 Pablo Vidal
Pablo Vidal Kristin I. Stanford
Kristin I. Stanford
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Endocrinol., 29 April 2020
Sec. Obesity
Volume 11 - 2020 | https://doi.org/10.3389/fendo.2020.00270
This article is part of the Research TopicCurrent Challenges for Targeting Brown Fat Thermogenesis to Combat ObesityView all 16 articles
Exercise training results in beneficial adaptations to numerous tissues and offers protection against metabolic disorders including obesity and type 2 diabetes. Multiple studies have indicated that both white (WAT) and brown (BAT) adipose tissue may play an important role to mediate the beneficial effects of exercise. Studies from both rodents and humans have identified exercise-induced changes in WAT including increased mitochondrial activity and glucose uptake, an altered endocrine profile, and in rodents, a beiging of the WAT. Studies investigating the effects of exercise on BAT have resulted in conflicting data in terms of mitochondrial activity, glucose uptake, and thermogenic activity in rodents and humans, and remain an important area of investigation. This review discusses the exercise-induced adaptations to white and brown adipose tissue, distinguishing important differences between rodents and humans and highlighting the latest studies in the field and their implications.
Exercise training is an important non-pharmacological strategy to prevent and treat metabolic diseases, including obesity and type 2 diabetes. Exercise results in adaptations to almost all tissues in the body that contribute to the beneficial effects of exercise to improve whole-body metabolic health. A single bout of moderate intensity exercise has dramatic effects on glucose metabolism, lowering circulating insulin concentrations and improving skeletal muscle insulin sensitivity (1). Exercise training, defined as repeated bouts of exercise over a period of weeks, months, or years can decrease insulin concentrations and improve glucose tolerance (1, 2).
While it is well-established that exercise induces adaptations to skeletal muscle (2) and the cardiovascular system (3), several studies have now determined that exercise also results in adaptations to adipose tissue that improve whole-body metabolic health (4–15). These exercise-induced adaptations to adipose tissue include increased mitochondrial activity (5, 10), decreased cell size and lipid content (11), reduced inflammation (12, 13), and, in rodents, increased presence of thermogenic brown-like adipocytes or “beige” cells (6, 10, 15). Exercise also alters the endocrine profile of adipose tissue, inducing the release of adipokines and lipokines that mediate tissue-to-tissue communication and contribute to the improved metabolic homeostasis seen with exercise (9, 14, 16). Here, we will discuss studies investigating the exercise-induced adaptations to white and brown adipose tissue in humans and rodents, with a particular focus on the adaptations that contribute to thermogenesis.
Adipose tissue is a type of connective tissue consisting primarily of mature adipocytes (~65–90% in volume) (17, 18), a cell type whose defining characteristic is accumulation of internal fat droplets (19). In addition to the mature adipocytes, adipose tissue consists of a stromal vascular fraction (SVF). The SVF is immensely heterogeneous, containing pre-adipocytes, mesenchymal stem cells, endothelial cells, and a variety of immune cells, including macrophages and natural killer T cells (20). The SVF is very dynamic and can respond and adapt to stimulus such as β-adrenergic stimulation (20) and exercise (8).
Adipose tissue can be broadly classified into two different types, white adipose tissue and brown adipose tissue (21). Certain stimuli such as cold, sympathetic activation (22), exercise (6, 23) or an enriched environment (24) can give rise to a third type of adipocytes, beige adipocytes, within the WAT.
White adipose tissue (WAT) is composed of white adipocytes and its primary function is energy storage. Energy is stored by mature adipocytes in the form of triglycerides as one unilocular lipid droplet which occupies most of the cell volume and can vary in size (25). Adipose tissue is very dynamic, it can expand in size via hyperplasia or hypertrophy of the adipocytes (26, 27). WAT can be further subdivided into two different depots with distinct functions based on anatomical location, subcutaneous and visceral WAT (28).
Subcutaneous WAT (scWAT) is located beneath the skin. In mice, scWAT is located in the inguinal, anterior axillary and interscapular regions (28–30). In humans, scWAT locations can be divided into lower-body, comprising gluteal and leg depots, and upper-body, in the anterior abdominal wall region (28). These distinct locations of scWAT adapt differently to the same stimulus (26, 31). Under obesogenic conditions, lower-body adipocytes tend to expand via hyperplasia, which has been associated with improved metabolic adaptations (32), while upper-body adipocytes expand via hypertrophy (26). Increases in upper-body scWAT are correlated with decreased insulin sensitivity and impaired glucose tolerance (31).
Visceral WAT (vWAT) surrounds internal organs. In mice, vWAT is found in the perigonadal, mesenteric, perirenal, retroperitoneal, cardiac, and triceps-associated regions (8, 28–30). In humans, vWAT is located in the intraabdominal (omental and mesenteric) as well as in the cardiac region (28). In lean individuals, vWAT accounts for 10–20% of the total fat mass in males and 5–8% in females (33).
There are distinct differences between scWAT and vWAT. These two adipose tissue depots behave and adapt differently to the same stimuli (26, 28, 34). Adipocytes in scWAT are smaller, have higher avidity for free fatty acid and triglyceride uptake, and are more sensitive to insulin compared to adipocytes from the vWAT (33, 35). Subcutaneous WAT has elevated expression of genes involved in glucose and lipid metabolism, and insulin signaling, compared to vWAT (36). Conversely, increases in vWAT are correlated with impaired glucose tolerance and increased insulin resistance (31) while increases in scWAT are correlated with improved metabolism (37).
Brown adipose tissue (BAT) is a metabolically active tissue that burns carbohydrates and lipids to generate heat (38–40). Brown adipocytes are characterized by multilocular lipid droplets, a central nucleus and a high density of mitochondria (41, 42). The most distinctive feature of brown adipocytes is the high expression of the thermogenic protein uncoupling protein 1 (UCP1) (43). UCP1 is located in the inner membrane of mitochondria and uncouples the proton gradient potential generated by the electron transport chain. Release of this chemical gradient results in the dissipation of energy in the form of heat. In rodents, BAT is found in the interscapular, mediastinal, perirenal, axillary, and cervical regions (29, 30, 44). BAT is a mammal-specific tissue and in humans, it was long thought to be present only in infants. In 2009, multiple studies demonstrated that BAT is also present in adult individuals (45–48). In humans, BAT is found in the cervical, supraclavicular, axillary, and paravertebral regions (45, 49), as well as in the perirenal region in infants (50). Perirenal BAT consists mainly of dormant brown adipocytes that can be stimulated to give rise to active brown adipocytes (51). Brown adipose tissue mass is negatively correlated with BMI and age in humans (45). Given this, and the functional role of BAT, targeting BAT as a therapeutic to combat obesity and metabolic disorders has become increasingly important.
Beige or brite (brown in white) adipocytes are a particular type of adipocytes within scWAT. Over 100 different stimuli are known to induce beiging, and most of them act through activation of the sympathetic nervous system (SNS) (52). Beige adipocytes have multilocular lipid droplets, a central nucleus, and a high density of mitochondria, similar to brown adipocytes. However, while brown adipocytes arise from Pax7 and Myf5 positive cells (53, 54), beige adipocytes arise from Myf5 negative adipogenic stem cells within the adipose tissue (55, 56). White adipocyte tissue that has undergone beiging can be distinguished by the specific beiging markers CD137, TBX1, and TMEM26 (30). Beige adipocytes function similarly to brown adipocytes in that they directly generate energy in the form of heat, contributing to thermogenesis. Beige adipocytes deviate from brown adipocytes in that they have a high degree of plasticity. In the absence of beiging stimuli, UCP1 expression, and mitochondrial content of beige adipocytes decrease and beige adipocytes transition to a white adipocyte phenotype (49). Increasing beige adipocytes has significant potential to combat obesity and type 2 diabetes.
Exercise is an important therapeutic to prevent and treat metabolic diseases, including obesity and type 2 diabetes. Exercise results in adaptations to almost all tissues in the body, including adipose tissue. Exercise increases whole-body energy expenditure as chemical energy is converted into kinetic energy. During acute exercise, WAT has an important role in supplying this additional energy requirement from the triglyceride stores within the mature adipocytes (57). Independent from its role during acute exercise, chronic exercise leads to several metabolic adaptations in WAT (Figure 1). In this section, we will be reviewing the different metabolic adaptations that occur in WAT with exercise in both rodents and humans, including thermogenesis, mitochondrial adaptations, glucose metabolism, lipid metabolism, and endocrine adaptations.
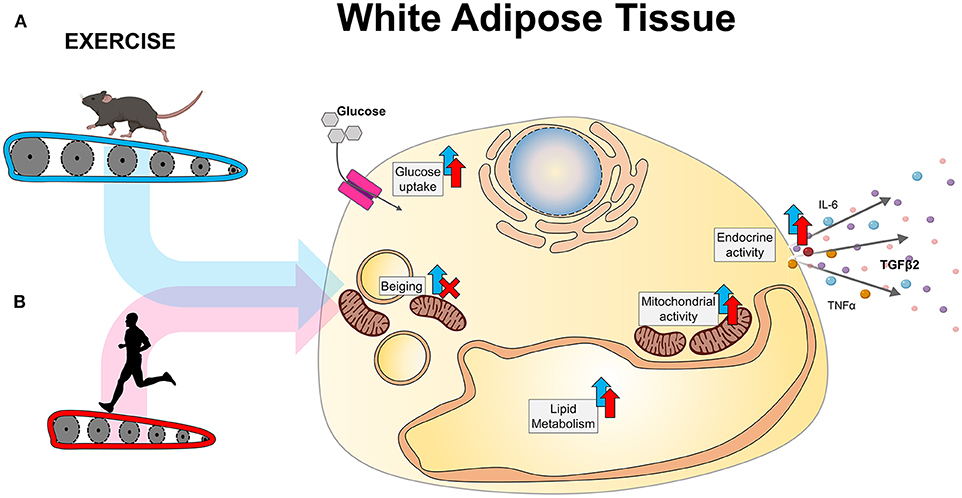
Figure 1. Exercise-induced adaptations to WAT in (A) rodents and (B) humans.
An important exercise-induced adaptation to scWAT in rodents is the beiging of scWAT. Exercise induces an upregulation of thermogenic genes such as Prdm16 and Ucp1 in inguinal scWAT (6, 15, 58, 59) and an increased presence of adipocytes with multilocular lipid droplets (6, 60). The appearance of beige adipocytes does not occur homogeneously, as some regions of the inguinal scWAT are more prone to beiging than others (58, 61). This exercise-induced beiging is specific to scWAT, in particular the inguinal scWAT (8), and does not occur in vWAT (23, 60, 62). Beiging of scWAT is the molecular mechanism that leads to increased thermogenesis in WAT with exercise, as beige adipocytes increase non-shivering thermogenesis.
While beiging is an important adaptation to exercise, it is unclear why exercise induces a beiging of scWAT. Beiging of scWAT by non-exercise stimuli, including through cold-exposure, environmental factors or pharmaceuticals, is thought to be induced through a heat compensatory mechanism in which adrenergic stimulation compensates for heat loss with the upregulation of UCP1 (44, 63–65). This explanation does not make sense in the context of exercise-induced beiging, because exercise itself increases heat production (66, 67). Several hypotheses have been proposed as the underlying mechanism, one of which is an increase in sympathetic innervation, which occurs in scWAT during exercise (52, 68) Other hypotheses have indicated that beiging occurs in response to the exercise-induced release of myokines, such as irisin (23), myostatin (69), meteorin-like 1 (Metrnl) (70), lactate (71), and β-aminoisobutyric acid (BAIBA) (72), or other secreted factors released during exercise, including brain-derived neurotrophic factor (BDNF) (24). More investigation is needed to fully understand this complex mechanism. These hypotheses are all important and plausible, but the most likely explanation is that the exercise-induced beiging of scWAT occurs because exercise decreases the adipocyte size and lipid content in scWAT, decreasing insulation of the body and necessitating heat production, which results in the beiging of scWAT (52, 73). The fact that mice are commonly housed at 20–22°C, the habitual indoor temperatures for humans, which itself contributes to mice being under chronic cold stress (74), provides further support for this explanation.
To address the hypothesis that beiging occurs in response to a loss of fat mass in a cold stress environment, multiple studies have investigated the effects of exercise at thermoneutrality (30°C) (14, 75, 76). Interestingly, when mice are housed at thermoneutral conditions, the exercise-induced increase of thermogenic gene expression and appearance of multilocular adipocytes exercise is blunted in male and female mice (75, 76), and this occurred independent of changes in body mass, fat mass, or running distance. Interestingly, one of these studies investigated female mice and determined that total running distance was lower at thermoneutrality (~40%) (76) and observed no differences in body weight or adiposity compared to sedentary mice, while another study determined that running distance was increased at thermoneutrality in male mice (~50%) compared to mice at room temperature (75). These mice also had lower body mass compared to sedentary mice and mice housed at room temperature. While these discrepancies make some of the nuances between these studies difficult to interpret, each study determined that exercise-induced increase in thermogenic genes was blunted at thermoneutrality. These data suggest that the exercise-induced beiging is not a direct consequence of exercise, it is indirectly induced through other stimuli such as increased cold stress due to loss of WAT mass.
Several human studies have determined that exercise in humans does not induce beiging of scWAT (77–80). In lean or obese individuals, 10–16 weeks of endurance training did not change the expression of thermogenic genes including UCP1, PRDM16, and PGC1A in scWAT in males and females (77, 80–82). Studies conducted in highly exercise-trained populations and individuals with a more active lifestyle have also not observed any differences in UCP1 expression in scWAT compared to sedentary controls (83, 84). These results collectively indicate that exercise does not induce beiging in humans.
The mechanistic reason as to why rodents and humans have opposite thermogenic adaptations in WAT is currently unknown. Similar to what has been discussed earlier, it is likely a result of cold stress; since rodents are smaller, they have a higher surface to volume ratio that makes them more susceptible to cold stress. Exercise decreases WAT accumulation, increasing cold stress, and thermogenic adaptations are increased to counter this effect. This would not be the case in humans, so the loss of WAT may not induce the same thermogenic response. However, most human studies investigating the effects of exercise on WAT have been conducted indoors in controlled environments. Investigating human subjects who exercise in the cold (i.e., skiers, open water swimmers) might result in a thermogenic response to human WAT.
Exercise increases mitochondrial activity and density in scWAT and vWAT in rodents (5–8, 10, 58, 60, 85–87). Eleven days of voluntary wheel cage running increases the oxygen consumption rate of scWAT (6) and upregulates mitochondrial genes in both scWAT (6, 86) and vWAT (7, 8, 10, 58, 85). Importantly, exercise at thermoneutrality also results in upregulation of electron transport chain proteins (76), indicating that the increase in mitochondrial activity after exercise is independent of the beiging of WAT. In vitro studies indicate that exercise increases basal oxygen consumption rate of adipocytes differentiated from the SVF of scWAT (inguinal) or vWAT (perigonadal) of exercised mice (8), however maximal respiratory capacity only increased in adipocytes derived from scWAT (8). These data indicate that mitochondrial adaptations with exercise occur in both scWAT and vWAT in rodents, independent of beiging.
Exercise induces mitochondrial adaptations in human scWAT in lean male subjects (83, 88, 89) or young obese female subjects (77). Six weeks of high-intensity interval training (HIIT) increased mitochondrial respiration of scWAT (88). Ten to eighteen sessions of alternating continuous moderate-intensity training and HIIT did not change expression of genes involved in oxidative phosphorylation such as PGC1A or COXIV (78, 83, 90), but long term aerobic exercise-training increased expression of several genes involved in oxidative phosphorylation (89) and mitochondrial biogenesis (83). Exercise induced mitochondrial adaptations in vWAT have not been investigated in humans. Together these data indicate that exercise or increased physical activity increases mitochondrial activity in mouse and human WAT.
Exercise improves whole-body glucose homeostasis in rodents (91) and humans (1). Exercise increases glucose uptake and insulin sensitivity of scWAT (6, 15) and induces upregulation of genes and proteins involved in glucose metabolism in scWAT and vWAT (7, 8). These data indicate that exercise improves glucose metabolism in WAT in rodents. Here, we will focus on the effects of exercise in glucose homeostasis in WAT.
Recent studies have investigated the effects of exercise at thermoneutrality on glucose metabolism, with conflicting results. One study found that exercise still resulted in improvements in whole-body glucose tolerance (75), whereas another found no effect of exercise on whole-body glucose homeostasis at thermoneutrality (76). Interestingly, the latter found that there was an increase in in vivo insulin-stimulated 3H-2DG uptake in vWAT at thermoneutrality, but no changes were found in scWAT (76). In the latter study, the lack of exercise-induced changes to glucose metabolism can likely be attributed to the fact that mice at thermoneutrality ran ~40% less than mice at room temperature (76). As the results from these two studies are conflicting, the effects of exercise on glucose metabolism at thermoneutrality are unclear. Further research is essential to elucidate the effects of exercise at thermoneutrality on glucose metabolism and determine which adaptations arise at a systemic level and which are specific to the WAT.
Studies investigating exercise-induced adaptations to glucose homeostasis in human WAT are less comprehensive. One study determined that 6 months of exercise upregulated genes involved in glucose metabolism in lower-body scWAT (89). Two weeks of exercise increased insulin-stimulated glucose uptake in lower-body scWAT, but not upper-body scWAT or vWAT (92). These data indicate that scWAT and vWAT, and even upper-body and lower-body scWAT, have distinct adaptations to glucose metabolism with exercise. This is of particular interest to human physiology as humans with a higher proportion of upper-body WAT have been correlated with impaired glucose tolerance, while humans with a higher proportion of lower-body WAT are associated with improved glucose levels (32). These data indicate the lower scWAT has a prominent role on the effect on whole-body glucose homeostasis and is more susceptible to exercise.
Exercise effects lipid metabolism in WAT during exercise. Moderate exercise (40–65% VO2 max) acutely increases whole-body lipolysis two to three times over basal rates after exercising for 30 min, and increases lipolysis up to 5-fold over basal after 4 h of exercise (93). Here, we will focus on the chronic adaptations of exercise to WAT with regard to lipid metabolism.
In rodents, exercise induces several adaptations that affect lipid metabolism including changes in gene expression (6, 8, 94), post-translational modifications (7) and an altered lipidomic profile (94). Two to three weeks of voluntary wheel cage running upregulates genes involved in fatty acid oxidation in scWAT and vWAT (6, 8), and genes involved in phospholipid metabolism in scWAT (94). Twelve days of voluntary wheel cage exercise increases phosphorylation of hormone sensitive lipase (HSL) (86), and exercise over a longer duration (6 weeks) increases phosphorylation of adipose triglyceride lipase (ATGL) (7). These post-translational modifications result in increased lipolytic activity of ATGL and HSL (95–97). Another study demonstrated that chronic treadmill training (8 weeks) did not increase the rate of lipolysis in isolated adipocytes under basal conditions, but when these adipocytes were stimulated by a β-adrenergic agonist, lipolysis was significantly increased in adipocytes isolated from exercised mice compared to adipocytes isolated from sedentary mice (98). Together, these results suggest that exercise induces adaptations that increase lipolysis.
Exercise also induces extensive adaptations to the lipidomic profile of scWAT in rodents. Previous work in our laboratory demonstrated that 3 weeks of exercise dramatically alters the lipidome of scWAT. Exercise significantly decreased the overall abundance of triacylglycerol (TAG), phosphatidylserines (PS) lysophosphatidylglycerols and lysophosphatidylinositols (LPI) (94). In addition to the changes in overall lipid classes, there were also decreases in several specific molecular species of phosphatidic acid, phosphatidylethanolamines (PE), and PS. These changes corresponded with a significant upregulation of several genes involved in phospholipid metabolism. These data suggest molecular species-specific remodeling of phospholipids and TAGs in scWAT in response to exercise (66, 94). The functional consequence of the exercise-induced changes to the lipidome of scWAT have not been identified, but that will be the focus of future investigation.
Research on the effects of chronic exercise on lipid metabolism in humans has not been thoroughly investigated. Studies have shown that active individuals (self-reported exercise >3x per week) have increased levels of CPT1B, the rate-limiting enzyme in fatty acid oxidation, in scWAT compared to sedentary individuals (83), and 6 months of exercise upregulates several genes involved in lipid metabolism (89). These data indicate that long-term exercise increases fatty acid oxidation in human WAT. However, shorter duration exercise interventions do not alter adaptations to lipid metabolism in WAT (82, 83). Three weeks of exercise in sedentary individuals did not change CPT1B levels (83), and 12 weeks of exercise in obese subjects did not change expression levels of ATGL, HSL, or other lipolytic enzymes (82). Taken together, these data indicate that exercise upregulates lipid metabolism in WAT of both rodents and humans.
Exercise induces considerable adaptations to the secretory profile of several tissues, including adipose tissue (13, 99). Secretory factors released from adipose tissue have been labeled as adipokines. Four or more weeks of exercise in rodents decreases leptin and adiponectin mRNA levels in scWAT (100) and circulation (87, 100, 101) in rodents and humans. Exercise also increases expression of other factors such as TNF-α and IL-6 in both WAT depots and in circulation (85, 100).
Recent work in our laboratory determined that transplantation of scWAT from exercised donor mice into sedentary recipient mice resulted in improved whole-body glucose tolerance. Glucose uptake was also increased in BAT, soleus and tibialis anterior, indicating that an endocrine factor is released from exercise-trained scWAT to mediate these effects (6). TGF-β2 was recently identified as the adipokine responsible for these beneficial effects on glucose metabolism (14). TGF-β2 is an adipokine secreted in response to exercise in both rodents and humans from WAT. In rodents, acute treatment with TGF-β2 increased glucose uptake in soleus, heart and BAT, and increased fatty acid uptake in skeletal muscle. Notably, adipose tissue specific TGF-β2 knockout mice did not have exercise-induced improvements in systemic glucose uptake (14).
Exercise can also induce adaptations in WAT through myokines such as myostatin and BAIBA. Myostatin is a well-known factor that inhibits skeletal muscle growth (102). Exercise decreases myostatin levels in skeletal muscle and serum (103). Reduced levels of myostatin promote beiging of the scWAT in rodents (104) and are correlated with improved insulin sensitivity in humans (103). During exercise, increase in PGC1α triggers the secretion of β-aminoisobutyric acid (BAIBA) in both rodents and humans. BAIBA promotes beiging of scWAT in rodents while it is inversely correlated with serum glucose and insulin levels in humans (72). These data indicate that exercise stimulates release of secretory factors, from WAT as well as other tissues like skeletal muscle, that result in positive metabolic systemic and WAT specific adaptations.
Exercise can be broadly divided into endurance (aerobic) and resistance (strength) training (2). There have been several studies investigating the different adaptations of endurance and resistance training in skeletal muscle (2, 105), but this is not the case with adipose tissue. Most studies have investigated the effects of endurance training on adipose tissue, using treadmill or voluntary wheel cage running in rodents, and running or cycling for human studies. Some studies have compared the effects of different intensities, moderate (MIT) or high-intensity (HIT) endurance training on adipose tissue and found that MIT and HIT had similar effects on WAT in rodents (106, 107) and humans (92, 108). Meta-analysis comparing the effect of MIT or HIT on adiposity in humans found HIT resulted in a greater decrease in total fat mass (109). A few human studies have mixed endurance and resistance training in their exercise protocols, without finding any striking differences when compared to just endurance training (14, 77, 79, 82). However, to our knowledge, the direct effect of resistance compared to endurance exercise in adipose tissue has not been investigated.
BAT accounts for a small percentage of total fat mass than WAT, but it is a much more metabolically active tissue than WAT (110). Exercise increases energy expenditure, thus indirectly increasing in thermogenesis (111). BAT and WAT functions are different, and so are their exercise-induced adaptations. Here, we will discuss the different metabolic adaptations that occur in BAT with exercise in both rodents and humans (Figure 2).
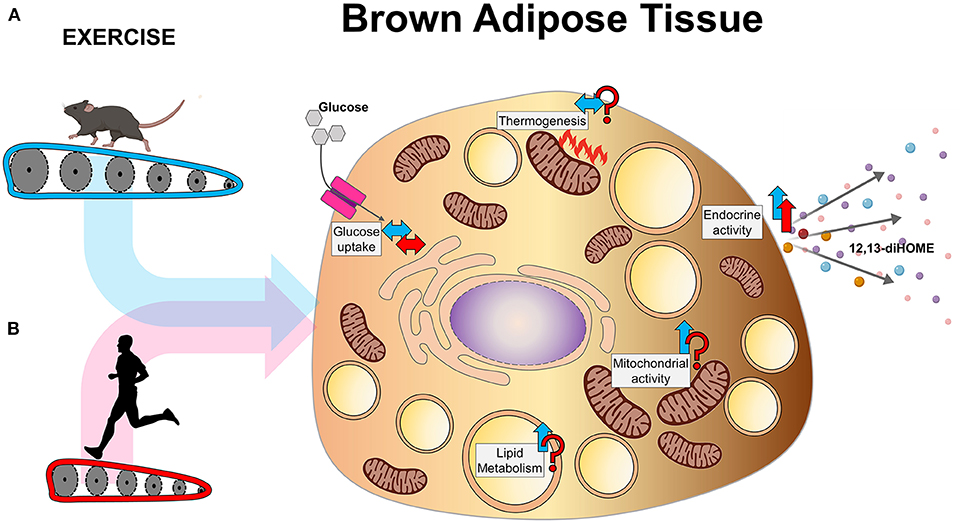
Figure 2. Exercise-induced adaptations to BAT in (A) rodents and (B) humans.
The thermogenic effects of exercise on BAT in rodents have been thoroughly investigated, with conflicting results. Eleven weeks of swimming (6 days/week; 2 h per day) increased blood flow and oxygen consumption in response to acute injection with norepinephrine (NE) (112, 113), indicating that exercise may increase sensitivity to adrenergic stimulation in BAT. These data are difficult to interpret because swimming as an exercise modality indirectly results in cold stress. Interestingly, these studies found that when the water temperature is 32, 36, or 38°C, acute injection of NE had the same response to increase blood flow and oxygen consumption, but BAT mass was only increased when the water temperature was 32°C (112). Other studies investigated the effects of exercise on BAT using 6 weeks of treadmill training as the exercise protocol (114). Interestingly, there was no effect of treadmill exercise to affect oxygen consumption or blood flow at rest or after NE injection (114, 115). Furthermore, BAT mass and protein content were decreased with 6 weeks of treadmill training (115, 116), regardless of the ambient temperature of the exercise (room temperature or 4°C) (116). In female rats, 6 weeks of treadmill exercise increased BAT mass and total protein content (117), but 9 weeks of treadmill training reduced BAT mass and decreased UCP1 expression (118). The reason for this is unclear, but it is possible that the discrepancies between these two studies could be explained by differences in the rat strain studied, as the first study used Sprague-Dawley while the latter used F-344 NNia. These data indicate that different exercise modalities, or different animal strains, could result in different adaptations to BAT.
More recent studies have indicated that exercise does not affect, or even decreases, BAT activity (58, 86, 119). Twelve days of voluntary wheel cage running in mice did not alter BAT mass (86), and 6 weeks of treadmill training in rats did not affect BAT mass, brown adipocyte size or Ucp1 expression (58, 119). Oxidation of palmitate was also reduced in BAT ex vivo after 6 weeks of treadmill training, indicating exercise decreases fatty acid oxidation in BAT (58). Exercise at thermoneutrality also reduced BAT mass and did not alter markers of thermogenesis (75). These data indicate that exercise does not increase thermogenic activity in BAT in rodents in the absence of a cold stress (i.e., swimming).
There is currently a paucity of data that has investigated the thermogenic adaptations of BAT with exercise in humans. Studies have determined that endurance trained athletes subjected to cold exposure have decreased glucose uptake in BAT compared to sedentary subjects (84, 120). It is important to note that the current gold standard to measure BAT activity in humans is 18FDG-PET/CT (121), and humans studies have only determined BAT mass and activity in the context of its ability to take up glucose. Moreover, cold exposure is frequently needed to activate BAT for detection by 18FDG-PET/CT scans. Other methods like infrared thermography (122) and T2 mapping (123) have been developed to evaluate BAT presence, but they have not yet been used to assess differences in BAT activity with exercise. Fat T2 relaxation time mapping is based on BAT having higher water content than WAT. This technique measures BAT activity and does not require cold exposure for detection (123). The use of these new techniques will be important to truly ascertain the effects of exercise on BAT in humans in vivo.
The effects of exercise on mitochondrial activity in BAT have also been investigated. In rodents, 2–8 weeks of exercise did not change or decreased expression of mitochondrial genes (8, 58, 75, 76). Recent work in our laboratory determined that 11 days of voluntary wheel cage running (VWR) in male mice decreased basal oxygen consumption rate (OCR) in brown adipocytes differentiated from the SVF of BAT (8), but cells from both sedentary and exercise-trained BAT were able to respond to pharmacological stimulation to a similar extent. Eleven days of VWR decreased NADH autofluorescence, an indirect marker of metabolism, compared to the sedentary controls (8). In contrast, 6–8 weeks of treadmill training in rats significantly increased expression of proteins involved in mitochondrial biogenesis, such as PGC1α, NRF1, or TFAM (119, 124). The reason for the discrepancies in these studies are unclear, although duration, exercise modality, or species investigated (rat or mouse), could contribute to these different responses to exercise.
Studies on the effect of exercise in BAT mitochondria in humans are limited. One study found no differences on PGC1α expression in BAT between endurance athletes and sedentary males (84). Overall, exercise appears to decrease mitochondrial activity in BAT in mice, but more human studies are needed to elucidate the effects of exercise on mitochondrial activity in BAT.
The effects of exercise on glucose uptake in BAT in rodents are conflicting. On one hand, some studies have shown that 2–8 weeks of exercise upregulates expression of genes involved in insulin signaling, glucose and fatty acid oxidation in BAT (8, 124, 125). However, 2 weeks of exercise decreased basal glucose uptake in brown adipocytes differentiated from SVF (8). Another study indicated that 6 weeks of exercise did not effect in vivo glucose uptake in BAT at room temperature or thermoneutrality, measured by in vivo insulin-stimulated 3H-2DG uptake (76). These data reveal that, although exercise results in an upregulation of genes involved in glucose metabolism, in vivo data in rodents indicates that exercise does not increase glucose uptake in BAT.
Several studies have indicated that exercise does not alter glucose uptake in BAT in humans. As little as 6 sessions of HIIT or moderate-intensity exercise-training in a 2 week period decreased insulin-stimulated glucose uptake in BAT (92), and 6 weeks of moderate-intensity continuous training did not affect cold-stimulated glucose uptake measured by 18FDG-PET/CT (126). In addition, endurance athletes have reduced glucose uptake in BAT when subjected to cold stimulation compared to sedentary subjects (measured by 18FDG-PET/CT) (84). Another study determined that there was no association of BAT mass or activity to physical activity in a cohort of 130 healthy, sedentary subjects (127). These data indicate that exercise or increased physical activity does not increase glucose metabolism in human BAT.
The effects of exercise on lipid metabolism in BAT has not been thoroughly investigated. Eleven days of exercise increased expression of genes involved in fatty acid oxidation (8), but decreased expression of genes involved in fatty acid biosynthesis (94), phospholipid metabolism (94) and lipolysis (8, 75). Oxidation of palmitate was also reduced in BAT ex vivo after 6 weeks of treadmill training (58).
Exercise affects the lipidomic profile of BAT by increasing total abundance of TAGs phosphatidylcholines (PC) and cholesterol esters, while decreasing cardiolipins and lysophosphatidylglycerols (94). Exercise also significantly increased several specific molecular species of PC and PE in BAT. These data show that exercise decreases lipid metabolism in BAT. To our knowledge, there are currently no studies analyzing the effect of exercise on lipid metabolism in human BAT. While it is clear that BAT lipid metabolism changes with exercise, the role of the exercise-induced decrease in lipolysis or changes in BAT lipidome have not been identified and will be the topic of future investigations.
It is important to note that in most cases, particularly in human studies, BAT activity, and mass are measured by glucose uptake. This is important in most settings, however, since exercise is a thermogenic activity it is unlikely that exercise would increase glucose uptake in BAT. This has led several groups to hypothesize that exercise may alter the endocrine activity of BAT. In fact, multiple studies have identified an endocrine role for BAT in response to exercise (13, 16, 128). Recent work in our laboratory identified the lipokine, 12,13-diHOME, to be released from BAT in response to exercise in mice and humans (9) Upregulation of 12,13-diHOME activates fatty acid uptake and oxidation in skeletal muscle without affecting glucose homeostasis (9). This data shows a direct role of BAT to improve metabolic health with exercise. These are the first data to identify a secreted factor from BAT with exercise to mediate skeletal muscle metabolic adaptations.
Exercise results in positive metabolic adaptations in both white and brown adipose tissue. Exercise increases mitochondrial activity, glucose metabolism, and endocrine activity in WAT in both rodents and humans. Notably, beiging of WAT only occurs with exercise in rodents, but both humans and rodents have increased mitochondrial activity independent of beiging of WAT. Exercise increases endocrine activity of BAT but does not affect glucose uptake in rodents and humans. Additionally, exercise does not affect thermogenesis and decreases mitochondrial activity in BAT in rodents.
An important point of investigation has been the effects of exercise-induced beiging in WAT. While this adaptation has been clearly identified in rodents, studies in humans have not identified the same effects. More recent studies have begun to investigate the effects of exercise at thermoneutrality to parse apart the direct effects of exercise on beiging, and have demonstrated that exercise at thermoneutrality blunts the effects of exercise on thermogenic gene expression (75, 76). Expanding these studies will provide greater insight and translational relevance for determining the effects of exercise on WAT (and potentially BAT).
Most of the studies discussed in this review have been conducted in either males or females. This is of particular importance as there are clear sex differences in adipose tissue depots among males and females, with females having a higher percentage of WAT (27) and higher BAT activity at rest (45). Another important issue in the field of BAT thermogenesis, especially in human studies, is the measurement of BAT activity. 18FDG-PET/CT is the gold standard for measurement of BAT mass and activity in humans, however, this analysis is solely based on the ability of BAT to uptake glucose to use it as a substrate. This highlights the importance of new techniques to accurately measure BAT activity and establish in vivo measurements of BAT thermogenic capacity, including in the context of exercise. Newer techniques such as infrared thermography and T2 mapping are potential mechanisms to elucidate the adaptations of BAT to exercise.
There is a need for the comprehensive understanding of the mechanisms underlying the chronic adaptations of adipose tissue with exercise. A single session of exercise leads to acute changes in expression of several genes (129). Successive bouts of exercise most lead to a cumulative effect of these acute changes resulting in chronic adaptations, which contribute to changes in glucose metabolism, fatty acid metabolism, and mitochondrial activity. Post-translational modifications such as protein phosphorylation regulate protein activity (130), and chronic exercise increases overall phosphorylation of proteins such as HSL and ATGL, which result in increased lipolytic activity (7, 75). Epigenetic modifications may also be underlying drivers of exercise-induce adaptations to exercise; studies have shown that exercise results in changes to the genome-wide DNA methylation pattern of human WAT (131, 132). These studies indicate that epigenetic modifications could oversee the chronic adaptations to adipose tissue with exercise by promoting or inhibiting expression of metabolic genes. Understanding factors that trigger exercise-induced adaptations remains an open field that will be an important for future investigations.
Together these studies highlight the importance of exercise to alter function of WAT and BAT that could provide important targets to improve metabolic health and reduce obesity. Future studies will investigate other mechanisms by which exercise exerts metabolic adaptations on adipose tissue such as increased mitochondrial function, improved glucose homeostasis or endocrine function, providing important translational relevance for exercise as a therapeutic tool.
PV and KS designed, wrote, and edited the manuscript.
This work was supported by National Institutes of Health Grants R01-HL138738 to KS, R01-AG060542 to KS.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Goodyear LJ, Kahn BB. Exercise, glucose transport, and insulin sensitivity. Annu Rev Med. (1998) 49:235–61. doi: 10.1146/annurev.med.49.1.235
2. Egan B, Zierath JR. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. (2013) 17:162–84. doi: 10.1016/j.cmet.2012.12.012
3. Hellsten Y, Nyberg M. Cardiovascular adaptations to exercise training. Compr Physiol. (2015) 6:1–32. doi: 10.1002/cphy.c140080
4. Blond MB, Rosenkilde M, Gram AS, Tindborg M, Christensen AN, Quist JS, et al. How does 6 months of active bike commuting or leisure-time exercise affect insulin sensitivity, cardiorespiratory fitness and intra-abdominal fat? A randomised controlled trial in individuals with overweight and obesity. Br J Sports Med. (2019) 53:1183–92. doi: 10.1136/bjsports-2018-100036
5. Stallknecht B, Vinten J, Ploug T, Galbo H. Increased activities of mitochondrial enzymes in white adipose tissue in trained rats. Am J Physiol. (1991) 261(3 Pt 1):E410–4. doi: 10.1152/ajpendo.1991.261.3.E410
6. Stanford KI, Middelbeek RJ, Townsend KL, Lee MY, Takahashi H, So K, et al. A novel role for subcutaneous adipose tissue in exercise-induced improvements in glucose homeostasis. Diabetes. (2015) 64:2002–14. doi: 10.2337/db14-0704
7. Stephenson EJ, Lessard SJ, Rivas DA, Watt MJ, Yaspelkis BB III, Koch LG, et al. Exercise training enhances white adipose tissue metabolism in rats selectively bred for low- or high-endurance running capacity. Am J Physiol Endocrinol Metab. (2013) 305:E429–38. doi: 10.1152/ajpendo.00544.2012
8. Lehnig AC, Dewal RS, Baer LA, Kitching KM, Munoz VR, Arts PJ, et al. Exercise training induces depot-specific adaptations to white and brown adipose tissue. iScience. (2019) 11:425–39. doi: 10.1016/j.isci.2018.12.033
9. Stanford KI, Lynes MD, Takahashi H, Baer LA, Arts PJ, May FJ, et al. 12,13-diHOME: an exercise-induced lipokine that increases skeletal muscle fatty acid uptake. Cell Metab. (2018) 27:1111–20.e3. doi: 10.1016/j.cmet.2018.04.023
10. Sutherland LN, Bomhof MR, Capozzi LC, Basaraba SA, Wright DC. Exercise and adrenaline increase PGC-1{alpha} mRNA expression in rat adipose tissue. J Physiol. (2009) 587(Pt 7):1607–17. doi: 10.1113/jphysiol.2008.165464
11. Craig BW, Hammons GT, Garthwaite SM, Jarett L, Holloszy JO. Adaptation of fat cells to exercise: response of glucose uptake and oxidation to insulin. J Appl Physiol Respir Environ Exerc Physiol. (1981) 51:1500–6. doi: 10.1152/jappl.1981.51.6.1500
12. Geng L, Liao B, Jin L, Huang Z, Triggle CR, Ding H, et al. Exercise alleviates obesity-induced metabolic dysfunction via enhancing FGF21 sensitivity in adipose tissues. Cell Rep. (2019) 26:2738–52.e4. doi: 10.1016/j.celrep.2019.02.014
13. Lee S, Norheim F, Langleite TM, Gulseth HL, Birkeland KI, Drevon CA. Effects of long-term exercise on plasma adipokine levels and inflammation-related gene expression in subcutaneous adipose tissue in sedentary dysglycaemic, overweight men and sedentary normoglycaemic men of healthy weight. Diabetologia. (2019) 62:1048–64. doi: 10.1007/s00125-019-4866-5
14. Takahashi H, Alves CRR, Stanford KI, Middelbeek RJW, Pasquale N, Ryan RE, et al. TGF-β2 is an exercise-induced adipokine that regulates glucose and fatty acid metabolism. Nat Metab. (2019) 1:291–303. doi: 10.1038/s42255-018-0030-7
15. Trevellin E, Scorzeto M, Olivieri M, Granzotto M, Valerio A, Tedesco L, et al. Exercise training induces mitochondrial biogenesis and glucose uptake in subcutaneous adipose tissue through eNOS-dependent mechanisms. Diabetes. (2014) 63:2800–11. doi: 10.2337/db13-1234
16. Rodriguez A, Becerril S, Ezquerro S, Mendez-Gimenez L, Fruhbeck G. Crosstalk between adipokines and myokines in fat browning. Acta Physiol. (2017) 219:362–81. doi: 10.1111/apha.12686
17. Garrow JS. New approaches to body composition. Am J Clin Nutr. (1982) 35(Suppl. 5):1152–8. doi: 10.1093/ajcn/35.5.1152
18. Goglia F, Geloen A, Lanni A, Minaire Y, Bukowiecki LJ. Morphometric-stereologic analysis of brown adipocyte differentiation in adult mice. Am J Physiol. (1992) 262(4 Pt 1):C1018–23. doi: 10.1152/ajpcell.1992.262.4.C1018
19. Ali AT, Hochfeld WE, Myburgh R, Pepper MS. Adipocyte and adipogenesis. Eur J Cell Biol. (2013) 92:229–36. doi: 10.1016/j.ejcb.2013.06.001
20. Burl RB, Ramseyer VD, Rondini EA, Pique-Regi R, Lee YH, Granneman JG. Deconstructing adipogenesis induced by β3-adrenergic receptor activation with single-cell expression profiling. Cell Metab. (2018) 28:300–9e4. doi: 10.1016/j.cmet.2018.05.025
21. Fawcett DW. Differences in physiological activity in brown and white fat as revealed by histochemical reactions. Science. (1947) 105:123–5. doi: 10.1126/science.105.2718.123
22. Labbé SM, Caron A, Chechi K, Laplante M, Lecomte R, Richard D. Metabolic activity of brown, “beige,” and white adipose tissues in response to chronic adrenergic stimulation in male mice. Am J Physiol Endocrinol Metab. (2016) 311:E260–8. doi: 10.1152/ajpendo.00545.2015
23. Bostrom P, Wu J, Jedrychowski MP, Korde A, Ye L, Lo JC, et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature. (2012) 481:463–8. doi: 10.1038/nature10777
24. Cao L, Choi EY, Liu X, Martin A, Wang C, Xu X, et al. White to brown fat phenotypic switch induced by genetic and environmental activation of a hypothalamic-adipocyte axis. Cell Metab. (2011) 14:324–38. doi: 10.1016/j.cmet.2011.06.020
25. Roncari DA, Hamilton BS. Cellular and molecular factors in adipose tissue growth and obesity. Adv Exp Med Biol. (1993) 334:269–77. doi: 10.1007/978-1-4615-2910-1_20
26. Tchoukalova YD, Votruba SB, Tchkonia T, Giorgadze N, Kirkland JL, Jensen MD. Regional differences in cellular mechanisms of adipose tissue gain with overfeeding. Proc Natl Acad Sci USA. (2010) 107:18226–31. doi: 10.1073/pnas.1005259107
27. Tchoukalova YD, Koutsari C, Karpyak MV, Votruba SB, Wendland E, Jensen MD. Subcutaneous adipocyte size and body fat distribution. Am J Clin Nutr. (2008) 87:56–63. doi: 10.1093/ajcn/87.1.56
28. Tchkonia T, Thomou T, Zhu Y, Karagiannides I, Pothoulakis C, Jensen MD, et al. Mechanisms and metabolic implications of regional differences among fat depots. Cell Metab. (2013) 17:644–56. doi: 10.1016/j.cmet.2013.03.008
29. Cinti S. The adipose organ. Prostaglandins Leukot Essent Fatty Acids. (2005) 73:9–15. doi: 10.1016/j.plefa.2005.04.010
30. de Jong JM, Larsson O, Cannon B, Nedergaard J. A stringent validation of mouse adipose tissue identity markers. Am J Physiol Endocrinol Metab. (2015) 308:E1085–105. doi: 10.1152/ajpendo.00023.2015
31. Kissebah AH, Krakower GR. Regional adiposity and morbidity. Physiol Rev. (1994) 74:761–811. doi: 10.1152/physrev.1994.74.4.761
32. Karpe F, Pinnick KE. Biology of upper-body and lower-body adipose tissue–link to whole-body phenotypes. Nat Rev Endocrinol. (2015) 11:90–100. doi: 10.1038/nrendo.2014.185
33. Wajchenberg BL. Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocr Rev. (2000) 21:697–738. doi: 10.1210/edrv.21.6.0415
34. Zuriaga MA, Fuster JJ, Gokce N, Walsh K. Humans and mice display opposing patterns of “Browning” gene expression in visceral and subcutaneous white adipose tissue depots. Front Cardiovasc Med. (2017) 4:27. doi: 10.3389/fcvm.2017.00027
35. Ibrahim MM. Subcutaneous and visceral adipose tissue: structural and functional differences. Obes Rev. (2010) 11:11–8. doi: 10.1111/j.1467-789X.2009.00623.x
36. Atzmon G, Yang XM, Muzumdar R, Ma XH, Gabriely I, Barzilai N. Differential gene expression between visceral and subcutaneous fat depots. Horm Metab Res. (2002) 34:622–8. doi: 10.1055/s-2002-38250
37. Pinnick KE, Nicholson G, Manolopoulos KN, McQuaid SE, Valet P, Frayn KN, et al. Distinct developmental profile of lower-body adipose tissue defines resistance against obesity-associated metabolic complications. Diabetes. (2014) 63:3785–97. doi: 10.2337/db14-0385
38. Himms-Hagen J. Brown adipose tissue thermogenesis, energy balance, and obesity. Can J Biochem Cell Biol. (1984) 62:610–7. doi: 10.1139/o84-081
39. Bartelt A, Bruns OT, Reimer R, Hohenberg H, Ittrich H, Peldschus K, et al. Brown adipose tissue activity controls triglyceride clearance. Nat Med. (2011) 17:200–5. doi: 10.1038/nm.2297
40. Lowell BB, Spiegelman BM. Towards a molecular understanding of adaptive thermogenesis. Nature. (2000) 404:652–60. doi: 10.1038/35007527
41. Chu M, Sampath H, Cahana DY, Kahl CA, Somwar R, Cornea A, et al. Spatiotemporal dynamics of triglyceride storage in unilocular adipocytes. Mol Biol Cell. (2014) 25:4096–105. doi: 10.1091/mbc.e14-06-1085
42. Cohen P, Spiegelman BM. Cell biology of fat storage. Mol Biol Cell. (2016) 27:2523–7. doi: 10.1091/mbc.e15-10-0749
43. Golozoubova V, Hohtola E, Matthias A, Jacobsson A, Cannon B, Nedergaard J. Only UCP1 can mediate adaptive nonshivering thermogenesis in the cold. FASEB J. (2001) 15:2048–50. doi: 10.1096/fj.00-0536fje
44. Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiol Rev. (2004) 84:277–359. doi: 10.1152/physrev.00015.2003
45. Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, et al. Identification and importance of brown adipose tissue in adult humans. N Engl J Med. (2009) 360:1509–17. doi: 10.1056/NEJMoa0810780
46. Saito M, Okamatsu-Ogura Y, Matsushita M, Watanabe K, Yoneshiro T, Nio-Kobayashi J, et al. High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. Diabetes. (2009) 58:1526–31. doi: 10.2337/db09-0530
47. van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JM, Kemerink GJ, Bouvy ND, et al. Cold-activated brown adipose tissue in healthy men. N Engl J Med. (2009) 360:1500–8. doi: 10.1056/NEJMoa0808718
48. Virtanen KA, Lidell ME, Orava J, Heglind M, Westergren R, Niemi T, et al. Functional brown adipose tissue in healthy adults. N Engl J Med. (2009) 360:1518–25. doi: 10.1056/NEJMoa0808949
49. Ikeda K, Maretich P, Kajimura S. The common and distinct features of brown and beige adipocytes. Trends Endocrinol Metab. (2018) 29:191–200. doi: 10.1016/j.tem.2018.01.001
50. Lean ME, James WP, Jennings G, Trayhurn P. Brown adipose tissue uncoupling protein content in human infants, children and adults. Clin Sci. (1986) 71:291–7. doi: 10.1042/cs0710291
51. Jespersen NZ, Feizi A, Andersen ES, Heywood S, Hattel HB, Daugaard S, et al. Heterogeneity in the perirenal region of humans suggests presence of dormant brown adipose tissue that contains brown fat precursor cells. Mol Metab. (2019) 24:30–43. doi: 10.1016/j.molmet.2019.03.005
52. Nedergaard J, Cannon B. The browning of white adipose tissue: some burning issues. Cell Metab. (2014) 20:396–407. doi: 10.1016/j.cmet.2014.07.005
53. Seale P, Bjork B, Yang W, Kajimura S, Chin S, Kuang S, et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature. (2008) 454:961–7. doi: 10.1038/nature07182
54. Lepper C, Fan CM. Inducible lineage tracing of Pax7-descendant cells reveals embryonic origin of adult satellite cells. Genesis. (2010) 48:424–36. doi: 10.1002/dvg.20630
55. Berry DC, Jiang Y, Graff JM. Mouse strains to study cold-inducible beige progenitors and beige adipocyte formation and function. Nat Commun. (2016) 7:10184. doi: 10.1038/ncomms10184
56. Wu J, Bostrom P, Sparks LM, Ye L, Choi JH, Giang AH, et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell. (2012) 150:366–76. doi: 10.1016/j.cell.2012.05.016
57. Jones NL, Heigenhauser GJ, Kuksis A, Matsos CG, Sutton JR, Toews CJ. Fat metabolism in heavy exercise. Clin Sci. (1980) 59:469–78. doi: 10.1042/cs0590469
58. Wu MV, Bikopoulos G, Hung S, Ceddia RB. Thermogenic capacity is antagonistically regulated in classical brown and white subcutaneous fat depots by high fat diet and endurance training in rats: impact on whole-body energy expenditure. J Biol Chem. (2014) 289:34129–40. doi: 10.1074/jbc.M114.591008
59. Knudsen JG, Murholm M, Carey AL, Bienso RS, Basse AL, Allen TL, et al. Role of IL-6 in exercise training- and cold-induced UCP1 expression in subcutaneous white adipose tissue. PLoS ONE. (2014) 9:e84910. doi: 10.1371/journal.pone.0084910
60. Xu X, Ying Z, Cai M, Xu Z, Li Y, Jiang SY, et al. Exercise ameliorates high-fat diet-induced metabolic and vascular dysfunction, and increases adipocyte progenitor cell population in brown adipose tissue. Am J Physiol Regul Integr Comp Physiol. (2011) 300:R1115–25. doi: 10.1152/ajpregu.00806.2010
61. Chi J, Wu Z, Choi CHJ, Nguyen L, Tegegne S, Ackerman SE, et al. Three-dimensional adipose tissue imaging reveals regional variation in beige fat biogenesis and PRDM16-dependent sympathetic neurite density. Cell Metab. (2018) 27:226–36.e3. doi: 10.1016/j.cmet.2017.12.011
62. Seale P, Conroe HM, Estall J, Kajimura S, Frontini A, Ishibashi J, et al. Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J Clin Invest. (2011) 121:96–105. doi: 10.1172/JCI44271
63. Ghorbani M, Claus TH, Himms-Hagen J. Hypertrophy of brown adipocytes in brown and white adipose tissues and reversal of diet-induced obesity in rats treated with a beta3-adrenoceptor agonist. Biochem Pharmacol. (1997) 54:121–31. doi: 10.1016/S0006-2952(97)00162-7
64. Ghorbani M, Himms-Hagen J. Appearance of brown adipocytes in white adipose tissue during CL 316,243-induced reversal of obesity and diabetes in Zucker fa/fa rats. Int J Obes Relat Metab Disord. (1997) 21:465–75. doi: 10.1038/sj.ijo.0800432
65. Cousin B, Cinti S, Morroni M, Raimbault S, Ricquier D, Penicaud L, et al. Occurrence of brown adipocytes in rat white adipose tissue: molecular and morphological characterization. J Cell Sci. (1992) 103(Pt 4):931–42.
66. Lehnig AC, Stanford KI. Exercise-induced adaptations to white and brown adipose tissue. J Exp Biol. (2018) 221(Pt Suppl. 1):jeb161570. doi: 10.1242/jeb.161570
67. Saugen E, Vollestad NK. Nonlinear relationship between heat production and force during voluntary contractions in humans. J Appl Physiol. (1995) 79:2043–9. doi: 10.1152/jappl.1995.79.6.2043
68. Ranallo RF, Rhodes EC. Lipid metabolism during exercise. Sports Med. (1998) 26:29–42. doi: 10.2165/00007256-199826010-00003
69. Feldman BJ, Streeper RS, Farese RV Jr, Yamamoto KR. Myostatin modulates adipogenesis to generate adipocytes with favorable metabolic effects. Proc Natl Acad Sci USA. (2006) 103:15675–80. doi: 10.1073/pnas.0607501103
70. Rao RR, Long JZ, White JP, Svensson KJ, Lou J, Lokurkar I, et al. Meteorin-like is a hormone that regulates immune-adipose interactions to increase beige fat thermogenesis. Cell. (2014) 157:1279–91. doi: 10.1016/j.cell.2014.03.065
71. Carriere A, Jeanson Y, Berger-Muller S, Andre M, Chenouard V, Arnaud E, et al. Browning of white adipose cells by intermediate metabolites: an adaptive mechanism to alleviate redox pressure. Diabetes. (2014) 63:3253–65. doi: 10.2337/db13-1885
72. Roberts LD, Bostrom P, O'Sullivan JF, Schinzel RT, Lewis GD, Dejam A, et al. beta-Aminoisobutyric acid induces browning of white fat and hepatic beta-oxidation and is inversely correlated with cardiometabolic risk factors. Cell Metab. (2014) 19:96–108. doi: 10.1016/j.cmet.2013.12.003
73. Stanford KI, Goodyear LJ. Exercise regulation of adipose tissue. Adipocyte. (2016) 5:153–62. doi: 10.1080/21623945.2016.1191307
74. Fischer AW, Cannon B, Nedergaard J. Optimal housing temperatures for mice to mimic the thermal environment of humans: an experimental study. Mol Metab. (2018) 7:161–70. doi: 10.1016/j.molmet.2017.10.009
75. McKie GL, Medak KD, Knuth CM, Shamshoum H, Townsend LK, Peppler WT, et al. Housing temperature affects the acute and chronic metabolic adaptations to exercise in mice. J Physiol. (2019) 597:4581–600. doi: 10.1113/JP278221
76. Raun SH, Henriquez-Olguín C, Karavaeva I, Ali M, Møller LLV, Kot W, et al. Housing temperature influences exercise training adaptations in mice. Nat Commun. (2020) 11:1560. doi: 10.1038/s41467-020-15311-y
77. Brandao CFC, de Carvalho FG, Souza AO, Junqueira-Franco MVM, Batitucci G, Couto-Lima CA, et al. Physical training, UCP1 expression, mitochondrial density, and coupling in adipose tissue from women with obesity. Scand J Med Sci Sports. (2019) 29:1699–706. doi: 10.1111/sms.13514
78. Camera DM, Anderson MJ, Hawley JA, Carey AL. Short-term endurance training does not alter the oxidative capacity of human subcutaneous adipose tissue. Eur J Appl Physiol. (2010) 109:307–16. doi: 10.1007/s00421-010-1356-3
79. Norheim F, Langleite TM, Hjorth M, Holen T, Kielland A, Stadheim HK, et al. The effects of acute and chronic exercise on PGC-1α, irisin and browning of subcutaneous adipose tissue in humans. FEBS J. (2014) 281:739–49. doi: 10.1111/febs.12619
80. Tsiloulis T, Carey AL, Bayliss J, Canny B, Meex RCR, Watt MJ. No evidence of white adipocyte browning after endurance exercise training in obese men. Int J Obes (Lond). (2018) 42:721–7. doi: 10.1038/ijo.2017.295
81. Nakhuda A, Josse AR, Gburcik V, Crossland H, Raymond F, Metairon S, et al. Biomarkers of browning of white adipose tissue and their regulation during exercise- and diet-induced weight loss. Am J Clin Nutr. (2016) 104:557–65. doi: 10.3945/ajcn.116.132563
82. Stinkens R, Brouwers B, Jocken JW, Blaak EE, Teunissen-Beekman KF, Hesselink MK, et al. Exercise training-induced effects on the abdominal subcutaneous adipose tissue phenotype in humans with obesity. J Appl Physiol. (2018) 125:1585–93. doi: 10.1152/japplphysiol.00496.2018
83. Pino MF, Parsons SA, Smith SR, Sparks LM. Active individuals have high mitochondrial content and oxidative markers in their abdominal subcutaneous adipose tissue. Obesity. (2016) 24:2467–70. doi: 10.1002/oby.21669
84. Vosselman MJ, Hoeks J, Brans B, Pallubinsky H, Nascimento EB, van der Lans AA, et al. Low brown adipose tissue activity in endurance-trained compared with lean sedentary men. Int J Obes. (2015) 39:1696–702. doi: 10.1038/ijo.2015.130
85. Castellani L, Root-Mccaig J, Frendo-Cumbo S, Beaudoin MS, Wright DC. Exercise training protects against an acute inflammatory insult in mouse epididymal adipose tissue. J Appl Physiol. (2014) 116:1272–80. doi: 10.1152/japplphysiol.00074.2014
86. Knuth CM, Peppler WT, Townsend LK, Miotto PM, Gudiksen A, Wright DC. Prior exercise training improves cold tolerance independent of indices associated with non-shivering thermogenesis. J Physiol. (2018) 596:4375–91. doi: 10.1113/JP276228
87. Laye MJ, Rector RS, Warner SO, Naples SP, Perretta AL, Uptergrove GM, et al. Changes in visceral adipose tissue mitochondrial content with type 2 diabetes and daily voluntary wheel running in OLETF rats. J Physiol. (2009) 587(Pt 14):3729–39. doi: 10.1113/jphysiol.2009.172601
88. Dohlmann TL, Hindso M, Dela F, Helge JW, Larsen S. High-intensity interval training changes mitochondrial respiratory capacity differently in adipose tissue and skeletal muscle. Physiol Rep. (2018) 6:e13857. doi: 10.14814/phy2.13857
89. Ronn T, Volkov P, Tornberg A, Elgzyri T, Hansson O, Eriksson KF, et al. Extensive changes in the transcriptional profile of human adipose tissue including genes involved in oxidative phosphorylation after a 6-month exercise intervention. Acta Physiol. (2014) 211:188–200. doi: 10.1111/apha.12247
90. Flores-Opazo M, Boland E, Garnham A, Murphy RM, McGee SL, Hargreaves M. Exercise and GLUT4 in human subcutaneous adipose tissue. Physiol Rep. (2018) 6:e13918. doi: 10.14814/phy2.13918
91. Ikemoto S, Thompson KS, Itakura H, Lane MD, Ezaki O. Expression of an insulin-responsive glucose transporter (GLUT4) minigene in transgenic mice: effect of exercise and role in glucose homeostasis. Proc Natl Acad Sci USA. (1995) 92:865–9. doi: 10.1073/pnas.92.3.865
92. Motiani P, Virtanen KA, Motiani KK, Eskelinen JJ, Middelbeek RJ, Goodyear LJ, et al. Decreased insulin-stimulated brown adipose tissue glucose uptake after short-term exercise training in healthy middle-aged men. Diabetes Obes Metab. (2017) 19:1379–88. doi: 10.1111/dom.12947
93. Wolfe RR. Fat metabolism in exercise. Adv Exp Med Biol. (1998) 441:147–56. doi: 10.1007/978-1-4899-1928-1_14
94. May FJ, Baer LA, Lehnig AC, So K, Chen EY, Gao F, et al. Lipidomic adaptations in white and brown adipose tissue in response to exercise demonstrate molecular species-specific remodeling. Cell Rep. (2017) 18:1558–72. doi: 10.1016/j.celrep.2017.01.038
95. Bartz R, Zehmer JK, Zhu M, Chen Y, Serrero G, Zhao Y, et al. Dynamic activity of lipid droplets: protein phosphorylation and GTP-mediated protein translocation. J Proteome Res. (2007) 6:3256–65. doi: 10.1021/pr070158j
96. Zimmermann R, Strauss JG, Haemmerle G, Schoiswohl G, Birner-Gruenberger R, Riederer M, et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science. (2004) 306:1383–6. doi: 10.1126/science.1100747
97. Steinberg D. Interconvertible enzymes in adipose tissue regulated by cyclic AMP-dependent protein kinase. Adv Cyclic Nucleotide Res. (1976) 7:157–98.
98. Higa TS, Spinola AV, Fonseca-Alaniz MH, Evangelista FS. Remodeling of white adipose tissue metabolism by physical training prevents insulin resistance. Life Sci. (2014) 103:41–8. doi: 10.1016/j.lfs.2014.02.039
99. Hackney AC, Lane AR. Exercise and the regulation of endocrine hormones. Prog Mol Biol Transl Sci. (2015) 135:293–311. doi: 10.1016/bs.pmbts.2015.07.001
100. Gollisch KS, Brandauer J, Jessen N, Toyoda T, Nayer A, Hirshman MF, et al. Effects of exercise training on subcutaneous and visceral adipose tissue in normal- and high-fat diet-fed rats. Am J Physiol Endocrinol Metab. (2009) 297:E495–504. doi: 10.1152/ajpendo.90424.2008
101. Bradley RL, Jeon JY, Liu FF, Maratos-Flier E. Voluntary exercise improves insulin sensitivity and adipose tissue inflammation in diet-induced obese mice. Am J Physiol Endocrinol Metab. (2008) 295:E586–94. doi: 10.1152/ajpendo.00309.2007
102. McPherron AC, Lawler AM, Lee SJ. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature. (1997) 387:83–90. doi: 10.1038/387083a0
103. Hittel DS, Axelson M, Sarna N, Shearer J, Huffman KM, Kraus WE. Myostatin decreases with aerobic exercise and associates with insulin resistance. Med Sci Sports Exerc. (2010) 42:2023–9. doi: 10.1249/MSS.0b013e3181e0b9a8
104. Shan T, Liang X, Bi P, Kuang S. Myostatin knockout drives browning of white adipose tissue through activating the AMPK-PGC1α-Fndc5 pathway in muscle. FASEB J. (2013) 27:1981–9. doi: 10.1096/fj.12-225755
105. Pillon NJ, Gabriel BM, Dollet L, Smith JAB, Sardon Puig L, Botella J, et al. Transcriptomic profiling of skeletal muscle adaptations to exercise and inactivity. Nat Commun. (2020) 11:470. doi: 10.1038/s41467-019-13869-w
106. Martinez-Huenchullan SF, Ban LA, Olaya-Agudo LF, Maharjan BR, Williams PF, Tam CS, et al. Constant-Moderate and high-intensity interval training have differential benefits on insulin sensitive tissues in high-fat fed mice. Front Physiol. (2019) 10:459. doi: 10.3389/fphys.2019.00459
107. Groussard C, Maillard F, Vazeille E, Barnich N, Sirvent P, Otero YF, et al. Tissue-Specific oxidative stress modulation by exercise: a comparison between MICT and HIIT in an obese rat model. Oxid Med Cell Longev. (2019) 2019:1965364. doi: 10.1155/2019/1965364
108. Lunt H, Draper N, Marshall HC, Logan FJ, Hamlin MJ, Shearman JP, et al. High intensity interval training in a real world setting: a randomized controlled feasibility study in overweight inactive adults, measuring change in maximal oxygen uptake. PLoS ONE. (2014) 9:e83256. doi: 10.1371/journal.pone.0083256
109. Viana RB, Naves JPA, Coswig VS, de Lira CAB, Steele J, Fisher JP, et al. Is interval training the magic bullet for fat loss? A systematic review and meta-analysis comparing moderate-intensity continuous training with high-intensity interval training (HIIT). Br J Sports Med. (2019) 53:655–64. doi: 10.1136/bjsports-2018-099928
110. Bartelt A, Heeren J. Adipose tissue browning and metabolic health. Nat Rev Endocrinol. (2014) 10:24–36. doi: 10.1038/nrendo.2013.204
111. Richard D, Rivest S. The role of exercise in thermogenesis and energy balance. Can J Physiol Pharmacol. (1989) 67:402–9. doi: 10.1139/y89-064
112. Hirata K. Blood flow to brown adipose tissue and norepinephrine- induced calorigenesis in physically trained rats. Jpn J Physiol. (1982) 32:279–91. doi: 10.2170/jjphysiol.32.279
113. Hirata K. Enhanced calorigenesis in brown adipose tissue in physically trained rats. Jpn J Physiol. (1982) 32:647–53. doi: 10.2170/jjphysiol.32.647
114. Wickler SJ, Stern JS, Glick Z, Horwitz BA. Thermogenic capacity and brown fat in rats exercise-trained by running. Metabolism. (1987) 36:76–81. doi: 10.1016/0026-0495(87)90067-9
115. Moriya K, Leblanc J, Arnold J. Effects of exercise and intermittent cold exposure on shivering and nonshivering thermogenesis in rats. Jpn J Physiol. (1987) 37:715–27. doi: 10.2170/jjphysiol.37.715
116. Richard D, Arnold J, Leblanc J. Energy balance in exercise-trained rats acclimated at two environmental temperatures. J Appl Physiol. (1986) 60:1054–9. doi: 10.1152/jappl.1986.60.3.1054
117. Yoshioka K, Yoshida T, Wakabayashi Y, Nishioka H, Kondo M. Effects of exercise training on brown adipose tissue thermogenesis in ovariectomized obese rats. Endocrinol Jpn. (1989) 36:403–8. doi: 10.1507/endocrj1954.36.403
118. Scarpace PJ, Yenice S, Tumer N. Influence of exercise training and age on uncoupling protein mRNA expression in brown adipose tissue. Pharmacol Biochem Behav. (1994) 49:1057–9. doi: 10.1016/0091-3057(94)90264-X
119. De Matteis R, Lucertini F, Guescini M, Polidori E, Zeppa S, Stocchi V, et al. Exercise as a new physiological stimulus for brown adipose tissue activity. Nutr Metab Cardiovasc Dis. (2013) 23:582–90. doi: 10.1016/j.numecd.2012.01.013
120. Singhal V, Maffazioli GD, Ackerman KE, Lee H, Elia EF, Woolley R, et al. Effect of chronic athletic activity on brown fat in young women. PLoS ONE. (2016) 11:e0156353. doi: 10.1371/journal.pone.0156353
121. Carpentier AC, Blondin DP, Virtanen KA, Richard D, Haman F, Turcotte EE. Brown adipose tissue energy metabolism in humans. Front Endocrinol. (2018) 9:447. doi: 10.3389/fendo.2018.00447
122. Nirengi S, Wakabayashi H, Matsushita M, Domichi M, Suzuki S, Sukino S, et al. An optimal condition for the evaluation of human brown adipose tissue by infrared thermography. PLoS ONE. (2019) 14:e0220574. doi: 10.1371/journal.pone.0220574
123. Holstila M, Pesola M, Saari T, Koskensalo K, Raiko J, Borra RJ, et al. MR signal-fat-fraction analysis and T2* weighted imaging measure BAT reliably on humans without cold exposure. Metabolism. (2017) 70:23–30. doi: 10.1016/j.metabol.2017.02.001
124. de Las Heras N, Klett-Mingo M, Ballesteros S, Martin-Fernandez B, Escribano O, Blanco-Rivero J, et al. Chronic exercise improves mitochondrial function and insulin sensitivity in brown adipose tissue. Front Physiol. (2018) 9:1122. doi: 10.3389/fphys.2018.01122
125. Barbosa MA, Guerra-Sa R, De Castro UGM, de Lima WG, Dos Santos RAS, Campagnole-Santos MJ, et al. Physical training improves thermogenesis and insulin pathway, and induces remodeling in white and brown adipose tissues. J Physiol Biochem. (2018) 74:441–54. doi: 10.1007/s13105-018-0637-x
126. Motiani P, Teuho J, Saari T, Virtanen KA, Honkala SM, Middelbeek RJ, et al. Exercise training alters lipoprotein particles independent of brown adipose tissue metabolic activity. Obes Sci Pract. (2019) 5:258–72. doi: 10.1002/osp4.330
127. Acosta FM, Martinez-Tellez B, Sanchez-Delgado G, Migueles JH, Contreras-Gomez MA, Martinez-Avila WD, et al. Association of objectively measured physical activity with brown adipose tissue volume and activity in young adults. J Clin Endocrinol Metab. (2019) 104:223–33. doi: 10.1210/jc.2018-01312
128. Stanford KI, Goodyear LJ. Muscle-Adipose tissue cross talk. Cold Spring Harb Perspect Med. (2018) 8:a029801. doi: 10.1101/cshperspect.a029801
129. Shen Y, Zhou H, Jin W, Lee HJ. Acute exercise regulates adipogenic gene expression in white adipose tissue. Biol Sport. (2016) 33:381–91. doi: 10.5604/20831862.1224395
130. Krebs EG. Historical perspectives on protein phosphorylation and a classification system for protein kinases. Philos Trans R Soc Lond B Biol Sci. (1983) 302:3–11. doi: 10.1098/rstb.1983.0033
131. Ronn T, Volkov P, Davegardh C, Dayeh T, Hall E, Olsson AH, et al. A six months exercise intervention influences the genome-wide DNA methylation pattern in human adipose tissue. PLoS Genet. (2013) 9:e1003572. doi: 10.1371/journal.pgen.1003572
Keywords: exercise, obesity, white adipose tissue (WAT), brown adipose tissue (BAT), thermogenesis
Citation: Vidal P and Stanford KI (2020) Exercise-Induced Adaptations to Adipose Tissue Thermogenesis. Front. Endocrinol. 11:270. doi: 10.3389/fendo.2020.00270
Received: 10 February 2020; Accepted: 14 April 2020;
Published: 29 April 2020.
Edited by:
Takeshi Yoneshiro, University of California, San Francisco, United StatesReviewed by:
Alexander Bartelt, Ludwig Maximilian University of Munich, GermanyCopyright © 2020 Vidal and Stanford. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kristin I. Stanford, a3Jpc3Rpbi5zdGFuZm9yZEBvc3VtYy5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.