 Reidun Aesoy
Reidun Aesoy Colin D. Clyne
Colin D. Clyne Ashwini L. Chand
Ashwini L. Chand- 1Cancer Drug Discovery, Hudson Institute of Medical Research, Melbourne, VIC, Australia
- 2Department of Biomedicine, University of Bergen, Bergen, Norway
- 3Department of Molecular and Translational Science, Monash University, Clayton, VIC, Australia
- 4Cancer and Inflammation Laboratory, Olivia Newton-John Cancer Research Institute, Melbourne, VIC, Australia
- 5School of Cancer Medicine, La Trobe University, Melbourne, VIC, Australia
There is emerging evidence asserting the importance of orphan nuclear receptors (ONRs) in cancer initiation and progression. In breast cancer, there is a lot unknown about ONRs in terms of their expression profile and their transcriptional targets in the various stages of tumor progression. With the classification of breast tumors into distinct molecular subtypes, we assess ONR expression in the different breast cancer subtypes and with patient outcomes. Complementing this, we review evidence implicating ONR-dependent molecular pathways in breast cancer progression to identify candidate ONRs as potential prognostic markers and/or as therapeutic targets.
Introduction
Breast cancer is the most common cancer in women worldwide. Nearly 1.7 million new cases diagnosed in 2012 accounted for 12% of all new cancer cases and 25% of all cancers in women (1). The incidence of breast cancer is expected to increase primarily due to changes to the demographic with increasing aging populations and obesity, key risk factors for breast cancer. While patients with primary breast cancer are treated successfully, it is in the treatment of recurrent and invasive tumors where there is a clear need for the development of new therapies. Here, we discuss the potential of members of the nuclear receptor (NR) family as druggable targets for the discovery of new treatments in breast cancer.
The growth of the primary tumor in the breast is predominantly dependent on estrogen (2), via its interaction with its target receptor, the estrogen receptor (ER), expressed in 75% of all breast cancers. Hence, endocrine therapy is currently the most effective adjuvant treatment for ER-positive breast cancer. These include selective ER modulators (SERMs), selective ER down-regulators (SERDs), and aromatase inhibitors (AIs) all of which target the estrogen action and biosynthesis pathways (3–5). Despite the success of endocrine treatments in breast cancer, patients often present with resistance to endocrine therapy, despite high tumor ER expression. Furthermore, approximately 70% of patients with advanced disease ultimately acquire resistance to endocrine therapy [reviewed by Musgrove and Sutherland (6)]. The mechanisms for de novo or acquired endocrine therapy resistance are still poorly understood and may involve mechanisms of crosstalk between ER and other cell signaling networks, e.g., the Human Epidermal Growth Factor Receptor (HER) pathways. The actions of ER, Progesterone Receptor (PR), and Androgen Receptor (AR) in mediating breast cancer growth are well described in the literature. However, often overlooked in this context are the other members of the NR superfamily that function as transcription factors in regulating steroidogenesis, proliferation, and invasion and migration properties of cancer cells. The focus of the current review is to interrogate the expression patterns of the orphan nuclear receptors (ONR) and “adopted ONRs” in breast cancer, identify correlations with patient outcomes, and review the literature for functional evidence that may identify new ONRs as potentially linked to breast cancer in a prognostic sense or potential therapeutic targets.
Orphan NRs, referred to as ONRs throughout this review (Table 1), form a subgroup within the NR superfamily (7). Unlike the NRs, that are ligand activated; the ONRs have been classified together due to the lack of known endogenous ligands or other interacting synthetic compounds and drugs. Table 1 provides an overview of the ONR and adopted ONRs along with the potential endogenous or synthetic ligands that have been identified. While functioning in a similar manner to NRs, the ONRs can dimerize and bind to NR response elements (consisting of two palindromic NR half sites) to mediate transcription. As with NRs, ONRs can also hetero-dimerize, however it is ONRs that bind to NR half sites as monomers. Within the ONRs, the term “adopted ONRs” describes ONRs for which endogenous ligand or synthetic compounds have now been identified and shown to modulate activity (7–9) (Table 1). Structural analysis of ONRs also demonstrates that certain NRs, such as NR5A1 and NR5A2 are held in a constitutively active confirmation to maintain ligand-independent activity (10–14). Furthermore, modulation of the activity of NRs and ONRs by co-regulator proteins such as Nuclear Receptor Co-activators (NCOAs 1–3, also termed steroid receptor co-activators or SRCs 1–3), form an essential component of functional modulation to dictate whether genes are actively transcribed or repressed (15–17). The identification of synthetic compounds to modulate ONR activity is testament to the druggability of this class of transcription factors making them an attractive target for drug development in cancer. Due to their druggability and wide-ranging cellular functions, there is considerable interest in identifying novel ligands for ONRs as a therapeutic tool.
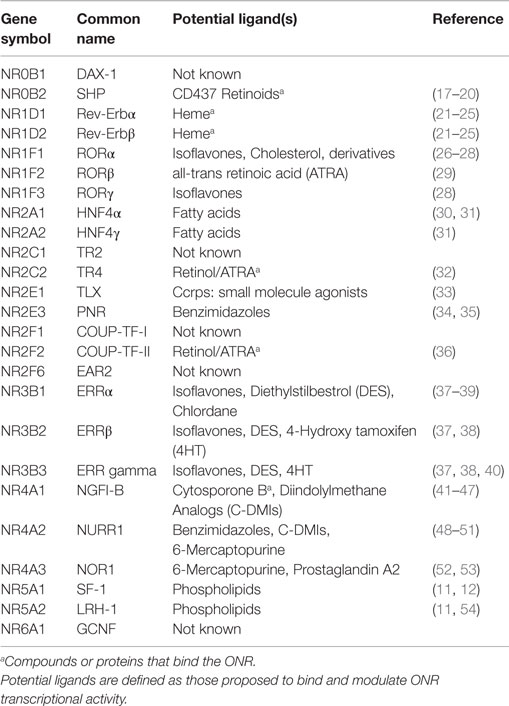
Table 1. Orphan nuclear receptors and proposed ligands.
Given the roles of ONRs in regulating transcription of genes involved in processes important for development, metabolism, immunity, angiogenesis, steroidogenesis, and fertility, their actions have also been implicated in multiple diseases including diverse types of cancers. Although not widely reported, there is emerging but clear evidence of ONR-dependent mechanisms in the regulation of tumor growth and progression. Recent work quantifying the expression profiles of the NR superfamily members in well curated tumor biopsy tissue and isolated cancer-associated fibroblast (CAFs) now allow a good insight into the possible actions of NRs in the development and progression of breast cancer (55–57).
We analyzed datasets available online to establish correlations between ONR expression and clinical parameters that define breast cancer outcomes. We also present an overview of the current understanding of ONR-related cellular mechanisms in breast cancer development.
Molecular Signatures Defining Breast Cancers
There are now several intrinsic molecular signatures acquired by genome-wide expression profiling of breast tumors that are used to stratify the heterogeneity of breast cancers (58–62). A clear delineation of these multi-gene signatures will ultimately allow personalized therapies. Breast tumors are classified into a variety of molecular subtypes with the four major subtypes being Luminal A, Luminal B, HER2-enriched, and basal-like tumors. Luminal A and Luminal B subtypes are both ER-positive tumors; Luminal A tumors being less aggressive than the Luminal B subtype, and more responsive to endocrine therapies (63–66). HER2-enriched tumors show amplification of the HER2 gene and these patients respond well to treatment that target HER2. High ER and PR levels, the lack of HER2 amplification in low-grade tumors and specific gene signatures, are now used as indicators of good effectiveness with endocrine therapy (61, 62). Basal-like tumors, also known as Triple-Negative Breast Cancers (TNBCs, lacking expression of ER, PR and HER2) are aggressive tumors with the only treatment options being surgery and chemotherapy.
In this vein, to address the question that other NRs could be used to further classify tumor subtypes, recently, three key studies provide a comprehensive expression profile of the NR superfamily and their interacting co-regulatory proteins in tumor and associated stroma in breast cancer (55–57). Based on these publications, akin to ER status, the expression of other NRs and ONRs could very likely provide additional prognostic power in the classification of breast tumor subtypes.
Correlation of ONR mRNA Transcript Expression with Patient Survival Outcomes
We used publically available Kaplan–Meier Plotter1 and BreastMark2 algorithms to correlate ONR gene expression to clinical outcomes within the datasets available within the two platforms as indicated in Tables 2 and 3. Kaplan–Meier Plot analysis was used specifically to assess ONR expression in relation to relapse free survival (RFS) in breast cancer patients; analysis also stratified to ER-positive and ER-negative status from gene expression data and survival information of 1,809 patients collated from GEO (Affymetrix HGU133A and HGU133 Plus 2.0 microarrays) EGA and TCGA databases (67). With the BreastMark algorithm, mRNA expression patterns of the genes of interest can be classified within breast cancer subtypes according to the PAM50 molecular signature (68). The database integrates gene expression and survival data from 26 datasets from 12 microarray platforms corresponding to 4,738 samples. We utilized the database to identify ONRs in which mRNA expression levels were positively or negatively correlated with disease progression in all breast cancer or stratified into the various subtypes.
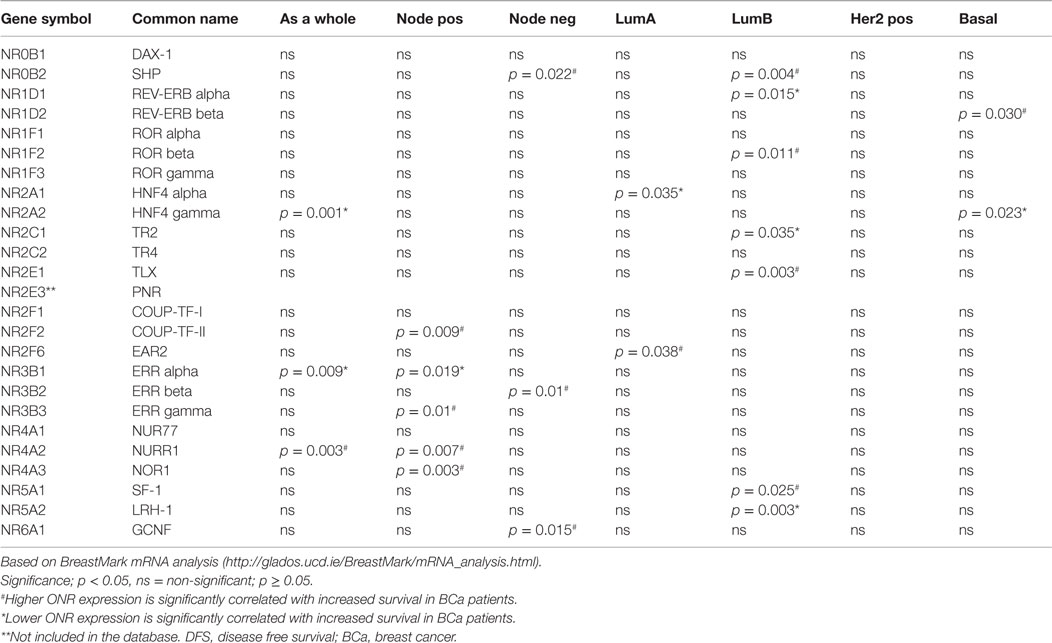
Table 2. Correlation of ONR expression to DFS in breast cancer patients.
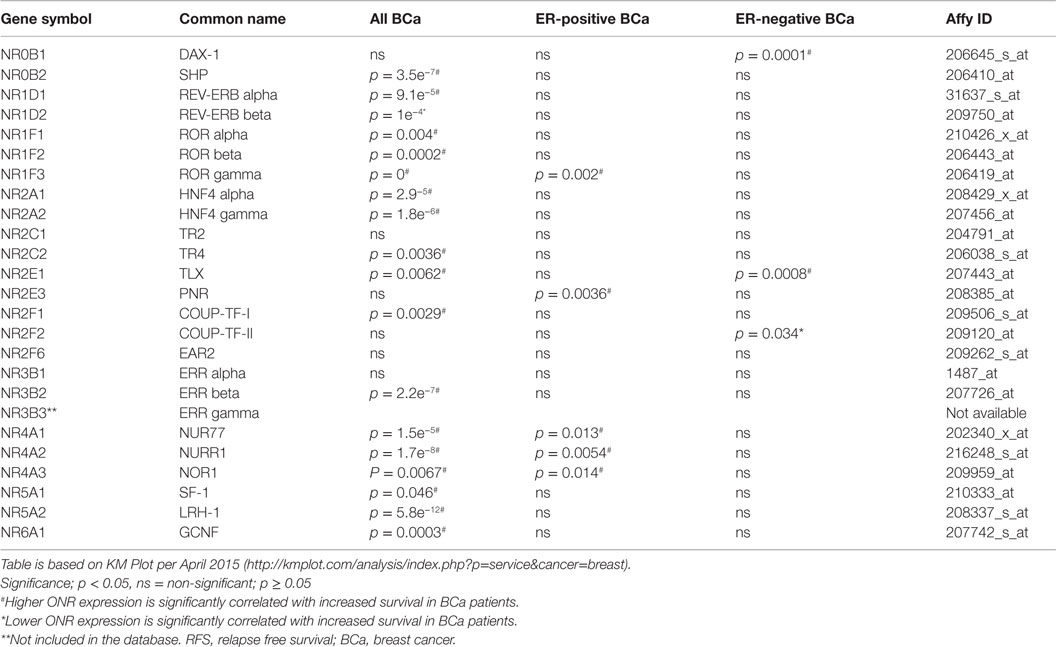
Table 3. Correlation of ONR expression to RFS in breast cancer patients.
ONRs Associated with the Luminal A Subtype
Lower expression of Hepatocyte Nuclear Factor 4 alpha (HNFα, NR2A1) and higher expression of V-erbA related protein (EAR2, NR2F6) were associated with increased DFS in patients with the Luminal A subtype of breast cancer (p = 0.035 and p = 0.038, respectively, Table 2). The functions of these two ONRs in the context of breast cancer are not defined as yet, although HNF4α, a key transcriptional regulator of hepatocyte differentiation and function in liver, acts as a tumor suppressor in the diethylinitrosamine-induced rat model of liver carcinogenesis, and inhibits epithelial-mesenchymal transition (EMT) (69). In a recent study focusing on expression patterns of genes associated with altered metabolism of breast cancer cells (using the TCGA database), the analysis of 556 transcription factor sequence motifs identified HNF4α-specific response elements as enriched in differentially co-expressed gene targets (70). Interestingly, this data suggest a potential function for this ONR in breast cancer cells in mediating tumor proliferation via the regulation of cellular metabolism, and warrants further investigation.
EAR2 is a member of the COUP-TF orphan subfamily and its established functions include negative regulation of renin and the luteinizing hormone receptor gene transcription and brain function including memory and learning (71–74). EAR2 expression is highly expressed in both ER-positive and ER-negative tumors relative to normal breast tissue (57). Additionally, EAR2 has been shown to directly interact with the aromatase promoter and down-regulate the expression of aromatase in breast cancer cell lines (75).
ONRs Associated with the Luminal B Subtype
Within the BreastMark criteria, reduced expression of REV-ERBα, TR2, LRH-1, and increased RORβ and SHP expression are significantly associated with greater DFS in patients in Luminal B tumors (p = 0.015, p = 0.035, p = 0.003, p = 0.011, p = 0.004, p = 0.025 and p = 0.003, respectively, Table 2). In contrast to Luminal A tumors, a greater number ONRs are associated with the Luminal B subtype, a more aggressive tumor and patients are often resistant to treatment. The functional implications of these expression profiles for each ONR associations are discussed in detail below.
REV-ERBα (also known as NR1D1) is a transcriptional repressor, abundantly expressed in adipose tissue and its functions are linked mainly to adipocyte differentiation, regulation of cell metabolism, and thermogenic responses (76) REV-ERBα lacks a transcriptional activation domain and represses target genes via its interaction with the transcriptional corepressor N-CoR. REV-ERBα has been shown to have a prosurvival function in HER2-positive breast cancer cells (77). However, recently it has been shown that REV-ERBβ (NR1D2) is also expressed in breast cancer cell lines and that its expression not correlated with HER2 or ER expression (9). Furthermore, the identification of a novel REV-ERBβ inhibitor, ARN5187, results in tumor cell cytotoxicity suggesting anticancer effects of REV-ERBβ suppression (9). We observed higher expression of REV-ERBβ as significantly associated with increased DFS in the basal cancer subtype (p = 0.03). Further determination of the protein expression and function of the two REV-ERB isoforms would allow a better understanding of the roles these ONRs play in the different breast cancer subtypes particularly with regards to HER2 status. Given its significant interaction with multiple co-regulators in ER-negative breast tumors, and a significant correlation to predict poorer outcome for patients, further investigation of the two REV-ERB isoforms in breast cancer is warranted.
Similarly to REV-ERB isoforms, TR2 (NR2C1) is a transcriptional repressor, and inhibits ERα-mediated transcription to regulate cell proliferation in breast cancer cells (78). In transgenic mouse models where TR2 is either over-expressed or down regulated, TR2 was identified as a key transcriptional repressor of GATA1 (79). Therefore, reduced TR2 in tumors may be the mechanism via which increased GATA1 promotes a more aggressive tumor phenotype in breast cancers (80). However, it is the lower expression of TR2 that was associated with increased survival in women with Luminal B tumors (Table 2, p = 0.035). Further investigation of the protein localization and correlation to TR2 mRNA expression profiles and the identification of breast cancer specific gene targets are required to further understand this observation.
In contrast to the REV-ERBs and TR2, Liver Receptor Homolog-1 (LRH-1, NR5A2) is a constitutive transcriptional activator, binding DNA as a monomer to promote transcription (11, 12, 14). The activity of LRH-1 was previously thought to be mainly under the control of co-regulators (14, 54, 81, 82, 83); however potential endogenous ligands identified include phosphatidylinositols, in particular, phosphatidylinositol-3-phosphate (PIP3) (11, 54), small molecule agonists (84) and antagonists (85).
Low LRH-1 expression significantly correlated with increased DFS in Luminal B tumor patients (Table 2, p = 0.003). Additionally, a strong correlation with co-regulator expression in ER-negative breast tumors suggests that increased LRH-1 activity may correlate with predicted poorer outcome for those patients (55). The expression of LRH-1 in both ER-positive and ER-negative tumors indicate that more functional evidence is require to determine its contribution in these varying tumor contexts. There is strong functional evidence linking LRH-1 to estrogen-regulated pathways in breast cancer tumor and associated stromal cell types [reviewed in Ref. (86)].
LRH-1 stimulates transcription of the aromatase gene, thereby promoting estrogen synthesis within CAFs (87–89). Given this strong association with tumor-associated estrogen production, most functional studies have been directed toward understanding the interplay between LRH-1 and estrogen-dependent proliferation pathways within breast tumors. While LRH-1 expression itself is under estrogen regulation, LRH-1 in a positive feedback manner induces estrogen-mediated cell proliferation (90). This is primarily via the co-operative regulation of gene-transcription of ERα and LRH-1 target genes (91–93). Both ERα and LRH-1 directly stimulate the transcription of one another, adding to the close functional relationship between the two NRs (90, 94). There are five transcript variants of LRH-1 that give rise to protein isoforms of three different sizes. There is some evidence that transcript variant four may be the active LRH-1 isoform in ER-positive breast cancer cells (94).
LRH-1 expression is detectable in both ER-positive and negative tumors (95). Despite low mRNA, LRH-1 protein levels are readily detected in ER-negative breast cancer cell lines and tumor tissue (56, 95–97) and this discrepancy is partly explained by increased transcript stability in ER-negative tumor cells (97). Knockdown of LRH-1 expression with shRNA constructs in ER-negative cells reduced cell migration and invasion in ER-negative breast cancer cells (98). The functional importance of LRH-1 in ER-negative cells and the ability of LRH-1 to regulate cell proliferation gene expression in endocrine-resistant breast cancer cells (91) as well as in the presence of SERDs (92) suggests a possible role in promoting tumor-aggressiveness. Given the breadth of knowledge on the tumor promoting effects of LRH-1 in various cancers, LRH-1 is a strong candidate as a therapeutic target. However, there is a need to demonstrate its contribution to mammary tumor initiation and formation using in vivo mouse models.
As age is the most common risk factor in breast cancer, it is interesting to note that the expression of RAR-related orphan receptor β (RORβ, also known as NR1F2) is greater in the postmenopausal breast compared to premenopausal tissue. However, its expression is significantly downregulated in ER-negative breast cancer implicating a potential role in the early stages of tumor growth (57). Above median expression of RORβ is correlated with greater DFS rates in patients with Luminal B cancers (Table 2, p = 0.011). Although its function is unclear, the ligands for RORβ may provide additional functional evidence of its roles in breast cancer cells.
ONRs Associated with Lymph Node-Positive Breast Cancer
Lymph node status is one of the most important predictors of breast cancer recurrence and survival. Data from the Surveillance, Epidemiology, and End Results (SEER) Program shows that women with lymph node negative tumors have a better prognosis compared to those with tumors in the lymph nodes.
In addition to overall DFS for all breast cancer cases, low ERRα and high NURR1 expression was associated with improved survival in patients with lymph node positive tumors (p = 0.019 and p = 0.007, respectively, Table 2). Structurally and functionally related to the ERs, ERRα (Estrogen-related receptor (ERR) alpha, also known as NR3B1) over-expression is associated with a poorer outcome in patients with ER-negative breast cancer (99). Functional in vitro studies in breast cancer cell lines show that the suppression of ERRα inhibits cellular proliferation and migration (100). Expression of ERRα positively correlates with HER2 status in breast tumors and ERRα transcriptional activity is positively modulated by EGFR/HER2 signaling in breast cancer cells. This suggests a role for ERRα in mediating the transition from ER-positive luminal toward more aggressive HER2-expressing tumor subtype (101, 102).
NURR1 (the NR-related 1 protein, also known as NR4A2), nerve growth factor I B (NGFIB, also known as Nur77 or NR4A1), and the neuron-derived orphan receptor 1 (NOR1, NR4A3) together constitute the NR4A subfamily of NRs (103–105). No endogenous ligands have been identified for these ONRs and their ability to transactivate genes appears to be constitutive. The NR4A receptors are however targeted by several hormones and xenobiotic compounds that affect the expression and/or activity of these stress early response genes (Table 1) (106). A recent study profiling all NRs in ER-positive and ER-negative breast tumors revealed that while most NR mRNA levels were significantly lower in breast cancer versus normal tissue, the expression of all of the NR4A family members (Nur77, NURR1, and NOR1) were significantly up-regulated in ER-positive breast cancer (57). Additionally in ER-negative breast tumors, when compared to normal breast tissue, NUR77 and NOR-1, together with EAR2, were found to have significantly increased (57).
In primary tumors, protein levels of NURR1 are inversely correlated with lymph node metastasis (107). This observation is mirrored in our analysis with the BreastMark dataset showing increased NURR1 mRNA expression is significantly correlated with increased survival in lymph node positive breast cancer. Data analysis from BreastMark and Kaplan–Meier Plotter indicate above median expression of NURR1 mRNA is associated with better prognosis and RFS in all breast cancer patients (107). However, functional in vivo experiments indicate the contrary. When NURR1 expression is knocked down by shRNA, growth of xenografts of highly invasive MDA-MB-231 and MDA-MB-468 breast cancer cells in athymic nude mice is significantly attenuated (107). Interestingly, high cytoplasmic expression of NURR1 is significantly correlated with advanced pathologic stage and higher tumor grade of bladder tumors as well as an increased number of distant metastases and decreased recurrence free survival (108). Thus, not only the expression level of NURR1 but the sub-cellular localization in which it is expressed may be of importance in its contribution toward tumor growth and metastatic potential.
Increased expression of NOR1 is correlated with better clinical outcomes in patients with lymph node positive tumors and ER-positive tumors (p = 0.003 and p = 0.014 Tables 2 and 3, respectively). Little is known about the actions of NOR1 in breast cancer, however in the ER-positive breast cancer cell line MCF7, NOR1 mRNA is up-regulated by a pro-apoptotic compound A23187 (109), shown to cause apoptotic cell death, suggesting a possible role in tumor cell apoptosis. NOR1 and NUR77 are thought to form heterodimers with NR retinoid X receptor RXR to modulate retinoic acid (RA) signaling (110, 111). Additionally in a positive feedback loop, RAs are involved in the positive regulation of NOR1 and NURR1 and the downregulation of NUR77 in breast cancer cells (112) correlating to decrease cell proliferation (113–115). Taken together these studies indicate that the NR4A family members may exert multiple functions in cancer, through their genomic functions in promoting tumor growth and survival, and, paradoxically, through non-genomic functions that may potentially exert tumor suppression by induction of apoptosis.
We observed an association of increased COUP-TFII expression with improved survival in patients with lymph node-positive tumors (Table 2, p = 0.009). The mRNA levels of the COUP transcription factors COUP-TFI (NR2F1) and COUP-TFII (NR2F2) were reported to be significantly decreased in both ER-positive and ER-negative breast cancer tissue compared to normal breast (57). COUP-TFII mRNA showed a discriminating role in the classification of breast cancer grades as it was more highly expressed in better-differentiated, lower-grade lesions (57). COUP-TFII was, in addition to the NR members TRβ, MR and PPARγ, reported to be a predictor of improved metastasis-free survival in tamoxifen-treated patients after adjustment for expression of ERα (57).
However, previously the amounts of protein of both these receptors have been reported to be higher in human breast cancer tissue than normal breast (116, 117). While the results are uncertain for COUP-TFI as it was only examined in a very small set of samples (116), COUP-TFII protein expression has been examined in a larger cohort of human breast cancers (117). Of 119 invasive ductal carcinoma of the breast, 59% stained positive for COUP-TFII in the nuclei of carcinoma cells (117). Higher nuclear expression of COUP-TFII was correlated with decreased disease-free and overall survival of the patient (117). Additionally, COUP-TFII was correlated to clinical stage (significantly higher expressed in stage IV tumors than stage I), histological grade (significantly higher expression with grade) and ERα status (significantly higher expressed in ERα positive than negative breast carcinomas) (117). Several groups have looked at the function of COUP-TFI and COUP-TFII in breast cancer cell lines. COUP-TFI may promote estrogen-independent transcriptional activity of ERα in breast cancer cells (116, 118). Furthermore, COUP-TFI mediates its actions through CXCL12/CXCR4 signaling. The over-expression of COUP-TFI results in down-regulation of CXCL12 and up-regulation of CXCR4 expression, to promote motility of MCF-7 cells (119). Furthermore, COUP-TFI mRNA expression is significantly higher in grade 1 (but not in grade 2 or 3) tumors than in normal breast tissue and that CXCR4 mRNA is significantly higher in cancerous tissue than in normal breast tissue and its expression is increasing with tumor grade, whereas transcripts of CXCL12 was significantly decreased in all the tumor grades compared to normal tissues (119).
ONRs Associated with Lymph Node-Negative Breast Cancer
Higher SHP, ERRβ and GCNF expression were associated with improved survival in patients with lymph node-negative tumors (p = 0.022, p = 0.01, p = 0.015, respectively, Table 2). Like Dax-1 (NROB1), the other member of this subgroup, short heterodimer partner (SHP, also termed NROB2) lacks a DNA-binding domain (DBD) and represses the action of NRs, including ERα through direct interaction with the receptor (120). SHP has been shown to inhibit aromatase expression by repressing LRH-1 activity on the promoter II element of Cyp19, the gene encoding aromatase (121). The actions of SHP in the repression of ER, LRH-1 and other NRs illustrates the importance of assessing the expression profile of this ONR in relation to other NRs to gage NR functionality. Above median expression of SHP also correlated with increased DFS in the luminal B subtype (Table 2, p = 0.004). These findings indicate that SHP may have a protective function in breast cancers and may be specifically so in lymph node-negative and luminal B subtype tumors. The expression and further characterization of this ONR are important due to its interactions with a range of NRs; whether SHP binds preferentially to ONRs over other co-regulators would also assist in defining its contribution to ONR-dependent actions in the cancer context.
Estrogen-related receptors are a subgroup of ONRs that have a strong homology with ER. We have previously discussed the roles of ERRα in lymph node positive tumors. Above median expression of ERRα is associated with greater survival in patients with lymph node-negative tumors. In other studies, the overexpression of ERRβ has been correlated with improved prognosis and longer relapse-free survival in breast cancer patients, and its mRNA levels inversely correlated with tumor cells in S-phase in patient-derived tumor samples (122). The functions of ERRs are tightly related to ER-mediated actions in breast cancer cells mainly due to the high degree of structural similarity to one another and to ER (123, 124), there is very little known about the details of ERR family members in this context. Recently though, in a detailed comparison of the ERRs on estradiol-stimulated ER transcription it was identified that it is only ERRβ that co-localized into the nucleus with ERα and with fluorescence resonance energy transfer revealed that ERRβ directly interacted with ERα to repress its transcriptional activity (125). The findings further suggest a unique inhibitory role for ERRβ in estrogen-dependent cellular function such as cancer cell proliferation via the regulation of cell cycle progression.
Similarly above median expression of Germ Cell Nuclear Factor Receptor (GCNF) is correlated with improved patient outcomes compared to those with below median expression (Table 2, p = 0.015). Initially identified as exclusively expressed in germ cells (126), GCNF is well characterized for its importance in embryonic development in regulating neural development and gastrulation (127) and oocyte function (128). Its unlikely role in breast cancer is suggested from a recent analysis of Affymetrix, Illumina and RNAseq microarray data to perform gene enrichment analysis identified shared common promoter motifs for GNCF in TNBC samples (129).
ONRs in ER-Positive and ER-Negative Breast Cancer
The Kaplan–Meier Plot analysis was used to assess ONR expression in relation to RFS in breast cancer patients; stratified to ER-positive and ER-negative tumors. Here, gene expression data and survival information of 1,809 patients have been collated from GEO (Affymetrix HGU133A and HGU133 + 2 microarrays) EGA and TCGA (130). Above median expression of 16 out of 24 ONRs was significantly correlated with increased RFS when analyzing all breast cancer patients in the database (Table 3, n = 3554). As an exception, lower expression of REV-ERBβ was correlated with increased patient RFS (p = 1.0e−4). In ER-positive breast cancer, above median expression of five ONRs including RORγ, PNR, Nurr77, Nurr1 and NOR1 was associated with better RFS (p = 0.002, p = 0.0036, p = 0.013, p = 0.0054 and p = 0.014, respectively, Table 3). In ER-negative breast cancer patients, higher expression of Dax-1 and TLX, and lower COUP-TFII expression were found to significantly correlate with increased RFS of (p = 0.0001, p = 0.0008, and p = 0.034, respectively, Table 3). This pattern or association with ER status may be important in identifying the regulatory mechanisms via which these ONRs are expressed; whether these ONRs modify/enhance the responses to endocrine therapies directed at ER such as SERMs are yet to be delineated.
Summary
The NR family of transcription factors plays diverse and important roles in development and the regulation of normal physiological functions in a tissue-specific manner. By understanding the contribution of ONRs in breast cancer subtypes, many novel mechanistic insights into tumor progression can be gleaned. From our data analysis, we observe complex expression patterns of various ONRs that could be selectively assessed in Luminal A, Luminal B, Lymph node positive and negative tumor types. Indeed within the specific subtype, the function of a particular ONR may possess a completely unique transcriptional imprint, therefore with varied effects on tumor proliferation and invasiveness; as is observed for AR, which can have pro- and anti-proliferative effects that is dependent on ER status of the breast cancer cell.
There is a lot that is unknown about ONRs in terms of their basic expression profile, mode of regulation of expression, cell subtype expression patterns, roles in normal mammary gland development processes, and transcriptional targets in the mammary gland and tumor scenarios. Our review attempted to identify “candidates” that were closely associated with breast cancer survival outcomes and whether molecular subtypes of breast cancer could be linked to ONR expression patterns. In addition to expression patterns, large-scale ChIP-seq experiments in well defined in breast cancer tissue samples and in in vivo tumor models will identify ONRs that are critical in this process. In addition, determining changes in ONR expression linked to key risk factors in breast cancer may also assist in understanding the mechanisms of this disease. The ligand-dependent actions of NRs provide extensive information that could potentially allow us to design new ligands for the ONRs in order to modulate their function. By the pharmacological targeting critical ONRs, we may in future identify effective therapies that could complement existing therapies in the treatment of breast cancer.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge the support of Victorian Government’s Opera-tional Infrastructure Support Program and the National Health and Medical Research Council of Australia Grants. ALC is supported by an NHMRC Career Development Award (ID 1062247).
Footnotes
References
1. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer (2015) 136:E359–86. doi: 10.1002/ijc.29210
2. Missmer SA, Eliassen AH, Barbieri RL, Hankinson SE. Endogenous estrogen, androgen, and progesterone concentrations and breast cancer risk among postmenopausal women. J Natl Cancer Inst (2004) 96:1856–65. doi:10.1093/jnci/djh336
3. Brodie AM, Dowsett M, Coombes RC. Aromatase inhibitors as new endocrine therapy for breast cancer. Cancer Treat Res (1988) 39:51–65. doi:10.1007/978-1-4613-1731-9_4
4. Katzenellenbogen BS, Katzenellenbogen JA. Estrogen receptor transcription and transactivation: estrogen receptor alpha and estrogen receptor beta: regulation by selective estrogen receptor modulators and importance in breast cancer. Breast Cancer Res (2000) 2:335–44. doi:10.1186/bcr78
5. Simpson ER, Dowsett M. Aromatase and its inhibitors: significance for breast cancer therapy. Recent Prog Horm Res (2002) 57:317–38. doi:10.1210/rp.57.1.317
6. Musgrove EA, Sutherland RL. Biological determinants of endocrine resistance in breast cancer. Nat Rev Cancer (2009) 9:631–43. doi:10.1038/nrc2713
7. Benoit G, Cooney A, Giguere V, Ingraham H, Lazar M, Muscat G, et al. International union of pharmacology. LXVI. Orphan nuclear receptors. Pharmacol Rev (2006) 58:798–836. doi:10.1124/pr.58.4.10
8. Chen T. Nuclear receptor drug discovery. Curr Opin Chem Biol (2008) 12:418–26. doi:10.1016/j.cbpa.2008.07.001
9. De Mei C, Ercolani L, Parodi C, Veronesi M, Vecchio CL, Bottegoni G, et al. Dual inhibition of REV-ERBbeta and autophagy as a novel pharmacological approach to induce cytotoxicity in cancer cells. Oncogene (2015) 34(20):2597–608. doi:10.1038/onc.2014.203
10. Benoit G, Malewicz M, Perlmann T. Digging deep into the pockets of orphan nuclear receptors: insights from structural studies. Trends Cell Biol (2004) 14:369–76. doi:10.1016/j.tcb.2004.05.007
11. Krylova IN, Sablin EP, Moore J, Xu RX, Waitt GM, MacKay JA, et al. Structural analyses reveal phosphatidyl inositols as ligands for the NR5 orphan receptors SF-1 and LRH-1. Cell (2005) 120:343–55. doi:10.1016/j.cell.2005.01.024
12. Li Y, Choi M, Cavey G, Daugherty J, Suino K, Kovach A, et al. Crystallographic identification and functional characterization of phospholipids as ligands for the orphan nuclear receptor steroidogenic factor-1. Mol Cell (2005) 17:491–502. doi:10.1016/j.molcel.2005.02.002
13. Li Y, Lambert MH, Xu HE. Activation of nuclear receptors: a perspective from structural genomics. Structure (2003) 11:741–6. doi:10.1016/S0969-2126(03)00133-3
14. Sablin EP, Krylova IN, Fletterick RJ, Ingraham HA. Structural basis for ligand-independent activation of the orphan nuclear receptor LRH-1. Mol Cell (2003) 11:1575–85. doi:10.1016/S1097-2765(03)00236-3
15. Lahusen T, Henke RT, Kagan BL, Wellstein A, Riegel AT. The role and regulation of the nuclear receptor co-activator AIB1 in breast cancer. Breast Cancer Res Treat (2009) 116:225–37. doi:10.1007/s10549-009-0405-2
16. Oh AS, Lahusen JT, Chien CD, Fereshteh MP, Zhang X, Dakshanamurthy S, et al. Tyrosine phosphorylation of the nuclear receptor coactivator AIB1/SRC-3 is enhanced by Abl kinase and is required for its activity in cancer cells. Mol Cell Biol (2008) 28:6580–93. doi:10.1128/MCB.00118-08
17. Dawson MI, Xia Z, Jiang T, Ye M, Fontana JA, Farhana L, et al. Adamantyl-substituted retinoid-derived molecules that interact with the orphan nuclear receptor small heterodimer partner: effects of replacing the 1-adamantyl or hydroxyl group on inhibition of cancer cell growth, induction of cancer cell apoptosis, and inhibition of SRC homology 2 domain-containing protein tyrosine phosphatase-2 activity. J Med Chem (2008) 51:5650–62. doi:10.1021/jm800456k
18. Dawson MI, Xia Z, Liu G, Ye M, Fontana JA, Farhana L, et al. An adamantyl-substituted retinoid-derived molecule that inhibits cancer cell growth and angiogenesis by inducing apoptosis and binds to small heterodimer partner nuclear receptor: effects of modifying its carboxylate group on apoptosis, proliferation, and protein-tyrosine phosphatase activity. J Med Chem (2007) 51(18):2622–39. doi:10.1021/jm800456k
19. Farhana L, Dawson MI, Dannenberg JH, Xu L, Fontana JA. SHP and Sin3A expression are essential for adamantyl-substituted retinoid-related molecule-mediated nuclear factor-kappaB activation, c-Fos/c-Jun expression, and cellular apoptosis. Mol Cancer Ther (2009) 8:1625–35. doi:10.1158/1535-7163.MCT-08-0964
20. Farhana L, Dawson MI, Leid M, Wang L, Moore DD, Liu G, et al. Adamantyl-substituted retinoid-related molecules bind small heterodimer partner and modulate the Sin3A repressor. Cancer Res (2007) 67:318–25. doi:10.1158/0008-5472.CAN-06-2164
21. Kojetin D, Wang Y, Kamenecka TM, Burris TP. Identification of SR8278, a synthetic antagonist of the nuclear heme receptor REV-ERB. ACS Chem Biol (2011) 6:131–4. doi:10.1021/cb1002575
22. Kumar N, Solt LA, Wang Y, Rogers PM, Bhattacharyya G, Kamenecka TM, et al. Regulation of adipogenesis by natural and synthetic REV-ERB ligands. Endocrinology (2010) 151:3015–25. doi:10.1210/en.2009-0800
23. Raghuram S, Stayrook KR, Huang P, Rogers PM, Nosie AK, McClure DB, et al. Identification of heme as the ligand for the orphan nuclear receptors REV-ERBalpha and REV-ERBbeta. Nat Struct Mol Biol (2007) 14:1207–13. doi:10.1038/nsmb1344
24. Solt LA, Wang Y, Banerjee S, Hughes T, Kojetin DJ, Lundasen T, et al. Regulation of circadian behaviour and metabolism by synthetic REV-ERB agonists. Nature (2012) 485:62–8. doi:10.1038/nature11030
25. Yin L, Wu N, Curtin JC, Qatanani M, Szwergold NR, Reid RA, et al. Rev-erbalpha, a heme sensor that coordinates metabolic and circadian pathways. Science (2007) 318:1786–9. doi:10.1126/science.1150179
26. Kallen J, Schlaeppi JM, Bitsch F, Delhon I, Fournier B. Crystal structure of the human RORalpha ligand binding domain in complex with cholesterol sulfate at 2.2 A. J Biol Chem (2004) 279:14033–8. doi:10.1074/jbc.M400302200
27. Kallen JA, Schlaeppi JM, Bitsch F, Geisse S, Geiser M, Delhon I, et al. X-ray structure of the hRORalpha LBD at 1.63 A: structural and functional data that cholesterol or a cholesterol derivative is the natural ligand of RORalpha. Structure (2002) 10:1697–707. doi:10.1016/S0969-2126(02)00912-7
28. Kojima H, Takeda Y, Muromoto R, Takahashi M, Hirao T, Takeuchi S, et al. Isoflavones enhance interleukin-17 gene expression via retinoic acid receptor-related orphan receptors alpha and gamma. Toxicology (2015) 329:32–9. doi:10.1016/j.tox.2015.01.007
29. Stehlin-Gaon C, Willmann D, Zeyer D, Sanglier S, Van Dorsselaer A, Renaud JP, et al. All-trans retinoic acid is a ligand for the orphan nuclear receptor ROR beta. Nat Struct Biol (2003) 10:820–5. doi:10.1038/nsb979
30. Hertz R, Magenheim J, Berman I, Bar-Tana J. Fatty acyl-CoA thioesters are ligands of hepatic nuclear factor-4alpha. Nature (1998) 392:512–6. doi:10.1038/33185
31. Wisely GB, Miller AB, Davis RG, Thornquest AD Jr, Johnson R, Spitzer T, et al. Hepatocyte nuclear factor 4 is a transcription factor that constitutively binds fatty acids. Structure (2002) 10:1225–34. doi:10.1016/S0969-2126(02)00829-8
32. Zhou XE, Suino-Powell KM, Xu Y, Chan CW, Tanabe O, Kruse SW, et al. The orphan nuclear receptor TR4 is a vitamin A-activated nuclear receptor. J Biol Chem (2011) 286:2877–85. doi:10.1074/jbc.M110.168740
33. Benod C, Villagomez R, Filgueira CS, Hwang PK, Leonard PG, Poncet-Montange G, et al. The human orphan nuclear receptor tailless (TLX, NR2E1) is druggable. PLoS One (2014) 9:e99440. doi:10.1371/journal.pone.0099440
34. Qin Q, Knapinska A, Dobri N, Madoux F, Chase P, Hodder P, et al. In pursuit of synthetic modulators for the orphan retina-specific nuclear receptor NR2E3. J Ocul Pharmacol Ther (2013) 29:298–309. doi:10.1089/jop.2012.0135
35. Wolkenberg SE, Zhao Z, Kapitskaya M, Webber AL, Petrukhin K, Tang YS, et al. Identification of potent agonists of photoreceptor-specific nuclear receptor (NR2E3) and preparation of a radioligand. Bioorg Med Chem Lett (2006) 16:5001–4. doi:10.1016/j.bmcl.2006.07.056
36. Kruse SW, Suino-Powell K, Zhou XE, Kretschman JE, Reynolds R, Vonrhein C, et al. Identification of COUP-TFII orphan nuclear receptor as a retinoic acid-activated receptor. PLoS Biol (2008) 6:e227. doi:10.1371/journal.pbio.0060227
37. Coward P, Lee D, Hull MV, Lehmann JM. 4-Hydroxytamoxifen binds to and deactivates the estrogen-related receptor gamma. Proc Natl Acad Sci U S A (2001) 98:8880–4. doi:10.1073/pnas.151244398
38. Suetsugi M, Su L, Karlsberg K, Yuan YC, Chen S. Flavone and isoflavone phytoestrogens are agonists of estrogen-related receptors. Mol Cancer Res (2003) 1(13):981–91.
39. Yang C, Chen S. Two organochlorine pesticides, toxaphene and chlordane, are antagonists for estrogen-related receptor alpha-1 orphan receptor. Cancer Res (1999) 59:4519–24.
40. Greschik H, Flaig R, Renaud JP, Moras D. Structural basis for the deactivation of the estrogen-related receptor gamma by diethylstilbestrol or 4-hydroxytamoxifen and determinants of selectivity. J Biol Chem (2004) 279:33639–46. doi:10.1074/jbc.M402195200
41. Chintharlapalli S, Burghardt R, Papineni S, Ramaiah S, Yoon K, Safe S. Activation of Nur77 by selected 1,1-Bis(3’-indolyl)-1-(p-substituted phenyl)methanes induces apoptosis through nuclear pathways. J Biol Chem (2005) 280:24903–14. doi:10.1074/jbc.M500107200
42. Cho SD, Lee SO, Chintharlapalli S, Abdelrahim M, Khan S, Yoon K, et al. Activation of nerve growth factor-induced B alpha by methylene-substituted diindolylmethanes in bladder cancer cells induces apoptosis and inhibits tumor growth. Mol Pharmacol (2010) 77:396–404. doi:10.1124/mol.109.061143
43. Cho SD, Lei P, Abdelrahim M, Yoon K, Liu S, Guo J, et al. 1,1-bis(3’-indolyl)-1-(p-methoxyphenyl)methane activates Nur77-independent proapoptotic responses in colon cancer cells. Mol Carcinog (2008) 47:252–63. doi:10.1002/mc.20378
44. Cho SD, Yoon K, Chintharlapalli S, Abdelrahim M, Lei P, Hamilton S, et al. Nur77 agonists induce proapoptotic genes and responses in colon cancer cells through nuclear receptor-dependent and nuclear receptor-independent pathways. Cancer Res (2007) 67:674–83. doi:10.1158/0008-5472.CAN-06-2907
45. Lee SO, Abdelrahim M, Yoon K, Chintharlapalli S, Papineni S, Kim K, et al. Inactivation of the orphan nuclear receptor TR3/Nur77 inhibits pancreatic cancer cell and tumor growth. Cancer Res (2010) 70:6824–36. doi:10.1158/0008-5472.CAN-10-1992
46. Liu JJ, Zeng HN, Zhang LR, Zhan YY, Chen Y, Wang Y, et al. A unique pharmacophore for activation of the nuclear orphan receptor Nur77 in vivo and in vitro. Cancer Res (2010) 70:3628–37. doi:10.1158/0008-5472.CAN-09-3160
47. Zhan Y, Du X, Chen H, Liu J, Zhao B, Huang D, et al. Cytosporone B is an agonist for nuclear orphan receptor Nur77. Nat Chem Biol (2008) 4:548–56. doi:10.1038/nchembio.106
48. Dubois C, Hengerer B, Mattes H. Identification of a potent agonist of the orphan nuclear receptor Nurr1. ChemMedChem (2006) 1:955–8. doi:10.1002/cmdc.200600078
49. Hintermann S, Chiesi M, von Krosigk U, Mathe D, Felber R, Hengerer B. Identification of a series of highly potent activators of the Nurr1 signaling pathway. Bioorg Med Chem Lett (2007) 17:193–6. doi:10.1016/j.bmcl.2006.09.062
50. Inamoto T, Papineni S, Chintharlapalli S, Cho SD, Safe S, Kamat AM. 1,1-Bis(3’-indolyl)-1-(p-chlorophenyl)methane activates the orphan nuclear receptor Nurr1 and inhibits bladder cancer growth. Mol Cancer Ther (2008) 7:3825–33. doi:10.1158/1535-7163.MCT-08-0730
51. Ordentlich P, Yan Y, Zhou S, Heyman RA. Identification of the antineoplastic agent 6-mercaptopurine as an activator of the orphan nuclear hormone receptor Nurr1. J Biol Chem (2003) 278:24791–9. doi:10.1074/jbc.M302167200
52. Kagaya S, Ohkura N, Tsukada T, Miyagawa M, Sugita Y, Tsujimoto G, et al. Prostaglandin A2 acts as a transactivator for NOR1 (NR4A3) within the nuclear receptor superfamily. Biol Pharm Bull (2005) 28:1603–7. doi:10.1248/bpb.28.1603
53. Wansa KD, Harris JM, Yan G, Ordentlich P, Muscat GE. The AF-1 domain of the orphan nuclear receptor NOR-1 mediates trans-activation, coactivator recruitment, and activation by the purine anti-metabolite 6-mercaptopurine. J Biol Chem (2003) 278:24776–90. doi:10.1074/jbc.M300088200
54. Ortlund EA, Lee Y, Solomon IH, Hager JM, Safi R, Choi Y, et al. Modulation of human nuclear receptor LRH-1 activity by phospholipids and SHP. Nat Struct Mol Biol (2005) 12:357–63. doi:10.1038/nsmb910
55. Doan TB, Eriksson NA, Graham D, Funder JW, Simpson ER, Kuczek ES, et al. Breast cancer prognosis predicted by nuclear receptor-coregulator networks. Mol Oncol (2014) 8:998–1013. doi:10.1016/j.molonc.2014.03.017
56. Knower KC, Chand AL, Eriksson N, Takagi K, Miki Y, Sasano H, et al. Distinct nuclear receptor expression in stroma adjacent to breast tumors. Breast Cancer Res Treat (2013) 142:211–23. doi:10.1007/s10549-013-2716-6
57. Muscat GE, Eriksson NA, Byth K, Loi S, Graham D, Jindal S, et al. Research resource: nuclear receptors as transcriptome: discriminant and prognostic value in breast cancer. Mol Endocrinol (2013) 27:350–65. doi:10.1210/me.2012-1265
58. Perou CM, Sorlie T, Eisen MB, van de Rijn M, Jeffrey SS, Rees CA, et al. Molecular portraits of human breast tumours. Nature (2000) 406:747–52. doi:10.1038/35021093
59. Sorlie T, Perou CM, Tibshirani R, Aas T, Geisler S, Johnsen H, et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc Natl Acad Sci U S A (2001) 98:10869–74. doi:10.1073/pnas.191367098
60. Sorlie T, Tibshirani R, Parker J, Hastie T, Marron JS, Nobel A, et al. Repeated observation of breast tumor subtypes in independent gene expression data sets. Proc Natl Acad Sci U S A (2003) 100:8418–23. doi:10.1073/pnas.0932692100
61. van ‘t Veer LJ, Dai H, van de Vijver MJ, He YD, Hart AA, Bernards R, et al. Expression profiling predicts outcome in breast cancer. Breast Cancer Res (2003) 5:57–8. doi:10.1186/bcr716
62. van de Vijver MJ, He YD, van’t Veer LJ, Dai H, Hart AA, Voskuil DW, et al. A gene-expression signature as a predictor of survival in breast cancer. N Engl J Med (2002) 347:1999–2009. doi:10.1056/NEJMoa021967
63. Badve S, Turbin D, Thorat MA, Morimiya A, Nielsen TO, Perou CM, et al. FOXA1 expression in breast cancer – correlation with luminal subtype A and survival. Clin Cancer Res (2007) 13:4415–21. doi:10.1158/1078-0432.CCR-07-0122
64. Camp JT, Elloumi F, Roman-Perez E, Rein J, Stewart DA, Harrell JC, et al. Interactions with fibroblasts are distinct in Basal-like and luminal breast cancers. Mol Cancer Res (2011) 9:3–13. doi:10.1158/1541-7786.MCR-10-0372
65. Cheang MC, Chia SK, Voduc D, Gao D, Leung S, Snider J, et al. Ki67 index, HER2 status, and prognosis of patients with luminal B breast cancer. J Natl Cancer Inst (2009) 101:736–50. doi:10.1093/jnci/djp082
66. Gatza ML, Silva GO, Parker JS, Fan C, Perou CM. An integrated genomics approach identifies drivers of proliferation in luminal-subtype human breast cancer. Nat Genet (2014) 46(10):1051–9. doi:10.1038/ng.3073
67. Gyorffy B, Schafer R. Meta-analysis of gene expression profiles related to relapse-free survival in 1,079 breast cancer patients. Breast Cancer Res Treat (2009) 118:433–41. doi:10.1007/s10549-008-0242-8
68. Madden SF, Clarke C, Gaule P, Aherne ST, O’Donovan N, Clynes M, et al. BreastMark: an integrated approach to mining publicly available transcriptomic datasets relating to breast cancer outcome. Breast Cancer Res (2013) 15:R52. doi:10.1186/bcr3444
69. Ning BF, Ding J, Yin C, Zhong W, Wu K, Zeng X, et al. Hepatocyte nuclear factor 4 alpha suppresses the development of hepatocellular carcinoma. Cancer Res (2010) 70:7640–51. doi:10.1158/0008-5472.CAN-10-0824
70. Reznik E, Sander C. Extensive decoupling of metabolic genes in cancer. PLoS Comput Biol (2015) 11:e1004176. doi:10.1371/journal.pcbi.1004176
71. Kummer MP, Hammerschmidt T, Martinez A, Terwel D, Eichele G, Witten A, et al. Ear2 deletion causes early memory and learning deficits in APP/PS1 mice. J Neurosci (2014) 34:8845–54. doi:10.1523/JNEUROSCI.4027-13.2014
72. Liu X, Huang X, Sigmund CD. Identification of a nuclear orphan receptor (Ear2) as a negative regulator of renin gene transcription. Circ Res (2003) 92:1033–40. doi:10.1161/01.RES.0000071355.82009.43
73. Warnecke M, Oster H, Revelli JP, Alvarez-Bolado G, Eichele G. Abnormal development of the locus coeruleus in Ear2(Nr2f6)-deficient mice impairs the functionality of the forebrain clock and affects nociception. Genes Dev (2005) 19:614–25. doi:10.1101/gad.317905
74. Zhang Y, Dufau ML. EAR2 and EAR3/COUP-TFI regulate transcription of the rat LH receptor. Mol Endocrinol (2001) 15:1891–905. doi:10.1210/mend.15.11.0720
75. Yang S, Fang Z, Suzuki T, Sasano H, Zhou J, Gurates B, et al. Regulation of aromatase P450 expression in endometriotic and endometrial stromal cells by CCAAT/enhancer binding proteins (C/EBPs): decreased C/EBPbeta in endometriosis is associated with overexpression of aromatase. J Clin Endocrinol Metab (2002) 87:2336–45. doi:10.1210/jc.87.5.2336
76. Gerhart-Hines Z, Feng D, Emmett MJ, Everett LJ, Loro E, Briggs ER, et al. The nuclear receptor Rev-erbalpha controls circadian thermogenic plasticity. Nature (2013) 503:410–3. doi:10.1038/nature12642
77. Kourtidis A, Jain R, Carkner RD, Eifert C, Brosnan MJ, Conklin DS. An RNA interference screen identifies metabolic regulators NR1D1 and PBP as novel survival factors for breast cancer cells with the ERBB2 signature. Cancer Res (2010) 70:1783–92. doi:10.1158/0008-5472.CAN-09-1550
78. Hu YC, Shyr CR, Che W, Mu XM, Kim E, Chang C. Suppression of estrogen receptor-mediated transcription and cell growth by interaction with TR2 orphan receptor. J Biol Chem (2002) 277:33571–9. doi:10.1074/jbc.M203531200
79. Tanabe O, Shen Y, Liu Q, Campbell AD, Kuroha T, Yamamoto M, et al. The TR2 and TR4 orphan nuclear receptors repress Gata1 transcription. Genes Dev (2007) 21:2832–44. doi:10.1101/gad.1593307
80. Boidot R, Vegran F, Jacob D, Chevrier S, Cadouot M, Feron O, et al. The transcription factor GATA-1 is overexpressed in breast carcinomas and contributes to survivin upregulation via a promoter polymorphism. Oncogene (2010) 29:2577–84. doi:10.1038/onc.2009.525
81. Suzuki T, Kasahara M, Yoshioka H, Umesono K, Morohashi K. LXXLL motifs in Dax-1 have target specificity for the orphan nuclear receptors Ad4BP/SF-1 and LRH-1. Endocr Res (2002) 28:537. doi:10.1081/ERC-120016835
82. Yazawa T, Inaoka Y, Okada R, Mizutani T, Yamazaki Y, Usami Y, et al. PPAR-gamma coactivator-1alpha regulates progesterone production in ovarian granulosa cells with SF-1 and LRH-1. Mol Endocrinol (2010) 24:485–96. doi:10.1210/me.2009-0352
83. Li Y, Choi M, Suino K, Kovach A, Daugherty J, Kliewer SA, et al. Structural and biochemical basis for selective repression of the orphan nuclear receptor liver receptor homolog 1 by small heterodimer partner. Proc Natl Acad Sci U S A (2005) 102:9505–10. doi:10.1073/pnas.0501204102
84. Whitby RJ, Dixon S, Maloney PR, Delerive P, Goodwin BJ, Parks DJ, et al. Identification of small molecule agonists of the orphan nuclear receptors liver receptor homolog-1 and steroidogenic factor-1. J Med Chem (2006) 49:6652–5. doi:10.1021/jm060990k
85. Benod C, Carlsson J, Uthayaruban R, Hwang P, Irwin JJ, Doak AK, et al. Structure-based discovery of antagonists of nuclear receptor LRH-1. J Biol Chem (2013) 288:19830–44. doi:10.1074/jbc.M112.411686
86. Lazarus KA, Wijayakumara D, Chand AL, Simpson ER, Clyne CD. Therapeutic potential of liver receptor homolog-1 modulators. J Steroid Biochem Mol Biol (2012) 130:138–46. doi:10.1016/j.jsbmb.2011.12.017
87. Bouchard MF, Taniguchi H, Viger RS. Protein kinase A-dependent synergism between GATA factors and the nuclear receptor, liver receptor homolog-1, regulates human aromatase (CYP19) PII promoter activity in breast cancer cells. Endocrinology (2005) 146:4905–16. doi:10.1210/en.2005-0187
88. Clyne CD, Kovacic A, Speed CJ, Zhou J, Pezzi V, Simpson ER. Regulation of aromatase expression by the nuclear receptor LRH-1 in adipose tissue. Mol Cell Endocrinol (2004) 215:39–44. doi:10.1016/j.mce.2003.11.001
89. Clyne CD, Speed CJ, Zhou J, Simpson ER. Liver receptor homologue-1 (LRH-1) regulates expression of aromatase in preadipocytes. J Biol Chem (2002) 277:20591–7. doi:10.1074/jbc.M201117200
90. Annicotte JS, Chavey C, Servant N, Teyssier J, Bardin A, Licznar A, et al. The nuclear receptor liver receptor homolog-1 is an estrogen receptor target gene. Oncogene (2005) 24(55):8167–75. doi:10.1038/sj.onc.1208950
91. Bianco S, Brunelle M, Jangal M, Magnani L, Gevry N. LRH-1 governs vital transcriptional programs in endocrine-sensitive and -resistant breast cancer cells. Cancer Res (2014) 74:2015–25. doi:10.1158/0008-5472.CAN-13-2351
92. Chand AL, Wijayakumara DD, Knower KC, Herridge KA, Howard TL, Lazarus KA, et al. The orphan nuclear receptor LRH-1 and ERalpha activate GREB1 expression to induce breast cancer cell proliferation. PLoS One (2012) 7:e31593. doi:10.1371/journal.pone.0031593
93. Lai CF, Flach KD, Alexi X, Fox SP, Ottaviani S, Thiruchelvam PT, et al. Co-regulated gene expression by oestrogen receptor alpha and liver receptor homolog-1 is a feature of the oestrogen response in breast cancer cells. Nucleic Acids Res (2013) 41:10228–40. doi:10.1093/nar/gkt827
94. Thiruchelvam PT, Lai CF, Hua H, Thomas RS, Hurtado A, Hudson W, et al. The liver receptor homolog-1 regulates estrogen receptor expression in breast cancer cells. Breast Cancer Res Treat (2011) 127:385–96. doi:10.1007/s10549-010-0994-9
95. Miki Y, Clyne CD, Suzuki T, Moriya T, Shibuya R, Nakamura Y, et al. Immunolocalization of liver receptor homologue-1 (LRH-1) in human breast carcinoma: possible regulator of insitu steroidogenesis. Cancer Lett (2006) 244:24–33. doi:10.1016/j.canlet.2005.11.038
96. Chand AL, Herridge KA, Thompson EW, Clyne CD. The orphan nuclear receptor LRH-1 promotes breast cancer motility and invasion. Endocr Relat Cancer (2010) 17:965–75. doi:10.1677/ERC-10-0179
97. Lazarus KA, Zhao Z, Knower KC, To SQ, Chand AL, Clyne CD. Oestradiol reduces liver receptor homolog-1 mRNA transcript stability in breast cancer cell lines. Biochem Biophys Res Commun (2013) 438:533–9. doi:10.1016/j.bbrc.2013.07.101
98. Chand AL, Herridge KA, Howard TL, Simpson ER, Clyne CD. Tissue-specific regulation of aromatase promoter II by the orphan nuclear receptor LRH-1 in breast adipose stromal fibroblasts. Steroids (2011) 76:741–4. doi:10.1016/j.steroids.2011.02.024
99. Ariazi EA, Kraus RJ, Farrell ML, Jordan VC, Mertz JE. Estrogen-related receptor alpha1 transcriptional activities are regulated in part via the ErbB2/HER2 signaling pathway. Mol Cancer Res (2007) 5:71–85. doi:10.1158/1541-7786.MCR-06-0227
100. Gerdin AK, Surve VV, Jonsson M, Bjursell M, Bjorkman M, Edenro A, et al. Phenotypic screening of hepatocyte nuclear factor (HNF) 4-gamma receptor knockout mice. Biochem Biophys Res Commun (2006) 349:825–32. doi:10.1016/j.bbrc.2006.08.103
101. Deblois G, Chahrour G, Perry MC, Sylvain-Drolet G, Muller WJ, Giguere V. Transcriptional control of the ERBB2 amplicon by ERRalpha and PGC-1beta promotes mammary gland tumorigenesis. Cancer Res (2010) 70:10277–87. doi:10.1158/0008-5472.CAN-10-2840
102. Deblois G, Hall JA, Perry MC, Laganiere J, Ghahremani M, Park M, et al. Genome-wide identification of direct target genes implicates estrogen-related receptor alpha as a determinant of breast cancer heterogeneity. Cancer Res (2009) 69:6149–57. doi:10.1158/0008-5472.CAN-09-1251
103. Law SW, Conneely OM, DeMayo FJ, O’Malley BW. Identification of a new brain-specific transcription factor, NURR1. Mol Endocrinol (1992) 6:2129–35. doi:10.1210/mend.6.12.1491694
104. Milbrandt J. Nerve growth factor induces a gene homologous to the glucocorticoid receptor gene. Neuron (1988) 1:183–8. doi:10.1016/0896-6273(88)90138-9
105. Ohkura N, Hijikuro M, Yamamoto A, Miki K. Molecular cloning of a novel thyroid/steroid receptor superfamily gene from cultured rat neuronal cells. Biochem Biophys Res Commun (1994) 205:1959–65. doi:10.1006/bbrc.1994.2900
106. Deutsch AJ, Angerer H, Fuchs TE, Neumeister P. The nuclear orphan receptors NR4A as therapeutic target in cancer therapy. Anticancer Agents Med Chem (2012) 12:1001–14. doi:10.2174/187152012803529619
107. Llopis S, Singleton B, Duplessis T, Carrier L, Rowan B, Williams C. Dichotomous roles for the orphan nuclear receptor NURR1 in breast cancer. BMC Cancer (2013) 13:139. doi:10.1186/1471-2407-13-139
108. Inamoto T, Czerniak BA, Dinney CP, Kamat AM. Cytoplasmic mislocalization of the orphan nuclear receptor Nurr1 is a prognostic factor in bladder cancer. Cancer (2010) 116:340–6. doi:10.1002/cncr.24737
109. Ohkubo T, Ohkura N, Maruyama K, Sasaki K, Nagasaki K, Hanzawa H, et al. Early induction of the orphan nuclear receptor NOR-1 during cell death of the human breast cancer cell line MCF-7. Mol Cell Endocrinol (2000) 162:151–6. doi:10.1016/S0303-7207(00)00222-7
110. Forman BM, Umesono K, Chen J, Evans RM. Unique response pathways are established by allosteric interactions among nuclear hormone receptors. Cell (1995) 81:541–50. doi:10.1016/0092-8674(95)90075-6
111. Perlmann T, Jansson L. A novel pathway for vitamin A signaling mediated by RXR heterodimerization with NGFI-B and NURR1. Genes Dev (1995) 9:769–82. doi:10.1101/gad.9.7.769
112. Maruyama K, Tsukada T, Bandoh S, Sasaki K, Ohkura N, Yamaguchi K. Retinoic acids differentially regulate NOR-1 and its closely related orphan nuclear receptor genes in breast cancer cell line MCF-7. Biochem Biophys Res Commun (1997) 231:417–20. doi:10.1006/bbrc.1997.6122
113. Lacroix A, Lippman ME. Binding of retinoids to human breast cancer cell lines and their effects on cell growth. J Clin Invest (1980) 65:586–91. doi:10.1172/JCI109703
114. Rubin M, Fenig E, Rosenauer A, Menendez-Botet C, Achkar C, Bentel JM, et al. 9-Cis retinoic acid inhibits growth of breast cancer cells and down-regulates estrogen receptor RNA and protein. Cancer Res (1994) 54:6549–56.
115. van der Burg B, van der Leede BM, Kwakkenbos-Isbrucker L, Salverda S, de Laat SW, van der Saag PT. Retinoic acid resistance of estradiol-independent breast cancer cells coincides with diminished retinoic acid receptor function. Mol Cell Endocrinol (1993) 91:149–57. doi:10.1016/0303-7207(93)90267-N
116. Le Dily F, Metivier R, Gueguen MM, Le Peron C, Flouriot G, Tas P, et al. COUP-TFI modulates estrogen signaling and influences proliferation, survival and migration of breast cancer cells. Breast Cancer Res Treat (2008) 110:69–83. doi:10.1007/s10549-007-9693-6
117. Nagasaki S, Suzuki T, Miki Y, Akahira J, Shibata H, Ishida T, et al. Chicken ovalbumin upstream promoter transcription factor II in human breast carcinoma: possible regulator of lymphangiogenesis via vascular endothelial growth factor-C expression. Cancer Sci (2009) 100:639–45. doi:10.1111/j.1349-7006.2008.01078.x
118. Metivier R, Gay FA, Hubner MR, Flouriot G, Salbert G, Gannon F, et al. Formation of an hER alpha-COUP-TFI complex enhances hER alpha AF-1 through Ser118 phosphorylation by MAPK. EMBO J (2002) 21:3443–53. doi:10.1093/emboj/cdf344
119. Boudot A, Kerdivel G, Lecomte S, Flouriot G, Desille M, Godey F, et al. COUP-TFI modifies CXCL12 and CXCR4 expression by activating EGF signaling and stimulates breast cancer cell migration. BMC Cancer (2014) 14:407. doi:10.1186/1471-2407-14-407
120. Johansson L, Thomsen JS, Damdimopoulos AE, Spyrou G, Gustafsson JA, Treuter E. The orphan nuclear receptor SHP inhibits agonist-dependent transcriptional activity of estrogen receptors ERalpha and ERbeta. J Biol Chem (1999) 274:345–53. doi:10.1074/jbc.274.1.345
121. Kovacic A, Speed CJ, Simpson ER, Clyne CD. Inhibition of aromatase transcription via promoter II by short heterodimer partner in human preadipocytes. Mol Endocrinol (2004) 18:252–9. doi:10.1210/me.2003-0211
122. Sengupta D, Bhargava DK, Dixit A, Sahoo BS, Biswas S, Biswas G, et al. ERRbeta signalling through FST and BCAS2 inhibits cellular proliferation in breast cancer cells. Br J Cancer (2014) 110:2144–58. doi:10.1038/bjc.2014.53
123. Giguere V. Orphan nuclear receptors: from gene to function. Endocr Rev (1999) 20:689–725. doi:10.1210/edrv.20.5.0378
124. Lu D, Kiriyama Y, Lee KY, Giguere V. Transcriptional regulation of the estrogen-inducible pS2 breast cancer marker gene by the ERR family of orphan nuclear receptors. Cancer Res (2001) 61(18):6755–61.
125. Tanida T, Matsuda KI, Yamada S, Hashimoto T, Kawata M. Estrogen-related receptor beta reduces the subnuclear mobility of estrogen receptor alpha and suppresses estrogen-dependent cellular function. J Biol Chem (2015) 290:12332–45. doi:10.1074/jbc.M114.619098
126. Chen F, Cooney AJ, Wang Y, Law SW, O’Malley BW. Cloning of a novel orphan receptor (GCNF) expressed during germ cell development. Mol Endocrinol (1994) 8:1434–44. doi:10.1210/me.8.10.1434
127. Wang Q, Cooney AJ. Revisiting the role of GCNF in embryonic development. Semin Cell Dev Biol (2013) 24:679–86. doi:10.1016/j.semcdb.2013.08.003
128. Lan ZJ, Gu P, Xu X, Jackson KJ, DeMayo FJ, O’Malley BW, et al. GCNF-dependent repression of BMP-15 and GDF-9 mediates gamete regulation of female fertility. EMBO J (2003) 22:4070–81. doi:10.1093/emboj/cdg405
129. Willis S, De P, Dey N, Long B, Young B, Sparano JA, et al. Enriched transcription factor signatures in triple negative breast cancer indicates possible targeted therapies with existing drugs. Meta Gene (2015) 4:129–41. doi:10.1016/j.mgene.2015.04.002
Keywords: orphan nuclear receptors, breast cancer, transcription factors, breast cancer subtypes, small molecule inhibitors
Citation: Aesoy R, Clyne CD and Chand AL (2015) Insights into orphan nuclear receptors as prognostic markers and novel therapeutic targets for breast cancer. Front. Endocrinol. 6:115. doi: 10.3389/fendo.2015.00115
Received: 04 June 2015; Accepted: 11 July 2015;
Published: 07 August 2015
Edited by:
Gabriella Castoria, Second University of Naples, ItalyCopyright: © 2015 Aesoy, Clyne and Chand. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ashwini L. Chand, ashwini.chand@onjcri.org.au