 Emese Domonkos
Emese Domonkos Július Hodosy
Július Hodosy Daniela Ostatníková
Daniela Ostatníková Peter Celec
Peter Celec- 1Faculty of Medicine, Institute of Molecular Biomedicine, Comenius University, Bratislava, Slovakia
- 2Faculty of Medicine, Institute of Physiology, Comenius University, Bratislava, Slovakia
- 3Faculty of Medicine, Institute of Pathophysiology, Comenius University, Bratislava, Slovakia
- 4Department of Molecular Biology, Faculty of Natural Sciences, Comenius University, Bratislava, Slovakia
Testosterone affects brain functions and might explain some of the observed behavioral sex differences. Animal models may help in elucidating the possible involvement of sex hormones in these sex differences. The effects of testosterone have been intensively investigated, especially in anxiety models. Numerous experiments have brought inconsistent results with either anxiolytic or anxiogenic effects. Besides methodological variations, contradictory findings might be explained by the divergent metabolism of testosterone and its recognition by neurons during prenatal and postnatal development. Gonadectomy and subsequent supplementation have been used to study the role of sex hormones. However, the variable duration of hypogonadism might affect the outcomes and the effect of long-term androgen deficiency is understudied. Testosterone can be metabolized to dihydrotestosterone strengthening the androgen signaling, but also to estradiol converting the androgen to estrogen activity. Moreover, some metabolites of testosterone can modulate γ-aminobutyric acid and serotonergic neurotransmission. Here we review the currently available experimental data in experimental rodents on the effects of testosterone on anxiety during development. Based on the experimental results, females are generally less anxious than males from puberty to middle-age. The anxiety-like behavior of females and males is likely influenced by early organizational effects, but might be modified by activational effects of testosterone and its metabolites. The effects of sex hormones leading to anxiogenesis or anxiolysis depend on factors affecting hormonal status including age. The biological and several technical issues make the study of effects of testosterone on anxiety very complex and should be taken into account when interpreting experimental results.
Introduction
Besides depression, anxiety disorders, including generalized anxiety disorder, panic disorder, phobias, social anxiety disorder, separation anxiety disorder, obsessive-compulsive disorder and post-traumatic stress disorder, are the most common of all mental disorders (1). According to a survey performed in 2010, 14% of the population in Europe suffers from an anxiety disorder each year (2). The prevalence rate of anxiety disorders is influenced by various factors, including gender (3). It is known that sex hormones affect brain morphology as well as brain functions. Based on the sex differences in prevalence of anxiety disorders in human, the role of sex hormones has been intensively investigated. A large body of animal experiments has brought contradictory results showing either anxiolytic or anxiogenic effects of testosterone (4, 5). These inconsistent findings might be due to differences in organizational and activational actions of testosterone affecting the neuronal circuits during prenatal and postnatal development. Therefore, in the present paper, the currently available experimental data on the sex differences in anxiety-like behavior and the potential role of testosterone affecting these differences across different period of life are reviewed.
Sex Differences in Anxiety-Like Behavior in Rodent Models
Anxiety is interconnected with behavioral and brain functions such as fear response. In clinical practice, anxiety is considered to be a consequence of overestimated response to threat in uncertain situations associated with prolonged hyper-vigilance and hyper-arousal (6). In neuropsychiatric studies, rodents are the most commonly used animal models (7). While in humans, anxiety might be assessed by self-report questionnaire, in rodents anxiety is evaluated based on temporary behavioral responses to various threatening stimuli. Therefore the translational value of such evaluation is disputable. On the other hand, animal experiments are crucial to elucidate causality behind observed associations. Although mice and rats may differ in some behavioral traits, the behavioral tests assessing anxiety in rodents are based on their common innate avoidance of certain conditions, such as bright illuminations (8) or open space (9–11). Therefore the most of the anxiety tests are suitable for both, mice and rats (7). In anxiety assessment it should be also considered that anxiety is associated with other behavioral phenotypes, such as aggression and violence (12–19). These exhibit sex differences and positive correlations with testosterone concentration (20, 21). Furthermore, anxiety-related behavior assessed in experimental rodents is often related to their cognitive performance, which might be affected by sex hormones (22, 23) and dependent on androgen signaling (24).
It should be noted that some behavioral traits, including anxiety, might be influenced by estrous cycle in females. It has been shown that female rat in proestrus display more explorative behavior in a novel environment and less freezing behavior following shock in defensive burying test, they spent more time in the open arm of the elevated plus maze and interact longer with an unfamiliar social partner in comparison to males but also to females during other phases of the estrous cycle (25). Although, to cover whole estrous cycle, females in the experiments are often selected for each stages and a heterogeneous group is used, but it leads to high inter-individual variability (26–28). Therefore, the assessment of sex differences in anxiety behavior is problematic.
Women are twice more likely to experience anxiety as men (26). However, it seems that sex differences in the behavioral stress responses mostly point to the opposite direction. Despite of the evidence suggesting the effect of hormonal fluctuation, the behavior of the females is often generalized without considering the estrous cycle, especially in the earlier experiments investigating sex differences. Such results show that adult female rats move in the open field more than males (29), while male rats display more freezing behavior and defecation, longer-lasting grooming reaction, less rearing as well as less time spent in the center zone of the arena (28). Females, mice as well as rats, make more entries into the open arms and spent greater time in open arms of the elevated plus maze, interact more with a same-sex social partner in a novel environment and bury fewer marbles than males (27, 28, 30, 31). On the contrary, in the Vogel conflict test, male rats seems to be less anxious than females indicated by more licking of the drinking spout during the punish period (30). In time spent in the bright chamber of the light-dark box, contradictory results have been found. In some experiments, females show higher anxiety than males (32), in other experiments, the opposite sex difference was proved (33), while the lack of sex differences is published rarely (27). It should be noted that sex differences in experimental animals in anxiety-like behavior might depend on many factors, such as strain (34) or breeding condition (32), and other sources of inter-individual variability are likely not identified yet (35). The sex differences in frequently used behavioral tests for anxiety changing with age, as well as the short- and long-term consequences of testosterone deprivation or exposure in males in different periods of life are illustrated in Figure 1.
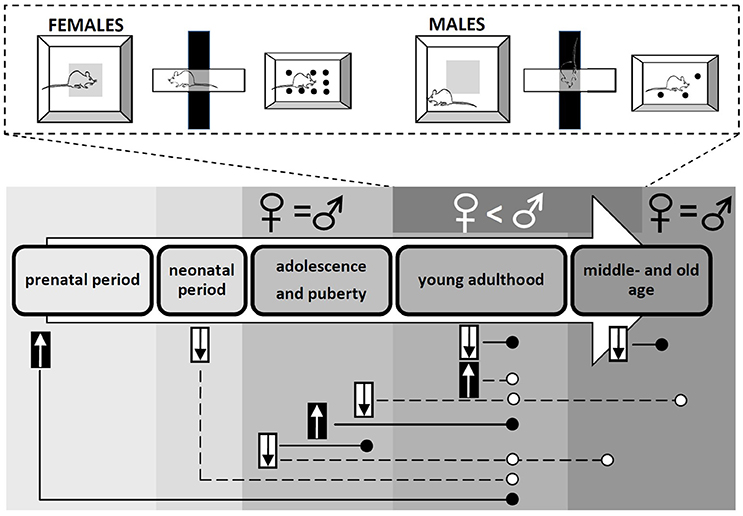
Figure 1. Sex differences in anxiety-like behavior assessed using the open field, elevated plus maze and marble-burying tests. The consequences of testosterone exposure and deprivation in males are shown across the life.  - testosterone exposure;
- testosterone exposure;  - testosterone deprivation; ° - anxiolytic effect; • - anxiogenic effect.
- testosterone deprivation; ° - anxiolytic effect; • - anxiogenic effect.
The Role of Testosterone in Anxiety-Like Behavior of Rodents Across Different Life Periods
The sexually dimorphic behaviors related to anxiety can be influenced by both, organizational and activational effects of gonadal hormones. Organization of the neuronal circuits by testosterone and/or its metabolites predetermines the behavioral responses to the activational effects of sex hormones later in life. Besides the sensitive period of early—prenatal and perinatal development (15, 27), increasing amount of evidence supports the hypothesis that pubertal hormones (re)organize the brain and determine a variety of adult behaviors during adolescence (36, 37) or outside of the classic critical periods of neurodevelopment (38). An overview of the results of some experiments investigating sex differences and the role of testosterone in anxiety-like behavior, with particular regard to the age of animals with hormone manipulation and analysis of the outcomes, is provided in Table 1.
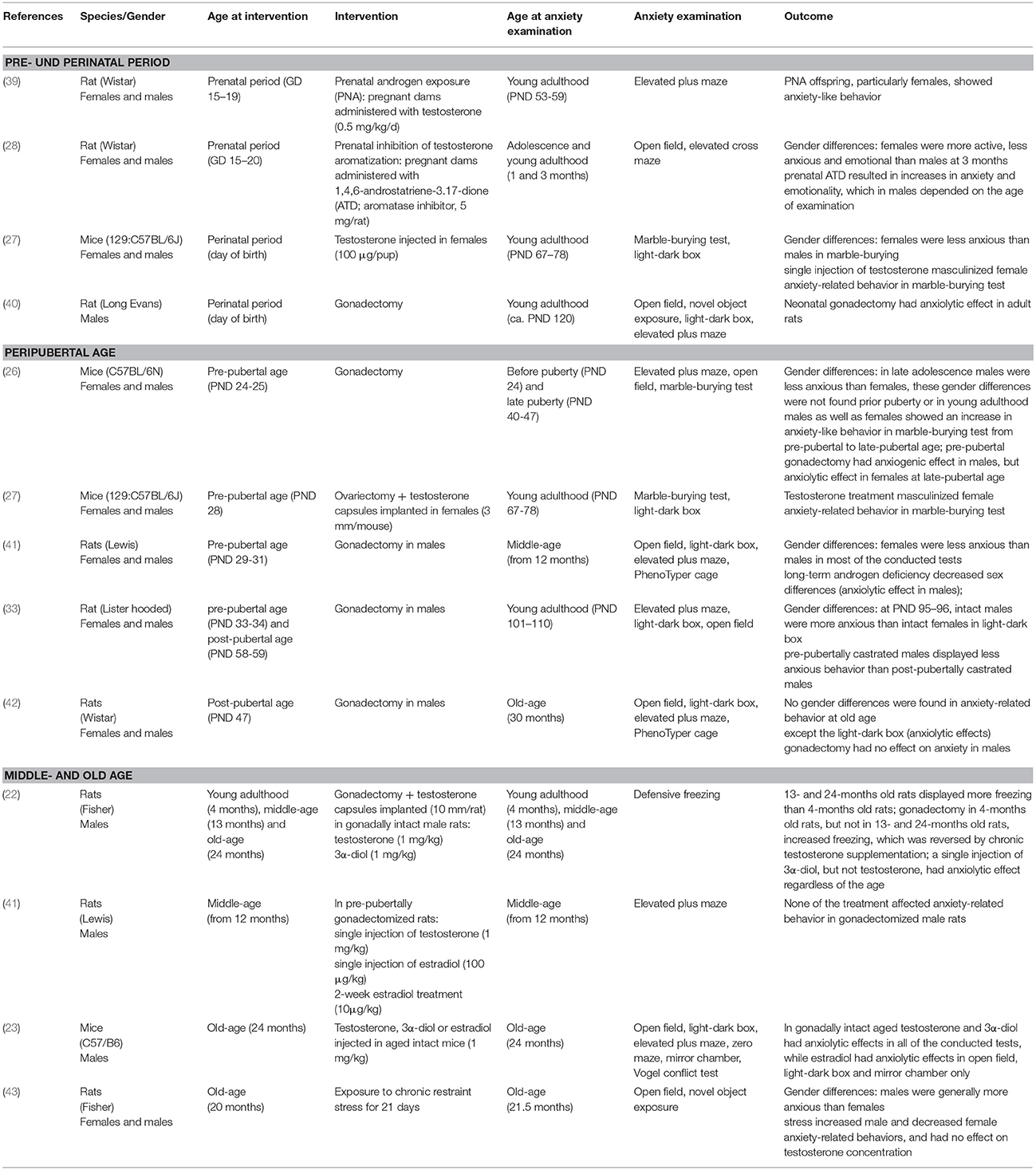
Table 1. Overview of some experimental results on the effect of testosterone on anxiety-like behavior in regard to age at hormone manipulation.
Prenatal and Neonatal Period
Early development is a critical period for brain formation and also for its modulation by sex hormones contributing to sexual dimorphism. Prenatal exposure to testosterone excess is associated with impaired neural development and mental functions (44, 45). Offspring of women with polycystic ovary syndrome is exposed to high concentration of androgens in utero. As shown in a rat model, maternal hypergonadism may result in increased risk for developing anxiety disorder in offspring, especially in female offspring (39). Inhibition of testosterone aromatization to estradiol in pregnant dams may induce increased defecation and freezing in the open field, as well as decreased number of hangings from and time spent on the open arms, pointing out an increased emotionality and anxiety in both, female and male offspring (28, 46). Accordingly, exposure to high concentration of testosterone during prenatal development leads to anxiogenesis in the adulthood, and females seems to be more sensitive to this effect of prenatal testosterone.
In addition to prenatal, also perinatal exposure to testosterone may predetermine anxiety-related behavior later during life. As found in adult rats and mice, testosterone injected on the day of birth or neonatal ovariectomy can masculinize anxiety-related behavior of females increasing the number of the marbles buried (27) and decreasing the time spent on the open arms (47). On contrary, adult male rats, castrated on the day of birth have a higher locomotor activity in the open arena, spent more time in the center zone, in the light chamber, in the open arms, and exhibit a higher number of novel object visits than their sham operated counterparts (40, 48). These results suggest that the absence of estrogens in females and testosterone in males during the perinatal period restrain the normal differentiation of gender-specific adult anxiety responses to the particular stimuli.
Adolescence
Puberty is a restricted period in adolescence characterized by a rapid elevation of circulating sex hormones accompanied by reproductive maturation. Adolescence, as the age between childhood and adulthood, represents a sensitive period in neurodevelopment as well. Numerous evidence proves that the remodeling of the adolescent brain occurs under the influence of sex hormones (36, 37). A large body of animal experiments has shown that sex differences in several anxiety tests arise from the organizational effects of testosterone at peri-pubertal age. In rats, male gonadectomy, either at puberty onset or shortly thereafter, results in lower anxiety in the open area, the behavioral response typical for females (29). Similar effects of pre-pubertal castration were found in the elevated plus maze and light-dark box, where adult male rats that had been gonadectomized prior to puberty exhibit a higher frequency of head-dipping and more time spent in the open arms or in the light part of the light-dark box (33). Environment-related social anxiety in males is organized also by pubertal sex hormones, where the aromatization of testosterone seems to be critical (49). In addition, middle- (12-months old) as well as old-aged male rats (30-months old) that have been castrated before puberty onset or at late adolescent age, respectively, display lower or similar anxiety in comparison to intact aged-matched females in a test-specific manner (41, 42). Pre-pubertal gonadectomy has anxiogenic consequences in male mice in the elevated plus maze, but anxiolytic effect in female mice in the marble-burying test (26). On the other side, testosterone exposure and social experience during adolescence equally increase anxiety-like behavior, as shown in adult male hamsters (50). In females rats, single testosterone administration at the day of birth (organizational effect) as well as continual testosterone exposure from the puberty onset (activational effect) increases activity in marble-burying test of anxiety-like behavior, abolishing sex differences in adulthood (27). Furthermore, while in 1-month-old intact rats no sex differences were found in anxiety-like behavior, testing at 3 months of age revealed clear sex differences in reaction to the environmental novelty, females being more active and less anxious than males in the open field and in the elevated plus maze (28). Interestingly, in late adolescence, male mice spent more time on open arms than females. However, this sex difference in open-arm time is not present prior to puberty onset and does not persist into young adulthood (26).
Adulthood
Most published experiments examine the role of testosterone in anxiety-like behavior and the underlying mechanisms in young adult animals. Unlike the anxiolytic effect of peri-pubertal castration, castration of adult males is frequently associated with increased anxiety-like behavior in a battery of behavioral tests, such as open field, elevated plus maze, and defensive-burying test (51–58). However, in some experiments in rats, no effect of adult gonadectomy was found (29, 33). Presumably, the different time interval between hormone deprivation and behavioral examination among the studies may result in these differences. The short-term effect of gonadectomy has been investigated in a large number of experiments, while the effects of the long-term hypogonadism are understudied.
The anxiogenic effect of adult castration might be reversed by hormone supplementation (52, 54, 59–63). The anxiolytic-like effect of testosterone has been revealed in gonadally intact healthy animals, as well (64–66). However, testosterone can be either reduced by 5α-reductase to the more potent androgen dihydrotestosterone, or aromatized by aromatase to estradiol converting the androgen to estrogen activity. In brain, dihydrotestosterone can be further metabolized to 5α-androstane-3α,17β-diol (3α-diol) and to 5α-androstane-3β,17β-diol (3β-diol), neuroactive steroids possessing neuromodulatory activity (67). One of the most cited study investigating the rapid effects of testosterone on anxiety-like behavior in mice has suggested that the anxiolytic effect of testosterone is mediated by its 5α-reduced metabolites (64). Likewise, it has been proved that administration of 3α-diol decreases anxiety-like behavior in male rats (52, 59, 60, 63) as well as in female rats (68), while inhibition of testosterone metabolism to 3α-diol increases anxiety in male rats (62). Similar to effect of hormone administration, sexual experience and the exposure of intact male rats or mice to female subjects may decrease anxiety-like behavior associated with increased concentration of testosterone in plasma and hippocampus, as well as increased hypothalamic testosterone and 3α-diol concentration (64, 69). On the other hand, estradiol (61) and another metabolite of testosterone, androsterone (70), may cause anxiolysis, as well. Fernández-Guasti and Martínez-Mota (54) have shown that repeated administration of testosterone, but not a single injection of testosterone, nor treatment with 3α-diol or androsterone produced anxiolysis in male gonadectomized rats (54). According to published data, the manifestation of the anxiolytic-like effect of testosterone or its metabolites is highly dependent on dose and duration of treatment (54, 58, 64, 65, 71).
Due to its complex metabolism, the effects of testosterone might be mediated through different mechanisms of action. Testosterone and dihydrotestosterone are ligands of the androgen receptor. The latter binds with a greater affinity to the receptor and activates gene transcription resulting in an increased androgen activity (72). The role of androgen signaling in the regulation of anxiety-related behavior was demonstrated by administration of the androgen receptor antagonist flutamide (52, 54, 66), but also using the animal model of testicular feminization mutation (73–76) and androgen receptor knockout mice (77). In addition, it has been shown that some selective androgen receptor modulators may exert neuroprotective effects (78) and may affect some behavioral and brain functions (79–83). However, experimental studies examining their effects on anxiety important for the deeper understanding of the association between testosterone and anxiety, but potentially also for therapeutic applications are lacking. Testosterone can exert its effects also via estrogen receptors following aromatization of testosterone to estradiol, as well as by conversion of dihydrotestosterone to 3α-diol and 3β-diol (23, 71). Furthermore, it was shown that testosterone may modulate γ-aminobutyric acid (GABA)-stimulated chloride influx (84). This might be due to the fact that 3α-diol and androsterone can bind to GABAA/benzodiazepine receptor. Unlike to GABAC receptor, which seems not to be involved in the regulation of anxiety by testosterone (85), inhibition of GABAA receptor by bicucculine or picrotoxin (64), as well as by flumazenil (70) diminishes the anxiolytic effect of testosterone metabolites. In females, inhibition of GABAA and ERβ receptor diminishes the anxiolytic effect of testosterone, but not when aromatase is inhibited. Thus, the anxiolytic effect of testosterone might be mediated by its reduced metabolites via GABAA with participation of ERβ (86). On the other side, using βER-knockout mice, it has been shown that the anxiolytic effect of testosterone metabolites mediated by the GABAA receptor may be not as robust as via activation of ERβ receptor (70). In addition, it was shown that testosterone can regulate serotonin neurotransmission. Gonadectomy results in decreased expression of 5-HT2A serotonergic receptors in hippocampus, testosterone replacement enhances 5-HT2A expression accompanied by reduced anxiety (58). On the contrary, inhibition of 5-HT1A receptor increases efficacy of testosterone treatment leading to enhanced anxiolytic-like effect (53). As shown by observation of inter-individual differences, anxious animals display higher expression of tryptophan hydroxylase—an enzyme involved in serotonin synthesis, and higher concentration of serotonin in amygdala than their non-anxious counterparts (35). Interestingly, while in gonadally intact rats, anxiety-like behavior positively correlates with the expression of tryptophan hydroxylase, this association is abolished by gonadectomy. As serotonin depletion diminishes anxiety level in anxious individuals, it also reverses the anxiogenic effect of gonadectomy (57).
Aging
Aging is associated with a progressive decline of circulating testosterone resulting from decreased function of the hypothalamo-pituitary-gonadal axis. Unlike menopause in women, andropuase in men is a long-lasting process. The age-related hypogonadism in men is considered to cause some of the senescence symptoms, including anxiety (87). This has been proved in rats. While gonadectomy in young adulthood and, partially, at middle-age induces anxiety, this effect is less apparent in aged rats (22). On the other hand, in a genetic mouse model of aging, no association was found between low testosterone and anxiety (88). It should be considered that andropause is via negative feedback associated with increased concentration of luteinizing hormone. In adult female rats, agonist of gonadotropin-releasing hormone triptorelin, which inhibits luteinizing hormone release, alone as well as in combination with estradiol treatment reduces gonadectomy-induced anxiety (89). Further experiments are needed to examine the impact of luteinizing hormone modulation on anxiety in aging males.
In our laboratory, sex differences in anxiety-like behavior of middle-aged (12-months old) rats have been shown. Aging females were less anxious than aging males in a battery of behavioral tests (41). Similarly, male rats at older age (21-months old) have proved to be generally more anxious than females. In addition, stress in males increased, while in females it decreased anxiety-like behavior (43). However, we have shown that at very old age (30-months old) female and male rats do not differ any more in anxiety-like behavior in elevated plus maze and light-dark box, or in open-field ambulation, except rearing behavior (42). These results suggest that the causal role of endogenous testosterone in anxiety-related behavior at is doubtful at least in aging animals. However, it should be considered that the anxiety-like behavior in these rats might be biased by age-related decline in locomotor activity (90). Presumably, the sex differences, observed in young and middle-aged animals, result from the organizational effect of testosterone occurring at early development and puberty. These differences, however, do not persist into the old age, which can be caused by age-related cognitive and affective decline.
There is some evidence suggesting the anti-anxiety properties of exogenous testosterone as well as its metabolites, 3α-diol and estradiol, in old gonadally intact rats (23, 91) or in rats gonadectomized at old age (22). On contrary, we have demonstrated that neither single administration of testosterone or estradiol, nor short-term treatment with estradiol affect anxiety-like behavior in middle-aged rats, when they suffer from long-term hypogonadism initiated before puberty onset (41). Therefore, it seems to be possible that the length of testosterone deficiency as well as the age of animals might determine the activational effect of supplemented hormones on anxiety-like behavior. Furthermore, the age, dose and duration of hormone replacement therapy as well as their interactions were shown to influence the action of testosterone leading to either anxiolysis or anxiogenesis (91).
Conclusion
Sex differences in anxiety-like behavior origin from organizational effect and might be modified by activational effect of sex hormones. There are critical periods across life that are crucial in organization of neuronal circuits and pre-programming the activational effect of testosterone on anxiety-like behavior. According to the current experimental studies, testosterone exposure during brain development predetermines higher anxiety in males. Furthermore, the absence of testosterone during the activation of neuronal circuits involved in anxiety results in anxiogenesis, as well. Although it seems that testosterone might have anxiolytic effect in adult males following short-term androgen deficiency, the impact of testosterone on anxiety-related behavior following long-term hypogonadal condition is understudied, mainly in aged animals, and needs further examinations. Based on the experimental results, age is a crucial factor that modulates the effect of both, endogenous and exogenous sex hormones. Thus, studying the expression of the various androgen receptors in different brain regions during development will be important to understand the contradictory findings from different experiments. Testosterone exposure or even deficiency in critical periods of development may have long-lasting consequences persisting into middle- or old age. This should be considered when interpreting experimental results and potentially hormonal treatments in human patients.
Author Contributions
ED drafted the manuscript, prepared the table and the figure. JH corrected the draft. DO corrected the draft. PC designed the review and corrected the draft. All authors approved the final version of the manuscript.
Funding
The authors are supported by Slovak research and development agency (grant number APVV-15-0045).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Kendrick T, Pilling S. Common mental health disorders — identification and pathways to care: NICE clinical guideline. Br J Gen Pract. (2012) 62:47–9. doi: 10.3399/bjgp12X616481
2. Wittchen HU, Jacobi F, Rehm J, Gustavsson A, Svensson M, Jönsson B, et al. The size and burden of mental disorders and other disorders of the brain in Europe 2010. Eur Neuropsychopharmacol. (2011) 21:655–79. doi: 10.1016/j.euroneuro.2011.07.018
3. Alonso J, Angermeyer MC, Bernert S, Bruffaerts R, Brugha TS, Bryson H, et al. Prevalence of mental disorders in Europe: results from the European Study of the Epidemiology of Mental Disorders (ESEMeD) project. Acta Psychiatr Scand. (2004) 109:21–7. doi: 10.1111/j.1600-0047.2004.00327.x
4. Celec P, Ostatníková D, Hodosy J. On the effects of testosterone on brain behavioral functions. Front. Neurosci. (2015) 9:12. doi: 10.3389/fnins.2015.00012
5. McHenry J, Carrier N, Hull E, Kabbaj M. Sex differences in anxiety and depression: role of testosterone. Front Neuroendocrinol. (2014) 35:42–57. doi: 10.1016/j.yfrne.2013.09.001
6. Sylvers P, Lilienfeld SO, LaPrairie JL. Differences between trait fear and trait anxiety: Implications for psychopathology. Clin Psychol Rev. (2011) 31:122–37. doi: 10.1016/j.cpr.2010.08.004
7. Padurariu M, Antioch I, Balmus I, Ciobica A, El-lethey HS. Describing some behavioural animal models of anxiety and their mechanistics with special reference to oxidative stress and oxytocin relevance. Int J Vet Sci. Med. (2017) 5:98–104. doi: 10.1016/j.ijvsm.2017.08.003
8. Crawley J, Goodwin FK. Preliminary report of a simple animal behavior model for the anxiolytic effects of benzodiazepines. Pharmacol Biochem Behav. (1980) 13:167–70.
9. Pellow S, Chopin P, File SE, Briley M. Validation of open : closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J Neurosci Methods (1985) 14:149–67. doi: 10.1016/0165-0270(85)90031-7
10. Schmitt U, Hiemke C. Combination of open field and elevated plus-maze: a suitable test battery to assess strain as well as treatment differences in rat behavior. Prog Neuro-Psychopharmacol Biol Psychiatry (1998) 22:1197–215. doi: 10.1016/S0278-5846(98)00051-7
11. Seibenhener ML, Wooten MC. Use of the open field maze to measure locomotor and anxiety-like behavior in mice. J Vis Exp. (2015) 8. doi: 10.3791/52434
12. Bacqué-Cazenave J, Cattaert D, Delbecque JP, Fossat P. Social harassment induces anxiety-like behaviour in crayfish. Sci Rep. (2017) 7:1–7. doi: 10.1038/srep39935
13. Birger M, Swartz M, Cohen D, Alesh Y, Grishpan C, Kotelr M. Aggression: the testosterone-serotonin link. Isr Med Assoc J. (2003) 5:653–8.
14. Bondar NP, Lepeshko AA, Reshetnikov VV. Effects of early-life stress on social and anxiety-like behaviors in adult mice: sex-specific effects. Behav Neurol. (2018) 2018:32–4. doi: 10.1155/2018/1538931
15. Dela Cruz C, Pereira OCM. Prenatal testosterone supplementation alters puberty onset, aggressive behavior, and partner preference in adult male rats. J Physiol Sci. (2012) 62:123–31. doi: 10.1007/s12576-011-0190-7
16. Giammanco M, Tabacchi G, Giammanco S, Di Majo D, La Guardia M. Testosterone and aggressiveness. Med Sci Monit. (2005) 11:RA136–45. doi: 10.5812/ijem.3661
17. Kudryavtseva NN, Bondar NP, Avgustinovich DF. Effects of repeated experience of aggression on the aggressive motivation and development of anxiety in male mice. Neurosci Behav Physiol. (2004) 34:721–30. doi: 10.1023/B:NEAB.0000036013.11705.25
18. Ma XC, Jiang D, Jiang WH, Wang F, Jia M, Wu J, et al. Social isolation-induced aggression potentiates anxiety and depressive-like behavior in male mice subjected to unpredictable chronic mild stress. PLoS ONE (2011) 6:e20955. doi: 10.1371/journal.pone.0020955
19. Neumann ID, Veenema AH, Beiderbeck DI. Aggression and anxiety: social context and neurobiological links. Front Behav Neurosci. (2010) 4:12. doi: 10.3389/fnbeh.2010.00012
20. Assari S, Caldwell CH, Zimmerman MA. Sex differences in the association between testosterone and violent behaviors. Trauma Mon. (2014) 19:26–32. doi: 10.5812/traumamon.18040
21. Harris JA, Rushton JP, Hampson E, Jackson DN. Salivary testosterone and self report aggressive and pro social personality characteristics in men and women. AggressBehav. (1996) 22:321–31. doi: 10.1002/(SICI)1098-2337(1996)22:5<321::AID-AB1>3.0.CO;2-M
22. Frye C, Edinger K, Lephart E, Walf A. 3alpha-androstanediol, but not testosterone, attenuates age-related decrements in cognitive, anxiety, and depressive behavior of male rats. Front Aging Neurosci. (2010) 2:15. doi: 10.3389/fnagi.2010.00015
23. Frye CA, Edinger K, Sumida K. Androgen administration to aged male mice increases anti-anxiety behavior and enhances cognitive performance. Neuropsychopharmacology (2008) 33:1049–61. doi: 10.1038/sj.npp.1301498
24. Jones BA, Watson NV. Spatial memory performance in androgen insensitive male rats. Physiol Behav. (2005) 85:135–41. doi: 10.1016/j.physbeh.2005.03.023
25. Frye CA, Petralia SM, Rhodes ME. Estrous cycle and sex differences in performance on anxiety tasks coincide with increases in hippocampal progesterone and 3 alpha, 5 alpha –THP. Pharmacol Biochem Behav. (2000). 67:587–96. doi: 10.1016/S0091-3057(00)00392-0
26. Boivin JR, Piekarski DJ, Wahlberg JK, Wilbrecht L. Age, sex, and gonadal hormones differently influence anxiety- and depression-related behavior during puberty in mice. Psychoneuroendocrinology (2017) 85:78–87. doi: 10.1016/j.psyneuen.2017.08.009
27. Goel N, Bale TL. Organizational and activational effects of testosterone on masculinization of female physiological and behavioral stress responses. Endocrinology (2008) 149:6399–405. doi: 10.1210/en.2008-0433
28. Ordyan NÉ, Pivina SG, Akulova VK. Effects of impaired testosterone metabolism during prenatal ontogenesis on the level of anxiety and behavior of rats in a novel environment. Neurosci Behav Physiol. (2007) 37:435–41. doi: 10.1007/s11055-007-0032-5
29. Brand T, Slob AK. Peripubertal castration of male rats, adult open field ambulation and partner preference behavior. Behav Brain Res. (1988) 30:111–7. doi: 10.1016/0166-4328(88)90141-6
30. Johnston AL, File SE. Sex differences in animal tests of anxiety. Physiol Behav. (1991) 49:245–50. doi: 10.1016/0031-9384(91)90039-Q
31. Primus RJ, Kellogg CK. Gonadal hormones during puberty organize environment-related social interaction in the male rat. Horm Behav. (1990) 24:311–23. doi: 10.1016/0018-506X(90)90012-M
32. Guo M, Wu CF, Liu W, Yang JY, Chen D. Sex difference in psychological behavior changes induced by long-term social isolation in mice. Prog Neuro Psychopharmacology Biol Psychiatry (2004) 28:115–21. doi: 10.1016/j.pnpbp.2003.09.027
33. Brown GR, Kulbarsh KD, Spencer KA, Duval C. Peri-pubertal exposure to testicular hormones organizes response to novel environments and social behaviour in adult male rats. Horm Behav. (2015) 73:135–41. doi: 10.1016/j.yhbeh.2015.07.003
34. Burke NN, Coppinger J, Deaver DR, Roche M, Finn DP, Kelly J. Sex differences and similarities in depressive- and anxiety-like behaviour in the Wistar-Kyoto rat. Physiol Behav. (2016) 167:28–34. doi: 10.1016/j.physbeh.2016.08.031
35. Näslund J, Studer E, Pettersson R, Hagsäter M, Nilsson S, Nissbrandt H, et al. Differences in anxiety-like behavior within a batch of wistar rats are associated with differences in serotonergic transmission, enhanced by acute sri administration, and abolished by serotonin depletion. Int J Neuropsychopharmacol. (2015) 18:1–9. doi: 10.1093/ijnp/pyv018
36. Schulz K, Molenda-Figueira H, Sisk C. Back to the future: the organizational–activational hypothesis adapted to puberty and adolescence. Horm Behav. (2009) 55:597–604. doi: 10.1016/j.yhbeh.2009.03.010.Back
37. Sisk CL, Zehr JL. Pubertal hormones organize the adolescent brain and behavior. Front Neuroendocrinol. (2005) 26:163–74. doi: 10.1016/j.yfrne.2005.10.003
38. Palanza P. Animal models of anxiety and depression: how are females different? Neurosci Biobehav Rev. (2001) 25:219–33. doi: 10.1016/S0149-7634(01)00010-0
39. Hu M, Richard JE, Maliqueo M, Kokosar M, Fornes R, Benrick A, et al. Maternal testosterone exposure increases anxiety-like behavior and impacts the limbic system in the offspring. Proc Natl Acad Sci USA. (2015) 112:201507514. doi: 10.1073/pnas.1507514112
40. Zuloaga DG, Jordan CL, Breedlove SM. The organizational role of testicular hormones and the androgen receptor in anxiety-related behaviors and sensorimotor gating in rats. Endocrinology (2011a) 152:1572–81. doi: 10.1210/en.2010-1016
41. Domonkos E, Borbélyová V, Csongová M, Bosý M, Kačmárová M, Ostatníková D, et al. Sex differences and sex hormones in anxiety-like behavior of aging rats. Horm Behav. (2017) 93:159–65. doi: 10.1016/j.yhbeh.2017.05.019
42. Borbélyová V, Domonkos E, Bábícková J, Tóthová L, Bos? M, Hodosy J, et al. No effect of testosterone on behavior in aged Wistar rats. Aging (2016) 8:2848–61. doi: 10.18632/aging.101096
43. Bowman RE, Maclusky NJ, Diaz SE, Zrull MC, Luine VN. Aged rats : sex differences and responses to chronic stress. Brain Res. (2006) 6:156–66. doi: 10.1016/j.brainres.2006.07.047
44. Baron-Cohen S, Knickmeyer RC, Belmonte MK. Sex differences in the brain: implications for explaining autism. Science (2005) 310:819–23. doi: 10.1126/science.1115455
45. Xu X-J, Zhang H-F, Shou X-J, Li J, Jing W-L, Zhou Y, et al. Prenatal hyperandrogenic environment induced autistic-like behavior in rat offspring. Physiol Behav. (2015) 138:13–20. doi: 10.1016/j.physbeh.2014.09.014
46. Pivina SG, Akulova VK, Ordyan NÉ. Characteristics of behavior and stress reactivity of the hypophyseal-adrenocortical system in rats with prenatal inhibition of testosterone metabolism. Neurosci Behav Physiol. (2007) 37:53–8. doi: 10.1007/s11055-007-0149-6
47. Leret ML, Molina-Holgado F, González MI. The effect of perinatal exposure to estrogens on the sexually dimorphic response to novelty. Physiol Behav. (1994) 55:371–3. doi: 10.1016/0031-9384(94)90148-1
48. Lucion AB, Charchat H, Pereira GA, Rasia-Filho AA. Influence of early postnatal gonadal hormones on anxiety in adult male rats. Physiol Behav. (1996) 60:1419–23.
49. Kellogg CK, Lundin A. Brain androgen-inducible aromatase is critical for adolescent organization of environment-specific social interaction in male rats. Horm Behav. (1999) 35:155–62. doi: 10.1006/hbeh.1998.1508
50. Morris JS, Weil ZM, Nelson RJ. Sexual experience and testosterone during adolescence alter adult neuronal morphology and behavior. Horm Behav. (2013) 64:454–60. doi: 10.1016/j.yhbeh.2013.08.001
51. Carrier N, Saland SK, Duclot F, He H, Mercer R. The anxiolytic and antidepressant-like effects of testosterone and estrogen in gonadectomized male rats. Biol Psychiatry (2015) 78:259–69. doi: 10.1016/j.biopsych.2014.12.024
52. Edinger KL, Frye CA. Intrahippocampal administration of an androgen receptor antagonist, flutamide, can increase anxiety-like behavior in intact and DHT-replaced male rats. Horm Behav. (2006) 50:216–22. doi: 10.1016/j.yhbeh.2006.03.003
53. Fedotova J, Hritcu L. Testosterone promotes anxiolytic-like behavior in gonadectomized male rats via blockade of the 5-HT1Areceptors. Gen Comp Endocrinol. (2017) 254:14–21. doi: 10.1016/j.ygcen.2017.09.006
54. Fernández-Guasti A, Martínez-Mota L. Anxiolytic-like actions of testosterone in the burying behavior test: Role of androgen and GABA-benzodiazepine receptors. Psychoneuroendocrinology (2005) 30:762–70. doi: 10.1016/j.psyneuen.2005.03.006
55. Frye CA, Seliga AM. Testosterone increases analgesia, anxiolysis, and cognitive performance of male rats. Cogn Affect Behav Neurosci. (2001) 1:371–81.
56. Khakpai F. The effect of opiodergic system and testosterone on anxiety behavior in gonadectomized rats. Behav Brain Res. (2014) 263:9–15. doi: 10.1016/j.bbr.2014.01.013
57. Näslund J, Studer E, Johansson E, Eriksson E. Effects of gonadectomy and serotonin depletion on inter-individual differences in anxiety-like behaviour in male Wistar rats. Behav Brain Res. (2016) 308:160–5. doi: 10.1016/j.bbr.2016.04.015
58. Nikmahzar E, Jahanshahi M, Ghaemi A, Naseri GR, Moharreri AR, Lotfinia AA. Hippocampal serotonin-2A receptor-immunoreactive neurons density increases after testosterone therapy in the gonadectomized male mice. Anat Cell Biol. (2016) 49:259. doi: 10.5115/acb.2016.49.4.259
59. Edinger KL, Frye CA. Testosterone's anti-anxiety and analgesic effects may be due in part to actions of its 5alpha-reduced metabolites in the hippocampus. Psychoneuroendocrinology (2005) 30:418–30. doi: 10.1016/j.psyneuen.2004.11.001
60. Edinger KL, Frye CA. Testosterone's analgesic, anxiolytic, and cognitive-enhancing effects may be due in part to actions of its 5alpha-reduced metabolites in the hippocampus. Behav Neurosci. (2004) 118:1352–64. doi: 10.1037/0735-7044.118.6.1352
61. Filova B, Malinova M, Babickova J, Tothova L, Ostatnikova D, Celec P, et al. Effects of testosterone and estradiol on anxiety and depressive-like behavior via a non-genomic pathway. Neurosci Bull. (2015) 31:288–96. doi: 10.1007/s12264-014-1510-8
62. Frye CA, Edinger KL. Testosterone's metabolism in the hippocampus may mediate its anti-anxiety effects in male rats. Pharmacol Biochem Behav. (2004) 78:473–81. doi: 10.1016/j.pbb.2004.04.019
63. Osborne DM, Edinger K, Frye CA. Chronic administration of androgens with actions at estrogen receptor beta have anti-anxiety and cognitive-enhancing effects in male rats. Age (2009) 31:191–8. doi: 10.1007/s11357-009-9114-3
64. Aikey JL, Nyby JG, Anmuth DM, James PJ. Testosterone rapidly reduces anxiety in male house mice (Mus musculus). Horm Behav. (2002) 42:448–60. doi: 10.1006/hbeh.2002.1838
65. Buddenberg TE, Komorowski M, Ruocco LA, Silva MA, Topic B. Attenuating effects of testosterone on depressive-like behavior in the forced swim test in healthy male rats. Brain Res Bull. (2009) 79:182–6. doi: 10.1016/j.brainresbull.2009.02.008
66. Hodosy J, Zelmanová D, Majzúnová M, Filová B, Malinová M, Ostatníková D, et al. The anxiolytic effect of testosterone in the rat is mediated via the androgen receptor. Pharmacol Biochem Behav. (2012) 102:191–5. doi: 10.1016/j.pbb.2012.04.005
67. Durdiakova J, Ostatnikova D, Celec P. Testosterone and its metabolites–modulators of brain functions. Acta Neurobiol Exp. (2011) 71:434–54.
68. Frye CA, Lacey EH. Posttraining androgens' enhancement of cognitive performance is temporally distinct from androgens' increases in affective behavior. Cogn Affect Behav Neurosci. (2001) 1:172–82. doi: 10.3758/CABN.1.2.172
69. Edinger KL, Frye CA. Sexual experience of male rats influences anxiety-like behavior and androgen levels. Physiol Behav. (2007) 92:443–53. doi: 10.1016/j.physbeh.2007.04.018
70. Frye CA, Koonce CJ, Edinger KL, Osborne DM, Walf AA. Androgens with activity at estrogen receptor beta have anxiolytic and cognitive-enhancing effects in male rats and mice. Horm Behav. (2008) 54:726–34. doi: 10.1016/j.yhbeh.2008.07.013
71. Pak TR, Chung WCJ, Lund TD, Hinds LR, Clay CM, Handa RJ. The androgen metabolite, 5α-androstane-3β, 17β-diol, is a potent modulator of estrogen receptor-beta1-mediated gene transcription in neuronal cells. Endocrinology (2005) 146:147–55. doi: 10.1210/en.2004-0871
72. Brinkmann AO. Molecular mechanisms of androgen action–a historical perspective. Methods Mol Biol. (2011) 776:3–24. doi: 10.1007/978-1-61779-243-4_1.
73. Chen CV, Brummet JL, Lonstein JS, Jordan CL, Breedlove SM. New knockout model confirms a role for androgen receptors in regulating anxiety-like behaviors and HPA response in mice. Horm Behav. (2014) 65:211–8. doi: 10.3851/IMP2701.Changes
74. Hamson DK, Jones BA, Csupity AS, Ali FM, Watson NV. Androgen insensitive male rats display increased anxiety-like behavior on the elevated plus maze. Behav Brain Res. (2014) 259:158–63. doi: 10.1016/j.bbr.2013.11.021
75. Zuloaga DG, Poort JE, Jordan CL, Breedlove SM. Male rats with the testicular feminization mutation of the androgen receptor display elevated anxiety-related behavior and corticosterone response to mild stress. Horm Behav. (2011) 60:380–8. doi: 10.1016/j.yhbeh.2011.07.008
76. Zuloaga DG, Morris JA, Jordan CL, Breedlove SM. Mice with the testicular feminization mutation demonstrate a role for androgen receptors in the regulation of anxiety-related behaviors and the hypothalamic-pituitary-adrenal axis. Horm Behav. (2008) 54:758–66. doi: 10.1016/j.yhbeh.2008.08.004
77. Chen CV, Brummet JL, Jordan CL, Breedlove SM. Down, but not out : partial elimination of androgen receptors in the male mouse brain does not affect androgenic regulation of anxiety or HPA activity (2015) 157:764–73. doi: 10.1210/en.2015-1417
78. Jayaraman A, Christensen A, Moser VA, Vest RS, Miller CP, Hattersley G, et al. Selective androgen receptor modulator RAD140 is neuroprotective in cultured neurons and kainate-lesioned male rats. Endocrinology (2014) 155:1398–406. doi: 10.1210/en.2013-1725
79. Akita K, Harada K, Ichihara J, Takata N, Takahashi Y, Saito K. A novel selective androgen receptor modulator, NEP28, is efficacious in muscle and brain without serious side effects on prostate. Eur J Pharmacol. (2013) 720:107–14. doi: 10.1016/j.ejphar.2013.10.042
80. Hengevoss J, Piechotta M, Müller D, Hanft F, Parr MK, Schänzer W, et al. Combined effects of androgen anabolic steroids and physical activity on the hypothalamic-pituitary-gonadal axis. J Steroid Biochem Mol Biol. (2015) 150:86–96. doi: 10.1016/j.jsbmb.2015.03.003
81. Kudwa AE, López FJ, McGivern RF, Handa RJ. A selective androgen receptor modulator enhances male-directed sexual preference, proceptive behavior, and lordosis behavior in sexually experienced, but not sexually naive, female rats. Endocrinology (2010) 151:2659–68. doi: 10.1210/en.2009-1289
82. Miner JN, Chang W, Chapman MS, Finn PD, Mei HH, López FJ, et al. An orally active selective androgen receptor modulator is efficacious on bone, muscle, and sex function with reduced impact on prostate. Endocrinology (2007) 148:363–73. doi: 10.1210/en.2006-0793
83. Morimoto M, Amano Y, Oka M, Harada A, Fujita H, Hikichi Y, et al. Amelioration of sexual behavior and motor activity deficits in a castrated rodent model with a selective androgen receptor modulator SARM-2f. PLoS ONE (2017) 12:e0189480. doi: 10.1371/journal.pone.0189480
84. Bitran D, Kellogg CK, Hilvers RJ. Treatment with an anabolic-androgenic steroid affects anxiety-related behavior and alters the sensitivity of cortical GABAA receptors in the rat. Horm Behav. (1993) 27:568–83. doi: 10.1006/hbeh.1993.1041
85. Roohbakhsh A, Moghaddam AH, Delfan KM. Anxiolytic-like effect of testosterone in male rats: GABAC receptors are not involved. Iran J Basic Med Sci. (2011) 14:376–82. doi: 10.22038/ijbms.2011.5027
86. Gutiérrez-García AG, Contreras CM, Vásquez-Hernández DI, Molina-Jiménez T, Jacome-Jacome E. Testosterone reduces cumulative burying in female Wistar rats with minimal participation of estradiol. Pharmacol Biochem Behav. (2009) 93:406–12. doi: 10.1016/j.pbb.2009.06.002
87. Zhang XW, Liu ZH, Hu XW, Yuan YQ, Bai WJ, Wang XF, et al. Androgen replacement therapy improves psychological distress and health-related quality of life in late onset hypogonadism patients in Chinese population. Chin. Med. J. (2012) 125:3806–10. doi: 10.3760/cma.j.issn.0366-6999.2012.21.011
88. Egashira N, Koushi E, Okuno R, Shirakawa A, Mishima K, Iwasaki K, et al. Depression-like behavior and reduced plasma testosterone levels in the senescence-accelerated mouse. Behav Brain Res. (2010) 209:142–7. doi: 10.1016/j.bbr.2010.01.030
89. Arfa-Fatollahkhani P, Nahavandi A, Abtahi H, Anjidani S, Borhani S, Jameie SB, et al. The effect of luteinizing hormone reducing agent on anxiety and novel object recognition memory in gonadectomized rats. Basic Clin Neurosci. (2017) 8:113–20. doi: 10.18869/nirp.bcn.8.2.113
90. Hamezah HS, Durani LW, Ibrahim NF, Yanagisawa D, Kato T, Shiino A, et al. Volumetric changes in the aging rat brain and its impact on cognitive and locomotor functions. Exp Gerontol. (2017) 99:69–79. doi: 10.1016/j.exger.2017.09.008
Keywords: anxiety-like behavior, perinatal, adolescence, puberty, aging
Citation: Domonkos E, Hodosy J, Ostatníková D and Celec P (2018) On the Role of Testosterone in Anxiety-Like Behavior Across Life in Experimental Rodents. Front. Endocrinol. 9:441. doi: 10.3389/fendo.2018.00441
Received: 31 January 2018; Accepted: 17 July 2018;
Published: 06 August 2018.
Edited by:
Ishwar Parhar, Monash University Malaysia, MalaysiaReviewed by:
Jacob Raber, Oregon Health and Science University, United StatesCarla Mucignat, Università degli Studi di Padova, Italy
Copyright © 2018 Domonkos, Hodosy, Ostatníková and Celec. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter Celec, cGV0ZXJjZWxlY0BnbWFpbC5jb20=