 Sophia Kristina Rupp
Sophia Kristina Rupp Ellen Wölk
Ellen Wölk Andreas Stengel
Andreas Stengel- 1Department of Psychosomatic Medicine and Psychotherapy, University Hospital Tübingen, Tübingen, Germany
- 2Charité Center for Internal Medicine and Dermatology, Department for Psychosomatic Medicine, Charité - Universitätsmedizin Berlin, Corporate Member of Freie Universität Berlin, Humboldt-Universität zu Berlin and Berlin Institute of Health, Berlin, Germany
Background: Nesfatin-1 is an 82-amino acid polypeptide, cleaved from the 396-amino acid precursor protein nucleobindin-2 (NUCB2) and discovered in 2006 in the rat hypothalamus. In contrast to the growing body of evidence for the pleiotropic effects of the peptide, the receptor mediating these effects and the exact signaling cascades remain still unknown.
Methods: This systematic review was conducted using a search in the Embase, PubMed, and Web of Science databases. The keywords “nesfatin-1” combined with “receptor”, “signaling”, “distribution”, “pathway”, g- protein coupled receptor”, and “binding” were used to identify all relevant articles reporting about potential nesfatin-1 signaling and the assumed mediation via a Gi protein-coupled receptor.
Results: Finally, 1,147 articles were found, of which 1,077 were excluded in several steps of screening, 70 articles were included in this systematic review. Inclusion criteria were studies investigating nesfatin-1’s putative receptor or signaling cascade, observational preclinical and clinical studies, experimental studies, registry-based studies, cohort studies, population-based studies, and studies in English language. After screening for eligibility, the studies were assigned to the following subtopics and discussed regarding intracellular signaling of nesfatin-1 including the potential receptor mediating these effects and downstream signaling of the peptide.
Conclusion: The present review sheds light on the various effects of nesfatin-1 by influencing several intracellular signaling pathways and downstream cascades, including the peptide’s influence on various hormones and their receptors. These data point towards mediation via a Gi protein-coupled receptor. Nonetheless, the identification of the nesfatin-1 receptor will enable us to better investigate the exact mediating mechanisms underlying the different effects of the peptide along with the development of agonists and antagonists.
1 Introduction
Since the discovery of nesfatin-1 in 2006 in the rat hypothalamus (1), there has been a vast amount of research to further explore the pleiotropic physiological functions of this peptide. Nesfatin-1 is an 82-amino acid polypeptide generated via post-translational processing of hypothalamic nucleobindin-2 (NUCB2), a protein of 396 amino acids, whose sequence is highly conserved from fish to mammals (2) pointing towards its physiological relevance. The peptide is predominantly localized in food intake-regulatory nuclei e.g., the arcuate nucleus (Arc), the paraventricular nucleus (PVN) and the nucleus of the solitary tract (3, 4). Accordingly, the peptide became first known for its anorexigenic effects; however, subsequent studies unraveled its many other properties, such as cardiovascular effects, lipid metabolism, reproduction functions, and emotion- related functions (5–7). Subsequently, the peptides’ central part, nesfatin-130-59, which has been identified as the active core of the peptide, was reported to reduce food intake after intracerebroventricular (icv) injection in mice (8, 9) and in rats (10).
However, multiple studies have indicated that nesfatin-1 is also secreted by peripheral tissues such as adipose tissue, gastric mucosa, pancreatic endocrine beta cells and testes and its expression level was found to be 20 times higher in endocrine cells of the oxyntic gastric mucosa than in the brain (11–13).
Numerous studies have reported multifunctional effects of nesfatin-1: peripheral nesfatin-1 affects glucose homeostasis (14) and shows anti-apoptotic and anti-inflammatory effects (15). In addition, peripherally administered nesfatin-1 induces vasoconstriction and elevates blood pressure (16), and moreover decreases antral and duodenal motility (17) and reduces gastric emptying (18). Taken together, the role of nesfatin-1 goes far beyond its initially observed function as a negative modulator of food intake.
In contrast to the growing body of evidence of the pleiotropic peptide’s effects, the receptor mediating these effects is still largely unknown. Although several studies suggest that the peptide activates extracellular and intracellular regulatory pathways involving multiple putative binding sites (19), a specific nesfatin-1 receptor has not yet been identified. Therefore, the present paper was designed to systematically review the findings about nesfatin-1 signaling, especially by focusing on its potential receptor. This systematic review will discuss the state of knowledge on nesfatin-1 signaling, the putative mediation via a Gi protein-coupled receptor and highlight respective direct and indirect evidence. Lastly, gaps in knowledge will be highlighted to encourage further research hopefully leading to the identification of the nesfatin-1 receptor.
2 Methods
We followed the preferred reporting items for systematic reviews and meta-analyses (PRISMA) (20) to report the results of this review.
2.1 Search Strategy
This article is a systematic review; articles related to the topic, were searched, collected, and screened. Articles released from the earliest day of publication to the day the search was performed were included. The search was conducted on November 22nd in 2020. We searched Embase, PubMed and Web of Science using the following keywords: “nesfatin-1” combined with “receptor”, “signaling”, “distribution”, “pathway”, G-protein coupled receptor”, and “binding” (Table 1). All studies that contained material applicable to the topic were considered. We investigated the reference lists of the included studies to find other potential articles for inclusion. Local experts in the field were consulted for additional studies. Retrieved manuscripts were extracted using a standardized collection tool. Gray literature was not considered due to lack of essential information usually affecting this type of research. There was no date (all studies until November 2020) or species restriction in the search, but the search was limited to texts in English.

Table 1 The search strategy of this review.
2.2 Inclusion and Exclusion Criteria
The inclusion criteria were as follows: (1) observational preclinical and clinical studies, experimental studies, registry-based studies, cohort studies, population-based studies; (2) Studies that were written in English (3) Studies investigating nesfatin-1 putative receptor or signaling cascade.
The exclusion criteria were as follows: (1) Review articles, surveys, case reports, comments, letters, conference abstracts or posters, and economic evaluation; (2) Studies for which abstracts or full-text articles were not available; (3) Studies that were not available in English; (4) Studies with absence of outcome data.
2.3 Study Selection and Data Extraction
One reviewer screened all titles and abstracts retrieved from the electronic searches to identify potentially eligible articles. Full texts of the potentially eligible articles were retrieved. Two reviewers (S.R. and E.W.) screened all full text articles and potentially eligible or unclear full-text articles, determined whether they were eligible or not eligible and recorded the reason for exclusion. Any disagreements between both reviewers were resolved through discussion.
The quality of the studies included in this review was assessed with respect to risk of bias within individual and across studies by thoroughly evaluating the study designs, selection of population/species, methodological procedures applied as well as presentation of the results. Following a full-text review of the eligible studies, one reviewer (E.W.) extracted the relevant data. From each included study the following information was extracted: first author of the publication, year of publication, title, population/species, size of the respective population/species, study type, research question/purpose, and key findings (Table 2).
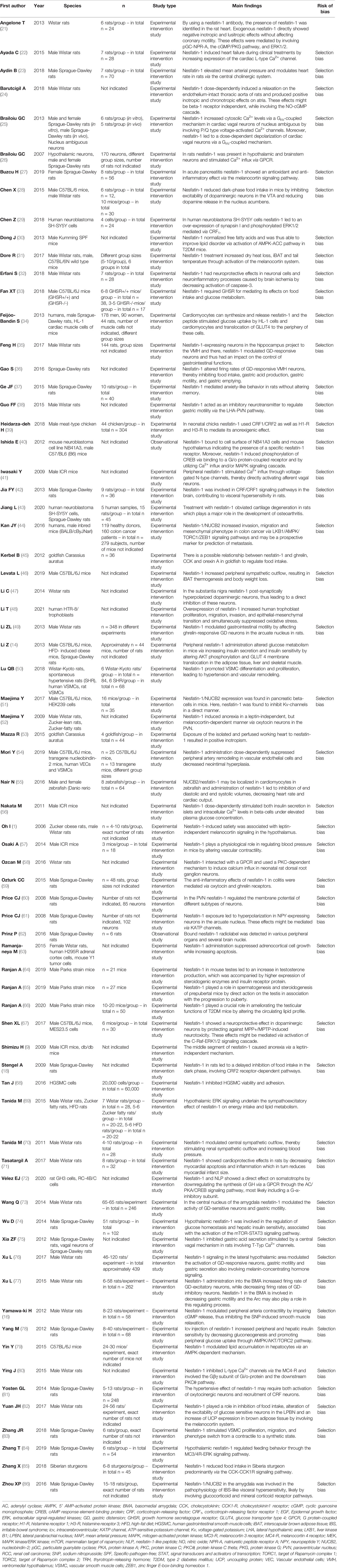
Table 2 Main findings of articles (in alphabetical order) included and discussed in this systematic review.
3 Results
1,147 articles were identified after searching the databases using the keywords mentioned above. Reviews, surveys, case reports, comments, letters, conference abstracts or posters, as well as economic evaluation and non-English language studies or studies with no full text available, and duplicates were excluded. This deceased the number to 425 articles. Next, title and abstract were screened and studies which deviated from the main topic were excluded; thus, 128 studies required full-text screening. Here, studies not related to the review topic were excluded. Ultimately, 70 articles were selected for this systematic review. The PRISMA flow diagram schematically depicts the article selection process (Figure 1). Table 2 shows the main results of these articles.
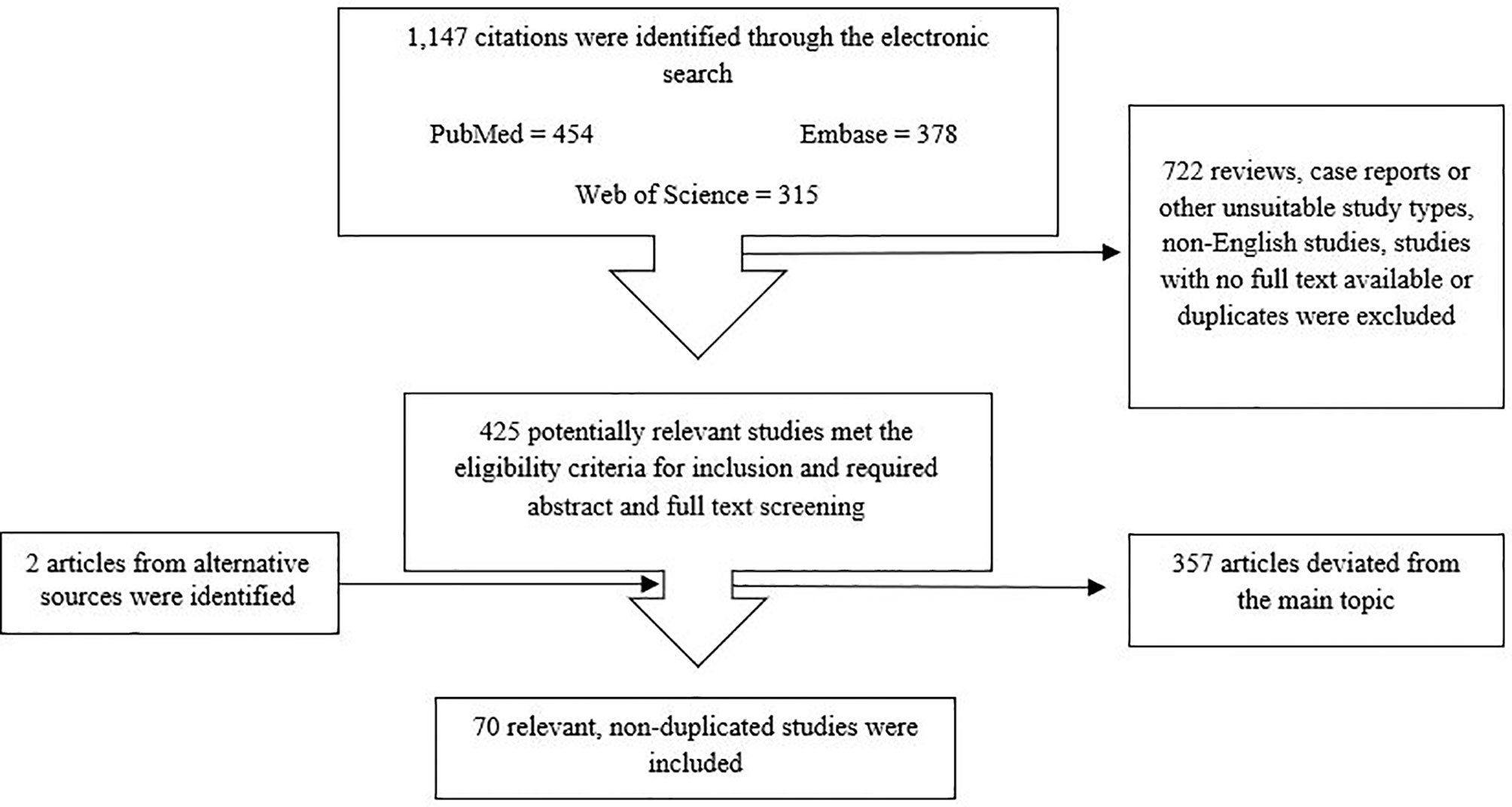
Figure 1 Flowchart for article screening and selection.
3.1 Quality Assessment
The current review consists of observational preclinical and clinical studies or experimental studies, assuming a risk for bias. There are several issues that may contribute to limited comparability between studies and thus limited transferability of study results in general. First, the studies included different species or population, which may lead to limited comparability. Second, the respective study populations differed in sample size, while the study methods, treatments and observation periods used were also very heterogeneous. Furthermore, we only reviewed the abstracts and full texts published in English. Studies in other languages have not been included. Additionally, we searched only three databases for potentially eligible studies. Taken together, bias cannot be ruled out for the studies included in this systematic review; thus, these limitations should be kept in mind when interpreting the results discussed here.
4 Discussion
The aim of this systematic review was to identify and summarize the state of knowledge on signaling cascades of nesfatin-1 and to shed light on the peptide’s potential receptor. Nesfatin-1 affects multiple sites in the organism and elicits a variety of intracellular effects, which accounts for the pleiotropic nature of this peptide. The multiple intracellular signaling cascades triggered by nesfatin-1 (Figure 2) only reinforce the multifaceted nature of this peptide and highlight that it may be of great importance to understand the exact cascades by further investigating the particular type and localization of the potential receptor in the future.
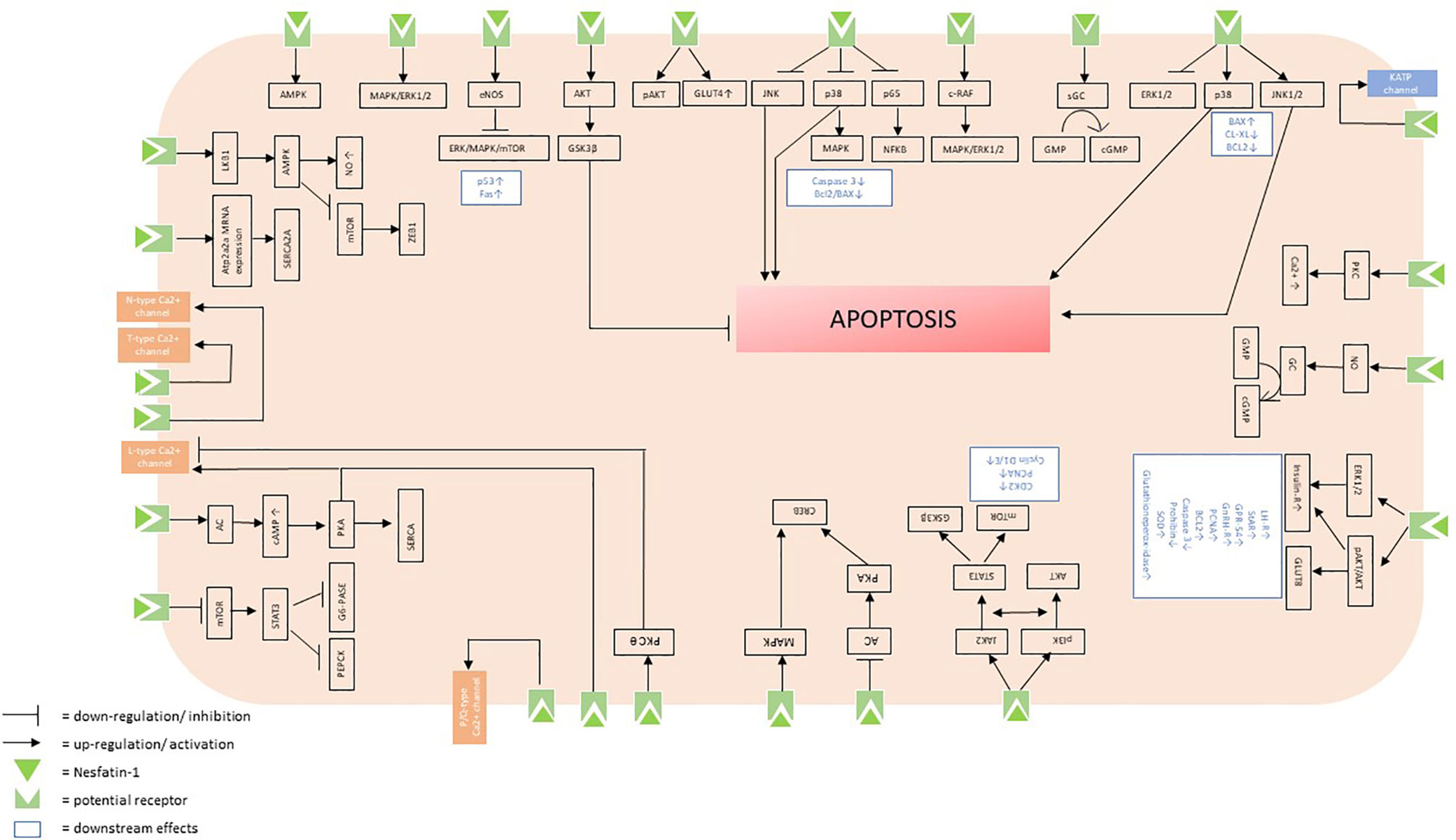
Figure 2 Putative intracellular signaling cascades initiated by the binding of nesfatin-1 to its receptor.
4.1 Intracellular Signaling and Potential Receptor
By using autoradiography binding of radiolabeled nesfatin-1 in the gastric mucosa of corpus and antrum, in duodenum, jejunum and ileum and centrally in the cortex, paraventricular nucleus of the hypothalamus, area postrema, dorsal motor nucleus of the vagus nerve and cerebellum was shown, giving rise to respective expression sited of the receptor (62). Since nesfatin-1 is known to be able to cross the blood-brain barrier, peripheral nesfatin-1 might access and then bind to these central receptors (87). In addition, NUCB2/Nesfatin-1 was found to be widely distributed in the central nervous system of mice (88) and rats (3, 89).
4.1.1 AMPK Signaling Pathway
Converging evidence points towards a mediation of nesfatin-1’s effects via a G-protein coupled receptor: Previous results suggest the presence of a nesfatin-1 receptor in hepatocytes whose activation stimulates the phosphorylation of AMPK, reducing hepatic lipid accumulation. These alterations were associated with a significant attenuation of peroxisome proliferator-activated receptor γ (PPARγ) and sterol-regulatory element-binding protein-1 (SREBP1), which are known lipogenesis-related transcription factors (79). Furthermore, in diabetic mice low-dose injection of nesfatin-1 also regulated fatty acid metabolism via activation of an AMPK-ACC pathway (30). The assumption of an influence of nesfatin-1 on AMPK signaling cascades is consistent with recent findings, highlighting that nesfatin-1/NUCB2 enhances invasion, migration, and mesenchymal phenotype in colon cancer via liver kinase B1 (LKB1)/AMPK/target of rapamycin kinase complex I (TORC1)/ZEB1 signaling pathways (44). In line with this, it was further shown that nesfatin-1 suppresses peripheral artery remodeling likely by increasing NO production and LKB1-mediated activation of AMPK in vascular endothelial cells in mice (54). However, in the testis of mice nesfatin-1-induces an increase in G protein-coupled receptor 54 (GPR-54) accompanied by increases in PCNA, Bcl2, androgen receptor (AR), GLUT8, insulin receptor and gonadotropin-releasing hormone receptor (GnRH-R) (64, 65). In addition, a potential role of protein kinase B (AKT) as a signaling mechanism for nesfatin-1-induced glucose uptake and activation of the AMP-activated protein kinase (AMPK)/extracellular signal-regulated kinase 1/2 (ERK1/2) system was found through which nesfatin-1 enhances insulin sensitivity in the testis of mice (64, 66). Supporting these findings, icv injection of nesfatin-1 increased peripheral and hepatic insulin sensitivity by decreasing gluconeogenesis and promoting peripheral glucose uptake through the AMPK/AKT/TORC2 pathway (78).
4.1.2 MAPK Signaling Pathway
Furthermore, previous results assumed the presence of a nesfatin-1-specific-receptor, likely a Gi/o protein-coupled receptor, on the cell surface of a murine neuroblastoma cell line (NB41A3 cells) and mouse hypothalamus. Here, binding of nesfatin-1 activates the cAMP-response reporter (CRE) and further phosphorylates CREB in these cells by utilizing Ca2+ influx and/or the MAPK signaling pathway (40). Other studies also point to a crucial role of nesfatin-1 in the ERK/MAPK/mechanistic Target of Rapamycin (mTOR) signaling pathway by upregulating endothelial NO synthase (eNOS) levels in human gastrointestinal smooth muscle cells (HGSMC). Here, nesfatin-1 increases pro-apoptotic factors p53 and Fas, thereby accelerating apoptosis in HGSMC. Furthermore, nesfatin-1 led to reduced expression of ERK1/2, p38, MAPK and mTOR in HGSMC (68). Moreover, in adrenocortical cells nesfatin-1 increases apoptosis by involving ERK1/2, p38, c-Jun N-terminal kinase 1/2 (JNK1/2) signaling pathways, Bcl-2-associated X protein (Bax), B-cell lymphoma-extra-large (BCL-XL), and Bcl-2 genes (63). In accordance, nesfatin-1 decreased apoptosis rate in rat chondrocytes also through the Bax/Bcl-2 signaling pathway and further through suppressing both NF-κB and MAPK signaling pathways (43). Furthermore, it was found that nesfatin-1 ameliorates levels of human gene for type H procollagen (CoI2a1) while it decreases the IL-1beta induced expression of matrix metalloproteinases (MMP), a disintegrin and metalloproteinase with thrombospondin motifs 5 (ADAMTS5), cyclooxygenase 2 (COX-2), caspase-3, nitric oxide (NO), inducible nitric oxide synthase (iNOS), prostaglandin E2 (PGE2), and interleukin-6. The nesfatin-1-induced decrease of caspase-3 activity was also detected in the hippocampus accompanied by fewer ionized calcium binding adaptor molecule 1 (Iba-1)-positive cells (32). Interestingly, consecutive intraperitoneal (ip) administration of nesfatin-1 for three weeks decreases brain-derived neurotrophic factor (BDNF) and phosphorylated-ERK levels in the hippocampus and prefrontal cortex (PFC), thereby downregulating the ERK signaling pathway (37). In accordance, it was demonstrated that nesfatin-1 protects dopaminergic neurons from 1-Methyl-4-phenylpyridinium (MPP+)- and 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridin (MPTP)-induced neurotoxicity both in vivo and in vitro, probably via activation of the C-Raf/ERK1/2 signaling cascade (67). Regarding nesfatin-1’s influence on the ERK signaling cascade, it was further found that the peptide regulates the autonomic nervous system through ERK signaling in corticotropin-releasing factor (CRF) positive neurons in the paraventricular nucleus (PVN). Precisely, central nesfatin-1 administration stimulated MAPK activity thereby enhancing ERK1/2 phosphorylation in CRF neurons of the PVN (69). Consistent with these findings, also in human neuroblastoma SH-SY5Y cells, it was observed that the stimulatory effects of nesfatin-1 on synapsin I expression are mediated by CRF1 through the cAMP/MAPK/ERK pathway (29).
4.1.3 CRF Signaling Pathway
Regarding nesfatin-1’s influence on CRF signaling pathways, gastric distension (GD)-responsive neurons were found in ventromedial hypothalamic nucleus (VMH), whose firing rate was altered by nesfatin-1, most likely through interactions with CRF signaling pathways (35). This hypothesis was strengthened by the observed association of nesfatin-1 and irritable bowel syndrome-like visceral hypersensitivity that is also mediated, at least in part, by CRF/CRF1 signaling pathways in the brain (42). In consistence with these findings, it was reported that nesfatin-1 reduced dark-phase food intake via CRF pathways (18). Furthermore, in neonatal chicks nesfatin-1 used CRF1/CRF2 as well as histamine (H)1 and H3 receptors to mediate its anorexigenic effect. Interestingly, co-injection of nesfatin-1 and chlorpheniramine (H1 antagonist) blunted nesfatin-1-induced hypophagia, while co-administration with thioperamide (H3 antagonist) reinforced the hypophagic effect of nesfatin-1. This divergence in the effects of H1 and H3 receptors can be explained by the fact that H1 is a postsynaptic receptor, while H3 is a presynaptic autoreceptor, leading to the hypothesis that H1 and H3 play a different regulatory role in the feeding behavior influenced by nesfatin-1 (39).
4.1.4 Melanocortin Signaling Pathway
Using the melanocortin signaling pathway, nesfatin-1 was found to alter firing rates of GD-responsive neurons in the VMH (36), baso-medial amygdala (BMA) (77), arcuate nucleus (49) and central nucleus of the amygdala (73). Also, by involving the melanocortin system nesfatin-1 alters the excitability of glucose sensitive neurons in the lateral parabrachial nucleus (LPBN) and increases the level of uncoupling protein (UCP) expression in brown adipose tissue (BAT) (82). In accordance, it was assumed that nesfatin-1-induced satiety is associated with leptin-independent melanocortin signaling in the hypothalamus (1). This hypothesis was strengthened by a study showing that hypothalamic nesfatin-1 regulates feeding behavior through the MC3/4-ERK signaling pathway. Interestingly, in this study, hypothalamic nesfatin-1 mediates its effects via MC3/4, while not altering expression of MC3/4 (84). Furthermore, through activation of the melanocortin system icv nesfatin-1 treatment increases dry heat loss, interscapular (i) BAT and tail temperature. Moreover, nesfatin-1 administration upregulated the expression of POMC and MC3 mRNA in the hypothalamus along with an elevation of iodothyronine deiodinase 2 (Dio2), UCP1 and PPARγ-1 alpha mRNA in the iBAT (31). Furthermore, the nesfatin-1-induced elevation in the mRNA expression of the cAMP-responsive gene Dio2 strengthens the hypothesis of an activation of the β-adrenergic/cAMP signaling pathway, which is in line with other findings emphasizing the established role of nesfatin-1 in sympathetic nerve activity by activating the central melanocortin system (70). On the other hand, only the midsegment of nesfatin-1 proves effective and leads to a higher activation of c-Fos in the brainstem nucleus of the solitary tract (NTS). Furthermore, administration of the middle segment of nesfatin-1 results in an increase in the expression of proopiomelanocortin (POMC), cocaine- and amphetamine-regulated transcript (CART) mRNA in the NTS (9). In adult ventricular myocytes nesfatin-1 targets MC4, sequentially coupling to the Gβγ subunits of Gi/o, leading to the subsequent activation of the novel protein kinase C (PKC) θ isoform, subsequently resulting in an inhibition of L-Type Ca2+ channels associated with a hyperpolarizing shift in the voltage-dependence of inactivation. Notable the nesfatin-1 mediated inhibition of calcium-influx was not affected by Kt-5720, a PKA antagonist indicating that the cAMP/PKA pathway is not involved in the nesfatin-1-induced L-Type Ca2+ channel response in ventricular myocytes (80). In contrast, it was observed that both nesfatin-1 and nesfatin-1-like peptide directly affect somatotrophs via binding to a GPCR containing a G-α-i subunit and utilizing the adenylyl cyclase (AC)/protein kinase A (PKA)/CREB signaling pathway subunit, thereby downregulating the synthesis of ghrelin hormone (72).
4.1.5 Ion Currents
However, in cardiac vagal neurons nesfatin-1 may increase cytosolic Ca2+ levels via a Gi/o-coupled mechanism by involving P/Q-type voltage-activated Ca2+ channels (25), while in hypothalamic neurons the peptide might mediate Ca2+ influx also via a GPCR, most likely by affecting both L- and P/Q- Ca2+ channels. Surprisingly, Ca2+ influx was significantly reduced by a PKA blocker, indicating an involvement of PKA in hypothalamic neurons (26). The effects of nesfatin-1 in sensory neurons may be mediated through the participation of a Gi/o protein coupled receptor as it is speculated that the nesfatin-1-induced Ca2+ increase may result from a Ca2+ influx from both extracellular and intracellular sources (58). Furthermore, several other studies proposed an influence of nesfatin-1 on calcium levels in various tissues: Exposure of the isolated and perfused working heart to nesfatin-1 results in positive inotropism mediated through cAMP, PKA, L-type Ca2+ channels, sarcoplasmic/endoplasmic reticulum calcium ATPase 2a (SERCA2a) pumps, ERK1/2 and phospholamban (PLN) (53). On the other hand, it was found that nesfatin-1 leads to increased expression of Atp2a2a mRNA encoding SERCA2a, while there were no changes observed in the expression of ryanodine receptor 1b (RyR1b) encoding mRNA (55). Furthermore, a different study demonstrated that nesfatin-1 induces heart failure during clinical treatments by increasing expression of the cardiac L-type Ca2+ channel a1c subunit (22). Likewise, it was shown that in mice, nesfatin-1 dose-dependently stimulates intracellular Ca2+ levels via L-type Ca2+ channels independently of PKA and phospholipase A2 (PLA2) in beta-cells under elevated plasma glucose concentration (56). One mechanism of the elevated Ca2+ influx could be an increase in membrane Na+ permeability, which depolarizes the membrane to open voltage-gated calcium channels (VDCC) (90), a mechanism used by glucagon-like peptide 1 (GLP-1) (91). Another potential explanation is an involvement of protein kinase C, which has been reported to increase glucose-stimulated Ca2+ influx through VDCC and insulin secretion (92). In contrast, another study reported that peripheral nesfatin-1 stimulates Ca2+ influx through voltage-gated N-type channels, thereby directly activating afferent vagal neurons (41). On the other hand, nesfatin-1 showed an effect on vagal neurons by mediating Ca2+ signaling through T-type channels, which are low-voltage activated channels localized in different areas of the central nervous system (75, 93, 94). The ultimate role of T-type Ca2+ channel activation in the response of vagal neurons to nesfatin-1 and the downstream target/effect of this signaling pathway remain unknown.
However, nesfatin-1 not only regulates the calcium influx via various mechanisms, but also influences the levels of other ions: More precisely, nesfatin-1 directly induces an inhibition of the voltage-dependent potassium (Kv) current by directly binding to Kv2.1 channels to exert its effect on pancreatic beta cells, as these Kv channels have been identified as the major contributors to Kv currents in these cells. Since nesfatin-1 is localized in beta cells, it has been hypothesized that nesfatin-1 may affect beta cell function in an autocrine/paracrine manner (51). Another finding assumes that nesfatin-1 activates KATP channels, thereby inhibiting orexigenic neuropeptide Y (NPY) neurons of the arcuate nucleus and consequently leading to satiety (61).
4.1.6 NO-cGMP System
Interestingly, nesfatin-1 also targets the NO-cGMP system; again, the particular receptor(s) is/are still unknown and require further research: In atrial tissue nesfatin-1 might increase the sensitivity of smooth muscle to cGMP-mediated relaxing mechanisms rather than increasing the NO levels. Interestingly, these positive chronotropic effects in the atrial tissue were independent of the β1-adrenergic receptor (24). On the other hand, nesfatin-1 also impairs the SNP-induced cGMP production, thereby inhibiting the NO donor-induced smooth muscle relaxations. Since these effects were persistent in the presence of IBMX, a phosphodiesterase (PDE) inhibitor, it can be suggested that these effects may be mediated via inhibition of soluble guanylate cyclase activity rather than activation of PDEs (16). However, other findings indicated that by recruiting guanylyl cyclase-linked receptors, namely natriuretic peptide receptor type A (NPR-A), and thereby involving the cGMP/protein kinase G pathway and ERK1/2, nesfatin-1 mediates negative inotropic and lusitropic effects in rats. Since the use of pertussistoxin did not inhibit the effects of nesfatin-1, the hypothesis that nesfatin-1 binds to a Gi/o cannot be confirmed by these observations (21).
4.1.7 AKT Signaling Pathway
Also involved in multiple aspects of the effects of nesfatin-1 is the AKT signaling pathway: In the myocardium of mice with myocardial infection, nesfatin-1 increases expressions of phosphorylated-AKT/AKT and phosphorylated-glycogen synthase kinase 3 beta (GSK-3β)/GSK-3β, thereby protecting cardiac tissue (71). Other findings indicate that overexpression of nesfatin-1 in human trophoblasts influences the expression of phosphoinositide-3-kinase (PI3K)/AKT/mTOR and AKT/GSK3β pathway, contributing to trophoblast dysfunction simultaneously suppressing oxidative stress by reducing reactive oxygen species (ROS), malondialdehyde (MDA), and superoxide dismutase (SOD) levels (48). Furthermore, in vascular smoot muscle cells (VSMC) nesfatin-1 modulates the crosstalk between PI3K/AKT/mTOR and Janus kinase 2 (JAK2)/STAT3 signaling, leading to hypertension and vascular remodeling (50). Nesfatin-1 also dose-dependently increased MMP-2 and MMP-9 levels, while it decreased PPARγ gene expression in VSMCS contributing to vascular remodeling and neointimal hyperplasia (83). Regarding STAT3 signaling, hypothalamic nesfatin-1 in rats activates the mTOR-STAT3 pathway, thereby regulating glucose homeostasis and hepatic insulin sensitivity (74). Moreover, studies involving HL-1 cells and cardiomyocytes indicate a nesfatin-1-induced phosphorylation of ERK1/2, AKT and the substrate of AKT, AS160, thereby stimulating peripheral GLUT-4 translocation (34). These findings are in line with the results of a recent study reporting nesfatin-1 to affect glucose metabolism by affecting AKT phosphorylation and GLUT4 membrane translocation in adipose tissue, liver, and skeletal muscle (14). The specific receptor to which nesfatin-1 binds to trigger these effects is still unknown.
4.2 Colocalization With Other Peptides
Furthermore, PVN neurons in rats are directly depolarized by nesfatin-1, suggesting a mediation of these effects through the activation of a GPCR. Since PVN neurons respond to nesfatin-1 and further produce nesfatin-1, there might be potential interactions between various subgroups of PVN neurons engaged in the control of several autonomic outputs. The specificity of the effects may also arise from the co-expression of other receptors on nesfatin-1-sensitive neurons. An alternative explanation is that multiple nesfatin-1 receptors exist that are separately responsible for hyperpolarizing or depolarizing responses (60). It was shown that in the PVN, nesfatin-1 targets magnocellular and parvocellular Oxt neurons as well as nesfatin-1 neurons themselves, further stimulating Oxt release. Furthermore, the presence of nesfatin-1 specifically in the secretory vesicles of PVN neurons was found, indicating paracrine/autocrine actions of nesfatin-1 (52). On the other hand, it was suggested that the hypertensive effect of nesfatin-1 may require both activation of Oxt neurons and recruitment of CRF neurons (81). Moreover, by targeting ghrelin and Oxt receptors nesfatin-1 suppresses neutrophil infiltration and improves glutathione levels in acute pancreatitis (27) and colitis (59). Interestingly, the effects of nesfatin-1 on myeloperoxidase activity, lipid peroxidation and glutathione level were abrogated by application of melanocortin receptor antagonist. Despite previously demonstrating that nesfatin-1 utilizes the melanocortin signaling pathway for its effects on food intake, these findings indicate that nesfatin-1 may also utilize the receptors mediating its anorexigenic effect for anti-inflammatory effects (59). Since the effects of nesfatin-1 on GD- excitatory neurons and GD-inhibitory neurons in the PVN were attenuated in the presence of H4928, an Oxt receptor antagonist, it can be assumed that nesfatin-1involves Oxt receptors to modulate gastric function (38). Moreover, in goldfish nesfatin-1 inhibits ghrelin mRNA expression in the brain and vice versa, leading to the assumption of a negative feedback inhibition of endogenous ghrelin and nesfatin-1 (45). Another study indicates that peripheral nesfatin-1 most likely exerts part of its effect directly via ghrelin receptor (GHSR) signaling. Furthermore, the results here show that the effects of nesfatin-1 in mice fed a normal chow diet, such as improvement of glucose tolerance, upregulation and phosphorylation of AKT-kinase mRNA and GLUT4 membrane translocation, were dependent on the presence of GHSR. In high-fat diet (HFD) fed mice, nesfatin-1 additionally led to a raise of AKT levels in liver tissues, which is also a GHSR-dependent mechanism (33). Therefore, it is possible that nesfatin-1 acts as an endogenous inverse agonist of the GHSR and influences the structure or activity of the GHSR. Furthermore, it was found that nesfatin-1 inhibits the excitability of dopaminergic neurons in the ventral tegmental area (VTA) (28) and the substantia nigra (47), thereby decreasing dopamine release (28). In addition, nesfatin-1 signaling in the lateral hypothalamic area (LHA) or electric stimulation of the Arc modulate the activation of GD-responsive neurons, gastric motility and gastric secretion involving melanin-concentrating hormone signaling indicating the potential presence of nesfatin-1 specific receptors in these neurons (76). Recent findings also provide evidence that nesfatin-1 administered into the amygdala targets glucocorticoid and mineral corticoid receptor pathways, thereby being involved in the pathophysiology of irritable bowel syndrome (IBS)-like visceral hypersensitivity (86). Lastly, predominantly via the CCK-CCK1R signaling pathway nesfatin-1 reduces food intake in Siberia sturgeon (85).
Involving cholinergic pathways, central administration of nesfatin-1 elevates mean arterial pressure and modulates heart rate in rats. These cardiovascular effects were mediated by both nicotinic and muscarinic receptors. Considering that recent studies have shown that both muscarinic and nicotinic acetylcholine receptors interact with G-coupled proteins (95), it may be that nesfatin-1 mediates its cardiovascular effects via central muscarinic and nicotinic receptors because of its affinity to GPCRs (23).
Furthermore, it was found that central nesfatin-1 can increase peripheral sympathetic outflow and thus β-adrenergic activation, resulting in iBAT thermogenesis and body weight loss. It was demonstrated that the thermogenetic effect of nesfatin-1 mainly depends on β3-adrenergic stimulation. In addition, levels of Dio2 and cell death inducing DFFA like effector A (CIDEA) mRNA were increased in brown adipose tissue after nesfatin-1 administration, which is plausible as both are involved in regulating the thermogenic program (46). Since the direct effects of peripherally injected nesfatin-1 on vascular smooth muscle were attenuated by pretreatment with propranolol, an involvement of the β-adrenergic system can also be suspected here (57).
4.3 Implications for Future Research
The described various distribution of the nesfatin-1 receptor further supports the assumption that nesfatin-1 may be involved in the regulation of various homeostatic functions, and the various signaling pathways underlying the actions of nesfatin-1 emphasize its multiple effects. The recent discoveries about the diverse effects of nesfatin-1 indicate the importance of evaluating and determining the potential use of nesfatin-1 in a therapeutic context in the future. For this reason, further research is needed to fully understand the exact cascades of the peptide and its complex interplay with other hormones.
5 Conclusion
This present review highlights nesfatin-1 as a pleotropic peptide that acts at multiple levels in the organism, thereby eliciting a wide variety of effects. As diverse as the effects of the peptide, so are the intracellular signaling pathways and downstream effects summarized here, including the influence of nesfatin-1 on various hormones and their receptors. The identification of the so far unknown nesfatin-1 receptor will represent a major leap forward in our understanding of the physiology of nesfatin-1 and will allow us to better investigate the precise mechanisms underlying the many different effects of the peptide. Further research is needed so we might be able to consider a therapeutic use of nesfatin-1 in the future.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author Contributions
EW performed the systematic search. SR and EW screened the papers and SR wrote the first draft of the manuscript. AS planned the study and gave critical input throughout the study. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We acknowledge support by Deutsche Forschungsgemeinschaft and the Open Access Publishing Fund of the University of Tübingen.
References
1. Oh-I S, Shimizu H, Satoh T, Okada S, Adachi S, Inoue K, et al. Identification of Nesfatin-1 as a Satiety Molecule in the Hypothalamus. Nature (2006) 443(7112):709–12. doi: 10.1038/nature05162
2. Dore R, Levata L, Lehnert H, Schulz C. Nesfatin-1: Functions and Physiology of a Novel Regulatory Peptide. J Endocrinol (2017) 232(1):R45–65. doi: 10.1530/JOE-16-0361
3. Goebel M, Stengel A, Wang L, Lambrecht NW, Taché Y. Nesfatin-1 Immunoreactivity in Rat Brain and Spinal Cord Autonomic Nuclei. Neurosci Lett (2009) 452(3):241–6. doi: 10.1016/j.neulet.2009.01.064
4. Shimizu H, Oh-I S, Okada S, Mori M. Nesfatin-1: An Overview and Future Clinical Application. Endocr J (2009) 56(4):537–43. doi: 10.1507/endocrj.k09e-117
5. Prinz P, Stengel A. Nesfatin-1: Current Status as a Peripheral Hormone and Future Prospects. Curr Opin Pharmacol (2016) 31:19–24. doi: 10.1016/j.coph.2016.08.011
6. Gao X, Zhang K, Song M, Li X, Luo L, Tian Y, et al. Role of Nesfatin-1 in the Reproductive Axis of Male Rat. Sci Rep (2016) 6:32877. doi: 10.1038/srep32877
7. Wei Y, Li J, Wang H, Wang G. NUCB2/nesfatin-1: Expression and Functions in the Regulation of Emotion and Stress. Prog Neuropsychopharmacol Biol Psychiatry (2018) 81:221–7. doi: 10.1016/j.pnpbp.2017.09.024
8. Stengel A, Goebel-Stengel M, Wang L, Kato I, Mori M, Taché Y. Nesfatin-1(30-59) But Not the N- and C-Terminal Fragments, Nesfatin-1(1-29) and Nesfatin-1(60-82) Injected Intracerebroventricularly Decreases Dark Phase Food Intake by Increasing Inter-Meal Intervals in Mice. Peptides (2012) 35(2):143–8. doi: 10.1016/j.peptides.2012.03.015
9. Shimizu H, Oh-I S, Hashimoto K, Nakata M, Yamamoto S, Yoshida N, et al. Peripheral Administration of Nesfatin-1 Reduces Food Intake in Mice: The Leptin-Independent Mechanism. Endocrinology (2009) 150(2):662–71. doi: 10.1210/en.2008-0598
10. Prinz P, Teuffel P, Lembke V, Kobelt P, Goebel-Stengel M, Hofmann T, et al. Nesfatin-130-59 Injected Intracerebroventricularly Differentially Affects Food Intake Microstructure in Rats Under Normal Weight and Diet-Induced Obese Conditions. Front Neurosci (2015) 9:422. doi: 10.3389/fnins.2015.00422
11. Stengel A, Taché Y. Nesfatin-1-Role as Possible New Potent Regulator of Food Intake. Regul Pept (2010) 163(1-3):18–23. doi: 10.1016/j.regpep.2010.05.002
12. Mohan H, Unniappan S. Ontogenic Pattern of Nucleobindin-2/Nesfatin-1 Expression in the Gastroenteropancreatic Tissues and Serum of Sprague Dawley Rats. Regul Pept (2012) 175(1-3):61–9. doi: 10.1016/j.regpep.2012.02.006
13. Stengel A, Goebel M, Yakubov I, Wang L, Witcher D, Coskun T, et al. Identification and Characterization of Nesfatin-1 Immunoreactivity in Endocrine Cell Types of the Rat Gastric Oxyntic Mucosa. Endocrinology (2009) 150(1):232–8. doi: 10.1210/en.2008-0747
14. Li Z, Gao L, Tang H, Yin Y, Xiang X, Li Y, et al. Peripheral effects of nesfatin-1 on glucose homeostasis. PLoS One (2013) 8(8):e71513. doi: 10.1371/journal.pone.0071513
15. Özsavcí D, Erşahin M, Şener A, Özakpinar ÖB, Toklu HZ, Akakín D, et al. The Novel Function of Nesfatin-1 as an Anti-Inflammatory and Antiapoptotic Peptide in Subarachnoid Hemorrhage-Induced Oxidative Brain Damage in Rats. Neurosurgery (2011) 68(6):1699–708. doi: 10.1227/NEU.0b013e318210f258 discussion 1708.
16. Yamawaki H, Takahashi M, Mukohda M, Morita T, Okada M, Hara Y. A Novel Adipocytokine, Nesfatin-1 Modulates Peripheral Arterial Contractility and Blood Pressure in Rats. Biochem Biophys Res Commun (2012) 418(4):676–81. doi: 10.1016/j.bbrc.2012.01.076
17. Atsuchi K, Asakawa A, Ushikai M, Ataka K, Tsai M, Koyama K, et al. Centrally Administered Nesfatin-1 Inhibits Feeding Behaviour and Gastroduodenal Motility in Mice. Neuroreport (2010) 21(15):1008–11. doi: 10.1097/WNR.0b013e32833f7b96
18. Stengel A, Goebel M, Wang L, Rivier J, Kobelt P, Mönnikes H, et al. Central Nesfatin-1 Reduces Dark-Phase Food Intake and Gastric Emptying in Rats: Differential Role of Corticotropin-Releasing Factor2 Receptor. Endocrinology (2009) 150(11):4911–9. doi: 10.1210/en.2009-0578
19. Angelone T, Rocca C, Pasqua T. Nesfatin-1 in Cardiovascular Orchestration: From Bench to Bedside. Pharmacol Res (2020) 156:104766. doi: 10.1016/j.phrs.2020.104766
20. Moher D, Liberati A, Tetzlaff J, Altman DG, PRISMA Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. BMJ (2009) 339:b2535. doi: 10.1136/bmj.b2535
21. Angelone T, Filice E, Pasqua T, Amodio N, Galluccio M, Montesanti G, et al. Nesfatin-1 as a Novel Cardiac Peptide: Identification, Functional Characterization, and Protection Against Ischemia/Reperfusion Injury. Cell Mol Life Sci (2013) 70(3):495–509. doi: 10.1007/s00018-012-1138-7
22. Ayada C, Turgut G, Turgut S. The Effect of Nesfatin-1 on Heart L-Type Ca²⁺ Channel α1c Subunit in Rats Subjected to Chronic Restraint Stress. Bratisl Lek Listy (2015) 116(5):326–9. doi: 10.4149/bll_2015_061
23. Aydin B, Guvenc G, Altinbas B, Niaz N, Yalcin M. Modulation of Nesfatin-1-Induced Cardiovascular Effects by the Central Cholinergic System. Neuropeptides (2018) 70:9–15. doi: 10.1016/j.npep.2018.05.001
24. Barutcigil A, Tasatargil A. Effects of Nesfatin-1 on Atrial Contractility and Thoracic Aorta Reactivity in Male Rats. Clin Exp Hypertens (2018) 40(5):414–20. doi: 10.1080/10641963.2017.1384487
25. Brailoiu GC, Deliu E, Tica AA, Rabinowitz JE, Tilley DG, Benamar K, et al. Nesfatin-1 Activates Cardiac Vagal Neurons of Nucleus Ambiguus and Elicits Bradycardia in Conscious Rats. J Neurochem (2013) 126(6):739–48. doi: 10.1111/jnc.12355
26. Brailoiu GC, Dun SL, Brailoiu E, Inan S, Yang J, Chang JK, et al. Nesfatin-1: Distribution and Interaction With a G Protein-Coupled Receptor in the Rat Brain. Endocrinology (2007) 148(10):5088–94. doi: 10.1210/en.2007-0701
27. Buzcu H, Ozbeyli D, Yuksel M, Cilingir Kaya OT, Kasimay Cakir O. Nesfatin-1 Protects From Acute Pancreatitis: Role of Melanocortin Receptors. J Physiol Pharmacol (2019) 70(6):839–48. doi: 10.26402/jpp.2019.6.03
28. Chen X, Shu X, Cong ZK, Jiang ZY, Jiang H. Nesfatin-1 Acts on the Dopaminergic Reward Pathway to Inhibit Food Intake. Neuropeptides (2015) 53:45–50. doi: 10.1016/j.npep.2015.07.004
29. Chen Z, Xu YY, Ge JF, Chen FH. CRHR1 Mediates the Up-Regulation of Synapsin I Induced by Nesfatin-1 Through ERK 1/2 Signaling in SH-SY5Y Cells. Cell Mol Neurobiol (2018) 38(3):627–33. doi: 10.1007/s10571-017-0509-x
30. Dong J, Xu H, Xu H, Wang PF, Cai GJ, Song HF, et al. Nesfatin-1 Stimulates Fatty-Acid Oxidation by Activating AMP-Activated Protein Kinase in STZ-Induced Type 2 Diabetic Mice. PloS One (2013) 8(12):e83397. doi: 10.1371/journal.pone.0083397
31. Dore R, Levata L, Gachkar S, Jöhren O, Mittag J, Lehnert H, et al. The Thermogenic Effect of Nesfatin-1 Requires Recruitment of the Melanocortin System. J Endocrinol (2017) 235(2):111–22. doi: 10.1530/JOE-17-0151
32. Erfani S, Moghimi A, Aboutaleb N, Khaksari M. Nesfatin-1 Improve Spatial Memory Impairment Following Transient Global Cerebral Ischemia/Reperfusion via Inhibiting Microglial and Caspase-3 Activation. J Mol Neurosci (2018) 65(3):377–84. doi: 10.1007/s12031-018-1105-3
33. Fan XT, Tian Z, Li SZ, Zhai T, Liu JL, Wang R, et al. Ghrelin Receptor Is Required for the Effect of Nesfatin-1 on Glucose Metabolism. Front Endocrinol (2018) 9:633. doi: 10.3389/fendo.2018.00633
34. Feijóo-Bandín S, Rodríguez-Penas D, García-Rúa V, Mosquera-Leal A, Otero MF, Pereira E, et al. Nesfatin-1 in Human and Murine Cardiomyocytes: Synthesis, Secretion, and Mobilization of GLUT-4. Endocrinology (2013) 154(12):4757–67. doi: 10.1210/en.2013-1497
35. Feng H, Wang Q, Guo F, Han X, Pang M, Sun X, et al. Nesfatin-1 Influences the Excitability of Gastric Distension-Responsive Neurons in the Ventromedial Hypothalamic Nucleus of Rats. Physiol Res (2017) 66(2):335–44. doi: 10.33549/physiolres.933347
36. Gao S, Guo F, Sun X, Zhang N, Gong Y, Xu L. The Inhibitory Effects of Nesfatin-1 in Ventromedial Hypothalamus on Gastric Function and Its Regulation by Nucleus Accumbens. Front Physiol (2017) 7:634. doi: 10.3389/fphys.2016.00634
37. Ge JF, Xu YY, Qin G, Pan XY, Cheng JQ, Chen FH. Nesfatin-1, a Potent Anorexic Agent, Decreases Exploration and Induces Anxiety-Like Behavior in Rats Without Altering Learning or Memory. Brain Res (2015) 1629:171–81. doi: 10.1016/j.brainres.2015.10.027
38. Guo FF, Xu L, Gao SL, Sun XR, Li ZL, Gong YL. The Effects of Nesfatin-1 in the Paraventricular Nucleus on Gastric Motility and its Potential Regulation by the Lateral Hypothalamic Area in Rats. J Neurochem (2015) 132(3):266–75. doi: 10.1111/jnc.12973
39. Heidarzadeh H, Zendehdel M, Babapour V, Gilanpour H. The Effect of Nesfatin-1 on Food Intake in Neonatal Chicks: Role of CRF1/CRF2 and H1/H3 Receptors. Vet Res Commun (2018) 42(1):39–47. doi: 10.1007/s11259-017-9706-9
40. Ishida E, Hashimoto K, Shimizu H, Okada S, Satoh T, Kato I, et al. Nesfatin-1 Induces the Phosphorylation Levels of cAMP Response Element-Binding Protein for Intracellular Signaling in a Neural Cell Line. PloS One (2012) 12):e50918. doi: 10.1371/journal.pone.0050918
41. Iwasaki Y, Nakabayashi H, Kakei M, Shimizu H, Mori M, Yada T. Nesfatin-1 Evokes Ca2+ Signaling in Isolated Vagal Afferent Neurons via Ca2+ Influx Through N-Type Channels. Biochem Biophys Res Commun (2009) 390(3):958–62. doi: 10.1016/j.bbrc.2009.10.085
42. Jia FY, Li XL, Li TN, Wu J, Xie BY, Lin L. Role of Nesfatin-1 in a Rat Model of Visceral Hypersensitivity. World J Gastroenterol (2013) 19(22):3487–93. doi: 10.3748/wjg.v19.i22.3487
43. Jiang L, Xu K, Li J, Zhou X, Xu L, Wu Z, et al. Nesfatin-1 Suppresses Interleukin-1β-Induced Inflammation, Apoptosis, and Cartilage Matrix Destruction in Chondrocytes and Ameliorates Osteoarthritis in Rats. Aging (2020) 12(2):1760–77. doi: 10.18632/aging.102711
44. Kan JY, Yen MC, Wang JY, Wu DC, Chiu YJ, Ho YW, et al. Nesfatin-1/Nucleobindin-2 Enhances Cell Migration, Invasion, and Epithelial-Mesenchymal Transition via LKB1/AMPK/TORC1/ZEB1 Pathways in Colon Cancer. Oncotarget (2016) 7(21):31336–49. doi: 10.18632/oncotarget.9140
45. Kerbel B, Unniappan S. Nesfatin-1 Suppresses Energy Intake, Co-Localises Ghrelin in the Brain and Gut, and Alters Ghrelin, Cholecystokinin and Orexin mRNA Expression in Goldfish. J Neuroendocrinol (2012) 24(2):366–77. doi: 10.1111/j.1365-2826.2011.02246.x
46. Levata L, Dore R, Jöhren O, Schwaninger M, Schulz C, Lehnert H. Nesfatin-1 Acts Centrally to Induce Sympathetic Activation of Brown Adipose Tissue and Non-Shivering Thermogenesis. Horm Metab Res (2019) 51(10):678–85. doi: 10.1055/a-0985-4272
47. Li C, Zhang F, Shi L, Zhang H, Tian Z, Xie J, et al. Nesfatin-1 Decreases Excitability of Dopaminergic Neurons in the Substantia Nigra. J Mol Neurosci (2014) 52(3):419–24. doi: 10.1007/s12031-013-0169-3
48. Li T, Wei S, Fan C, Tang D, Luo D. Nesfatin-1 Promotes Proliferation, Migration and Invasion of HTR-8/SVneo Trophoblast Cells and Inhibits Oxidative Stress via Activation of PI3K/AKT/mTOR and AKT/Gsk3β Pathway. Reprod Sci (2021) 28(2):550–61. doi: 10.1007/s43032-020-00324-1
49. Li ZL, Xu L, Sun XR, Guo FF, Gong YL, Gao SL. Central Nesfatin-1 Influences the Excitability of Ghrelin-Responsive Gastric Distension Neurons in the Arcuate Nucleus and Reduces Gastric Motility in Rats. Eur J Neurosci (2013) 38(11):3636–43. doi: 10.1111/ejn.12366
50. Lu QB, Wang HP, Tang ZH, Cheng H, Du Q, Wang YB, et al. Nesfatin-1 Functions as a Switch for Phenotype Transformation and Proliferation of VSMCs in Hypertensive Vascular Remodeling. Biochim Biophys Acta Mol Basis Dis (2018) 1864(6 Pt A):2154–68. doi: 10.1016/j.bbadis.2018.04.002
51. Maejima Y, Horita S, Kobayashi D, Aoki M, O’hashi R, Imai R, et al. Nesfatin-1 Inhibits Voltage Gated K+ Channels in Pancreatic Beta Cells. Peptides (2017) 95:10–5. doi: 10.1016/j.peptides.2017.07.001
52. Maejima Y, Sedbazar U, Suyama S, Kohno D, Onaka T, Takano E, et al. Nesfatin-1-Regulated Oxytocinergic Signaling in the Paraventricular Nucleus Causes Anorexia Through a Leptin-Independent Melanocortin Pathway. Cell Metab (2009) 10(5):355–65. doi: 10.1016/j.cmet.2009.09.002
53. Mazza R, Gattuso A, Filice M, Cantafio P, Cerra MC, Angelone T, et al. Nesfatin-1 as a New Positive Inotrope in the Goldfish (Carassius Auratus) Heart. Gen Comp Endocrinol (2015) 224:160–7. doi: 10.1016/j.ygcen.2015.08.003
54. Mori Y, Shimizu H, Kushima H, Saito T, Hiromura M, Terasaki M, et al. Nesfatin-1 Suppresses Peripheral Arterial Remodeling Without Elevating Blood Pressure in Mice. Endocr Connect (2019) 8(5):536–46. doi: 10.1530/EC-19-0120
55. Nair N, Gerger C, Hatef A, Weber LP, Unniappan S. Ultrasonography Reveals In Vivo Dose-Dependent Inhibition of End Systolic and Diastolic Volumes, Heart Rate and Cardiac Output by Nesfatin-1 in Zebrafish. Gen Comp Endocrinol (2016) 234:142–50. doi: 10.1016/j.ygcen.2016.02.015
56. Nakata M, Manaka K, Yamamoto S, Mori M, Yada T. Nesfatin-1 Enhances Glucose-Induced Insulin Secretion by Promoting Ca(2+) Influx Through L-Type Channels in Mouse Islet β-Cells. Endocr J (2011) 58(4):305–13. doi: 10.1507/endocrj.k11e-056
57. Osaki A, Shimizu H. Peripheral Administration of Nesfatin-1 Increases Blood Pressure in Mice. Hypertens Res (2014) 37(2):185–6. doi: 10.1038/hr.2013.122
58. Ozcan M, Gok ZB, Kacar E, Serhatlioglu I, Kelestimur H. Nesfatin-1 Increases Intracellular Calcium Concentration by Protein Kinase C Activation in Cultured Rat Dorsal Root Ganglion Neurons. Neurosci Lett (2016) 619:177–81. doi: 10.1016/j.neulet.2016.03.018
59. Ozturk CC, Oktay S, Yuksel M, Akakin D, Yarat A, Kasimay Cakir O. Anti-Inflammatory Effects of Nesfatin-1 in Rats With Acetic Acid - Induced Colitis and Underlying Mechanisms. J Physiol Pharmacol (2015) 66(5):741–50.
60. Price CJ, Hoyda TD, Samson WK, Ferguson AV. Nesfatin-1 Influences the Excitability of Paraventricular Nucleus Neurones. J Neuroendocrinol (2008) 20(2):245–50. doi: 10.1111/j.1365-2826.2007.01641.x
61. Price CJ, Samson WK, Ferguson AV. Nesfatin-1 Inhibits NPY Neurons in the Arcuate Nucleus. Brain Res (2008) 1230:99–106. doi: 10.1016/j.brainres.2008.06.084
62. Prinz P, Goebel-Stengel M, Teuffel P, Rose M, Klapp BF, Stengel A. Peripheral and Central Localization of the Nesfatin-1 Receptor Using Autoradiography in Rats. Biochem Biophys Res Commun (2016) 470(3):521–7. doi: 10.1016/j.bbrc.2016.01.113
63. Ramanjaneya M, Tan BK, Rucinski M, Kawan M, Hu J, Kaur J, et al. Nesfatin-1 Inhibits Proliferation and Enhances Apoptosis of Human Adrenocortical H295R Cells. J Endocrinol (2015) 226(1):1–11. doi: 10.1530/JOE-14-0496
64. Ranjan A, Choubey M, Yada T, Krishna A. Direct Effects of Neuropeptide Nesfatin-1 on Testicular Spermatogenesis and Steroidogenesis of the Adult Mice. Gen Comp Endocrinol (2019) 271:49–60. doi: 10.1016/j.ygcen.2018.10.022
65. Ranjan A, Choubey M, Yada T, Krishna A. Immunohistochemical Localization and Possible Functions of Nesfatin-1 in the Testis of Mice During Pubertal Development and Sexual Maturation. J Mol Histol (2019) 50(6):533–49. doi: 10.1007/s10735-019-09846-8
66. Ranjan A, Choubey M, Yada T, Krishna A. Nesfatin-1 Ameliorates Type-2 Diabetes-Associated Reproductive Dysfunction in Male Mice. J Endocrinol Invest (2020) 43(4):515–28. doi: 10.1007/s40618-019-01136-0
67. Shen XL, Song N, Du XX, Li Y, Xie JX, Jiang H. Nesfatin-1 Protects Dopaminergic Neurons Against MPP+/MPTP-Induced Neurotoxicity Through the C-Raf-ERK1/2-Dependent Anti-Apoptotic Pathway. Sci Rep (2017) 7:40961. doi: 10.1038/srep40961
68. Tan J, Jin L, Wang Y, Cao B, Wang S, Zhang F, et al. Nesfatin-1 Acts as an Inhibitory Factor in Human Gastrointestinal Smooth Muscle Cells in Diabetes Mellitus-Induced Delayed Gastric Emptying. Int J Clin Exp Pathol (2016) 9(11):11214–21.
69. Tanida M, Gotoh H, Yamamoto N, Wang M, Kuda Y, Kurata Y, et al. Hypothalamic Nesfatin-1 Stimulates Sympathetic Nerve Activity via Hypothalamic ERK Signaling. Diabetes (2015) 64(11):3725–36. doi: 10.2337/db15-0282
70. Tanida M, Mori M. Nesfatin-1 Stimulates Renal Sympathetic Nerve Activity in Rats. Neuroreport (2011) 22(6):309–12. doi: 10.1097/WNR.0b013e328346107f
71. Tasatargil A, Kuscu N, Dalaklioglu S, Adiguzel D, Celik-Ozenci C, Ozdem S, et al. Cardioprotective Effect of Nesfatin-1 Against Isoproterenol-Induced Myocardial Infarction in Rats: Role of the Akt/GSK-3β Pathway. Peptides (2017) 95:1–9. doi: 10.1016/j.peptides.2017.07.003
72. Vélez EJ, Unniappan S. Nesfatin-1 and Nesfatin-1-Like Peptide Suppress Growth Hormone Synthesis via the AC/PKA/CREB Pathway in Mammalian Somatotrophs. Sci Rep (2020) 10(1):16686. doi: 10.1038/s41598-020-73840-4
73. Wang Q, Guo F, Sun X, Gao S, Li Z, Gong Y, et al. Effects of Exogenous Nesfatin-1 on Gastric Distention-Sensitive Neurons in the Central Nucleus of the Amygdala and Gastric Motility in Rats. Neurosci Lett (2014) 582:65–70. doi: 10.1016/j.neulet.2014.09.003
74. Wu D, Yang M, Chen Y, Jia Y, Ma ZA, Boden G, et al. Hypothalamic Nesfatin-1/NUCB2 Knockdown Augments Hepatic Gluconeogenesis That Is Correlated With Inhibition of mTOR-STAT3 Signaling Pathway in Rats. Diabetes (2014) 63(4):1234–47. doi: 10.2337/db13-0899
75. Xia ZF, Fritze DM, Li JY, Chai B, Zhang C, Zhang W, et al. Nesfatin-1 Inhibits Gastric Acid Secretion via a Central Vagal Mechanism in Rats. Am J Physiol Gastrointest Liver Physiol (2012) 303(5):G570–7. doi: 10.1152/ajpgi.00178.2012
76. Xu L, Wang H, Gong Y, Pang M, Sun X, Guo F, et al. Nesfatin-1 Regulates the Lateral Hypothalamic Area Melanin-Concentrating Hormone-Responsive Gastric Distension-Sensitive Neurons and Gastric Function via Arcuate Nucleus Innervation. Metabolism (2017) 67:14–25. doi: 10.1016/j.metabol.2016.10.010
77. Xu L, Wang Q, Guo F, Pang M, Sun X, Gao S, et al. Nesfatin-1 Signaling in the Basom Edial Amygdala Modulates the Gastric Distension-Sensitive Neurons Discharge and Decreases Gastric Motility via Melanocortin 3/4 Receptors and Modified by the Arcuate Nucleus. Eur J Pharmacol (2015) 764:164–72. doi: 10.1016/j.ejphar.2015.07.002
78. Yang M, Zhang Z, Wang C, Li K, Li S, Boden G, et al. Nesfatin-1 Action in the Brain Increases Insulin Sensitivity Through Akt/AMPK/TORC2 Pathway in Diet-Induced Insulin Resistance. Diabetes (2012) 61(8):1959–68. doi: 10.2337/db11-1755
79. Yin Y, Li Z, Gao L, Li Y, Zhao J, Zhang W. AMPK-Dependent Modulation of Hepatic Lipid Metabolism by Nesfatin-1. Mol Cell Endocrinol (2015) 417:20–6. doi: 10.1016/j.mce.2015.09.006
80. Ying J, Zhang Y, Gong S, Chang Z, Zhou X, Li H, et al. Nesfatin-1 Suppresses Cardiac L-Type Ca²⁺ Channels Through Melanocortin Type 4 Receptor and the Novel Protein Kinase C Theta Isoform Pathway. Cell Physiol Biochem (2015) 36(2):555–68. doi: 10.1159/000430120
81. Yosten GL, Samson WK. Neural Circuitry Underlying the Central Hypertensive Action of Nesfatin-1: Melanocortins, Corticotropin-Releasing Hormone, and Oxytocin. Am J Physiol Regul Integr Comp Physiol (2014) 306(10):R722–7. doi: 10.1152/ajpregu.00396.2013
82. Yuan JH, Chen X, Dong J, Zhang D, Song K, Zhang Y, et al. Nesfatin-1 in the Lateral Parabrachial Nucleus Inhibits Food Intake, Modulates Excitability of Glucosensing Neurons, and Enhances UCP1 Expression in Brown Adipose Tissue. Front Physiol (2017) 8:235. doi: 10.3389/fphys.2017.00235
83. Zhang JR, Lu QB, Feng WB, Wang HP, Tang ZH, Cheng H, et al. Nesfatin-1 Promotes VSMC Migration and Neointimal Hyperplasia by Upregulating Matrix Metalloproteinases and Downregulating Pparγ. BioMed Pharmacother (2018) 102:711–7. doi: 10.1016/j.biopha.2018.03.120
84. Zhang T, Wang M, Liu L, He B, Hu J, Wang Y. Hypothalamic Nesfatin-1 Mediates Feeding Behavior via MC3/4R-ERK Signaling Pathway After Weight Loss in Obese Sprague-Dawley Rats. Peptides (2019) 119:170080. doi: 10.1016/j.peptides.2019.04.007
85. Zhang X, Qi J, Tang N, Wang S, Wu Y, Chen H, et al. Intraperitoneal Injection of Nesfatin-1 Primarily Through the CCK-CCK1R Signal Pathway Affects Expression of Appetite Factors to Inhibit the Food Intake of Siberian Sturgeon (Acipenser Baerii). Peptides (2018) 109:14–22. doi: 10.1016/j.peptides.2018.09.008
86. Zhou XP, Sha J, Huang L, Li TN, Zhang RR, Tang MD, et al. Nesfatin-1/NUCB2 in the Amygdala Influences Visceral Sensitivity via Glucocorticoid and Mineralocorticoid Receptors in Male Maternal Separation Rats. Neurogastroenterol Motil (2016) 28(10):1545–53. doi: 10.1111/nmo.12853
87. Pan W, Hsuchou H, Kastin AJ. Nesfatin-1 Crosses the Blood-Brain Barrier Without Saturation. Peptides (2007) 28(11):2223–8. doi: 10.1016/j.peptides.2007.09.005
88. Goebel-Stengel M, Wang L, Stengel A, Taché Y. Localization of Nesfatin-1 Neurons in the Mouse Brain and Functional Implication. Brain Res (2011) 1396:20–34. doi: 10.1016/j.brainres.2011.04.031
89. Foo KS, Brismar H, Broberger C. Distribution and Neuropeptide Coexistence of Nucleobindin-2 mRNA/Nesfatin-Like Immunoreactivity in the Rat CNS. Neuroscience (2008) 156(3):563–79. doi: 10.1016/j.neuroscience.2008.07.054
90. Kato M, Ma HT, Tatemoto K. GLP-1 Depolarizes the Rat Pancreatic Beta Cell in a Na(+)-Dependent Manner. Regul Pept (1996) 62(1):23–7. doi: 10.1016/0167-0115(95)00164-6
91. Hiriart M, Matteson DR. Na Channels and Two Types of Ca Channels in Rat Pancreatic B Cells Identified With the Reverse Hemolytic Plaque Assay. J Gen Physiol (1988) 91(5):617–39. doi: 10.1085/jgp.91.5.617
92. Arkhammar P, Juntti-Berggren L, Larsson O, Welsh M, Nånberg E, Sjöholm A, et al. Protein Kinase C Modulates the Insulin Secretory Process by Maintaining a Proper Function of the β-Cell Voltage- Activated Ca2+ Channels. J Biol Chem (1994) 269:2743–9. doi: 10.1016/S0021-9258(17)42006-0
93. Iftinca MC. Neuronal T-Type Calcium Channels: What’s New? Iftinca: T-Type Channel Regulation. J Med Life (2011) 4(2):126–38.
94. Ross WN. Understanding Calcium Waves and Sparks in Central Neurons. Nat Rev Neurosci (2012) 13(3):157–68. doi: 10.1038/nrn3168
Keywords: gut-brain axis, food intake, NUCB2, nucleobindin-2, stress, nesfatin-1
Citation: Rupp SK, Wölk E and Stengel A (2021) Nesfatin-1 Receptor: Distribution, Signaling and Increasing Evidence for a G Protein-Coupled Receptor – A Systematic Review. Front. Endocrinol. 12:740174. doi: 10.3389/fendo.2021.740174
Received: 12 July 2021; Accepted: 26 August 2021;
Published: 10 September 2021.
Edited by:
Liliane Schoofs, KU Leuven, BelgiumReviewed by:
Tommaso Angelone, University of Calabria, ItalyHendrik Lehnert, University of Salzburg, Austria
Copyright © 2021 Rupp, Wölk and Stengel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andreas Stengel, YW5kcmVhcy5zdGVuZ2VsQG1lZC51bmktdHVlYmluZ2VuLmRl