
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci., 01 December 2023
Sec. Quaternary Science, Geomorphology and Paleoenvironment
Volume 11 - 2023 | https://doi.org/10.3389/feart.2023.1302419
This article is part of the Research TopicBehaviors and Adaptations of Prehistoric Hunter-Gatherers in the (Sub)Tropical Rainforest Area—Archaeology, Chronology and PaleoenvironmentView all 5 articles
 Devara Anil1*
Devara Anil1* Monika Devi2,3
Monika Devi2,3 Neha Ati1Mukesh C. P1Zakir Khan4
Neha Ati1Mukesh C. P1Zakir Khan4 Vrushab Mahesh1
Vrushab Mahesh1 P. Ajithprasad1Naveen Chauhan2
P. Ajithprasad1Naveen Chauhan2 Akash Pandey1
Akash Pandey1 Gopesh Jha5,6*
Gopesh Jha5,6*The chronology and hominin association of the South Asian Middle Palaeolithic have attracted much attention in the last few decades. The emergence of Middle Palaeolithic culture in the region has been debated between the local origins (behavioural change) model based on an early date around 380 ka and the diffusion (biological change) model based on Homo sapiens dispersals from Africa around 120–80 ka. The latter has more consensus, whereas the former requires a more robust chronological framework to attribute the emergence of the Middle Palaeolithic to behavioural changes. In the absence of hominin remains, the presence of Middle Palaeolithic technological trajectories are frequently used as behavioural markers of Homo sapiens. Homo sapiens fossil remains from the regions between Africa and South Asia dated to ∼ 200 ka presents more convincing support for the latter model. Here we present contextual, chronological and technological analysis of Middle Palaeolithic assemblages dated to 52 ka from Motravulapadu, Andhra Pradesh, India. Morphometrical analysis of the lithic assemblage indicates diverse Levallois core reductions were practised at the site at the onset of MIS 3. Further this evidence highlights the significance of MIS 3 cultural diversity in South Asia, likely related to changing population dynamics, cultural drift, and the highly variable climatic context of MIS 3.
Explaining the Homo sapiens dispersals from their African origin to Eurasia has become more complex than earlier, despite new discoveries (Liu et al., 2015; Harvati et al., 2019; Bergström et al., 2021; Slimak et al., 2022; Freidline et al., 2023). This is because the current archaeological, genetic and fossil records across Eurasia and Australia do not match with any existing model that the explains Homo sapiens dispersals (Reyes-Centeno, 2016; Bae et al., 2017; Sahle et al., 2018; Harvati and Reyes-Centeno, 2022; Vahdati et al., 2022). In addition, the absence of fossil records and lack of robust chronological characterization of the Late Pleistocene archaeological record from the regions in the Homo sapiens dispersal route further complicates the models. One of such regions, South Asia, strategically situated between Africa and East Asia could have been a major corridor for the Homo sapiens dispersals from West to East (Blinkhorn and Petraglia, 2017). Besides, the Palaeolithic record of South Asia has a long history of research and shows rich and diverse cultural evidence. The region’s role in the Homo sapiens dispersals route was highlighted in the recent times where it attracted much attention among the paleoanthropological communities (James and Petraglia, 2005; Field et al., 2007; Petraglia et al., 2007; Petraglia et al., 2009; Boivin et al., 2013; Bae et al., 2017). Following Petraglia et al.’s (2007) work at Jwalapuram, Andhra Pradesh that proposes Homo sapiens migrated to South Asia with a Middle Palaeolithic tool kit, between 120 and 80 ka—prior to the Toba super-eruption of 75 ka and survived the catastrophic event, many other discoveries across South Asia suggested similar narratives (Blinkhorn et al., 2019; Clarkson et al., 2020). This model, popularly known as the pre-Toba/MIS-5 model, is supported by further evidences, such as 1) South Asian Middle Palaeolithic culture is datable to the beginning of the Late Pleistocene (Blinkhorn et al., 2013; Blinkhorn et al., 2019; Clarkson et al., 2020); 2) the revised ages for the presence of Homo sapiens in Eurasia to ∼ 200 ka (Harvati et al., 2019) and in Australia to ∼ 65 ka (Clarkson et al., 2017). However, recently the antiquity of the South Asian Middle Palaeolithic was pushed back to the Middle Pleistocene (Akhilesh, et al., 2018; Anil et al., 2022). Further the early presence of the Homo sapiens in West Asia and Caucasus was seen as failed dispersal event (Harvati et al., 2019).
Here we present the securely dated Levallois dominated Middle Palaeolithic assemblages from Motravulapadu, Prakasam District, Andhra Pradesh, India (Figure 1). Motravulapadu (15° 8′10.82″North latitudes, 79°25′16.08″East longitudes) is situated on the banks of Nachua vagu, a tributary of the Manneru River (Figure 2). The stream flows for 15 km southwards before joining the Manneru River which drains into the Mopadu reservoir. The site was initially reported by Mishra and Singaraju (2009) and later explored by (Srinivasulu, 2012). The current team conducted a thorough investigation of the site, recording stratigraphy and Palaeolithic material at various locations. Motravulapadu is a large, 4 km x 2 km site where rill and sheet erosional events have exposed artefacts, fossilised faunal remains and several vertical sections of Quaternary formations. Furthermore, current mining activities have exposed parts of the bedrock in a few spots. The site has a long record of hominin occupation ranging from Middle Palaeolithic to the Microlithic. Large deposits of volcanic tephra (YTT) were observed over a vast area, belonging to ∼75 ka Toba eruption. Co-occurrence of YTT, mammalian fauna, and Middle Palaeolithic assemblages provides a unique opportunity to understand hominin ecology and past environment during Late Pleistocene.
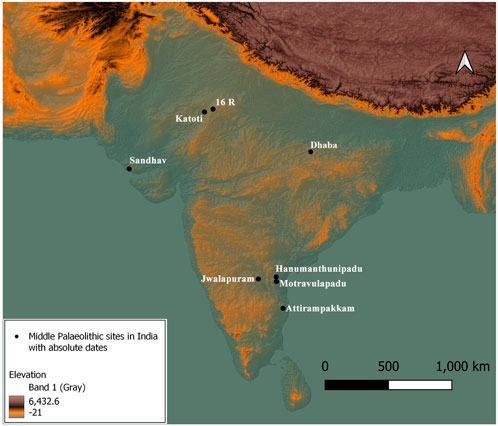
FIGURE 1. Map showing the location of the site Motravualapdu and other dated Middle Palaeolithic sites in India.

FIGURE 2. Locations of the trenches excavated at Motravulapadu.
Excavations were conducted at this site for one field-season in 2021. Trenches were opened at six localities within an area of 0.5 km x 0.6 km. These localities were chosen with the intention of understanding the relationship between the sediments exposed at these localities and building a complete lithostratigraphic history of the site. However, the current paper presents the stratigraphy, luminescence chronology and lithic assemblage recovered from Trench 1.
Excavations were carried out employing a single context recording system, with discrete sediment units separated into 10 cm spits as required. Sediments were screened using a 2 mm sieve to recover micro artefacts. Excavated sections and artefact-bearing horizons were photographed for photogrammetric reconstruction. OSL samples were collected from all sediment units, with the recovery of sediment samples supported by in situ background radiation measurements.
Sediment samples were analysed for luminescence ages in Physical Research Laboratory, Ahmedabad, India using the procedures discussed in (Anil et al., 2022). We measured 12 aliquots per sample using multigrain p-IR-IRSL on single grain disc. Preheat test was conducted on the natural sample, and an arithmetic mean of De values of 3 aliquots for each preheat was used. The doses for 280°C, 300°C, 320°C, and 340°C fell within 5% of the estimated paleo-dose and 320°C was used as pre-heat temperature (Supplementary Figure S1). The overdispersion (OD) in estimated De values was relatively low (∼8%), indicating well bleaching of the sediments before burial. Thus, for De estimation central age model (CAM) was used. Typical feldspar shine-down growth curves are shown in Supplementary Figures S2–S4 for Unit 3 and Unit 4 samples, respectively. A residual dose of 5.1 ± 0.6 Gy, which is the value of the dose remaining in the sample after 5 hours extended solar lamp bleaching, was subtracted. The dose reproducibility of the used p-IR-IRSL-SAR protocol was tested using the dose recovery test (ratio of recovered to given dose <10% of unity). For the dose recovery test, a known dose of 94 ± 1 Gy was given in the reader after the solar lamp beaching for 5 hours. The given dose was immediately recovered as an unknown dose. The recovered doses were further corrected for residual doses. The observed dose recovery ratio was 0.90 ± 0.03, indicating that the used p-IR-IRSL-SAR protocol has good dose reproducibility. The fading rate measurements were done using the prescription of Auclair et al. (2003) and (Huntley and Lamothe, 2001). The estimated g—values for MVP_T1_OSL3, MVP_T1_OSL4, and MVP_T1_OSL5 are −0.6 ± 0.5, −0.6 ± 0.5, and 0.6 ± 0.5, respectively. These values are either less than 1% or negative with ∼100% errors (Supplementary Figure S5), suggesting the p-IR-IRSL signal is not affected by the anomalous fading (Buylaert et al., 2011). Therefore, no fading corrections were applied to the samples to correct the ages. The concentration of Uranium (238U), Thorium (232Th) and Potassium (40K) nuclides were measured using High Purity Germanium (HPGe) detector. Further, these concentrations were used to estimate the total dose rate assuming infinite matrix assumption, and secular equilibrium for all the nuclides (Beck and de Planque, 1985). An internal potassium (40K) of 12.5% ± 0.5% (Huntley and Baril, 1997) and 400 ± 100 ppm Rubidium (87 Rb) (Huntley and Hancock, 2001) were considered for dose rate estimations.
Quantitative and qualitative attributes were recorded for each artefact in the assemblage, except for broken flakes, flaked pieces, and core fragments. Artefacts were categorized into standard technological groups and more formal typological classes wherever suitable (e.g., Core, Flake, Retouched flake, Blade). For typological and technological descriptions of the artefacts, standard terminologies used across South Asia and beyond (e.g. (Akhilesh et al., 2018; Blinkhorn, 2012; Jones, 2007; Zaidner, 2018), were employed. The same thing applies to the terms used in the formal descriptions of key technologies such as Levallois (Van Peer, 1992b; Boëda, 1995; Bordes, 1980). For each artefact, a number of metric and non-metric attributes were recorded (Supplementary Tables S1, S2). Representative artefacts were photographed for illustration purposes.
Trench 1 is the largest among the six trenches that measures 4 m x 3.5 m (Figure 3A). Excavations were conducted to a depth of 3.5 m where the digging ceased after encountering the water table. Five distinct stratigraphic units were identified in Trench 1 (Figure 3B). The bottom most Unit (Unit 5) in the trench is of a brownish red coloured silty sandy sediment with relatively high moisture content and only the top 40 cm of this Unit was excavated below which water table was encountered. Unit 5 is overlain by ∼ 80 cm thick, dark brownish red coloured sandy silt sediment (Unit 4). This Unit yielded Middle Palaeolithic artefacts. A light brown coloured clayey silt (Unit 3) of uneven thickness overlays Unit 4 throughout the trench. Lithic artefacts were also recovered from the topmost part of this Unit. Unit 3 is overlain by Unit 2, a pale yellow coloured sandy silty sediment distributed unevenly throughout the trench.
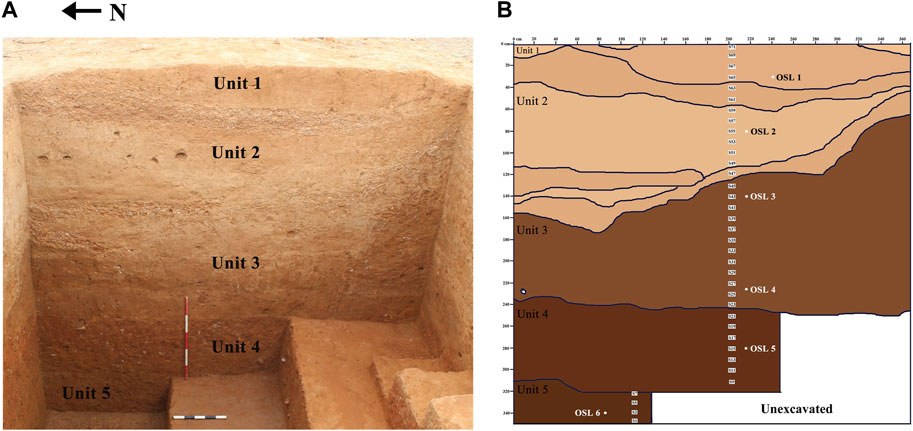
FIGURE 3. (A) Section facing west, trench 1; (B) Schematic sketch of the section.
Notably, at least three different episodes of high energy erosional events represented by coarse material consisting of small sized pebbles and carbonate nodules were observed within Unit 2 (Figure 4). Interestingly, these subunits within Unit 2 yielded microlithic artefacts. Unit 1 is a yellow-coloured, silt dominated Unit that represents topsoil. The stratigraphy of Trench 1 is dominated by the presence of fine-grain sediments primarily consisting of silt and with less percentage of sand and clay. Except the artefacts in Unit 4, 3 and 2, no other pebbles or any other coarse material was observed in the trench. Such nature of the sediments indicates low energy depositional conditions, which are archaeologically ideal for better preservation of site/occupational features.

FIGURE 4. Coarse sediments within Unit 2.
Six samples were collected for OSL dating. However, only three of them were processed for age estimation, as the artefacts were found only in these Units. The three dated samples were collected at the depths of 1.40 and 2.25 m from Unit 3, and 2.80 m from Unit 4. The dates for Unit 3 are 35 ± 2 ka and 24 ± 2 ka, and Unit 4 is dated to 52 ± 3 ka (Table 1). Luminescence age estimations were done using p-IR-IRSL protocol. All the three samples showed less over dispersion indicating well bleaching before the burial. No significant fading was observed in the fading estimations and therefore ages were not corrected.
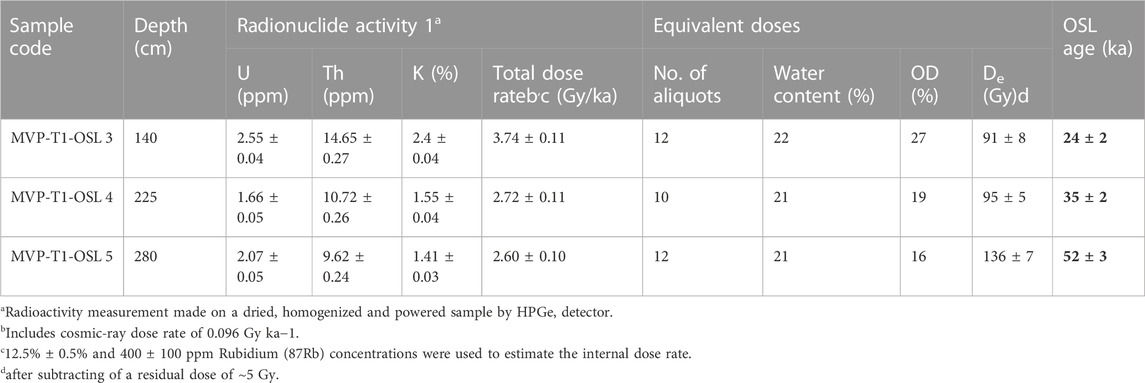
TABLE 1. Dose rate data, De values and OSL ages for the sediment samples from Trench 1, Motravulapadu.
Trench 1 has yielded three artefact bearing horizons: Unit 2, 3 and 4. The Unit 2 artefacts are not discussed here as no luminescence age of this sediment was assessed. Besides, the artefacts from Unit 2 are microlithic in nature consisting of microblade cores and blades, backed blades, geometric microliths such as lunates, triangles, and trapeziums. The typo-technology of the artefact assemblages of Unit 3 and Unit 4 is discussed below.
Unit 3 yielded 121 artefacts within the top 35 cm associated with fine grain sediments (Figure 5). All the artefacts were made of fine grain quartzites that are easily available in the alluvial gravel beds. Among the 121 artefacts recovered, 47 are of debitage, 17 flaked pieces, and 4 are of worked nodule (Table 2).

FIGURE 5. In situ artefacts excavated from the Unit 3 of Trench 1.
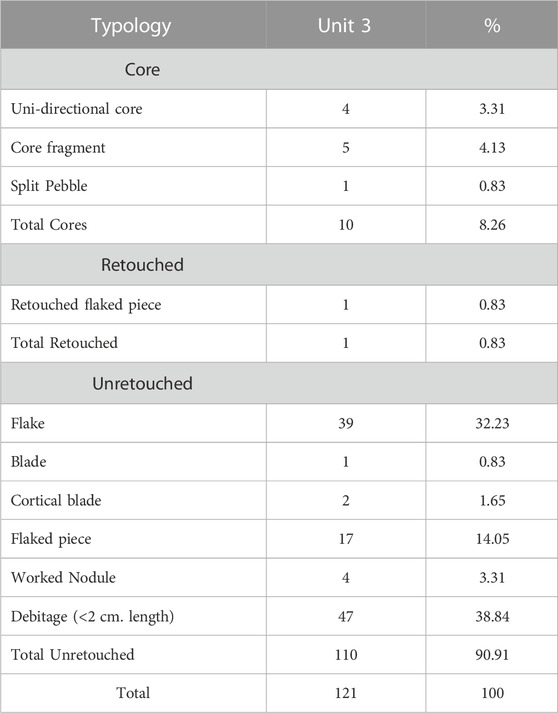
TABLE 2. Composition of the Lithic assemblage from Unit 3, Trench 1.
These three categories which are a crucial part of the lithic reduction sequence form 56.20% of the total assemblage. All the below-mentioned lithic analyses and technological descriptions are based on the rest of 53 artefacts.
Lithic debitage (<2 cm in length) constituted a large portion (38.8%, n = 47) of the assemblage. Unretouched flakes, both complete and broken, are the next common class (32.23%, n = 39) followed by flaked pieces (14.05%, n = 17). Core component in the assemblage consists of only four unidirectional cores (Figure 6), five core fragments and one split pebble (Supplementary Figure S6). Overall, 80% of the assemblage, mostly cores (19.23%) and flakes (61.54%), retain cortex. The cortex is of cobble cortex type indicating its riverine source.

FIGURE 6. Unidirectional cores from Unit 3 of Trench 1 at Motravulapadu.
Generally, discarded flakes with cortex would indicate the initial stage reduction of cores and the subsequent use of unidirectional core technology. Initial stages of reduction sequences in the assemblage are evident by the presence of cortex on flake platforms, the dorsal surface, and lateral margins, which account for half of the complete flakes (n = 15). Intact flakes with two or more dorsal flake scars (n = 28) are dominated by the flakes with radial and weakly radial scarring forming 71.42% (n = 20). Examples of unidirectional flake scarring (from proximal) makes up 28.57% (n = 8). Three-quarters of the flakes have cortical platform, and a few instances of faceting (14.28%) and overhang removal (10.71%) are also observed. Flakes from different stages of the reduction sequence such as cortical flakes having cortical platforms and entire dorsal surface with cortex, flakes having cortical platforms with a few preparation scars on the dorsal surface, flakes having prepared platforms with cortex on the dorsal surface, and platform rejuvenation flakes are present in the assemblage (Supplementary Figure S7). The presence of aforementioned flake types along with high percentage of debitage indicates the primary nature of lithic assemblage. The lithic assemblage recovered from Unit 3 of Trench 1 lacks characteristic diagnostic features that would allow it to be assigned to any known specific reduction sequence. Excavations at Trench 4 located 500 m north of Trench 1 yielded blade based lithic assemblages (details of which are not discussed in this paper) dated to 29 ka (Personal communication with Monica Devi). Similarities were observed between the typo-technological attributes of debitage from Unit 3 of Trench 1 and Trench 4 debitage. Based on these similarities and the luminescence age of 24 ka of Unit 3, which is close to the age (29 ka) of the Trench 4 artefact-bearing horizon, it can be safely concluded that the assemblage is part of a blade core reduction process.
Unit 4 has yielded 357 artefacts dominated by the products of Levallois reduction (Table 3). Most artefacts retain cortex (65.47%), indicating that the initial preparation of cores must have taken place on-site. Pebbles from the stream bed must have served as a raw material source. The cortex type is observed to be cobble type, which further corroborates the aforesaid observation. None of the artefacts show any evidence of abrasion and are in mint condition.
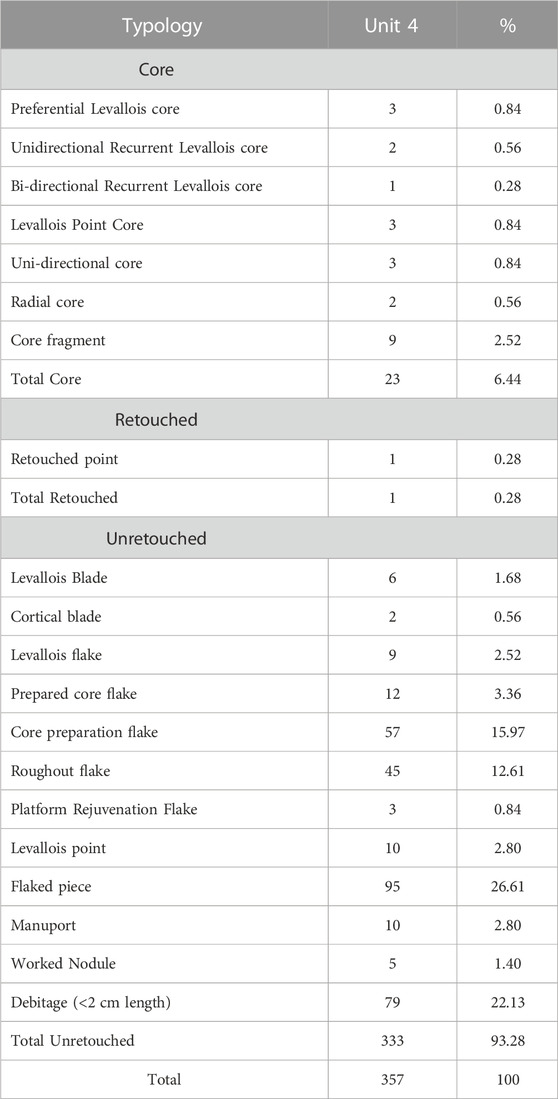
TABLE 3. Composition of the lithic assemblage from Unit 4 of Trench 1.
The number of cores in the assemblage accounts for 14; all belong to formal reduction strategies. The formal core reductions include Levallois, radial, and unidirectional. Notably, except for 9 core fragments, no other informal cores are present in the assemblage. Among Levallois core reduction strategies, recurrent and preferential (Figure 7) systems are present along with Levallois point cores (Supplementary Figures S8, S9). Three unidirectional and two radial cores are present in the assemblage.

FIGURE 7. Levallois Cores from Unit 4 of Trench 1 at Motravulaapdu. (A) Preferential Levallois core; (B) Recurrent Levallois core.
A maximum of two and a minimum of one core rotations are observed in Levallois cores, whereas unidirectional and radial cores show a maximum of three and a minimum of one rotation. Most cores show one (n = 5) and two (n = 7) major scars (>1/3 core length), with two cores exhibiting three and four scars each. Cores’ mean length, medial width and medial thickness are 56.35 mm x 66.11 mm x 32.35 mm when oriented along the flaking axis (last flake scar) (Supplementary Table S3). The core’s mean proximal, medial, and distal widths measured along the flaking axis are 64.68 mm x 66.11 mm x 46.85 mm, respectively. The proximal shape of the cores is relatively straight (mean 0.98 mm), whereas the distal shape is tapering (mean 1.55 mm). No considerable variation in core elongation (ranging between 0.63 and 1.22) is observed, maybe because there is no significant variation in the cores’ mean length and mean medial width. The core flatness index ranges from 0.88 to 3.69, where most cores are wider than thick. Five cores have cortical platforms, six have single conchoidal platforms, and 3 show multiple conchoidal platforms. Platforms were facetted on 9 cores; in five instances, no preparation was observed. The last scar-face-length of cores is less than the axial core length, indicating the flaking face is limited to the smaller axial surface. The last scar lengths range from 26.42 to 61.61 mm, with an average of 39.19 mm, and the last scar widths range between 23.57 and 40.75 mm, and a mean of 32.64 mm. On average, the last flake scar elongation (mean = 1.26) indicates that relatively squarish flakes were removed. Majority of the last major flake scars exhibit feather terminations (73.33%), with non-feather terminations accounting for 26.66%.
One hundred forty-five flakes, including both retouched and unretouched, have been recorded in the assemblage, including 92 complete flakes. Flakes were classified according to technological type to understand their position in the reduction sequence (Supplementary Table S4). The typo-technological description of the flakes below is based on the complete flakes in the assemblage. A wide range of technological diversity is evident amongst the flakes, from core preparation flakes to end products. Considerable number of roughout flakes (31.03%) along with core preparation flakes (39.31%) overall forming 70.34% of the flake component indicate that initial preparation of the cores was done on the site. Flakes from different stages of the core reduction sequence are noted in the assemblage, from complete cortical flakes to end products such as Levallois points and flakes (Supplementary Figures S10–S12). The presence of one platform rejuvenation flake also corroborates the observation mentioned above. End products such as Levallois flakes including Levallois points, prepared core flakes and Levallois blades account for 27.59% of the flake component. The most common dorsal scar patterns present on complete flakes in the assemblage are proximal (41.30%), followed by weakly radial (32.61%) and radial (14.13%). Flakes with no dorsal scars (cortical flakes) were present in considerable numbers forming 11.96%. Cortical coverage ranges from 0%–100%, with 45.65% of flakes recorded with no cortex and 28.26% with less than 50% cortex, and 9.78%, 4.35%, 11.96% with 60%, 80%, and 100% cortex, respectively. Considerable number of flakes with only cortical platforms (28.63%) and 100% cortical cover of the dorsal and platform surface (11.96%) are observed. Notably, all the complete flakes show feather terminations.
Mean axial flake dimensions are 48.20 x 37.77 x 11.72 (LxWxT) mm; on average, flakes are squarish/oval (mean elongation = 1.44) (Supplementary Table S5). Typically, flakes are more than four times as wide as they are thick (mean flatness = 3.52), with a flatness range of 1.44%–6.82.75% of the flakes exhibit slightly expanding proximal margins (mean proximal shape = 0.80), with 25% exhibiting contracting proximal margins, leading to an upper proximal shape index of 1.12. In contrast, the distal shape indicates that 84.78% of the flakes exhibit distal contracting margins (mean = 2.23). Single conchoidal platforms are the most common type (38.04%), followed by cortical (28.26%), multiple conchoidal (23.91%), dihedral (7.61%), and crushed (2.17%) types. While half of the flakes (51.09%) have no platform preparation, faceting and overhang removal were seen on 35.87% and 13.04% respectively. A wide range in platform size is evident, with platform width ranging from 7.57 to 64.32 mm and platform thickness ranging from 3.03 to 29.30 mm. The platform shape index indicates that platforms are typically elongated (mean elongation = 2.90), with 81.52% being two times wider than thick. A total of 95 artefacts categorized as flaked pieces are present, which bear no precise ventral morphologies or negative flake scars originating from the margins of the artefacts but have clearly undergone some reduction. Except for one retouched point, no other retouched artefacts are noted in the assemblage. However, considerable number of Levallois points (n = 9) are present in the assemblage (Figure 8).

FIGURE 8. Levallois points from Unit 4 of Trench 1 at Motravulapadu.
Trench 1 at Motravulapadu reveals three artefacts bearing horizons belonging to distinct lithic reduction sequences. Microlithic artefacts in Unit 2 include micro-blade cores, backed blades, and geometric microliths. No chronometric age for Unit 2 is generated. But based on the context and composition of the sediment and its general features of occurrence around the landscape this Unit can be tentatively assigned to the Holocene age. Unit 3 has lithic assemblage aimed at producing elongated flakes and blades dated to 24 ka. Even though, the assemblage from Unit 3 is undiagnostic, based on the presence of debitage that resembles the debitage of Trench 4 lithic assemblage dated to 29 ka (personal communication with Monica Devi), it is reasonable to assume that the former is a blade technology-based assemblage.
Unit 4 from Trench 1 reveals lithic assemblages consisting of Levallois and prepared core reduction strategies dated to 52 ka. The presence of debitage smaller than 2 cm in length and flakes from different stages of core reduction indicates the in-situ nature of the assemblage. Variations in Levallois core reductions including Preferential, Recurrent, Uni and Bi-directional recurrent Levallois and Levallois point cores are present in the assemblage. Pebbles and cobbles available from the alluvial gravel beds were exploited to make artefacts. Most of the Levallois cores have cortex on the platform surface side. The pebbles were split in to two-halves which resulted in each half having natural hierarchical surfaces, exploited further as Levallois and other prepared cores. Typical examples of Levallois points and Levallois flakes are present in the assemblage, indicating widespread use of Levallois technology by 52 ka at the site. A few samples of Levallois blade elements are also noted in the assemblage, though retouched artefacts are minimal.
This 52-ka old Middle Palaeolithic assemblage from Motravulapadu with rich and diverse Levallois technology adds significant new information on the evolution of Middle Palaeolithic culture in South Asia. Further, this assemblage presents one of the securely dated Middle Palaeolithic assemblages dated to the beginning of MIS 3 timeframe in South Asia. The archaeological records of South Asia during the MIS 3 exhibit evidence of complex behavioural patterns, some of which were probably due to cultural transformations. The co-existence of multiple technological trajectories not only shows the diversification and regionalization of stone tool technology but also highlights the complexity of cultural processes (like convergence, displacement, and diffusion). It is extremely crucial to define the nature of the relationship and interaction between these techno-cultural practices. It seems clear that at least two technological trajectories existed during this time frame in South Asia. One is the microlithic technology dated back to 48 ka (Mishra et al., 2013; Clarkson et al., 2020) and the other is the Middle Palaeolithic technology continuing up to 38 ka (Clarkson et al., 2012; Petraglia et al., 2012) showing the potential temporal overlap between the two. However, it has been argued that South Asian microlithic technology was indigenously developed by Homo sapiens populations from Middle Palaeolithic technology (Clarkson et al., 2009; Clarkson et al., 2012; Clarkson, 2018; Clarkson et al., 2020; Haslam et al., 2010; M; Petraglia et al., 2007). These conclusions were made based on the excavations conducted in the Jwalapuram 9 rock shelter which yielded microlithic assemblages dated to 34 ka and in Jwalapuram 20 where the Middle Palaeolithic assemblages were dated to 38 ka. Similar claims were also made from the site Dhaba in the middle Son valley, Madhya Pradesh where Middle Palaeolithic technology continued up to 48 ka till the emergence of microlithic technology at 48 ka (Clarkson et al., 2020). The aforesaid studies attribute this independent innovation of microlithic technology in South Asia to Homo sapiens who migrated to the region from Africa between 120–70 ka with Middle Palaeolithic technology. However, microlithic assemblages stratigraphically and chronologically overlying the Middle Palaeolithic assemblages do not necessarily signify the transition between the two. In addition, the aforementioned studies (Clarkson et al., 2012; Clarkson et al., 2020) do not explain the presence of blade-based assemblages that usually date back to around ∼ 40 ka (Murty, 1979; Mishra, 2013) in the region. This temporal overlap of three distinct lithic technological trajectories (Prepared core/Levallois, Blade assemblages and Microlithic) in South Asia highlights the significance of MIS 3 cultural sequence in understanding the human cultural evolution in the region. Does this temporal overlap indicate regional variation in the cultural behaviour of hominins (possibly Homo sapiens)? Probably not, because the variations appear to be deep rooted as the three lithic technologies are based on very distinct lithic reduction sequences. Alternatively, could it be due to transitions between Late Middle Palaeolithic and Late Palaeolithic wherein the blade-based assemblages are a part of the transitional phase? The difficulty here is that the microlithic assemblages are older than the blade assemblages, and the Late Middle Palaeolithic assemblages are younger than microlithic assemblages by at least 10 ka. Does this temporal overlap, therefore, indicate presence of multiple hominin species practising distinct lithic technologies? The lack of hominin fossils from the region is the biggest hurdle in solving this issue. Notably, this scenario is significant as recent research in the fields of genetics and paleoanthropology not only contributed much to our understanding of hominin evolution and dispersals but also made it more complex than previously thought. The discovery of a few new hominin species with mixed features has changed the landscapes of paleoanthropological research and highlighted the complexities of evolution, dispersals, and diversity of the genus Homo. The discovery of Homo floresiensis, a small-bodied, hominin species from Liang Bua (Flores, Indonesia), for instance, has generated wide interest and debates among the scientific communities (Brown et al., 2004; Morwood et al., 2004; Morwood et al., 2005). The skeletal remains of Homo floresiensis were found in sediments that are dated between 100 and 60 ka and the associated lithic artefacts were dated to 190–50 ka (Sutikna et al., 2016). Further, the presence of another new hominin species, Homo luzonensis, whose remains were discovered in Callao cave (Northern Luzon, the Philippines) dated to 67 ka highlights the importance of Island Southeast Asia in the evolution of genus Homo (Mijares et al., 2010; Détroit et al., 2019). Another previously unknown hominin species with mixed features, Homo naledi, was discovered in the Rising Star cave system, South Africa dated to around 285 ka (Berger et al., 2015; Dirks et al., 2017). The date of this primitive species suggests that at least three hominin species existed across the African continent at ∼ 300 ka with the other two being the Homo sapiens and Homo heidelbergensis (Galway-Witham et al., 2019). Denisovan fossils are known from the Denisova cave in Altai mountains (Siberia, Russia) and Baishiya Karst cave on the Tibetan plateau, Xiahe, China dated between 200–50 ka (Chen et al., 2019; Douka et al., 2019). Two other well-known species of genus Homo, the Neanderthals, and Homo erectus persisted until about 40–45 ka in Europe and Indonesia respectively (Stringer, 2002). Besides, the recent advancements in the field of genetics and aDNA reconstructions help us to access the previously unknown facets of human evolution and dispersals. While colonizing Eurasia around 60 ka, Homo sapiens interbred with multiple extinct hominin species (Skoglund and Mathieson, 2018). The traces of such interbreeding events were recorded in the genomes of the modern-day population. These interbreeding events first occurred between Homo sapiens populations and Neanderthals in western Eurasia around 50–55 ka and contributed 2% of introgressed Neanderthal DNA that is now found in non-African populations (Green et al., 2010). In Asia, at least three different hominin groups appear to have been involved in the interbreeding events of which the Denisovans are currently known (Teixeira and Cooper, 2019). The two unknown hominin species are from South Asia and East Asia.
The aforementioned fossil and genetic data suggest that during the Late Pleistocene epoch particularly around 60 ka multiple hominin species have existed in different parts of the Old World of which only our species survived. In addition, Homo sapiens interacted with the Neanderthals, Denisovans, and possibly with two other unknown hominin species in Asia. However, the nature of these interactions is still unclear concerning the behavioral similarities and differences between these multiple hominin species. How exactly other hominin species (e.g., Neanderthals and Denisovans) acted at a biological and behavioral level in relation to our species is an important question. Because, if the other distinct species of genus Homo exhibits behaviors similar to our species, should they be considered as modern as we are on a cultural and cognitive level? Similarly, is it the differences in behaviors that helped our species to survive while others become extinct? Therefore, it is crucial to reconstruct the behaviors of these multiple species to answer these questions. In this regard, the MIS 3 archaeological record of South Asia can throw significant light on the hominin introgression events and behavioral evolution.
We present securely dated Middle Palaeolithic and blade-based assemblages from Motravulapadu, Andhra Pradesh, India, associated with fine grain sediments. Trench 1 excavations at the site revealed three distinct artefacts bearing horizons associated with low energy, silt dominated sedimentary contexts. Luminescence ages for the artefact bearing horizons yielded 52 ka for the Middle Palaeolithic assemblage and 24 ka for the blade-based assemblage. Typo-technological analysis of the Middle Palaeolithic assemblages indicates widespread use of Levallois technology at 52 ka at the site with diverse Levallois reductions. This 52-ka old Middle Palaeolithic assemblages adds significant data to the diverse lithic technological trajectories of MIS 3-time frame.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
DA: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Visualization, Writing–original draft, Software, Writing–review and editing. MD: Data curation, Formal Analysis, Methodology, Writing–review and editing. NA: Data curation, Formal Analysis. MC: Data curation., Formal Analysis. ZK: Investigation, Data curation, Formal analysis. VM: Writing–review and editing. PA: Investigation, Supervision, Validation, Writing–review and editing. NC: Methodology, Supervision, Validation. AP: Data curation, Formal Analysis. GJ: Writing–review and editing.
The authors declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by the Wenner-Gren foundation awarded to DA as Dissertation Fieldwork Grant entitled “Archaeological remains from Pre and Post Youngest Toba Tuff Deposits from Manneru River Basin, Andhra Pradesh. Initial survyes at Motravulapadu were funded by the National Geographic Society Early Career Grant (Grant number #HJ-163ER-17) entitled “Investigating Palaeolithic sites Associated with Youngest Toba Tuff deposits, Southeast India,” awarded to DA.
The authors acknowledge the Archaeological Survey of India and the Department of Archaeology and Museums, Andhra Pradesh, for granting permission to conduct field surveys in the region. Devara Anil thanks residents of Motravulapadu village, for their cooperation.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2023.1302419/full#supplementary-material
Akhilesh, K., Pappu, S., Rajapara, H. M., Gunnell, Y., Shukla, A. D., and Singhvi, A. K. (2018). Early Middle Palaeolithic culture in India around 385–172 ka reframes Out of Africa models. Nature 554 (7690), 97–101. doi:10.1038/nature25444
Anil, D., Chauhan, N., Ajithprasad, P., Devi, M., Mahesh, V., and Khan, Z. (2022). An early presence of modern human or convergent evolution? A 247 ka middle palaeolithic assemblage from Andhra Pradesh, India. J. Archaeol. Sci. Rep. 45, 103565. doi:10.1016/j.jasrep.2022.103565
Auclair, M., Lamothe, M., and Huot, S. (2003). Measurement of anomalous fading for feldspar IRSL using SAR. Radiat. Meas. 37 (4–5), 487–492. doi:10.1016/S1350-4487(03)00018-0
Bae, C. J., Douka, K., and Petraglia, M. D. (2017). On the origin of modern humans: asian perspectives. Science 358 (6368), eaai9067. doi:10.1126/SCIENCE.AAI9067
Berger, L. R., Hawks, J., de Ruiter, D. J., Churchill, S. E., Schmid, P., Delezene, L. K., et al. (2015). Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife 4, e09560. doi:10.7554/eLife.09560
Bergström, A., Stringer, C., Hajdinjak, M., Scerri, E. M. L., and Skoglund, P. (2021). Origins of modern human ancestry. Nature 590, 229–237. doi:10.1038/s41586-021-03244-5
Blinkhorn, J., Achyuthan, H., Petraglia, M., and Ditchfield, P. (2013). Middle palaeolithic occupation in the thar desert during the upper Pleistocene: the signature of a modern human exit out of Africa? Quat. Sci. Rev. 77, 233–238. doi:10.1016/j.quascirev.2013.06.012
Blinkhorn, J., Ajithprasad, P., Mukherjee, A., Kumar, P., Durcan, J., and Roberts, P. (2019). The first directly dated evidence for Palaeolithic occupation on the Indian coast at Sandhav, Kachchh. Quat. Sci. Rev. 224, 105975. doi:10.1016/J.QUASCIREV.2019.105975
Blinkhorn, J., and Petraglia, M. D. (2017). Environments and cultural change in the Indian subcontinent. Curr. Anthropol. 58 (S17), S463–S479. doi:10.1086/693462
Blinkhorn, J. A. (2012). The palaeolithic occupation of the thar desert. Oxford: University of Oxford.
Boëda, E. (1995). “Levallois. A volumetric construction, methods, a technique,” in The definition and interpretation of Levallois technology. Editors H. L. Dibble, and O. Bar-Yosef (Ann Arbor: Prehistory Press), 41–68.
Boivin, N., Fuller, D. Q., Dennell, R., Allaby, R., and Petraglia, M. D. (2013). Human dispersal across diverse environments of Asia during the upper Pleistocene. Quat. Int. 300, 32–47. doi:10.1016/j.quaint.2013.01.008
Bordes, F. (1980). Le débitage Levallois et ses variantes. Bull. Société préhistorique française 77 (2), 45–49. doi:10.3406/bspf.1980.5242
Brown, P., Sutikna, T., Morwood, M. J., Soejono, R. P., Saptomo, E. W., and Due, R. A. (2004). A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431 (7012), 1055–1061
Buylaert, J.-P., Thiel, C., Murray, A., Vandenberghe, D., Yi, S., and Lu, H. (2011). IRSL and post-IR IRSL residual doses recorded in modern dust samples from the Chinese Loess Plateau. Geochronometria 38 (4), 432–440. doi:10.2478/s13386-011-0047-0
Chen, F., Welker, F., Shen, C. C., Bailey, S. E., Bergmann, I., Davis, S., et al. (2019). A late middle Pleistocene denisovan mandible from the Tibetan plateau. Nature 569 (7756), 409–412. doi:10.1038/s41586-019-1139-x
Clarkson, C. (2018). “The South Asian Microlithic: homo sapiens dispersal or adaptive response?,” in Lithic technological organization and paleoenvironmental change (Berlin Germany: Springer), 37–61.
Clarkson, C., Harris, C., Li, B., Neudorf, C. M., Roberts, R. G., Lane, C., et al. (2020). Human occupation of northern India spans the Toba super-eruption ∼74,000 years ago. Nat. Commun. 11 (1), 961. doi:10.1038/s41467-020-14668-4
Clarkson, C., Jacobs, Z., Marwick, B., Fullagar, R., Wallis, L., Smith, M., et al. (2017). Human occupation of northern Australia by 65,000 years ago. Nature 547 (7663), 306–310. doi:10.1038/nature22968
Clarkson, C., Jones, S., and Harris, C. (2012). Continuity and change in the lithic industries of the Jurreru Valley, India, before and after the Toba eruption. Quat. Int. 258, 165–179. doi:10.1016/j.quaint.2011.11.007
Clarkson, C., Petraglia, M., Korisettar, R., Haslam, M., Boivin, N., Crowther, A., et al. (2009). The oldest and longest enduring microlithic sequence in India: 35 000 years of modern human occupation and change at the Jwalapuram Locality 9 rockshelter. Antiquity 83 (320), 326–348. doi:10.1017/S0003598X0009846X
Détroit, F., Mijares, A. S., Corny, J., Daver, G., Zanolli, C., Dizon, E., et al. (2019). A new species of Homo from the Late Pleistocene of the Philippines. Nature 568 (7751), 181–186. doi:10.1038/s41586-019-1067-9
Dirks, P. H., Roberts, E. M., Hilbert-Wolf, H., Kramers, J. D., Hawks, J., Dosseto, A., et al. (2017). The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. eLife 6, e24231. doi:10.7554/eLife.24231
Douka, K., Slon, V., Jacobs, Z., Ramsey, C. B., Shunkov, M. V., Derevianko, A. P., et al. (2019). Age estimates for hominin fossils and the onset of the upper palaeolithic at Denisova cave. Nature 565 (7741), 640–644. doi:10.1038/s41586-018-0870-z
Field, J. S., Petraglia, M. D., and Lahr, M. M. (2007). The southern dispersal hypothesis and the South Asian archaeological record: examination of dispersal routes through GIS analysis. J. Anthropol. Archaeol. 26 (1), 88–108. doi:10.1016/j.jaa.2006.06.001
Freidline, S. E., Westaway, K. E., Joannes-Boyau, R., Duringer, P., Ponche, J. L., Morley, M. W., et al. (2023). Early presence of Homo sapiens in Southeast Asia by 86–68 kyr at tam pà ling, northern Laos. Nat. Commun. 14 (1), 3193. doi:10.1038/s41467-023-38715-y
Galway-Witham, J., Cole, J., and Stringer, C. (2019). Aspects of human physical and behavioural evolution during the last 1 million years. J. Quat. Sci. 34 (6), 355–378. doi:10.1002/jqs.3137
Green, R. E., Krause, J., Briggs, A. W., Maricic, T., Stenzel, U., Kircher, M., et al. (2010). A draft sequence of the neandertal genome. Science 328 (5979), 710–722. doi:10.1126/science.1188021
Harvati, K., and Reyes-Centeno, H. (2022). Evolution of Homo in the middle and late Pleistocene. J. Hum. Evol. 173, 103279. doi:10.1016/j.jhevol.2022.103279
Harvati, K., Röding, C., Bosman, A. M., Karakostis, F. A., Grün, R., Stringer, C., et al. (2019). Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia. Nature 571 (7766), 500–504. doi:10.1038/s41586-019-1376-z
Haslam, M., et al. (2010) ‘Indian lithic technology prior to the searching for an early modern human signature (7)’, The upper Palaeolithic revolution in global perspective: papers in honour of Sir Paul Mellars. Katherine V. Cambridge Boyle Clive Gamble Ofer Bar-Yosef James Barrett (Eds), pp. 71–82.
Huntley, D. J., and Baril, M. R. (1997). The K content of the K-feldspars being measured in optical dating or in thermoluminescence dating. Anc. TL 15 (1), 11–13.
Huntley, D. J., and Hancock, R. G. V. (2001). The Rb contents of the K-feldspar grains being measured in optical dating. Anc. TL 19 (2), 43–46.
Huntley, D. J., and Lamothe, M. (2001). Ubiquity of anomalous fading in K-feldspars and the measurement and correction for it in optical dating. Can. J. Earth Sci. 38 (7), 1093–1106. doi:10.1139/e01-013
James, H. A., and Petraglia, M. (2005). Modern human origins and the evolution of behavior in the later Pleistocene record of South Asia. Curr. Anthropol. 46 (S5), S3–S27. doi:10.1086/444365
Jones, S. C. (2007). A human catastrophe? The impact of the 74,000-year-old supervolcanic eruption of Toba on hominin populations in India. Cambridge: University of Cambridge.
Liu, W., Martinón-Torres, M., Cai, Y. j., Xing, S., Tong, H. w., Pei, S. w., et al. (2015). The earliest unequivocally modern humans in southern China. Nature 526 (7575), 696–699. doi:10.1038/nature15696
Mijares, A. S., Détroit, F., Piper, P., Grün, R., Bellwood, P., Aubert, M., et al. (2010). New evidence for a 67,000-year-old human presence at Callao cave, Luzon, Philippines. J. Hum. Evol. 59 (1), 123–132. doi:10.1016/j.jhevol.2010.04.008
Mishra, S. (2013). Early hunter-gatherers: the upper palaeolithic. J. Field Archaeol. 6, 321–330. doi:10.1371/journal.pone.0069280
Mishra, S., Chauhan, N., and Singhvi, A. K. (2013). Continuity of microblade technology in the Indian Subcontinent since 45 ka: implications for the dispersal of modern humans. PLoS One 8 (7), e69280. doi:10.1371/journal.pone.0069280
Morwood, M. J., Soejono, R. P., Roberts, R. G., Sutikna, T., Turney, C. S. M., Westaway, K. E., et al. (2004). Archaeology and age of a new hominin from Flores in eastern Indonesia. Nature 431, 1087–1091.
Morwood, M. J., Brown, P., Jatmiko Sutikna, T., Saptomo, E. W., Westaway, K. E., Rokus , A. D., et al. (2005). Further evidence for small-bodied hominins from the Late Pleistocene of Flores, Indonesia. Nature 437, 1012–1017.
Mishra, R., and Singaraju, V. (2009). Quaternary Geological Studies of Inland Dunes of Prakasam District. Andhra Pradesh.: Unpublsihed Progress Report for the Field Season 2004-05. Geological Survey of India.
Murty, M. L. K. (1979). Recent research on the upper palaeolithic phase in India. J. Field Archaeol. 6 (3), 301–320. doi:10.2307/529657
Petraglia, M., Clarkson, C., Boivin, N., Haslam, M., Korisettar, R., Chaubey, G., et al. (2009). Population increase and environmental deterioration correspond with microlithic innovations in South Asia ca. 35,000 years ago. Proc. Natl. Acad. Sci. 106 (30), 12261–12266. doi:10.1073/pnas.0810842106
Petraglia, M., Korisettar, R., Boivin, N., Clarkson, C., Ditchfield, P., Jones, S., et al. (2007). Middle paleolithic assemblages from the Indian subcontinent before and after the Toba super-eruption. Science 317 (5834), 114–116. doi:10.1126/science.1141564
Petraglia, M. D., Ditchfield, P., Jones, S., Korisettar, R., and Pal, J. (2012). The Toba volcanic super-eruption, environmental change, and hominin occupation history in India over the last 140,000 years. Quat. Int. 258, 119–134. doi:10.1016/j.quaint.2011.07.042
Reyes-Centeno, H. (2016). Out of Africa and into Asia: fossil and genetic evidence on modern human origins and dispersals. Quat. Int. 416, 249–262. doi:10.1016/J.QUAINT.2015.11.063
Sahle, Y., Reyes-Centeno, H., and Bentz, C. (2018). Modern human origins and dispersal: current state of knowledge and future directions. Evol. Anthropol. 27 (2), 64–67. doi:10.1002/evan.21573
Skoglund, P., and Mathieson, I. (2018). Ancient genomics of modern humans: the first decade. Annu. Rev. Genomics Hum. Genet. 19 (1), 381–404. doi:10.1146/annurev-genom-083117-021749
Slimak, L., Zanolli, C., Higham, T., Frouin, M., Schwenninger, J. L., Arnold, L. J., et al. (2022). Modern human incursion into Neanderthal territories 54,000 years ago at Mandrin, France. Sci. Adv. 8 (6), eabj9496. doi:10.1126/SCIADV.ABJ9496
Stringer, C. (2002). Modern human origins: progress and prospects. Philosophical Trans. R. Soc. Lond. Ser. B Biol. Sci. 357 (1420), 563–579. doi:10.1098/rstb.2001.1057
Sutikna, T., Tocheri, M. W., Morwood, M. J., Saptomo, E. W., Jatmiko, , Awe, R. D., et al. (2016). Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia. Nature 532 (7599), 366–369. doi:10.1038/nature17179
Teixeira, J. C., and Cooper, A. (2019). Using hominin introgression to trace modern human dispersals. Proc. Natl. Acad. Sci. 116 (31), 15327–15332. doi:10.1073/pnas.1904824116
Vahdati, A. R., Weissmann, J. D., Timmermann, A., Ponce de León, M., and Zollikofer, C. P. E. (2022). Exploring Late Pleistocene hominin dispersals, coexistence and extinction with agent-based multi-factor models. Quat. Sci. Rev. 279, 107391. doi:10.1016/J.QUASCIREV.2022.107391
Van Peer, P. (1992b). “The Levallois reduction strategy,” in Monographs in World Archaeology. 13th edn. (Madison, Wisconsin: Prehistory Press).
Keywords: Levallois technology, Middle Palaeolithic, luminescence chronology, South Asia, Homo sapiens dispersals
Citation: Anil D, Devi M, Ati N, C. P M, Khan Z, Mahesh V, Ajithprasad P, Chauhan N, Pandey A and Jha G (2023) Diversity of MIS 3 Levallois technology from Motravulapadu, Andhra Pradesh, India-implications of MIS 3 cultural diversity in South Asia. Front. Earth Sci. 11:1302419. doi: 10.3389/feart.2023.1302419
Received: 26 September 2023; Accepted: 14 November 2023;
Published: 01 December 2023.
Edited by:
Nathan D. Stansell, Northern Illinois University, United StatesReviewed by:
Huw Groucutt, Max Planck Institute for Chemical Ecology, GermanyCopyright © 2023 Anil, Devi, Ati, C. P, Khan, Mahesh, Ajithprasad, Chauhan, Pandey and Jha. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Devara Anil, ZGV2YXJhLmFuaWxrdW1hckBnbWFpbC5jb20=; Gopesh Jha, Z2poYUBzaGgubXBnLmRl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.