
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Endocrinol. , 22 June 2021
Sec. Obesity
Volume 12 - 2021 | https://doi.org/10.3389/fendo.2021.692677
This article is part of the Research Topic Next-Generation Sequencing in Aging-Related Diseases View all 4 articles
 Katherine M. Livingstone1*
Katherine M. Livingstone1* Mun Hua Tan2
Mun Hua Tan2 Gavin Abbott1
Gavin Abbott1 Rachel L. Duckham1,3
Rachel L. Duckham1,3 Larry Croft4
Larry Croft4 Joey Ward5
Joey Ward5 Mark McEvoy6
Mark McEvoy6 Michelle A. Keske1
Michelle A. Keske1 Christopher Austin4
Christopher Austin4 Steven J. Bowe7
Steven J. Bowe7Body composition (fat, skeletal muscle and bone mass) is an important determinant of overall health and risk of endocrine disorders such as type 2 diabetes and osteoporosis. Although diet and physical activity are strongly implicated, body composition is also heritable. We conducted a discovery genome-wide association study on 31 phenotypes from the three-compartment body composition model (fat, lean and bone mass) in a set of 4 386 individuals (n = 2 109 males, n = 2 294 females) from the UK Biobank pilot imaging enhancement program that underwent a dual energy X-ray absorptiometry (DXA) scan for assessment of body composition and genetic screening. From 6 137 607 imputed single nucleotide polymorphisms (SNPs) we identified 17 body composition loci (P<5.0 x 10-8). GWAS from the combined dataset identified four statistically significant SNPs (rs7592270, rs145972737, rs13212044, rs77772562). In sex-stratified GWAS, 10 male specific SNPs across all traits were identified and five female specific SNPs. Of the 17 SNPs, six were in or close to a gene where there was a plausible functional connection. Three SNPs (rs7592270, rs77772562 and rs7552312) were correlated with obesity phenotypes, one SNP (rs2236705) with lean phenotypes and two with bone mass phenotypes (rs112098641 and rs113380185). These results highlight candidate genes and biological pathways related to body composition, including glucose metabolism and estrogen regulation, that are of interest to replicate in future studies.
Body composition is implicated in the progression of many chronic diseases, including endocrine disorders such as type 2 diabetes (T2D) (1). The heritability of body composition is high, with total and regional distribution of fat, lean and bone mass governed by genetic susceptibility (2). The increasing availability of genetic data from large population-based cohorts provides an opportunity for Genome-Wide-Association Studies (GWAS) to identify causal Single Nucleotide Polymorphisms (SNPs) with multiple body composition phenotypes.
Most body composition GWAS research has focused on the genetics of body mass index (BMI) (2), waist and hip circumference (3) or singular compartments of body composition, such as lean or fat mass (4, 5). While previous GWAS have investigated the role of bone content of specific regions (6), few have investigated the genetics of total and regional bone mass. These studies have advanced understanding of the role and potential mechanisms of common genetic variations in BMI and body fat distribution, including sexual dimorphisms in the genetic regulation of these traits. Specifically, pathway analyses suggest that adipose tissue deposition and BMI are closely linked with insulin regulation and lipid biology, and thus share pathways with T2D and glycaemic traits (2, 3). These phenotypes have also been linked with skeletal growth processes (3), yet the common genetic variations in fat, lean and bone phenotypes remain unclear as they have not been investigation simultaneously. To our knowledge, no GWAS have investigated the genetics of all three body composition compartments simultaneously. Understanding the genetics of the three-compartment model of body composition will provide new insights into the shared heritability of and inter-connection between fat, lean and bone traits. Moreover, the majority of research has derived estimates of fat and lean mass from bio-electrical impedance analysis, which has inflexible hydration assumptions and is unable to assess bone mass (7). While dual energy X-ray absorptiometry (DXA) is the gold standard for assessing the three-compartment model of body composition, the high cost and exposure to radiation has largely precluded application in a large population cohort (8).
The availability of gold standard DXA data on 5 170 participants from the UK Biobank (UKB) pilot imaging enhancement program provided an opportunity to perform a discovery GWAS on the genetic determinants of fat, lean and bone body composition. Sex-stratified analyses were performed to identify SNPs that differed between males and females.
UKB is a population cohort of 500 000 individuals aged 40 to 69 years living in the United Kingdom (9). Individuals identified from National Health Service patient registers were invited to one of 22 assessment centres between 2006 and 2020 to provide deep phenotyping and molecular data. UKB received approval from the National Information Governance Board for Health and Social Care and the National Health Service North West Centre for Research Ethics Committee (Ref: 11/NW/0382). All participants provided informed consent to participate. UKB imaging enhancement program will make DXA data available for 100 000 individuals by 2023 (10). Between 2014 to 2015, UKB undertook a pilot study in 5 170 participants. For the present study, complete case analysis was used. Participants were excluded if they had missing call rate of > 0.1, unusual level of heterozygosity, sex mismatch, sex chromosome aneuploidy, had missing data on sex, or were not of ‘White British’ descent.
Total and regional fat, lean and bone mass were determined from a total body GE-Lunar iDXA scan (GE Healthcare, Madison, Wisconsin, USA) and analyzed with Encore software (version 11.0) using standardized procedures. All scans were collected and analyzed by a trained radiographer. For the purpose of this study, 31 fat, lean and bone phenotypes were included (Table 1).
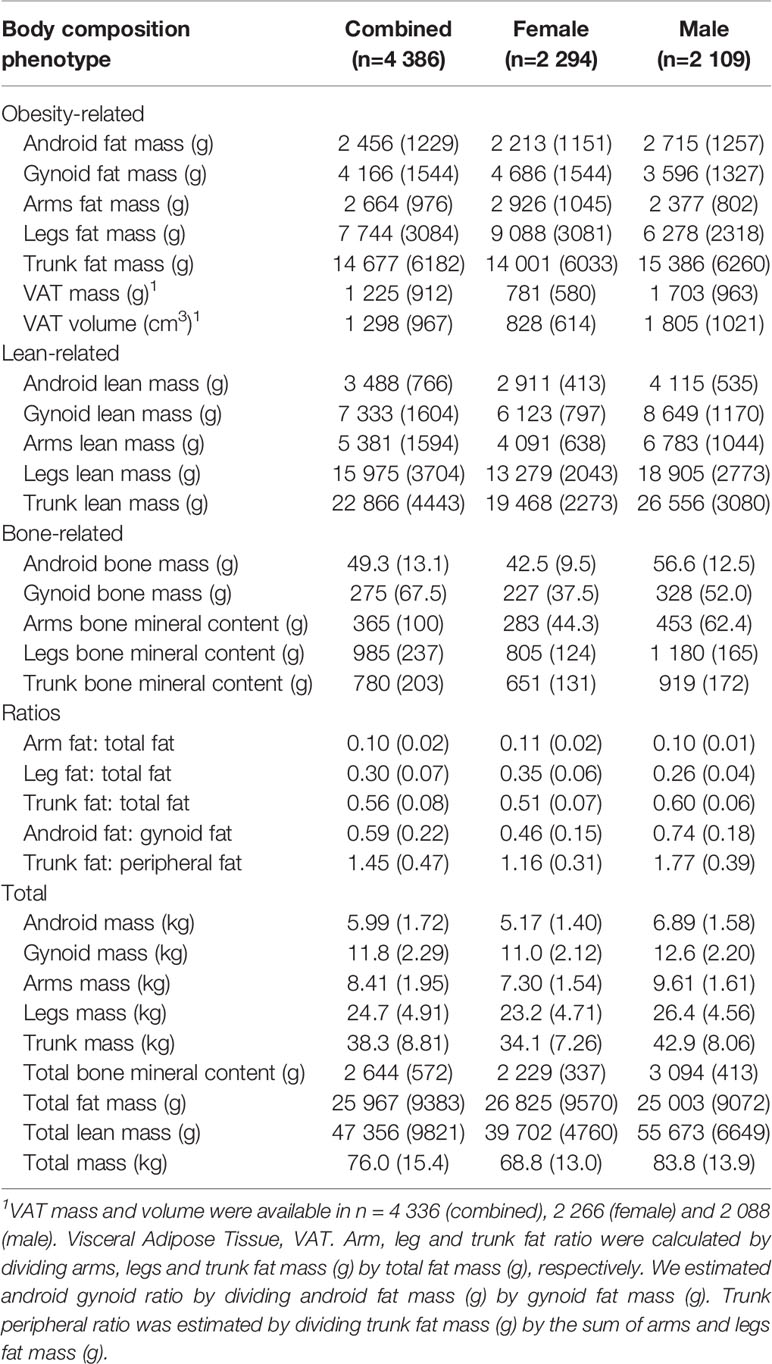
Table 1 Mean (SD) of body composition phenotypes in combined, female and male datasets.
The March 2018 release of the imputed genetic data (analyzed from blood samples) from UK Biobank (downloaded 11 November 2019) was used (11). Genome-wide association analysis for each of the 31 phenotypes was performed on imputed SNPs with PLINK v2.00a3LM, using the generalized linear model suitable for quantitative phenotypes. In combined analysis, genotyping platform, genetic sex and the first four principal genetic components were set as covariates. Resulting polymorphisms with missing call rates of greater than 0.05, Hardy-Weinberg equilibrium p-values of less than 1 x 10-6, allele frequencies lower than 0.01 and/or imputation scores of less than 0.8 were excluded from the final set variants. The ‘estlambda’ function in GenABEL library was used to estimate genomic inflation factor of each GWAS analysis. SNP associations with p-values < 5.0 x 10-8 were considered statistically significant. Bonferroni correction was used to correct for multiple testing. Related individuals were identified based on kinship coefficient values and, for any pair of individuals with valid phenotypic values, one was excluded at random. The LD Score Regression was used to estimate heritability and correlations of phenotypes. Correlations were deemed strong at r>0.6 and significant at P>0.05. We cross-referenced lead SNPs from all independent signals in our analyses with the NHGRI-EBI catalog of published GWAS to determine whether fat, lean and bone-associated signals overlapped with previously identified associations from previous GWAS (GWAS Catalog—downloaded 9 July 2020). For SNPs identified at p-values < 5.0 x 10-8, SNPs were inspected to ascertain proximity to a coding gene and thus potential plausible functional connections. Only SNPs in or very close to a gene where there was a plausible functional connection were further discussed.
A total of 4 386 participants (n=2 109 male, n=2 294 female) were included (Figure S1). Mean age at recruitment was 55.9 (SD 7.5). Mean (SD) of obesity-, lean-, and bone-related body composition phenotypes, as well as body composition ratios, are presented in Table 1. The genomic inflation factors for the combined and sex-stratified GWAS are presented in Table S1. For the combined dataset, genomic inflation factors ranged between 1.010 (SE 2.12 X 10-6) to 1.046 (SE 4.52 X 10-6). Heritability estimates for body composition phenotypes in the combined dataset are shown in Table S2 and ranged between 0.08 (SE 0.12) to 0.51 (SE 0.14). Correlations for body composition phenotypes are shown in Table S3. With the exception of leg fat ratio, android gynoid ratio, trunk bone mineral content, trunk peripheral ratio and Visceral Adipose Tissue (VAT) mass and volume, the majority of the 31 body composition phenotypes were strongly and significantly positively correlated with most other phenotypes.
As shown in Table 2, from 6 137 607 imputed SNPs, we identified 17 body composition loci (P<5.0 x 10-8). GWAS from the combined dataset identified four statistically significant SNPs (rs7592270, rs145972737, rs13212044, rs77772562; Figure S2). In sex-stratified GWAS, 10 male specific SNPs across all traits were identified and five female specific SNPs (Table 1). Of these, six SNPs were in or close to a gene where there was a plausible functional connection (rs7592270, rs77772562, rs7552312, rs2236705, rs112098641 and rs113380185; Figure 1, Table 1, Figures S3 and S4). Three SNPs were associated with obesity phenotypes in the combined dataset. SNP rs7592270 was correlated with female android fat mass, android total mass, trunk total mass, VAT mass and VAT volume, SNP rs7552312 was correlated with trunk total fat ratio and SNP rs77772562 was correlated with the trunk peripheral ratio. One SNP rs2236705 was correlated with lean phenotypes and two SNPs (rs112098641 and rs113380185) were correlated with bone phenotypes. In sex-stratified datasets, SNP rs2236705 was correlated with female lean leg mass, SNP rs112098641 was associated with female gynoid bone mass and SNP rs113380185 was correlated with male android bone mass (Table 1, Figures S3 and S4).
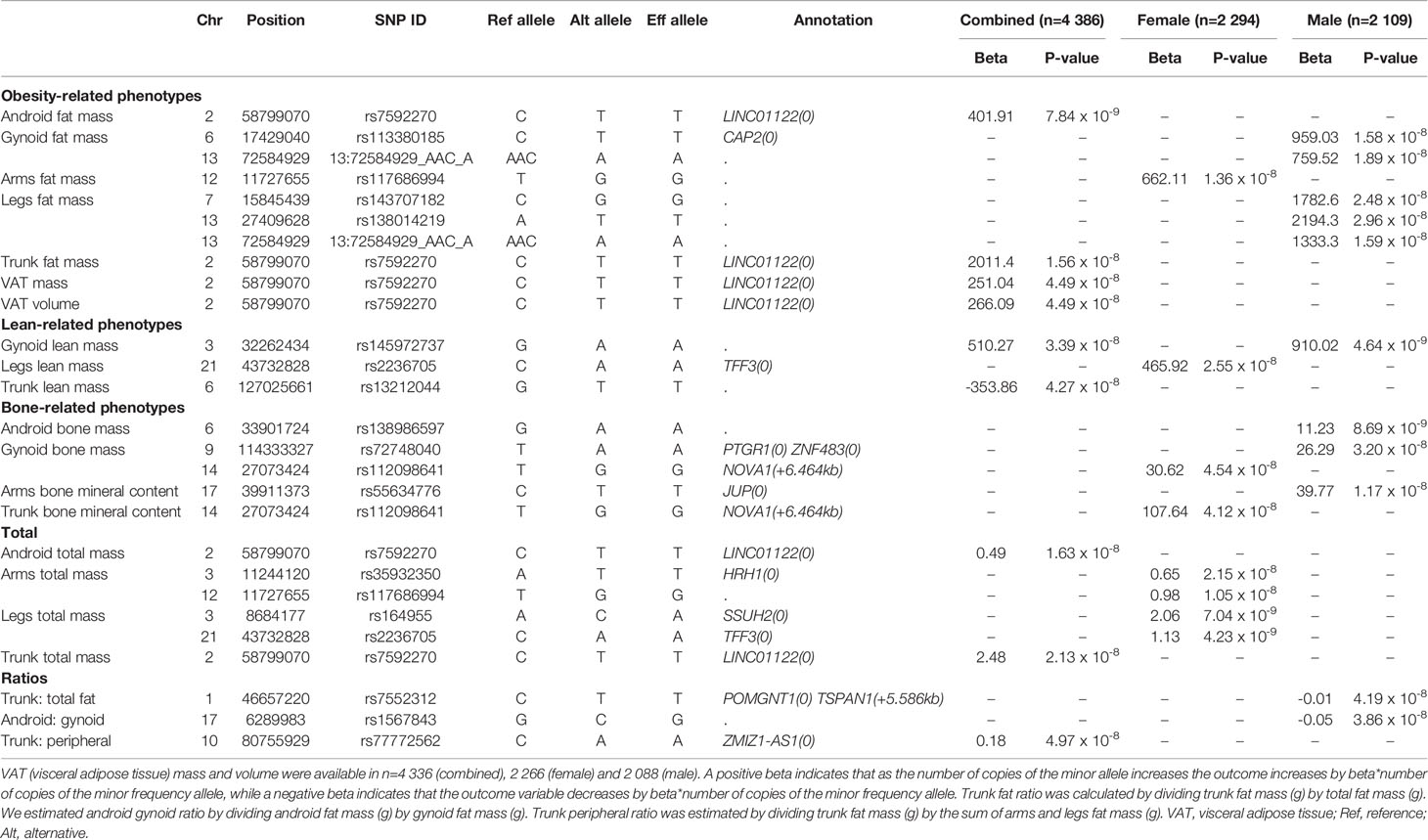
Table 2 List of SNPs in combined, female and male datasets significant at p< 5.0 x 10-8.
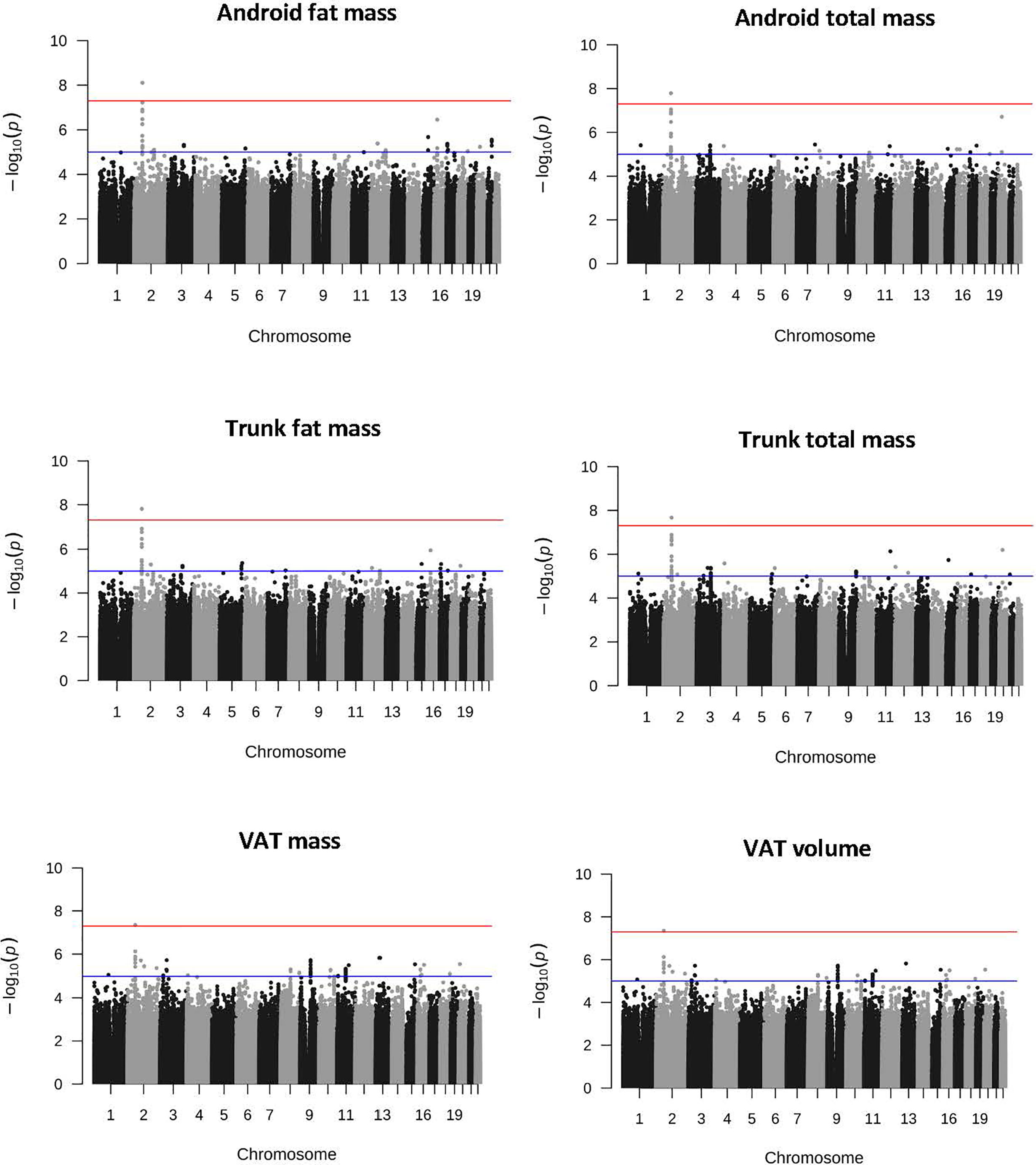
Figure 1 Manhattan plot of obesity-related SNP rs7592270 statistically significant above line at p-value 5.0 x 10-8 in the combined (male and female) GWAS (n = 4 386; VAT, n = 4 336) VAT, visceral adipose tissue.
The aim of this discovery GWAS was to provide preliminary evidence for the role of genetics in the three-compartment model of body composition. From the 6 137 607 imputed SNPs and 31 body composition phenotypes investigated, our main findings were that 17 SNPs were significantly associated with either fat, lean or bone phenotypes. Of these, six SNPs were linked to genes with known functional outcomes and thus help to explain the physiological mechanisms leading to body composition. These preliminary findings support the need for clinicians to consider the inter-connection between fat, lean and bone mass and provide new insights into biological pathways, including glucose metabolism and estrogen regulation, that will inform future research aimed at understanding the complex biology of body composition.
We identified three SNPs related to obesity that have been previously reported to be associated with risk of T2D (12, 13). Thus, these SNPs may exert their effects at increasing T2D risk by modifying body composition (specifically fat) and by altering glucose and insulin pathways (3). More specifically, SNP rs7592270 was correlated with female android fat mass, android total mass, trunk total mass, VAT mass and volume, all of which are indicators of harmful accumulation of central regional body fat often associated with increased risk of T2D (12). SNP rs7592270 is found in the noncoding gene LINC01122, which is correlated with anthropometric extremes and is near SNPs correlated to obesity class I (rs929641) and being overweight (rs887912) (14). SNPs rs7592270, rs929641 and rs887912 were in or very close to LINC01122. This SNP is found in the antisense noncoding gene ZMIZ1-AS1, sharing a bidirectional promoter with ZMIZ1. ZMIZ1, a transcription factor regulator, is involved in glucose regulation and diabetes (15). SNP rs7552312 was correlated with male trunk fat mass and is found in the POMGNT1 gene. Mutations in this gene are also associated with T2D (13), which provides another link between obesity and risk of T2D. T2D is often characterised by higher fat and lower skeletal muscle mass, increasing risk of inadequate glucose metabolism, while excess fat mass distribution within the central region may increase fat infiltration within the muscle and risk of insulin resistance (16), further supporting that these obesogenic SNPs are also linked to T2D risk.
We observed that three SNPs were related to lean and bone mass phenotypes and are likely to impact body composition via estrogen pathways. SNP rs2236705 was correlated with female lean leg mass. This SNP is found in the intron of the TFF3 gene, which is involved in skeletal metabolism, is estrogen regulated and regulates glucose metabolism (17). SNP rs112098641 is associated with female gynoid bone mass. The SNP is 6kb upstream from the NOVA1 gene in the intron of a lncRNA gene, which is estrogenically regulated (18). Moreover, estrogen plays a role in the development of lean tissue mass and bone mass regulation in adulthood (19). Estrogen acts through two receptors – the alpha and beta receptors – which are important in lean and bone metabolism, respectively. Through these receptors, estrogen protects bone metabolism by regulating the survival of osteoclasts, if circulating levels of estrogen are low, bone mass will be lost, such as that observed in post-menopausal women (19). Similarly, the estrogen-receptor alpha located on skeletal muscle may activate signalling pathways, such as Insulin-like Growth Factor-1, that mediates skeletal metabolism (20). SNP rs113380185, found in the CAP2 gene, was correlated with male android bone mass and has been associated with human height (21) (Table S1, Figures S3 and S4).
We acknowledge some limitations. Our findings are not generalizable to non-white populations. Our sample size is relatively small and we are likely underpowered to detect variants with small effect sizes. Nonetheless, these findings are from a pilot study and are designed to be replicated and to inform future research in the full 100 000 participants from the UKB imaging enhancement programme. Strengths of this study include the investigation of 31 phenotypes from the three-compartment model of body composition. These have been largely under-researched due to the high cost and participant burden of collecting DXA measures.
The present study identified six loci that help explain the physiological mechanisms leading to body composition. These preliminary findings support the connection between fat, lean and bone mass and the need for these compartments of body composition to be considered concurrently by clinicians. The biological pathways highlighted by this study, including glucose metabolism and estrogen regulation, will inform future research aimed at understanding the complex biology of body composition. Potential candidate genes, such as LINC01122 and POMGNT1, are of interest to investigate in future studies, including the full UKB imaging enhancement programme dataset when it becomes available.
The data analyzed in this study is subject to the following licenses/restrictions: The UK Biobank data are available on application to the UK Biobank (www.ukbiobank.ac.uk/). Requests to access these datasets should be directed to www.ukbiobank.ac.uk/.
UK Biobank received ethical approval from the National Information Governance Board for Health and Social Care and the National Health Service North West Centre for Research Ethics Committee (reference 13/NW/0382). All participants provided informed consent to participate. The present analyses were conducted under UK Biobank application number 34894. The patients/participants provided their written informed consent to participate in this study.
KL, JW, RD, MM, and SB designed the analysis. MT, GA, SB, JW, and LC conducted the statistical analysis. KL, MT, MK, and SB drafted the manuscript. All authors contributed to the article and approved the submitted version.
KL is supported by a National Health and Medical Research Council Emerging Leadership Investigator Grant (APP1173803). JW is funded by the Lister Prize Fellowship (173096). The funding source had no role in the design or conduct of the study; collection, management, analysis, and interpretation of the data; or preparation, review, or approval of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors acknowledge Dr Christopher McAvaney for contribution to data management.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2021.692677/full#supplementary-material
1. Al-Goblan AS, Al-Alfi MA, Khan MZ. Mechanism Linking Diabetes Mellitus and Obesity. Diabetes Metab syndrome Obes (2014) 7:587–91. doi: 10.2147/DMSO.S67400
2. Locke AE, Kahali B, Berndt SI, Justice AE, Pers TH, Day FR, et al. Genetic Studies of Body Mass Index Yield New Insights for Obesity Biology. Nature (2015) 518(7538):197–206. doi: 10.1038/nature14177
3. Shungin D, Winkler TW, Croteau-Chonka DC, Ferreira T, Locke AE, Mägi R, et al. New Genetic Loci Link Adipose and Insulin Biology to Body Fat Distribution. Nature (2015) 518(7538):187–96. doi: 10.1038/nature14132
4. Rask-Andersen M, Karlsson T, Ek WE, Johansson Å. Genome-Wide Association Study of Body Fat Distribution Identifies Adiposity Loci and Sex-Specific Genetic Effects. Nat Commun (2019) 10(1):339. doi: 10.1038/s41467-018-08000-4
5. Lu Y, Day FR, Gustafsson S, Buchkovich ML, Na J, Bataille V, et al. New Loci for Body Fat Percentage Reveal Link Between Adiposity and Cardiometabolic Disease Risk. Nat Commun (2016) 7:10495. doi: 10.1038/ncomms10495
6. Estrada K, Styrkarsdottir U, Evangelou E, Hsu Y-H, Duncan EL, Ntzani EE, et al. Genome-Wide Meta-Analysis Identifies 56 Bone Mineral Density Loci and Reveals 14 Loci Associated With Risk of Fracture. Nat Genet (2012) 44(5):491–501. doi: 10.1038/ng.2249
7. Davydov DM, Boev A, Gorbunov S. Making the Choice Between Bioelectrical Impedance Measures for Body Hydration Status Assessment. Sci Rep (2021) 11(1):7685. doi: 10.1038/s41598-021-87253-4
8. Comuzzie AG, Cole SA, Laston SL, Voruganti VS, Haack K, Gibbs RA, et al. Novel Genetic Loci Identified for the Pathophysiology of Childhood Obesity in the Hispanic Population. PloS One (2012) 7(12):e51954. doi: 10.1371/journal.pone.0051954
9. Sudlow C, Gallacher J, Allen N, Beral V, Burton P, Danesh J, et al. UK Biobank: An Open Access Resource for Identifying the Causes of a Wide Range of Complex Diseases of Middle and Old Age. PloS Med (2015) 12(3):e1001779. doi: 10.1371/journal.pmed.1001779
10. Littlejohns TJ, Holliday J, Gibson LM, Garratt S, Oesingmann N, Alfaro-Almagro F, et al. The UK Biobank Imaging Enhancement of 100,000 Participants: Rationale, Data Collection, Management and Future Directions. Nat Commun (2020) 11(1):2624. doi: 10.1038/s41467-020-15948-9
11. Bycroft C, Freeman C, Petkova D, Band G, Elliott LT, Sharp K, et al. Genome-Wide Genetic Data on ~500,000 UK Biobank Participants. bioRxiv (2017) 166298. doi: 10.1101/166298
12. Lee BJ, Ku B. A Comparison of Trunk Circumference and Width Indices for Hypertension and Type 2 Diabetes in a Large-Scale Screening: A Retrospective Cross-Sectional Study. Sci Rep (2018) 8(1):13284. doi: 10.1038/s41598-018-31624-x
13. Chien LC, Hsu FC, Bowden DW, Chiu YF. Generalization of Rare Variant Association Tests for Longitudinal Family Studies. Genet Epidemiol (2016) 40(2):101–12. doi: 10.1002/gepi.21951
14. Berndt SI, Gustafsson S, Mägi R, Ganna A, Wheeler E, Feitosa MF, et al. Genome-Wide Meta-Analysis Identifies 11 New Loci for Anthropometric Traits and Provides Insights Into Genetic Architecture. Nat Genet (2013) 45(5):501–12. doi: 10.1038/ng.2606
15. van de Bunt M, Manning Fox JE, Dai X, Barrett A, Grey C, Li L, et al. Transcript Expression Data From Human Islets Links Regulatory Signals From Genome-Wide Association Studies for Type 2 Diabetes and Glycemic Traits to Their Downstream Effectors. PloS Genet (2015) 11(12):e1005694. doi: 10.1371/journal.pgen.1005694
16. Jensen J, Rustad PI, Kolnes AJ, Lai YC. The Role of Skeletal Muscle Glycogen Breakdown for Regulation of Insulin Sensitivity by Exercise. Front Physiol (2011) 2:112. doi: 10.3389/fphys.2011.00112
17. Mhawech-Fauceglia P, Wang D, Samrao D, Liu S, DuPont NC, Pejovic T. Trefoil Factor Family 3 (TFF3) Expression and Its Interaction With Estrogen Receptor (ER) in Endometrial Adenocarcinoma. Gynecol Oncol (2013) 130(1):174–80. doi: 10.1016/j.ygyno.2013.03.030
18. Shults CL, Dingwall CB, Kim CK, Pinceti E, Rao YS, Pak TR. 17β-Estradiol Regulates the RNA-Binding Protein Nova1, Which Then Regulates the Alternative Splicing of Estrogen Receptor β in the Aging Female Rat Brain. Neurobiol Aging (2018) 61:13–22. doi: 10.1016/j.neurobiolaging.2017.09.005
19. Streicher C, Heyny A, Andrukhova O, Haigl B, Slavic S, Schüler C, et al. Estrogen Regulates Bone Turnover by Targeting RANKL Expression in Bone Lining Cells. Sci Rep (2017) 7(1):6460. doi: 10.1038/s41598-017-06614-0
20. Tiidus PM. Benefits of Estrogen Replacement for Skeletal Muscle Mass and Function in Post-Menopausal Females: Evidence From Human and Animal Studies. Eurasian J Med (2011) 43(2):109–14. doi: 10.5152/eajm.2011.24
Keywords: dual energy X-ray absorptiometry, genome-wide association study, loci, bone mass, fat mass, lean mass, body composition, genetics
Citation: Livingstone KM, Tan MH, Abbott G, Duckham RL, Croft L, Ward J, McEvoy M, Keske MA, Austin C and Bowe SJ (2021) Discovery Genome-Wide Association Study of Body Composition in 4,386 Adults From the UK Biobank’s Pilot Imaging Enhancement Study. Front. Endocrinol. 12:692677. doi: 10.3389/fendo.2021.692677
Received: 09 April 2021; Accepted: 03 June 2021;
Published: 22 June 2021.
Edited by:
Andrea P. Rossi, Integrated University Hospital Verona, ItalyReviewed by:
Marwan El Ghoch, Beirut Arab University, LebanonCopyright © 2021 Livingstone, Tan, Abbott, Duckham, Croft, Ward, McEvoy, Keske, Austin and Bowe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Katherine M. Livingstone, ay5saXZpbmdzdG9uZUBkZWFraW4uZWR1LmF1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.