 Zhenyu Xie
Zhenyu Xie Xin Li
Xin Li Yuzhen He
Yuzhen He Jian Zhang
Jian Zhang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Endocrinol. , 22 October 2020
Sec. Thyroid Endocrinology
Volume 11 - 2020 | https://doi.org/10.3389/fendo.2020.570604
Background: Papillary thyroid cancer has been associated with chronic inflammation. A systematic understanding of immune cell infiltration in PTC is essential for subsequent immune research and new diagnostic and therapeutic strategies.
Methods: Three different algorithms, single-sample gene set enrichment analysis (ssGSEA), immune cell marker and CIBERSORT, were used to evaluate immune cell infiltration levels (abundance and proportion) in 10 data sets (The Cancer Genome Atlas [TCGA], GSE3467, GSE3678, GSE5364, GSE27155, GSE33630, GSE50901, GSE53157, GSE58545, and GSE60542; a total of 799 PTC and 194 normal thyroid samples). Consensus unsupervised clustering divided PTC patients into low-immunity and high-immunity groups. Weighted gene coexpression network analysis (WGCNA) and gene set enrichment analysis (GSEA) were used to analyze the potential mechanisms causing differences in the immune response.
Results: Compared with normal tissues, PTC tissues had a higher overall immune level and higher abundance levels and proportions of M2 macrophages, Tregs, monocytes, neutrophils, dendritic cells (DCs), mast cells (MCs), and M0 macrophages. Compared with early PTC, advanced PTC showed higher immune infiltration and higher abundance levels and proportions of M2 macrophages, Tregs, monocytes, neutrophils, DCs, MCs, and M0 macrophages. Compared to the low-immunity group, the high-immunity group exhibited more advanced stages, larger tumor sizes, greater lymph node metastases, higher tall-cell PTCs, lower follicular PTC proportions, more BRAF mutations, and fewer RAS mutations. Epstein-Barr virus (EBV) infection was the most significantly enriched Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway for key module genes.
Conclusions: In human PTC, M2 macrophages, Tregs, monocytes, neutrophils, DCs, MCs, and M0 macrophages appear to play a tumor-promoting role, while M1 macrophages, CD8+ T cells, B cells, NK cells, and T follicular helper (TFH) cells (including eosinophils, γδ T cells, and Th17 cells with weak supporting evidence) appear to play an antitumor role. During the occurrence and development of PTC, the overall immune level was increased, and the abundance and proportion of tumor-promoting immune cells were significantly increased, indicating that immune escape had been aggravated. Finally, we speculate that EBV may play an important role in changing the immune microenvironment of PTC tumors.
Thyroid cancer (TC) is the fifth most common cancer in women in the USA, with an estimated 52,890 new TC cases nationwide in 2020 (1). The incidence of TC has been increasing in recent decades, and showing a trend of rejuvenation (2–7). TC also has the highest rate of growth among diagnosed tumors in the USA (8). Papillary thyroid carcinoma (PTC) belongs to differentiated thyroid carcinoma (DTC) and accounts for approximately 80–85% of all TCs (2, 9). Although the vast majority of PTCs can be cured by surgery and 131I treatment, 10% of patients still die from poorly differentiated and advanced tumors (10). A study with a median follow-up period of 27 years also showed that 28% of PTC patients relapsed and 9% died of PTC (11). The reason is partly because the interaction and collective effect of tumors with the surrounding immune microenvironment during the occurrence and development of the tumor promotes immune tolerance and then progresses to immune escape, resulting in the body’s inability to completely clear the tumor.
The immune editing hypothesis was proposed by Dunn in 2002 and divided the interaction between the immune system and the tumor into three stages: (1) elimination: tumor cells are cleared by the immune system before a clinical diagnosis is established. This stage is also called immune surveillance; (2) equilibrium: tumor cells with less immunogenicity gradually replace tumor cells with more immunogenicity; and (3) escape: tumor cells eventually escape the body’s immune surveillance (12). According to this hypothesis, the immune editing of tumors also sculpts the tumor’s immune phenotype while clearing the tumor, enabling the host to generate immune activity against the tumor. When the tumor is clinically diagnosed, it can not only tolerate the immune response but also manipulate the immune system to promote tumor progression. Poschke et al. (13) believe that tumors have two different methods of immune escape: camouflage and sabotage. Camouflage refers to the malformation and loss of the major histocompatibility complex class I (MHC-I) molecules on the surfaces of tumors, allowing tumors to escape the detection of the immune system. Sabotage refers to the ability of some tumors, including PTC, to manipulate part of the immune system to fight against the body’s immune response to protect themselves. In this process, tumors attract and even mediate some immune cells, such as tumor-associated macrophages (TAMs), myeloid-derived suppressor cells (MDSCs), Tregs, and mast cells (MCs). In general, MDSCs, Tregs, and MCs respond to uncontrolled inflammatory reactions in the body, but in tumor progression, they have created an immune microenvironment that allows this process. Existing evidence shows that Tregs, neutrophils, dendritic cells (DCs), MCs, and TAMs play a tumorigenic role in the PTC tumor microenvironment (TME), while CD8+ T cells, B cells, and NK cells play a protective role (14–16). However, the infiltration and role of some major immune cells in PTC are still controversial. Studies on nonprimary immune cells are scarce, and systematic research on the PTC immune microenvironment remains to be conducted. Therefore, accurate assessment of the specific pattern of immune cells in PTC, including not only its phenotype but also its function, is essential for subsequent immune research and new diagnostic and therapeutic strategies.
The purpose of this study was to systematically understand immune cell infiltration in the PTC microenvironment and to summarize the changes in the immune microenvironment during the occurrence and development of PTC. Our research found that M2 macrophages, Tregs, monocytes, neutrophils, DCs, MCs, and M0 macrophages play a tumor-promoting role, while M1 macrophages, CD8+ T cells, B cells, NK cells, and T follicular helper (TFH) cells play an antitumor role. During the occurrence and development of PTC, the overall immune level increased, and the abundance and proportion of tumor-promoting immune cells increased significantly. Our research will help researchers systematically understand PTC immune cell infiltration and provide a reference for subsequent basic and clinical research related to PTC immunity.
The RNA-seq data (level-three, FPKM) of the thyroid carcinoma (THCA) dataset were downloaded from The Cancer Genome Atlas (TCGA) (https://portal.gdc.cancer.gov/) and included 58 normal thyroid samples (N) and 512 PTC samples (T). The clinical data of THCA were downloaded from the University of California at Santa Cruz (UCSC) Xena platform (https://xena.ucsc.edu/).
The other nine PTC microarray datasets were downloaded from the Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo): GSE3467 (N = 9, T = 9), GSE3678 (N = 7, T = 7), GSE5364 (N = 16, T = 35), GSE27155 (N = 4, T = 51), GSE33630 (N = 45, T = 49), GSE50901 (N = 4, T = 61), GSE53157 (N = 3, T = 15), GSE58545 (N = 18, T = 27), and GSE60542 (N = 30, T = 33).
The ssGSEA score was used to quantify the enrichment level of 29 immune signatures in each THCA sample (17). To understand the overall situation of immune infiltration and facilitate visualization, we corrected the data as follows. For each ssGSEA score, Xi was replaced with Xi ′ with the equation Xi ′ = (Xi −Xmin)/(Xmax − Xmin), where Xmin and Xmax represent the minimum and maximum values of the ssGSEA score in the THCA sample, respectively (18). The same method was also applied to the other nine GEO datasets.
Additionally, we also measured the infiltration abundance of THCA immune cells through immune cell-specific markers (19). Given that its algorithm and results are highly similar to ssGSEA and are also measurements of immune cell abundance, we present the results in the form of a supplementary graph.
CIBERSORT is a deconvolution algorithm based on RNA-seq data to calculate the composition ratio of 22 immune cells in each sample (20). We performed 1,000 permutations and retained samples of p < 0.05 to ensure the reliability of the results, and the sum of various immune cells was 1. Considering the small sample size of each data set except TCGA after deconvolution, the results of nine GEO data sets were integrated into one validation set.
The Spearman correlation between each pair of cells was calculated and visualized using the R package “corrplot”. Hierarchical agglomerative clustering based on Ward’s linkage and Euclidean distance was used to group immune cells in the PTC microenvironment. An unsupervised clustering algorithm (K-means) was applied to identify immune infiltration patterns and classify patients for further analysis. The results were visualized in the form of an immune network (21).
Based on the ssGSEA scores of 29 immune signatures, the R package “ConsensusClusterPlus” was used to perform unsupervised clustering of the THCA samples (22). The cumulative distribution function (CDF), consensus heat map, and principal component analysis (PCA) (23) were applied to evaluate the optimal K (number of groups). We renamed the two groups to the low-immunity group (L-immunity) and to high-immunity group (H-immunity) based on the differences in the ssGSEA scores between the two groups after clustering.
ESTIMATE was used to evaluate the composition of the TME for each THCA sample (24) and included immune score (immune cell infiltration), stromal score (stromal content), ESTIMATE score (stromal-immune comprehensive score) and tumor purity.
First, differentially expressed genes (DEGs) between PTC and adjacent normal tissues in TCGA dataset were identified with the R package “limma” (|log2FC| > 1 and adjusted p-value < 0.05).
WGCNA aims to identify coexpressed gene modules, explore associations between gene networks and traits of interest, and discover hub genes in networks (25). DEGs were divided into different gene modules by the R package “WGCNA” (minModuleSize = 30).
The module with the highest absolute module significance was defined as the key module. A gene that satisfies the following conditions was defined as a hub gene.
1. The gene is in the key module.
2. The absolute module membership (MM) > 0.5, which represents Pearson’s correlation between the gene and the module.
3. The absolute gene significance (GS) > 0.5, which represents Pearson’s correlation between the gene and the target traits.
The genes in the key module were subjected to Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses with DAVID 6.8 (https://david.ncifcrf.gov/).
The hub genes were input into the STRING database (v11.0) (https://string-db.org/) to analyze the interaction network of the hub gene-encoded proteins. The minimum required interaction score was set to 0.4 (medium confidence), and the results were visualized by Cytoscape (v3.7.2).
GSEA software (version 4.0.3) (https://www.gsea-msigdb.org/gsea/) was used to explore the main biological function that caused differences between the L-immunity and H-immunity groups. c2.cp.kegg.v6.2.symbols.gmt was set to the reference gene set and permutations=1000 for each analysis. |normalized enrichment score (NES) |≥1.0, p-value ≤ 0.05, and false discovery rate (FDR) q-val ≤ 0.25 were considered statistically significant.
The t-test and Mann-Whitney U test were used for comparisons between two groups. The chi-square test was used to assess differences in clinical parameters between the L-immunity and H-immunity groups. The Pearson and Spearman method was used for correlation analysis. The log-rank method was used to calculate the significant survival p-values (progression-free survival (PFS) as the ending indicator). R software (version 3.6.1) and SPSS (v25.0) software were used to analyze the data. R, GraphPad Prism (v8.0) software and Excel were used for data visualization. p < 0.05 was considered significant.
ssGSEA evaluated 29 immune signatures between PTC and adjacent normal tissues in TCGA (Figure 1A), and the results showed that Tregs, Th1 cells, T helper cells, plasmacytoid DCs (pDCs),immmature DCs (iDCs), DCs, activated DCs (aDCs), neutrophils, MCs, and macrophages increased in PTC (p < 0.05), while CD8+ T cells decreased in PTC. There were no significant differences in TILs, TFH cells, NK cells, or B cells. Additionally, the type II IFN response, the type I IFN response, parainflammation, MHC class I, HLA, checkpoint, chemokine receptor (CCR) and APC costimulation were elevated in PTC, indicating overall enhancement of the immune system in PTC. The ssGSEA results of the nine GEO validation sets (GSE3467, GSE3678, GSE5364, GSE27155, GSE33630, GSE50901, GSE53157, GSE58545, and GSE60542) also supported the overall enhancement of the immune system in PTC (Figures 1B–J).
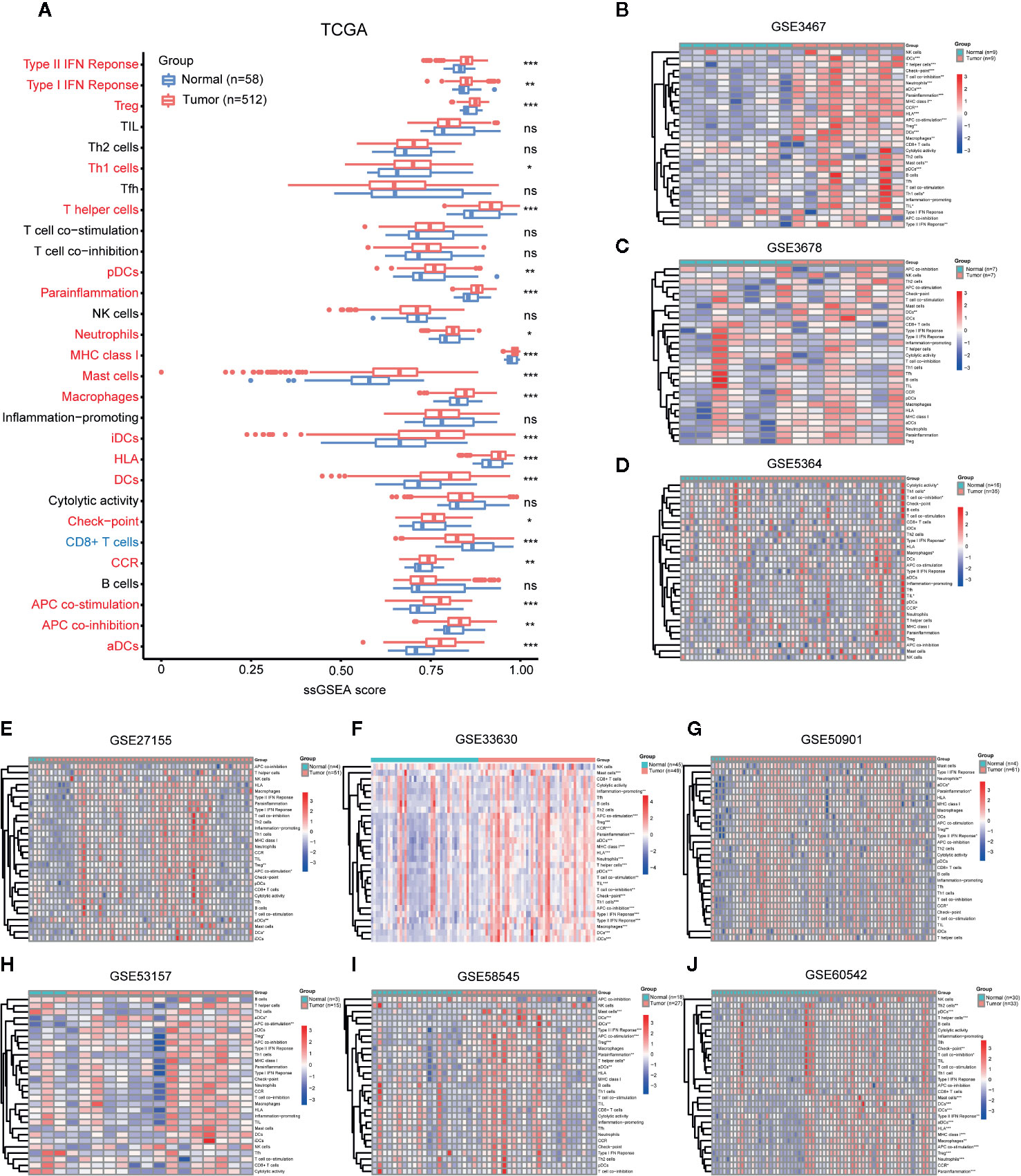
Figure 1 Comparison of immune cell infiltration (abundance) between PTC and normal tissues. Comparison of ssGSEA scores of 29 immune signatures between PTC and normal tissues in (A) TCGA, (B) GSE3467, (C) GSE3678, (D) GSE5364, (E) GSE27155, (F) GSE33630, (G) GSE50901, (H) GSE53157, (I) GSE58545, and (J) GSE60542. *p < 0.05, **p < 0.01, ***p < 0.001. ns, not significant.
In addition, immune cell markers in TCGA showed that TPSAB1 (MCs), IL3RA (pDCs), CD68 (macrophages), CD1A (iDCs), and B3GAT1 (NK cells) were overexpressed in PTC. PTPRC (memory T cells), MS4A1 (B cells), IL17A (Th17 cells), CXCR5 (TFH cells), and CD8A (cytotoxic T cells) decreased in PTC (Figure S1A).
To summarize the immune cell infiltration results (abundance) of ssGSEA and immune cell markers:
1. Compared with the immune system in normal tissues, the immune system in PTC appears to be enhanced overall.
2. Compared with normal tissues, PTC tissues have an increase in the number of tumor-promoting immune cells, which is particularly significant.
3. Compared with normal tissues, the two algorithms have slightly different evaluations of antitumor immune cells in PTC. The ssGSEA results show that no significant differences in antitumor immune cells, while the immune cell marker results show a downward trend for antitumor immune cell abundance in PTC compared with normal tissues.
CIBERSORT was used to calculate the proportion of each of the 22 immune cell types based on RNA-seq data. We performed measurements on the TCGA and GEO datasets (Figures 2A–D). The GEO dataset incorporates valid samples from nine datasets, including GSE3467, GSE3678, GSE5364, GSE27155, GSE33630, GSE50901, GSE53157, GSE58545, and GSE60542.
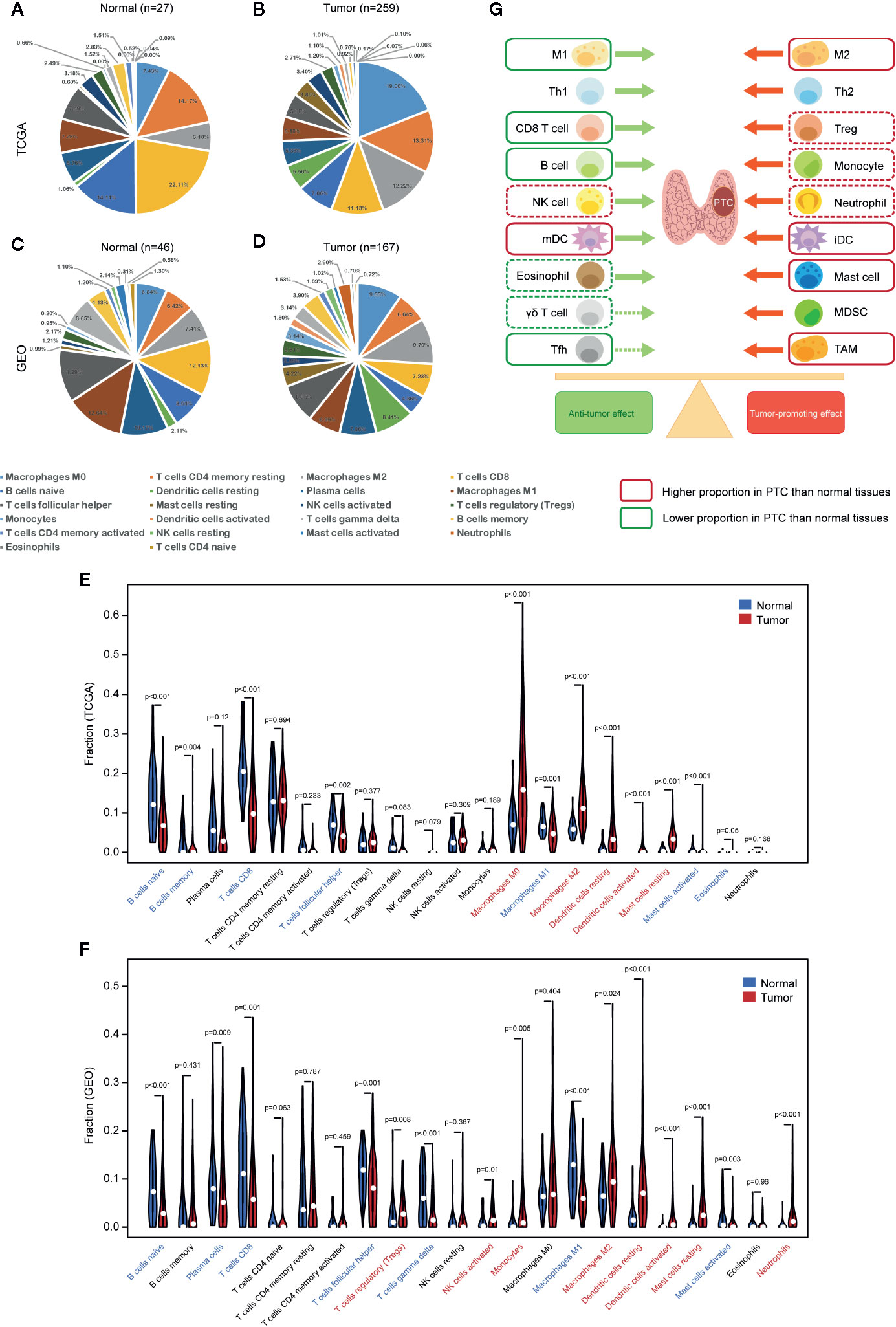
Figure 2 Comparison of immune cell infiltration (proportion) between PTC and normal tissues. Proportions of 22 types of immune cells (CIBERSORT) in (A) normal tissues (TCGA), (B) PTC tissues (TCGA), (C) normal tissues (GEO), and (D) PTC tissues (GEO). (E) Comparison of immune cell proportions between PTC and normal tissues (TCGA). (F) Comparison of immune cell proportions between PTC and normal tissues (GEO). (G) Balance chart of the differences in immune cell infiltration between PTC and normal tissues (TCGA and GEO). The dashed box indicates that the difference is significant in TCGA or GEO, while the solid box indicates that the difference is significant in both TCGA and GEO. GEO samples are from nine data sets, including GSE3467, GSE3678, GSE5364, GSE27155, GSE33630, GSE50901, GSE53157, GSE58545, and GSE60542. mDC, mature dendritic cell, iDC, immature dendritic cell.
We compared the proportions of immune cells between the PTC and normal tissues in both TCGA and GEO datasets. In TCGA, normal tissues had higher levels of naive B cells, memory B cells, CD8+ T cells, TFH cells, M1 macrophages, activated MCs, and eosinophils than PTC tissues, while PTC tissues had higher levels of M0 macrophages, M2 macrophages, resting DCs, activated DCs, and resting MCs than normal tissues (Figure 2E). In GEO, normal tissues had higher levels of naive B cells, plasma cells, CD8+ T cells, TFH cells, T cells gamma delta, M1 macrophages, and activated MCs than PTC tissues, while PTC tissues had higher levels of Tregs, activated NK cells, monocytes, M2 macrophages, resting DCs, activated DCs, resting MCs, and neutrophils than normal tissues (Figure 2F). To assist with the understanding of these results, we listed the results in a balance chart (Figure 2G). The construction of the balance chart mainly refers to the reviews by Varricchi et al. (14), Galdiero et al. (15) and Ferrari et al. (16) and the literature table that we created (Table 1).

Table 1 Immune cells in the PTC microenvironment.
In summary, compared with normal thyroid tissues, PTC tissues have a higher proportion of tumor-promoting immune cells and a lower proportion of antitumor immune cells.
ssGSEA was used to analyze the relationship between PTC progression and immune cell infiltration in TCGA. The results showed that patients with high tumor stage, lymph node metastasis, and larger tumor size had an increased number of immune cells, and the increase in tumor-promoting immune cells was particularly significant, including Th1 cells, macrophages, Tregs, monocytes, neutrophils, iDCs and MCs (Figures 3A–C). Patients with tumor metastases had lower immunity levels than patients without metastases (Figure 3D).
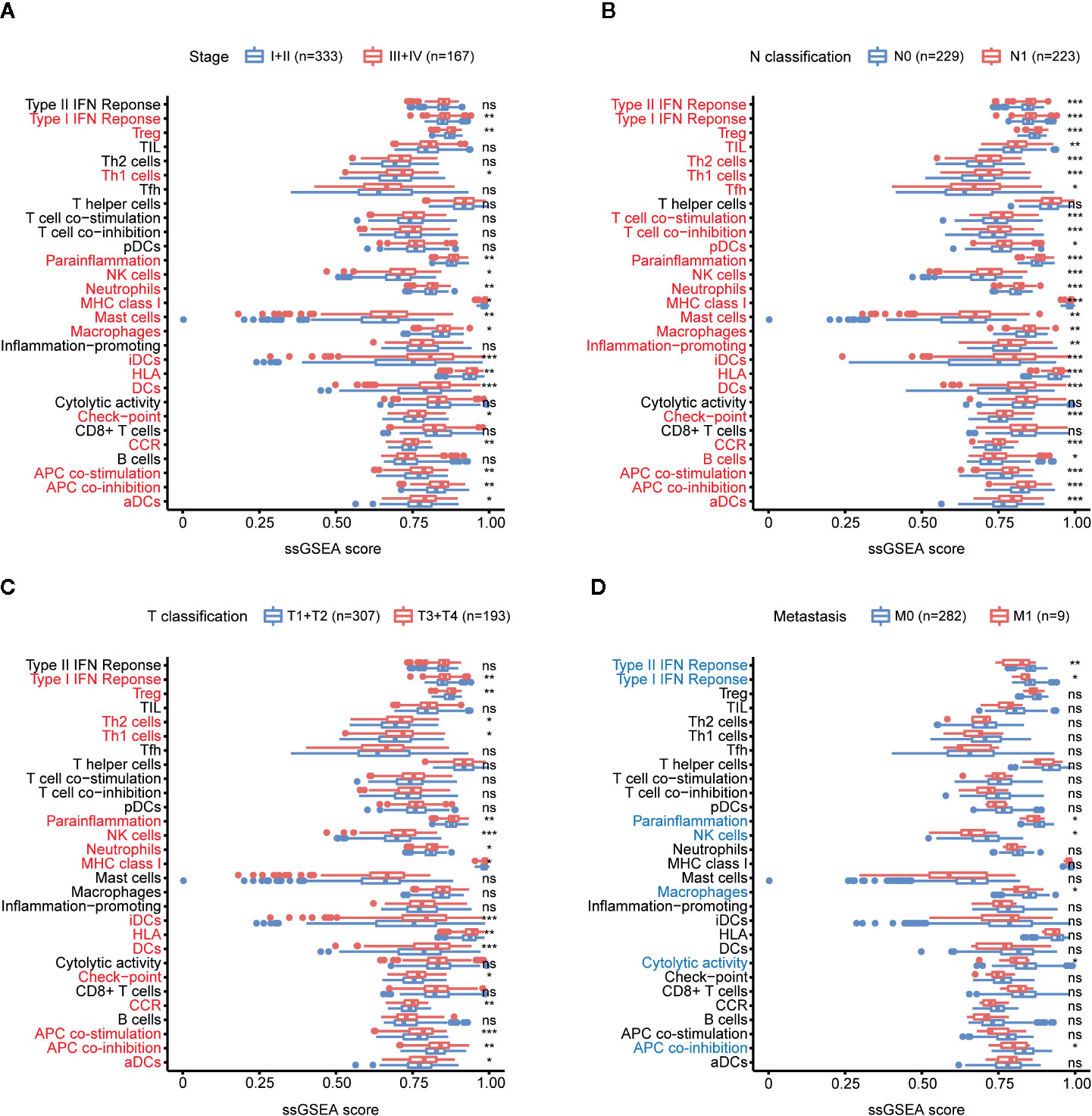
Figure 3 Relationship between immune cell infiltration (abundance) and PTC progression. (A) Comparison of the ssGSEA scores of 29 immune signatures between stages I and II PTC patients and stages III and IV PTC patients. (B) Comparison of the ssGSEA scores of 29 immune signatures between N0 and N1 PTC patients. (C) Comparison of the ssGSEA scores of 29 immune signatures between T1 and T2 PTC patients and T3 and T4 PTC patients. (D) Comparison of the ssGSEA scores of 29 immune signatures between PTC patients with and without tumor metastasis. *p < 0.05, **p < 0.01, ***p < 0.001. ns, not significant.
In addition, the results of immune cell markers also showed that PTC patients with a high tumor stage, lymph node metastasis, and a larger tumor size had an increased number of immune cells, and the increase in tumor-promoting immune cells was particularly significant, including TPSAB1 (MCs), FOXP3 (Tregs), CD1A (iDCs), and neutrophils (CXCR2s) (Figures S1B–D).
In summary, with the progression of PTC, immune cell infiltration increased comprehensively.
CIBERSORT analyzed the relationship between PTC progression and immune cell infiltration in TCGA. The results show that TFH cells, CD8+ T cells, plasma cells, and M1 macrophages have a higher proportion in stages I and II than in other stages. Resting memory CD4+ T cells, neutrophils, monocytes, resting MCs, M2 macrophages, M0 macrophages, resting DCs, and activated DCs were higher in stages III and IV (Figure 4A). CD8+ T cells were higher in N0 PTC, and T cells gamma delta, resting memory CD4+ T cells, resting DCs, and activated DCs were higher in N1 PTC (Figure 4B). TFH cells, CD8+ T cells, plasma cells, M1 macrophages, and memory B cells were higher in T1 + T2 PTC. Resting memory CD4+ T cells, neutrophils, monocytes, M2 macrophages, M0 macrophages, resting DCs, and activated DCs were higher in T3 + T4 PTC (Figure 4C). These results are further shown in a balance chart (Figure 4D).
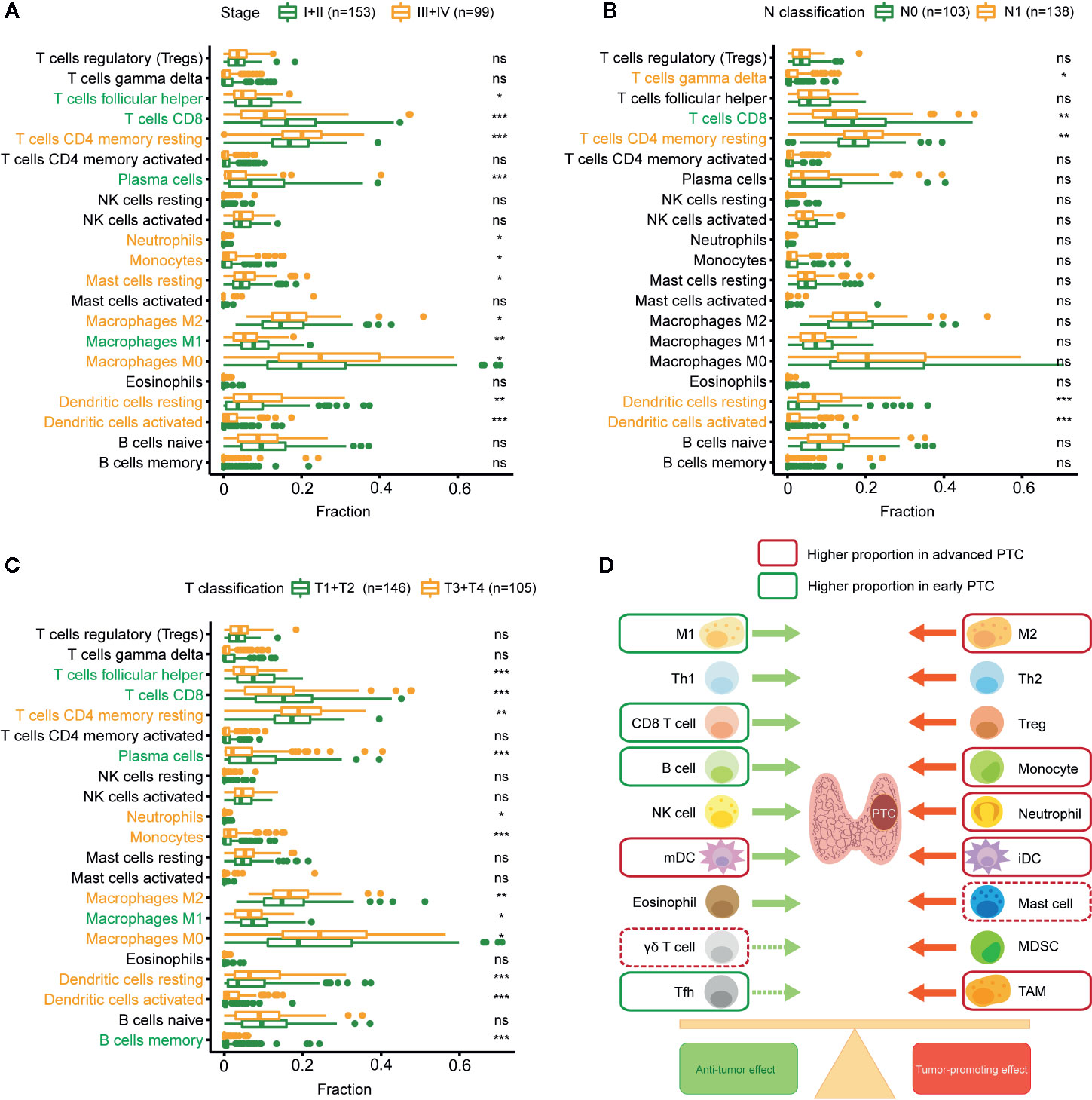
Figure 4 Relationship between immune cell infiltration (proportion) and PTC progression. (A) Comparison of immune cell proportion (CIBERSORT) between stages I and II PTC patients and stages III and IV PTC patients. (B) Comparison of immune cell proportions between N0 and N1 PTC patients. (C) Comparison of immune cell proportions between T1 and T2 PTC patients and T3 and T4 PTC patients. (D) Balance chart of the relationship between immune cell infiltration and PTC progression (TCGA). *p < 0.05, **p < 0.01, ***p < 0.001. ns, not significant.
In summary, compared with early PTC, advanced PTC has a higher proportion of tumor-promoting immune cells and a lower proportion of antitumor immune cells.
We performed correlation analysis on immune cells based on ssGSEA (abundance) (Figure 5A) and CIBERSORT (proportion) (Figure 5B). The ssGSEA results showed a high positive correlation among all immune cells. The CIBERSORT results showed a positive correlation among antitumor immune cells (TFH cells, M1 macrophages, plasma cells, and CD8+ T cells), and these antitumor immune cells were negatively correlated with immune cells with tumor-promoting effects (M0 macrophages, M2 macrophages, Tregs, activated DCs, and resting MCs).
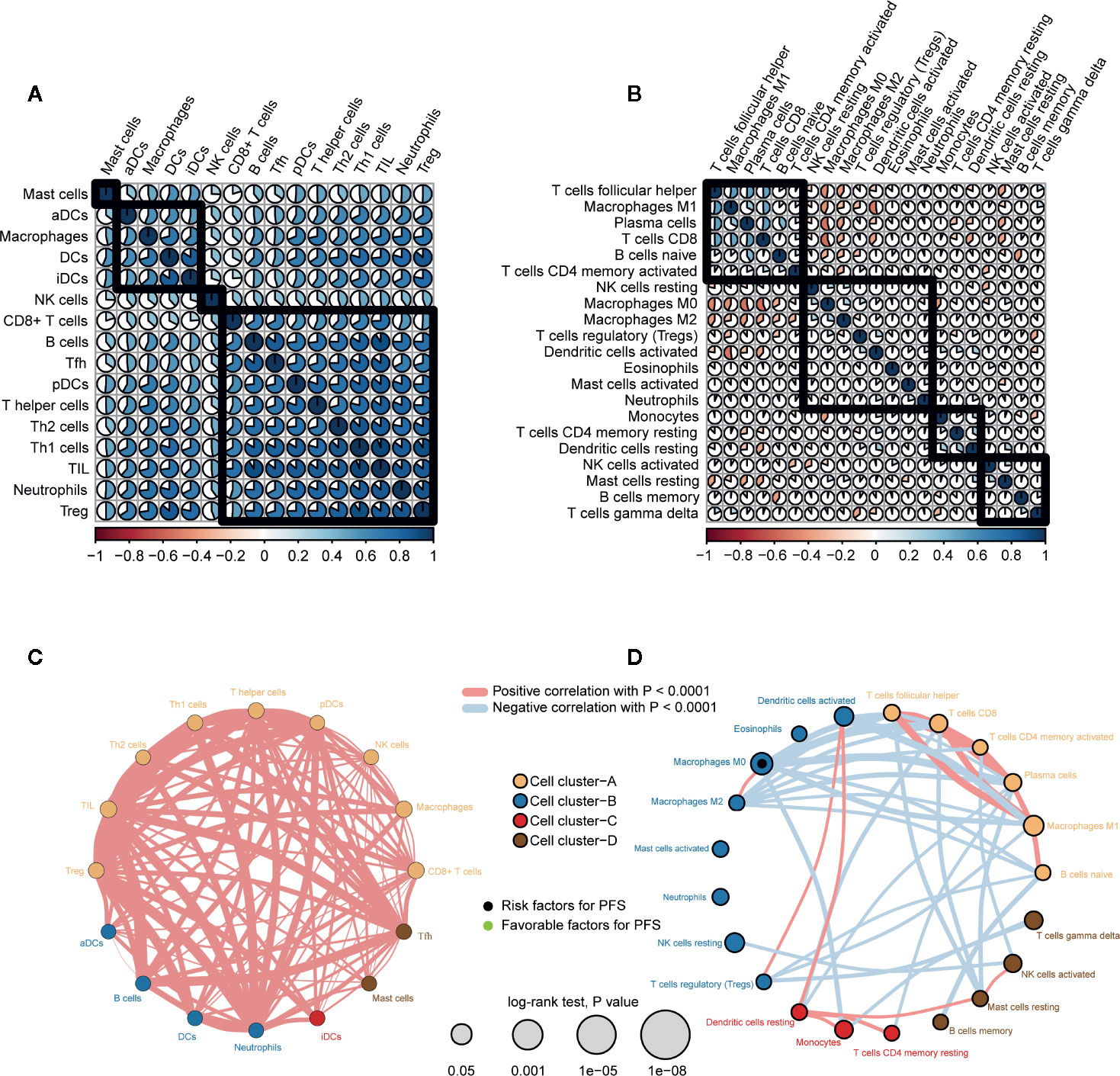
Figure 5 Correlation and clustering of immune cell types in PTC. (A) Spearman correlation analysis between immune cells types (abundance, ssGSEA) in PTC. (B) Spearman correlation analysis between immune cell types (proportion, CIBERSORT) in PTC. (C) Immune network of immune cell types (abundance). The size of each circle represents the effect of each immune cell type on prognosis (PFS). Thicker lines indicate higher Spearman correlations between immune cell types. (D) Immune network of immune cell types (proportion).
We also clustered immune cells from the perspective of abundance (Figure 5C) and proportion (Figure 5D) and examined the effect of each immune cell type on prognosis. CIBERSORT results in immune cells (TFH cells, CD8+ T cells, activated memory CD4+ T cells, plasma cells, M1 macrophages, naive B cells) clustered in cell cluster A. In addition, except for M0 macrophages (proportion) (HR = 11.8, log rank p = 0.032), the effects of other immune cells on PFS did not reach statistical significance.
In summary, there was a positive correlation among all immune cells. However, antitumor immune cells and tumor-promoting immune cells oppose each other at a proportional level of correlation.
Consensus unsupervised clustering was performed on 502 PTC patients based on 29 immune signatures of ssGSEA. We considered the relative change in area under the CDF curve at K = 2 to 9 (Figures 6A, B) and found that the consensus heatmaps showed stable partitions when the samples were clustered into two groups (k = 2) (Figure 6C). PCA of all genes revealed significant differences between the two groups (Figure 6D). We renamed the two groups to the H-immunity (n = 293) and L-immunity groups (n = 209) based on the differences in immune signatures (Figure 6E).
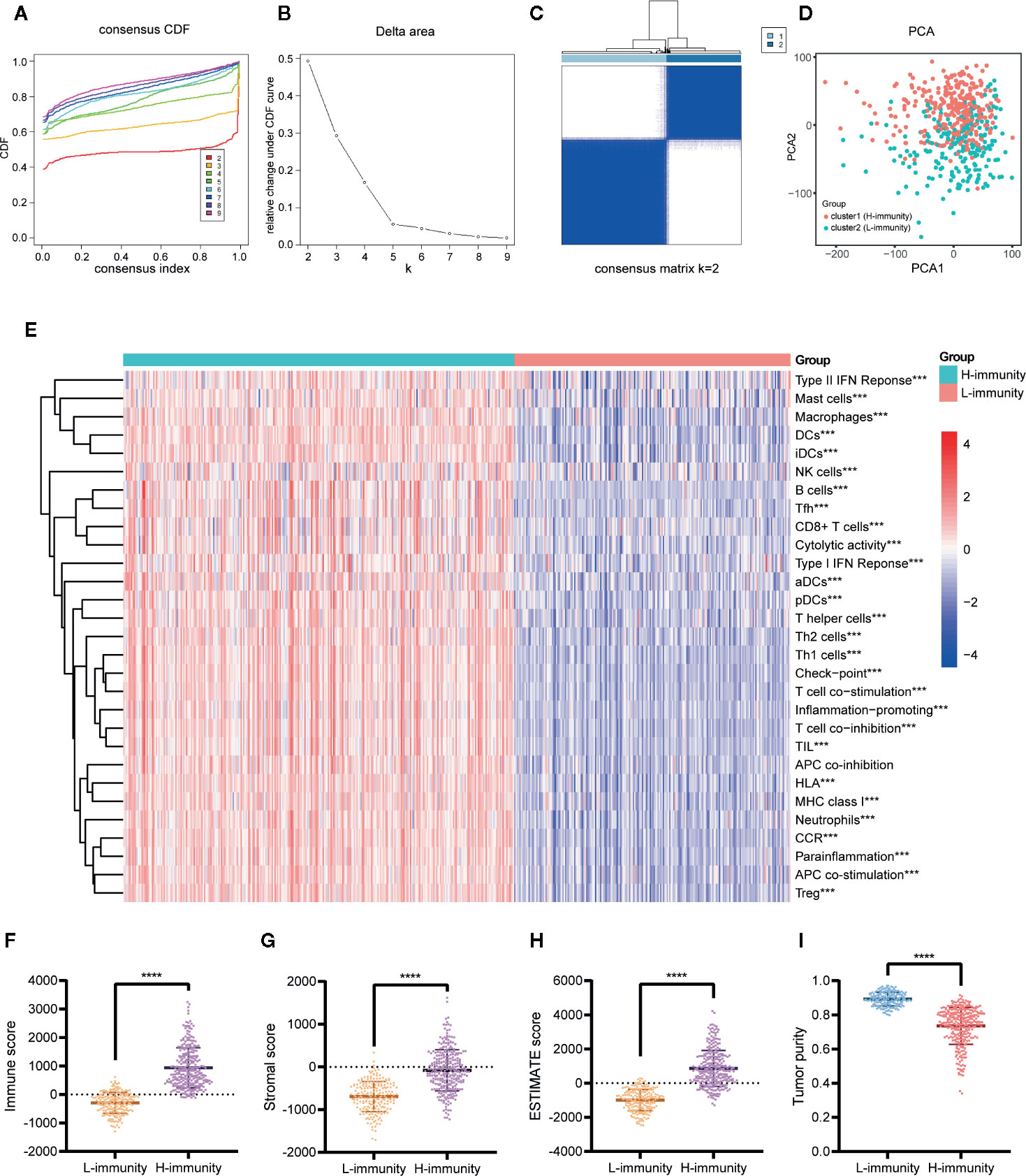
Figure 6 Consensus clustering of PTC patients based on immune signatures. (A) Consensus CDF curve (K = 2–9). (B) Relative change in area under the CDF curve (K = 2–9). (C) Consensus heatmap of K = 2. (D) PCA of all genes for the unsupervised clustering results (K = 2). (E) Comparison of the ssGSEA scores of 29 immune signatures between the cluster1 (H-immunity) and cluster2 (L-immunity) groups. Comparison of the (F) immune scores, (G) stromal scores, (H) ESTIMATE scores, and (I) tumor purity between the L-immunity and H-immunity groups. ***p < 0.001, ****p < 0.001.
The composition of the TME of PTC samples was analyzed by ESTIMATE. The results showed that the H-immunity group had higher immune scores (Figure 6F), stromal scores (Figure 6G), and ESTIMATE scores (Figure 6H) and lower tumor purity (Figure 6I) than the L-immunity group.
In summary, we divided PTC patients into L-immunity and H-immunity groups by consensus clustering and verified the excellent discrimination ability of this grouping for all genes, immune signatures, and TME compositions.
To understand the effects of immune levels on clinical traits in PTC patients, we compared clinical parameters between the H-immunity and L-immunity groups (Table 2). Compared to patients in the L-immunity group, patients in the H-immunity group had an advanced American Joint Committee on Cancer (AJCC) stage, advanced T classification, lymph node metastasis, higher tall-cell PTC and lower follicular PTC proportions, more BRAF mutations and fewer RAS mutations.
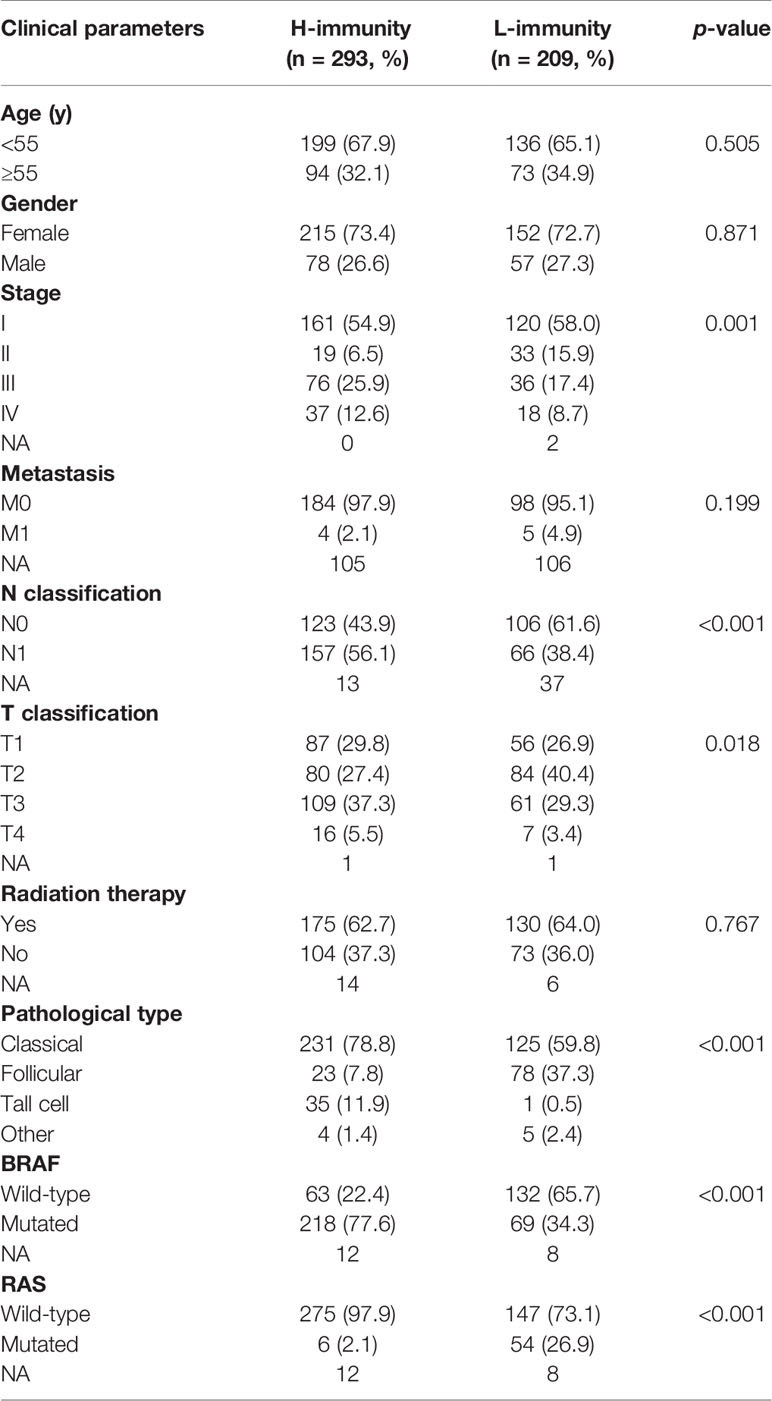
Table 2 Comparison of the clinical parameters between the H-immunity and L-immunity groups in PTC.
To construct a weighted coexpression network and screen for modules and genes related to PTC immunity, 6254 DEGs between PTC (n = 512) and normal tissues (n = 58) in TCGA were selected for subsequent WGCNA (502 patients with complete clinical information were selected) (Figure 7A). After a series of adjustments to the WGCNA parameters, the DEGs were divided into 19 modules by average linkage hierarchical clustering (Figures 7B–D). The correlation heatmap showed that the pink module, which contains 276 genes, had the highest correlation with the H-immunity group (Pearson’s correlation coefficient = 0.51, p < 0.001) (Figure 7E). The 34 genes in the pink module with MM > 0.5 and GS > 0.5 were defined as hub genes (Figure 7F).
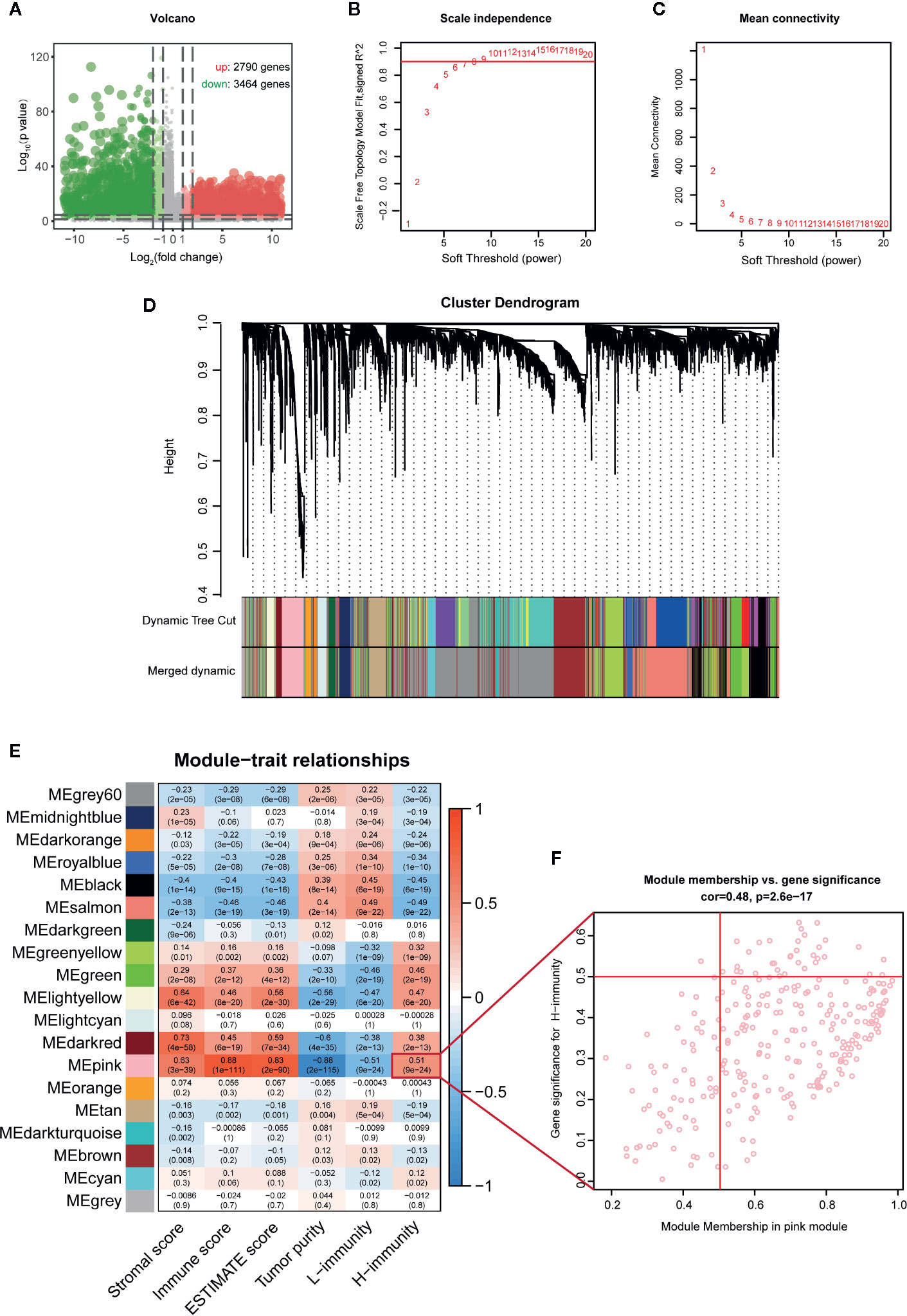
Figure 7 WGCNA screening key modules and hub genes related to PTC immunity. (A) Volcano map of DEGs between PTC and adjacent tissues (TCGA). (B) Calculation of the scale-free fit index of various soft-thresholding powers (β). (C) Analysis of the mean connectivity of various soft-thresholding powers (β). (D) Clustering dendrogram of 502 PTC patients, where 6254 DEGs were clustered based on the dissimilarity measure (1-TOM) and divided into 19 modules. (E) Correlation heatmap between module eigengenes and immune traits; the modules with high relevance to the H-immunity group are the key modules of mechanism research. (F) Scatter plot of the key module eigengenes (pink).
A total of 276 pink module eigengenes were used for functional and pathway enrichment analyses to explore potential mechanisms that cause differences in the immune microenvironment between PTCs. “Response to interferon-gamma”, “external side of plasma membrane”, and “chemokine receptor binding” were the most common GO terms for biological processes, cellular components, and molecular functions, respectively (Figure 8A). “Epstein-Barr virus infection” was the most significantly enriched pathway in KEGG analysis (p-adjust < 0.001) (Figure 8B). Additionally, we constructed a PPI network with the 34 hub genes and found that TYROBP, CTLA4, CSF1R, CTSS, and HLAs play key roles (Figure 8C).
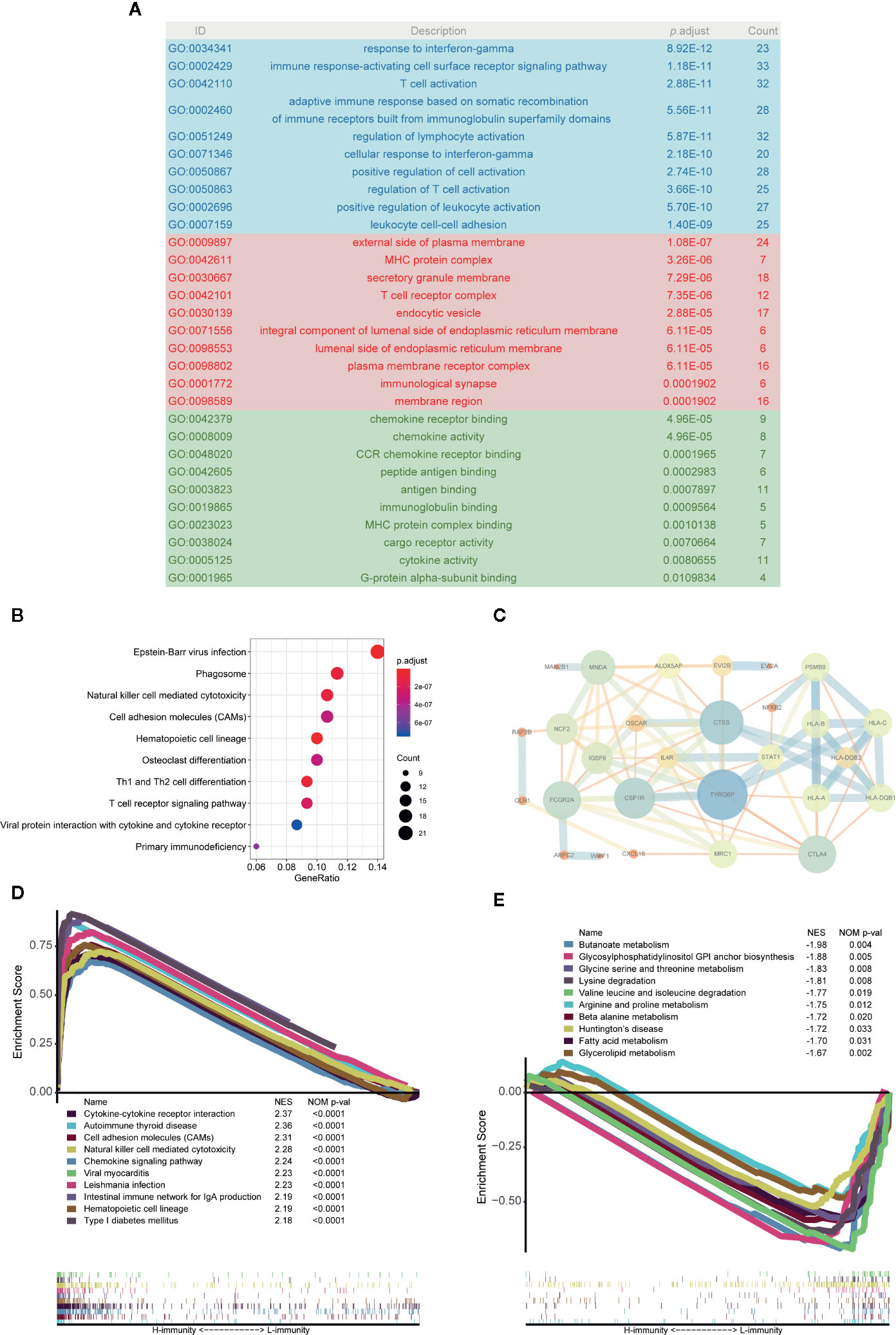
Figure 8 Analysis of the main mechanism of PTC immunity. (A) GO analysis of 276 pink module eigengenes; the blue, red and green areas represent biological processes, cellular components, and molecular functions, respectively. (B) KEGG analysis of pink module eigengenes: the top 10 significantly enriched pathways are shown. (C) Protein-protein interactions (PPIs) of hub genes. (D) Top 10 enriched pathways in the H-immunity group based on GSEA. (E) Top 10 enriched pathways in the L-immunity group based on GSEA.
GSEA was also used to identify the mechanism and functional differences between the L-immunity and H-immunity groups. The H-immunity group was enriched in the following terms: cytokine-cytokine receptor interaction, autoimmune thyroid disease, cell adhesion molecules (CAMs), natural killer cell-mediated cytotoxicity, chemokine signaling pathway, viral myocarditis, Leishmania infection, intestinal immune network for IgA production, hematopoietic cell lineage, and type I diabetes mellitus (Figure 8D). Conversely, the L-immunity group was enriched in the following metabolism-related pathways: butanoate metabolism; glycosylphosphatidylinositol (GPI) anchor biosynthesis; glycine, serine, and threonine metabolism; lysine degradation; valine, leucine, and isoleucine degradation; arginine and proline metabolism; beta alanine metabolism; Huntington’s disease; fatty acid metabolism; and glycerolipid metabolism (Figure 8E).
Our findings showed that during the occurrence and development of PTC, the immune system is enhanced; moreover, M2 macrophages, Tregs, monocytes, neutrophils, DCs, MCs, and M0 macrophages play a tumor-promoting role, and their abundances and proportions are significantly increased. M1 macrophages, CD8+ T cells, B cells, NK cells, and TFH cells (eosinophils, γδ T cells, and Th17 cells with weak supporting evidence) exert an antitumor effect, and the proportions decreased.
PTC is subdivided into “inflammatory” tumors (71). Extensive evidence shows that autoimmunity and inflammation are risk factors for TC (72). Increasing evidence also suggests that cancer-related inflammation may be a useful target for new diagnostic and therapeutic strategies in TCs (73). Therefore, to evaluate the specific pattern of immune cells involved in PTC, including not only its phenotype but also its function, it is essential to understand the immunological characteristics of PTC. Many major advances have been made in the study of immune infiltration in PTC, especially in major immune cells (B cells, TAMs, NK cells, neutrophils, DCs, MCs, CD8+ T cells and Tregs) (14–16). A considerable part of our research is repeating, verifying, and integrating previously reported results of immune cell phenotypes and functions in PTC. We used ssGSEA, immune cell marker and CIBERSORT to evaluate the infiltration level of immune cells in 10 datasets (a total of 799 PTC and 194 normal samples) from the perspective of abundance and proportion, respectively. Furthermore, it is essential to ensure that the results of the analysis are highly reliable. We also analyzed subtypes of major immune cells and immune cells that lacked attention (such as M1 macrophages, M2 macrophages, Th1, Th2, pDCs, iDCs, eosinophils, monocytes, TFH cells, T cells gamma delta, and Th17 cells) This will help researchers comprehensively understand the PTC immune microenvironment. Our research supports the evidence that M2 macrophages, Tregs, monocytes, neutrophils, DCs, MCs, and TAMs play a tumor-promoting role, while M1 macrophages, CD8+ T cells, B cells, NK cells, and TFH cells (eosinophils, γδ T cells, and Th17 cells with weak supporting evidence) exert an antitumor effect. Among them, related reports of the phenotype and function of monocytes, γδ T cells, TFH cells, and eosinophils in PTC patients are still lacking. Inflammation of monocyte recruitment and activation is associated with the development of TC in a mouse model (74). IL-17-producing γδT17 cells play a decisive role in the chemotherapy-induced anticancer immune response (75). TFH cells can regulate antibody production by B cells (76); the frequency of TFH cells increases in patients with autoimmune thyroid disease (AITD), and a significant number of TFH cells are also detected in the thyroid tissues of patients with Hashimoto’s thyroiditis (77). The above evidence also supports our conclusions about the function of immune cells.
Our research shows that compared with the immune level in normal tissues, the immune level in PTC tissues is generally higher, and the increase in tumor-promoting immune cells is particularly significant. The proportion of immune cells in the TME is also favoring tumor-promoting immune cells. The elevation of TAMs, DCs, MCs, neutrophils, NK cells, and Tregs in PTC has been reported (28–31, 36, 38, 41, 45, 55, 78). Our study found that these tumor-promoting immune cells were recruited in large numbers in PTC and exhibited a tumor-promoting immune microenvironment phenotype. This indicates that PTCs manipulate partial immune cells such as M2 macrophages, Tregs, monocytes, neutrophils, DCs, MCs, and M0 macrophages to fight against the body’s immune response to protect themselves, which is also called “sabotage” (13). Eventually, the tumor evaded immune surveillance and escaped the immune system.
In additional research, we found that immune cell infiltration increased during tumor progression, and the increase in tumor-promoting immune cells was particularly significant. The proportion of immune cells in the TME was further tilted towards tumor-promoting immune cells. Previous mainstream studies have shown that TAMs, DCs, MCs, neutrophils, and Tregs are positively correlated with PTC progression (24, 29–34, 38, 41, 42, 46, 48, 49, 51, 58, 61, 68, 69, 79), and B cells, CD8+ T cells, NK cells, and iDCs are negatively correlated with PTC progression (26, 37, 56, 58–62). Unlike previous studies, our research found that both the abundance and proportion of tumor-promoting immune cells were positively correlated with PTC progression, while the proportion of antitumor immune cells during the progression of PTC decreased (CIBERSORT), and the abundance increased (ssGSEA) or did not significantly change (immune cell marker). These results show that in escalating immune confrontation, immune escape is aggravated, and the tumor’s ability to fight the immune response of the body is further strengthened by recruiting immunosuppressive cells. Eventually, the dynamic balance between antitumor and tumor-promoting immune cells in the original immune system has allowed the irreversible development of tumors. The positive correlations among the H-immunity group, tumor stage, lymph node metastasis, and tumor size also support the above hypothesis.
Our analysis of the correlation of the abundances of immune cells in PTC showed a high positive correlation among all examined cells, indicating that the immune system as a whole has a high degree of consistency. However, antitumor immune cells and tumor-promoting immune cells oppose each other at a proportional level of correlation. Our clustering results on immune cells also show the opposition between immune cells that exert procancer and anticancer effects. We speculate that in the process of tumor progression, the confrontation between tumor-killing immune cells and tumor-controlled immunosuppressive cells in the human immune system continues to escalate, and eventually, the proportion of tumor-promoting immune cells in the TME will have an irreversible advantage. This indicates that the tumor cells escaped the body’s immune surveillance by “sabotage” and achieved immune escape. Except for M0 macrophages (proportion) in our prognosis studies, other immune cells did not reach significant levels. Previous studies in PTC have generally shown that TAMs and neutrophils are associated with a worse prognosis (28, 51–54), while CD8+ T cells and DCs are associated with a better prognosis (26, 39, 62, 63). In fact, the roles of various immune cells in prognosis are still controversial (26, 64), and there are many negative results (36, 48, 50, 64, 70). Considering the positive correlation between immune cell infiltration and tumor progression, we suspect that the insignificant prognosis of immune cells may be related to the lower number of outcomes, such as death, recurrence and metastasis of PTC.
We divided patients into L-immunity and H-immunity groups by consensus clustering and verified the excellent discrimination ability of this grouping for all genes, immune signatures, and TME composition. Compared to patients in the L-immunity group, patients in the H-immunity group showed a more advanced stage, larger tumor size, more lymph node metastasis, higher tall-cell PTC proportions, lower follicular PTC proportions, more BRAF mutations and fewer RAS mutations. The BRAF V600E mutation is related to the immunosuppressive mechanism of PTC; compared to the ratio in wild-type BRAF PTC, the CD8+/FoxP3+ ratio in BRAF V600E mutant PTC was significantly reduced, and the proportion of M2 type TAMs was increased (27, 80, 81). We speculate that somatic mutations in genes such as BRAF and RAS in PTC may initiate the changes in the tumor immune microenvironment.
WGCNA and GSEA were used to explore the underlying mechanisms that cause PTC immune differences, and a number of significantly different immune-related functions and pathways were found. What caught our attention was that Epstein-Barr virus (EBV) infection was the most significantly enriched KEGG pathway. Despite some controversy, many studies have pointed out that EBV is highly expressed in patients with TC and is associated with increasing development of thyroid tumors (82–85). Studies related to nasopharyngeal carcinoma (NPC), Burkitt lymphoma (BL) and gastric cancer have found that EBV can evade host immune recognition and latent infection in B lymphocytes, can epigenetically suppress host tumor suppressor genes, and can provide a potential “hit and run” mechanism for viral carcinogenesis (86–88). We speculate that EBV may play an important role in the change in the PTC immune microenvironment.
However, this study has some limitations. First, ssGSEA, immune cell markers, and CIBERSORT are algorithms based on RNA-seq data, and the abundant results of ssGSEA and immune cell markers are inconsistent in some antitumor immune cells. We have shown their respective results and common conclusions. Second, the research on the mechanism that causes immune differences was based on the inference of several algorithms and has not been experimentally verified.
In the next study, single-cell analysis of immune cells around and within tumors will be an important direction for PTC immune research to elucidate the function of immune cells in the PTC TME.
In conclusion, our research shows that M2 macrophages, Tregs, monocytes, neutrophils, DCs, MCs, and M0 macrophages play a tumor-promoting role in PTC, while M1 macrophages, CD8+ T cells, B cells, NK cells, TFH cells (eosinophils, γδ T cells, and Th17 cells with weak supporting evidence) play an antitumor role. During the occurrence and development of PTC, the overall immune level increased, and the abundance and proportion of tumor-promoting immune cells significantly increased. We speculate that EBV may play an important role in changing the immune microenvironment of PTC.
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding author/s.
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
ZX designed the analytical strategies, performed data analyses, and wrote the manuscript. YL, XL, YZH, SW, SYW, JS, and YCH performed data analyses. JZ conceived the research and wrote the manuscript. All authors contributed to the article and approved the submitted version.
This research was supported by the National Natural Science Foundation of China (81600602).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to thank Mr. Guangqi Li for his support with bioinformatics analysis and pay tribute to the contributions of public databases such as the TCGA and GEO to human medicine. This manuscript has been released as a pre-print at [ResearchSquare] (89).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2020.570604/full#supplementary-material
Supplementary Figure 1 | Changes in immune cell infiltration (immune cell-specific markers) during the occurrence and development of PTC. (A) Comparison of immune cell abundance between PTC and normal tissues. (B) Comparison of immune cell abundance between stages I and II PTC patients and stages III and IV PTC patients. (C) Comparison of immune cell abundance between N0 and N1 PTC patients. (D) Comparison of immune cell abundance between T1 and T2 PTC patients and T3 and T4 PTC patients.
aDCs, activated dendritic cells; AJCC, American Joint Committee on Cancer; BL, Burkitt lymphoma; CAMs, cell adhesion molecules; CCR, chemokine receptor; CDF, cumulative distribution function; DCs, dendritic cells; DEGs, differentially expressed genes; DTC, differentiated thyroid carcinoma; EBV, Epstein-Barr virus; FDR, false discovery rate; GEO, Gene Expression Omnibus; GO, Gene Ontology; GPI, glycosylphosphatidylinositol; GSEA, gene set enrichment analysis; iDCs, immmature dendritic cells; KEGG, Kyoto Encyclopedia of Genes and Genomes; MCs, mast cells; mDCs, mature dendritic cells; MDSCs, myeloid-derived suppressor cells; MHC-I, major histocompatibility complex class I; NES, normalized enrichment score; NPC, nasopharyngeal carcinoma; PCA, principal component analysis; pDCs, plasmacytoid dendritic cells; PFS, progression-free survival; PPI, Protein-protein interaction; PTC, Papillary thyroid carcinoma; ssGSEA, single-sample gene set enrichment analysis; TAMs, macrophages; TC, Thyroid cancer; TCGA, The Cancer Genome Atlas; TFH cell, T follicular helper cell; THCA, thyroid carcinoma in The Cancer Genome Atlas; TME, tumor microenvironment; UCSC, University of California at Santa Cruz; WGCNA, Weighted gene coexpression network analysis.
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA Cancer J Clin (2020) 70:7–30. doi: 10.3322/caac.21590
2. Schneider DF, Chen H. New developments in the diagnosis and treatment of thyroid cancer. CA Cancer J Clin (2013) 63:374–94. doi: 10.3322/caac.21195
3. Li N, Du XL, Reitzel LR, Xu L, Sturgis EM. Impact of enhanced detection on the increase in thyroid cancer incidence in the United States: review of incidence trends by socioeconomic status within the surveillance, epidemiology, and end results registry, 1980-2008. Thyroid (2013) 23:103–10. doi: 10.1089/thy.2012.0392
4. Kitahara CM, Sosa JA. The changing incidence of thyroid cancer. Nat Rev Endocrinol (2016) 12:646–53. doi: 10.1038/nrendo.2016.110
5. Lim H, Devesa SS, Sosa JA, Check D, Kitahara CM. Trends in Thyroid Cancer Incidence and Mortality in the United States, 1974-2013. Jama (2017) 317:1338–48. doi: 10.1001/jama.2017.2719
6. Gupta S, Harper A, Ruan Y, Barr R, Frazier AL, Ferlay J, et al. International trends in the incidence of cancer among adolescents and young adults. J Natl Cancer Inst (2020), djaa007. doi: 10.1093/jnci/djaa007
7. Carling T, Udelsman R. Thyroid cancer. Annu Rev Med (2014) 65:125–37. doi: 10.1146/annurev-med-061512-105739
8. Ferrari SM, Fallahi P, Ruffilli I, Elia G, Ragusa F, Paparo SR, et al. Molecular testing in the diagnosis of differentiated thyroid carcinomas. Gland Surg (2018) 7:S19–29. doi: 10.21037/gs.2017.11.07
9. Fagin JA, Wells SA. Biologic and clinical perspectives on thyroid cancer. N Engl J Med (2016) 375:1054–67. doi: 10.1056/NEJMra1501993
10. Albero A, Lopez JE, Torres A, de la Cruz L, Martin T. Effectiveness of chemotherapy in advanced differentiated thyroid cancer: a systematic review. Endocr Relat Cancer (2016) 23:R71–84. doi: 10.1530/ERC-15-0194
11. Grogan RH, Kaplan SP, Cao H, Weiss RE, Degroot LJ, Simon CA, et al. A study of recurrence and death from papillary thyroid cancer with 27 years of median follow-up. Surgery (2013) 154:1436–1446; discussion 1446-1437. doi: 10.1016/j.surg.2013.07.008
12. Dunn GP, Bruce AT, Ikeda H, Old LJ, Schreiber RD. Cancer immunoediting: from immunosurveillance to tumor escape. Nat Immunol (2002) 3:991–8. doi: 10.1038/ni1102-991
13. Poschke I, Mougiakakos D, Kiessling R. Camouflage and sabotage: tumor escape from the immune system. Cancer Immunol Immunother (2011) 60:1161–71. doi: 10.1007/s00262-011-1012-8
14. Varricchi G, Loffredo S, Marone G, Modestino L, Fallahi P, Ferrari SM, et al. The immune landscape of thyroid cancer in the context of immune checkpoint inhibition. Int J Mol Sci (2019) 20:E3934. doi: 10.3390/ijms20163934
15. Galdiero MR, Varricchi G, Marone G. The immune network in thyroid cancer. Oncoimmunology (2016) 5:e1168556. doi: 10.1080/2162402x.2016.1168556
16. Ferrari SM, Fallahi P, Elia G, Ragusa F, Ruffilli I, Paparo SR, et al. Thyroid autoimmune disorders and cancer. Semin Cancer Biol (2019) 64:135–46. doi: 10.1016/j.semcancer.2019.05.019
17. Barbie DA, Tamayo P, Boehm JS, Kim SY, Moody SE, Dunn IF, et al. Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1. Nature (2009) 462:108–12. doi: 10.1038/nature08460
18. He Y, Jiang Z, Chen C, Wang X. Classification of triple-negative breast cancers based on Immunogenomic profiling. J Exp Clin Cancer Res (2018) 37:327. doi: 10.1186/s13046-018-1002-1
19. Bindea G, Mlecnik B, Tosolini M, Kirilovsky A, Waldner M, Obenauf AC, et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity (2013) 39:782–95. doi: 10.1016/j.immuni.2013.10.003
20. Newman AM, Liu CL, Green MR, Gentles AJ, Feng W, Xu Y, et al. Robust enumeration of cell subsets from tissue expression profiles. Nat Methods (2015) 12:453–7. doi: 10.1038/nmeth.3337
21. Zeng D, Li M, Zhou R, Zhang J, Sun H, Shi M, et al. Tumor microenvironment characterization in gastric cancer identifies prognostic and immunotherapeutically relevant gene signatures. Cancer Immunol Res (2019) 7:737–50. doi: 10.1158/2326-6066.CIR-18-0436
22. Wilkerson MD, Hayes DN. ConsensusClusterPlus: a class discovery tool with confidence assessments and item tracking. Bioinformatics (2010) 26:1572–3. doi: 10.1093/bioinformatics/btq170
23. Ringner M. What is principal component analysis? Nat Biotechnol (2008) 26:303–4. doi: 10.1038/nbt0308-303
24. Yoshihara K, Shahmoradgoli M, Martinez E, Vegesna R, Kim H, Torres-Garcia W, et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat Commun (2013) 4:2612. doi: 10.1038/ncomms3612
25. Langfelder P, Horvath S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinf (2008) 9:559. doi: 10.1186/1471-2105-9-559
26. Cunha LL, Morari EC, Guihen AC, Razolli D, Gerhard R, Nonogaki S, et al. Infiltration of a mixture of immune cells may be related to good prognosis in patients with differentiated thyroid carcinoma. Clin Endocrinol (Oxf) (2012) 77:918–25. doi: 10.1111/j.1365-2265.2012.04482.x
27. Na KJ, Choi H. Immune landscape of papillary thyroid cancer and immunotherapeutic implications. Endocr Relat Cancer (2018) 25:523–31. doi: 10.1530/ERC-17-0532
28. Jung KY, Cho SW, Kim YA, Kim D, Oh BC, Park DJ, et al. Cancers with higher density of tumor-associated macrophages were associated with poor survival rates. J Pathol Transl Med (2015) 49:318–24. doi: 10.4132/jptm.2015.06.01
29. Ryder M, Ghossein RA, Ricarte-Filho JC, Knauf JA, Fagin JA. Increased density of tumor-associated macrophages is associated with decreased survival in advanced thyroid cancer. Endocr Relat Cancer (2008) 15:1069–74. doi: 10.1677/ERC-08-0036
30. Qing W, Fang WY, Ye L, Shen LY, Zhang XF, Fei XC, et al. Density of tumor-associated macrophages correlates with lymph node metastasis in papillary thyroid carcinoma. Thyroid (2012) 22:905–10. doi: 10.1089/thy.2011.0452
31. Proietti A, Ugolini C, Melillo RM, Crisman G, Elisei R, Santoro M, et al. Higher intratumoral expression of CD1a, tryptase, and CD68 in a follicular variant of papillary thyroid carcinoma compared to adenomas: correlation with clinical and pathological parameters. Thyroid (2011) 21:1209–15. doi: 10.1089/thy.2011.0059
32. Fang W, Ye L, Shen L, Cai J, Huang F, Wei Q, et al. Tumor-associated macrophages promote the metastatic potential of thyroid papillary cancer by releasing CXCL8. Carcinogenesis (2014) 35:1780–7. doi: 10.1093/carcin/bgu060
33. Cho JW, Kim WW, Lee YM, Jeon MJ, Kim WG, Song DE, et al. Impact of tumor-associated macrophages and BRAF(V600E) mutation on clinical outcomes in patients with various thyroid cancers. Head Neck (2019) 41:686–91. doi: 10.1002/hed.25469
34. Chang WC, Chen JY, Lee CH, Yang AH. Expression of decoy receptor 3 in diffuse sclerosing variant of papillary thyroid carcinoma: correlation with M2 macrophage differentiation and lymphatic invasion. Thyroid (2013) 23:720–6. doi: 10.1089/thy.2012.0261
35. Cho SW, Kim YA, Sun HJ, Kim YA, Oh BC, Yi KH, et al. CXCL16 signaling mediated macrophage effects on tumor invasion of papillary thyroid carcinoma. Endocr Relat Cancer (2016) 23:113–24. doi: 10.1530/ERC-15-0196
36. Hilly O, Koren R, Raz R, Rath-Wolfson L, Mizrachi A, Hamzany Y, et al. The role of s100-positive dendritic cells in the prognosis of papillary thyroid carcinoma. Am J Clin Pathol (2013) 139:87–92. doi: 10.1309/AJCPAKYDO56NKMYZ
37. Scouten WT, Francis GL. Thyroid cancer and the immune system: a model for effective immune surveillance. Expert Rev Endocrinol Metab (2006) 1:353–66. doi: 10.1586/17446651.1.3.353
38. Yu H, Huang X, Liu X, Jin H, Zhang G, Zhang Q, et al. Regulatory T cells and plasmacytoid dendritic cells contribute to the immune escape of papillary thyroid cancer coexisting with multinodular non-toxic goiter. Endocrine (2013) 44:172–81. doi: 10.1007/s12020-012-9853-2
39. Hilly O, Rath-Wolfson L, Koren R, Mizrachi A, Hamzany Y, Bachar G, et al. CD1a-positive dendritic cell density predicts disease-free survival in papillary thyroid carcinoma. Pathol Res Pract (2015) 211:652–6. doi: 10.1016/j.prp.2015.05.009
40. Kuwabara K, Nishishita T, Morishita M, Oyaizu N, Yamashita S, Kanematsu T, et al. Results of a phase I clinical study using dendritic cell vaccinations for thyroid cancer. Thyroid (2007) 17:53–8. doi: 10.1089/thy.2006.0178
41. Melillo RM, Guarino V, Avilla E, Galdiero MR, Liotti F, Prevete N, et al. Mast cells have a protumorigenic role in human thyroid cancer. Oncogene (2010) 29:6203–15. doi: 10.1038/onc.2010.348
42. Visciano C, Liotti F, Prevete N, Cali G, Franco R, Collina F, et al. Mast cells induce epithelial-to-mesenchymal transition and stem cell features in human thyroid cancer cells through an IL-8-Akt-Slug pathway. Oncogene (2015) 34:5175–86. doi: 10.1038/onc`.2014.441
43. Ari A, Gunver F. Comparison of neutrophil-lymphocyte ratio and platelet-lymphocyte ratio in patients with thyroiditis and papillary tumors. J Int Med Res (2019) 47:2077–83. doi: 10.1177/0300060519838392
44. Seretis C, Gourgiotis S, Gemenetzis G, Seretis F, Lagoudianakis E, Dimitrakopoulos G. The significance of neutrophil/lymphocyte ratio as a possible marker of underlying papillary microcarcinomas in thyroidal goiters: a pilot study. Am J Surg (2013) 205:691–6. doi: 10.1016/j.amjsurg.2012.08.006
45. Galdiero MR, Varricchi G, Loffredo S, Bellevicine C, Lansione T, Ferrara AL, et al. Potential involvement of neutrophils in human thyroid cancer. PloS One (2018) 13:e0199740. doi: 10.1371/journal.pone.0199740
46. Wen W, Wu P, Li J, Wang H, Sun J, Chen H. Predictive values of the selected inflammatory index in elderly patients with papillary thyroid cancer. J Transl Med (2018) 16:261. doi: 10.1186/s12967-018-1636-y
47. Liu JF, Ba L, Lv H, Lv D, Du JT, Jing XM, et al. Association between neutrophil-to-lymphocyte ratio and differentiated thyroid cancer: a meta-analysis. Sci Rep (2016) 6:38551. doi: 10.1038/srep38551
48. Lang BH, Ng CP, Au KB, Wong KP, Wong KK, Wan KY. Does preoperative neutrophil lymphocyte ratio predict risk of recurrence and occult central nodal metastasis in papillary thyroid carcinoma? World J Surg (2014) 38:2605–12. doi: 10.1007/s00268-014-2630-z
49. Manatakis DK, Tseleni-Balafouta S, Balalis D, Soulou VN, Korkolis DP, Sakorafas GH, et al. Association of baseline neutrophil-to-lymphocyte ratio with clinicopathological characteristics of papillary thyroid carcinoma. Int J Endocrinol (2017) 2017:8471235. doi: 10.1155/2017/8471235
50. Kim SM, Kim EH, Kim BH, Kim JH, Park SB, Nam YJ, et al. Association of the preoperative neutrophil-to-ymphocyte count ratio and platelet-to-lymphocyte count ratio with clinicopathological characteristics in patients with papillary thyroid cancer. Endocrinol Metab (Seoul) (2015) 30:494–501. doi: 10.3803/EnM.2015.30.4.494
51. Liu CL, Lee JJ, Liu TP, Chang YC, Hsu YC, Cheng SP. Blood neutrophil-to-lymphocyte ratio correlates with tumor size in patients with differentiated thyroid cancer. J Surg Oncol (2013) 107:493–7. doi: 10.1002/jso.23270
52. Lee F, Yang PS, Chien MN, Lee JJ, Leung CH, Cheng SP. An increased neutrophil-to-lymphocyte ratio predicts incomplete response to therapy in differentiated thyroid cancer. Int J Med Sci (2018) 15:1757–63. doi: 10.7150/ijms.28498
53. Lee KH, Seok EY, Kim EY, Yun JS, Park YL, Park CH. Different prognostic values of individual hematologic parameters in papillary thyroid cancer due to age-related changes in immunity. Ann Surg Treat Res (2019) 96:70–7. doi: 10.4174/astr.2019.96.2.70
54. Ozmen S, Timur O, Calik I, Altinkaynak K, Simsek E, Gozcu H, et al. Neutrophil-lymphocyte ratio (NLR) and platelet-lymphocyte ratio (PLR) may be superior to C-reactive protein (CRP) for predicting the occurrence of differentiated thyroid cancer. Endocr Regul (2017) 51:131–6. doi: 10.1515/enr-2017-0013
55. Modi J, Patel A, Terrell R, Tuttle RM, Francis GL. Papillary thyroid carcinomas from young adults and children contain a mixture of lymphocytes. J Clin Endocrinol Metab (2003) 88:4418–25. doi: 10.1210/jc.2003-030342
56. Gogali F, Paterakis G, Rassidakis GZ, Liakou CI, Liapi C. CD3(-)CD16(-)CD56(bright) immunoregulatory NK cells are increased in the tumor microenvironment and inversely correlate with advanced stages in patients with papillary thyroid cancer. Thyroid (2013) 23:1561–8. doi: 10.1089/thy.2012.0560
57. Xu X, Rao G, Gaffud MJ, Ding HG, Maki G, Klingemann HG, et al. Clinicopathological significance of major histocompatibility complex class I-related chain a and B expression in thyroid cancer. J Clin Endocrinol Metab (2006) 91:2704–12. doi: 10.1210/jc.2006-0492
58. Gogali F, Paterakis G, Rassidakis GZ, Kaltsas G, Liakou CI, Gousis P, et al. Phenotypical analysis of lymphocytes with suppressive and regulatory properties (Tregs) and NK cells in the papillary carcinoma of thyroid. J Clin Endocrinol Metab (2012) 97:1474–82. doi: 10.1210/jc.2011-1838
59. Lee EK, Sunwoo JB. Natural killer cells and thyroid diseases. Endocrinol Metab (Seoul) (2019) 34:132–7. doi: 10.3803/EnM.2019.34.2.132
60. Parhar RS, Zou M, Al-Mohanna FA, Baitei EY, Assiri AM, Meyer BF, et al. IL-12 immunotherapy of Braf(V600E)-induced papillary thyroid cancer in a mouse model. Lab Invest (2016) 96:89–97. doi: 10.1038/labinvest.2015.126
61. French JD, Weber ZJ, Fretwell DL, Said S, Klopper JP, Haugen BR. Tumor-associated lymphocytes and increased FoxP3+ regulatory T cell frequency correlate with more aggressive papillary thyroid cancer. J Clin Endocrinol Metab (2010) 95:2325–33. doi: 10.1210/jc.2009-2564
62. Aghajani MJ, Yang T, McCafferty CE, Graham S, Wu X, Niles N. Predictive relevance of programmed cell death protein 1 and tumor-infiltrating lymphocyte expression in papillary thyroid cancer. Surgery (2018) 163:130–6. doi: 10.1016/j.surg.2017.04.033
63. Pilli T, Toti P, Occhini R, Castagna MG, Cantara S, Caselli M, et al. Chronic lymphocytic thyroiditis (CLT) has a positive prognostic value in papillary thyroid cancer (PTC) patients: the potential key role of Foxp3+ T lymphocytes. J Endocrinol Invest (2018) 41:703–9. doi: 10.1007/s40618-017-0794-8
64. Cunha LL, Marcello MA, Nonogaki S, Morari EC, Soares FA, Vassallo J, et al. CD8+ tumour-infiltrating lymphocytes and COX2 expression may predict relapse in differentiated thyroid cancer. Clin Endocrinol (Oxf) (2015) 83:246–53. doi: 10.1111/cen.12586
65. Matsubayashi S, Kawai K, Matsumoto Y, Mukuta T, Morita T, Hirai K, et al. The correlation between papillary thyroid carcinoma and lymphocytic infiltration in the thyroid gland. J Clin Endocrinol Metab (1995) 80:3421–4. doi: 10.1210/jcem.80.12.8530576
66. Kuo C-Y, Liu T-P, Yang P-S, Cheng S-P. Characteristics of lymphocyte-infiltrating papillary thyroid cancer. J Cancer Res Pract (2017) 4:95–9. doi: 10.1016/j.jcrpr.2017.03.003
67. Shi J, Zhou J. Role of CD4+ CD25+ regulatory T cells in peripheral blood from patients with papillary thyroid carcinoma. Lin Chung Er Bi Yan Hou Tou Jing Wai Ke Za Zhi (2012) 26:965–9.
68. French JD, Kotnis GR, Said S, Raeburn CD, McIntyre RC, Klopper JP, et al. Programmed death-1+ T cells and regulatory T cells are enriched in tumor-involved lymph nodes and associated with aggressive features in papillary thyroid cancer. J Clin Endocrinol Metab (2012) 97:E934–43. doi: 10.1210/jc.2011-3428
69. Ugolini C, Elisei R, Proietti A, Pelliccioni S, Lupi C, Borrelli N, et al. FoxP3 expression in papillary thyroid carcinoma: a possible resistance biomarker to iodine 131 treatment. Thyroid (2014) 24:339–46. doi: 10.1089/thy.2012.0589
70. Jiang G, Ma S, Wei Y, Wu Y, Yu X, Liu H. The prevalence and distribution of Th17 and Tc17 cells in patients with thyroid tumor. Immunol Lett (2014) 162:68–73. doi: 10.1016/j.imlet.2014.07.005
71. Thorsson V, Gibbs DL, Brown SD, Wolf D, Bortone DS, Ou Yang TH, et al. The immune landscape of cancer. Immunity (2018) 48:812–830.e814. doi: 10.1016/j.immuni.2018.03.023
72. Ferrari SM, Fallahi P, Elia G, Ragusa F, Ruffilli I, Paparo SR, et al. Thyroid autoimmune disorders and cancer. Semin Cancer Biol (2020) 64:135–46. doi: 10.1016/j.semcancer.2019.05.019
73. Naoum GE, Morkos M, Kim B, Arafat W. Novel targeted therapies and immunotherapy for advanced thyroid cancers. Mol Cancer (2018) 17:51. doi: 10.1186/s12943-018-0786-0
74. Park S, Zhu J, Altan-Bonnet G, Cheng SY. Monocyte recruitment and activated inflammation are associated with thyroid carcinogenesis in a mouse model. Am J Cancer Res (2019) 9:1439–53.
75. Ma Y, Aymeric L, Locher C, Mattarollo SR, Delahaye NF, Pereira P, et al. Contribution of IL-17-producing gamma delta T cells to the efficacy of anticancer chemotherapy. J Exp Med (2011) 208:491–503. doi: 10.1084/jem.20100269
76. Vinuesa CG, Linterman MA, Yu D, MacLennan IC. Follicular helper T cells. Annu Rev Immunol (2016) 34:335–68. doi: 10.1146/annurev-immunol-041015-055605
77. Zhu C, Ma J, Liu Y, Tong J, Tian J, Chen J, et al. Increased frequency of follicular helper T cells in patients with autoimmune thyroid disease. J Clin Endocrinol Metab (2012) 97:943–50. doi: 10.1210/jc.2011-2003
78. Tsuge K, Takeda H, Kawada S, Maeda K, Yamakawa M. Characterization of dendritic cells in differentiated thyroid cancer. J Pathol (2005) 205:565–76. doi: 10.1002/path.1731
79. Ryu HS, Park YS, Park HJ, Chung YR, Yom CK, Ahn SH, et al. Expression of indoleamine 2,3-dioxygenase and infiltration of FOXP3+ regulatory T cells are associated with aggressive features of papillary thyroid microcarcinoma. Thyroid (2014) 24:1232–40. doi: 10.1089/thy.2013.0423
80. Smallridge RC, Chindris AM, Asmann YW, Casler JD, Serie DJ, Reddi HV, et al. RNA sequencing identifies multiple fusion transcripts, differentially expressed genes, and reduced expression of immune function genes in BRAF (V600E) mutant vs BRAF wild-type papillary thyroid carcinoma. J Clin Endocrinol Metab (2014) 99:E338–347. doi: 10.1210/jc.2013-2792
81. Kim K, Jeon S, Kim TM, Jung CK. Immune gene signature delineates a subclass of papillary thyroid cancer with unfavorable clinical outcomes. Cancers (Basel) (2018) 10:494. doi: 10.3390/cancers10120494
82. Homayouni M, Arabzadeh SAM, Nili F, Razi F, Amoli MM. Evaluation of the presence of Epstein-Barr virus (EBV) in Iranian patients with thyroid papillary carcinoma. Pathol Res Pract (2017) 213:854–6. doi: 10.1016/j.prp.2017.01.020
83. Almeida JFM, Campos AH, Marcello MA, Bufalo NE, Rossi CL, Amaral LHP, et al. Investigation on the association between thyroid tumorigeneses and herpesviruses. J Endocrinol Invest (2017) 40:823–9. doi: 10.1007/s40618-017-0609-y
84. Almeida JFM, Proenca-Modena JL, Bufalo NE, Peres KC, Teixeira EDS, Teodoro L, et al. Epstein-Barr virus induces morphological and molecular changes in thyroid neoplastic cells. Endocrine (2020) 69(2):321–30. doi: 10.1007/s12020-020-02253-0
85. Moghoofei M, Mostafaei S, Nesaei A, Etemadi A, Sadri Nahand J, Mirzaei H, et al. Epstein-Barr virus and thyroid cancer: the role of viral expressed proteins. J Cell Physiol (2019) 234:3790–9. doi: 10.1002/jcp.27144
86. Naseem M, Barzi A, Brezden-Masley C, Puccini A, Berger MD, Tokunaga R, et al. Outlooks on Epstein-Barr virus associated gastric cancer. Cancer Treat Rev (2018) 66:15–22. doi: 10.1016/j.ctrv.2018.03.006
87. Lieberman PM. Virology. Epstein-Barr virus turns 50. Science (2014) 343:1323–5. doi: 10.1126/science.1252786
88. Thorley-Lawson DA. Epstein-Barr virus: exploiting the immune system. Nat Rev Immunol (2001) 1:75–82. doi: 10.1038/35095584
Keywords: papillary thyroid carcinoma, immune cell infiltration, tumor microenvironment, immune escape, TCGA, CIBERSORT
Citation: Xie Z, Li X, He Y, Wu S, Wang S, Sun J, He Y, Lun Y and Zhang J (2020) Immune Cell Confrontation in the Papillary Thyroid Carcinoma Microenvironment. Front. Endocrinol. 11:570604. doi: 10.3389/fendo.2020.570604
Received: 04 August 2020; Accepted: 02 October 2020;
Published: 22 October 2020.
Edited by:
Yuji Nagayama, Nagasaki University, JapanReviewed by:
Alessandro Antonelli, University of Pisa, ItalyCopyright © 2020 Xie, Li, He, Wu, Wang, Sun, He, Lun and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jian Zhang, amlhbnpoYW5nQGNtdS5lZHUuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.