 Nicole Mohajer
Nicole Mohajer Chrislyn Y. Du2
Chrislyn Y. Du2 Bruce Blumberg
Bruce Blumberg- 1Deparment of Pharmaceutical Sciences, University of California, Irvine, CA, United States
- 2Deparment of Developmental and Cell Biology, University of California, Irvine, CA, United States
- 3Deparment of Biomedical Engineering, University of California, Irvine, CA, United States
Adult and childhood obesity have reached pandemic level proportions. The idea that caloric excess and insufficient levels of physical activity leads to obesity is a commonly accepted answer for unwanted weight gain. This paradigm offers an inconclusive explanation as the world continually moves towards an unhealthier and heavier existence irrespective of energy balance. Endocrine disrupting chemicals (EDCs) are chemicals that resemble natural hormones and disrupt endocrine function by interfering with the body’s endogenous hormones. A subset of EDCs called obesogens have been found to cause metabolic disruptions such as increased fat storage, in vivo. Obesogens act on the metabolic system through multiple avenues and have been found to affect the homeostasis of a variety of systems such as the gut microbiome and adipose tissue functioning. Obesogenic compounds have been shown to cause metabolic disturbances later in life that can even pass into multiple future generations, post exposure. The rising rates of obesity and related metabolic disease are demanding increasing attention on chemical screening efforts and worldwide preventative strategies to keep the public and future generations safe. This review addresses the most current findings on known obesogens and their effects on the metabolic system, the mechanisms of action through which they act upon, and the screening efforts through which they were identified with. The interplay between obesogens, brown adipose tissue, and the gut microbiome are major topics that will be covered.
Obesity Is a Serious Problem
Obesity has become a present-day pandemic affecting people of all ages across the world. According to the World Health Organization, the prevalence of global obesity has nearly tripled since 1975 with a continued upward trajectory (1). In 2016, the WHO reported more than 1.9 billion adults as overweight, with 650 million of those adults as obese. The prevalence of obesity in children has continued to rise in the U.S alone, despite the nation’s efforts to promote better nutrition practices and increase physical activity levels in the educational system (2). In 2019, a staggering 38.2 million children under the age of 5 were reported as overweight or obese, worldwide (1). Comorbidities associated with obesity affect nearly all physiological systems and lead to serious health complications including mortality and a lowered quality of life. Obesity contributes to a growing list of health complications including insulin resistance, cardiovascular diseases, airway dysfunctions, metabolic syndrome, kidney disease, osteoarthritis, skin diseases, reproductive disorders, and cancer (3, 4) and death from COVID-19 (5). In addition to physiological comorbidities, the burden of obesity affects the individual’s psychological well-being, leading to higher stress and depression. Obesity often presents together with depression and negative self-image in both children and adults, creating a vicious cycle where the conditions potentiate each other (3, 6). Those who suffer from depression are 58% more likely to develop obesity, and those who are obese are 55% more likely to develop chronic depression (7). Obesity makes it less likely for students to stay in school past the 12th grade, independently of their parent’s socioeconomic status (8). Similarly, lower education levels have been linked to higher weight gain and obesity (9). Obesity places a financial and emotional burden on individuals, their families, and the nation at large when loss of productivity and loss of work is considered. The CDC reported the national obesity-related cost to be $147 billion in 2008, however, more recent data from 2014 estimates the cost of obesity and its comorbidities to be closer to $2 trillion dollars (10, 11). It is estimated that the annual cost of obesity in the U.S will rise $48-66 billion each year throughout 2030 (4). Therefore, the severe consequences of obesity on both individual and population-level health demand that urgent attention be paid to this worsening pandemic.
Obesity Is More Than Calories In/Out
Obesity is a multifaceted disease, and its etiology remains widely misunderstood. Weight gain has primarily been blamed on high calorie diet and a sedentary lifestyle. Many types of fad diets have shown short-lived improvements in body weight, but the overall success for long-term weight loss through caloric restriction remains inefficacious and the global prevalence of obesity continues to rise. Recent research has highlighted the shortcomings of the energy balance, or “calories in versus calories out” paradigm of weight management. The idea that people must consume less calories than they burn in order to lose weight is self-evident, but is no longer an all-inclusive explanation for the increasing rate of obesity and long-term weight gain.
Some lower income countries have reported a decrease in exercise, other higher income countries, such as the U.S., have reported a consistent or even increased level of exercise over the last 30 years despite the continuous rise in obesity (12, 13). If the population is gaining weight despite recommended physical activity levels, then the problem must also include the nature of the foods ingested as well as energy expenditure. We must consider the quality of the calorie being consumed since not all calories are created equal. The quality of the calorie, and whether it is coming from healthy foods or unhealthy foods, influences the types of food we should and shouldn’t eat and how our bodies metabolize those calories for maximal benefit. Primates who were given calorically equal meals that only differed in the percentage of cis or trans-fats showed a disparity in weight gain after six years, with the trans-fat group showing an increase in visceral fat (14). The composition of our diets, more so than the caloric count of our daily diets, affects hormonal imbalances, metabolic efficiency, epigenetics, gut health, and fat accumulation (15). According to the carbohydrate-insulin model of obesity, the way we metabolize processed carbohydrates and foods that are higher on the glycemic index (such as starchy, refined, and sugary foods) promotes fat storage in fat cells and is driven by spikes in insulin levels (16). Therefore, eating the same number of calories in candy vs Brussel sprouts will be processed, metabolized, and stored in very different ways. Taken together, the current caloric models of obesity and weight gain are insufficient as stand-alone explanations for the sudden increase in global obesity over the past few decades.
EDCs and Obesogens
While environmental, nutritional, and socioeconomic factors may all contribute to weight gain, there are other components of our immediate environment that offer a more in-depth explanation for the etiopathology of obesity. Increasing evidence has linked chemical exposure, ingestion, and inhalation of industrial compounds to obesity and other metabolic and endocrine related diseases. As the world modernizes, more chemicals pollute our food, water, air, and soil, making exposure unavoidable. Endocrine disrupting chemicals (EDCs) are chemicals or mixtures that disrupt endocrine function and interfere with the body’s endogenous hormones (17). EDCs are structured like and act similarly to natural hormones and disrupt homeostasis by binding to hormone receptors (18). Naturally found EDC’s include plant phytoestrogens such as those in soy-based foods and dairy products. Phytoestrogens can behave as endocrine disruptors by affecting estrogen receptor-mediated pathways (19). Synthetic EDCs are found in common industrial products such as pesticides, fungicides, flame retardants, plastics, food wrappers, solvents, and metals. Both in vitro and in vivo studies have shown that synthetic EDCs exert effects on multiple systems including the reproductive system, the central nervous system, the immune system, and on metabolic function (20). In addition to endocrine pathways there are many non-hormonal cellular signaling pathways that could potentially be disrupted by chemical exposures. The concept of “signal toxicity” has been developed to account for this potential disruption of the thousands of cellular signaling pathways that could be targeted (21). Relevant examples of signal toxicity include disruption of neurotransmitter signaling, growth factor signaling pathways, receptor kinase signaling, etc. These should not be ignored in the developing discussion about environmental chemicals and obesity.
While EDCs can affect multiple physiological systems, recent research has placed much needed focus on chemicals that might be associated with the rising rates of metabolic syndrome and obesity. A subset of EDCs act as obesogens – chemicals that lead to increased fat storage, in vivo after exposure [reviewed in (22–24)]. The environmental obesogen model proposes that obesogens cause greater susceptibility to weight gain, lipid storage, and energy imbalances that lead to obesity (25). In 2015, the Parma consensus broadened the definition of obesogens to include EDCs that affect other obesity related metabolic conditions that drive metabolic syndrome, such as insulin resistance, hypertension, dyslipidemia, and hyperglycemia (26). This class of EDCs was denoted as metabolism disrupting chemicals (MDCs) [reviewed in (26, 27)]. Many chemical obesogens have been identified and numerous reviews have been written about them in recent years (22–24, 28). The total number of obesogens is currently unknown because no systematic attempt has been undertaken to identify them. This review will identify recent findings on possible obesogens, their effects on metabolism and lipid dysregulation and the mechanisms through which they act.
Adipogenesis, Nuclear Receptors, and Key Pathways
The obesogen hypothesis holds that exposure to obesogenic chemicals will lead to increased white adipose tissue (WAT) mass. Adipogenesis is the cellular process by which pluripotent stem cells or preadipocytes commit their fate to differentiating into adipocytes (29). WAT can be found subcutaneously or viscerally, and too much WAT can result in in excess lipid storage, altered adipocyte homeostasis, the disruption of energy balance, and changes in metabolic set points [reviewed in (30)]. In healthy individuals, WAT plays an important role in metabolism and energy homeostasis throughout the body. However, people with obesity and type two diabetes (T2D) experience an inflammatory response in their adipose tissue, particularly in visceral white fat that contains higher levels of reactive oxidative species (29, 31). WAT differentiation and cell functioning is primarily controlled by the peroxisome proliferator-activated receptor gamma (PPARγ), also known as the “master” regulator of adipogenesis (32). PPARγ is a ligand-activated transcription factor which is responsible for the growth and development of adipose tissue and acts as the receptor for antidiabetic drugs such as rosiglitazone (32). Some obesogenic EDCs can bind to PPARγ, creating downstream effects that influence multipotent mesenchymal stromal stem cells (MSCs) to favor the adipogenic pathway. EDCs can bind to other nuclear receptors as well, including estrogen, androgen, and progesterone receptors, thyroid receptors, and retinoid X receptors (18). EDC action is not limited to nuclear receptors; EDCs can also bind to nonnuclear receptors and nonsteroid receptors (18). Orphan nuclear receptors, such as estrogen related receptor alpha and estrogen related receptor beta, have been found to play a role in metabolic disease, weight gain, and obesity when exposed to EDCs such as Bisphenol AF (18, 33, 34). When EDCs bind receptors that are meant to regulate vital cellular functions and cell signaling, major health consequences can arise, disrupting homeostatic mechanisms and correct development.
in Vitro Assays for Obesogens
There are many in vitro models that can be used to assess the potential obesogenic properties of chemicals. A list of in vitro model systems and obesogens identified using these models is presented in Table 1. When using non-human cell types as obesity models, it is important to understand that the translational application to humans might sometimes be limited by differences between species (53). The first studies on adipogenesis and obesogens occurred in the early 2000’s on mouse 3T3-L1 cells derived from 3T3 cells [reviewed in (30)]. 3T3-L1 is a well-established preadipocyte cell line derived from 17–19-day old mouse embryos and has a fibroblastic morphology that can be readily induced into adipocyte differentiation (54, 55). To differentiate 3T3-L1 cells into adipocytes, the cells were treated with a minimum level of an adipogenic cocktail that often includes insulin, dexamethasone, and 3-isobutyl-1-methylxanthine (56). The benefits of this cell type include its ease of culture and cost effectiveness compared to mature adipocytes and other primary cells. 3T3-L1 cells have been used for toxicogenomic studies aimed at evaluating the efficacy of screening for obesogens (57). Although these cells can maintain high stability in transcription patterns, they produce differing lipid accumulation levels between tested compounds which can interfere with the interpretation of the mechanistic possibilities (57). 3T3-L1 cells are also highly sensitive to small perturbations in assay conditions such as brand of plastic plates, batches of bovine sera, origin and passage number of cells, and density at induction, all of which can impact their utility (58).

Table 1 In vitro model systems and associated obesogens.
Another cell type that has been used for a similar purpose is the C3H10T½ cell line, which was developed in 1973 from mouse embryonic stem cells (59). These cells can differentiate into various mesodermal cell types including myocytes, chondrocytes, and adipocytes. C3H10T½ cells exhibit a fibroblast like morphology resembling multipotent MSCs, leading some investigators to mistakenly refer to these cells as bona fide MSCs. The primary applications for C3H10T½ cells have centered around evaluating the impact of compounds on adipogenesis and the molecular mechanisms underlying adipogenic differentiation (60). Notable characteristics of these cells are their ability to maintain a relatively homogenous population of multipotent stem cell-like cells and their usefulness in assessing adipocyte commitment and differentiation.
The OP9 mouse stromal cell line was developed from the calvaria of newborn mice that were genetically modified to be deficient in macrophage colony stimulating factor. This bone marrow derived stromal cell line is known for its ability to collect large amounts of triglyceride droplets when stimulated towards adipogenesis after 72 hours, allowing these cells to be an appropriate model for rapidly screening chemicals for adipogenic effects (61, 62). A clonal derivative denoted as OP9-K cells was later developed that could differentiate rapidly and reproducibly. OP9-K cells are readily transfected with an efficiency of ~80% and were validated as a model system for microarray analysis of the differentiated transcriptome (63). In comparison with 3T3-L1 and mouse bone-marrow derived MSCs, OP9 cells were more sensitive to the induction of adipogenesis by chemicals known to activate PPARγ and RXR (51, 58). The OP9 cell line appears to be a promising in vitro model to study adipogenesis using mouse cell lines.
Multipotent mesenchymal stromal stem cells, also known mesenchymal stem cells (hMSCs) have been used to assess possible metabolic disruptors in vitro (64, 65). MSCs are used as an alternative to human pre-adipocytes which have reduced proliferative ability and can exhibit physiological differences related to the fat depot of origin within the body (66). MSCs are bona fide, multipotent precursors of a variety of cell types including adipocytes, chondroblasts, osteoblasts, and hematopoietic‐supporting stromal cells (67). MSCs can simultaneously express genes characteristic of various mesenchymal cell lineages while also maintaining osteogenic and adipogenic potential in vitro (68). A key advantage of the MSC model is that two important parameters of adipogenesis can be evaluated: commitment of stem cells to pre-adipocytes and the differentiation of pre-adipocytes to mature adipocytes (69, 70). The ability to evaluate both endpoints make MSCs a favorable system to study adipogenesis, although they are currently less utilized than the preadipocyte models such as 3T3-L1 cells. Using human MSCs may aid in understanding the effects of contaminants in humans as well as facilitate translational efforts (65).
In Vivo Assays for Obesogens
In vivo models allow the study of chemical effects on complex organisms in which multiple systems must work simultaneously in a natural physiological environment. Using in vivo models is a valuable tool in determining if and how chemicals act as EDCs in a way that is more translatable to how EDCs may act in humans. Various in vivo model systems and obesogens identified using these models are listed in Table 2. Rodents, particularly mice have been a very widely used model to study the effects of chemical exposures and infer possible effects in humans. Mouse models have allowed more focus to be placed on the developmental origins of disease, which aids in understanding the origins of chronic and adult-onset diseases. Mouse models have also allowed for the discovery of obesogens and endocrine disruptors.
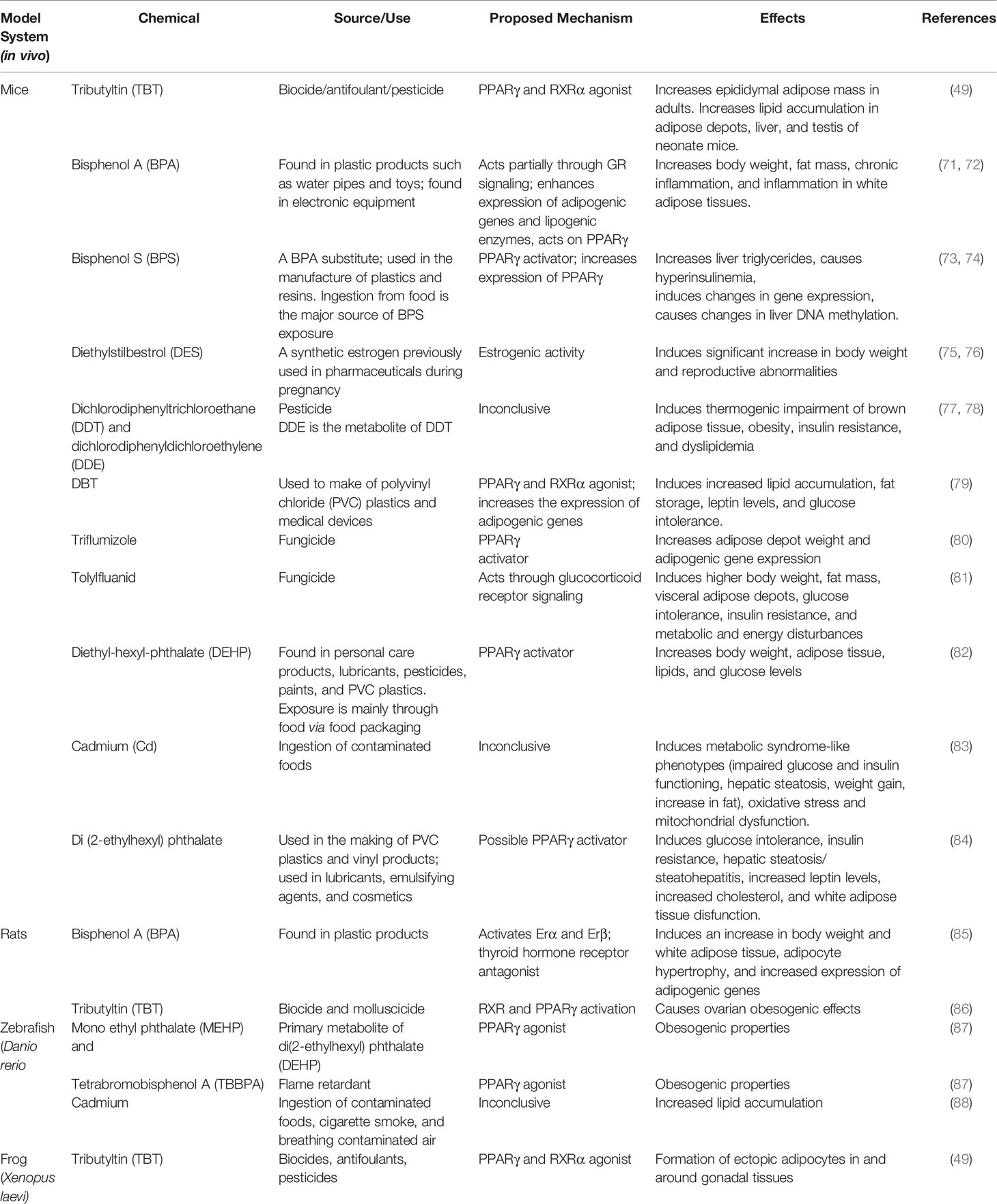
Table 2 In vivo model systems and associated obesogens.
Diethylstilbestrol (DES), a chemical that was once prescribed to prevent miscarriage, was discovered to be a dangerous EDC that causes lifelong health issues (75). DES was identified as a chemical that increased adiposity, in vivo, presumably as a result of its action on the estrogen receptor (76).
Mouse models also made it possible to show that tributyltin chloride (TBT) not only caused adipocyte differentiation in cell models but had adipogenic effects in vivo. TBT studies began with in vitro models using 3T3-L1 cell lines and were later extended into animal models. Triphenyltin and TBT were discovered to be activators of PPARγ and retinoid X receptor (RXR) via a preadipocyte 3T3-L1 cell model (42). A contemporaneous study showed that TBT could elicit adipocyte differentiation in 3T3-L1 cells and fat accumulation in mice treated prenatally (49). Dibutyltin (DBT) is an organotin used as a heat stabilizer in polyvinyl chloride (PVC) plastics and is also a metabolite of TBT. DBT was shown to be a PPARγ and RXR activator, in vitro, and inducer of adipogenesis in 3T3-L1 cells and in human and mouse MSCs (40, 41). Perinatal exposure of pregnant C57BL6/J mouse dams led to increased leptin levels, glucose intolerance, and increased fat storage in adulthood, confirming DBT’s obesogenic effects in a complex organism (40). Common fungicides have also become classified as obesogenic EDCs after being screened using mouse models. Triflumizole was found to be a PPARγ activator in vitro, and when tested in gestating female mice, it was found to increase adipose depot weight and shift MSC fate to favor adipogenesis (80). Another common fungicide, tolylfluanid, was shown to be a glucocorticoid receptor activator in vitro, and obesogen in mice (81).
Rodent studies have been used to confirm the obesogenic action of a number of plastic monomers, plasticizers and other additives [reviewed in 27](Table 2). More recent studies testing the safety of BPS, a BPA substitute used in the manufacturing of plastics, have used in vivo mouse models to determine if BPS exhibits obesogenic properties by affecting gene expression and DNA methylation. It was found, for the first time, that even low doses of BPS acted as an in vivo obesogen and caused epigenetic changes in genes related to metabolism (73). BPA and BPS were found to target PPARγ in human macrophages and were confirmed to cause metabolic abnormalities through PPARγ in mouse models (74). Heavy metal exposure was shown to be obesogenic in mice [reviewed in 27]. CD-1 female mice that were exposed to cadmium via their drinking water from gestation to postnatal day 10 showed delayed obesogenic properties in female offspring, revealing that cadmium exposure can contribute to obesity later in life (83). This has particular relevance for populations living in the desert southwest of the US where heavy metal exposure via dust is prevalent.
It is important to note that some of the effects of early life obesogen exposure can be transmitted to future generations. When adult male or female animals are exposed to a chemical, they (the F0) are exposed as are germ cells within the animals (F1) generation. Effects observed in the F2 generation and beyond are considered transgenerational because these generations were not exposed to the chemical (89). In contrast, exposure to a gestating female mammal will elicit direct effects in the F0 (adult), F1 (embryo) and F2 (germ cells in the embryo) generations. Therefore, the F3 generation is the first not to be exposed and effects seen in F3 and beyond are transgenerational. This topic has been reviewed extensively in recent years and will not be discussed further here for brevity (23, 90–92).
While rodents have been the primary model used to study EDCs, less complex organisms have also proven to be valuable models in studying chemical exposure (Table 2). The adipogenic pathways taking place in less complex organisms also show interspecies similarities to those in higher organisms while the assays themselves are less expensive, shorter, and higher throughput. Zebrafish larvae have been used as an exposure model to test the effects of obesogen exposure on lipid accumulation via Oil red-O staining and to activate PPARγ. Test compounds such as the halogenated BPA analog tetrabromobisphenol-A (TBBPA) were found to induce zebrafish larval lipid accumulation (93). Exposure to environmentally relevant TBT concentrations resulted in adipocyte hypertrophy within only hours after exposure in zebrafish larvae, demonstrating the potency of TBT toward adipogenic endpoints (94).
Other species have also been used to screen for potential obesogens in vivo, such as the African clawed frog, Xenopus laevis (Table 2). X. laevis larvae were used to create a PPARγ reporter model to compare the metabolic capacity to those in mammals. A transgenic approach was used to express both human PPARγ and a series of PPARγ Response Element (PPRE)-eGFP reporter genes simultaneously (87). This approach enabled the detection of PPARγ activators using an in vivo context.
The effects of MDCs can even be studied using invertebrate models to further assess the underlying mechanisms through which chemicals alter lipid homeostasis. These included species like the fruit fly, Drosophila melanogaster, the water flea, Daphnia magna, and the roundworm, Caenorhabditis elegans. Invertebrates possess organ systems that allow nutrient uptake, storage, and energy metabolism through forms like glycogen and lipids (95). Since many intermediary pathways of metabolism are conserved, invertebrates can be a valuable model system for measuring metabolic change. For example, exposure of D. melanogaster to the plasticizer, dibutyl phthalate (DBP) led to increased lipid storage, starvation resistance, hyperglycemia, and hyperphagia in males via evolutionarily conserved insulin and glucagon-like signaling pathways (96). Long term parental exposure of D. melanogaster to a typical plasticizer, Bis(2-ethylhexyl) phthalate (DEHP), elicited significant change in body weight of offspring. The specific changes depended on the exposure period, dose, and gender of the exposed parent. Paternal DEHP treatment resulted in increased body weight of male offspring, whereas maternal exposure led to weight loss in male offspring (97). Exposure of a variety of aquatic invertebrates (coral, rotifers, copepods, octopus, scallop, crab, urchins, and worms) to environmental chemicals (PBDEs, phthalates, organotins or nanoparticles) led to alterations in the expression of genes important for de novo lipogenesis, fatty acid modification and triacylglycerol synthesis (98). BDE-47 increased de novo lipogenesis in the copepod, Tigriopus japonicus after only 24 hours of exposure (99). Exposure of the water flea, Daphnia magna to known mammalian obesogens including TBT and BPA enhanced fat storage, whereas exposure to DEHP or triphenyltin impaired growth and reduced fat storage (100). Experiments such as these confirm the value of less complex model organisms to identify conserved mechanisms underlying metabolic disease.
Thermogenic Adipocytes and EDCs
Broadly speaking, there are two major types of adipose tissue found in the human body: white adipose tissue (WAT), which primarily stores lipids and is maintained throughout adulthood, and thermogenic brown adipose tissue (BAT) which “burns” lipids and is primarily found in newborns and infants. BAT was previously thought to be non-existent or very minimal in adult humans but there are indeed brown fat depots found in adults (101). CT and PET-CT scans revealed multiple locations of BAT within the adult body. Brown adipose tissue is found both subcutaneously and viscerally. The major BAT depot in adult humans is subcutaneous in the supraclavicular region with smaller deposits under the clavicles and in the axilla (102). Visceral BAT can be perivascular, perivisceral and around solid organs such as the pancreas, kidney, liver and spleen (102). White adipocytes make up most of our body fat mass and form the visceral and subcutaneous fat tissues that store energy in the form of triglycerides. White adipose cells typically contain a large unilocular lipid droplet while brown adipocytes contain smaller, multilocular droplets that are rich in mitochondria (103). Brown adipocytes exhibit thermogenic activity when uncoupling proteins such as UCP1 are activated in response to environmental stimuli, mainly exposure to cold temperatures (104). Due to their large number of mitochondria, brown adipocytes act as energy generators rather than energy storers and burn calories as heat is expended. In addition to these distinct types of fat cells, a third, hybrid type of fat known as beige or brite adipose tissue, can form past infancy into adulthood. This process, known as “browning” or “beiging”, occurs as WAT is exposed to stimuli such as cold temperatures, catecholamines, physical activity, or thiazolidinediones, transforming them into brown-like, mitochondria rich, thermogenic adipocytes (105).
The discovery that brown and beige adipocytes exist in adults has raised increased interest in possible therapeutic strategies to treat obesity and type 2 diabetes (T2D) through the browning of white adipose tissue. Abundance of brown adipose tissue is associated with lower levels of metabolic disorders such as T2D, and its presence is associated with the improvement of insulin resistance (106). Increased BAT resulting from 10 days of cold exposure in 14-15°C resulted in a 43% increase in insulin sensitivity in people with T2D (107). Enhancing the formation or function of thermogenic adipocytes appears to be a promising key in the future treatment of obesity and related metabolic diseases (108, 109).
Since brown and beige adipose tissues are important for preventing obesity and T2D, interest is growing in the effects of EDCs on the formation and function of these valuable forms of thermogenic fat cells (110). Below we discuss some recent findings in this area. A list of obesogens associated with the disruption of thermogenic fat and adipose tissue is presented in Table 3.
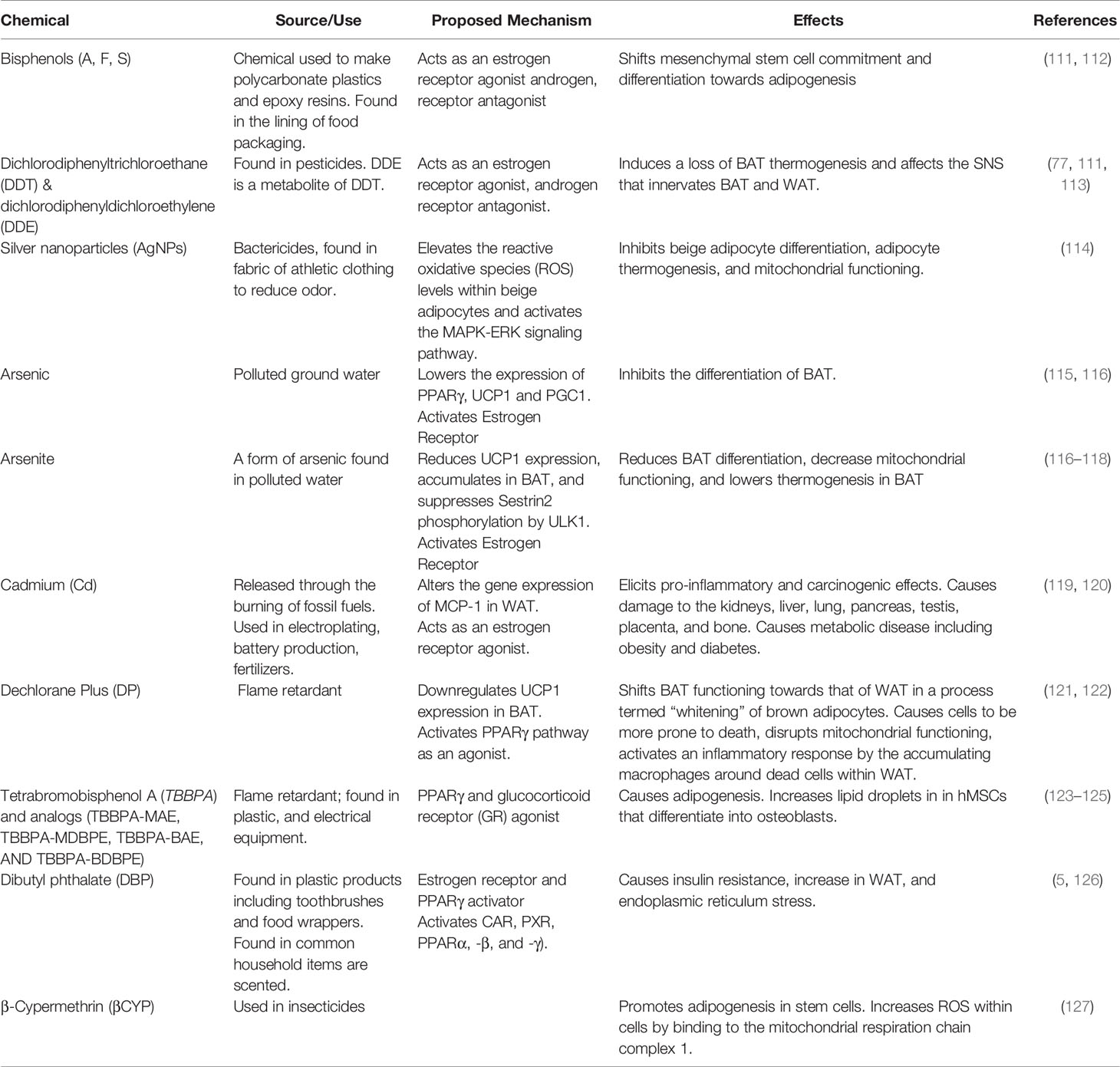
Table 3 Chemical obesogens and their effects on thermogenic fat and adipose tissue.
Bisphenols
The potential of BPA and its analogs as obesogens and their effects on brown thermogenic fat have become a topic of recent interest since many products labeled as “BPA free” likely contain its analog substitutes BPS and BPF. These compounds are found in products containing polycarbonate plastics and epoxy resins (including adhesives, plastics, paint, and sealants). BPS and BPF were shown to be as hormonally active as BPA, acting as estrogen receptor agonists and androgen receptor antagonists in vitro and in vivo [reviewed in (128)]. As obesogens, BPS and BPF elicited adipogenic differentiation in mouse preadipocytes and promoted the proliferation of fat cells, causing an increase in body mass (129). BPS and BPF exhibited obesogenic effects on human adipose-derived mesenchymal stromal stem cells (hADSC) in a dose dependent manner ranging from 0.01 to 25 μM (130). Pregnant F0 C57BL6/J mouse dams were exposed to a human-equivalent dose of BPS (1.5 μg/kg bw/day) throughout pregnancy and lactation. F1 pups were fed a high fat diet (HFD) over the next 15 weeks. Body weight was monitored weekly and body fat measured at euthanasia. Similar analyses were performed on the F2 and F3 generation offspring (131). Findings revealed interesting sex-dependent multigenerational effects, with multigenerational obesogenic effects found in both males and females, yet transgenerational effects only found in females, indicating that BPS is a likely sex-dependent obesogen. More discussion about differing effects on male and females can be found under sexual dimorphisms. Interestingly, BPA, BPS, and BPF have all been linked to a downregulation of the gene encoding micro RNA 26 (miR-26a) from in vitro studies. MiR-26a and miR-26b were shown to be key genetic regulators of the adipocyte browning process (132, 133). Both miR-26a and b are critical for adipogenesis and promote cellular pathways involving energy expenditure, mitochondrial formation, and the upregulation of uncoupling protein-1 (UCP-1) perhaps the key protein in thermogenesis (133).
Dichlorodiphenyltrichloroethane and Dichlorodiphenyldichloroethylene
Dichlorodiphenyltrichloroethane (DDT) and dichlorodiphenyl-dichloroethylene (DDE) are two widespread organochlorine compounds that have been shown to affect thermogenic BAT and WAT in mouse models (134). DDT is a pesticide that is now banned in the United States, however, it remains in use in other parts of the world as a mechanism to control the spread of malaria (113). Both are known to collect in adipose tissue due to their lipophilicity. DDT and its metabolites such as DDE bioaccumulate up the food chain. DDT and DDE act as nuclear estrogen receptor agonists (DDT), androgen receptor antagonist (DDE), and can also bind to certain GPCRs, which can cause alter estrogen signaling (111). DDT has been shown in mouse models to affect the sympathetic nervous system that innervates both BAT and WAT. The sympathetic nervous system controls thermogenesis in brown fat and is needed for the beiging of WAT (77, 135). Dual administration of DDT and DDE or single treatment with DDE induced a loss of thermogenic abilities in BAT in adult female mice by reducing their BAT sympathetic innervation and regulation (77).
Dechlorane Plus
The polychlorinated flame retardant, Dechlorane Plus (DP), is now known to impair BAT functioning and metabolic processes. DP has been found in humans in levels from 0.1 μg/kg to 1000 μg/kg, most concerningly in breast milk and umbilical cord serum (121). DP is a known PPARγ agonist, however, recent studies have shown that DP may also act via other pathways. DP exposure in vitro has shown an upregulation of adipogenic markers in the presence of a PPARγ antagonist, suggesting there could be an alternate pathway that DP may be acting on, independent of PPARγ (121). DP disrupted the function of BAT by down-regulating the expression of UCP1 mRNA, increasing lipid accumulation and disrupting mitochondrial functioning (121). These data were interpreted to indicate that BAT had been “whitened” (121). The whitening of BAT caused the affected cells to become more prone to cell death and activated an inflammatory response via the accumulation of macrophages around the dead cells within WAT (122). DP treatment led to WAT hypertrophy and dysfunction, in part by the inhibition of insulin signaling.
Dibutyl Phthalate
Dibutyl phthalate (DBP) is a plasticizer found in plastic products such as toothbrushes, food wrappers, and in common household items as a fragrance-enhancing additive. DBP is a known EDC and obesogen that can affect fat accumulation and metabolic processes. DBP activates multiple receptors including the estrogen receptor, constitutive androstane receptor (CAR), the pregnane X receptor (PXR), and peroxisome proliferator-activated receptor subtypes (PPARα, -β, and -γ), which regulate the expression of genes encoding metabolic enzymes (126). Pregnant mouse dams were exposed to DBP from the 12th day of gestation to one week after birth. 4- to 5-month-old mice from the DBP group exhibited higher body weight, lower expression of UCP1, insulin insensitivity, greater endoplasmic reticulum stress, and levels of inguinal and epididymal WAT that were twice as high as controls (136). DBP exposure caused insulin resistance, suggesting the presence of a pre-diabetic condition which is often a comorbidity in obese individuals. Lower UCP1 levels impair the production of heat by thermogenic tissues and increased endoplasmic reticulum stress, greatly affecting mitochondrial function (136). Offspring of DBP-treated mice also showed significantly higher levels of binding immunoglobulin protein (Bip), and CCAAT/enhancer-binding protein homologous protein (Chop), which are two markers of ER stress that were associated with lower UCP1 levels. Offspring of DBP-treated mice appeared to be obesity prone via the inhibition of UCP1 caused by ER stress in adipose tissue after protein extraction and western blot analysis revealed that Bip and Chop were increased compared to controls (136).
Tetrabromobisphenols
Brominated chemicals such as TBBPA are used in the production of plastics and electronics to reduce flammability as a safety precaution. However, TBBPA, along with its structural analogues TBBPA-MAE, TBBPA-MDBPE, TBBPA-BAE, AND TBBPA-BDBPE were shown to promote adipogenesis in 3T3-L1 cells by activating PPARγ and glucocorticoid receptors (137, 138), with TBBPA-MAE, and TBBPA-MDBPE acting as stronger promoters of adipogenesis than TBBPA itself (123). hMSCs were exposed to a mixture of TBBPA and TCDD (2,3,7,8-Tetrachlorodibenzo-p-dioxin) to mimic a more realistic scenario of chemical mixtures found in the natural environment. It was found that TBBPA alone predisposed hMSCs to differentiate into adipocytes and increased levels of PPARγ mRNA. In contrast, when TCDD was administered alone, it inhibited the differentiation process and PPARγ mRNA expression. However, when dually administered, TBBPA overrode TCDD’s inhibitory properties (124). Interestingly, TBBPA also increased the number of lipid droplets in hMSCs that differentiated into osteoblasts. While lipids in the bone marrow are an essential component of bone health, too many lipids can result in the impairment of stem and progenitor cell function (124, 125).
β-Cypermethrin
β-Cypermethrin (βCYP) is a widely used pyrethroid, a class of chemicals commonly used as insecticides. Like other pyrethroids (synthetic relatives of the naturally occurring pesticide pyrethrin), βCYP is an EDC and has been found in humans. High concentrations of βCYP (concentrations of 25, 50, and 100 μM) promoted adipogenesis in 3T3-L1 cells (127). Mechanistically, β-CYP acts on 3T3-L1 by increasing the reactive oxygen species within cells via binding to the mitochondrial respiration chain complex 1. This disruption in the complex reduces the mitochondrial membrane potential, which is required for the browning of WAT, increased autophagy, and the miR-34a mediated polarization of macrophages to M2 cells. While He et al. reported a βCYP mediated downregulation of intracellular and extracellular miR-34a in 3T3-L1 cells, upregulation and increased expression of miR-34a has also been linked to obesity (139). The upregulation of miR-34 decreased SIRT1 expression, reduced NAMPT expression which in turn decreased NAD+ levels, which are crucial to cellular health, redox reactions, metabolism, and the browning of WAT (140, 141). NAMPT is important for the synthesis of NAD+ and is known to decrease in aging and obesity, leading to lower levels of NAD+ (139). Notably, an increase in NAD+ was also found in 3T3-L1 murine preadipocytes that differentiated into adipocytes after in vitro exposure to the obesogen, monoethylhexyl phthalate (142). It will be interesting to fully understand the role of miR-34a in obesity and how this is affected by pyrethroid expression.
Metals
Heavy metals have been found to play a role in the disruption of the metabolic system including WAT, beige, and BAT imbalances. Cadmium (Cd) is a highly toxic heavy metal that can pollute water and food sources from burning fossil fuels and Cd is also present in tobacco and tobacco smoke [reviewed in (119)]. Cd is carcinogenic and proinflammatory, affecting multiple organs. Cd is a likely EDC that affects obesity and metabolism by altering adipose tissue through epigenetic changes [reviewed in 119]. In vitro exposure to cadmium altered the expression of MCP-1 (monocyte chemoattractant protein-1) in WAT, a chemokine for macrophage recruitment that plays a role in obesity and WAT inflammation [reviewed in (119, 120)]. Cd can bind to the estrogen receptor and it is also thought to bind to the androgen receptor, however, the exact mechanism through how Cd acts is unknown [reviewed in (119)].
Silver nanoparticles (AgNPs) have also become interesting as a potential EDC and obesogen. AgNPs are widely used as bactericides, and commonly found in the fabric of athletic clothing to eliminate odor. An exploration of the in vivo and in vitro role of silver nanoparticles in thermogenic beige adipocyte differentiation revealed that AgNP inhibited beige adipocyte differentiation, adipocyte thermogenesis, and mitochondrial functioning. AgNP elevated ROS levels within beige adipocytes and activated the MAPK-ERK signaling pathway which is involved in adipogenesis (114).
Arsenic is a metalloid that is widely found in the environment in soil, sediment, and water sources. Currently, more than 200 million people worldwide are exposed to drinking water containing dangerous levels of arsenic (143). Arsenic has been shown to affect adipogenesis in in vivo mouse models, as well as specifically target BAT (115). Arsenic exposure inhibited the differentiation of BAT and lowered expression of PPARγ, UCP1 and PGC1α, which are defining features of brown adipocytes (115). Arsenite, similarly reduced BAT differentiation, decreased mitochondrial functioning, and lowered thermogenesis in BAT by reducing UCP1 expression (117). Arsenite accumulates in BAT and suppressed Sestrin2 phosphorylation by ULK1, which under normal conditions, promotes the antioxidant effects of Sestrin2 against ROS that form within the cells (117, 118). Mice exposed to 300 μg/L of arsenite in drinking water for 9 weeks showed significant increases in inguinal WAT mass, lower thermogenic abilities when exposed to cold temperature, decreased beiging of WAT, and lowered expression of genes involved in regulating thermogenesis and metabolic functions (143).
Sexual Dimorphisms
Multigenerational, transgenerational, and cohort based longitudinal studies offer a valuable platform to explore sex-dependent effects of EDCs and obesogens on males and females from birth into later life. Both human and mouse models have revealed obesogenic related differences in males and females, predisposing one or the other to an increased risk of overweight/obesity. A recent study found that BPA exhibited sex-dependent effects on mouse fetuses when given a low dose of 2.5 micrograms/kg/day. These mice showed changes in hepatic processes, including inflammatory effects and alterations in proteins involved in cholesterol and fatty acid functioning in females only (144). Human cohort studies revealed that prenatal BPA exposure increased the risk of obesity in young girls, but not in boys. A study done in rural East China showed that urinary BPA concentrations were linked to increased adiposity measures in girls at 7 years of age (145). Similarly, a study involving 585 high school aged students from Spain found that BPA disproportionately affected females over males, with an association between dietary BPA exposure and overweight/obesity in females (146).
The BPA analog BPS demonstrated sexually dimorphic effects after exposing female F0 mouse dams to an environmentally relevant dose of BPS in their drinking water during gestation through post-natal day 21, followed by a high fat diet fed to all offspring. F1 male mice became overweight, while F1 females developed dyslipidemia. The F2 generation showed increases in body weight and fat mass for both sexes, however, this generation was impacted the greatest in terms of sexual dimorphisms. Notably, F2 females showed a 35% increase in body weight over their counterpart males. Cholesterol levels, blood glucose levels, and circulating triglycerides were higher in males while non-esterified fatty acids were higher in females. Both sexes showed a downregulation in genes involved in lipogenesis, however, only F2 females downregulated genes involved in lipolysis. The F3 generation showed striking sexual dimorphism in that increased visceral adipose tissue mass only occurred in females, as did the upregulation of genes involved in lipogenesis (131).
When TBT (50 nM) was given to pregnant F0 mouse dams from conception throughout lactation via their drinking water, greater transgenerational and sex-dependent effects were observed in F4 male offspring (147). TBT was found to have largely male dependent transgenerational effects in the F3-F4 group, significantly increasing their body weight, fat storage, and creating changes in the male metabolome (147, 148). F4 Males developed a “thrifty phenotype” and had a reduced ability to mobilize fat storage after overnight fasting. Male mice that were exposed to TBT and later a high fat diet showed an increase in body fat that persisted even after the mice were returned to a normal chow (148).
Sex-dependent and adipocyte related epigenetic effects were observed in F3 offspring of F0 rats exposed to DDT via intraperitoneal injection during pregnancy (149). This study compared DMRs between vehicle control rats, lean rats exposed to atrazine, and DDT exposed rats. Adipocytes isolated from gonadal fat pads were used to identify unique DNA methylation regions between the three groups. Different DMRs were observed between males and females in the F3 generation of DDT exposed rats, indicating sex-dependent obesogenic effects in regions associated with obesity, T2D, and metabolic syndrome. A human longitudinal study revealed striking differences in 12-year-old males and females from a birth cohort study. Data from 240 children, who were exposed to DDT and its metabolite, DDE, in utero, were collected from the Center for the Health Assessment of Mothers and Children of Salinas (CHAMACOS). Maternal environmental DDT and DDE exposure was determined by collecting maternal serum during pregnancy. Data analysis was consistent with previous DDT and DDE studies, and revealed that in utero exposure increased obesity related measures for males only, indicating a sex-specific effect and risk for obesity in human males and not females (150).
Participants and data from the Hamamatsu Birth Cohort for Mothers and Children study were used to identify potential sex-specific prenatal exposure effects of perfluoroalkyl substances. Umbilical cord serum was measured for perfluorooctane sulfonate (PFOS) and PFOA concentration levels. BMI repeated measurements were taken from the children at ages 1, 4, 10, 18, 24, 32, 40, 50, and 66 months. The results indicated a significant difference in the BMI and body weight of girls. Prenatal exposure was linked to a lower birthweight in the girls, yet this effect was reversed as they aged, resulting in increased BMI and body weight as they reached 5 years of age (151).
The Microbiome and Obesity
The gut microbiota is predominantly composed of bacteria from the phyla Bacteroidetes, Firmicutes, and Actinobacteria, which secrete enzymes that degrade dietary fiber (152). Individual differences in dietary habits can alter the diversity of microbial species within the intestinal tract (153–155). Moreover, the microbiome was found to differ markedly between normal weight and obese individuals. The obese microbiome was characterized by the predominance of Bacteroidetes over Firmicutes in the gut (155). Remarkably, transplant of the gut microbiome from obese mice to germ-free mice transferred the obese phenotype and microbiome and was characterized by a greater capacity to harvest energy (155). The gut microbiome is an important factor in energy harvest as it is a unique “organ” with microbial symbionts which help metabolize dietary polysaccharides and promote fat storage (156). Different individuals may share similar microbial genes which adapt to the unique metabolic needs of that individual, but have different bacteria lineages (157). Adult twins were observed to have similar bacterial communities than unrelated individuals, but there is no single bacterial phenotype, or “core” human gut microbiome defined by a set of microbial organismal lineages, shared by unrelated individuals. When samples of humanized mice (i.e. mice that received fecal microbiota transplantation of human feces) were fed the Western diet, they showed increased adiposity; a trait is transmissible via microbiota transplantation (158).
The Role of EDCs in the Gut Microbiome
EDCs have been implicated in disease states such as insulin resistance, glucose intolerance, type 2 diabetes, and obesity (159), breast and prostate cancer (160, 161), as well as reproductive development disorders (162). In addition to their direct effects on physiology, exposure to EDCs, such as BPA, disturbs gut microbial composition (163). This may lead to changes in host lipid metabolism, among other effects (163). It was proposed that environmental obesogens could cause gut dysbiosis which might lead to inflammation and insulin resistance (164). The gut employs a multilayered mucus structure to maintain distance between the gut epithelial cells and the gut microbiota as a protective mechanism (165). EDCs could increase the permeability of the small intestine, increasing the likelihood that bacterial pathogens will enter the body’s circulation and target other organs (166). Food additives, such as artificial sweeteners (167, 168), and contaminants such as pesticide residues (169) can interfere with the gut microbiota and gut barrier function which could lead to intestinal, metabolic, and autoimmune disorders. Notably, obesity and T2D were found to be associated with intestinal dysbiosis (170) and gut barrier disruption (171). A list of potential and known obesogenic chemicals and how they affect the gut microbiome is presented in Table 4.
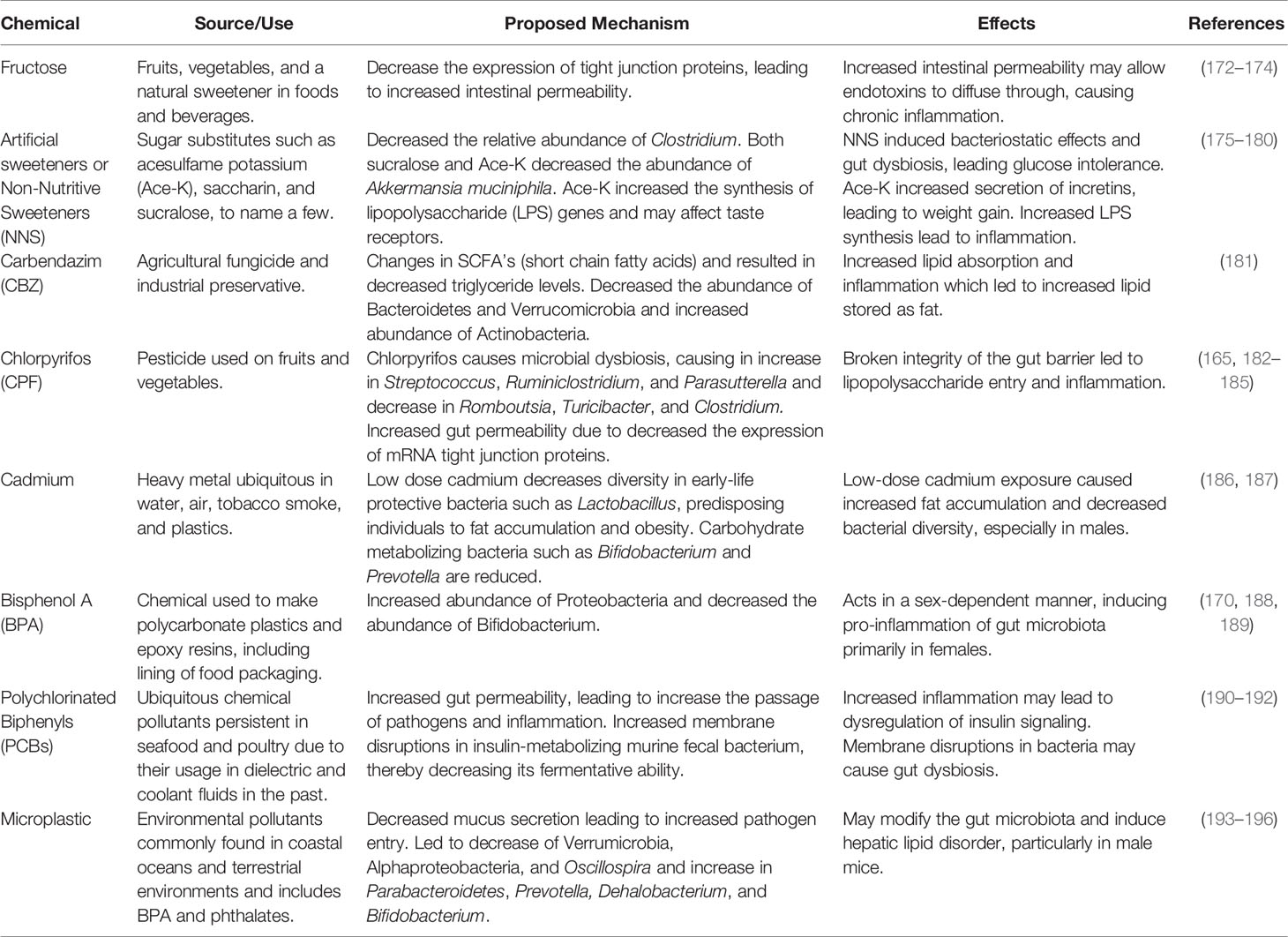
Table 4 Chemical obesogens and their effects on the microbiome.
Artificial Sweeteners
Non-nutritive sweeteners (NNS) are widely consumed dietary additives that could act as obesogens, impairing the growth of gut flora and inducing glucose intolerance (175). NNSs such as acesulfame potassium (Ace-K), saccharin, and sucralose induced bacteriostatic effects, which changed the composition of the gut microbiome (176) and induced glucose intolerance by promoting gut dysbiosis (177). Consumption of the NNS sucralose (a chlorinated derivative of sucrose) decreased the relative abundance of Clostridium, which converts primary bile acids into secondary bile acids (178). Both sucralose and Ace-K decreased the abundance of Akkermansia muciniphila (179), which is correlated with increased lipid metabolism and decreased inflammation (197). Sucralose may also affect taste receptors, thereby increasing the secretion of incretins glucose-dependent insulinotropic polypeptide (GIP) and GIP-1, leading to weight gain, hyperglycemia, hyperleptinemia, and hyperinsulinemia (180). Ace-K appears to have gender-specific effects as it decreased functional genes involved in energy metabolism in female mice but increased their expression in male mice (175). Ace-K also significantly increased the synthesis of lipopolysaccharide (LPS) genes, which may increase inflammation (175). Notwithstanding these results, the obesogenic properties of artificial sweeteners are highly disputed and future studies aimed at testing the links between artificial sweeteners, gut dysbiosis, and obesity will be required.
Fructose Consumption
The first scientist to put fructose on the table as a cause for obesity was Dr. Robert Lustig in the early 2000’s. Fructose was found to cause changes in liver metabolism and energy signaling, creating a feedback loop in which insulin resistance and overeating occurs (198). Fructose is found in many processed food and beverages and is an obesogen known to cause glucose intolerance and insulin resistance (198). Consequently, it has been found that fructose restriction results in increased glucose tolerance and decreased insulin levels (199). On the other hand, fructose consumption causes hepatic de novo lipogenesis (DNL), which is dependent on the metabolism of fructose to acetate by the gut microbiota (200). The depletion of microbiota suppresses the conversion of fructose into hepatic acetyl CoA and fatty acids. It is proposed that there is a dual mechanism for fructose DNL: hepatic fructose metabolism promotes DNL transcriptionally while microbial acetate fuels DNL. Additionally, male mice fed a high-fat, high fructose diet (HFrD) demonstrated increased expression of inflammatory cytokines monocyte chemoattractant protein -1 (MCP-1_, toll-like receptor 4 (TLR4), interleukin-1 beta (IL-1β), and tumor necrosis factor alpha (TNF-α), which were associated with glucose intolerance and lipid accumulation (201). Fructose may cause gut dysbiosis and the upregulation of genes important for fat transport and storage such as CD36, fatty acid synthase (FAS), and sterol regulatory element-binding protein 1 (SREBP1), leading to hepatic steatosis (202). This may be a possible contributor to fatty liver disease (201). Exposure of rats to high-fat and sucrose (HFS) resulted in animals with 51% fat mass and 24% lean mass compared with 40% fat mass and 48% lean mass in the high-fat and fructose (HFF) diet group (203). Thus, while both types of diets increased fat mass, the HFS elicited substantially more fat mass than did the HFF diet. In addition to body fat mass composition, compared to the HFF group, the HFS group showed higher metabolic dysregulation and glucose intolerance, greater levels of liposaccharides and insulin, an increase in ROS as well as in markers for ROS, and an increase in lipid synthesis transcription factor, Srebp-1 (203). The comparison between fructose and sucrose indicate that the type of carbohydrate can have different effects, and that a HFS diet created more negative results than did a HFF diet. At the species level, there was a significant decrease in the abundance of Coprococcus eutactus in the HFS and HFF groups and an increase in abundance of Lactobacillus reuteri and Bacteroides fragilis in the HFF group. These two species have been associated with compromised integrity of the intestinal epithelium (203). Dysbiosis in the gut microbiota may also cause increased production of LPS, which stimulates a systematic inflammatory response (204) that may lead to insulin resistance and glucose intolerance (203). Rats fed a high fructose diet for one week showed reduced gut flora levels, increased inflammatory markers TNF-α and IFN-γ, increased glycation of gut proteins, and the reduced ability of extracted gut microbiota to grow on non-glycated proteins, likely causing the reduction in gut microbial survival (205). Interestingly, the glycation rate of high fructose fed rats was found to be higher than in diabetic rats (205).
It was proposed that one of the mechanisms through which fructose consumption causes obesity is by increasing intestinal permeability, leading to lipid accumulation, inflammation, and metabolic endotoxemia (a type of low-grade inflammation) (201). Fructose consumption has also been shown to affect the maternal microbiome during pregnancy, which is thought to alter the offspring’s gut health as well. The effects of fructose consumption on the maternal microbiome were tested by feeding rats 10% fructose via their water from 8 weeks through pregnancy. It was found that the maternal microbiome was altered to produce less beneficial bacteria, Lactobacillus and Bacteroides. Offspring maintained on fructose exposure showed a decrease in genes responsible for gut barrier function and a dysregulation of genes responsible for epithelial tight junction (172), which are diffusion barriers that regulate the passage of solutes across the epithelia (206). Shorter small intestines, lower birth weight, and increased fat mass were also observed, resulting in an overall unhealthy phenotype. In a non-high-fat diet-induced obesity model, microbiome dysbiosis was also associated with decreased expression of tight junction proteins (207). It is thought that the resulting increased gut permeability might allow endotoxins such as LPS to diffuse through the tight junctions and interact with host immune cells to cause chronic inflammation (173). More studies are needed to establish a direct link between obesogen-induced gut dysbiosis and intestinal permeability.
Carbendazim
Carbendazim is a broad-spectrum, benzimidazole fungicide that is an EDC and an obesogen (181). Mice that were administered 0.1, 0.5, or 5mg/kg body weight per day carbenazim (CBZ) via their drinking water showed alterations in the gut microbiota that led to changes in the relative abundance of circulating short chain fatty acids (SCFAs) (181). Exposure to CBZ decreased the relative abundance of Bacteroidetes and Verrucomicrobia and increased the abundance of Actinobacteria, which resulted in altered levels of SCFAs (181). Microbial dysbiosis in the host resulted in downstream effects including decreased triglyceride (TG) synthesis in the liver, increased lipid absorption, and a multi-tissue inflammatory response (181). To compensate for the elevated lipid levels, the host reduced lipid synthesis in the liver and increased lipid storage in fat tissue (181). SCFAs and the gut microbiota work in conjunction to maintain optimal gut health, and the metabolite of SCFAs released by gut microbes play a role in epithelial cell health and intestinal barrier function. G-protein coupled receptors Gpr41 and Gpr43 have been discovered to work with SCFAs to mediate processes involved in host metabolism and intestinal epithelial functioning and health (181). Altered levels of SCFAs further change the expression of intestinal Gpr41 and Gpr43 as well as downregulates genes involved in host immune function (181). Taken together, these data are consistent with a model in which CBZ exposure caused gut dysbiosis that led to increased absorption of TG, leading to inflammation, hyperlipidemia, and increased fat storage. However, it is also possible that gut dysbiosis occurred in parallel with CBZ-altered lipid metabolism rather than causing it (181). The effects of CBZ have also been assessed in adult zebrafish who were exposed to 0, 30, and 100 μg/L of CBZ for 21 days (208). CBZ exposure altered the gut microbiota of the zebrafish, significantly decreasing phylum levels of Firmicutes, Bacteroidetes, Actinobacteria, α-Proteobacteria, γ-Proteobacteria and Verrucomicrobia (208).
Chlorpyrifos
Chlorpyrifos (CPF) is a pesticide that is widely used on fruits and vegetables (182–184). CPF is also an EDC that decreased the levels of epinephrine, luteinizing hormone (LH), and follicle-stimulating hormone (FSH) (185), and induced gut microbial dysbiosis at low doses (209, 210). CPF exposure led to increased relative abundance of Streptococcus, Ruminiclostridium, and Coriobacteriaceae and decreased abundance of Romboutsia, Turicibacter, and Clostridium (185). CPF led to enriched abundance of Parasutterella in both normal fat diet fed and high fat diet fed rats; similar to results found after antibiotic treatment (185). CPF treatment decreased the abundance of SCFA-producing bacteria such as Romboutsia, Turicibacter, and Clostridium and enriched the pathogenic genus Streptococcus, which can result in an altered gut barrier (211). Interestingly, the effects of CPF on gut hormones appear to be age specific, as CPF increases the serum levels of glucagon-like peptide -1(GLP-1), pancreatic polypeptide (PP), peptide YY (PYY), and GIP in the newly weaned 3 week old rats in comparison to only stimulating the release of PPY and ghrelin when CPF is exposed during adulthood at 8 weeks of age up to 28 weeks of age (185). Zebrafish that were exposed to 30, 100, and 300 μg/L of CPF for 21 days, showed higher levels of oxidative stress coupled with the decrease of microbial diversity within the gut (74). Specifically, levels of γ-Proteobacteria showed a significant decrease in the gut microbiota, along with significant changes in 25 other types of genus level bacteria within the gut (74).
One of the proposed mechanisms through which CPF may cause obesity is by interfering with the gut microbiota and increasing gut permeability (185). CPF may increase gut permeability by reducing the expression of tight junction proteins (165). It was proposed that CPF breaks the integrity of the gut barrier by decreasing expression of mRNA encoding tight junction proteins which may lead to LPS entry and low-grade inflammation (165). Thus, the imbalance of microbiota, coupled with increased gut permeability, causes an increase in LPS and leads to inflammation (165), which are characteristics of obesity and diabetes. However, the mechanism(s) through which chlorpyrifos acts to modulate the microbiome and promote obesity are areas which needs further investigation.
Cadmium
Cadmium is a heavy metal to which humans are readily exposed because it is ubiquitous in water, air, tobacco smoke, as well as plastics (186, 187). Exposure to cadmium during early life is especially detrimental since the gut microbiota is just being established (212). Cadmium decreases overall diversity in the gut microbiota, particularly in populations such as Lactobacillus, which are known to be protective early in life (213). This can reset the metabolic programming throughout life, leading to fat accumulation and obesity-related metabolic diseases (187). Low-dose cadmium exposure reduces the abundance of Bifidobacterium and Prevotella, which metabolize carbohydrates such as oligosaccharides and polysaccharides respectively (187, 214, 215). Interestingly, early life cadmium exposure elicited slightly more adiposity in males than in females (187, 212). However, the use of mostly male mice in metabolic studies of cadmium exposure (187) and metabolic variations between genders could contribute to these differences; thus, more research in this area is necessary (173). When three-week-old rats were administered cadmium at 0.1, 0.25, 1, or 4 mg/kg for eight weeks, changes of gut microbiota were observed (216). Mice in the 4 mg/kg group showed significant decreases in the beneficial genus Prevotellaceae_NK3B31_group, Prevotella_9, and Lachnoclostridium, and increases in Escherichia_Shigella, which increases oxidative stress and inflammation (216).
Bisphenols
BPA is a chemical obesogen that is ubiquitous in the environment. BPA is used for making polycarbonate plastics and epoxy resins including those that line food packaging, as well as in thermal papers (170, 217). BPA is an EDC that may cause gut dysbiosis, metabolic disorders, and eventually lead to T2D (170, 188). Oral exposure to BPA dissolved in water in mice showed an increased the abundance of bacterial phylum Proteobacteria and a decrease in phylum Tenericutes (188). The increase in Proteobacteria, which is a microbial marker of dysbiosis, was caused by the induction of Epsilon proteobacteria (188). The increase of Epsilon proteobacteria was due to the increase of Helicobacteraceae which is associated with inflammatory bowel disease (218). Exposure to BPA caused a decrease in Bifidobacterium, which may be significant as some strains are known to have anti-inflammatory properties in intestinal epithelial cells (170, 219). Interestingly, the effects of BPA may be sex dependent as it induced a shift towards more pro-inflammatory gut microbiota in adult female mice and anti-inflammatory microbiota in adult male mice (189). Females injected with the antibiotic vancomycin did not experience BPA-induced gut dysbiosis, which suggests that gram-positive bacteria may not be a causal mechanism (189). BPA decreased the production of immunoglobulin A (IgA) which plays a key role in gut barrier integrity and gut homeostasis, leading to gut dysbiosis (170). Overall, the BPA microbiome had a similar species profile to one shaped by a high-fat and high-sucrose diet (188). These were linked to inflammatory bowel disease, metabolic disorders, and colorectal cancer (166). When 200 Crohn’s Disease (CD) patients were studied to assess the relationship between EDCs and CD, higher levels of BPA were found in patients with active CD compared to those in remission (220). In both remission and active CD groups, BPA was found to alter inflammatory responses in patients with gut barrier disruption and in those who tested positive for bacterial DNA in their blood (220). Taken together, it is evident that BPA could be playing an active role in the pathology of CD by disrupting the gut microbiome.
More recently, it has been shown that Bisphenol S (BPS) and Bisphenol F (BPF), which are BPA alternatives, cause microbial dysbiosis and are inversely related to developmental toxicity and estrogenic activity (221). Zebrafish that were exposed to BPF, BPS, or a combination of both chemicals at 1, 10, 100, or 1000 μg/L showed oxidative stress and intestinal inflammation as well as changes in their gut microbiome (222). Exposure to BPS and BPF, both separately and in conjunction, were found to increase pathogenic bacteria in the intestinal tract of the zebrafish, including the genus Flavobacterium, Pseudomonas, and Stenotrophomonas (222). In addition to direct action on the microbiome, exposure to EDCs such as BPA might affect the brain by disrupting the neural programming through the microbiome-gut-brain axis (223).
Polychlorinated Biphenyls
PCBs (polychlorinated biphenyls) are ubiquitous environmental pollutants, and at environmentally relevant doses, may alter the abundance and diversity of the gut microbiome (224). PCBs may cause gut inflammation, which are implicated in metabolic diseases such as diabetes, through gut dysbiosis (190). Exposure to PCBs causes abdominal fat accumulation and obese individuals may be more vulnerable to PCB exposure than lean individuals (225). Male mice that were administered PCB via oral gavage had an increase in Clostridiales and decrease in Bifidobacterium, Lactobacillus, Ruminococcus, and Oscillospira in comparison to the controls. Decreases in Bifidobacterium and Lactobacillus are significant as these are probiotics that may reduce body weight, fat mass (226), and decrease endotoxemia (173). When exposed to a high dose of PCBs, there was an increase in Prevotella (227) which is associated with insulin resistance (228) and intestinal inflammation at mucosal sites (229), although Prevotella has been associated with beneficial health effects in another study (215). Additionally, when adult female mice were exposed to low-dose PCBs, there was an increase in A. muciniphila (227).
It is possible that PCB exposure may lead to obesity by disrupting the epithelial integrity and increasing gut permeability, rendering it vulnerable to pathogens in the gut mucosa (191). This may lead to high levels of inflammation, which is known to lead to dysregulation of insulin signaling, GLP-1 secretion from the intestine (190), and interfere with adipogenesis through the aryl hydrocarbon receptor (AhR) (230). Additionally, PCB exposure induced membrane disruptions in insulin-metabolizing murine fecal bacterium, decreasing its fermentative ability (192). Bacterial membrane disruptions were characterized by leakage of intracellular potassium, which is a central intracellular cation in bacterial cells (192). Therefore, bacterial membrane disruption may be a possible mechanism by which PCB exposure could cause gut dysbiosis (192). Recent studies have shown that PBC exposure induced deviations in the gut microbiome of mice, which further promoted non-alcoholic fatty liver disease in the animals. The obesity inducing receptor, PXR, and anti-obesity nuclear receptor, CAR, were knocked out in mice to assess the relationship between the receptors and the effects of PCB on metabolic functioning and the regulation of gut microbiome (231). It was previously shown that the PBC mixture, Aroclor1260, exacerbated NAFLD in mouse models via the activation of RXR and CAR pathways. It should be noted that mixtures of chemicals pose a challenge when studying the hormonal effects of individual chemicals, and conclusions cannot always be drawn from the effects of an individual chemical if it is a part of a mixture. Aroclor1260 was chosen as the test PCB mixture due to its relevance to human exposure, its resistance to metabolism, and its ability to bioaccumulate. PCB was further used to explore the role of PXR and CAR in relation to changes in the gut microbiome in diet induced obese mice and to determine what role PCR and CAR play in the gut-liver axis in relation to the gut microbiome. Both CAR and RXR knockout groups showed altered microbiome diversity, leading to greater hepatic and intestinal inflammation, dysregulations in energy metabolism, and nonalcoholic steatohepatitis. KO mice also showed a decrease in microbes related to lower inflammation levels and an increase in microbes related to inflammation, indicating that CAR and RXR may have a protective effect on the gut-liver-axis via the gut microbiome (231).
Microplastic
Microplastics, which can range from nanometers to larger than five millimeters, are environmental pollutants commonly found in coastal oceans and terrestrial environments (193). They are particularly dangerous as they can persist in the marine ecosystem, rendering them difficult to remove (193). Moreover, many microplastics contain obesogens such as BPA (194) and phalates (195). Although there is a paucity of information regarding microplastics and their effects on the microbiome, a recent study showed that polystyrene microplastic may modify the gut microbiota and induce hepatic lipid disorder in male mice (196). Polystyrene microplastics caused decreased mucus secretion (196), which may allow pathogens to penetrate the intestinal mucosa. The relative abundance of Verrucomicrobia, Alphaproteobacteria, and Oscillospira decreased whereas the abundance of Parabacteroidetes, Prevotella, Dehalobacterium, and Bifidobacterium increased (196). While polystyrene microplastics were found to have a direct effect on the populations of various gut microbiota, it was also observed to disrupt lipid metabolism on a molecular level by altering the expression of key genes related to hepatic lipogenesis. Levels of PPARγ mRNA were found to decrease, while the mRNA levels of PPARα mRNA and fatty acid synthase (Fas), increased (196). PPARα is largely responsible for β-oxidation (196), and PPARγ is known to play key roles in adipocyte differentiation, lipid metabolism (232) and the storage and mobilization of lipids (233). It was proposed that polystyrene microplastics may cause gut dysbiosis and affect lipid metabolism in the liver through an indirect and unknown pathway related to the gut microbiota, although the mechanism remains unclear (196). More studies are necessary to validate whether microplastics lead to hepatic lipid metabolism disorder, and whether the effects are dependent or independent to the populations of gut microbiota.
Conclusion
The study of obesogenic compounds is still in its early stages. However, the field has already shed ample light on factors that could contribute to the obesity pandemic beyond the energy balance paradigm. Metabolic disease has demanded increasing attention as the prevalence of worldwide obesity and related morbidities continues to rise (1). Thus far, ~50 obesogens have been recognized [reviewed in (30)], but more research must be done to discover those that remain unidentified and to test other candidate obesogens, in vivo. Reliable and replicable standardized detection assays and high throughput screening methods are needed to identify the remaining obesogens in the environment quickly and efficiently. The mechanisms of action through which known obesogens function by are still not well understood. In vitro studies (42) and in vivo studies (49) of TBT put the PPARγ and RXR pathways on the map as the major pathways of adipogenesis. Several obesogens have been found to function through the activation of this heterodimeric master regulator of adipogenesis. Other obesogens may function through pathways that remain to be identified. A more comprehensive understanding of obesogenic mechanisms and the mapping of their functional pathways will be key to implementing both preventative measures and therapeutic strategies against metabolic disease.
Current research on thermogenic fat, its function, and the role of its disruption in metabolic disease is calling for more attention on how modulating thermogenesis can be used in the treatment of metabolic diseases such as T2D and obesity. Increasing the activity of brown adipose tissue and promoting the beiging of WAT through genetic approaches, pharmacological methods, tissue-specific and cell type-specific strategies have become promising therapeutic avenues against obesity [reviewed in (234)]. There is a paucity of data concerning the role of obesogens and their effect on the microbiome, and more research is needed on how obesogens affect the gut. How the microbiome interacts with the gut-brain axis and how this influences appetite is a new field of research that may offer novel therapeutic strategies and directions for the treatment of obesity. The gut microbiota converts dietary nutrients into metabolites that can regulate appetite via vagal stimulation or through immune-neuroendocrine mechanisms (235). It is established that some obesogenic substances effect the gut microbiome, and when the gut microbiome moves away from its homeostatic levels, a variety of problems can arise, such as the dysregulation of orexigenic signals such as insulin and ghrelin which have effects on appetite and satiation (236). Potential therapeutics that target the gut microbiome, such as microbiome transplantation, may become viable future treatments for obesity.
The study of EDCs offers insights into how normal metabolic processes can be disrupted, and why the population is becoming unhealthier, particularly with respect to metabolic disease. As more MDCs are discovered, and more information is uncovered about currently used chemicals in industry, strategies to restrict usage and minimize exposure will become important. Avoidance of exposure through ingestion, inhalation, and direct contact is a definitive way to prevent metabolic disruption caused by EDCs before disease develops. In vivo transgenerational studies, which were only briefly discussed in this review, revealed epigenomic reprogramming effects and phenotypical metabolic effects caused by F0 chemical exposure on offspring into the F4 generation (147, 237). The existence of such “generational toxicity” demands further education about exposure prevention and transparency to keep the public and future generations safe from the effects of exposure to harmful chemicals.
Author Contributions
BB, NM, CD, and CC contributed to writing the manuscript. BB and NM performed final editing, revisions, and submission.
Funding
Supported by grants from the US National Institutes of Health, R01ES023316 and ES031139 to BB.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
3. Lakkis JI, Weir MR. Obesity and Kidney Disease. Prog Cardiovasc Dis (2018) 61:157–67. doi: 10.1016/j.pcad.2018.07.005
4. Jehan S, Zizi F, Pandi-Perumal SR, McFarlane SI, Jean-Louis G, Myers AK. Energy Imbalance: Obesity, Associated Comorbidities, Prevention, Management and Public Health Implications. Adv Obes Weight Manag Control (2020) 10:146–61. doi: 10.15406/aowmc.2020.10.00321
5. Anderson MR, Geleris J, Anderson DR, Zucker J, Nobel YR, Freedberg D, et al. Body Mass Index and Risk for Intubation or Death in SARS-CoV-2 Infection : A Retrospective Cohort Study. Ann Intern Med (2020) 173:782–90. doi: 10.7326/M20-3214
6. Sagar R, Gupta T. Psychological Aspects of Obesity in Children and Adolescents. Indian J Pediatr (2018) 85:554–9. doi: 10.1007/s12098-017-2539-2
7. Milano W, Ambrosio P, Carizzone F, De Biasio V, Di Munzio W, Foia MG, et al. Depression and Obesity: Analysis of Common Biomarkers. Diseases (2020) 8:23. doi: 10.3390/diseases8020023
8. Lindberg L, Persson M, Danielsson P, Hagman E, Marcus C. Obesity in Childhood, Socioeconomic Status, and Completion of 12 or More School Years: A Prospective Cohort Study. BMJ Open (2021) 11:e040432.
9. Hajian-Tilaki KO, Heidari B. Association of Educational Level With Risk of Obesity and Abdominal Obesity in Iranian Adults. J Public Health (Oxf) (2010) 32:202–9. doi: 10.1093/pubmed/fdp083
10. Tremmel M, Gerdtham UG, Nilsson PM, Saha S. Economic Burden of Obesity: A Systematic Literature Review. Int J Environ Res Public Health (2017) 14:435. doi: 10.3390/ijerph14040435
12. Mozaffarian D. Foods, Obesity, and Diabetes-are All Calories Created Equal? Nutr Rev (2017) 75:19–31. doi: 10.1093/nutrit/nuw024
13. Brown RE, Sharma AM, Ardern CI, Mirdamadi P, Mirdamadi P, Kuk JL. Secular Differences in the Association Between Caloric Intake, Macronutrient Intake, and Physical Activity With Obesity. Obes Res Clin Pract (2016) 10:243–55. doi: 10.1016/j.orcp.2015.08.007
14. Kavanagh K, Jones KL, Sawyer J, Kelley K, Carr JJ, Wagner JD, et al. Trans Fat Diet Induces Abdominal Obesity and Changes in Insulin Sensitivity in Monkeys. Obes (Silver Spring) (2007) 15:1675–84. doi: 10.1038/oby.2007.200
15. Hall KD, Chung ST. Low-Carbohydrate Diets for the Treatment of Obesity and Type 2 Diabetes. Curr Opin Clin Nutr Metab Care (2018) 21:308–12. doi: 10.1097/MCO.0000000000000470
16. Ludwig DS, Ebbeling CB. The Carbohydrate-Insulin Model of Obesity: Beyond "Calories In, Calories Out". JAMA Intern Med (2018) 178:1098–103. doi: 10.1001/jamainternmed.2018.2933
17. Zoeller RT, Brown TR, Doan LL, Gore AC, Skakkebaek NE, Soto AM, et al. Endocrine-Disrupting Chemicals and Public Health Protection: A Statement of Principles From The Endocrine Society. Endocrinology (2012) 153:4097–110. doi: 10.1210/en.2012-1422
18. Diamanti-Kandarakis E, Bourguignon JP, Giudice LC, Hauser R, Prins GS, Soto AM, et al. Endocrine-Disrupting Chemicals: An Endocrine Society Scientific Statement. Endocr Rev (2009) 30:293–342. doi: 10.1210/er.2009-0002
19. Sridevi V, Naveen P, Karnam VS, Reddy PR, Arifullah M. Beneficiary and Adverse Effects of Phytoestrogens: A Potential Constituent of Plant-Based Diet. Curr Pharm Des (2021) 27:802–15. doi: 10.2174/1381612826999200917154747
20. Street ME, Audouze K, Legler J, Sone H, Palanza P. Endocrine Disrupting Chemicals: Current Understanding, New Testing Strategies and Future Research Needs. Int J Mol Sci (2021) 22:933.
21. Kanno J. Introduction to the Concept of Signal Toxicity. J Toxicol Sci (2016) 41:SP105–9. doi: 10.2131/jts.41.SP105
22. Amato AA, Wheeler HB, Blumberg B. Obesity and Endocrine-Disrupting Chemicals. Endocr Connect (2021) 10:R87–105.
23. Egusquiza RJ, Blumberg B. Environmental Obesogens and Their Impact on Susceptibility to Obesity: New Mechanisms and Chemicals. Endocrinol (2020) 161:1–14. doi: 10.1210/endocr/bqaa024
24. Ren XM, Kuo Y, Blumberg B. Agrochemicals and Obesity. Mol Cell Endocrinol (2020) 515:110926. doi: 10.1016/j.mce.2020.110926
25. Grun F, Blumberg B. Environmental Obesogens: Organotins and Endocrine Disruption via Nuclear Receptor Signaling. Endocrinology (2006) 147:S50–5. doi: 10.1210/en.2005-1129
26. Heindel JJ, Vom Saal FS, Blumberg B, Bovolin P, Calamandrei G, Ceresini G, et al. Parma Consensus Statement on Metabolic Disruptors. Environ Health (2015) 14:54. doi: 10.1186/s12940-015-0042-7
27. Heindel JJ, Blumberg B, Cave M, Machtinger R, Mantovani A, Mendez MA, et al. Metabolism Disrupting Chemicals and Metabolic Disorders. Reprod Toxicol (2017) 68:3–33. doi: 10.1016/j.reprotox.2016.10.001
28. Griffin MD, Pereira SR, DeBari MK, Abbott RD. Mechanisms of Action, Chemical Characteristics, and Model Systems of Obesogens. BMC BioMed Eng (2020) 2:6. doi: 10.1186/s42490-020-00040-6
29. Jakab J, Miskic B, Miksic S, Juranic B, Cosic V, Schwarz D, et al. Adipogenesis as a Potential Anti-Obesity Target: A Review of Pharmacological Treatment and Natural Products. Diabetes Metab Syndr Obes (2021) 14:67–83. doi: 10.2147/DMSO.S281186
30. Heindel JJ, Blumberg B. Environmental Obesogens: Mechanisms and Controversies. Annu Rev Pharmacol Toxicol (2019) 59:89–106. doi: 10.1146/annurev-pharmtox-010818-021304
31. Lopez-Perez D, Redruello-Romero A, Garcia-Rubio J, Arana C, Garcia-Escudero LA, Tamayo F, et al. In Patients With Obesity, the Number of Adipose Tissue Mast Cells Is Significantly Lower in Subjects With Type 2 Diabetes. Front Immunol (2021) 12:664576. doi: 10.3389/fimmu.2021.664576
32. Tontonoz P, Spiegelman BM. Fat and Beyond: The Diverse Biology of PPARgamma. Annu Rev Biochem (2008) 77:289–312. doi: 10.1146/annurev.biochem.77.061307.091829
33. Tripathi M, Yen PM, Singh BK. Estrogen-Related Receptor Alpha: An Under-Appreciated Potential Target for the Treatment of Metabolic Diseases. Int J Mol Sci (2020) 21:1645. doi: 10.3390/ijms21051645
34. Matsushima A, Liu X, Okada H, Shimohigashi M, Shimohigashi Y. Bisphenol AF is a Full Agonist for the Estrogen Receptor ERalpha But a Highly Specific Antagonist for ERbeta. Environ Health Perspect (2010) 118:1267–72. doi: 10.1289/ehp.0901819
35. Sun Z, Yang X, Liu QS, Li C, Zhou Q, Fiedler H, et al. Butylated Hydroxyanisole Isomers Induce Distinct Adipogenesis in 3T3-L1 Cells. J Hazard Mater (2019) 379:120794. doi: 10.1016/j.jhazmat.2019.120794
36. Pereira-Fernandes A, Demaegdt H, Vandermeiren K, Hectors TL, Jorens PG, Blust R, et al. Evaluation of a Screening System for Obesogenic Compounds: Screening of Endocrine Disrupting Compounds and Evaluation of the PPAR Dependency of the Effect. PLoS One (2013) 8:e77481. doi: 10.1371/journal.pone.0077481
37. Pereira-Fernandes A, Vanparys C, Hectors TL, Vergauwen L, Knapen D, Jorens PG, et al. Unraveling the Mode of Action of an Obesogen: Mechanistic Analysis of the Model Obesogen Tributyltin in the 3T3-L1 Cell Line. Mol Cell Endocrinol (2013) 370:52–64. doi: 10.1016/j.mce.2013.02.011
38. Chamorro-Garcia R, Kirchner S, Li X, Janesick A, Casey SC, Chow C, et al. Bisphenol A Diglycidyl Ether Induces Adipogenic Differentiation of Multipotent Stromal Stem Cells Through a Peroxisome Proliferator-Activated Receptor Gamma-Independent Mechanism. Environ Health Perspect (2012) 120:984–9. doi: 10.1289/ehp.1205063
39. Helies-Toussaint C, Peyre L, Costanzo C, Chagnon MC, Rahmani R. Is Bisphenol S a Safe Substitute for Bisphenol A in Terms of Metabolic Function? Vitro Study. Toxicol Appl Pharmacol (2014) 280:224–35. doi: 10.1016/j.taap.2014.07.025
40. Chamorro-García R, Shoucri BM, Willner S, Käch H, Janesick A, Blumberg B. Effects of Perinatal Exposure to Dibutyltin Chloride on Fat and Glucose Metabolism in Mice, and Molecular Mechanisms. Environ Health Perspect (2018) 126:057006. doi: 10.1289/EHP3030
41. Milton FA, Lacerda MG, Sinoti SBP, Mesquita PG, Prakasan D, Coelho MS, et al. Dibutyltin Compounds Effects on PPARgamma/RXRalpha Activity, Adipogenesis, and Inflammation in Mammalians Cells. Front Pharmacol (2017) 8:507. doi: 10.3389/fphar.2017.00507
42. Kanayama T, Kobayashi N, Mamiya S, Nakanishi T, Nishikawa J. Organotin Compounds Promote Adipocyte Differentiation as Agonists of the Peroxisome Proliferator-Activated Receptor Gamma/Retinoid X Receptor Pathway. Mol Pharmacol (2005) 67:766–74. doi: 10.1124/mol.104.008409
43. Temkin AM, Bowers RR, Magaletta ME, Holshouser S, Maggi A, Ciana P, et al. Effects of Crude Oil/Dispersant Mixture and Dispersant Components on PPARgamma Activity in Vitro and in Vivo: Identification of Dioctyl Sodium Sulfosuccinate (DOSS; CAS #577-11-7) as a Probable Obesogen. Environ Health Perspect (2016) 124:112–9. doi: 10.1289/ehp.1409672
44. Park Y, Kim Y, Kim J, Yoon KS, Clark J, Lee J, et al. Imidacloprid, a Neonicotinoid Insecticide, Potentiates Adipogenesis in 3T3-L1 Adipocytes. J Agric Food Chem (2013) 61:255–9. doi: 10.1021/jf3039814
45. Mesnage R, Biserni M, Genkova D, Wesolowski L, Antoniou MN. Evaluation of Neonicotinoid Insecticides for Oestrogenic, Thyroidogenic and Adipogenic Activity Reveals Imidacloprid Causes Lipid Accumulation. J Appl Toxicol (2018) 38:1483–91. doi: 10.1002/jat.3651
46. Hurst CH, Waxman DJ. Activation of PPARalpha and PPARgamma by Environmental Phthalate Monoesters. Toxicol Sci (2003) 74:297–308. doi: 10.1093/toxsci/kfg145
47. Biserni M, Mesnage R, Ferro R, Wozniak E, Xenakis T, Mein CA, et al. Quizalofop-P-Ethyl Induces Adipogenesis in 3T3-L1 Adipocytes. Toxicol Sci (2019) 170:452–61. doi: 10.1093/toxsci/kfz097
48. Bowers RR, Temkin AM, Guillette LJ, Baatz JE, Spyropoulos DD. The Commonly Used Nonionic Surfactant Span 80 has RXRalpha Transactivation Activity, Which Likely Increases the Obesogenic Potential of Oil Dispersants and Food Emulsifiers. Gen Comp Endocrinol (2016) 238:61–8. doi: 10.1016/j.ygcen.2016.04.029
49. Grun F, Watanabe H, Zamanian Z, Maeda L, Arima K, Cubacha R, et al. Endocrine-Disrupting Organotin Compounds are Potent Inducers of Adipogenesis in Vertebrates. Mol Endocrinol (2006) 20:2141–55. doi: 10.1210/me.2005-0367
50. Inadera H, Shimomura A. Environmental Chemical Tributyltin Augments Adipocyte Differentiation. Toxicol Lett (2005) 159:226–34. doi: 10.1016/j.toxlet.2005.05.015
51. Andrews FV, Kim SM, Edwards L, Schlezinger JJ. Identifying Adipogenic Chemicals: Disparate Effects in 3T3-L1, OP9 and Primary Mesenchymal Multipotent Cell Models. Toxicol. In Vitro (2020) 67:104904. doi: 10.1016/j.tiv.2020.104904
52. Wang RY, Abbott RD, Zieba A, Borowsky FE, Kaplan DL. Development of a Three-Dimensional Adipose Tissue Model for Studying Embryonic Exposures to Obesogenic Chemicals. Ann BioMed Eng (2017) 45:1807–18. doi: 10.1007/s10439-016-1752-x
53. Lee MJ, Fried SK. Optimal Protocol for the Differentiation and Metabolic Analysis of Human Adipose Stromal Cells. Methods Enzymol (2014) 538:49–65. doi: 10.1016/B978-0-12-800280-3.00004-9
54. Green H, Kehinde O. Spontaneous Heritable Changes Leading to Increased Adipose Conversion in 3T3 Cells. Cell (1976) 7:105–13. doi: 10.1016/0092-8674(76)90260-9
55. Green H, Kehinde O. Sublines of Mouse 3t3 Cells That Accumulate Lipid. Cell (1974) 1:113–6. doi: 10.1016/0092-8674(74)90126-3
56. Kramer AH, Joos-Vandewalle J, Edkins AL, Frost CL, Prinsloo E. Real-Time Monitoring of 3T3-L1 Preadipocyte Differentiation Using a Commercially Available Electric Cell-Substrate Impedance Sensor System. Biochem Biophys Res Commun (2014) 443:1245–50. doi: 10.1016/j.bbrc.2013.12.123
57. Pereira-Fernandes A, Vanparys C, Vergauwen L, Knapen D, Jorens PG, Blust R. Toxicogenomics in the 3T3-L1 Cell Line, a New Approach for Screening of Obesogenic Compounds. Toxicol Sci (2014) 140:352–63. doi: 10.1093/toxsci/kfu092
58. Kassotis CD, Masse L, Kim S, Schlezinger JJ, Webster TF, Stapleton HM. Characterization of Adipogenic Chemicals in Three Different Cell Culture Systems: Implications for Reproducibility Based on Cell Source and Handling. Sci Rep (2017) 7:42104. doi: 10.1038/srep42104
59. Reznikoff CA, Brankow DW, Heidelberger C. Establishment and Characterization of a Cloned Line of C3H Mouse Embryo Cells Sensitive to Postconfluence Inhibition of Division. Cancer Res (1973) 33:3231–8.
60. Lee N, Kim I, Park S, Han D, Ha S, Kwon M, et al. Creatine Inhibits Adipogenesis by Downregulating Insulin-Induced Activation of the Phosphatidylinositol 3-Kinase Signaling Pathway. Stem Cells Dev (2015) 24:983–94. doi: 10.1089/scd.2014.0130
61. Lane JM, Doyle JR, Fortin JP, Kopin AS, Ordovas JM. Development of an OP9 Derived Cell Line as a Robust Model to Rapidly Study Adipocyte Differentiation. PLoS One (2014) 9:e112123. doi: 10.1371/journal.pone.0112123
62. Wolins NE, Quaynor BK, Skinner JR, Tzekov A, Park C, Choi K, et al. OP9 Mouse Stromal Cells Rapidly Differentiate Into Adipocytes: Characterization of a Useful New Model of Adipogenesis. J Lipid Res (2006) 47:450–60. doi: 10.1194/jlr.D500037-JLR200
63. Ruiz-Ojeda FJ, Ruperez AI, Gomez-Llorente C, Gil A, Aguilera CM. Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. Int J Mol Sci (2016) 17:1040. doi: 10.3390/ijms17071040
64. Janderová L, McNeil M, Murrell AN, Mynatt RL, Smith SR. Human Mesenchymal Stem Cells as an In Vitro Model for Human Adipogenesis. Obes Res (2003) 11:65–74. doi: 10.1038/oby.2003.11
65. Janesick AS, Dimastrogiovanni G, Vanek L, Boulos C, Chamorro-Garcia R, Tang W, et al. On the Utility of ToxCast and ToxPi as Methods for Identifying New Obesogens. Environ Health Perspect (2016) 124:1214–26. doi: 10.1289/ehp.1510352
66. Hauner H, Entenmann G. Regional Variation of Adipose Differentiation in Cultured Stromal-Vascular Cells From the Abdominal and Femoral Adipose Tissue of Obese Women. Int J Obes (1991) 15:121–6.
67. Conget PA, Minguell JJ. Phenotypical and Functional Properties of Human Bone Marrow Mesenchymal Progenitor Cells. J Cell Physiol (1999) 181:67–73. doi: 10.1002/(SICI)1097-4652(199910)181:1<67::AID-JCP7>3.0.CO;2-C
68. Tremain N, Korkko J, Ibberson D, Kopen GC, DiGirolamo C, Phinney DG. MicroSAGE Analysis of 2,353 Expressed Genes in a Single Cell-Derived Colony of Undifferentiated Human Mesenchymal Stem Cells Reveals mRNAs of Multiple Cell Lineages. Stem Cells (2001) 19:408–18. doi: 10.1634/stemcells.19-5-408
69. Shoucri BM, Hung VT, Chamorro-Garcia R, Shioda T, Blumberg B. Retinoid X Receptor Activation During Adipogenesis of Female Mesenchymal Stem Cells Programs a Dysfunctional Adipocyte. Endocrinology (2018) 159:2863–83. doi: 10.1210/en.2018-00056
70. Shoucri BM, Martinez ES, Abreo TJ, Hung VT, Moosova Z, Shioda T, et al. Retinoid X Receptor Activation Alters the Chromatin Landscape To Commit Mesenchymal Stem Cells to the Adipose Lineage. Endocrinology (2017) 158:3109–25. doi: 10.1210/en.2017-00348
71. Vom Saal FS, Nagel SC, Coe BL, Angle BM, Taylor JA. The Estrogenic Endocrine Disrupting Chemical Bisphenol A (BPA) and Obesity. Mol Cell Endocrinol (2012) 354:74–84. doi: 10.1016/j.mce.2012.01.001
72. Yang M, Chen M, Wang J, Xu M, Sun J, Ding L, et al. Bisphenol A Promotes Adiposity and Inflammation in a Nonmonotonic Dose-Response Way in 5-Week-Old Male and Female C57BL/6J Mice Fed a Low-Calorie Diet. Endocrinology (2016) 157:2333–45. doi: 10.1210/en.2015-1926
73. Brulport A, Vaiman D, Chagnon MC, Le Corre L. Obesogen Effect of Bisphenol S Alters mRNA Expression and DNA Methylation Profiling in Male Mouse Liver. Chemosphere (2020) 241:125092. doi: 10.1016/j.chemosphere.2019.125092
74. Gao P, Wang L, Yang N, Wen J, Zhao M, Su G, et al. Peroxisome Proliferator-Activated Receptor Gamma (Pparγ) Activation and Metabolism Disturbance Induced by Bisphenol A and its Replacement Analog Bisphenol S Using In Vitro Macrophages and In Vivo Mouse Models. Environ Int (2020) 134:105328. doi: 10.1016/j.envint.2019.105328
75. Newbold RR, McLachlan JA. Vaginal Adenosis and Adenocarcinoma in Mice Exposed Prenatally or Neonatally to Diethylstilbestrol. Cancer Res (1982) 42:2003–11.
76. Newbold RR, Padilla-Banks E, Snyder RJ, Phillips TM, Jefferson WN. Developmental Exposure to Endocrine Disruptors and the Obesity Epidemic. Reprod Toxicol (2007) 23:290–6. doi: 10.1016/j.reprotox.2006.12.010
77. vonderEmbse AN, Elmore SE, Jackson KB, Habecker BA, Manz KE, Pennell KD, et al. Developmental Exposure to DDT or DDE Alters Sympathetic Innervation of Brown Adipose in Adult Female Mice. Environ Health (2021) 20:37. doi: 10.1186/s12940-021-00721-2
78. La Merrill M, Karey E, Moshier E, Lindtner C, La Frano MR, Newman JW, et al. Perinatal Exposure of Mice to the Pesticide DDT Impairs Energy Expenditure and Metabolism in Adult Female Offspring. PLoS One (2014) 9:e103337. doi: 10.1371/journal.pone.0103337
79. Chamorro-Garcia R, Shoucri BM, Willner S, Kach H, Janesick A, Blumberg B. Effects of Perinatal Exposure to Dibutyltin Chloride on Fat and Glucose Metabolism in Mice, and Molecular Mechanisms, in Vitro. Environ Health Perspect (2018) 126:057006. doi: 10.1289/EHP3030
80. Li X, Pham HT, Janesick AS, Blumberg B. Triflumizole is an Obesogen in Mice That Acts Through Peroxisome Proliferator Activated Receptor Gamma (PPARgamma). Environ Health Perspect (2012) 120:1720–6. doi: 10.1289/ehp.1205383
81. Regnier SM, Kirkley AG, Ye H, El-Hashani E, Zhang X, Neel BA, et al. Dietary Exposure to the Endocrine Disruptor Tolylfluanid Promotes Global Metabolic Dysfunction in Male Mice. Endocrinology (2015) 156:896–910. doi: 10.1210/en.2014-1668
82. Hao C, Cheng X, Guo J, Xia H, Ma X. Perinatal Exposure to Diethyl-Hexyl-Phthalate Induces Obesity in Mice. Front Biosci (Elite Ed) (2013) 5:725–33. doi: 10.2741/E653
83. Jackson TW, Ryherd GL, Scheibly CM, Sasser AL, Guillette TC, Belcher SM. Gestational Cd Exposure in the CD-1 Mouse Induces Sex-Specific Hepatic Insulin Insensitivity, Obesity, and Metabolic Syndrome in Adult Female Offspring. Toxicol Sci (2020) 178:264–80. doi: 10.1093/toxsci/kfaa154
84. Hsu JW, Nien CY, Chen HW, Tsai FY, Yeh SC, Kao YH, et al. Di(2-Ethylhexyl)Phthalate Exposure Exacerbates Metabolic Disorders in Diet-Induced Obese Mice. Food Chem Toxicol (2021) 156:112439. doi: 10.1016/j.fct.2021.112439
85. Somm E, Schwitzgebel VM, Toulotte A, Cederroth CR, Combescure C, Nef S, et al. Perinatal Exposure to Bisphenol a Alters Early Adipogenesis in the Rat. Environ Health Perspect (2009) 117:1549–55. doi: 10.1289/ehp.11342
86. de Araujo JFP, Podratz PL, Sena GC, Merlo E, Freitas-Lima LC, Ayub JGM, et al. The Obesogen Tributyltin Induces Abnormal Ovarian Adipogenesis in Adult Female Rats. Toxicol Lett (2018) 295:99–114. doi: 10.1016/j.toxlet.2018.06.1068
87. Punzon I, Latapie V, Le Mevel S, Hagneau A, Jolivet P, Palmier K, et al. Towards a Humanized PPARgamma Reporter System for In Vivo Screening of Obesogens. Mol Cell Endocrinol (2013) 374:1–9. doi: 10.1016/j.mce.2013.04.004
88. Green AJ, Hoyo C, Mattingly CJ, Luo Y, Tzeng JY, Murphy SK, et al. Cadmium Exposure Increases the Risk of Juvenile Obesity: A Human and Zebrafish Comparative Study. Int J Obes (Lond) (2018) 42:1285–95. doi: 10.1038/s41366-018-0036-y
89. Skinner MK. Environmental Epigenetics and a Unified Theory of the Molecular Aspects of Evolution: A Neo-Lamarckian Concept That Facilitates Neo-Darwinian Evolution. Genome Biol Evol (2015) 7:1296–302. doi: 10.1093/gbe/evv073
90. King SE, Skinner MK. Epigenetic Transgenerational Inheritance of Obesity Susceptibility. Trends Endocrinol Metab (2020) 31:478–94. doi: 10.1016/j.tem.2020.02.009
91. Lee MK, Blumberg B. Transgenerational Effects of Obesogens. Basic Clin Pharmacol Toxicol (2019) 125 Suppl 3:44–57. doi: 10.1111/bcpt.13214
92. Nilsson EE, Maamar MB, Skinner MK. Environmentally Induced Epigenetic Transgenerational Inheritance and the Weismann Barrier: The Dawn of Neo-Lamarckian Theory. J Dev Biol (2020) 8:28. doi: 10.3390/jdb8040028
93. Riu A, McCollum CW, Pinto CL, Grimaldi M, Hillenweck A, Perdu E, et al. Halogenated Bisphenol-A Analogs Act as Obesogens in Zebrafish Larvae (Danio Rerio). Toxicol Sci (2014) 139:48–58. doi: 10.1093/toxsci/kfu036
94. Ouadah-Boussouf N, Babin PJ. Pharmacological Evaluation of the Mechanisms Involved in Increased Adiposity in Zebrafish Triggered by the Environmental Contaminant Tributyltin. Toxicol Appl Pharmacol (2016) 294:32–42. doi: 10.1016/j.taap.2016.01.014
95. Baker KD, Thummel CS. Diabetic Larvae and Obese Flies-Emerging Studies of Metabolism in Drosophila. Cell Metab (2007) 6:257–66. doi: 10.1016/j.cmet.2007.09.002
96. Williams MJ, Wiemerslage L, Gohel P, Kheder S, Kothegala LV, Schioth HB. Dibutyl Phthalate Exposure Disrupts Evolutionarily Conserved Insulin and Glucagon-Like Signaling in Drosophila Males. Endocrinology (2016) 157:2309–21. doi: 10.1210/en.2015-2006
97. Chen MY, Liu HP, Cheng J, Chiang SY, Liao WP, Lin WY. Transgenerational Impact of DEHP on Body Weight of Drosophila. Chemosphere (2019) 221:493–9. doi: 10.1016/j.chemosphere.2018.12.193
98. Lee MC, Park JC, Lee JS. Effects of Environmental Stressors on Lipid Metabolism in Aquatic Invertebrates. Aquat Toxicol (2018) 200:83–92. doi: 10.1016/j.aquatox.2018.04.016
99. Lee MC, Han J, Lee SH, Kim DH, Kang HM, Won EJ, et al. A Brominated Flame Retardant 2,2',4,4' Tetrabrominated Diphenyl Ether (BDE-47) Leads to Lipogenesis in the Copepod Tigriopus Japonicus. Aquat Toxicol (2016) 178:19–26. doi: 10.1016/j.aquatox.2016.07.002
100. Jordao R, Garreta E, Campos B, Lemos MF, Soares AM, Tauler R, et al. Compounds Altering Fat Storage in Daphnia Magna. Sci Total Environ (2016) 545–546:127–36. doi: 10.1016/j.scitotenv.2015.12.097
101. Nedergaard J, Bengtsson T, Cannon B. Unexpected Evidence for Active Brown Adipose Tissue in Adult Humans. Am J Physiol Endocrinol Metab (2007) 293:E444–52. doi: 10.1152/ajpendo.00691.2006
102. Sacks H, Symonds ME. Anatomical Locations of Human Brown Adipose Tissue: Functional Relevance and Implications in Obesity and Type 2 Diabetes. Diabetes (2013) 62:1783–90. doi: 10.2337/db12-1430
103. Sanchez-Gurmaches J, Guertin DA. Adipocyte Lineages: Tracing Back the Origins of Fat. Biochim Biophys Acta (2014) 1842:340–51. doi: 10.1016/j.bbadis.2013.05.027
104. Lizcano F, Arroyave F. Control of Adipose Cell Browning and Its Therapeutic Potential. Metabolites (2020) 10:471. doi: 10.3390/metabo10110471
105. Cohen P, Kajimura S. The Cellular and Functional Complexity of Thermogenic Fat. Nat Rev Mol Cell Biol (2021) 22:393–409. doi: 10.1038/s41580-021-00350-0
106. Maliszewska K, Kretowski A. Brown Adipose Tissue and Its Role in Insulin and Glucose Homeostasis. Int J Mol Sci (2021) 22:1530.
107. Hanssen MJ, Hoeks J, Brans B, van der Lans AA, Schaart G, van den Driessche JJ, et al. Short-Term Cold Acclimation Improves Insulin Sensitivity in Patients With Type 2 Diabetes Mellitus. Nat Med (2015) 21:863–5. doi: 10.1038/nm.3891
108. Becher T, Palanisamy S, Kramer DJ, Eljalby M, Marx SJ, Wibmer AG, et al. Brown Adipose Tissue is Associated With Cardiometabolic Health. Nat Med (2021) 27:58–65. doi: 10.1038/s41591-020-1126-7
109. Wibmer AG, Becher T, Eljalby M, Crane A, Andrieu PC, Jiang CS, et al. Brown Adipose Tissue Is Associated With Healthier Body Fat Distribution and Metabolic Benefits Independent of Regional Adiposity. Cell Rep Med (2021) 2:100332. doi: 10.1016/j.xcrm.2021.100332
110. Di Gregorio I, Busiello RA, Burgos Aceves MA, Lepretti M, Paolella G, Lionetti L. Environmental Pollutants Effect on Brown Adipose Tissue. Front Physiol (2018) 9:1891. doi: 10.3389/fphys.2018.01891
111. Amir S, Shah STA, Mamoulakis C, Docea AO, Kalantzi OI, Zachariou A, et al. Endocrine Disruptors Acting on Estrogen and Androgen Pathways Cause Reproductive Disorders Through Multiple Mechanisms: A Review. Int J Environ Res Public Health (2021) 18:1464.
112. Brandner JM, Zorn-Kruppa M, Yoshida T, Moll I, Beck LA, De Benedetto A. Epidermal Tight Junctions in Health and Disease. Tissue Barriers (2015) 3:e974451. doi: 10.4161/21688370.2014.974451
113. Rehwagen C. WHO Recommends DDT to Control Malaria. BMJ (2006) 333:622. doi: 10.1136/bmj.333.7569.622-b
114. Yue L, Zhao W, Wang D, Meng M, Zheng Y, Li Y, et al. Silver Nanoparticles Inhibit Beige Fat Function and Promote Adiposity. Mol Metab (2019) 22:1–11. doi: 10.1016/j.molmet.2019.01.005
115. Tinkov AA, Aschner M, Ke T, Ferrer B, Zhou JC, Chang JS, et al. Adipotropic Effects of Heavy Metals and Their Potential Role in Obesity. Fac Rev (2021) 10:32. doi: 10.12703/r/10-32
116. Huff MO, Todd SL, Smith AL, Elpers JT, Smith AP, Murphy RD, et al. Arsenite and Cadmium Activate MAPK/ERK via Membrane Estrogen Receptors and G-Protein Coupled Estrogen Receptor Signaling in Human Lung Adenocarcinoma Cells. Toxicol Sci (2016) 152:62–71. doi: 10.1093/toxsci/kfw064
117. Bae J, Jang Y, Kim H, Mahato K, Schaecher C, Kim IM, et al. Arsenite Exposure Suppresses Adipogenesis, Mitochondrial Biogenesis and Thermogenesis via Autophagy Inhibition in Brown Adipose Tissue. Sci Rep (2019) 9:14464. doi: 10.1038/s41598-019-50965-9
118. Ro SH, Semple I, Ho A, Park HW, Lee JH. Sestrin2, a Regulator of Thermogenesis and Mitohormesis in Brown Adipose Tissue. Front Endocrinol (Lausanne) (2015) 6:114. doi: 10.3389/fendo.2015.00114
119. Bimonte VM, Besharat ZM, Antonioni A, Cella V, Lenzi A, Ferretti E, et al. The Endocrine Disruptor Cadmium: A New Player in the Pathophysiology of Metabolic Diseases. J Endocrinol Invest (2021) 44:1363–77. doi: 10.1007/s40618-021-01502-x
120. Surmi BK, Hasty AH. Macrophage Infiltration Into Adipose Tissue: Initiation, Propagation and Remodeling. Future Lipidol (2008) 3:545–56. doi: 10.2217/17460875.3.5.545
121. Peshdary V, Styles G, Rigden M, Caldwell D, Kawata A, Sorisky A, et al. Exposure to Low Doses of Dechlorane Plus Promotes Adipose Tissue Dysfunction and Glucose Intolerance in Male Mice. Endocrinology (2020) 161:bqaa096. doi: 10.1210/endocr/bqaa096
122. Kotzbeck P, Giordano A, Mondini E, Murano I, Severi I, Venema W, et al. Brown Adipose Tissue Whitening Leads to Brown Adipocyte Death and Adipose Tissue Inflammation. J Lipid Res (2018) 59:784–94. doi: 10.1194/jlr.M079665
123. Liu QS, Sun Z, Ren X, Ren Z, Liu A, Zhang J, et al. Chemical Structure-Related Adipogenic Effects of Tetrabromobisphenol A and Its Analogues on 3T3-L1 Preadipocytes. Environ Sci Technol (2020) 54:6262–71. doi: 10.1021/acs.est.0c00624
124. Kakutani H, Yuzuriha T, Akiyama E, Nakao T, Ohta S. Complex Toxicity as Disruption of Adipocyte or Osteoblast Differentiation in Human Mesenchymal Stem Cells Under the Mixed Condition of TBBPA and TCDD. Toxicol Rep (2018) 5:737–43. doi: 10.1016/j.toxrep.2018.06.007
125. Rendina-Ruedy E, Rosen CJ. Lipids in the Bone Marrow: An Evolving Perspective. Cell Metab (2020) 31:219–31. doi: 10.1016/j.cmet.2019.09.015
126. Henley DV, Korach KS. Endocrine-Disrupting Chemicals Use Distinct Mechanisms of Action to Modulate Endocrine System Function. Endocrinology (2006) 147:S25–32. doi: 10.1210/en.2005-1117
127. He B, Wang X, Jin X, Xue Z, Zhu J, Wang C, et al. Beta -Cypermethrin Promotes the Adipogenesis of 3T3-L1 Cells via Inducing Autophagy and Shaping an Adipogenesis-Friendly Microenvironment. Acta Biochim Biophys Sin (Shanghai) (2020) 52:821–31. doi: 10.1093/abbs/gmaa049
128. Rochester JR, Bolden AL. Bisphenol S and F: A Systematic Review and Comparison of the Hormonal Activity of Bisphenol A Substitutes. Environ Health Perspect (2015) 123:643–50. doi: 10.1289/ehp.1408989
129. Ramskov Tetzlaff CN, Svingen T, Vinggaard AM, Rosenmai AK, Taxvig C. Bisphenols B, E, F, and S and 4-Cumylphenol Induce Lipid Accumulation in Mouse Adipocytes Similarly to Bisphenol A. Environ Toxicol (2020) 35:543–52. doi: 10.1002/tox.22889
130. Reina-Perez I, Olivas-Martinez A, Mustieles V, Ruiz-Ojeda FJ, Molina-Molina JM, Olea N, et al. Bisphenol F and Bisphenol S Promote Lipid Accumulation and Adipogenesis in Human Adipose-Derived Stem Cells. Food Chem Toxicol (2021) 152:112216. doi: 10.1016/j.fct.2021.112216
131. Brulport A, Le Corre L, Maquart G, Barbet V, Dastugue A, Severin I, et al. Multigenerational Study of the Obesogen Effects of Bisphenol S After a Perinatal Exposure in C57BL6/J Mice Fed a High Fat Diet. Environ Pollut (2021) 270:116243. doi: 10.1016/j.envpol.2020.116243
132. Karbiener M, Pisani DF, Frontini A, Oberreiter LM, Lang E, Vegiopoulos A, et al. MicroRNA-26 Family is Required for Human Adipogenesis and Drives Characteristics of Brown Adipocytes. Stem Cells (2014) 32:1578–90. doi: 10.1002/stem.1603
133. Verbanck M, Canouil M, Leloire A, Dhennin V, Coumoul X, Yengo L, et al. Low-Dose Exposure to Bisphenols A, F and S of Human Primary Adipocyte Impacts Coding and non-Coding RNA Profiles. PLoS One (2017) 12:e0179583. doi: 10.1371/journal.pone.0179583
134. Cano-Sancho G, Salmon AG, La Merrill MA. Association Between Exposure to P,P'-DDT and Its Metabolite P,P'-DDE With Obesity: Integrated Systematic Review and Meta-Analysis. Environ Health Perspect (2017) 125:096002. doi: 10.1289/EHP527
135. Cao Q, Jing J, Cui X, Shi H, Xue B. Sympathetic Nerve Innervation is Required for Beigeing in White Fat. Physiol Rep (2019) 7:e14031. doi: 10.14814/phy2.14031
136. Li H, Li J, Qu Z, Qian H, Zhang J, Wang H, et al. Intrauterine Exposure to Low-Dose DBP in the Mice Induces Obesity in Offspring via Suppression of UCP1 Mediated ER Stress. Sci Rep (2020) 10:16360. doi: 10.1038/s41598-020-73477-3
137. Riu A, Grimaldi M, le Maire A, Bey G, Phillips K, Boulahtouf A, et al. Peroxisome Proliferator-Activated Receptor Gamma is a Target for Halogenated Analogs of Bisphenol a. Environ Health Perspect (2011) 119:1227–32. doi: 10.1289/ehp.1003328
138. Chappell VA, Janesick A, Blumberg B, Fenton SE. Tetrabromobisphenol-A Promotes Early Adipogenesis and Lipogenesis in 3T3-L1 Cells. Toxicol Sci (2018) 166:332–44. doi: 10.1093/toxsci/kfy209
139. Choi JJ, Eum SY, Rampersaud E, Daunert S, Abreu MT, Toborek M. Exercise Attenuates PCB-Induced Changes in the Mouse Gut Microbiome. Environ Health Perspect (2013) 121:725–30. doi: 10.1289/ehp.1306534
140. Verdin E. NAD(+) in Aging, Metabolism, and Neurodegeneration. Science (2015) 350:1208–13. doi: 10.1126/science.aac4854
141. Benzi A, Sturla L, Heine M, Fischer AW, Spinelli S, Magnone M, et al. CD38 Downregulation Modulates NAD(+) and NADP(H) Levels in Thermogenic Adipose Tissues. Biochim Biophys Acta Mol Cell Biol Lipids (2021) 1866:158819.
142. Ellero-Simatos S, Claus SP, Benelli C, Forest C, Letourneur F, Cagnard N, et al. Combined Transcriptomic-(1)H NMR Metabonomic Study Reveals That Monoethylhexyl Phthalate Stimulates Adipogenesis and Glyceroneogenesis in Human Adipocytes. J Proteome Res (2011) 10:5493–502. doi: 10.1021/pr200765v
143. Castriota F, Zushin PH, Sanchez SS, Phillips RV, Hubbard A, Stahl A, et al. Chronic Arsenic Exposure Impairs Adaptive Thermogenesis in Male C57BL/6J Mice. Am J Physiol Endocrinol Metab (2020) 318:E667–77. doi: 10.1152/ajpendo.00282.2019
144. Tonini C, Segatto M, Bertoli S, Leone A, Mazzoli A, Cigliano L, et al. Prenatal Exposure to BPA: The Effects on Hepatic Lipid Metabolism in Male and Female Rat Fetuses. Nutrients (2021) 13:1970.
145. Guo J, Zhang J, Wu C, Xiao H, Lv S, Lu D, et al. Urinary Bisphenol A Concentrations and Adiposity Measures at Age 7 Years in a Prospective Birth Cohort. Chemosphere (2020) 251:126340. doi: 10.1016/j.chemosphere.2020.126340
146. Robles-Aguilera V, Galvez-Ontiveros Y, Rodrigo L, Salcedo-Bellido I, Aguilera M, Zafra-Gomez A, et al. Factors Associated With Exposure to Dietary Bisphenols in Adolescents. Nutrients (2021) 13:1553.
147. Chamorro-Garcia R, Diaz-Castillo C, Shoucri BM, Kach H, Leavitt R, Shioda T, et al. Ancestral Perinatal Obesogen Exposure Results in a Transgenerational Thrifty Phenotype in Mice. Nat Commun (2017) 8:2012. doi: 10.1038/s41467-017-01944-z
148. Chamorro-Garcia R, Poupin N, Tremblay-Franco M, Canlet C, Egusquiza R, Gautier R, et al. Transgenerational Metabolomic Fingerprints in Mice Ancestrally Exposed to the Obesogen TBT. Environ Int (2021) 157:106822. doi: 10.1016/j.envint.2021.106822
149. King SE, Nilsson E, Beck D, Skinner MK. Adipocyte Epigenetic Alterations and Potential Therapeutic Targets in Transgenerationally Inherited Lean and Obese Phenotypes Following Ancestral Exposures. Adipocyte (2019) 8:362–78. doi: 10.1080/21623945.2019.1693747
150. Warner M, Ye M, Harley K, Kogut K, Bradman A, Eskenazi B. Prenatal DDT Exposure and Child Adiposity at Age 12: The CHAMACOS Study. Environ Res (2017) 159:606–12. doi: 10.1016/j.envres.2017.08.050
151. Horikoshi T, Nishimura T, Nomura Y, Iwabuchi T, Itoh H, Takizawa T, et al. Umbilical Cord Serum Concentrations of Perfluorooctane Sulfonate, Perfluorooctanoic Acid, and the Body Mass Index Changes From Birth to 5 1/2 Years of Age. Sci Rep (2021) 11:19789. doi: 10.1038/s41598-021-99174-3
152. Defois C, Ratel J, Garrait G, Denis S, Le Goff O, Talvas J, et al. Food Chemicals Disrupt Human Gut Microbiota Activity And Impact Intestinal Homeostasis As Revealed By In Vitro Systems. Sci Rep (2018) 8:11006. doi: 10.1038/s41598-018-29376-9
153. Chaplin A, Parra P, Laraichi S, Serra F, Palou A. Calcium Supplementation Modulates Gut Microbiota in a Prebiotic Manner in Dietary Obese Mice. Mol Nutr Food Res (2016) 60:468–80. doi: 10.1002/mnfr.201500480
154. David LA, Maurice CF, Carmody RN, Gootenberg DB, Button JE, Wolfe BE, et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature (2014) 505:559–63. doi: 10.1038/nature12820
155. Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An Obesity-Associated Gut Microbiome With Increased Capacity for Energy Harvest. Nature (2006) 444:1027–31. doi: 10.1038/nature05414
156. Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, et al. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage. Proc Natl Acad Sci USA (2004) 101:15718–23. doi: 10.1073/pnas.0407076101
157. Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, et al. A Core Gut Microbiome in Obese and Lean Twins. Nature (2009) 457:480–4. doi: 10.1038/nature07540
158. Turnbaugh PJ, Ridaura VK, Faith JJ, Rey FE, Knight R, Gordon JI. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci Trans Med (2009) 1:6ra14–4. doi: 10.1126/scitranslmed.3000322
159. Gore AC, Chappell VA, Fenton SE, Flaws JA, Nadal A, Prins GS, et al. EDC-2: The Endocrine Society's Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocrine Rev (2015) 36:E1–150. doi: 10.1210/er.2015-1010
160. Burks H, Pashos N, Martin E, McLachlan J, Bunnell B, Burow M. Endocrine Disruptors and the Tumor Microenvironment: A New Paradigm in Breast Cancer Biology. Mol Cell Endocrinol (2017) 457:13–9. doi: 10.1016/j.mce.2016.12.010
161. Hu W-Y, Shi G-B, Hu D-P, Nelles JL, Prins GS. Actions of Estrogens and Endocrine Disrupting Chemicals on Human Prostate Stem/Progenitor Cells and Prostate Cancer Risk. Mol Cell Endocrinol (2012) 354:63–73. doi: 10.1016/j.mce.2011.08.032
162. Sifakis S, Androutsopoulos VP, Tsatsakis AM, Spandidos DA. Human Exposure to Endocrine Disrupting Chemicals: Effects on the Male and Female Reproductive Systems. Environ Toxicol Pharmacol (2017) 51:56–70. doi: 10.1016/j.etap.2017.02.024
163. Liu Y, Yao Y, Li H, Qiao F, Wu J, Du Z-Y, et al. Influence of Endogenous and Exogenous Estrogenic Endocrine on Intestinal Microbiota in Zebrafish. PLoS One (2016) 11:e0163895–e0163895. doi: 10.1371/journal.pone.0163895
164. Jiao N, Baker SS, Nugent CA, Tsompana M, Cai L, Wang Y, et al. Gut Microbiome may Contribute to Insulin Resistance and Systemic Inflammation in Obese Rodents: A Meta-Analysis. Physiol Genomics (2018) 50:244–54. doi: 10.1152/physiolgenomics.00114.2017
165. Liang Y, Zhan J, Liu D, Luo M, Han J, Liu X, et al. Organophosphorus Pesticide Chlorpyrifos Intake Promotes Obesity and Insulin Resistance Through Impacting Gut and Gut Microbiota. Microbiome (2019) 7:19–9. doi: 10.1186/s40168-019-0635-4
166. Javurek AB, Spollen WG, Johnson SA, Bivens NJ, Bromert KH, Givan SA, et al. Effects of Exposure to Bisphenol A and Ethinyl Estradiol on the Gut Microbiota of Parents and Their Offspring in a Rodent Model. Gut Microbes (2016) 7:471–85. doi: 10.1080/19490976.2016.1234657
167. Lerner A, Matthias T. Changes in Intestinal Tight Junction Permeability Associated With Industrial Food Additives Explain the Rising Incidence of Autoimmune Disease. Autoimmun Rev (2015) 14:479–89. doi: 10.1016/j.autrev.2015.01.009
168. Csáki KF. Synthetic Surfactant Food Additives can Cause Intestinal Barrier Dysfunction. Med Hypotheses (2011) 76:676–81. doi: 10.1016/j.mehy.2011.01.030
169. Dirinck E, Jorens PG, Covaci A, Geens T, Roosens L, Neels H, et al. Obesity and Persistent Organic Pollutants: Possible Obesogenic Effect of Organochlorine Pesticides and Polychlorinated Biphenyls. Obesity (2011) 19:709–14. doi: 10.1038/oby.2010.133
170. Malaisé Y, Menard S, Cartier C, Gaultier E, Lasserre F, Lencina C, et al. Gut Dysbiosis and Impairment of Immune System Homeostasis in Perinatally-Exposed Mice to Bisphenol A Precede Obese Phenotype Development. Sci Rep (2017) 7:14472–2. doi: 10.1038/s41598-017-15196-w
171. Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, et al. Cross-Talk Between Akkermansia Muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc Natl Acad Sci USA (2013) 110:9066–71. doi: 10.1073/pnas.1219451110
172. Astbury S, Song A, Zhou M, Nielsen B, Hoedl A, Willing BP, et al. High Fructose Intake During Pregnancy in Rats Influences the Maternal Microbiome and Gut Development in the Offspring. Front Genet (2018) 9:203–3. doi: 10.3389/fgene.2018.00203
173. Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, et al. Selective Increases of Bifidobacteria in Gut Microflora Improve High-Fat-Diet-Induced Diabetes in Mice Through a Mechanism Associated With Endotoxaemia. Diabetologia (2007) 50:2374–83. doi: 10.1007/s00125-007-0791-0
174. Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic Endotoxemia Initiates Obesity and Insulin Resistance. Diabetes (2007) 56:1761–72. doi: 10.2337/db06-1491
175. Bian X, Chi L, Gao B, Tu P, Ru H, Lu K. The Artificial Sweetener Acesulfame Potassium Affects the Gut Microbiome and Body Weight Gain in CD-1 Mice. PLoS One (2017) 12:e0178426–e0178426. doi: 10.1371/journal.pone.0178426
176. Wang QP, Browman D, Herzog H, Neely GG. Non-Nutritive Sweeteners Possess a Bacteriostatic Effect and Alter Gut Microbiota in Mice. PLoS One (2018) 13:e0199080. doi: 10.1371/journal.pone.0199080
177. Suez J, Korem T, Zeevi D, Zilberman-Schapira G, Thaiss CA, Maza O, et al. Artificial Sweeteners Induce Glucose Intolerance by Altering the Gut Microbiota. Nature (2014) 514:181–6. doi: 10.1038/nature13793
178. Uebanso T, Ohnishi A, Kitayama R, Yoshimoto A, Nakahashi M, Shimohata T, et al. Effects of Low-Dose Non-Caloric Sweetener Consumption on Gut Microbiota in Mice. Nutrients (2017) 9:560. doi: 10.3390/nu9060560
179. Olivier-Van Stichelen S, Rother KI, Hanover JA. Maternal Exposure to Non-Nutritive Sweeteners Impacts Progeny's Metabolism and Microbiome. Front Microbiol (2019) 10:1360–0. doi: 10.3389/fmicb.2019.01360
180. Sánchez-Tapia M, Martínez-Medina J, Tovar AR, Torres N. Natural and Artificial Sweeteners and High Fat Diet Modify Differential Taste Receptors, Insulin, and TLR4-Mediated Inflammatory Pathways in Adipose Tissues of Rats. Nutrients (2019) 11:880. doi: 10.3390/nu11040880
181. Jin C, Zeng Z, Wang C, Luo T, Wang S, Zhou J, et al. Insights Into a Possible Mechanism Underlying the Connection of Carbendazim-Induced Lipid Metabolism Disorder and Gut Microbiota Dysbiosis in Mice. Toxicol Sci (2018) 166:382–93. doi: 10.1093/toxsci/kfy205
182. Liu P, Wu C-h, Chang X-l, Qi X-j, Zheng M-l, Zhou Z-j. Assessment of Chlorpyrifos Exposure and Absorbed Daily Doses Among Infants Living in an Agricultural Area of the Province of Jiangsu, China. Int Arch Occup Environ Health (2014) 87:753–62. doi: 10.1007/s00420-013-0918-1
183. Saunders M, Magnanti BL, Correia Carreira S, Yang A, Alamo-Hernández U, Riojas-Rodriguez H, et al. Chlorpyrifos and Neurodevelopmental Effects: A Literature Review and Expert Elicitation on Research and Policy. Environ Health (2012) 11:S5. doi: 10.1186/1476-069X-11-S1-S5
184. Zhang Y, Han S, Liang D, Shi X, Wang F, Liu W, et al. Prenatal Exposure to Organophosphate Pesticides and Neurobehavioral Development of Neonates: A Birth Cohort Study in Shenyang, China. PLoS One (2014) 9:e88491. doi: 10.1371/journal.pone.0088491
185. Li JW, Fang B, Pang GF, Zhang M, Ren FZ. Age- and Diet-Specific Effects of Chronic Exposure to Chlorpyrifos on Hormones, Inflammation and Gut Microbiota in Rats. Pestic Biochem Physiol (2019) 159:68–79. doi: 10.1016/j.pestbp.2019.05.018
186. O. World Health. Preventing Disease Through Healthy Environments: Exposure to Cadmium: A Major Public Health Concern. Geneva: World Health Organization (2019).
187. Ba Q, Li M, Chen P, Huang C, Duan X, Lu L, et al. Sex-Dependent Effects of Cadmium Exposure in Early Life on Gut Microbiota and Fat Accumulation in Mice. Environ Health Perspect (2017) 125:437–46. doi: 10.1289/EHP360
188. Lai K-P, Chung Y-T, Li R, Wan H-T, Wong CK-C. Bisphenol A Alters Gut Microbiome: Comparative Metagenomics Analysis. Environ Pollution (2016) 218:923–30. doi: 10.1016/j.envpol.2016.08.039
189. Xu J, Huang G, Nagy T, Teng Q, Guo TL. Sex-Dependent Effects of Bisphenol A on Type 1 Diabetes Development in non-Obese Diabetic (NOD) Mice. Arch Toxicol (2019) 93:997–1008. doi: 10.1007/s00204-018-2379-5
190. Petriello MC, Hoffman JB, Vsevolozhskaya O, Morris AJ, Hennig B. Dioxin-Like PCB 126 Increases Intestinal Inflammation and Disrupts Gut Microbiota and Metabolic Homeostasis. Environ Pollut (2018) 242:1022–32. doi: 10.1016/j.envpol.2018.07.039
191. Choi YJ, Seelbach MJ, Pu H, Eum SY, Chen L, Zhang B, et al. Polychlorinated Biphenyls Disrupt Intestinal Integrity via NADPH Oxidase-Induced Alterations of Tight Junction Protein Expression. Environ Health Perspect (2010) 118:976–81. doi: 10.1289/ehp.0901751
192. Hoffman JB, Flythe MD, Hennig B. Environmental Pollutant-Mediated Disruption of Gut Microbial Metabolism of the Prebiotic Inulin. Anaerobe (2019) 55:96–102. doi: 10.1016/j.anaerobe.2018.11.008
193. Trainic M, Flores JM, Pinkas I, Pedrotti ML, Lombard F, Bourdin G, et al. Airborne Microplastic Particles Detected in the Remote Marine Atmosphere. Commun Earth Environ (2020) 1:64. doi: 10.1038/s43247-020-00061-y
194. Barboza LGA, Cunha SC, Monteiro C, Fernandes JO, Guilhermino L, Bisphenol A. And its Analogs in Muscle and Liver of Fish From the North East Atlantic Ocean in Relation to Microplastic Contamination. Exposure Risk to Hum Consumers. J Hazard Mater (2020) 393:122419. doi: 10.1016/j.jhazmat.2020.122419
195. Takdastan A, Niari MH, Babaei A, Dobaradaran S, Jorfi S, Ahmadi M. Occurrence and Distribution of Microplastic Particles and the Concentration of Di 2-Ethyl Hexyl Phthalate (DEHP) in Microplastics and Wastewater in the Wastewater Treatment Plant. J Environ Manage (2020) 280:111851. doi: 10.1016/j.jenvman.2020.111851
196. Lu L, Wan Z, Luo T, Fu Z, Jin Y. Polystyrene Microplastics Induce Gut Microbiota Dysbiosis and Hepatic Lipid Metabolism Disorder in Mice. Sci Total Environ (2018), 631–632:449–58. doi: 10.1016/j.scitotenv.2018.03.051
197. Schneeberger M, Everard A, Gómez-Valadés AG, Matamoros S, Ramírez S, Delzenne NM, et al. Akkermansia Muciniphila Inversely Correlates With the Onset of Inflammation, Altered Adipose Tissue Metabolism and Metabolic Disorders During Obesity in Mice. Sci Rep (2015) 5:16643. doi: 10.1038/srep16643
198. Lustig RH. Fructose: Metabolic, Hedonic, and Societal Parallels With Ethanol. J Am Diet Assoc (2010) 110:1307–21. doi: 10.1016/j.jada.2010.06.008
199. Lustig RH, Mulligan K, Noworolski SM, Tai VW, Wen MJ, Erkin-Cakmak A, et al. Isocaloric Fructose Restriction and Metabolic Improvement in Children With Obesity and Metabolic Syndrome. Obes (Silver Spring) (2016) 24:453–60. doi: 10.1002/oby.21371
200. Zhao S, Jang C, Liu J, Uehara K, Gilbert M, Izzo L, et al. Dietary Fructose Feeds Hepatic Lipogenesis via Microbiota-Derived Acetate. Nature (2020) 579:586–91. doi: 10.1038/s41586-020-2101-7
201. Do MH, Lee E, Oh MJ, Kim Y, Park HY. High-Glucose or -Fructose Diet Cause Changes of the Gut Microbiota and Metabolic Disorders in Mice Without Body Weight Change. Nutrients (2018) 10:761. doi: 10.3390/nu10060761
202. Moon JS, Lee S, Park MA, Siempos II, M. Haslip, Lee PJ, et al. UCP2-Induced Fatty Acid Synthase Promotes NLRP3 Inflammasome Activation During Sepsis. J Clin Invest (2015) 125:665–80. doi: 10.1172/JCI78253
203. Rosas-Villegas A, Sánchez-Tapia M, Avila-Nava A, Ramírez V, Tovar AR, Torres N. Differential Effect of Sucrose and Fructose in Combination With a High Fat Diet on Intestinal Microbiota and Kidney Oxidative Stress. Nutrients (2017) 9:393. doi: 10.3390/nu9040393
204. Yücel G, Zhao Z, El-Battrawy I, Lan H, Lang S, Li X, et al. Lipopolysaccharides Induced Inflammatory Responses and Electrophysiological Dysfunctions in Human-Induced Pluripotent Stem Cell Derived Cardiomyocytes. Sci Rep (2017) 7:2935. doi: 10.1038/s41598-017-03147-4
205. Patil R, Arvindekar A. Glycation of Gut Proteins Initiates Microbial Dysbiosis and can Promote Establishment of Diabetes in Experimental Animals. Microb Pathog (2021) 152:104589. doi: 10.1016/j.micpath.2020.104589
206. Balda MS, Flores-Maldonado C, Cereijido M, Matter K. Multiple Domains of Occludin Are Involved in the Regulation of Paracellular Permeability. J Cell Biochem (2000) 78:85–96. doi: 10.1002/(SICI)1097-4644(20000701)78:1<85::AID-JCB8>3.0.CO;2-F
207. Nagpal R, Newman TM, Wang S, Jain S, Lovato JF, Yadav H. Obesity-Linked Gut Microbiome Dysbiosis Associated With Derangements in Gut Permeability and Intestinal Cellular Homeostasis Independent of Diet. J Diabetes Res (2018) 2018:3462092–3462092. doi: 10.1155/2018/3462092
208. Bao Z, Zhao Y, Wu A, Lou Z, Lu H, Yu Q, et al. Sub-Chronic Carbendazim Exposure Induces Hepatic Glycolipid Metabolism Disorder Accompanied by Gut Microbiota Dysbiosis in Adult Zebrafish (Daino Rerio). Sci Total Environ (2020) 739:140081. doi: 10.1016/j.scitotenv.2020.140081
209. Joly C, Gay-Quéheillard J, Léké A, Chardon K, Delanaud S, Bach V, et al. Impact of Chronic Exposure to Low Doses of Chlorpyrifos on the Intestinal Microbiota in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME®) and in the Rat. Environ Sci Pollution Res (2013) 20:2726–34. doi: 10.1007/s11356-012-1283-4
210. Zhao Y, Zhang Y, Wang G, Han R, Xie X. Effects of Chlorpyrifos on the Gut Microbiome and Urine Metabolome in Mouse (Mus Musculus). Chemosphere (2016) 153:287–93. doi: 10.1016/j.chemosphere.2016.03.055
211. Tsatsaronis JA, Walker MJ, Sanderson-Smith ML. Host Responses to Group A Streptococcus: Cell Death and Inflammation. PLoS Pathog (2014) 10:e1004266. doi: 10.1371/journal.ppat.1004266
212. Cox LM, Yamanishi S, Sohn J, Alekseyenko AV, Leung JM, Cho I, et al. Altering the Intestinal Microbiota During a Critical Developmental Window has Lasting Metabolic Consequences. Cell (2014) 158:705–21. doi: 10.1016/j.cell.2014.05.052
213. Pantoja-Feliciano IG, Clemente JC, Costello EK, Perez ME, Blaser MJ, Knight R, et al. Biphasic Assembly of the Murine Intestinal Microbiota During Early Development. ISME J (2013) 7:1112–5. doi: 10.1038/ismej.2013.15
214. Sela DA, Chapman J, Adeuya A, Kim JH, Chen F, Whitehead TR, et al. The Genome Sequence of Bifidobacterium Longum Subsp. Infantis Reveals Adaptations for Milk Utilization Within the Infant Microbiome. Proc Natl Acad Sci USA (2008) 105:18964–9. doi: 10.1073/pnas.0809584105
215. Kovatcheva-Datchary P, Nilsson A, Akrami R, Lee YS, De Vadder F, Arora T, et al. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated With Increased Abundance of Prevotella. Cell Metab (2015) 22:971–82. doi: 10.1016/j.cmet.2015.10.001
216. Yang J, Chen W, Sun Y, Liu J, Zhang W. Effects of Cadmium on Organ Function, Gut Microbiota and its Metabolomics Profile in Adolescent Rats. Ecotoxicol Environ Saf (2021) 222:112501. doi: 10.1016/j.ecoenv.2021.112501
217. Rice P, Aungst J. Updated Review of Literature and Data on Bisphenol A (CAS RN 80-05-7). (2014). p. 2014.
218. Bohr URM, Glasbrenner B, Primus A, Zagoura A, Wex T, Malfertheiner P. Identification of Enterohepatic Helicobacter Species in Patients Suffering From Inflammatory Bowel Disease. J Clin Microbiol (2004) 42:2766–8. doi: 10.1128/JCM.42.6.2766-2768.2004
219. Khokhlova EV, Smeianov VV, Efimov BA, Kafarskaia LI, Pavlova SI, Shkoporov AN. Anti-Inflammatory Properties of Intestinal Bifidobacterium Strains Isolated From Healthy Infants. Microbiol Immunol (2012) 56:27–39. doi: 10.1111/j.1348-0421.2011.00398.x
220. Linares R, Fernandez MF, Gutierrez A, Garcia-Villalba R, Suarez B, Zapater P, et al. Endocrine Disruption in Crohn's Disease: Bisphenol A Enhances Systemic Inflammatory Response in Patients With Gut Barrier Translocation of Dysbiotic Microbiota Products. FASEB J (2021) 35:e21697.
221. Catron T, Keely S, Brinkman N, Zurlinden T, Wood C, Wright J, et al. Host Developmental Toxicity of BPA and BPA Alternatives Is Inversely Related to Microbiota Disruption in Zebrafish. Toxicological Sci Off J Soc Toxicol (2018) 167:468–83.
222. Wang Y, Wang B, Wang Q, Liu Y, Liu X, Wu B, et al. Intestinal Toxicity and Microbial Community Disorder Induced by Bisphenol F and Bisphenol S in Zebrafish. Chemosphere (2021) 280:130711. doi: 10.1016/j.chemosphere.2021.130711
223. Kaur S, Sarma SJ, Marshall BL, Liu Y, Kinkade JA, Bellamy MM, et al. Developmental Exposure of California Mice to Endocrine Disrupting Chemicals and Potential Effects on the Microbiome-Gut-Brain Axis at Adulthood. Sci Rep (2020) 10:10902. doi: 10.1038/s41598-020-67709-9
224. Boucher O, Muckle G, Jacobson JL, Carter RC, Kaplan-Estrin M, Ayotte P, et al. Domain-Specific Effects of Prenatal Exposure to PCBs, Mercury, and Lead on Infant Cognition: Results From the Environmental Contaminants and Child Development Study in Nunavik. Environ Health Perspect (2014) 122:310–6. doi: 10.1289/ehp.1206323
225. Chi Y, Lin Y, Zhu H, Huang Q, Ye G, Dong S. PCBs–high-Fat Diet Interactions as Mediators of Gut Microbiota Dysbiosis and Abdominal Fat Accumulation in Female Mice. Environ Pollution (2018) 239:332–41. doi: 10.1016/j.envpol.2018.04.001
226. Crovesy L, Ostrowski M, Ferreira DMTP, Rosado EL, Soares-Mota M. Effect of Lactobacillus on Body Weight and Body Fat in Overweight Subjects: A Systematic Review of Randomized Controlled Clinical Trials. Int J Obes (2017) 41:1607–14. doi: 10.1038/ijo.2017.161
227. Cheng SL, Li X, Lehmler H-J, Phillips B, Shen D, Cui JY. Gut Microbiota Modulates Interactions Between Polychlorinated Biphenyls and Bile Acid Homeostasis. Toxicological Sci (2018) 166:269–87. doi: 10.1093/toxsci/kfy208
228. Pedersen HK, Gudmundsdottir V, Nielsen HB, Hyotylainen T, Nielsen T, Jensen BAH, et al. Human Gut Microbes Impact Host Serum Metabolome and Insulin Sensitivity. Nature (2016) 535:376–81. doi: 10.1038/nature18646
229. Iljazovic A, Roy U, Gálvez EJC, Lesker TR, Zhao B, Gronow A, et al. Perturbation of the Gut Microbiome by Prevotella Spp. Enhances Host Susceptibility to Mucosal Inflammation. Mucosal Immunol (2020) 14:113–24. doi: 10.1038/s41385-020-0296-4
230. Gourronc FA, Robertson LW, Klingelhutz AJ. A Delayed Proinflammatory Response of Human Preadipocytes to PCB126 is Dependent on the Aryl Hydrocarbon Receptor. Environ Sci Pollut Res Int (2018) 25:16481–92. doi: 10.1007/s11356-017-9676-z
231. Wahlang B, Alexander NC 2nd, Li X, Rouchka EC, Kirpich IA, Cave MC. Polychlorinated Biphenyls Altered Gut Microbiome in CAR and PXR Knockout Mice Exhibiting Toxicant-Associated Steatohepatitis. Toxicol Rep (2021) 8:536–47. doi: 10.1016/j.toxrep.2021.03.010
232. Larsen TM, Toubro S, Astrup A. PPARgamma Agonists in the Treatment of Type II Diabetes: Is Increased Fatness Commensurate With Long-Term Efficacy? Int J Obes (2003) 27:147–61. doi: 10.1038/sj.ijo.802223
233. Janani C, Ranjitha Kumari BD. PPAR Gamma Gene – A Review. Diabetes Metab Syndrome: Clin Res Rev (2015) 9:46–50. doi: 10.1016/j.dsx.2014.09.015
234. Wang CH, Wei YH. Therapeutic Perspectives of Thermogenic Adipocytes in Obesity and Related Complications. Int J Mol Sci (2021) 22:7177.
235. Torres-Fuentes C, Schellekens H, Dinan T, Cryan J. The Microbiota–Gut–Brain Axis in Obesity. Lancet Gastroenterol Hepatol (2017) 2:747–56. doi: 10.1016/S2468-1253(17)30147-4
236. Gupta A, Osadchiy V, Mayer EA. Brain–gut–microbiome Interactions in Obesity and Food Addiction. Nat Rev Gastroenterol Hepatol (2020) 17:655–72. doi: 10.1038/s41575-020-0341-5
Keywords: EDC, MDC, obesity, endocrine disrupting chemical, obesogens, adipogenesis, metabolism disrupting chemicals
Citation: Mohajer N, Du CY, Checkcinco C and Blumberg B (2021) Obesogens: How They Are Identified and Molecular Mechanisms Underlying Their Action. Front. Endocrinol. 12:780888. doi: 10.3389/fendo.2021.780888
Received: 21 September 2021; Accepted: 23 October 2021;
Published: 25 November 2021.
Edited by:
Marco Aurelio Romano, State University of Midwest Paraná, BrazilReviewed by:
Ronald Biemann, University Medical Center Leipzig, GermanyPatricia Cristina Lisboa, Rio de Janeiro State University, Brazil
Copyright © 2021 Mohajer, Du, Checkcinco and Blumberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bruce Blumberg, Ymx1bWJlcmdAdWNpLmVkdQ==