev.128.2.147
74. Ferris HA, Shupnik MA. Mechanisms for Pulsatile Regulation of the Gonadotropin Subunit Genes by GNRH1. Biol Reprod (2006) 74(6):993–8. doi: 10.1095/biolreprod.105.049049
75. Zheng W, Yang J, Jiang Q, He Z, Halvorson LM. Liver Receptor Homologue-1 Regulates Gonadotrope Function. J Mol Endocrinol (2007) 38(1-2):207–19. doi: 10.1677/JME-06-0001
76. Fortin J, Kumar V, Zhou X, Wang Y, Auwerx J, Schoonjans K, et al. NR5A2 Regulates Lhb and Fshb Transcription in Gonadotrope-Like Cells In Vitro, But Is Dispensable for Gonadotropin Synthesis and Fertility In Vivo. PloS One (2013) 8(3):e59058. doi: 10.1371/journal.pone.0059058
77. McCosh RB, Szeligo BM, Bedenbaugh MN, Lopez JA, Hardy SL, Hileman SM, et al. Evidence That Endogenous Somatostatin Inhibits Episodic, But Not Surge, Secretion of LH in Female Sheep. Endocrinology (2017) 158(6):1827–37. doi: 10.1210/en.2017-00075
78. Marques P, Skorupskaite K, George JT, Anderson RA, Rozario KS, Feingold KR. Physiology of GNRH and Gonadotropin Secretion. In: Feingold KR, Anawalt B, Boyce A, Chrousos G, de Herder WW, Dhatariya K, editors. Endotext. South Dartmouth (MA): MDText.com, Inc. (2000).
79. Maguire CA, Song YB, Wu M, Leon S, Carroll RS, Alreja M, et al. Tac1 Signaling Is Required for Sexual Maturation and Responsiveness of GnRH Neurons to Kisspeptin in the Male Mouse. Endocrinology (2017) 158(7):2319–29. doi: 10.1210/en.2016-1807
80. Young J, Metay C, Bouligand J, Tou B, Francou B, Maione L, et al. SEMA3A Deletion in a Family With Kallmann Syndrome Validates the Role of Semaphorin 3A in Human Puberty and Olfactory System Development. Hum Reprod (2012) 27(5):1460–5. doi: 10.1093/humrep/des022
81. Falardeau J, Chung WC, Beenken A, Raivio T, Plummer L, Sidis Y, et al. Decreased FGF8 Signaling Causes Deficiency of Gonadotropin-Releasing Hormone in Humans and Mice. J Clin Invest (2008) 118(8):2822–31. doi: 10.1172/JCI34538
82. Kim T, Lawson MA. GnRH Regulates Gonadotropin Gene Expression Through NADPH/Dual Oxidase-Derived Reactive Oxygen Species. Endocrinology (2015) 156(6):2185–99. doi: 10.1210/en.2014-1709
83. Kim HD, Choe HK, Chung S, Kim M, Seong JY, Son GH, et al. Class-C SOX Transcription Factors Control GnRH Gene Expression via the Intronic Transcriptional Enhancer. Mol Endocrinol (2011) 25(7):1184–96. doi: 10.1210/me.2010-0332
84. Hou H, Uuskula-Reimand L, Makarem M, Corre C, Saleh S, Metcalf A, et al. Gene Expression Profiling of Puberty-Associated Genes Reveals Abundant Tissue and Sex-Specific Changes Across Postnatal Development. Hum Mol Genet (2017) 26(18):3585–99. doi: 10.1093/hmg/ddx246
85. Trarbach EB, Abreu AP, Silveira LF, Garmes HM, Baptista MT, Teles MG, et al. Nonsense Mutations in FGF8 Gene Causing Different Degrees of Human Gonadotropin-Releasing Deficiency. J Clin Endocrinol Metab (2010) 95(7):3491–6. doi: 10.1210/jc.2010-0176
86. Biran J, Golan M, Mizrahi N, Ogawa S, Parhar IS, Levavi-Sivan B. Direct Regulation of Gonadotropin Release by Neurokinin B in Tilapia (Oreochromis Niloticus). Endocrinology (2014) 155(12):4831–42. doi: 10.1210/en.2013-2114
87. Nagae M, Uenoyama Y, Okamoto S, Tsuchida H, Ikegami K, Goto T, et al. Direct Evidence That KNDy Neurons Maintain Gonadotropin Pulses and Folliculogenesis as the GnRH Pulse Generator. Proc Natl Acad Sci USA (2021) 118(5):e2009156118. doi: 10.1073/pnas.2009156118
88. Navarro VM, Bosch MA, Leon S, Simavli S, True C, Pinilla L, et al. The Integrated Hypothalamic Tachykinin-Kisspeptin System as a Central Coordinator for Reproduction. Endocrinology (2015) 156(2):627–37. doi: 10.1210/en.2014-1651
89. Pennefather JN, Lecci A, Candenas ML, Patak E, Pinto FM, Maggi CA. Tachykinins and Tachykinin Receptors: A Growing Family. Life Sci (2004) 74(12):1445–63. doi: 10.1016/j.lfs.2003.09.039
90. Ebner K, Singewald N. The Role of Substance P in Stress and Anxiety Responses. Amino Acids (2006) 31(3):251–72. doi: 10.1007/s00726-006-0335-9
91. Zieglgansberger W. Substance P and Pain Chronicity. Cell Tissue Res (2019) 375(1):227–41. doi: 10.1007/s00441-018-2922-y
92. Simavli S, Thompson IR, Maguire CA, Gill JC, Carroll RS, Wolfe A, et al. Substance P Regulates Puberty Onset and Fertility in the Female Mouse. Endocrinology (2015) 156(6):2313–22. doi: 10.1210/en.2014-2012
93. Ogawa S, Ramadasan PN, Anthonysamy R, Parhar IS. Sexual Dimorphic Distribution of Hypothalamic Tachykinin1 Cells and Their Innervations to GnRH Neurons in the Zebrafish. Front Endocrinol (Lausanne) (2020) 11:534343. doi: 10.3389/fendo.2020.534343
94. Ciani E, Fontaine R, Maugars G, Nourizadeh-Lillabadi R, Andersson E, Bogerd J, et al. Gnrh Receptor Gnrhr2bbalpha Is Expressed Exclusively in Lhb-Expressing Cells in Atlantic Salmon Male Parr. Gen Comp Endocrinol (2020) 285:113293. doi: 10.1016/j.ygcen.2019.113293
95. Golan M, Zelinger E, Zohar Y, Levavi-Sivan B. Architecture of GnRH-Gonadotrope-Vasculature Reveals a Dual Mode of Gonadotropin Regulation in Fish. Endocrinology (2015) 156(11):4163–73. doi: 10.1210/en.2015-1150
96. Somoza GM, Mechaly AS, Trudeau VL. Kisspeptin and GnRH Interactions in the Reproductive Brain of Teleosts. Gen Comp Endocrinol (2020) 298:113568. doi: 10.1016/j.ygcen.2020.113568
97. Golan M, Biran J, Levavi-Sivan B. A Novel Model for Development, Organization, and Function of Gonadotropes in Fish Pituitary. Front Endocrinol (Lausanne) (2014) 5:182. doi: 10.3389/fendo.2014.00182
98. Liu D, Xiong F, Hew CL. Functional Analysis of Estrogen-Responsive Elements in Chinook Salmon (Oncorhynchus Tschawytscha) Gonadotropin II Beta Subunit Gene. Endocrinology (1995) 136(8):3486–93. doi: 10.1210/endo.136.8.7628385
99. Cavaco JE, Bogerd J, Goos H, Schulz RW. Testosterone Inhibits 11-Ketotestosterone-Induced Spermatogenesis in African Catfish (Clarias Gariepinus). Biol Reprod (2001) 65(6):1807–12. doi: 10.1095/biolreprod65.6.1807
100. Hellqvist A, Schmitz M, Borg B. Effects of Castration and Androgen-Treatment on the Expression of FSH-Beta and LH-Beta in the Three-Spine Stickleback, Gasterosteus Aculeatus–Feedback Differences Mediating the Photoperiodic Maturation Response? Gen Comp Endocrinol (2008) 158(2):178–82. doi: 10.1016/j.ygcen.2008.07.004
101. Borg B. Androgens in Teleost Fishes. Comp Biochem Physiol Part C: Pharmacol Toxicol Endocrinol (1994) 109(3):219–45. doi: 10.1016/0742-8413(94)00063-G
102. Crowder CM, Lassiter CS, Gorelick DA. Nuclear Androgen Receptor Regulates Testes Organization and Oocyte Maturation in Zebrafish. Endocrinology (2018) 159(2):980–93. doi: 10.1210/en.2017-00617
103. Sato T, Suzuki A, Shibata N, Sakaizumi M, Hamaguchi S. The Novel Mutant Scl of the Medaka Fish, Oryzias Latipes, Shows No Secondary Sex Characters. Zoolog Sci (2008) 25(3):299–306. doi: 10.2108/zsj.25.299
104. Larsen DA, Swanson P. Effects of Gonadectomy on Plasma Gonadotropins I and II in Coho Salmon, Oncorhynchus Kisutch. Gen Comp Endocrinol (1997) 108(1):152–60. doi: 10.1006/gcen.1997.6958
105. Safian D, Bogerd J, Schulz RW. Regulation of Spermatogonial Development by Fsh: The Complementary Roles of Locally Produced Igf and Wnt Signaling Molecules in Adult Zebrafish Testis. Gen Comp Endocrinol (2019) 284:113244. doi: 10.1016/j.ygcen.2019.113244
106. Muriach B, Cerda-Reverter JM, Gomez A, Zanuy S, Carrillo M. Molecular Characterization and Central Distribution of the Estradiol Receptor Alpha (ERalpha) in the Sea Bass (Dicentrarchus Labrax). J Chem Neuroanat (2008) 35(1):33–48. doi: 10.1016/j.jchemneu.2007.05.010
107. Guzman JM, Luckenbach JA, da Silva DAM, Hayman ES, Ylitalo GM, Goetz FW, et al. Seasonal Variation of Pituitary Gonadotropin Subunit, Brain-Type Aromatase and Sex Steroid Receptor mRNAs, and Plasma Steroids During Gametogenesis in Wild Sablefish. Comp Biochem Physiol A Mol Integr Physiol (2018) 219-220:48–57. doi: 10.1016/j.cbpa.2018.02.010
108. Andersson E, Borg B, Lambert JG. Aromatase Activity in Brain and Pituitary of Immature and Mature Atlantic Salmon (Salmo Salar L.) Parr. Gen Comp Endocrinol (1988) 72(3):394–401. doi: 10.1016/0016-6480(88)90161-X
109. Kazeto Y, Trant JM. Molecular Biology of Channel Catfish Brain Cytochrome P450 Aromatase (CYP19A2): Cloning, Preovulatory Induction of Gene Expression, Hormonal Gene Regulation and Analysis of Promoter Region. J Mol Endocrinol (2005) 35(3):571–83. doi: 10.1677/jme.1.01805
110. Paez-Pereda M, Kuchenbauer F, Arzt E, Stalla GK. Regulation of Pituitary Hormones and Cell Proliferation by Components of the Extracellular Matrix. Braz J Med Biol Res (2005) 38(10):1487–94. doi: 10.1590/S0100-879X2005001000005
111. Paez Pereda M, Ledda MF, Goldberg V, Chervin A, Carrizo G, Molina H, et al. High Levels of Matrix Metalloproteinases Regulate Proliferation and Hormone Secretion in Pituitary Cells. J Clin Endocrinol Metab (2000) 85(1):263–9. doi: 10.1210/jc.85.1.263
112. Liu YZ, Guo YF, Wang L, Tan LJ, Liu XG, Pei YF, et al. Genome-Wide Association Analyses Identify SPOCK as a Key Novel Gene Underlying Age at Menarche. PloS Genet (2009) 5(3):e1000420. doi: 10.1371/journal.pgen.1000420
113. Hui P, Xu X, Xu L, Hui G, Wu S, Lan Q. Expression of MMP14 in Invasive Pituitary Adenomas: Relationship to Invasion and Angiogenesis. Int J Clin Exp Pathol (2015) 8(4):3556–67.
114. Pan Y, Han C, Wang C, Hu G, Luo C, Gan X, et al. ADAM10 Promotes Pituitary Adenoma Cell Migration by Regulating Cleavage of CD44 and L1. J Mol Endocrinol (2012) 49(1):21–33. doi: 10.1530/JME-11-0174
115. Aroua S, Maugars G, Jeng SR, Chang CF, Weltzien FA, Rousseau K, et al. Pituitary Gonadotropins FSH and LH Are Oppositely Regulated by the Activin/Follistatin System in a Basal Teleost, the Eel. Gen Comp Endocrinol (2012) 175(1):82–91. doi: 10.1016/j.ygcen.2011.10.002
116. Poon SK, So WK, Yu X, Liu L, Ge W. Characterization of Inhibin Alpha Subunit (Inha) in the Zebrafish: Evidence for a Potential Feedback Loop Between the Pituitary and Ovary. Reproduction (2009) 138(4):709–19. doi: 10.1530/REP-09-0198
117. Burger LL, Dalkin AC, Aylor KW, Haisenleder DJ, Marshall JC. GnRH Pulse Frequency Modulation of Gonadotropin Subunit Gene Transcription in Normal Gonadotropes-Assessment by Primary Transcript Assay Provides Evidence for Roles of GnRH and Follistatin. Endocrinology (2002) 143(9):3243–9. doi: 10.1210/en.2002-220216
Keywords: puberty, pituitary, Atlantic salmon, follicle-stimulating hormone, transcriptomics
Citation: Crespo D, Skaftnesmo KO, Kjærner-Semb E, Yilmaz O, Norberg B, Olausson S, Vogelsang P, Bogerd J, Kleppe L, Edvardsen RB, Andersson E, Wargelius A, Hansen TJ, Fjelldal PG and Schulz RW (2022) Pituitary Gonadotropin Gene Expression During Induced Onset of Postsmolt Maturation in Male Atlantic Salmon: In Vivo and Tissue Culture Studies. Front. Endocrinol. 13:826920. doi: 10.3389/fendo.2022.826920
Received: 01 December 2021; Accepted: 17 February 2022;
Published: 15 March 2022.
Edited by:
Abdel Halim Harrath, King Saud University, Saudi ArabiaReviewed by:
Juan Ignacio Fernandino, Institute of Biotechnological Research (IIB-INTECH)(CONICET), ArgentinaCopyright © 2022 Crespo, Skaftnesmo, Kjærner-Semb, Yilmaz, Norberg, Olausson, Vogelsang, Bogerd, Kleppe, Edvardsen, Andersson, Wargelius, Hansen, Fjelldal and Schulz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Diego Crespo, ZGllZ28uY3Jlc3BvQGhpLm5v
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.