 Mariana Ferreira Bizzi
Mariana Ferreira Bizzi Graeme B. Bolger2,3†‡
Graeme B. Bolger2,3†‡ Márta Korbonits
Márta Korbonits
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Endocrinol. , 19 March 2019
Sec. Pituitary Endocrinology
Volume 10 - 2019 | https://doi.org/10.3389/fendo.2019.00141
This article is part of the Research Topic Molecular network study of pituitary adenomas View all 16 articles
Human phosphodiesterases (PDEs) comprise a complex superfamily of enzymes derived from 24 genes separated into 11 PDE gene families (PDEs 1–11), expressed in different tissues and cells, including heart and brain. The isoforms PDE4, PDE7, and PDE8 are specific for the second messenger cAMP, which is responsible for mediating diverse physiological actions involving different hormones and neurotransmitters. The cAMP pathway plays an important role in the development and function of endocrine tissues while phosphodiesterases are responsible for ensuring the appropriate intensity of the actions of this pathway by hydrolyzing cAMP to its inactive form 5'-AMP. PDE1, PDE2, PDE4, and PDE11A are highly expressed in the pituitary, and overexpression of some PDE4 isoforms have been demonstrated in different pituitary adenoma subtypes. This observed over-expression in pituitary adenomas, although of unknown etiology, has been considered a compensatory response to tumorigenesis. PDE4A4/5 has a unique interaction with the co-chaperone aryl hydrocarbon receptor-interacting protein (AIP), a protein implicated in somatotroph tumorigenesis via germline loss-of-function mutations. Based on the association of low PDE4A4 expression with germline AIP-mutation-positive samples, the available data suggest that lack of AIP hinders the upregulation of PDE4A4 protein seen in sporadic somatotrophinomas. This unique disturbance of the cAMP-PDE pathway observed in the majority of AIP-mutation positive adenomas could contribute to their well-described poor response to somatostatin analogs and may support a role in tumorigenesis.
Human phosphodiesterases (PDEs) comprise a complex superfamily of enzymes classified into 11 families, encoded by 24 genes representing over 100 different proteins. Many of these genes express several different mRNAs, and the resulting proteins vary widely in their distribution in various tissues and in various intracellular compartments (1).
PDE isoforms differ in their kinetics, distribution, and susceptibility to pharmacological inhibition, as well as selectivity for their different substrates, 3',5' cyclic monophosphate (cAMP) and 3',5' cyclic guanosine monophosphate (cGMP) (1). PDEs share some common structural characteristics: all PDE isoforms have a conserved catalytic domain of ~300 amino acids, located in the C-terminal portion of the protein, and most PDE isoforms contain family-specific regulatory regions in their N-terminal portions (2).
The catalytic regions of each family member differ in amino acid sequence and tertiary structure, which accounts for their specificity for substrate (cAMP and/or cGMP) and their ability to be inhibited by family- and isoform-specific inhibitors. PDE 4, 7, and 8 selectively hydrolyze cAMP, PDE 5, 6, and 9 are selective for cGMP, while PDEs 1, 2, 3, 10, and 11 hydrolyze both, although the specificity is variable (1, 3, 4).
The expression pattern of PDE isoforms varies between tissues and reflects their proliferative state and hormonal stimuli. In this mini review we aim to highlight the important role of these enzymes in pituitary diseases, especially the PDE4A4/5 isoform, encoded by the PDE4A gene, which has been implicated in GH-secreting adenomas due to its selective interaction with aryl hydrocarbon receptor-interacting protein (AIP), a known tumor suppressor gene (5).
The pituitary gland is a target of different neuroendocrine hormones, which play a crucial role in the control of cell differentiation and proliferation, in addition to hormone secretion, through specific interactions with members of the superfamily of G protein-coupled receptors (GPCRs) (6, 7) (Figure 1). The regulatory, usually hypothalamic, hormone couples to the G protein-coupled receptor in the cell of interest and a conformational change results in activation of the G protein complex. In the case of GHRH, the Gsα subunit is released from the αβγ G protein complex and binds to adenylyl cyclase, which then catalyzes the conversion of ATP into the second messenger cAMP. cAMP activates a cascade of other enzymes, thus amplifying the cellular reaction (3). Following GHRH activation of somatotrophs cAMP binds the regulatory subunit of protein kinase A (PKA) (3, 6). The activated catalytic subunit of PKA then phosphorylate a series of targets that regulate effector enzymes, ion channels, and activate the transcription of specific genes that mediate cell growth and differentiation. Additional effectors of cAMP include the exchange factor regulated by cAMP (EPAC) protein, cyclic nucleotide-gated ion channels, Popeye proteins, and possibly additional targets that are still under investigation (1, 8).
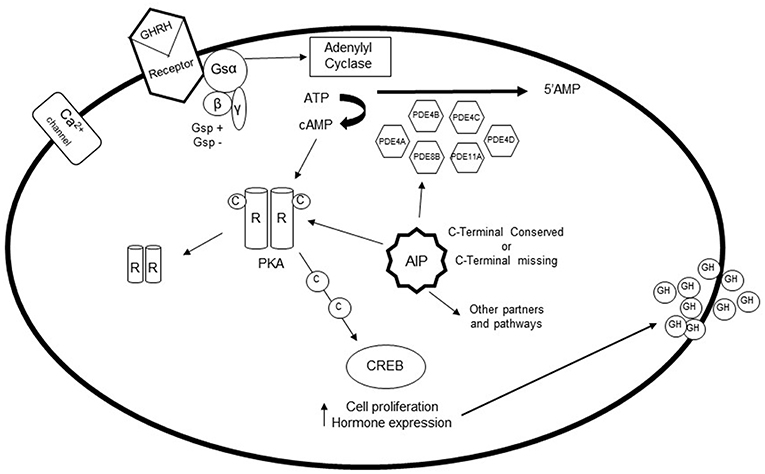
Figure 1. The role of phosphodiesterases (PDEs) in the pituitary gland: After stimulation of somatotroph cells via GHRH, the G protein coupled receptor is activated, which causes a conformational change of the receptor. The Gsα subunit detaches from the complex, and binds to adenylyl cyclase, which catalyzes the conversion of ATP to cAMP. Elevation of intracellular cAMP leads to dissociation of the catalytic subunit and the regulatory subunit of protein kinase A (PKA). Activation of protein kinase A can then phosphorylate a number of targets that regulate effector enzymes and ion channels as well as activates gene transcription that play a role in cell growth and differentiation. PDEs are fundamental in regulating this pathway, since they are the only enzymes capable of hydrolyzing cAMP to its inactive 5'-AMP form. PDE4A, PDE4B, PDE4C, PDE4D, PDE8B, and PDE11A are increased in GH-secreting adenomas, possibly as a compensatory mechanism. However, gsp and AIP mutations interfere with the expression of these PDEs.
PDEs act as regulators of the cAMP pathway, as they are capable of hydrolyzing cAMP to its inactive 5'-AMP form, which is the main pathway for inactivation of cAMP (3, 6). As a consequence, cAMP can either suppress cell proliferation and the mitogenic action of growth factors in some cell types, or conversely, promote the transition from cell cycle G0 to G1 and stimulate cell growth in others (9, 10). It is unclear, for example, why cAMP has a proliferative role in the somatotroph cells while an anti-proliferative role in gonadotroph cells (6, 9, 10). cAMP signaling is temporally, spatially, and functionally regulated by compartmentalization and influenced by a complex network of cell- and tissue-specific downstream effectors and regulators (11). In the pituitary, cAMP acts as a key signaling molecule that controls responsiveness to mitogens and secretagogues, such as hypothalamic hormones, neurotransmitters, and other peripheral factors (7) and a dysregulated cAMP-pathway is involved in the pathogenesis and response to therapy of pituitary adenomas (11).
PDEs are directly implicated in various endocrine disorders affecting the pituitary, adrenals, thyroid, testes, and ovaries (3). Little is known about the expression of PDE isoforms in the pituitary gland, especially in humans, since the vast majority of studies on the association between PDEs and endocrine functions have been performed in vitro or in animals. mRNA studies have implicated PDE1, PDE2, PDE4, and PDE11A as being the most highly expressed PDEs in the pituitary (3, 12–14). Interestingly, PDE4 is the only selective PDE for cAMP. The discovery of the physiological role of PDEs in the human pituitary has been hindered due to the lack of availability of specific antibodies. In addition, mRNA does not always reflect the protein amount or function due to variations in translation, protein stability, or posttranslational modifications.
PDE4 isoforms in mammals are encoded by four different genes (PDE4A, PDE4B, PDE4C, and PDE4D) and each of these genes encodes multiple isoforms, through the use of specific promoters for each isoform and alternative messenger RNA processing (15–17). PDE4s differ from the other PDE families by their specific catalytic regions (15–17), as well as by the presence of two “signature” regions called upstream conserved regions (UCRs), which are located in the N-terminal third of the proteins and referred to as UCR1 and UCR2 (18). The various isoforms encoded by each of the PDE4A, PDE4B, PDE4C, and PDE4D genes are divided into three groups: 'long' isoforms that contain both UCR1 and UCR2, “short” isoforms that do not have UCR1 but include UCR2, and “super short” isoforms that do not have UCR1 and contain a truncated UCR2 (18).
PDE4A8 is a long isoform of the PDE4A family with an N-terminal region distinct from the other PDE4A long isoforms PDE4A4, PDE4A10, and PDE4A11 (19–22). It is expressed at significant levels in various regions of the brain, especially in regions involved in coordination, sensation and higher cognitive functions (12, 20). It is also expressed in the pituitary gland (23).
PDE4A4, the human analog of rodent PDE4A5, is an isoform expressed in a wide variety of tissues, including lung and various brain regions (18–20, 24, 25). This isoform has UCR1 and UCR2 as well as a unique N-terminal region, which is highly conserved in mammals and has 88% similarity to the N-terminal region of rat PDE4A5 (20). This high degree of conservation between species suggests that the unique amino-terminal region of the PDE4A4 isoform has specific functions (26). The truncation of the PDE4A4 N-terminal region alters its enzymatic activities, its intracellular targeting, and its interaction with other proteins (27, 28). PDE4A5 interacts with AIP and has a reduced expression in AIP-mutation-positive adenomas (5, 26, 29, 30). PDE4A4 is expressed in the human pituitary (23). Furthermore, reduced AIP levels were shown to disproportionally enhance the PKA pathway activity under PDE4-specific inhibition in pituitary somatotrophs, pointing out to a link with the disease process involved in Carney complex (31).
By semi quantitative RT-PCR, PDE4C, and PDE4D were also shown to be expressed in the normal pituitary while no expression was detected for PDE8B (14).
Both PDE4A4 and PDE4A8 expression is increased in GH-, PRL-, ACTH- and FSH-secreting adenoma cells compared to their respective normal pituitary cells (23). Interestingly, the augmentation of PDEs observed in pituitary adenomas reflects a consequent increase in PKA activating transcription of cell growth promoting genes, suggesting that these phosphodiesterases might be increased as a possible adaptation or compensation to tumorigenesis, in an attempt to suppress the proliferative drive (23).
Protein-protein interaction between AIP and PDE2A (PDE2A1, PDE2A2, and PDE2A3) has been described (32). Although PDE2As has cGMP as their preferred substrate it may also hydrolyze cAMP (33).
PDE11A has higher expression in GH-secreting adenomas when compared with normal GH-cells (13), which is also described as a phenotype modifier in patients with Carney complex due to PRKAR1A mutations (34). The presence and role of PDE11A expression and variants were studied in somatotroph adenomas. Although nonsense and missense PDE11A variants were found in 20% of patients with acromegaly, there was no significant difference in variant frequency compared with controls, suggesting that these variants are unlikely to contribute to the pathogenesis of GH-secreting adenomas since the conservation of the wild-type allele of PDE11A remains in the majority of tumor samples and no significant clinical phenotype could be observed in patients with variant PDE11A (13).
Interestingly, although PDE8B was not detected in normal pituitary, this isoform was shown to be overexpressed in all GH-secreting adenomas, especially higher levels were observed in gsp-positive tumors (14). This study also showed that while PDE4C and 4D RNA expression is not increased in gsp-mutation negative GH-secreting adenomas compared to normal pituitary, gsp-positive samples had seven times higher expression (14). As cAMP-responsive element-binding protein represents the main endpoint of the cAMP pathway, the observed enhanced phosphodiesterase activities may significantly impact the phenotypic expression of gsp mutations in somatotrophinomas (14).
Compared to other PDE isoforms, human PDE4A4 is specifically associated with AIP (also called XAP2 or ARA9), a co-chaperone of HSP90 and HSC70 (26, 35). AIP has several partners, including the aryl hydrocarbon receptor (AhR), PDE4A5, PDE2A, survivin, Tom20, hepatitis B virus protein X, thyroid hormone receptor 1 (TRβ1), Epstein-Barr virus encoded nuclear antigen 3 and peroxisome proliferator-activated receptor—PPARα (36). This gene is described as a tumor suppressor gene in the pituitary (37, 38). Loss-of-function germline mutations predispose to pituitary adenomas, while reduced expression could be lead to altered epigenetic regulation via microRNAs alterations (39, 40).
AIP is expressed in GH- and PRL-cells and electron microscopy studies have identified AIP in the secretory vesicles (5). AIP is abundant in NFPAs (non-functioning pituitary adenomas), and has been shown in corticotrophinomas, although not in the secretory vesicles. However, no AIP expression has been detected in normal gonadotroph and corticotroph cells. Interestingly, it has been demonstrated that the overexpression of wild-type AIP reduces the cell proliferation in three different types of cell lines: GH3 cells, HEK293 cells, and TIG3 fibroblasts (5). These data confirmed that AIP has tumor suppressor gene properties (37, 41). Loss of interaction between AIP and PDE4A5 was seen in a β-galactosidase quantitative two-hybrid assay for pathogenic AIP mutations (R81*, Q164*, K103R, Q217*, C238Y, Y248del, R271W, V291M, and R304* (5, 23). For the K241E and R304Q variants, a borderline statistical significance was found for this interaction. For the R16H, V49M, I257V, and A229V variants, there were no clear reduction in their binding (23, 29). Many of the changes disrupting PDE4A5—AIP interaction are known to be important to the stability of the TPR structure of the AIP (42). Clinical data suggest that the R16H, V49M, and A229V may be polymorphisms while the I257V variant affects the TPR structure and clinical data would support a functional impact (29). We summarized data from variants tested in the PDE4A4/5—AIP interaction assay or with PDE4A4 or PDE4A8 immunostaining (Table 1), gathering clinical, frequency, prediction, and experimental data. We note that few variants were tested with more than one functional assay.

Table 1. Summary of data for variants tested in in the PDE4A4/5—AIP interaction assays.
There are different PDE4A4 and PDE4A8 expression patterns in somatotroph adenomas from patients with AIP mutations compared to patients with wild-type AIP (Table 2). It has been previously shown that the C-terminal part of AIP is a key for its functional effects. Mutations affecting the C-terminal end lead either to nonsense-mediated decay of the abnormal RNA (probably relevant for p.E222*), create a protein with significantly shortened half-life [as shown for p.F269_H275dup (44) and p.R304* (35)], or lose interaction with protein partners (91). AIP mutation-positive samples had significantly decreased PDE4A4 expression compared to sporadic somatotroph adenomas, suggesting that AIP mutation-positive somatotroph cells are unable to upregulate PDE4A4 expression (30).
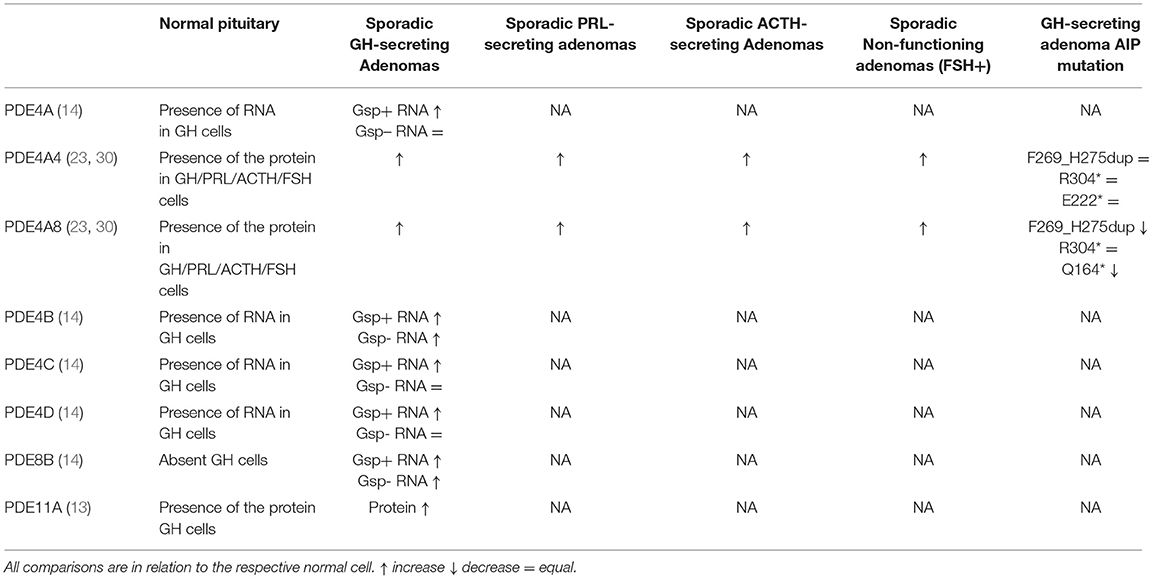
Table 2. Phosphodiesterases (PDE) isoforms and their respective protein/RNA expression in different pituitary cells types.
For PDE4A8, although no interaction with AIP has been shown due to the fact that this protein cannot be produced in vitro for the two-hybrid assay, a reduced protein expression was observed in AIP mutation-positive samples (30). These differences in PDE4A8 protein expression suggest that, similarly to PDE4A4/5, AIP may support expression or stability of PDE4A8, leading to closely regulated cAMP pathway activity (30).
The use of PDEs inhibitors, either selective or nonselective, represents an effective targeted strategy for the treatment of many human diseases, such as respiratory disorders, erectile dysfunction, prostate cancer and inflammatory diseases (92–95).
The inhibitory effect of heterologously expressed AIP on cAMP levels has not been altered by the general inhibition of phosphodiesterases (by IBMX) or the PDE4-specific inhibitor rolipram. Furthermore, the GH secretion was not altered by the use of these inhibitors (45). However, it has been shown that in rat somatotrophinoma GH3 cells, AIP regulates cAMP signaling and GH secretion independently of the AIP–PDE interaction. In the rat somatotrophinoma GH3 cells treated with forskolin, a drug that increases the cAMP levels, it was shown that the AIP overexpression could attenuate the cAMP response to the drug, even in the absence of PDE activity, while AIP knockdown activates the cAMP pathway. Although these effects are not observed in untreated cells, these results suggest that AIP may itself act as a tumor suppressor by reducing cAMP signaling (38, 45). However, GH-secreting adenomas with positive AIP mutation show reduced phosphorylation of the mitogen-activated protein kinases (MAPKs) 3 and 1 as well as reduction of phosphorylation of the cAMP response element binding protein (CREB). Also, AIP knockout causes reduced CREB phosphorylation in mouse embryonic fibroblasts although AIP knockdown rat somatotrophinoma GH3 cells do not show any of these changes on cAMP effectors (38, 96). To this point, the binding between PDE4A5-AIP does not seem to be the only regulator of this pathway.
In rat corticotroph cells, cAMP levels are related to selective activity of PDE1 (PDE1A or PDE1C) or PDE4, depending on the type and intensity of stress conditions (97). On the other hand, mouse corticotroph cell line AtT-20 with forskolin-induced elevated cAMP levels showed no response to IBMX with or without rolipram (98). Further studies are needed to clarify the possible therapeutic role of PDE manipulation in pituitary adenomas.
The cAMP pathway plays a key role in somatotroph tumorigenesis, as suggested by altered cAMP pathway in GNAS, PRKAR1A, AIP, and GPR101 mutated samples. Targeted therapies influencing this pathway may have a key role in the medical treatment of these currently often treatment-resistant conditions.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful for the support by Fundação de Amparo à Pesquisa de Minas Gerais—Fapemig (AR-O), Conselho Nacional de Desenvolvimento Científico e Tecnológico—CNPq (AR-O, MB) and the Medical Research Council UK (MK), and the NIH, USA (GB).
1. Francis SH, Blount MA, Corbin JD. Mammalian cyclic nucleotide phosphodiesterases: molecular mechanisms and physiological functions. Physiol Rev. (2011) 91:651–90. doi: 10.1152/physrev.00030.2010
2. Conti M, Beavo J. Biochemistry and physiology of cyclic nucleotide phosphodiesterases: essential components in cyclic nucleotide signaling. Annu Rev Biochem. (2007) 76:481–511. doi: 10.1146/annurev.biochem.76.060305.150444
3. Vezzosi D, Bertherat J. Phosphodiesterases in endocrine physiology and disease. Eur J Endocrinol. (2011) 165:177–88. doi: 10.1530/EJE-10-1123
4. Tian Y, Cui W, Huang M, Robinson H, Wan Y, Wang Y, et al. Dual specificity and novel structural folding of yeast phosphodiesterase-1 for hydrolysis of second messengers cyclic adenosine and guanosine 3',5'-monophosphate. Biochemistry. (2014) 53:4938–45. doi: 10.1021/bi500406h
5. Leontiou CA, Gueorguiev M, van der Spuy J, Quinton R, Lolli F, Hassan S, et al. The role of the aryl hydrocarbon receptor-interacting protein gene in familial and sporadic pituitary adenomas. J Clin Endocrinol Metab. (2008) 93:2390–401. doi: 10.1210/jc.2007-2611
6. Lania A, Mantovani G, Spada A. cAMP pathway and pituitary tumorigenesis. Ann Endocrinol. (2012) 73:73–5. doi: 10.1016/j.ando.2012.03.027
7. Peverelli E, Mantovani G, Lania AG, Spada A. cAMP in the pituitary: an old messenger for multiple signals. J Mol Endocrinol. (2014) 52:R67–77. doi: 10.1530/JME-13-0172
8. Maurice DH, Ke H, Ahmad F, Wang Y, Chung J, Manganiello VC. Advances in targeting cyclic nucleotide phosphodiesterases. Nat Rev Drug Discov. (2014) 13:290–314. doi: 10.1038/nrd4228
9. Mantovani G, Bondioni S, Ferrero S, Gamba B, Ferrante E, Peverelli E, et al. Effect of cyclic adenosine 3',5'-monophosphate/protein kinase a pathway on markers of cell proliferation in nonfunctioning pituitary adenomas. J Clin Endocrinol Metab. (2005) 90:6721–4. doi: 10.1210/jc.2005-0977
10. Pertuit M, Barlier A, Enjalbert A, Gérard C. Signalling pathway alterations in pituitary adenomas: involvement of Gsalpha, cAMP and mitogen-activated protein kinases. J Neuroendocrinol. (2009) 21:869–77. doi: 10.1111/j.1365-2826.2009.01910.x
11. Hernández-Ramírez LC, Trivellin G, Stratakis CA. Cyclic 3',5'-adenosine monophosphate (cAMP) signaling in the anterior pituitary gland in health and disease. Mol Cell Endocrinol. (2017) 463:72–86. doi: 10.1016/j.mce.2017.08.006
12. Mackenzie KF, Topping EC, Bugaj-Gaweda B, Deng C, Cheung YF, Olsen AE, et al. Human PDE4A8, a novel brain-expressed PDE4 cAMP-specific phosphodiesterase that has undergone rapid evolutionary change. Biochem J. (2008) 411:361–9. doi: 10.1042/BJ20071251
13. Peverelli E, Ermetici F, Filopanti M, Elli FM, Ronchi CL, Mantovani G, et al. Analysis of genetic variants of phosphodiesterase 11A in acromegalic patients. Eur J Endocrinol. (2009) 161:687–94. doi: 10.1530/EJE-09-0677
14. Persani L, Borgato S, Lania A, Filopanti M, Mantovani G, Conti M, et al. Relevant cAMP-specific phosphodiesterase isoforms in human pituitary: effect of Gs(alpha) mutations. J Clin Endocrinol Metab. (2001) 86:3795–800. doi: 10.1210/jcem.86.8.7779
15. Conti M, Richter W, Mehats C, Livera G, Park JY, Jin C. Cyclic AMP-specific PDE4 phosphodiesterases as critical components of cyclic AMP signaling. J Biol Chem. (2003) 278:5493–6. doi: 10.1074/jbc.R200029200
16. Houslay MD, Sullivan M, Bolger GB. The multienzyme PDE4 cyclic adenosine monophosphate-specific phosphodiesterase family: intracellular targeting, regulation, and selective inhibition by compounds exerting anti-inflammatory and antidepressant actions. Adv Pharmacol. (1998) 44:225–342. doi: 10.1016/S1054-3589(08)60128-3
17. Houslay MD, Adams DR. PDE4 cAMP phosphodiesterases: modular enzymes that orchestrate signalling cross-talk, desensitization and compartmentalization. Biochem J. (2003) 370:1–18. doi: 10.1042/bj20021698
18. Beard MB, Olsen AE, Jones RE, Erdogan S, Houslay MD, Bolger GB. UCR1 and UCR2 domains unique to the cAMP-specific phosphodiesterase family form a discrete module via electrostatic interactions. J Biol Chem. (2000) 275:10349–58. doi: 10.1074/jbc.275.14.10349
19. Bolger GB, McPhee I, Houslay MD. Alternative splicing of cAMP-specific phosphodiesterase mRNA transcripts. Characterization of a novel tissue-specific isoform, RNPDE4A8. J Biol Chem. (1996) 271:1065–71. doi: 10.1074/jbc.271.2.1065
20. Bolger GB, Rodgers L, Riggs M. Differential CNS expression of alternative mRNA isoforms of the mammalian genes encoding cAMP-specific phosphodiesterases. Gene. (1994) 149:237–44. doi: 10.1016/0378-1119(94)90155-4
21. Rena G, Begg F, Ross A, MacKenzie C, McPhee I, Campbell L, et al. Molecular cloning, genomic positioning, promoter identification, and characterization of the novel cyclic AMP-specific phosphodiesterase PDE4A10. Mol Pharmacol. (2001) 59:996–1011. doi: 10.1124/mol.59.5.996
22. Wallace DA, Johnston LA, Huston E, MacMaster D, Houslay TM, Cheung YF, et al. Identification and characterization of PDE4A11, a novel, widely expressed long isoform encoded by the human PDE4A cAMP phosphodiesterase gene. Mol Pharmacol. (2005) 67:1920–34. doi: 10.1124/mol.104.009423
23. Bolger GB, Bizzi MF, Pinheiro SV, Trivellin G, Smoot L, Accavitti MA, et al. cAMP-specific PDE4 phosphodiesterases and AIP in the pathogenesis of pituitary tumors. Endocr Relat Cancer. (2016) 23:419–31. doi: 10.1530/ERC-15-0205
24. McPhee I, Cochran S, Houslay MD. The novel long PDE4A10 cyclic AMP phosphodiesterase shows a pattern of expression within brain that is distinct from the long PDE4A5 and short PDE4A1 isoforms. Cell Signal. (2001) 13:911–8. doi: 10.1016/S0898-6568(01)00217-0
25. Bolger G, Michaeli T, Martins T, St John T, Steiner B, Rodgers L, et al. A family of human phosphodiesterases homologous to the dunce learning and memory gene product of Drosophila melanogaster are potential targets for antidepressant drugs. Mol Cell Biol. (1993) 13:6558–71. doi: 10.1128/MCB.13.10.6558
26. Bolger GB, Peden AH, Steele MR, MacKenzie C, McEwan DG, Wallace DA, et al. Attenuation of the activity of the cAMP-specific phosphodiesterase PDE4A5 by interaction with the immunophilin XAP2. J Biol Chem. (2003) 278:33351–63. doi: 10.1074/jbc.M303269200
27. Huston E, Beard M, McCallum F, Pyne NJ, Vandenabeele P, Scotland G, et al. The cAMP-specific phosphodiesterase PDE4A5 is cleaved downstream of its SH3 interaction domain by caspase-3. Consequences for altered intracellular distribution. J Biol Chem. (2000) 275:28063–74. doi: 10.1074/jbc.M906144199
28. Beard MB, Huston E, Campbell L, Gall I, McPhee I, Yarwood S, et al. In addition to the SH3 binding region, multiple regions within the N-terminal noncatalytic portion of the cAMP-specific phosphodiesterase, PDE4A5, contribute to its intracellular targeting. Cell Signal. (2002) 14:453–65. doi: 10.1016/S0898-6568(01)00264-9
29. Igreja S, Chahal HS, King P, Bolger GB, Srirangalingam U, Guasti L, et al. Characterization of aryl hydrocarbon receptor interacting protein (AIP) mutations in familial isolated pituitary adenoma families. Hum Mutat. (2010) 31:950–60. doi: 10.1002/humu.21292
30. Bizzi MF, Pinheiro SVB, Bolger GB, Schweizer JROL, Giannetti AV, Dang MN, et al. Reduced protein expression of the phosphodiesterases PDE4A4 and PDE4A8 in AIP mutation positive somatotroph adenomas. Mol Cell Endocrinol. (2018) 476:103–9. doi: 10.1016/j.mce.2018.04.014
31. Schernthaner-Reiter MH, Trivellin G, Stratakis CA. Interaction of AIP with protein kinase A (cAMP-dependent protein kinase). Hum Mol Genet. (2018). doi: 10.1093/hmg/ddy166
32. de Oliveira SK, Hoffmeister M, Gambaryan S, Müller-Esterl W, Guimaraes JA, Smolenski AP. Phosphodiesterase 2A forms a complex with the co-chaperone XAP2 and regulates nuclear translocation of the aryl hydrocarbon receptor. J Biol Chem. (2007) 282:13656–63. doi: 10.1074/jbc.M610942200
33. Martinez SE, Wu AY, Glavas NA, Tang XB, Turley S, Hol WG, et al. The two GAF domains in phosphodiesterase 2A have distinct roles in dimerization and in cGMP binding. Proc Natl Acad Sci USA. (2002) 99:13260–5. doi: 10.1073/pnas.192374899
34. Libé R, Horvath A, Vezzosi D, Fratticci A, Coste J, Perlemoine K, et al. Frequent phosphodiesterase 11A gene (PDE11A) defects in patients with Carney complex (CNC) caused by PRKAR1A mutations: PDE11A may contribute to adrenal and testicular tumors in CNC as a modifier of the phenotype. J Clin Endocrinol Metab. (2011) 96:E208–14. doi: 10.1210/jc.2010-1704
35. Hernández-Ramírez LC, Martucci F, Morgan RM, Trivellin G, Tilley D, Ramos-Guajardo N, et al. Rapid proteasomal degradation of mutant proteins is the primary mechanism leading to tumorigenesis in patients with missense AIP mutations. J Clin Endocrinol Metab. (2016) 101:3144–54. doi: 10.1210/jc.2016-1307
36. Trivellin G, Korbonits M. AIP and its interacting partners. J Endocrinol. (2011) 210:137–55. doi: 10.1530/JOE-11-0054
37. Vierimaa O, Georgitsi M, Lehtonen R, Vahteristo P, Kokko A, Raitila A, et al. Pituitary adenoma predisposition caused by germline mutations in the AIP gene. Science. (2006) 312:1228–30. doi: 10.1126/science.1126100
38. Hernández-Ramírez LC, Trivellin G, Stratakis CA. Role of phosphodiesterases on the function of Aryl Hydrocarbon Receptor-Interacting Protein (AIP) in the pituitary gland and on the evaluation of AIP gene variants. Horm Metab Res. (2017) 49:286–95. doi: 10.1055/s-0043-104700
39. Dénes J, Kasuki L, Trivellin G, Colli LM, Takiya CM, Stiles CE, et al. Regulation of aryl hydrocarbon receptor interacting protein (AIP) protein expression by MiR-34a in sporadic somatotropinomas. PLoS ONE. (2015) 10:e0117107. doi: 10.1371/journal.pone.0117107
40. Trivellin G, Butz H, Delhove J, Igreja S, Chahal HS, Zivkovic V, et al. MicroRNA miR-107 is overexpressed in pituitary adenomas and inhibits the expression of aryl hydrocarbon receptor-interacting protein in vitro. Am J Physiol Endocrinol Metab. (2012) 303:E708–19. doi: 10.1152/ajpendo.00546.2011
41. Gadelha MR, Prezant TR, Une KN, Glick RP, Moskal SF, Vaisman M, et al. Loss of heterozygosity on chromosome 11q13 in two families with acromegaly/gigantism is independent of mutations of the multiple endocrine neoplasia type I gene. J Clin Endocrinol Metab. (1999) 84:249–56.
42. Morgan RM, Hernández-Ramírez LC, Trivellin G, Zhou L, Roe SM, Korbonits M, et al. Structure of the TPR domain of AIP: lack of client protein interaction with the C-terminal α-7 helix of the TPR domain of AIP is sufficient for pituitary adenoma predisposition. PLoS ONE. (2012) 7:e53339. doi: 10.1371/journal.pone.0053339
43. Aflorei ED, Klapholz B, Chen C, Radian S, Dragu AN, Moderau N, et al. In vivo bioassay to test the pathogenicity of missense human AIP variants. J Med Genet. (2018) 55:522–9. doi: 10.1136/jmedgenet-2017-105191
44. Salvatori R, Radian S, Diekmann Y, Iacovazzo D, David A, Gabrovska P, et al. In-frame seven amino-acid duplication in AIP arose over the last 3000 years, disrupts protein interaction and stability and is associated with gigantism. Eur J Endocrinol. (2017) 177:257–66. doi: 10.1530/EJE-17-0293
45. Formosa R, Xuereb-Anastasi A, Vassallo J. Aip regulates cAMP signalling and GH secretion in GH3 cells. Endocr Relat Cancer. (2013) 20:495–505. doi: 10.1530/ERC-13-0043
46. Formosa R, Vassallo J. Aryl Hydrocarbon Receptor-Interacting Protein (AIP) N-terminus gene mutations identified in pituitary adenoma patients alter protein stability and function. Horm Cancer. (2017) 8:174–84. doi: 10.1007/s12672-017-0288-3
47. Formosa R, Farruga C, Xuereb-Anastasi A, Korbonits M, Vassallo J. Aryl hydrocarbon receptor- interacting protein: mutational analysis and functional validation in primary pituitary cell cultures. Endocr Abstr. (2010) 22:P436.
48. Cazabat L, Bouligand J, Salenave S, Bernier M, Gaillard S, Parker F, et al. Germline AIP mutations in apparently sporadic pituitary adenomas: prevalence in a prospective single-center cohort of 443 patients. J Clin Endocrinol Metab. (2012) 97:E663–70. doi: 10.1210/jc.2011-2291
49. Farrugia DJ, Agarwal MK, Pankratz VS, Deffenbaugh AM, Pruss D, Frye C, et al. Functional assays for classification of BRCA2 variants of uncertain significance. Cancer Res. (2008) 68:3523–31. doi: 10.1158/0008-5472.CAN-07-1587
50. Oriola J, Lucas T, Halperin I, Mora M, Perales MJ, Alvarez-Escolá C, et al. Germline mutations of AIP gene in somatotropinomas resistant to somatostatin analogues. Eur J Endocrinol. (2013) 168:9–13. doi: 10.1530/EJE-12-0457
51. Salvatori R, Daly AF, Quinones-Hinojosa A, Thiry A, Beckers A. A clinically novel AIP mutation in a patient with a very large, apparently sporadic somatotrope adenoma. Endocrinol Diabetes Metab Case Rep. (2014) 2014:140048. doi: 10.1530/EDM-14-0048
52. Georgitsi M, Karhu A, Winqvist R, Visakorpi T, Waltering K, Vahteristo P, et al. Mutation analysis of aryl hydrocarbon receptor interacting protein (AIP) gene in colorectal, breast, and prostate cancers. Br J Cancer. (2007) 96:352–6. doi: 10.1038/sj.bjc.6603573
53. Georgitsi M, Raitila A, Karhu A, Tuppurainen K, Mäkinen MJ, Vierimaa O, et al. Molecular diagnosis of pituitary adenoma predisposition caused by aryl hydrocarbon receptor-interacting protein gene mutations. Proc Natl Acad Sci USA. (2007) 104:4101–5. doi: 10.1073/pnas.0700004104
54. Cazabat L, Libè R, Perlemoine K, René-Corail F, Burnichon N, Gimenez-Roqueplo AP, et al. Germline inactivating mutations of the aryl hydrocarbon receptor-interacting protein gene in a large cohort of sporadic acromegaly: mutations are found in a subset of young patients with macroadenomas. Eur J Endocrinol. (2007) 157:1–8. doi: 10.1530/EJE-07-0181
55. Daly AF, Vanbellinghen JF, Khoo SK, Jaffrain-Rea ML, Naves LA, Guitelman MA, et al. Aryl hydrocarbon receptor-interacting protein gene mutations in familial isolated pituitary adenomas: analysis in 73 families. J Clin Endocrinol Metab. (2007) 92:1891–6. doi: 10.1210/jc.2006-2513
56. Buchbinder S, Bierhaus A, Zorn M, Nawroth PP, Humpert P, Schilling T. Aryl hydrocarbon receptor interacting protein gene (AIP) mutations are rare in patients with hormone secreting or non-secreting pituitary adenomas. Exp Clin Endocrinol Diabetes. (2008) 116:625–8. doi: 10.1055/s-2008-1065366
57. Guaraldi F, Salvatori R. Familial isolated pituitary adenomas: from genetics to therapy. Clin Transl Sci. (2011) 4:55–62. doi: 10.1111/j.1752-8062.2010.00254.x
58. Raitila A, Georgitsi M, Bonora E, Vargiolu M, Tuppurainen K, Mäkinen MJ, et al. Aryl hydrocarbon receptor interacting protein mutations seem not to associate with familial non-medullary thyroid cancer. J Endocrinol Invest. (2009) 32:426–9. doi: 10.1007/BF03346480
59. Tichomirowa MA, Barlier A, Daly AF, Jaffrain-Rea ML, Ronchi C, Yaneva M, et al. High prevalence of AIP gene mutations following focused screening in young patients with sporadic pituitary macroadenomas. Eur J Endocrinol. (2011) 165:509–15. doi: 10.1530/EJE-11-0304
60. Baciu I, Capatana C, Aflorei D, Botusan I, Coculescu M, Radian S. Screening of AIP mutations in young Romanian patients with sporadic pituitary adenomas. In: 15th International Congress of Endocrinology (Florence) (2012). p. P786.
61. Baciu I, Radian S, Capatina C, Botusan I, Aflorei D, Stancu C, et al. The P.R16H (C.47G > A) AIP gene variant in a case with invasive non-functioning pituitary macroadenoma and screening of a control cohort. Acta Endocrinol Bucharest. (2013) 9:97–108. doi: 10.4183/aeb.2013.97
62. Zatelli MC, Torre ML, Rossi R, Ragonese M, Trimarchi F, degli Uberti E, et al. Should aip gene screening be recommended in family members of FIPA patients with R16H variant? Pituitary. (2013) 16:238–44. doi: 10.1007/s11102-012-0409-5
63. Dinesen PT, Dal J, Gabrovska P, Gaustadnes M, Gravholt CH, Stals K, et al. An unusual case of an ACTH-secreting macroadenoma with a germline variant in the aryl hydrocarbon receptor-interacting protein (AIP) gene. Endocrinol Diabetes Metab Case Rep. (2015) 2015:140105. doi: 10.1530/EDM-14-0105
64. Iwata T, Yamada S, Mizusawa N, Golam HM, Sano T, Yoshimoto K. The aryl hydrocarbon receptor-interacting protein gene is rarely mutated in sporadic GH-secreting adenomas. Clin Endocrinol. (2007) 66:499–502. doi: 10.1111/j.1365-2265.2007.02758.x
65. Beckers A, Aaltonen LA, Daly AF, Karhu A. Familial isolated pituitary adenomas (FIPA) and the pituitary adenoma predisposition due to mutations in the aryl hydrocarbon receptor interacting protein (AIP) gene. Endocr Rev. (2013) 34:239–77. doi: 10.1210/er.2012-1013
66. Toledo RA, Sekiya T, Longuini VC, Coutinho FL, Lourenço DM, Toledo SP. Narrowing the gap of personalized medicine in emerging countries: the case of multiple endocrine neoplasias in Brazil. Clinics. (2012) 67 (Suppl. 1):3–6. doi: 10.6061/clinics/2012(Sup01)02
67. Guaraldi F, Corazzini V, Gallia GL, Grottoli S, Stals K, Dalantaeva N, et al. Genetic analysis in a patient presenting with meningioma and familial isolated pituitary adenoma (FIPA) reveals selective involvement of the R81X mutation of the AIP gene in the pathogenesis of the pituitary tumor. Pituitary. (2012) 15 (Suppl. 1):S61–7. doi: 10.1007/s11102-012-0391-y
68. Hernández-Ramírez LC, Gabrovska P, Dénes J, Stals K, Trivellin G, Tilley D, et al. Landscape of familial isolated and young-onset pituitary adenomas: prospective diagnosis in AIP mutation carriers. J Clin Endocrinol Metab. (2015) 100:E1242–54. doi: 10.1210/jc.2015-1869
69. Caimari F, Hernández-Ramírez LC, Dang MN, Gabrovska P, Iacovazzo D, Stals K, et al. Risk category system to identify pituitary adenoma patients with. J Med Genet. (2018) 55:254–60. doi: 10.1136/jmedgenet-2017-104957
70. Stratakis CA, Tichomirowa MA, Boikos S, Azevedo MF, Lodish M, Martari M, et al. The role of germline AIP, MEN1, PRKAR1A, CDKN1B and CDKN2C mutations in causing pituitary adenomas in a large cohort of children, adolescents, and patients with genetic syndromes. Clin Genet. (2010) 78:457–63. doi: 10.1111/j.1399-0004.2010.01406.x
71. Cai F, Zhang YD, Zhao X, Yang YK, Ma SH, Dai CX, et al. Screening for AIP gene mutations in a Han Chinese pituitary adenoma cohort followed by LOH analysis. Eur J Endocrinol. (2013) 169:867–84. doi: 10.1530/EJE-13-0442
72. Georgitsi M, De Menis E, Cannavò S, Mäkinen MJ, Tuppurainen K, Pauletto P, et al. Aryl hydrocarbon receptor interacting protein (AIP) gene mutation analysis in children and adolescents with sporadic pituitary adenomas. Clin Endocrinol. (2008) 69:621–7. doi: 10.1111/j.1365-2265.2008.03266.x
73. Daly AF, Tichomirowa MA, Petrossians P, Heliövaara E, Jaffrain-Rea ML, Barlier A, et al. Clinical characteristics and therapeutic responses in patients with germ-line AIP mutations and pituitary adenomas: an international collaborative study. J Clin Endocrinol Metab. (2010) 95:E373–83. doi: 10.1210/jc.2009-2556
74. Jennings JE, Georgitsi M, Holdaway I, Daly AF, Tichomirowa M, Beckers A, et al. Aggressive pituitary adenomas occurring in young patients in a large Polynesian kindred with a germline R271W mutation in the AIP gene. Eur J Endocrinol. (2009) 161:799–804. doi: 10.1530/EJE-09-0406
75. De Sousa SM, McCabe MJ, Wu K, Roscioli T, Gayevskiy V, Brook K, et al. Germline variants in familial pituitary tumour syndrome genes are common in young patients and families with additional endocrine tumours. Eur J Endocrinol. (2017) 176:635–44. doi: 10.1530/EJE-16-0944
76. Karaca Z, Taheri S, Tanriverdi F, Unluhizarci K, Kelestimur F. Prevalence of AIP mutations in a series of Turkish acromegalic patients: are synonymous AIP mutations relevant? Pituitary. (2015) 18:831–7. doi: 10.1007/s11102-015-0659-0
77. Radian S, Diekmann Y, Gabrovska P, Holland B, Bradley L, Wallace H, et al. Increased population risk of AIP-related acromegaly and gigantism in Ireland. Hum Mutat. (2017) 38:78–85. doi: 10.1002/humu.23121
78. Bell DR, Poland A. Binding of Aryl Hydrocarbon Receptor (AhR) to AhR-interacting protein. J Biol Chem. (2000) 275:36407–14. doi: 10.1074/jbc.M004236200
79. Occhi G, Jaffrain-Rea ML, Trivellin G, Albiger N, Ceccato F, De Menis E, et al. The R304X mutation of the aryl hydrocarbon receptor interacting protein gene in familial isolated pituitary adenomas: mutational hot-spot or founder effect? J Endocrinol Invest. (2010) 33:800–5. doi: 10.1007/BF03350345
80. Chahal HS, Stals K, Unterländer M, Balding DJ, Thomas MG, Kumar AV, et al. AIP mutation in pituitary adenomas in the 18th century and today. N Engl J Med. (2011) 364:43–50. doi: 10.1056/NEJMoa1008020
81. Cuny T, Pertuit M, Sahnoun-Fathallah M, Daly A, Occhi G, Odou MF, et al. Genetic analysis in young patients with sporadic pituitary macroadenomas: besides AIP don't forget MEN1 genetic analysis. Eur J Endocrinol. (2013) 168:533–41. doi: 10.1530/EJE-12-0763
82. de Lima DS, Martins CS, Paixao BM, Amaral FC, Colli LM, Saggioro FP, et al. SAGE analysis highlights the putative role of underexpression of ribosomal proteins in GH-secreting pituitary adenomas. Eur J Endocrinol. (2012) 167:759–68. doi: 10.1530/EJE-12-0760
83. Niyazoglu M, Sayitoglu M, Firtina S, Hatipoglu E, Gazioglu N, Kadioglu P. Familial acromegaly due to aryl hydrocarbon receptor-interacting protein (AIP) gene mutation in a Turkish cohort. Pituitary. (2014) 17:220–6. doi: 10.1007/s11102-013-0493-1
84. Williams F, Hunter S, Bradley L, Chahal HS, Storr HL, Akker SA, et al. Clinical experience in the screening and management of a large kindred with familial isolated pituitary adenoma due to an aryl hydrocarbon receptor interacting protein (AIP) mutation. J Clin Endocrinol Metab. (2014) 99:1122–31. doi: 10.1210/jc.2013-2868
85. Pardi E, Marcocci C, Borsari S, Saponaro F, Torregrossa L, Tancredi M, et al. Aryl hydrocarbon receptor interacting protein (AIP) mutations occur rarely in sporadic parathyroid adenomas. J Clin Endocrinol Metab. (2013) 98:2800–10. doi: 10.1210/jc.2012-4029
86. Occhi G, Trivellin G, Ceccato F, De Lazzari P, Giorgi G, Demattè S, et al. Prevalence of AIP mutations in a large series of sporadic Italian acromegalic patients and evaluation of CDKN1B status in acromegalic patients with multiple endocrine neoplasia. Eur J Endocrinol. (2010) 163:369–76. doi: 10.1530/EJE-10-0327
87. Vargiolu M, Fusco D, Kurelac I, Dirnberger D, Baumeister R, Morra I, et al. The tyrosine kinase receptor RET interacts in vivo with aryl hydrocarbon receptor-interacting protein to alter survivin availability. J Clin Endocrinol Metab. (2009) 94:2571–8. doi: 10.1210/jc.2008-1980
88. Mothojakan N, Ferrau F, Dang M, Barlier A, Chanson P, Occhi G. Polymorphism or mutation? - the role of the R304Q missense AIP mutation in the predisposition to pituitary adenoma. Endocr Abstr Biosci. (2016) 44:P167. doi: 10.1530/endoabs.44.P167
89. Imran SA, Aldahmani KA, Penney L, Croul SE, Clarke DB, Collier DM, et al. Unusual AIP mutation and phenocopy in the family of a young patient with acromegalic gigantism. Endocrinology Diabetes Metab Case Rep. (2018) 2018:17-0092. doi: 10.1530/EDM-17-0092
90. Richards S, Aziz N, Bale S, Bick D, Das S, Gastier-Foster J, et al. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American college of medical genetics and genomics and the association for molecular pathology. Genet Med. (2015) 17:405–24. doi: 10.1038/gim.2015.30
91. Kazlauskas A, Poellinger L, Pongratz I. Two distinct regions of the immunophilin-like protein XAP2 regulate dioxin receptor function and interaction with hsp90. J Biol Chem. (2002) 277:11795–801. doi: 10.1074/jbc.M200053200
92. Bolger GB. The PDE4 cAMP-specific phosphodiesterases: targets for drugs with antidepressant and memory-enhancing action. Adv Neurobiol. (2017) 17:63–102. doi: 10.1007/978-3-319-58811-7_4
93. Mokra D, Mokry J, Matasova K. Phosphodiesterase inhibitors: potential role in the respiratory distress of neonates. Pediatr Pulmonol. (2018) 53:1318–25. doi: 10.1002/ppul.24082
94. Moustafa F, Feldman SR. A review of phosphodiesterase-inhibition and the potential role for phosphodiesterase 4-inhibitors in clinical dermatology. Dermatol Online J. (2014) 20:22608. Available online at: https://escholarship.org/uc/item/2hx1m6kv
95. Hamilton TK, Hu N, Kolomitro K, Bell EN, Maurice DH, Graham CH, et al. Potential therapeutic applications of phosphodiesterase inhibition in prostate cancer. World J Urol. (2013) 31:325–30. doi: 10.1007/s00345-012-0848-7
96. Tuominen I, Heliövaara E, Raitila A, Rautiainen MR, Mehine M, Katainen R, et al. AIP inactivation leads to pituitary tumorigenesis through defective Gαi-cAMP signaling. Oncogene. (2015) 34:1174–84. doi: 10.1038/onc.2014.50
97. Ang KL, Antoni FA. Functional plasticity of cyclic AMP hydrolysis in rat adenohypophysial corticotroph cells. Cell Signal. (2002) 14:445–52. doi: 10.1016/S0898-6568(01)00267-4
Keywords: phosphodiesterases, cAMP pathway, pituitary, AIP (Aryl hydrocarbon receptor interacting protein), acromegaly, gigantism
Citation: Bizzi MF, Bolger GB, Korbonits M and Ribeiro-Oliveira A Jr (2019) Phosphodiesterases and cAMP Pathway in Pituitary Diseases. Front. Endocrinol. 10:141. doi: 10.3389/fendo.2019.00141
Received: 02 October 2018; Accepted: 15 February 2019;
Published: 19 March 2019.
Edited by:
Xianquan Zhan, Central South University, ChinaReviewed by:
Kjetil Taskén, Oslo University Hospital, NorwayCopyright © 2019 Bizzi, Bolger, Korbonits and Ribeiro-Oliveira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antonio Ribeiro-Oliveira Jr., YW50b25pb3JvanJAZ21haWwuY29t
†These authors have contributed equally to this work
‡Present Address: Graeme B. Bolger, BZI Pharma LLC, Birmingham, AL, United States
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.