 Emily E. Morin
Emily E. Morin Xiang-An Li
Xiang-An Li Anna Schwendeman
Anna Schwendeman
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Endocrinol. , 30 November 2018
Sec. Cancer Endocrinology
Volume 9 - 2018 | https://doi.org/10.3389/fendo.2018.00715
This article is part of the Research Topic Cholesterol and Oxysterols as Signal Molecules in Human Pathophysiology and Cancer: Implications for New Therapeutic Strategies View all 9 articles
High-density lipoprotein (HDL) have long been studied for their protective role against cardiovascular diseases, however recently relationship between HDL and cancer came into focus. Several epidemiological studies have shown an inverse correlation between HDL-cholesterol (HDL-C) and cancer risk, and some have even implied that HDL-C can be used as a predictive measure for survival prognosis in for specific sub-population of certain types of cancer. HDL itself is an endogenous nanoparticle capable of removing excess cholesterol from the periphery and returning it to the liver for excretion. One of the main receptors for HDL, scavenger receptor type B-I (SR-BI), is highly upregulated in endocrine cancers, notably due to the high demand for cholesterol by cancer cells. Thus, the potential to exploit administration of cholesterol-free reconstituted or synthetic HDL (sHDL) to deplete cholesterol in endocrine cancer cell and stunt their growth of use chemotherapeutic drug loaded sHDL to target payload delivery to cancer cell has become increasingly attractive. This review focuses on the role of HDL and HDL-C in cancer and application of sHDLs as endocrine cancer therapeutics.
Endocrine cancers are defined as those affecting the hormone secreting tissues of our body, including cancers of the adrenal, thyroid, parathyroid, prostate, pancreatic, and reproductive tissues. A rare subset of endocrine cancers, called neuroendocrine tumors (NETs), are neoplasms originating in endocrine tissue that migrate to form hormone-secreting tumors in other organs of the body, including intestine, lung, and pancreas (1). While specific molecular signatures may vary among the different types of endocrine cancers, they all share a common modality which is essential for tumor cell proliferation and overall survival: a high demand for cholesterol (2–4).
Cholesterol is a precursor molecule for steroid synthesis and bile acid production, making it essential for hormone production by endocrine tissue (5). Cholesterol is also an important component of cellular membranes, offering structure and rigidity to the plasma membrane as well as clustering with sphingolipids and glycerophospholipids to form highly-stable membrane microdomains or “lipid rafts” that host a number of proteins and lipids involved in key cell signaling pathways (6). In cancer, rapid cellular division is needed for the growth and survival of the tumor. Hence, a large demand for cholesterol is needed to facilitate the rapid formation of new membranes (3). Endocrine cancers in particular, display an even higher demand for cholesterol due to increased hormone and steroid production by these cells (7, 8).
A hydrophobic molecule, cholesterol has very poor aqueous solubility and thus is transported throughout the body by lipoproteins. Under normal conditions, HDL is a key participant in the reverse cholesterol transport (RCT) pathway, a process by which excess cholesterol from peripheral tissue is taken up by HDL and transported back to the liver for secretion in the bile or for redistribution to endocrine tissue for steroid production. Historically, HDL-C has been the focus of lipid metabolism modulating therapeutics for cardiovascular diseases, as high HDL-C or “the good cholesterol” and low LDL-C “bad cholesterol” have been well established as markers of cardiovascular health. Specifically, nascent HDL has the ability to reduce the burden of atherosclerosis by depleting foam-cell macrophages of their cholesterol and reducing inflammation and oxidation in the surrounding atheroma environment (9, 10). In this review, we will summarize what is known about the association between HDL-C levels and cancer and examine the utility of reconstituted or synthetic HDL as a potential therapeutic and drug delivery vehicle for endocrine cancers.
HDL is an endogenous, nanosized particle composed apolipoproteins, and lipids (11). Naturally, these particles range in shape, size, density, and charge depending on their lipid composition, protein cargo, and degree of maturation (11). The main protein component of HDL, apolipoprotein A-I (ApoA-I), is initially synthesized in the liver where it is subsequently secreted into the circulation. Once secreted, ApoA-I picks up a small amount of lipid to form pre-β HDL particles. These nascent, cholesterol-poor discs can then further interact with cholesterol-rich cells of the periphery to take up and deliver that cholesterol back to the liver where it is taken up via scavenger receptor type B-I (SR-BI) for secretion or further processing. Once picked up by HDL, cholesterol is esterified by lecithin:acyl cholesterol transferase (LCAT) to form cholesterol ester. Cholesterol ester is then buried within HDL's hydrophobic lipid core, inducing the maturation and formation of larger, spherical HDL particles. Spherical HDL particles contain not only ApoA-I, but also ApoE, which facilitates the growing load of CE into the hydrophobic core since ApoA-I can only facilitate a limited amount of CE in the HDL core. ApoE is also useful in that it is a substrate for low-density lipoprotein receptor (LDLR) and can deliver HDL cargo to hepatic LDLR for biliary excretion or to endocrine tissue expressing LDLR or SR-BI for use in steroid production (12, 13).
HDL is highly heterogeneous and is present in a variety of different forms depending on its size, shape, density, and lipid/protein composition. This is a result of HDL remodeling, which is a continuous process involving several endogenous enzymes (14). Put simply, HDL can be continually and reversibly recycled between lipid-poor apoA1, discoidal HDL, and small/large/larger spherical HDLs. These subsets of HDL are classified into two groups, HDL2 and HDL3, based on their densities (15, 11). HDL2 is lipid-rich and less dense (1.063–1.125 g/mL) than its HDL3 counterpart, which is dense (1.125–1.21 g/mL) protein-rich in comparison (11). Both HDL2 and HDL3 can be further divided into 2 and 3 subclasses, respectively, based on their size; HDL3 ranges in size from roughly 7–9 nm in diameter while HDL2 ranges from about 9–12 nm (11). To further complicate things, HDL can also be classified according to its surface charge and shape. Spherical, more neutral HDL particles are classified as α-HDL, while nascent, discoidal HDL particles, known as β-HDL, are poorly lipidated and more negative in overall charge.
In addition to the existing variety of subpopulations in healthy individuals, HDL particle makeup can vary significantly among patients of different disease states (16). Particularly, recent studies have identified changes in the diverse proteome of HDL particles in the various disease states (17, 18). While ApoA-I is the main protein in HDL, other proteins including ApoA-II, ApoC, paraoxanase (PON), ApoM, and serum amyloid A (SAA) have been identified and can be altered under disease conditions (19–21). The lipid composition of HDL particles can also vary with disease (22), and chronic changes in the HDL lipidome have been attributed to the high inflammatory state of various diseases, including the presence of lysophosphatidic acid (LPA), a phospholipid implicated in the progression of several endocrine cancers (23–25). Under such conditions, including atherosclerosis and lupus, HDL isolated from patients is said to be dysfunctional or proinflammatory, and its abilities to carry out cholesterol efflux and exert anti-inflammatory properties are lost (26–31). Similarly, studies have shown that HDL can promote breast cancer metastasis, which is attributed to the alterations in HDL's lipid and protein compositions under inflammatory and oxidative conditions (32, 33).
A number of observational studies and retrospective study analyses have shown that plasma HDL-C and ApoA-I levels are significantly reduced in cancer patients, including those with breast, ovarian, colon, prostate, and pancreatic carcinomas (34–50). These studies are summarized in Table 1. A number of studies also sought to investigate the predictive power of HDL-C or ApoA-I levels in subsets of cancers and found that, when combined with other traditional cancer biomarkers cancer antigen 125 (CA125) and transthyretin (TTR), either ApoA-I or HDL-C levels significantly increased the power of these panels to predict patient prognosis (52, 54–58). In some cases, however, there were no significant associations between HDL-C, ApoA-I, and cancer risk (51). This is likely due to differences in study design and evaluation as well as the methods used to quantify HDL-C and ApoA-I. For example, direct measurements of HDL-C are generally performed by mass precipitation and can be confounded by the presence of ApoE and other proteins. Other methods directly measure HDL particles via size and charge separation using density gradient ultracentrifugation, gel filtration, high performance liquid chromatography (HPLC), and nuclear magnetic resonance (NMR) among others. Each of these techniques has its drawbacks, and is generally bias toward one or more subpopulation of HDL or risks chemical modification of the particles during sample preparation (61). In addition other confounding factors such as lifestyle factors, co-morbidities, and physiological factors (i.e., pre- vs. post-menopausal women), all contributed to heterogeneity of the results since most of the analyses were done retrospectively using the existing body of publicly available clinical trial data. On the other hand, there are studies describing positive correlations between HDL-C and cancer risk, namely in breast cancers (60). However, given the high heterogeneity in HDL proteome, lipidome, and subclass distribution between patients in different disease settings, it is reasonable that such variability exists between studies. While the verdict is still out on the utility of HDL-C and ApoA-I as predictive biomarkers in cancers, there is clearly a role for HDL in this complex disease which will be discussed in more detail to follow.
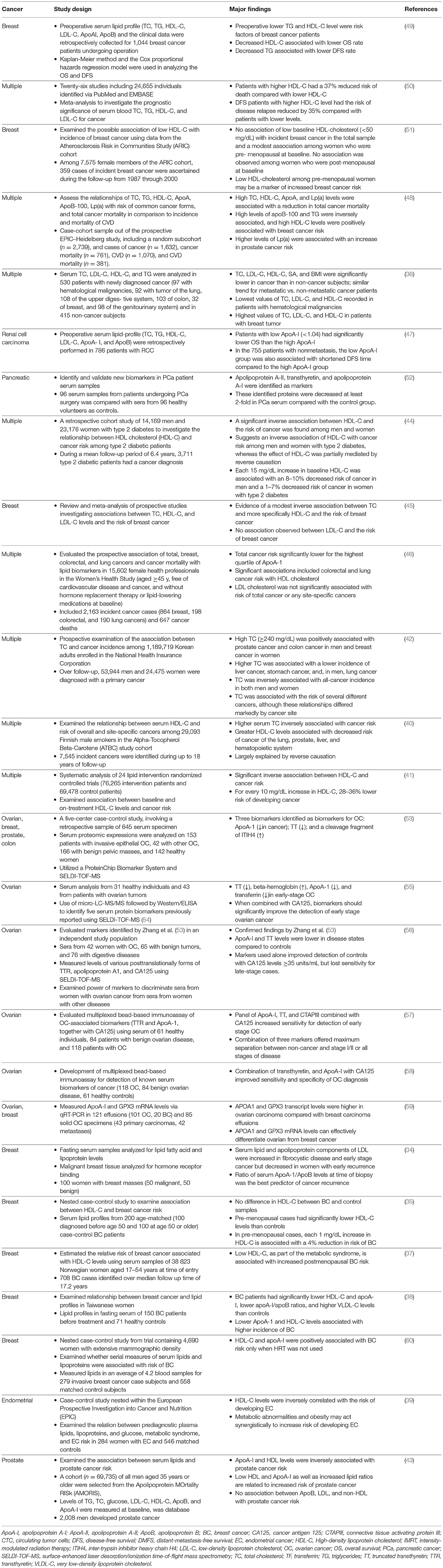
Table 1. Clinical relationships between HDL-C and endocrine cancers.
Whether decreased levels of HDL-C are a causal or consequential factor to cancer progression is yet to be elucidated, however we are logically drawn to the latter. It is known that cancer cells, in particular prostate, adrenal, and breast cancer cells, highly express the SR-BI on their plasma membrane (62–64). Because of their high-demand for cholesterol, cancer cell upregulation of SR-BI is likely a survival mechanism to increase HDL-C recruitment and, thus, increase cholesterol uptake needed for proliferation and hormone production while consequently decreasing circulating HDL-C. That being said, this argument could also be used to explain why, in some reports, HDL-C is associated with an increased risk of cancer, as it continues to provide additional cholesterol to and fuel the growth of the tumor. Regardless, we can take away several key points from these findings: (i) SR-BI is overexpressed in cancer cells, (ii) HDL-C levels are significantly affected by the presence and development of cancer, and (iii) the high affinity between SR-BI and HDL facilitates the transport of cholesterol to/from HDL and the cancer cell. In addition, HDL is known to have potent antioxidant activity and both endogenous and reconstituted HDL particles were shown to inhibit oxidative-stress induced proliferation of pancreatic cells in vitro (65). And although the details of the epidemiology can be disputed, there are clear opportunities for therapeutic intervention by utilizing the HDL/SR-BI axis, of which will be discussed below.
As previously mentioned, the main focus of HDL research over the past several decades has been surrounding its role in cardiovascular disease. Because of its role in facilitating RCT, several “HDL mimetics” have been developed and tested clinically in humans for their ability to reduce the burden of atherosclerosis and number of events following an acute coronary event (66–69). These HDL-mimicking particles, termed reconstituted HDL (rHDL) or synthetic HDL (sHDL), are cholesterol-free HDL particles prepared from plasma purified or recombinantly expressed ApoA-I or short synthetic ApoA-I mimetic peptides complexed with phospholipids. Since they lack cholesterol, these “empty” particles are highly effective in effluxing cholesterol from lipid-laiden cells both in vitro and in vivo (9, 70, 71). In addition to their augmented efflux capacity, these particles offer a natural ability to target SR-BI-expressing cells. When combined, the ability of rHDL/sHDL to deplete cellular cholesterol, target SR-BI expressing cells, along with the biocompatibility of the individual components and proven clinical safety make the application of sHDL for cancer therapy increasingly attractive.
The concept of utilizing sHDL and ApoA-I mimetic peptides as a cholesterol depletion therapy has been around for decades, but focused primarily in the context of cardiovascular diseases. In fact, several sHDL therapies have been developed and tested in various stages of human clinical trials (66, 69, 72). The purpose of such sHDL infusion therapies was to efflux cholesterol and reduce plaque size and vulnerability following an initial coronary event, in order to decrease the occurance of secondary events. Early sHDL clinical trials utilized lipid-free ApoA-I protein or mimetic peptides, such as ApoA-I milano, D-4F, and L-4F, however it was shown that the naked proteins and peptides themselves had a very short plasma half-life, and their pharmacological effect suffered as a consequence (73, 74). Formulation of peptide or full-length ApoA-I protein with phospholipid, forming sHDL, was shown to markedly improve plasma half-life and thus its overall therapeutic effect (75). Moreover, studies have shown that the phospholipid component of sHDL therapies is a driving determinant of the overall pharmacokinetic and pharmacodynamic effect (71).
Measurable improvements in the pharmacokinetic and pharmacodynamic effects of sHDL therapies has led to their progression from bench to bedside in both early and late stage clinical trials. Such sHDL products include peptide-based sHDLs, including ETC-642 (22A peptide/dipalmitoylphosphatidylcholine/sphingomyelin), and ApoA-I protein based sHDLs, including ETC-216 (recombinant ApoA-I/palmitoyloleoylphosphatidylcholine) and CER-001 (recombinant ApoA-I/sphingomyelin/dipalmitoylphosphatidylglycerol), among others. These products were all shown to be safe at high doses of up to 100 mg/kg in humans and possess potent cholesterol efflux abilities (76, 77). More recently CSL-112, reconstituted ApoA-I/soybean phosphatidylcholine, has advanced to a 17,000 patient Phase III clinical trial after showing promising ability to reduce atheroma burden and decrease secondary coronary events in earlier trials (78–80). Given their proven clinical safety and ability to facilitate cholesterol removal, sHDL products could be easily translated for use as cholesterol depleting therapies in cancer.
Given the dependence of endocrine cancers on cholesterol, cholesterol-targeting therapies have gained increasing attention. One approach is to directly deplete cholesterol from cells using cholesterol scavenging therapies. In addition to cholesterol being essential for the formation of new membranes during cell division, it is also vital for the formation of lipid-rafts in the plasma membrane. These lipid rafts are rich in cholesterol and sphingolipids and house many proteins and transporters involved in key signaling pathways, including the Akt signaling pathway implicated in the migration, proliferation, and survival of cancer cells (81). By depleting cholesterol from cells, lipid rafts are disrupted and the proteins they house internalized, drastically reducing the cell's ability to carry out its functions and often triggering cell death (4). In cancer cell lines, treatment with cyclodextrins induced marked cell death, and that cells with a higher abundance of lipid rafts were more susceptible to such treatments (82–84). More recently, Taylor et al. showed that HAC15 adrenal carcinoma cells treated with ETC-642, a clinically tested sHDL, displayed marked reduction in cellular cholesterol levels in addition to inhibition of aldosterone, cortisol, and androstenedione production (85). Thus, the application of cholesterol-depleting therapies, namely sHDL, for endocrine cancer deserves further investigation.
Several recent studies have investigated the use of HDL-mimetics, including sHDL, ApoA-I protein, and ApoA-I mimetic peptides, for cancer treatment, outlined in Table 2. In addition, treatment of cells with both sHDL and chemotherapeutic drugs was able to reduce the overall effective dose (91). Other studies utilizing ApoA-I protein or mimetic peptides L-4F, L-5F, and D-4F have shown that treatment of tumor-bearing mice with either protein or peptide can reduce both tumor volume and angiogenesis in tumor tissues when compared to control mice (87–90, 92–94). For example, in a mouse model of ovarian cancer, Gao et. al. show that L-4F peptide, when administered subcutaneously at 10 mg/kg/day for 3 weeks, could reduce overall angiogenesis and vessel number within the tumor tissue, which was connected to a decrease in expression levels of hypoxia inducible factor-1α (HIF-1α) (88). In a similar study, they also show that L-5F peptide could exert similar anti-angiogensis effects and led to decreased levels of serum vascular endothelial growth factor (VEGF) (87). In a separate study utilizing a similar ovarian cancer mouse model, Su et. al. demonstrated that both L-4F and L-5F peptides could decrease the overall volume of tumors in both flank and intraperitoneal tumors when given 10 mg/kg/day over the course of 5 or 9 weeks, respectively (86). They postulate that the antitumorogenic effect could be related to peptides' abilities to reduce circulating levels of LPA, and they were found to have significantly greater binding affinity for LPA when compared to full length ApoA-I protein. In a study by Peng et. al. L-4F peptide given at 10 mg/kg/day for 1 week was able to not only reduce size and weight of H7 pancreatic tumors in mice, but also significantly reduce several markers of inflammation within the tissue as well (90).
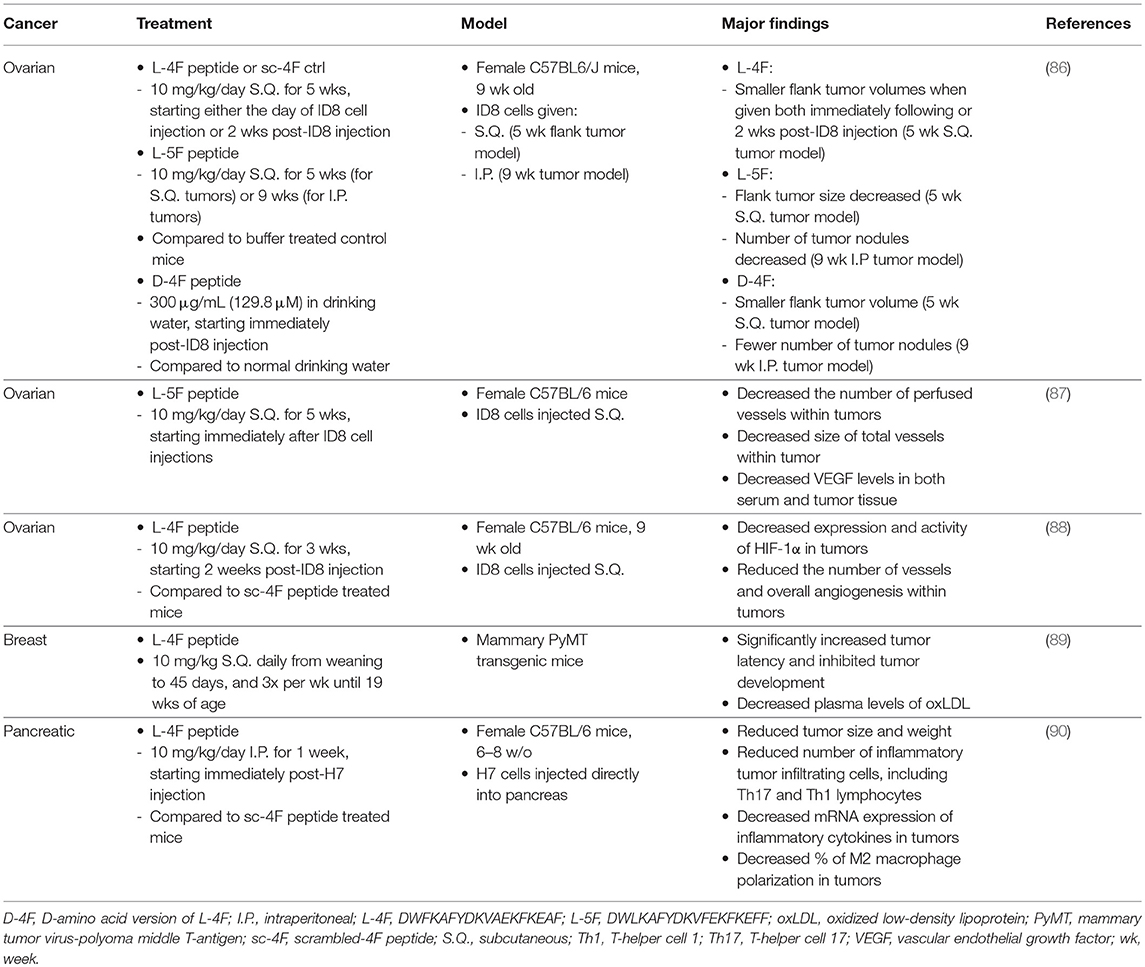
Table 2. Experimental studies utilizing HDL for endocrine cancer therapy.
While the above studies also included extensive screening of ApoA-I and mimetic peptides for viability and anti-proliferative activity in a broad range of cancer cell lines in vitro, there are also reports describing the ability of HDL to induce proliferation, migration, and survival in cancer cell cultures (32, 33, 95, 96). Consistent with previously mentioned clinical findings showing a positive association between HDL-C and cancer risk, these studies support the notion that HDL-C may promote the progression of cancer by supplying the tumor cells with their increasing demand for cholesterol. However, a distinction should be made between the epidemiology of HDL and cancer and the utility of HDL in cancer treatment: namely, that the use of HDL in cancer therapy referred to in this review involves the administration of “empty” cholesterol-free particles. These particles are the nascent, discoidal HDL particles with high cholesterol efflux activity as proven both in basic and clinical research. Of course, studies utilizing plasma purified HDLs should be considered differently. Plasma HDLs contain a variety of different components, including signaling lipids responsible for many of HDL's pro-angiogenic and Akt-activating properties (97, 22), and namely cholesterol capable of being delivered to cells (98, 99). While such studies are integral to understanding the role of endogenous HDL in cancer pathogenesis, they should not be confused with therapeutic implications utilizing sHDL or mimetic peptides with a defined molecular makeup and superior cholesterol efflux capacity.
Given the very poor solubility of many chemotherapeutic drugs, the hydrophobic lipid core of HDL presents an attractive environment and alternative strategy for delivery and formulation of this class of drugs. Not only is it possible to lower the overall dose of drug given by improving its solubility, but the SR-BI targeting ability of these sHDL nanoparticles affords the additional benefit of site-specific, cytosolic drug delivery to SR-BI over-expressing tumor cells while subsequently reducing systemic toxicity (94, 72, 100). Others have, with varying success, shown anti-tumorigenic by introducing HDL surface modifications to augment the targeting capacity and to extend particle half-life (101). The use of HDL-mimetics for targeted drug delivery has been extensively reviewed elsewhere (102, 72) and is beyond the scope of this review, however, its importance and growing relevance warrant mentioning.
Decades of epidemiological evidence suggests that, notably -C, plays a role in the incidence and progression of cancer. Whether or not this role is causal or consequential, or whether the risk association is positive or negative under specific conditions is still left for debate. Despite, we know from years of clinical and basic cardiovascular research that is an intimate player in the RCT process and has specific and potent cholesterol efflux ability both in vitro and in vivo. We also know that cholesterol is a vital resource for cancer cells, which require a constant supply to maintain and facilitate their rapid proliferation and overall survival. Endocrine cancers, in particular, are at an increased demand for cholesterol given their additional need for steroid production making them even more susceptible to cholesterol depletion interventions and targeting by due to upregulation of SR-BI.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
This publication was made possible by Grant Number R01GM113832 (to AS and X-AL) from NIGMS/NIH and NIH T32-HL125242 (EM). Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NIGMS or NIH.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Klimstra DS, Modlin IR, Coppola D, Lloyd RV, Suster S. The pathologic classification of neuroendocrine tumors: a review of nomenclature, grading, and staging systems. Pancreas (2010) 39:707–12. doi: 10.1097/MPA.0b013e3181ec124e
2. Hager MH, Solomon KR, Freeman MR. The role of cholesterol in prostate cancer. Curr Opin Clin Nutr Metab Care (2006) 9:379–85. doi: 10.1097/01.mco.0000232896.66791.62
3. Cruz PM, Mo H, McConathy WJ, Sabnis N, Lacko AG. The role of cholesterol metabolism and cholesterol transport in carcinogenesis: a review of scientific findings, relevant to future cancer therapeutics. Front Pharmacol. (2013). 4:119. doi: 10.3389/fphar.2013.00119
4. Beloribi-Djefaflia S, Vasseur S, Guillaumond F. Lipid metabolic reprogramming in cancer cells. Oncogenesis 5:e189. doi: 10.1038/oncsis.2015.49
5. Hu J, Zhang Z, Shen WJ, Azhar S. Cellular cholesterol delivery, intracellular processing and utilization for biosynthesis of steroid hormones. Nutri Metab. (2010) 7:47. doi: 10.1186/1743-7075-7-47
6. Pike LJ. Lipid rafts: bringing order to chaos. J Lipid Res. (2003) 44:655–67. doi: 10.1194/jlr.R200021-JLR200
7. Foster PA. Steroid metabolism in breast cancer. Minerva Endocrinol. (2008) 33:27–37. Available online at: http://cebp.aacrjournals.org/content/13/7/1185
8. Purohit A, Foster PA. Steroid sulfatase inhibitors for estrogen- and androgen-dependent cancers. J Endocrinol. (2012) 212:99–110. doi: 10.1530/JOE-11-0266
9. von Eckardstein A, Nofer JR, Assmann G. High density lipoproteins and arteriosclerosis: role of cholesterol efflux and reverse cholesterol transport. Arterioscl Thromb Vasc Biol. (2001) 21:13–27. doi: 10.1161/01.ATV.21.1.13
10. Rye KA, Barter PJ. Cardioprotective functions of HDLs. J Lipid Res. (2014) 55:168–79. doi: 10.1194/jlr.R039297
11. Kontush A, Lindahl M, Lhomme M, Calabresi L, Chapman MJ, Davidson WS. Structure of HDL: particle subclasses and molecular components. In: von Eckardstein A, Kardassis D, editors. High Density Lipoproteins, vol. 224. Cham; Heidelberg; New York, NY; Dordrecht; London: Springer International Publishing (2015). p. 3–51.
12. Lund-Katz S, Phillips MC. High density lipoprotein structure–function and role in reverse cholesterol transport. In: Harris JR, editor. Cholesterol Binding and Cholesterol Transport Proteins, vol. 51. Netherlands: Springer (2010). p. 183–227.
13. Favari E, Chroni A, Tietge UJ, Zanotti I, Escolà-Gil JC, Bernini F. Cholesterol Efflux and Reverse Cholesterol Transport. In: von Eckardstein A, Kardassis D, editors. High Density Lipoproteins, vol. 224. Cham; Heidelberg; New York, NY; Dordrecht; London: Springer International Publishing. (2015). p. 181–206.
14. Zannis VI, Fotakis P, Koukos G, Kardassis D, Ehnholm C, Jauhiainen M, et al. HDL biogenesis, remodeling, and Catabolism. In: von Eckardstein A, Kardassis D, editors. High Density Lipoproteins, vol. 224. Cham; Heidelberg; New York, NY; Dordrecht; London: Springer International Publishing (2015). p. 53–111.
15. Kontush A, Chapman MJ. High-Density Lipoproteins: Structure, Metabolism, Function and Therapeutics. Hoboken, NJ: John Wiley & Sons (2011).
16. Kratzer A, Giral H, Landmesser U. High-density lipoproteins as modulators of endothelial cell functions: alterations in patients with coronary artery disease. Cardiovasc Res. (2014) 103:350–61. doi: 10.1093/cvr/cvu139
17. Heinecke JW. The HDL proteome: a marker–and perhaps mediator–of coronary artery disease. J Lipid Res. (2009) 50(Suppl.):S167–71. doi: 10.1194/jlr.R800097-JLR200
18. Alwaili K, Bailey D, Awan Z, Bailey SD, Ruel I, Hafiane A, et al. The HDL proteome in acute coronary syndromes shifts to an inflammatory profile. Biochim Biophys Acta (2012) 1821:405–15. doi: 10.1016/j.bbalip.2011.07.013
19. Van Lenten BJ, Hama SY, de Beer FC, Stafforini DM, McIntyre TM, Prescott SM, et al. Anti-inflammatory HDL becomes pro-inflammatory during the acute phase response. Loss of protective effect of HDL against LDL oxidation in aortic wall cell cocultures. J Clin Invest. (1995) 96:2758–67. doi: 10.1172/JCI118345
20. Shah AS, Tan L, Long JL, Davidson WS. Proteomic diversity of high density lipoproteins: our emerging understanding of its importance in lipid transport and beyond. J Lipid Res. (2013) 54:2575–85. doi: 10.1194/jlr.R035725
21. Birner-Gruenberger R, Schittmayer M, Holzer M, Marsche G. Understanding high-density lipoprotein function in disease: recent advances in proteomics unravel the complexity of its composition and biology. Prog Lipid Res. (2014) 56:36–46. doi: 10.1016/j.plipres.2014.07.003
22. Kontush A, Lhomme M, Chapman MJ. Unraveling the complexities of the HDL lipidome. J Lipid Res. (2013) 54:2950–63. doi: 10.1194/jlr.R036095
23. Baker DL, Morrison P, Miller B, Riely CA, Tolley B, Westermann AM, et al. Plasma lysophosphatidic acid concentration and ovarian cancer. JAMA (2002) 287:3081–2. doi: 10.1001/jama.287.23.3081
24. Sutphen R, Xu Y, Wilbanks GD, Fiorica J, Grendys EC, LaPolla JP, et al. Lysophospholipids are potential biomarkers of ovarian cancer. Cancer Epidemiol Biomarkers Prev. (2004) 13:1185–91.
25. Willier S, Butt E, Grunewald TG. Lysophosphatidic acid (LPA) signalling in cell migration and cancer invasion: a focussed review and analysis of LPA receptor gene expression on the basis of more than 1700 cancer microarrays. Biol Cell (2013) 105:317–33. doi: 10.1111/boc.201300011
26. McMahon M, Grossman J, FitzGerald J, Dahlin-Lee E, Wallace DJ, Thong BY, et al. Proinflammatory high-density lipoprotein as a biomarker for atherosclerosis in patients with systemic lupus erythematosus and rheumatoid arthritis. Arthritis Rheum. (2006) 54:2541–9. doi: 10.1002/art.21976
27. Navab M, Anantharamaiah GM, Reddy ST, Van Lenten BJ, Fogelman AM. HDL as a biomarker, potential therapeutic target, and therapy. Diabetes (2009) 58:2711–7. doi: 10.2337/db09-0538
28. Namiri-Kalantari R, Gao F, Chattopadhyay A, Wheeler AA, Navab KD, Farias-Eisner R, et al. The dual nature of HDL: anti-inflammatory and pro-inflammatory. Biofactors (2015) 41:153–9. doi: 10.1002/biof.1205
29. Rosenson RS, Brewer HB Jr, Ansell BJ, Barter P, Chapman MJ, Heinecke JW. Dysfunctional HDL and atherosclerotic cardiovascular disease. Nat Rev Cardiol. (2015) 13:48–60. doi: 10.1038/nrcardio.2015.124
30. Vaisar T, Tang C, Babenko I, Hutchins P, Wimberger J, Suffredini AF, et al. Inflammatory remodeling of the HDL proteome impairs cholesterol efflux capacity. J Lipid Res. (2015) 56:1519–30. doi: 10.1194/jlr.M059089
31. Smith CK, Seto NL, Vivekanandan-Giri A, Yuan W, Playford MP, Manna Z, et al. Lupus high-density lipoprotein induces proinflammatory responses in macrophages by binding lectin-like oxidised low-density lipoprotein receptor 1 and failing to promote activating transcription factor 3 activity. Ann Rheum Dis. (2016). 76:602–11. doi: 10.1136/annrheumdis-2016-209683
32. Pan B, Ren H, He Y, Lv X, Ma Y, Li J, et al. HDL of patients with type 2 diabetes mellitus elevates the capability of promoting breast cancer metastasis. Clin Cancer Res. (2012) 18:1246–56. doi: 10.1158/1078-0432.CCR-11-0817
33. Pan B, Ren H, Lv X, Zhao Y, Yu B, He Y, et al. Hypochlorite-induced oxidative stress elevates the capability of HDL in promoting breast cancer metastasis. J Transl Med. (2012) 10:65. doi: 10.1186/1479-5876-10-65
34. Lane DM, Boatman KK, McConathy WJ. Serum lipids and apolipoproteins in women with breast masses. Breast Cancer Res Treat. (1995) 34:161–9. doi: 10.1007/BF00665788
35. Moorman PG, Hulka BS, Hiatt RA, Krieger N, Newman B, Vogelman JH, et al. Association between high-density lipoprotein cholesterol and breast cancer varies by menopausal status. Cancer Epidemiol Biomark Prev. (1998) 7:483–8.
36. Fiorenza AM, Branchi A, Sommariva D. Serum lipoprotein profile in patients with cancer. A comparison with non-cancer subjects. Int J Clin Lab Res. (2000) 30:141–5. doi: 10.1007/s005990070013
37. Furberg AS, Veierød MB, Wilsgaard T, Bernstein L, Thune I. Serum high-density lipoprotein cholesterol, metabolic profile, and breast cancer risk. JNCI (2004) 96:1152–60. doi: 10.1093/jnci/djh216
38. Chang SJ, Hou MF, Tsai SM, Wu SH, Hou LA, Ma H, et al. The association between lipid profiles and breast cancer among Taiwanese women. Clin Chem Lab Med. 45:1219–23. doi: 10.1515/CCLM.2007.263
39. Cust AE, Kaaks R, Friedenreich C, Bonnet F, Laville M, Tjønneland A, et al. Metabolic syndrome, plasma lipid, lipoprotein and glucose levels, and endometrial cancer risk in the European Prospective Investigation into Cancer and Nutrition (EPIC). Endocrine Relat Cancer (2007) 14:755–67. doi: 10.1677/ERC-07-0132
40. Ahn J, Lim U, Weinstein SJ, Schatzkin A, Hayes RB, Virtamo J, et al. Prediagnostic total and high-density lipoprotein cholesterol and risk of cancer. Cancer Epidemiol Biomark Prev. (2009) 18:2814–21. doi: 10.1158/1055-9965.EPI-08-1248
41. Jafri H, Alsheikh-Ali AA, Karas RH. Baseline and on-treatment high-density lipoprotein cholesterol and the risk of cancer in randomized controlled trials of lipid-altering therapy. J Am College Cardiol. (2010) 55:2846–54. doi: 10.1016/j.jacc.2009.12.069
42. Kitahara CM, Berrington de González A, Freedman ND, Huxley R, Mok Y, Jee SH, et al. Total cholesterol and cancer risk in a large prospective study in Korea. J Clin Oncol. (2011) 29:1592–8. doi: 10.1200/JCO.2010.31.5200
43. Van Hemelrijck M, Walldius G, Jungner I, Hammar N, Garmo H, Binda E, et al. Low levels of apolipoprotein A-I and HDL are associated with risk of prostate cancer in the Swedish AMORIS study. Cancer Causes Control (2011) 22:1011–9. doi: 10.1007/s10552-011-9774-z
44. Zhao W, Guan J, Horswell R, Li W, Wang Y, Wu X, et al. HDL cholesterol and cancer risk among patients with type 2 diabetes. Diabetes Care (2014) 37:3196–203. doi: 10.2337/dc14-0523
45. Touvier M, Fassier P, His M, Norat T, Chan DS, Blacher J, et al. Cholesterol and breast cancer risk: a systematic review and meta-analysis of prospective studies. Br J Nutr. (2015) 114:347–57. doi: 10.1017/S000711451500183X
46. Chandler PD, Song Y, Lin J, Zhang S, Sesso HD, Mora S, et al. Lipid biomarkers and long-term risk of cancer in the Women's Health Study. Am J Clin Nutr. (2016) 103:1397–407. doi: 10.3945/ajcn.115.124321
47. Guo S, He X, Chen Q, Yang G, Yao K, Dong P, et al. The effect of preoperative apolipoprotein A-I on the prognosis of surgical renal cell carcinoma: a retrospective large sample study. Medicine (2016) 95:e3147. doi: 10.1097/MD.0000000000003147
48. Katzke VA, Sookthai D, Johnson T, Kühn T, Kaaks R. Blood lipids and lipoproteins in relation to incidence and mortality risks for CVD and cancer in the prospective EPIC–Heidelberg cohort. BMC Med. (2017) 15:218. doi: 10.1186/s12916-017-0976-4
49. Li X, Tang H, Wang J, Xie X, Liu P, Kong Y, et al. The effect of preoperative serum triglycerides and high-density lipoprotein-cholesterol levels on the prognosis of breast cancer. Breast (2017) 32:1–6. doi: 10.1016/j.breast.2016.11.024
50. Zhou P, Li B, Liu B, Chen T, Xiao J. Prognostic role of serum total cholesterol and high-density lipoprotein cholesterol in cancer survivors: a systematic review and meta-analysis. Clin Chim Acta (2018) 477:94–104. doi: 10.1016/j.cca.2017.11.039
51. Kucharska-Newton AM, Rosamond WD, Mink PJ, Alberg AJ, Shahar E, Folsom AR. HDL-cholesterol and incidence of breast cancer in the ARIC cohort study. Ann Epidemiol. (2008) 18:671–7. doi: 10.1016/j.annepidem.2008.06.006
52. Ehmann M, Felix K, Hartmann D, Schnölzer M, Nees M, Vorderwülbecke S, et al. Identification of potential markers for the detection of pancreatic cancer through comparative serum protein expression profiling. Pancreas (2007) 34:205–14. doi: 10.1097/01.mpa.0000250128.57026.b2
53. Zhang Z, Bast RC, Yu Y, Li J, Sokoll LJ, Rai AJ, et al. Three biomarkers identified from serum proteomic analysis for the detection of early stage ovarian cancer. Cancer Res. (2004) 64:5882–90. doi: 10.1158/0008-5472.CAN-04-0746
54. Kozak KR, Amneus MW, Pusey SM, Su F, Luong MN, Luong SA, et al. Identification of biomarkers for ovarian cancer using strong anion-exchange ProteinChips: potential use in diagnosis and prognosis. Proc Natl Acad Sci USA. (2003) 100:12343–8. doi: 10.1073/pnas.2033602100
55. Kozak KR, Su F, Whitelegge JP, Faull K, Reddy S, Farias-Eisner R. Characterization of serum biomarkers for detection of early stage ovarian cancer. Proteomics (2005) 5:4589–96. doi: 10.1002/pmic.200500093
56. Moore LE, Fung ET, McGuire M, Rabkin CC, Molinaro A, Wang Z, et al. Evaluation of apolipoprotein A1 and posttranslationally modified forms of transthyretin as biomarkers for ovarian cancer detection in an independent study population. Cancer Epidemiol Biomark Prev. (2006) 15:1641–6. doi: 10.1158/1055-9965.EPI-05-0980
57. Clarke CH, Yip C, Badgwell D, Fung ET, Coombes KR, Zhang Z, et al. Proteomic biomarkers apolipoprotein A1, truncated transthyretin and connective tissue activating protein III enhance the sensitivity of CA125 for detecting early stage epithelial ovarian cancer. Gynecol Oncol. (2011) 122:548–53. doi: 10.1016/j.ygyno.2011.06.002
58. Kim YW, Bae SM, Lim H, Kim YJ, Ahn WS. Development of multiplexed bead-based immunoassays for the detection of early stage ovarian cancer using a combination of serum biomarkers. PLoS ONE (2012) 7:e44960. doi: 10.1371/journal.pone.0044960
59. Tuft Stavnes H, Nymoen DA, Hetland Falkenthal TE, Kærn J, Tropé CG, Davidson B. (APOA1 mRNA expression in ovarian serous carcinoma effusions is a marker of longer survival. Am J Clin Pathol. 2014) 142:51–7. doi: 10.1309/AJCPD8NBSHXRXQL7
60. Martin LJ, Melnichouk O, Huszti E, Connelly PW, Greenberg CV, Minkin S, et al. Serum lipids, lipoproteins, and risk of breast cancer: a nested case-control study using multiple time points. JNCI J Natl Cancer Institute (2015) 107:djv032. doi: 10.1093/jnci/djv032
61. Hafiane A, Genest J. High density lipoproteins: measurement techniques and potential biomarkers of cardiovascular risk. BBA Clin. (2015) 3:175–88. doi: 10.1016/j.bbacli.2015.01.005
62. Gutierrez-Pajares JL, Ben Hassen C, Chevalier S, Frank PG. SR-BI: linking cholesterol and lipoprotein metabolism with breast and prostate cancer. Front Pharmacol. (2016) 7:338. doi: 10.3389/fphar.2016.00338
63. Li J, Wang J, Li M, Yin L, Li XA, Zhang TG. Up-regulated expression of scavenger receptor class B type 1 (SR-B1) is associated with malignant behaviors and poor prognosis of breast cancer. Pathol Res Pract. (2016) 212:555–9. doi: 10.1016/j.prp.2016.03.011
64. Yuan B, Wu C, Wang X, Wang D, Liu H, Guo L, et al. High scavenger receptor class B type I expression is related to tumor aggressiveness and poor prognosis in breast cancer. Tumour Biol. (2016) 37:3581–8. doi: 10.1007/s13277-015-4141-4
65. Ruscica M, Botta M, Ferri N, Giorgio E, Macchi C, Franceschini G, et al. High density lipoproteins inhibit oxidative stress-induced prostate cancer cell proliferation. Sci Rep. (2018) 8:2236. doi: 10.1038/s41598-018-19568-8
66. Nissen SE, Tsunoda T, Tuzcu EM, Schoenhagen P, Cooper CJ, Yasin M, et al. Effect of recombinant apoa-i milano on coronary atherosclerosis in patients with acute coronary syndromes: a randomized controlled trial. JAMA (2003) 290:2292–300. doi: 10.1001/jama.290.17.2292
67. Remaley AT, Amar M, Sviridov D. HDL-replacement therapy: mechanism of action, types of agents and potential clinical indications. Exp Rev Cardiovasc Ther. (2008) 6:1203–15. doi: 10.1586/14779072.6.9.1203
68. Dasseux JL, Schwendeman AS, Zhu L. Apolipoprotein A-I Mimics. USPTO. United States, Cerenic Therapeutics Holding SA (2013).
69. Krause BR, Remaley AT. Reconstituted HDL for the acute treatment of acute coronary syndrome. Curr Opin Lipidol. (2013) 24:480–6. doi: 10.1097/MOL.0000000000000020
70. Di Bartolo BA, Nicholls SJ, Bao S, Rye KA, Heather AK, Barter PJ, et al. The apolipoprotein A-I mimetic peptide ETC-642 exhibits anti-inflammatory properties that are comparable to high density lipoproteins. Atherosclerosis (2011) 217:395–400. doi: 10.1016/j.atherosclerosis.2011.04.001
71. Schwendeman A, Sviridov DO, Yuan W, Guo Y, Morin EE, Yuan Y, et al. The effect of phospholipid composition of reconstituted HDL on its cholesterol efflux and anti-inflammatory properties. J Lipid Res. (2015) 56:1727–37. doi: 10.1194/jlr.M060285
72. Kuai R, Li D, Chen YE, Moon JJ, Schwendeman A. High-density lipoproteins: nature's multifunctional nanoparticles. ACS Nano (2016) 10:3015–41. doi: 10.1021/acsnano.5b07522
73. Nanjee MN, Crouse JR, King JM, Hovorka R, Rees SE, Carson ER, et al. Effects of intravenous infusion of lipid-free apo A-I in humans. Arterioscler Thromb Vasc Biol. (1996) 16:1203–14. doi: 10.1161/01.ATV.16.9.1203
74. Li D, Gordon S, Schwendeman A, Remaley A. Apolipoprotein mimetic peptides for stimulating cholesterol efflux. In: Anantharamaiah GM and Goldberg D, editors. Apolipoprotein Mimetics in the Management of Human Disease. Cham; Heidelberg; New York, NY; Dordrecht; London: Springer International Publishing (2015). p. 29–42.
75. Tang J, Li D, Drake L, Yuan W, Deschaine S, Morin EE, et al. Influence of route of administration and lipidation of apolipoprotein A-I peptide on pharmacokinetics and cholesterol mobilization. J Lipid Res. (2017) 58:124–36. doi: 10.1194/jlr.M071043
76. Khan MLN, Drake SL, Crockatt JG, Dasseux JLH. Single-dose intravenous infusion of ETC-642, a 22-Mer ApoA-I analogue and phospholipids complex, elevates HDL-C in atherosclerosis patients. Circulation (2003) 108(Suppl. IV):563–4.
77. Miles J, Khan M, Painchaud C, Lalwani N, Drake S, Dasseux J. Single-dose tolerability, pharmacokinetics, and cholesterol mobilization in HDL-C fraction following intravenous administration of ETC-642, a 22-mer ApoA-I analogue and phospholipids complex, in atherosclerosis patients. Arterioscler Thromb Vasc Biol. (2004), E19–E19.
78. CSL (2018). CSL Behring Announces First Patient Enrollment in Phase 3 Clinical Trial of CSL112 to Assess Reduction of Early Recurrent Cardiovascular Events in Heart Attack Survivors. King of Prussia, PA: PR Newswire.
79. Easton R, Gille A, D'Andrea D, Davis R, Wright SD, Shear C. A multiple ascending dose study of CSL112, an infused formulation of ApoA-I. J Clin Pharmacol. (2014) 54:301–10. doi: 10.1002/jcph.194
80. Gille A, D'Andrea D, Tortorici MA, Hartel G, Wright SD. CSL112 (apolipoprotein A-I [human]) enhances cholesterol efflux similarly in healthy individuals and stable atherosclerotic disease patients. Arterioscler Thromb Vasc Biol. (2018) 38:953–63. doi: 10.1161/ATVBAHA.118.310538
81. Badana A, Chintala M, Varikuti G, Pudi N, Kumari S, Kappala VR, et al. Lipid raft integrity is required for survival of triple negative breast cancer cells. J Breast Cancer (2016) 19:372–84. doi: 10.4048/jbc.2016.19.4.372
82. Li YC, Park MJ, Ye SK, Kim CW, Kim YN. Elevated levels of cholesterol-rich lipid rafts in cancer cells are correlated with apoptosis sensitivity induced by cholesterol-depleting agents. Am J Pathol. (2006) 168:1107–18; quiz 1404–1105. doi: 10.2353/ajpath.2006.050959
83. Resnik N, Repnik U, Kreft ME, Sepčić K, Maček P, Turk B, et al. Highly selective anti-cancer activity of cholesterol-interacting agents methyl-beta-cyclodextrin and ostreolysin A/pleurotolysin b protein complex on urothelial cancer cells. PLoS ONE (2015) 10:e0137878. doi: 10.1371/journal.pone.0137878
84. Yamaguchi R, Perkins G, Hirota K. Targeting cholesterol with beta-cyclodextrin sensitizes cancer cells for apoptosis. FEBS Lett. (2015) 589 (24 Pt B):4097–105. doi: 10.1016/j.febslet.2015.11.009
85. Taylor MJ, Sanjanwala AR, Morin EE, Rowland-Fisher E, Anderson K, Schwendeman A, et al. Synthetic high-density lipoprotein (sHDL) inhibits steroid production in HAC15 adrenal cells. Endocrinology (2016) 157:3122–9. doi: 10.1210/en.2014-1663
86. Su F, Kozak KR, Imaizumi S, Gao F, Amneus MW, Grijalva V, et al. Apolipoprotein A-I (apoA-I) and apoA-I mimetic peptides inhibit tumor development in a mouse model of ovarian cancer. Proc Natl Acad Sci USA. (2010) 107:19997–20002. doi: 10.1073/pnas.1009010107
87. Gao F, Vasquez SX, Su F, Roberts S, Shah N, Grijalva V, et al. L-5F, an apolipoprotein A-I mimetic, inhibits tumor angiogenesis by suppressing VEGF/basic FGF signaling pathways(). Integr Biol. (2011) 3:479–89. doi: 10.1039/c0ib00147c
88. Gao F, Chattopadhyay A, Navab M, Grijalva V, Su F, Fogelman AM, et al. Apolipoprotein A-I mimetic peptides inhibit expression and activity of hypoxia-inducible factor-1α in human ovarian cancer cell lines and a mouse ovarian cancer model. J Pharmacol Exp Ther. (2012) 342:255–62. doi: 10.1124/jpet.112.191544
89. Cedó L, García-León A, Baila-Rueda L, Santos D, Grijalva V, Martínez-Cignoni MR, et al. ApoA-I mimetic administration, but not increased apoA-I-containing HDL, inhibits tumour growth in a mouse model of inherited breast cancer. Sci Rep. (2016) 6:36387. doi: 10.1038/srep36387
90. Peng M, Zhang Q, Cheng Y, Fu S, Yang H, Guo X, et al. Apolipoprotein A-I mimetic peptide 4F suppresses tumor-associated macrophages and pancreatic cancer progression. Oncotarget (2017) 8:99693–706. doi: 10.18632/oncotarget.21157
91. Subramanian C, Kuai R, Zhu Q, White P, Moon J, Schwendeman A, et al. sHDL nanoparticles: a novel therapeutic strategy for adrenocortical carcinomas. Surgery (2016) 159:284–95. doi: 10.1016/j.surg.2015.08.023
92. Su F, Grijalva V, Navab K, Ganapathy E, Meriwether D, Imaizumi S, et al. HDL mimetics inhibit tumor development in both induced and spontaneous mouse models of colon cancer. Mol Cancer Ther. (2012) 11:1311–9. doi: 10.1158/1535-7163.MCT-11-0905
93. Zamanian-Daryoush M, Lindner D, Tallant TC, Wang Z, Buffa J, Klipfell E, et al. The cardioprotective protein apolipoprotein a1 promotes potent anti-tumorigenic effects. J Biol Chem. (2013) 288:21237–52. doi: 10.1074/jbc.M113.468967
94. Zheng Y, Liu Y, Jin H, Pan S, Qian Y, Huang C, et al. Scavenger receptor B1 is a potential biomarker of human nasopharyngeal carcinoma and its growth is inhibited by HDL-mimetic nanoparticles. Theranostics (2013) 3:477–86. doi: 10.7150/thno.6617
95. Sekine Y, Demosky SJ, Stonik JA, Furuya Y, Koike H, Suzuki K, et al. High-density lipoprotein induces proliferation and migration of human prostate androgen-independent cancer cells by an ABCA1-dependent mechanism. Mol Cancer Res. (2010) 8:1284–94. doi: 10.1158/1541-7786.MCR-10-0008
96. Danilo C, Gutierrez-Pajares JL, Mainieri MA, Mercier I, Lisanti MP, Frank PG. Scavenger receptor class B type I regulates cellular cholesterol metabolism and cell signaling associated with breast cancer development. Breast Cancer Res. (2013) 15:R87. doi: 10.1186/bcr3483
97. Nofer JR, Kehrel B, Fobker M, Levkau B, Assmann G, von Eckardstein A. HDL and arteriosclerosis: beyond reverse cholesterol transport. Atherosclerosis (2002) 161:1–16. doi: 10.1016/S0021-9150(01)00651-7
98. Pussinen PJ, Karten B, Wintersperger A, Reicher H, McLean M, Malle E, et al. The human breast carcinoma cell line HBL-100 acquires exogenous cholesterol from high-density lipoprotein via CLA-1 (CD-36 and LIMPII analogous 1)-mediated selective cholesteryl ester uptake. Biochem J. (2000) 349 (Pt 2):559–66. doi: 10.1042/bj3490559
99. Connelly MA, Williams DL. SR-BI and cholesterol uptake into steroidogenic cells. Trends Endocrinol Metab. (2003) 14:467–72. doi: 10.1016/j.tem.2003.10.002
100. Mooberry LK, Sabnis NA, Panchoo M, Nagarajan B, Lacko AG. Targeting the SR-B1 receptor as a gateway for cancer therapy and imaging. Front Pharmacol. (2016) 7:466. doi: 10.3389/fphar.2016.00466
101. Tang J, Kuai R, Yuan W, Drake L, Moon JJ, Schwendeman A. Effect of size and pegylation of liposomes and peptide-based synthetic lipoproteins on tumor targeting. Nanomedicine (2017) 13:1869–78. doi: 10.1016/j.nano.2017.04.009
Keywords: High-density lipoprotein (HDL), Apolipoprotein A-I (ApoA-I), endocrine cancer, cholesterol, cancer therapy
Citation: Morin EE, Li X-A and Schwendeman A (2018) HDL in Endocrine Carcinomas: Biomarker, Drug Carrier, and Potential Therapeutic. Front. Endocrinol. 9:715. doi: 10.3389/fendo.2018.00715
Received: 22 September 2018; Accepted: 12 November 2018;
Published: 30 November 2018.
Edited by:
Vincenzo Pezzi, Università della Calabria, ItalyReviewed by:
Paolo Magni, Università degli Studi di Milano, ItalyCopyright © 2018 Morin, Li and Schwendeman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Emily E. Morin, ZW1tb3JpbkBtZWQudW1pY2guZWR1
Anna Schwendeman, YW5uYXNjaHdAbWVkLnVtaWNoLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.