 Sven Sachs
Sven Sachs Stefan Eggmaier2
Stefan Eggmaier2 Daniel Madzia
Daniel Madzia- 1Abteilung Geowissenschaften, Naturkunde-Museum Bielefeld, Bielefeld, Germany
- 2Urwelt-Museum Oberfranken, Bayreuth, Germany
- 3Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland
Plesiosaurs are Mesozoic reptiles fully adapted to an aquatic lifestyle. Throughout their evolutionary history exceeding 140 million years plesiosaurs dispersed globally, achieved substantial diversity, occupied a variety of ecological niches, and experienced multiple faunal turnovers. Of those, the Early/Middle Jurassic transition event (∼175–171 Mya) has recently became of increased interest because it apparently profoundly affected all three major lineages of plesiosaurs. Once dominant Rhomaleosauridae started to vanish, while Pliosauridae and Plesiosauroidea diversified and gave rise to several clades that flourished for tens of millions of years. Here, we report exquisite, three-dimensionally preserved skeletons of a new plesiosaur from the Lower Jurassic of Germany. Franconiasaurus brevispinus gen. et sp. nov. lived during the late Toarcian (∼175 Mya), near the onset of the Early–Middle Jurassic turnover. Franconiasaurus displays an intriguing mixture of features, combining characters almost uniformly distributed among early plesiosaurs with those typically observed in later-diverging members of the clade. Phylogenetic analyses firmly place Franconiasaurus as the sister taxon to Cryptoclidia, bridging an evolutionary gap between early plesiosauroids, such as Plesiosaurus-like forms and microcleidids, and later-diverging representatives of the clade, such as cryptoclidids, leptocleidians, and elasmosaurids.
Introduction
Plesiosauroidea was a species-rich clade of plesiosaurs and one of their three major lineages (Benson and Druckenmiller, 2014). They existed from the Late Triassic to the Cretaceous/Paleogene mass extinction event (Benson and Druckenmiller, 2014; Madzia and Cau, 2020) and occupied a variety of ecological niches (e.g., O’Keefe et al., 2017). During their evolutionary history, plesiosauroids experienced several events of intense community changes. Of those, the Early/Middle Jurassic faunal turnover (∼175–171 Mya) has recently became of increased attention (e.g., Fischer et al., 2021; Sachs et al., 2023). During that time, plesiosauroids switched from early faunas, comprising Plesiosaurus-like forms and microcleidids to cryptoclidian faunas that had represented the globally dominant plesiosaurs since the Middle Jurassic until the end-Cretaceous (Benson and Druckenmiller, 2014; Madzia and Cau, 2020). However, the transition from “archaic”, Early Jurassic, to “derived”, Middle Jurassic and younger, plesiosauroid communities has been poorly understood because of the scarcity of plesiosaur fossil record from the Lower/Middle Jurassic boundary interval (see, e.g., Fischer et al., 2021; Sachs et al., 2022: Supplementary Table S2).
From the phylogenetic perspective, only a single taxon [Plesiopterys wildi from the lower Toarcian Posidonia Shale of Germany (O’Keefe, 2004)] has been reconstructed at this “transitional” position “between” microcleidids and cryptoclidians (e.g., Benson et al., 2012; Benson and Druckenmiller, 2014; Fischer et al., 2018; Madzia and Cau, 2020; Roberts et al., 2020). However, that taxon is based on an early juvenile individual that is additionally partly reconstructed and in a need of redescription. As such, its placement is doubtful.
In the present study we report two exquisite, three-dimensionally preserved plesiosaur skeletons from the Lower/Middle Jurassic boundary interval of Bavaria, Germany, that represent a new taxon of plesiosauroid, Franconiasaurus brevispinus gen. et sp. nov. Both specimens derive from the upper Toarcian (uppermost Lower Jurassic) Jurensismergel Formation and have been excavated by the Urwelt-Museum Oberfranken in a now-abandoned clay-pit in Mistelgau.
We present our first results of the study of these specimens; we introduce the new taxon, provide its initial description, illustrate it (Figures 1, 2), and reconstruct its phylogenetic placement (Figure 3).

FIGURE 1. Diagnostic skeletal elements of the type specimen of Franconiasaurus brevispinus gen. et sp. nov. (BT 011224.00). (A) main slab showing partly articulated skeleton in dorsal view. Posterior part of right mandibular ramus in (B) lateral and (C) dorsal aspect; anterior cervical vertebra (D) in articular and (E) lateral view; (F) posterior cervical vertebra in lateral view; (G) cervical rib in dorsolateral view; (H) dorsal vertebra in posterior view; (I) left scapula in medial view; (J) right coracoid in supposed dorsal view; (K) left humerus in dorsal view; (L) left femur with adjacent epipodials in dorsal view; and (M) left ilium in lateral view.

FIGURE 2. Overview of the referred specimen of Franconiasaurus brevispinus gen. et sp. nov. (BT 011241.00). (A) posterior cervical vertebra in posterior view; (B) tibia in supposed dorsal view; (C) anterior portions of coracoids in dorsal view; (D) left humerus in ventral view; (E) right humerus and radius in dorsal view. Scale bars equal 50 mm.
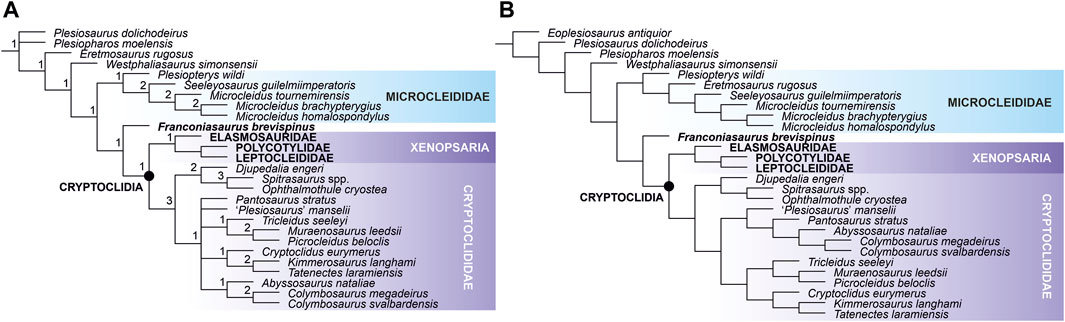
FIGURE 3. The phylogenetic placement of Franconiasaurus brevispinus gen. et sp. nov. showed on non-xenopsarian segment of Plesiosauroidea. (A) Reduced strict consensus tree reconstructed through parsimony analysis using equal weights. Numbers at nodes indicate Bremer support values. (B) A tree based on strict consensus trees reconstructed through weighted parsimony analyses with K set to 6, 9, and 12. The results of particular parsimony analyses with the implied weighting function differ in placements of certain taxa. However, the non-xenopsarian segments of Plesiosauroidea show the same topologies. The results of Symmetric Resampling are provided in Electronic Supplementary Material S3.
Franconiasaurus brevispinus gen. et sp. nov. (Figure 4) is a significant addition to the Early Jurassic vertebrate record in general because it helps filling an important gap in the evolutionary history of a major clade of Mesozoic aquatic tetrapods.

FIGURE 4. Life reconstruction of Franconiasaurus brevispinus gen. et sp. nov. (artwork by Joschua Knüppe).
Methods
Protocol for phylogenetic analyses
Our phylogenetic analyses were conducted using the dataset of Sachs et al. (2021) which is a substantially modified version of the data first assembled by Benson and Druckenmiller (2014). The taxon sampling was modified with addition of first-hand scores for Franconiasaurus brevispinus, obtained from BT 011224.00 (type specimen) and BT 011241.00 (referred specimen), both housed at the Urwelt-Museum Oberfranken, Bayreuth, Germany (BT), and with rescoring of Plesiopterys wildi based on preliminary assessment of the type specimen. In order to expand our sample of Early and Middle Jurassic plesiosaurs, we have additionally included the scores of the early-diverging plesiosauroid Plesiopharos moelensis (Puértolas-Pascual et al., 2021) and the early (non-thalassophonean) pliosaurids Arminisaurus schuberti and Cryonectes neustriacus (Sachs et al., 2023). As in Roberts et al. (2017), “Colymbosaurus trochantericus” was replaced by the two currently recognized Colymbosaurus species, C. megadeirus and C. svalbardensis; and Djupedalia engeri, Kimmerosaurus langhami, Muraenosaurus leedsi, and Spitrasaurus spp. were modified (see Roberts et al., 2017). Following Benson and Bowdler (2014), we have changed character 243 in Colymbosaurus spp. to 2. Finally, we have also added Ophthalmothule cryostea based on Roberts et al. (2020). The final version of the dataset comprised 130 operational taxonomic units (OTUs) and 270 characters; 67 of which were set as “additive” (=“ordered”) following Madzia et al. (2019). The character list and a TNT-executable matrix are provided in Electronic Supplementary Material S1.
Our analyses were performed using TNT 1.6 (Goloboff and Morales, 2023). Neusticosaurus pusillus was used as the outgroup. Four parsimony analyses were conducted; one based on equal weights only and three using the implied weighting function (Goloboff, 1993; Goloboff, 1995; Goloboff et al., 2008; Goloboff et al., 2018) with the concavity parameter (K) set to 6, 9, and 12. Before each analysis, we fixed the maximum number of most parsimonious trees to 200,000 trees (command “hold 200000;”). Our initial “New Technology” search involved 500 addition sequences and default settings activated for sectorial searches, ratchet, drift, and tree fusing. Then, we performed a “Traditional Search” with tree bisection-reconnection (TBR) branch-swapping on trees saved to RAM. For the parsimony analysis using equal weights, we calculated the Bremer support with TBR and retaining sub-optimal trees incorporating up to 3 additional steps. Nodal support for weighted parsimony analyses was determined using Symmetric Resampling with a “Traditional” search, 1,000 replicates, a default change probability (set at 33), and output expressed as frequency differences (GC).
Nomenclatural acts
This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the International Code of Zoological Nomenclature (ICZN). The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSIDs to the prefix https://zoobank.org/. The LSIDs are urn:lsid:zoobank.org:pub:90826206-FF90-4244-8C51-3768DA984249 for this publication, urn:lsid:zoobank.org:act:7F2F0C92-A50E-4690-B529-8218AC2C8B01 for the new genus Franconiasaurus, and urn:lsid:zoobank.org:act:AB189251-475E-4604-99C9-EF6A19F45565 for the new species F. brevispinus.
Systematic paleontology
Plesiosauria de Blainville, 1835.
Plesiosauroidea Gray, 1825.
Franconiasaurus gen. nov.
Type species—Franconiasaurus brevispinus sp. nov.
Etymology—After the region of Franconia (Franken in German), located in the northern part of Bavaria in southern Germany, where the specimens were found; and “σαῦρος” (sauros), Greek for “reptile”.
Diagnosis—As for the type and only species.
Franconiasaurus brevispinus sp. nov.
Type specimen—BT 011224.00, nearly complete, three-dimensionally preserved, and partly articulated skeleton.
Referred material—BT 011241.00, incomplete, largely disarticulated, three-dimensionally preserved skeleton.
Horizon and locality—The specimens were excavated in 2005 (BT 011241.00) and between 2014 and 2018 (BT 011224.00) by one of us (S.E.) in the lower section of the Jurensismergel Formation (Grammoceras thouarsense Zone), upper Toarcian, Lower Jurassic, in the Mistelgau fossil pit, Bayreuth District, Bavaria, Germany. For detailed information on the geological and stratigraphic settings see Electronic Supplementary Material S2.
Etymology—The name brevispinus refers to the low neural spines in the posterior cervical and dorsal vertebrae.
Diagnosis—A plesiosauroid diagnosed through the following unique combination of characters: surangular with transversely narrow dorsal side and convex lateral side, lacking a longitudinal lateral through; cervical centra amphicoelous, being wider than long/high; cervical zygapophyses narrower than centra, having planar facets and being not connected for most of their length; anterior cervical neural spines curve posterodorsally, posterior cervical neural spines inclined straight posterodorsally; posterior cervical and dorsal neural spines only moderately higher than long (height/length ratio approximately 1.6); cervical ribs with reduced anterior processes; dorsal process of scapula with slightly convex medial side, lacking a buttress like medial enforcement; long intercoracoid symphysis, coracoids being only slightly separated posteromedially; coracoid cornu does not extend to the level of the glenoid; ilium shaft straight, dorsal and ventral ends of ilium perpendicular to one another; femora and humeri subequal in length; propodials moderately elongate (length/width ratio 1.7); radius and tibia subquadratic with concave pre- and postaxial sides.
Description and comparison
A detailed anatomical description of Franconiasaurus is in preparation and will be published as part of a larger project elsewhere. Franconiasaurus is established based on two exquisite, three-dimensionally preserved specimens. The holotype (BT 011224.00) includes a largely complete skeleton which, however, lacks most of the skull. The referred specimen (BT 011241.00) represents a larger individual and comprises vertebrae, ribs, and some girdle and limb elements.
Part of the mandible is present in BT 011224.00 (Figures 1B, C). The preserved mandibular portions include a left dentary section, likely originating from the mid-section, and the posterior-most segments of both mandibular rami (Electronic Supplementary Material S3, Supplementary Figures S9A,B, S10). The dentary fragment bears 8 narrowly-spaced circular alveoli with the anterior and posterior alveoli being smaller than those from mid-section. Laterally, there are several large vascular foramina.
The surangular has a transversely narrow dorsal side (Figure 1C). On the medial side it lacks both a fossa and a longitudinal crest (Electronic Supplementary Material S3, Supplementary Figure S9C). Among Early Jurassic plesiosaurs, this condition is shared with the early pliosaurids Arminisaurus and Hauffiosaurus, as well as Plesiopterys and most microcleidids (O’Keefe, 2004; Großmann, 2007; Benson et al., 2011; Brown et al., 2013; Sachs and Kear, 2017). However, in many rhomaleosaurids a prominent crest accompanied by a deep surangular fossa is present on the medial side of the surangular (e.g., Vincent and Benson, 2012; Benson et al., 2015a). In some Early Jurassic plesiosaurs, such as Hauffiosaurus and most rhomaleosaurids, the lateral side of the mandible anterior to the glenoid additionally shows a well-formed longitudinal trough (Benson and Druckenmiller, 2014, Appendix 2, character 121). Such a through is absent in Franconiasaurus. Laterally, the preglenoid section of the mandible is gently convex (Figure 1B), resembling the condition in the Pliensbachian pliosaurids Arminisaurus and Cryonectes (Vincent et al., 2013; Sachs and Kear, 2017).
The articular surface of the glenoid fossa is only slightly dorsomedially-inclined and shorter than the retroarticular process (Figure 1B). The retroarticular process shows a slight inclination in posteromedial and posterodorsal direction; a condition shared with some pliosaurids, and plesiosauroids such as Plesiosaurus, Plesiopterys or cryptoclidids like Tricleidus or Kimmerosaurus (Benson and Druckenmiller, 2014, Appendix 2, characters 122 and 123). A relatively shorter retroarticular process is present in the Early Jurassic rhomaleosaurids Meyerasaurus and Rhomaleosaurus (Smith, 2007; Smith and Vincent, 2010).
On the medial side of the well preserved right mandibular fragment, a V-shaped posterior opening of the Meckelian canal is exposed (Electronic Supplementary Material S3, Supplementary Figure S9C). A lingual mandibular fenestra is not visible.
The axial skeleton of BT 011224.00 is largely preserved and partly articulated, comprising 24 cervical, 2 pectoral, 22 dorsal, 3 sacral, and 26 caudal vertebrae. Additional isolated posterior cervical, pectoral and dorsal vertebrae are present in BT 011241.00.
The cervical series in BT 011224.00 is almost complete, but the atlas-axis and some distal cervicals are missing. Judging from the small size of the cervical centrum preserved in the anteriormost position we think that only one or two post-axis cervicals are missing and that the neck comprised less than 30 cervical vertebrae.
All cervical centra are amphicoelous and wider than long and high (Figures 1D, E). The anterior cervicals are also longer than high, whereas the height and length of the posterior cervicals is about equal (Figure 1F). A similar condition is observable in cervicals of some rhomaleosaurids, such as Anningasaura and Stratesaurus or in the cryptoclidid Muraenosaurus (Andrews, 1910; Vincent and Benson, 2012; Benson et al., 2015a). Neurocentral sutures are completely fused in both specimens, supporting the adult ontogenetic stage for BT 011224.00 (sensu Brown, 1981). Ventrally, the cervical centra show flat and rounded midline ridges, separating two foramina subcentralia. The pre- and postzygapophyses are facing dorsomedially and are distinctly narrower than the centra. The zygapophyses are not connected for most of their length like in Microcleidus tournemirensis and M. homalospondylus (Bardet et al., 1999; Benson and Druckenmiller, 2014, Appendix 2, character 168) and have planar faces (Figure 1D).
The neural spines of the anterior cervical vertebrae curve posterodorsally, whereas those of the posterior cervicals are inclined straight posterodorsally (Figures 1E, F). This distinguishes Franconiasaurus from most Early Jurassic plesiosaurs with the exception of Attenborosaurus, Plesiosaurus, and Eoplesiosaurus (Benson and Druckenmiller, 2014; Appendix 2, characters 157 and 158).
The height/length ratio of the anterior neural spines varies between 1.4 and 1.5, whereas in the posterior cervical neural spines the ratio is 1.6–1.7. The neural spines are thus considerably shorter than those in the Toarcian microcleidids Seeleyosaurus and Microcleidus where the posterior cervical neural spines are more than twice as high as they are long (Dames, 1895; Großmann, 2006; Großmann, 2007; Vincent et al., 2019). A similar condition is observable in Plesiopterys (O’Keefe, 2004).
Both, the anterior and posterior cervical centra bear two co-joined and slightly posterolaterally-inclined rib facets (Figures 1D, F), a trait found in most Lower Jurassic plesiosauroids (Benson and Druckenmiller, 2014; Appendix 2, characters 160 and 161). The cervical ribs have a posteroventrally-oriented long axis and bear reduced anterior processes (Figure 1G), similar to the condition in the basal pliosaurid Arminisaurus (Sachs and Kear, 2017). In Plesiopterys both the anterior and posterior cervical rib processes are reduced (S. Sachs pers. obs. 2022) whereas most other Early Jurassic plesiosaurs have marked anterior and posterior cervical rib processes (Benson and Druckenmiller, 2014, Appendix 2, character 163).
All dorsal vertebrae have low neural arches with dorsolaterally-inclined transverse processes (Figure 1H). They are placed adjacent to the neural canal and bear a single rib facet. The dorsal neural spines are low, being only approximately 1.6 times as high as long. These neural spine proportions plus the absence of a strong anteroposterior constriction at the base of the dorsal neural spines distinguish Franconiasaurus from microcleidids (Benson and Druckenmiller, 2014). In Plesiopterys the dorsal neural spines are not constricted at the base, but more than twice as high as long (S. Sachs pers. obs. 2022). Dorsal and gastric ribs are present in both specimens.
The sacral vertebrae are preserved in articulation. The sacral ribs are placed adjacent to the centra, have a cylindrical shape and are only slightly expanded laterally (Electronic Supplementary Material S3, Supplementary Figure S9D).
The caudal centra have a subrectangular shape in articular view with a dished midsection in the articular surfaces (Electronic Supplementary Material S3, Supplementary Figure S9E). The length/height ratio in the proximal caudals varies between 0.75 and 0.83 and the caudal rib facets are placed dorsolaterally on the centra. Some isolated caudal ribs and hemapophyses are preserved in the holotype specimen. The hemapophyseal facets are situated on the posterior sides of the centra and do not markedly protrude ventrally. A similar morphology has been described for the caudal vertebrae of the rhomaleosaurids Eurycleidus, Rhomaleosaurus thorntoni, and “Trematospondylus” (Benson and Druckenmiller, 2014; Appendix 2, character 192; Smith and Benson, 2014; Sachs et al., 2022).
From the pectoral girdle, an almost complete but isolated coracoid and scapula, as well as several fragments, are preserved in BT 011224.00 and incomplete coracoids are likewise preserved in BT 011241.00 (Figures 1I, J, Figure 2C). The scapula is triradiate; the dorsal process is missing its dorsal end, but it has a broad ventral base and shows an undulated and posterodorsally-inflected anterior margin (Figure 1I), which distinguishes Franconiasaurus from Plesiopterys where the anterior margin is concave (O’Keefe, 2004). The dorsal process is posterodorsally-inclined relative to the ventral plate by 55°. The medial side of the dorsal process is gently convex and lacks a buttress-like enforcement that is present in some Early Jurassic plesiosauroids such as Microcleidus tournemirensis, and Microcleidus homalospondylus, but absent in Middle Jurassic cryptoclidids, such as Cryptoclidus, Tricleidus or Muraenosaurus (Andrews, 1910; Bardet et al., 1999; Benson and Druckenmiller, 2014, Appendix 2, character 206).
The ventral side of the dorsal process is anteroposteriorly expanded relative to the mid-shaft (Figure 1I). Among Early Jurassic plesiosaurs this condition is only shared with microcleidids and the rhomaleosaurid Avalonnectes (Benson and Druckenmiller, 2014, Appendix 2, character 203).
The right coracoids are almost complete (Figure 1J) and exposed in dorsal view in both specimens, whereas the according left coracoids are fragmentarily preserved. The anterior process is partly damaged in BT 011224.00 but complete in BT 011241.00 (Figure 2C). The process is rectangular and slightly longer than it is broad at its base, a condition that differs in Plesiopterys where the anterior process is much shorter and more anterolaterally inclined (see Electronic Supplementary Material S4, Supplementary Figure S3).
The well-defined semicircular posterolateral cornu, present in the holotype, does not exceed to the level of the glenoid which resembles the condition in some rhomaleosaurids (e.g., Meyerasaurus), pliosaurids (Hauffiosaurus zanoni), and Early Jurassic plesiosauroids (Plesiosaurus, Eoplesiosaurus, Westphaliasaurus) (Smith and Vincent, 2010; Benson et al., 2011; Schwermann and Sander, 2011; Benson and Druckenmiller, 2014). In microcleidids, however, the cornu extends to or beyond the level of the glenoid (Dames, 1895; Fraas, 1910; Großmann, 2006) and in Plesiopterys a cornu is absent (see Electronic Supplementary Material S4, Supplementary Figure S3).
The medial intercoracoid suture is straight and extends over nearly the entire length of the coracoid (Figure 1J). Only at the posteromedialmost part, the elements were slightly split which is well-visible in BT 011224.00. The posterior margin of the coracoid is slightly posterolaterally oriented.
From the pelvic girdle, the associated left ilium and ischium are preserved in BT 011224.00 and part of the ischium seems to be present in BT 011241.00. The ilium has a straight shaft and the dorsal and ventral ends of the element are perpendicular to one another (Figure 1M). This is similar to the condition observable in some pliosaurids (e.g., Peloneustes) or the plesiosauroid Plesiosaurus (Andrews, 1913; Benson and Druckenmiller, 2014, Appendix 2, characters 221 and 222). The shaft of the ilium appears to be transversely compressed and the dorsal side of the ilium is prominently expanded relative to the mid-shaft width.
The ischium has a broadly expanded medial side and a straight midline suture. The element has a length/width ratio of approximately 1.2. Though, the anteromedial end of the ischium is obscured. This condition is shared with most Early Jurassic plesiosaurs (Benson and Druckenmiller, 2014, Appendix 2, character 231), except for the basal rhomaleosaurid Macroplata where the ischia are relatively shorter (Ketchum and Smith, 2010).
All propodials are preserved in BT 011224.00 (Figures 1K, L) and both humeri and one femur are also present in BT 011241.00 (Figures 2D, E). The propodials are subequal in length; the humeri of BT 011224.00 are slightly shorter than the femora. All propodials in both individuals have a length to width ratio of 1.7 (in BT 011224.00: proximodistal length to distal width in the right humerus 22/12.48 cm and right femur 24/13.80 cm).
Humeri and femora have a dorsally and ventrally uniformly convex distal portion. The distal ends are convex.
The humeri have almost straight preaxial margins that lack a sharp longitudinal ridge (Figure 1K), resembling the condition in the basal plesiosauroid Plesiopharos and in xenopsarians, such as Brancasaurus and Eopolycotylus (Albright et al., 2007; Sachs et al., 2016; Puértolas-Pascual et al., 2021).
The proximal ends of the humeri are not inclined. They have a narrow dorsally-projecting tuberosity that has an undulated surface which bears several pits. The humeral capitulum is not exposed. The capitulum in the femur is likewise obscured and the morphology of the trochanter matches that of the tuberosity, being narrow and projecting dorsally.
Epipodials are preserved in both specimens (Figure 1L; Figures 2B, E, Electronic Supplementary Material S4, Supplementary Figure S3F, F). The radius and tibia are sub-quadratic and both show a concave preaxial margin. The ulna is crescent-shaped and the fibula has a circular morphology. The spatium interossum is large in the forelimb epipodials and narrow in the hindlimb elements. Some isolated metapodial elements and phalanges are preserved in BT 011224.00. The phalanges are elongate and slender.
Results of phylogenetic analyses
Numerical results (including lengths and numbers of most parsimonious trees) and full tree topologies (with node support values) are provided in Electronic Supplementary Material S3. All four phylogenetic analyses reconstruct broadly similar topologies for the early plesiosauroid segment of the plesiosaur phylogenetic tree and infer Franconiasaurus as the sister taxon to Cryptoclidia and as a sole representative of Plesiosauroidea to bridge the gap between early plesiosauroids and cryptoclidians (Figure 3). Following these results, we have performed two additional analyses. These analyses were based on equal weights and used the same settings but the first forced Franconiasaurus to fall within Cryptoclididae and the second constrained it as a microcleidid. The most parsimonious trees resulting from the former analysis were eleven steps longer than those reconstructed through the unconstrained parsimony with equal weights (2096 vs. 2085 steps), while the latter produced trees that were twelve steps longer (2097 vs. 2085 steps; Electronic Supplementary Material S3, Supplementary Table S1). As such, we consider it unlikely that Franconiasaurus is either a cryptoclidid or a microcleidid, and treat its placement to be “between” microcleidids and cryptoclidians as very well supported.
Only Plesiopterys wildi has previously been inferred at that position (e.g., Benson et al., 2012; Benson and Druckenmiller, 2014; Fischer et al., 2018; Madzia and Cau, 2020; Roberts et al., 2020). However, these results are doubtful. The taxon was established based on an early juvenile specimen that is additionally partly reconstructed. As such, some of its characters are obscured. We performed a preliminary reassessment of the Plesiopterys material which resulted in rescoring of 22 character states (Electronic Supplementary Material S4). When re-introduced in our matrix, the new Plesiopterys OTU was placed within Microcleididae, rather than as the sister taxon to Cryptoclidia, by all four analyses. Pending restudy of P. wildi, and possibly discovery of an adult individual of that taxon, such results need to be treated with caution.
Discussion
Character distribution in Franconiasaurus
Franconiasaurus displays a mixture of features that combine characters almost uniformly distributed among early plesiosaurs with those typically observed in later-diverging members of the clade. For example, as in earliest plesiosauroids and microcleidids, Franconiasaurus displays two co-joined rib facets in the anterior and middle cervical vertebrae or cervical zygapophyses with planar facets that are not connected medially, coracoid with a posterolateral cornu that does not exceed the level of the glenoid and a posterior margin that is posterolaterally oriented, ilium with a straight shaft that is mediolaterally compressed in the midsection, and radius and tibia with concave preaxial margin (e.g., Dames, 1895; Storrs, 1997; Großmann, 2006; Schwermann and Sander, 2011; Benson et al., 2012; Benson and Druckenmiller, 2014; Vincent et al., 2019). However, as is common in cryptoclidians (such as cryptoclidids) but not early-diverging plesiosauroids, Franconiasaurus shows reduced cervical rib processes with posteroventrally-oriented long axes and subrectangular caudal centra with a flat ventral side and widely-spaced hemapophyseal facets, scapula that lacks a medial enforcement, ilium that lacks a pronounced dorsal fossa, humerus with uninclined proximal end, and radius that lacks a pronounced anteroproximal flange (Andrews, 1910; O’Keefe and Wahl, 2003; O’Keefe and Street, 2009; Knutsen et al., 2012a; Benson and Bowdler, 2014; Benson and Druckenmiller, 2014; Roberts et al., 2020).
Marine reptile turnover at the Early/Middle Jurassic transition
The emergence of cryptoclidians, a diverse and globally-dispersed clade of later-diverging plesiosauroids, was associated with a major restructuring of marine reptile faunas near the Early/Middle Jurassic transition, probably between 175 and 171 Mya (Aalenian) (Fischer et al., 2021). This turnover apparently markedly affected several clades of marine tetrapods, including “fish-like” ichthyosaurs, marine crocodile relatives (thalattosuchians), as well as all three major lineages of plesiosaurs. The predatory plesiosaur clade Rhomaleosauridae that dominated in Early Jurassic seas started to disappear, meeting its final demise in the Callovian (late Middle Jurassic; ∼161.5 Ma) (e.g., Benson et al., 2015b), while Pliosauridae and Plesiosauroidea diversified. Pliosaurids established a successful clade of macropredators (Thalassophonea) that had regulated the upper tier of marine ecosystems since at least the early Bajocian (∼171 Ma) (Sachs et al., 2023) until the Turonian (early Late Cretaceous; ∼90 Ma; Madzia, 2016; Madzia et al., 2019) and comprised some of the largest aquatic predatory tetrapods that have ever lived, with forms exceeding lengths of 10 m (Knutsen et al., 2012b; Benson et al., 2013; Martill et al., 2023). Plesiosauroids, in turn, switched from microcleidid-rich communities common for Toarcian European epeiric seas to cryptoclidid faunas that represented the globally dominant plesiosauroid components in the Middle and Late Jurassic until they were replaced by leptocleidians and elasmosaurids near the Jurassic-Cretaceous transition (Benson and Druckenmiller, 2014).
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
SS: Conceptualization, Data curation, Formal Analysis, Investigation, Project administration, Resources, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing. SE: Data curation, Funding acquisition, Visualization, Writing–original draft, Writing–review and editing. DM: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
We are indebted to Joschua Knüppe (Ibbenbüren, Germany) for producing the artwork, Martín D. Ezcurra (CONICET–Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina) for handling our and José O'Gorman (Universidad Nacional de La Plata, Argentina) and Valentin Fischer (University of Liège, Belgium) for their constructive reviews that helped us to improve our manuscript. TNT is made available with the sponsorship of the Willi Hennig Society.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2024.1341470/full#supplementary-material
References
Albright, L. B., Gillette, D. D., and Titus, A. L. (2007). Plesiosaurs from the Upper Cretaceous (Cenomanian-Turonian) Tropic Shale of southern Utah, part 2: Polycotylidae. J. Vertebr. Paleontol. 27, 41–58. doi:10.1671/0272-4634(2007)27[41:pftucc]2.0.co;2
Andrews, C. W. (1910). A catalogue of the marine reptiles of the Oxford Clay, Part I. London: British Museum Natural History.
Andrews, C. W. (1913). A catalogue of the marine reptiles of the Oxford Clay, Part II. London: British Museum Natural History.
Bardet, N., Godefroit, P., and Sciau, J. (1999). A new elasmosaurid plesiosaur from the Lower Jurassic of Southern France. Palaeontology 42, 927–952. doi:10.1111/1475-4983.00103
Benson, R. B. J., and Bowdler, T. (2014). Anatomy of Colymbosaurus megadeirus (Reptilia, Plesiosauria) from the Kimmeridge Clay Formation of the U.K., and high diversity among Late Jurassic plesiosauroids. J. Vertebr. Paleontol. 34, 1053–1071. doi:10.1080/02724634.2014.850087
Benson, R. B. J., and Druckenmiller, P. S. (2014). Faunal turnover of marine tetrapods during the Jurassic-Cretaceous transition. Biol. Rev. 89, 1–23. doi:10.1111/brv.12038
Benson, R. B. J., Evans, M., Druckenmiller, P. S., and Lalueza-Fox, C. (2012). High diversity, low disparity and small body size in plesiosaurs (Reptilia, Sauropterygia) from the Triassic–Jurassic boundary. PLoS One 7, e31838. doi:10.1371/journal.pone.0031838
Benson, R. B. J., Evans, M., Smith, A. S., Sassoon, J., Moore-Faye, S., Ketchum, H. F., et al. (2013). A giant pliosaurid skull from the Late Jurassic of England. PLoS ONE 8, e65989. doi:10.1371/journal.pone.0065989
Benson, R. B. J., Evans, M., and Taylor, M. A. (2015a). The anatomy of Stratesaurus (Reptilia, Plesiosauria) from the lowermost Jurassic of Somerset, United Kingdom. J. Vertebr. Paleontol. 35, 9337399–e933826. doi:10.1080/02724634.2014.933739
Benson, R. B. J., Ketchum, H. F., Noè, L. F., and Gómez-Pérez, M. (2011). New information on Hauffiosaurus (Reptilia, Plesiosauria) based on a new species from the Alum Shale Member (Lower Toarcian: Lower Jurassic) of Yorkshire, UK. Palaeontology 54, 547–571. doi:10.1111/j.1475-4983.2011.01044.x
Benson, R. B. J., Zverkov, N. G., and Arkhangelsky, M. S. (2015b). Youngest occurrences of rhomaleosaurid plesiosaurs indicate survival of an archaic marine reptile clade at high palaeolatitudes. Acta Palaeontol. Pol. 60, 769–780. doi:10.4202/app.00167.2015
Brown, D. S. (1981). The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria. Bull. Br. Mus. Nat. Hist. Geol. 35, 253–347.
Brown, D. S., Vincent, P., and Bardet, N. (2013). Osteological redescription of the skull of Microcleidus homalospondylus (Sauropterygia, Plesiosauria) from the Lower Jurassic of england. J. Paleontol. 87, 537–549. doi:10.1666/11-104
Dames, W. B. (1895). Die Plesiosaurier der süddeutschen Liasformation. Abh. Königlich Preuss. Akad. Wiss. Berl., 1–81.
de Blainville, H. D. (1835). Description de quelques espéces de reptiles de la Californie, précédée de l’analyse d’un systéme général d’Erpétologie et d’Amphibiologie. Nouv. Ann. Muséum Natl. d’Histoire Nat. Paris 4, 233–296.
Fischer, V., Benson, R. B. J., Druckenmiller, P. S., Ketchum, H. F., and Bardet, N. (2018). The evolutionary history of polycotylid plesiosaurians. R. Soc. Open Sci. 5, 172177. doi:10.1098/rsos.172177
Fischer, V., Weis, R., and Thuy, B. (2021). Refining the marine reptile turnover at the Early–Middle Jurassic transition. PeerJ 9, e10647. doi:10.7717/peerj.10647
Goloboff, P. A. (1993). Estimating character weights during tree search. Cladistics 9, 83–91. doi:10.1111/j.1096-0031.1993.tb00209.x
Goloboff, P. A. (1995). Parsimony and weighting: a reply to Turner and Zandee. Cladistics 11, 91–104. doi:10.1111/j.1096-0031.1995.tb00006.x
Goloboff, P. A., Carpenter, J. M., Arias, J. S., and Esquivel, D. F. M. (2008). Weighting against homoplasy improves phylogenetic analysis of morphological data sets. Cladistics 24, 758–773. doi:10.1111/j.1096-0031.2008.00209.x
Goloboff, P. A., and Morales, M. E. (2023). TNT, version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics 39, 144–153. doi:10.1111/cla.12524
Goloboff, P. A., Torres, A., and Arias, J. S. (2018). Weighted parsimony outperforms other methods of phylogenetic inference under models appropriate for morphology. Cladistics 34, 407–437. doi:10.1111/cla.12205
Großmann, F. (2006). “Taxonomy, phylogeny and palaeoecology of the plesiosauroids (Sauropterygia, Reptilia) from the Posidonia Shale (Toarcian, Lower Jurassic) of Holzmaden, south west Germany,” (Tübingen: Eberhard-Karls-Universität Tübingen). Ph.D. thesis.
Großmann, F. (2007). The taxonomic and phylogenetic position of the Plesiosauroidea from the Lower Jurassic Posidonia Shale of south-west Germany. Palaeontology 50, 545–564. doi:10.1111/j.1475-4983.2007.00654.x
Ketchum, H. F., and Smith, A. S. (2010). The anatomy and taxonomy of Macroplata tenuiceps (Sauropterygia, Plesiosauria) from the Hettangian (Lower Jurassic) of Warwickshire, United Kingdom. J. Vertebr. Paleontol. 30, 1069–1081. doi:10.1080/02724634.2010.483604
Knutsen, E. M., Druckenmiller, P. S., and Hurum, J. H. (2012a). A new plesiosauroid (Reptilia: Sauropterygia) from the Agardhfjellet Formation (Middle Volgian) of central Spitsbergen, Norway. Norw. J. Geol. 92, 213–234.
Knutsen, E. M., Druckenmiller, P. S., and Hurum, J. H. (2012b). A new species of Pliosaurus (Sauropterygia: Plesiosauria) from the Middle Volgian of central Spitsbergen, Norway. Norw. J. Geol. 92, 235–258.
Madzia, D. (2016). A reappraisal of Polyptychodon (Plesiosauria) from the Cretaceous of England. PeerJ 4, e1998. doi:10.7717/peerj.1998
Madzia, D., and Cau, A. (2020). Estimating the evolutionary rates in mosasauroids and plesiosaurs: discussion of niche occupation in Late Cretaceous seas. PeerJ 8, e8941. doi:10.7717/peerj.8941
Madzia, D., Sachs, S., and Lindgren, J. (2019). Morphological and phylogenetic aspects of the dentition of Megacephalosaurus eulerti, a pliosaurid from the Turonian of Kansas, USA, with remarks on the cranial anatomy of the taxon. Geol. Mag. 156, 1201–1216. doi:10.1017/s0016756818000523
Martill, D. M., Jacobs, M. L., and Smith, R. E. (2023). A truly gigantic pliosaur (Reptilia, Sauropterygia) from the Kimmeridge Clay Formation (Upper Jurassic, Kimmeridgian) of England. Proc. Geol. Assoc. 134, 361–373. doi:10.1016/j.pgeola.2023.04.005
O’Keefe, F. R., Otero, R. A., Soto-Acuña, S., O’Gorman, J. P., Godfrey, S. J., and Chatterjee, S. (2017). Cranial anatomy of Morturneria seymourensis from Antarctica, and the evolution of filter feeding in plesiosaurs of the Austral Late Cretaceous. J. Vertebr. Paleontol. 37, e1347570. doi:10.1080/02724634.2017.1347570
O’Keefe, F. R., and Street, H. P. (2009). Osteology of the cryptocleidoid plesiosaur Tatenectes laramiensis, with comments on the taxonomic status of the Cimoliasauridae. J. Vertebr. Paleontol. 29, 48–57. doi:10.1671/039.029.0118
O’Keefe, F. R. (2004). Preliminary description and phylogenetic position of a new Plesiosaur (Reptilia: Sauropterygia) from the Toarcian of Holzmaden, Germany. J. Paleontol. 78, 973–988.
O’Keefe, F. R., and Wahl, W. (2003). Preliminary report on the osteology and relationships of a new aberrant cryptocleidoid plesiosaur from the Sundance Formation, Wyoming. Paludicola 4, 48–68.
Puértolas-Pascual, E., Marx, M., Mateus, O., Saleiro, A., Fernandes, A. E., Marinheiro, J., et al. (2021). A new plesiosaur from the Lower Jurassic of Portugal and the early radiation of Plesiosauroidea. Acta Palaeontol. Pol. 66, 369–388. doi:10.4202/app.00815.2020
Roberts, A. J., Druckenmiller, P. S., Cordonnier, B., Delsett, L. L., and Hurum, J. H. (2020). A new plesiosaurian from the Jurassic–Cretaceous transitional interval of the Slottsmøya Member (Volgian), with insights into the cranial anatomy of cryptoclidids using computed tomography. PeerJ 8, e8652. doi:10.7717/peerj.8652
Roberts, A. J., Druckenmiller, P. S., Delsett, L. L., and Hurum, J. H. (2017). Osteology and relationships of Colymbosaurus seeley, 1874, based on new material of C. svalbardensis from the Slottsmøya Member, Agardhfjellet Formation of central spitsbergen. J. Vertebr. Paleontol. 37, e1278381. doi:10.1080/02724634.2017.1278381
Sachs, S., Abel, P., and Madzia, D. (2022). A “long-forgotten” plesiosaur provides evidence of large-bodied rhomaleosaurids in the Middle Jurassic of Germany. J. Vertebr. Paleontol. 42, e2205456. doi:10.1080/02724634.2023.2205456
Sachs, S., Hornung, J. J., and Kear, B. P. (2016). Reappraisal of Europe’s most complete Early Cretaceous plesiosaurian Brancasaurus brancai Wegner, 1914 from the “Wealden Facies” of Germany. PeerJ 4, e2813. doi:10.7717/peerj.2813
Sachs, S., and Kear, B. P. (2017). A rare new Pliensbachian plesiosaurian from the Amaltheenton Formation of Bielefeld in northwestern Germany. Alcheringa 42, 487–500. doi:10.1080/03115518.2017.1367419
Sachs, S., Lindgren, J., Madzia, D., and Kear, B. P. (2021). Cranial osteology of the mid-Cretaceous elasmosaurid Thalassomedon haningtoni from the Western Interior Seaway of North America. Cret. Res. 123, 104769. doi:10.1016/j.cretres.2021.104769
Sachs, S., Madzia, D., Thuy, B., and Kear, B. P. (2023). The rise of macropredatory pliosaurids near the Early-Middle Jurassic transition. Sci. Rep. 13, 17558. doi:10.1038/s41598-023-43015-y
Schwermann, L., and Sander, P. M. (2011). Osteologie und Phylogenie von Westphaliasaurus simonsensii: ein neuer Plesiosauride (Sauropterygia) aus dem Unteren Jura (Pliensbachium) von Sommersell (Kreis Höxter), Nordrhein-Westfalen, Deutschland. Geol. und Paläontologie Westfalen 79, 1–56.
Smith, A. S. (2007). “Anatomy and systematics of the Rhomaleosauridae (sauropterygia: Plesiosauria),” (Dublin: University of Dublin). Ph.D. thesis.89
Smith, A. S., and Benson, R. B. J. (2014). Osteology of Rhomaleosaurus thorntoni (Sauropterygia: Rhomaleosauridae) from the Lower Jurassic (Toarcian) of Northamptonshire, England. Monogr. Palaeontogr. Soc. Lond. 168, 1–40. doi:10.1080/02693445.2014.11963953
Smith, A. S., and Vincent, P. (2010). A new genus of pliosaur (Reptilia: Sauropterygia) from the Lower Jurassic of Holzmaden, Germany. Palaeontology 53, 1049–1063. doi:10.1111/j.1475-4983.2010.00975.x
Storrs, G. W. (1997). “Morphological and taxonomic clarification of the genus Plesiosaurus,” in Ancient marine reptiles. Editors J. M. Callaway, and E. L. Nicholls (San Diego: Academic Press), 145–190.
Vincent, P., Bardet, N., and Mattioli, E. (2013). A new pliosaurid from the Pliensbachian (Early Jurassic) of Normandy (northern France). Acta Palaeontol. Pol. 58, 471–485. doi:10.4202/app.2011.0113
Vincent, P., and Benson, R. B. J. (2012). Anningasaura, a basal plesiosaurian (Reptilia, Plesiosauria) from the Lower Jurassic of Lyme Regis, United Kingdom. J. Vertebr. Paleontol. 32, 1049–1063. doi:10.1080/02724634.2012.686467
Keywords: faunal turnover, phylogenetic diversity, Plesiosauria, Plesiosauroidea, Early/Middle Jurassic transition
Citation: Sachs S, Eggmaier S and Madzia D (2024) Exquisite skeletons of a new transitional plesiosaur fill gap in the evolutionary history of plesiosauroids. Front. Earth Sci. 12:1341470. doi: 10.3389/feart.2024.1341470
Received: 20 November 2023; Accepted: 29 January 2024;
Published: 16 February 2024.
Edited by:
Martin Daniel Ezcurra, Museo Argentino de Ciencias Naturales Bernardino Rivadavia, ArgentinaReviewed by:
José O'Gorman, Universidad Nacional de La Plata, ArgentinaValentin Fischer, University of Liège, Belgium
Copyright © 2024 Sachs, Eggmaier and Madzia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel Madzia, ZGFuaWVsLm1hZHppYUBnbWFpbC5jb20=