 Michael J. Orchard
Michael J. Orchard- Geological Survey of Canada, Vancouver, BC, Canada
The Upper Triassic conodont genus Primatella is important for recognizing the Carnian-Norian stage boundary in Panthalassan and Tethyan domains. Originally based on Canadian material from Black Bear Ridge in the Western Canada Sedimentary Basin, two key species are highlighted, and their occurrence is documented in the allochthonous Wrangellia and Alexander terranes in the North American Cordillera; in the Great Basin in Nevada, western United States; at Pizzo Mondello, Sicily, in western Tethys; and in Timor-Leste on the southeastern margin of the Meso-Tethys Ocean (northeastern Gondwana). The correlation potential of Primatella species is compromised by a differing nomenclature employed in Tethyan regions of Europe where Primatella species have been assigned to other genera, namely, Ancyrogondolella (formerly Epigondolella), Carnepigondolella, and Metapolygnathus. This has obscured phyletic relationships and impacted the stratigraphic utility of two demonstrably cosmopolitan species, P. asymmetrica and P. bifida. These species are shown to occur widely in a variety of paleoenvironments and have a restricted stratigraphic range around the CNB for which they can be adopted as useful indices.
1 Introduction
Conodonts have proven to be a pre-eminent stratigraphic tool for dating and correlating Paleozoic and Triassic strata. As index fossils, they are invaluable in the recognition of events and for the definition of stratigraphic boundaries. In the Triassic, they define the base of the period as well as play an important role in at least supporting its constituent stage boundaries. Recent deliberations of the Carnian-Norian boundary (CNB) have been no exception and conodonts featured prominently in studies of the two candidate sections for the Global Stratigraphic Section and Point (GSSP) at Pizzo Mondello, Sicily (Mazza et al., 2012), and Black Bear Ridge, British Columbia (Orchard, 2014). Although both sections contain the distinctive Metapolygnathus parvus faunal interval that in North America corresponds to the latest Carnian Macrolobatus ammonoid zone, the associated conodont taxa, including those that characterize the basal Norian stage immediately above, are controversial.
Recent reclassification of Upper Triassic conodonts based on rich faunas from Black Bear Ridge (Orchard, 2013; Orchard, 2014) included the separation of a new genus, Primatella, that spans the CNB interval. This genus largely ranges within and helps define the primitia Zone sensu Orchard (Orchard, 1983; Orchard, 1991a), which was internally subdivided by Orchard (2014). The now agreed definition of the CNB is at the appearance of the bivalve Halobia austriaca (Hounslow et al., 2021), which corresponds broadly to the base of the P. asymmetrica-Norigondolella Subzone at Black Bear Ridge, and of the “Carnepigondolella” gulloae Zone in Pizzo Modello. The latter has been interpreted as a probable Primatella species by Orchard (2019).
Although P. primitia sensu stricto is uncommon and to date only known from Canada, the zonal name is retained for the sake of stability. The volume of the genus, which was originally assigned to a single species of Epigondolella, includes many similar species that have been assigned to a variety of other genera in Europe (Orchard, 2019), including the older Carnepigondolella (e.g., C. orchardi in Balini et al., 2010, pl. 3, fig. 30), the younger Ancyrogondolella (e.g., E. = A. rigoi in Mazza et al., 2010, pl. II, fig. 5 = Primatella triangulare), and the partly contemporaneous Metapolygnathus (e.g., Me. mersinensis in Mazza et al., 2012, pl. 4, fig. 7, 9). This paper reasserts the value of Primatella as a distinct genus for both intercontinental correlation and the recognition of the CNB by documenting the global distribution of two representative species established in Canada and now recognized in the United States, Europe, and Southeast Asia.
In this study, North American occurrences of Primatella asymmetrica and P. bifida are documented from northeastern British Columbia (B.C.), their type area; from Haida Gwaii, B.C. and Keku Islands, southeast Alaska, representative of, respectively, the allochthonous Wrangel and Alexander terranes that occupied an unknown but probably more southerly paleolatitude in the Late Triassic Panthalassan Ocean; and from lower latitude Nevada in the Great Basin of western United States. These successions are from various parts of Panthalassa, whereas records from the GSSP at Pizzo Mondello in Sicily are from western Tethys, and new collections from Timor-Leste come from the southeastern margin of Tethys, which was rifting from the northern edge of Gondwana in the Late Triassic (Metcalfe, 2021) (Figure 1). The two distinctive species are the focus of the present study, particularly as they occur widely but have not been consistently identified. Other Primatella species are thought to be equally widespread, and the genus is clearly a valuable indicator of the CNB.
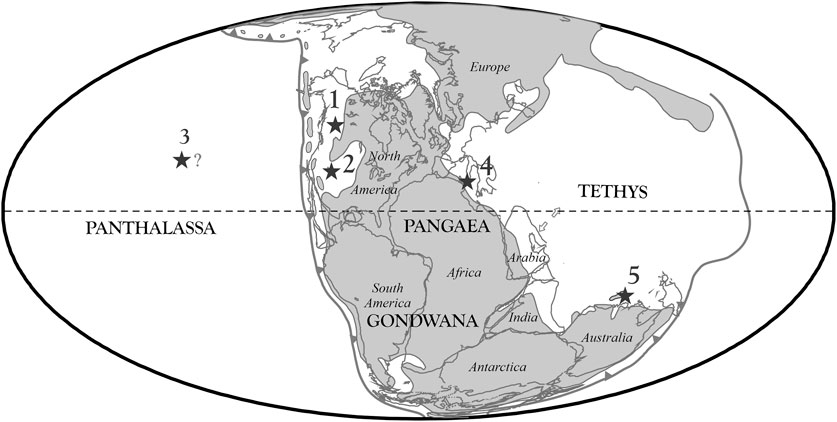
FIGURE 1. Simplified paleogeographic world map for the Triassic showing the locations of the cited areas: 1. Black Bear Ridge, northeast B.C.; 2. Berlin, western Nevada; 3. Sadler Point, Haida Gwaii—Kuiu Island, Alaska; 4. Pizzo Mondello, Sicily; 5. Timor-Leste [Based on Scotese (1998)]. Note that the northern Tethys margin differs in the updated reconstruction of Kocsis and Scotese (2021), but this does not impact the location of the studied sections.
2 Materials and methods
2.1 The locations
This report arises from the investigation over several decades of numerous Upper Triassic conodont collections from several areas of western North America, where studies have recently focused on defining a GSSP for the CNB. In particular, the Black Bear Ridge (BBR) candidate and nearby sites were sampled in considerable detail, with closely spaced bulk limestone samples subjected to standard processing procedures (Jeppsson et al., 1999). Numerous rich faunas were recovered, providing an outstanding conodont record through the boundary interval. This resulted in a substantially revised taxonomy for the interval (Orchard, 2013; Orchard, 2014) that has had some, but not universal, uptake. The taxonomy was successfully applied in both Haida Gwaii in the Wrangel terrane (Carter and Orchard, 2013) and in Nevada (Balini et al., 2015). Together, the conodont collections from these Panthalassan locations, which incidentally represent the sites from which much of the North American Carnian and Norian ammonoid zonation was developed (Silberling, 1959; Tozer, 1994), demonstrate the broad applicability of the BBR classification.
Further afield, conodont research at Pizzo Mondello (PM) in Sicily, Italy (Mazza et al., 2012), pre-dated the taxonomic revisions of Orchard (2014) but later descriptions of conodont faunas from the Tethys Realm (e.g., Rigo et al., 2018) have employed a nomenclature that has generally remained unchanged. Nevertheless, published illustrations from Europe enabled a preliminary revision of PM conodonts by Orchard (2019), who recognized several species of Primatella among those illustrated by Nicora et al. (2007), Balini et al. (2010), Mazza et al. (2010, 2012), and Mazza and Martinez-Pérez (2015) as species of Carnepigondolella, Epigondolella, and Metapolygnathus.
Far from both regions, in the southeastern Tethys on the margins of Gondwanaland, extensive investigations in Timor-Leste (McCartain, 2014; Haig et al., 2021) included the processing of numerous small conodont samples. The recovered collections have been studied by the present author, who has identified many species of Primatella, including examples of the two focus species.
2.2 The conodonts
Primatella accommodates a comparatively ornate clade that arose from Quadralella concurrent with the disappearance of Carnepigondolella in the late Carnian (Tuvalian) (Orchard, 2013). The youngest species of the latter genus (Orchard, 2014, figs. 17, 18) have been regarded by Mazza et al. (2012) as the earliest representatives of Epigondolella (now called Ancyrogondolella). However, in western Canada, a succession of Primatella species follows the disappearance of Carnepigondolella, which shows progressive diminution in size prior to its demise at the end of the samueli Zone (Orchard, 2014, figs. 17, 18). This trend is repeated later in the parvus Zone, which also contains diminutive taxa with reduced platforms. Epigondolella sensu stricto is now recognized as a middle Norian (Alaunian) genus, and similar lower Norian (Lacian) species have been reassigned to Ancyrogondolella (Orchard, 2018). The first representatives of the latter genus appear near the end of the Lower Norian Kerri ammonoid Zone in North America (Orchard, 1991a). In the author’s view, the most distinctly ornate taxa of the latest Tuvalian and early Lacian are probably species of Primatella (e.g., see Orchard, 2019).
Primatella asymmetrica and P. bifida are typical representatives of the genus in that they bear well-differentiated anterior nodes that rise above the platform margin as high apically blunt nodes compared with the low broadly rounded or incised nodes of some Quadralella species, and the high and sharply terminated denticles of Ancyrogondolella species (Orchard, 2019; fig. 4). Apart from the anterior ornament, the diagnostic morphology of P. asymmetrica is the asymmetric posterior platform with a strongly convex outer margin and inturned posterior. P. bifida has a bifurcated keel and an indented posterior margin. The supporting taxonomy is provided below.
3 Results
3.1 Northeast British Columbia
Both Primatella asymmetrica and P. bifida were first described from the CNB candidate section at BBR in northeastern B.C., located in the Peace River valley where the North American Norian zonation of ammonoids (Tozer, 1967; 1994) and conodonts (Orchard, 1983; Orchard, 1991a) was developed in the Pardonet Formation. This formation is primarily composed of gray carbonaceous limestone, calcareous and silty dolostone and shale indicative of deposition in an offshore deep marine setting on the northwestern margin of the Pangea supercontinent (Zonneveld et al., 2010).
At BBR and nearby localities, Primatella asymmetrica and P. bifida appear in upper Tuvalian strata near the base of the parvus Subzone and disappear in the lowest zone of the Norian, which is in the North American ammonoid Zone of Stikinoceras kerri and within the asymmetrica-Norigondolella conodont Subzone (Orchard, 2014). The nominal species of this conodont subzone ranges throughout the interval, whereas P. bifida disappears low in the subzone, a little above the CNB (Figure 2). Conodont associates of these Primatella species in the Pardonet Formation include diverse Primatella species as well as Acuminatella spp. and, in the Norian, common Norigondolella. In the Tuvalian part of their range, they are accompanied by Kraussodontus, Metapolygnathus, Parapetella, and Quadralella species, most of which disappear by the CNB.
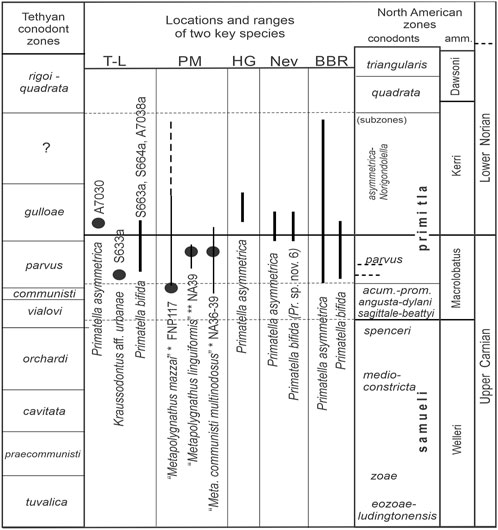
FIGURE 2. Occurrence and stratigraphic range of Primatella asymmetrica and P. bifida around the CNB in the five cited locations. T-L, Timor-Leste; PM, Pizzo Mondello; HG, Haida Gwaii; Nev, Nevada; BBR, Black Bear Ridge. Thicker bars are the observed ranges. Dots are from illustrations. For PM, thin bars are given ranges of specified taxa that are re-assigned to * = P. asymmetrica and ** = P. bifida, which originated in the sample number following it; it may not apply to all specimens. Sample numbers are given for T-L and PM.
3.2 Nevada
Upper Triassic conodonts from the Berlin-Ichthyosaur State Park in the Shoshone Mountains of central Nevada were recovered from samples collected during a restudy of the CNB in the Luning Formation (Balini et al., 2015). This site is important for both its articulated vertebrate remains and as the type section for the latest Carnian Klamathites macrolobatus ammonoid Zone (Silberling, 1959). The Triassic succession lies within the Berlin Allochthon of the Paradise terrane, a lithotectonic assemblage linked with several others thought to have originally lain along the same continental margin but possibly separated by major structural dislocations (Stewart et al., 1997).
Carbonate strata of the Upper Triassic Luning Formation are considered to have been deposited in both shallow-water and relatively deep-water marine environments. In West Union Canyon, the calcareous shale member is dominated by marls with rare intercalations of thin bedded limestones that carry ammonoids of both the Carnian Macrolobatus and Norian Kerri zones. Conodonts faunas include Primatella bifida, provisionally identified earlier as P. sp. nov. 6, and P. asymmetrica, both of which are common in the Brick Pile and North sections (Balini et al., 2015, figs. 5, 6). These species were mostly recovered from strata recognized as lower Norian, based on their ammonoids, or from underlying undated strata.
In total, the Nevadan conodont fauna is dominated by Primatella species as is commonly the case in the asymmetrica-Norigondolella Subzone of the primitia Zone in Canada; however, Norigondolella is absent, presumably reflecting some ecological barrier. Other associates are the same as in Canada, including Acuminatella. As discussed by Balini et al. (2015), the lowest collections at the Brick Pile section may date from the uppermost Carnian, whereas those from some 50 m higher and above, are typical of the Norian part of the primitia Zone.
Overlying the Luning Formation, the Gabbs Formation includes Rhaetian strata bearing Misikella posthernsteini and Zieglericonus rhaeticus (Orchard et al., 2007a), taxa that are unknown in northeast B.C. but are common in the Eurasian Tethyan successions and in allochthonous terranes in North America, including Wrangellia (Orchard, 1991b), Cache Creek (Golding et al., 2016), and Baja California (Orchard et al., 2007b). This suggests a more southerly paleolatitude for the Canadian terranes, although their longitude is uncertain. Despite differing Rhaetian provinciality, Primatella is common in all these areas around the CNB.
3.3 Haida Gwaii—Keku Islands
The North American Cordillera is composed of many allochthonous tectonostratigraphic terranes, of which Wrangellia and the Alexander terranes are the largest. The former includes both Vancouver Island and Haida Gwaii, whereas the Alexander terrane is well displayed in the Keku Islands area of southeast Alaska. The Alexander terrane is characterized by a long Paleozoic history culminating in amalgamation with Wrangellia as a composite terrane (Beranek et al., 2014), which later accreted to the western margin of the Intermontane terranes by the Middle Jurassic but was exotic with respect to the Laurentian margin during the Late Triassic. At that time, the Alaskan portion of Wrangellia in the Wrangel Mountains was estimated to lie at approximately 12°N (Trop et al., 2002). All these regions include Upper Triassic conodont-bearing successions in which Primatella is a common component.
The Upper Triassic conodont succession in Haida Gwaii was first summarized by Orchard (1991b) and later by Carter and Orchard (2013), who focussed on integrating the conodont and radiolarian successions around the CNB. These faunas occur in the Peril Formation, which mainly consists of thinly bedded siliceous dark gray to black limestone (Desrochers and Orchard, 1991) that accumulated on an intraoceanic carbonate platform built on a remnant volcanic arc. The Peril Formation carries a wholly pelagic fauna that implies deposition in deep water under low oxygen conditions. At Sadler Point in northwest Graham Island, Primatella asymmetrica occurs in association with radiolarian Assemblage 5 (Carter and Orchard, 2013), which is assigned to the “Upper primitia Zone”, or in current terminology the asymmetrica-Norigondolella Subzone of the primitia Zone. In common with BBR, associated species around the CNB in Haida Gwaii include additional Primatella species plus Acuminatella, Kraussodontus, Metapolygnathus, Norigondolella, Parapetella, and Quadralella species. As noted about Nevada, younger Rhaetian strata in Haida Gwaii include ‘Tethyan’ species, in this case, Misikella posthernsteini and Oncodella paucidentata.
Upper Triassic strata in the Keku Islands in southeast Alaska are assigned to the Hyd Group (Muffler, 1967; Katvala and Stanley, 2008). In that area, CNB Primatella occurs in the Cornwallis Limestone on Kuiu Island (Katvala and Stanley, 2008), an often oolitic fossiliferous limestone formed in a shallow-marine environment receiving clastic sediment input from terrestrial erosion. This is a very different sedimentary environment from that of the Peril Formation on Haida Gwaii, yet Primatella species dominate some collections and include P. bifida (see Supporting Taxonomy).
3.4 Pizzo Mondello
The Pizzo Mondello (PM) section in the Sicani Mountains of western Sicily, Italy, is the newly agreed locality for the CNB GSSP. It exposes the Scillato Formation, a Carnian-Rhaetian pelagic-hemipelagic succession of cherty limestones that accumulated in the western Tethyan realm (Balini et al., 2010). There, conodont zones recognized around the CNB are based on a variety of taxa (Nicora et al., 2007; Mazza et al., 2012; Rigo et al., 2018) and include Neocavitella cavitata, Norigondolella trinacriae, and Zieglericonus sp., which are unknown from contemporaneous strata in North America. Despite this provinciality, many conodont taxa are common to both regions, although neither Quadralella nor Primatella have featured in the PM literature. Orchard (2019) recognized both genera among species identified by authors as Carnepigondolella, Epigondolella, Metapolygnathus, and Paragondolella.
For example, an element of Primatella asymmetrica was regarded as a growth stage (GS5) of Metapolygnathus mazzai by Mazza and Martinez-Pérez (2015). It came from a sample from the communisti Zone immediately beneath the parvus Zone occurrence of a similar element included in M. communisti by those authors. These align with the Tuvalian appearance of P. asymmetrica at BBR.
The occurrence of Primatella bifida at PM has been discussed by Orchard (2014, 2019). An element identified as Metapolygnathus linguiformis by Balini et al. (2010) and Mazza et al. (2012) clearly has anterior nodes unlike the holotype of that species from Japan (Hayashi, 1968). On that basis, two species can be distinguished (see Supporting Taxonomy). As at BBR, this specimen of P. bifida from PM occurs in the parvus Zone.
3.5 Timor-Leste
Recent work in Timor-Leste has involved a wide-ranging study of Triassic strata (McCartain, 2014; Haig et al., 2021) that accumulated in marine basins along northeastern Gondwana on the southeastern margin of the Meso-Tethys Ocean. Upper Triassic strata include the Aitutu Formation (Audley-Charles, 1968), which includes a range of different lithological and facies associations, such as dark gray bioturbated mud-wackestone, interbedded siliciclastic mudstone beds, grainstone, and conglomerates.
Strata of the Aitutu Formation have yielded diverse CNB faunas, including most of the taxa known from the other locations described above, namely, species of Kraussodontus, Metapolygnathus, Norigondolella, Primatella, and Quadralella. Included among these are Primatella asymmetrica and P. bifida, which mostly occur in Primatella-dominated faunas that are interpreted, in the absence of typical Tuvalian taxa, to date from the lower Lacian (see Supporting Taxonomy). Both species occur in eastern Timor, while two additional faunas with P. bifida have been recovered from the central part of the country, where a nearby locality has yielded the slightly older but similar Kraussodontus aff. urbanae.
Unlike North American faunas, Acuminatella and Parapetella have not been found in Timor-Leste, but the presence of both Neocavitella cavitata and Norigondolella trinacriae in the CNB faunas mirror the Sicilian Tethyan record. Younger collections from the upper Norian and Rhaetian include Tethyan Misikella and Bajadontus, the latter being previously reported only from peri-Pacific allochthonous terranes in Mexico (Orchard et al., 2007b) and Japan (Zhang et al., 2017).
4 Conclusion
The conodont genus Primatella is a distinctive indicator for the Carnian-Norian boundary that bridges Tuvalian Carnepigondolella and Lacian Ancyrogondolella. Two representative species, P. asymmetrica and P. bifida, are shown to have a broad environmental tolerance in shallow and deep water and a cosmopolitan distribution that spans a range of North American (Pangean) paleolatitudes, distant locations in western and eastern Tethys, and in allochthonous Panthalassan terranes occupying an intermediate location.
In North America, the two species occur in the Pardonet Formation of northeastern British Columbia within the late Carnian (Tuvalian) parvus and succeed Norian (Lacian) asymmetrica-Norigondolella conodont subzones, which are equivalent to the upper Macrolobatus and Kerri ammonoid zones. The same age is given by collections in which these Primatella species occur in the Peril Formation of Haida Gwaii and the Cornwallis Limestone of Keku Islands, parts of the allochthonous Wrangell and Alexander terranes, and in the Luning Formation in the Great Basin of western Nevada. Acuminatella species also occur around the CNB in all these areas but not further afield, leading to the suggestion that the genus is an eastern Panthalassan endemic.
Some younger Tethyan conodont elements found in Haida Gwaii support a low paleolatitude (as is the case with younger Rhaetian strata in Nevada) and/or a more westerly longitude for Wrangellia. Typical Tethyan CNB successions in Sicily and Timor include some elements unknown in North America, but Primatella is an important element in all regions.
5 Supporting taxonomy
Taxonomic data for the genus and focus species, including their distribution is provided below. This is accompanied by photographic illustration (Figures 3, 4) from Orchard (2014) for BBR; Carter and Orchard (2013) for Haida Gwaii; Balini et al. (2015) for Nevada; and Mazza et al. (2012) and Mazza and Martinez-Pérez (2015) for PM. Illustrations from Timor-Leste are original unpublished images provided by E. McCartain (The University of Western Australia).
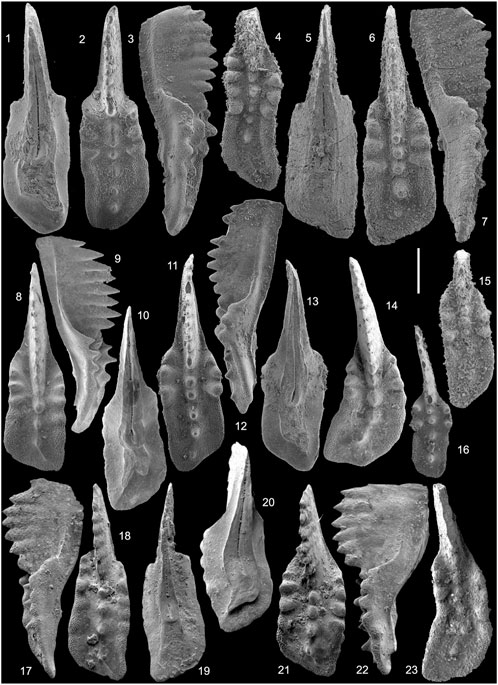
FIGURE 3. 1-22. Primatella asymmetrica Orchard, 2013. 1-3. GSC 136630 from sample 90SP-U11 (GSC cur. no. C-176937), Peril Formation, Sadler Point, Haida Gwaii; 4. NMMNH P-67736 from sample from sample BIS36, Brick Pile section, Union Canyon, Nevada; 5-7. NMMNH P-67735 from sample BIS4, Luning Formation at Berlin-Ichthyosaur State Park, central Nevada; 8-10. Micro-Unimi no. 2007 from sample FNP117, Scillato Formation, Pizzo Mondello, Sicily; 11-13. GSC 132616 from sample 18 h, Pardonet Formation, Black Bear Ridge, NE B.C.; ?14. Micro-Unimi no. 2006 from sample NA36-NA39, Scillato Formation, Pizzo Mondello, Sicily; 15. NMMNH P-67736 from sample BIS34, Luning Formation, Berlin-Ichthyosaur State Park, central Nevada; 16. GSC 132944 from sample 21h, Pardonet Formation, Black Bear Ridge, NE B.C.; 17-19. Element T507 from sample 7030b, Aitutu Formation, Timor-Leste; 20-22. Element T505 from sample 7030b, Aitutu Formation, Timor-Leste. 23. Kraussodontus aff. urbanae Orchard. T190 from sample S633a, Aitutu Formation, Timor-Leste. Scale bar = 200 µm.
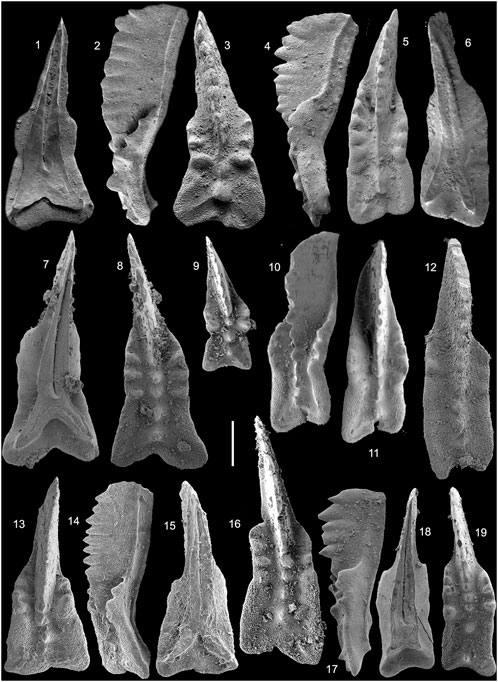
FIGURE 4. 1-19. Primatella bifida Orchard, 2014. 1-3. Element T498 from sample A7038a, Aitutu Formation, Timor-Leste; 4-6. Element T209 from sample S664a, Aitutu Formation, Timor-Leste; 7, 8. GSC 132955 from sample 21g, Pardonet Formation, Black Bear Ridge, NE B.C.; 9. GSC 132951 from sample 23, Pardonet Formation, Black Bear Ridge, NE B.C.; 10, 11. Element T205 from sample S663a, Aitutu Formation, Timor-Leste; 12. NMMNH P-67724 from sample BIS45, Luning Formation, Berlin-Ichthyosaur State Park, central Nevada; 13-15. Sample NA39, Scillato Formation. Pizzo Mondello, Sicily; 16. GSC 132954 from sample 23, Pardonet Formation, Black Bear Ridge, NE B.C.; 17-19. GSC 132956 from sample 07-22, Pardonet Formation, Black Bear Ridge, NE B.C. Scale bar = 200 µm.
5.1 Genus: Primatella Orchard, 2013
2013 Primatella gen. nov.—Orchard, p. 452-3.
2014 Primatella Orchard–Orchard, p. 87-8.
Type species: Epigondolella primitia Mosher, 1970, p. 740-41, pl. 110, figs. 8, 11, 12 (type number GSC 25051). From GSC curation number O-64654, asymmetrica-Norigondolella Subzone (Kerri Zone) of the Pardonet Formation at Brown Hill, northeast B.C.
Diagnosis-Description-Comparisons: see Orchard, 2014.
Remarks: Early studies of the Pardonet Formation conodonts led to the characterization of an “Epigondolella primitia population” in the latest Carnian and earliest Norian (Orchard, 1983; fig. 2), a composite of diverse morphotypes that commonly co-occur during this interval. These are united by the nature of the anterior platform ornament that consists of moderately high and well-differentiated but blunted nodes covered in compact microreticulation (op. cit., fig. 3). Initially, Orchard (Orchard, 1991a; Orchard, 1991b) excluded these elements from younger more strongly ornate epigondolellids (Ancyrogondolella) and re-assigned the holotype to Metapolygnathus because its pit was anteriorly shifted like that of the type species of that genus, M. communisti Hayashi. However, further study showed that several Late Triassic clades show pit migration through time, so this feature alone is judged to be insufficient for genus definition. Metapolygnathus species, which are far more common in the Tethys Realm, are now distinguished by an anterior pit and a minimal or absent platform ornament.
Orchard (2013) reviewed the classification and differentiation of Upper Triassic genera and proposed that members of the “Epigondolella primitia population” be assigned to a new genus, Primatella. This genus is thought to have evolved from Quadralella through the acquisition of a more differentiated anterior ornament of increasing amplitude (Orchard, 2014, fig. 23; 2019; fig. 4) concomitant with anterior pit migration. This trajectory later led to Ancyrogondolella in the late Kerri ammonoid Zone, represented by the “Epigondolella abneptis A” population (Orchard, 1983), in part later renamed E. quadrata (Orchard, 1991a) and now A. quadrata (Orchard, 2018). Compared with Primatella, the younger Lacian Ancyrogondolella has larger, more elevated, and sharper anterior denticles devoid of compact microreticulae (Orchard, 1983), whereas the older Carnepigondolella species generally have smaller more rounded nodes (as in C. zoae), small sharp denticles (as in C. samueli), or both (as in C. anitae), developed on less steep anterior platform margins.
The recognition of “Epigondolella” quadrata and other species of that genus (e.g., E. heinzi and E. miettoi) in the Tuvalian in Europe (Mazza et al., 2012; Rigo et al., 2018) is judged to be based on the last representatives of Carnepigondolella (e.g., C. spenceri; Orchard, 2014, figs. 17, 18) and the occurrence of unidentified Primatella species.
5.2 Primatella asymmetrica Orchard, 2013
Figure 3. 1-22.
aff. 2010 Metapolygnathus mersinensis Kozur and Moix—Balini et al., pl. 3, fig. 2a-c.
2013 Primatella asymmetrica sp. nov.—Orchard, p. 454-55, fig. 4, 19-21.
2013 Primatella asymmetrica Orchard—Carter and Orchard, fig. 7, 6-18.
2014 Primatella asymmetrica Orchard—Orchard, p. 88, fig. 61, 4-27.
2015 Primatella asymmetrica Orchard—Balini et al., fig. 11 k1-3, l1-3.
p2015 Metapolygnathus communisti Hayashi, Morphotype B—Mazza and Martinez-Pérez, pl. 6. 15.
p2015 Metapolygnathus mazzai Karádi, Kozur, and Görög—Mazza and Martinez-Pérez, . pl. 7. 15, 16.
Holotype: Orchard, 2013, p. 554-55, fig. 4 19-21 (type number GSC 132616). From bed 18h (=GSC cur. no. C-307817), middle parvus Subzone of the primitia Zone (=Macrolobatus Zone) of the Pardonet Formation at Black Bear Ridge, northeast B.C.
Diagnosis-Comparisons: see Orchard, 2014.
Remarks: Primatella asymmetrica is characterized by an asymmetric posterior platform that is expanded on its outer margin and incurved beyond that, often carrying a weak carina; the basal attachment reflects this asymmetry in the development of either a short secondary keel or lobe beneath the expansion. The pit lies at platform midlength. Mazza and Martinez-Pérez (2015) presented a growth series of Metapolygnathus mazzai from the communisti Zone of Pizzo Mondello (Figure 2). One specimen, characterized as GS5, corresponds to Primatella asymmetrica. Note that the holotype of M. mazzai (M. cf. primitius sensu Mazza et al., 2012, pl. 8, fig. 12) and additional elements assigned to that species from the Csovár borehole, Hungary (Karádi et al., 2013), do differ.
Another element illustrated from PM as a growth stage of M. communisti Morphotype B may belong here, although its posterior platform is shorter and more strongly curved than in typical P. asymmetrica (but see Orchard, 2014, fig 61. 19–21) The element figured as M. mersinensis by Balini et al. (2010), which pre-dates the known range of the present species, has a similar platform asymmetry but the anterior nodes are smaller and not as well differentiated.
In one sample from Timor-Leste, an element similar to Kraussodontus urbanae (Figure 3. 23) co-occurs with Primatella species. This element is uniformly narrow and more elongate than P. asymmetrica and has less differentiated anterior nodes, but it does have a slightly expanded outer posterior platform margin, unlike K. urbanae sensu stricto. The latter species is common in the angusta-dylani through the lower subdivision of the parvus subzones of the primitia Zone at BBR, a range that overlaps with the appearance of P. asymmetrica. The specimen is included here as a further example of similarities between the Canadian and Timor faunas.
Stratigraphic occurrence: At its type locality at Black Bear Ridge, Primatella asymmetrica occurs in at least twelve collections from the base of the Tuvalian parvus Subzone through the lower part of the Lacian asymmetrica-Norigondolella Subzone of the primitia Zone (Orchard, 2014, tables 7, 8). The inferred range of occurrences in Haida Gwaii and Nevada are consistent with this, as shown in Figure 2. In Nevada, P. asymmetrica occurs in seven collections, mostly from the Brick Pile section (Balini et al., 2015). At Pizzo Mondello, a slightly older occurrence of the species is confirmed in the communisti Zone but younger records of ‘Metapolygnathus’ mazzai in the Norian are of undetermined identity. The specimen of Metapolygnathus communisti questionably included here came from the parvus Zone at PM. In eastern Timor-Leste, sample A7030 (latitude −8.796°; longitude 126.6026°) contains several additional Primatella species but no typical Carnian taxa, so is judged to be Lacian in age. The slightly older Timor collection (sample S633a) with K. aff. urbanae occurs in the central part of the country (−8.8945°; 125.5515°).
5.3 Primatella bifida Orchard, 2014
Figure 4. 1–19.
p2008 Metapolygnathus primitius (Mosher, 1970). Katvala and Stanley, fig 40. 14 (only).
2012 Metapolygnathus linguiformis Hayashi—Mazza et al., p. 114, 116, pl. 8, fig 11.
2014 Primatella bifida sp. nov. —Orchard, p. 89-90a, fig 62. 1-15.
2015 Primatella sp. nov. 6—Balini et al., fig. 10 j1.
Holotype: Orchard, 2014, p. 89–90, fig 62. 13–15 (type number GSC 132956). From bed 07–22 (=GSC cur. no. C-307831), asymmetrica-Norigondolella Subzone of the primitia Zone (=Kerri Zone) of the Pardonet Formation at Black Bear Ridge, northeast B.C.
Diagnosis-Comparisons: See Orchard, 2014.
Remarks: Primatella bifida is characterized by a bifid variably expanded posterior platform margin and distinct well-differentiated anterior platform nodes. The basal pit lies beneath the center of the platform, posterior to which the basal scar divides and extends close to each posterolateral corner. Illustrated specimens show variation in posterior platform expansion, anterior node formation, and keel bifurcation.
The holotype of Metapolygnathus linguiformis Hayashi, 1968, despite being damaged, clearly lacks well-differentiated anterior marginal nodes, unlike the specimen illustrated by Mazza et al. (2012). Hayashi (1968) made no mention of ornament in M. linguiformis, and that species appears to lie closer to two others in which ornament is minimal: “Neogondolella” bisecta Igo, 1989, from a mixed fauna in the Mino Terrane in central Japan, and “Metapolygnathus” slovenicus Ramovs, 1994, from northwest Slovenia. The few illustrations of these taxa show incipient nodes or marginal undulation like many species of Quadralella. These two species may be synonyms, and both may be assignable to M. linguiformis, but all are excluded from P. bifida. The character of nodose ornament, or its absence, is the basis for distinguishing many Triassic conodonts and is regarded as sufficient for separating P. bifida from M. linguiformis.
Stratigraphic occurrence: At the type locality at Black Bear Ridge, Primatella bifida is common in the Tuvalian parvus Subzone through the lower part of the Lacian asymmetrica-Norigondolella sp. Subzone of the primitia Zone. Mazza et al. (2012) recorded their element of M. linguiformis from “lower Lacian” strata. The identity of unillustrated specimens is unknown, although, like P. bifida, these elements all occur in the “boundary interval” (op. cit., Figure 2). In West Union Canyon, Nevada P. bifida (provisionally identified as P. sp. nov. 6) occurs in six collections from the Brick Pile and North sections (Balini et al., 2015, figs. 5, 6), all of them within strata recognized as early Norian based on ammonoids, or from stratigraphically older but undated strata above Carnian beds. In the Keku Islands, southeast Alaska, the single specimen figured by Katvala and Stanley (2008) came from the Cornwallis Limestone, sample CPE-F2 at their locality #70 (56°56.099', 134°14.534') on the northeasternmost shore of Cornwallis Peninsula on Kuiu Island: there, it also occurs with other Primatella species and is regarded as Lacian. In Timor-Leste, three occurrences of P. bifida are recorded from the Aitutu Formation: sample S663a (−8.8663°; 125.6355°) from the center of the country appears Tuvalian as it also contains Quadralella, and nearby sample S664a (−8.8658°, 125.6353°) is probably Lacian as it contains only Primatella species, as does a third sample A7038a (−8.6344, 126.8395) from eastern Timor-Leste.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Acknowledgments
Thanks to Eujay McCartain (The University of Western Australia), whose wide-ranging study of the Triassic in Timor-Leste included the recovery, documentation, and photography of diverse conodont faunas that were shared for this study and are the subject of a future paper in preparation. This compilation represents a legacy contribution to Triassic Time and trans-Panthalassan correlations, the International Geological Correlation Program (IGCP) 467 (2003-08), and Geological Survey of Canada-funded programs to understand the evolution and assembly of the Canadian Cordillera. Viktor Karádi, Dmitry Ruban, Guichun Wu, Weiping Zeng, and one anonymous reviewer prompted improvement of this manuscript.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Audley-Charles, M. G. (1968). The Geology of Portuguese Timor. London, England: Memoir of the Geological Society of London.
Balini, M., Bertinelli, A., Di Stefano, P., Guaiumi, C., Levera, M., Mazza, M., et al. (2010). The late Carnian-Rhaetian succession at Pizzo Mondello (Sicani Mountains). Albertiana 39, 36–61.
Balini, M., Jenks, J. F., Martin, R., McRoberts, C. A., Orchard, M. J., and Silberling, N. J. (2015). The Carnian/Norian boundary succession at Berlin-Ichthyosaur State Park (Upper Triassic, central Nevada, USA). Paläontologische Z. 89, 399–433. doi:10.1007/s12542-014-0244-2
Carter, E. S., and Orchard, M. J. (2013). “Intercalibration of conodont and radiolarian faunas from the Carnian-Norian boundary interval in Haida Gwaii, British Columbia, Canada,” in The Triassic System. Editors L. H. Tanner, J. A. Spielman, and S. G. Lucas (Albuquerque, NM, United States: New Mexico Museum of Natural History and Science Bulletin) 64, 139.
Desrochers, A., and Orchard, M. J. (1991). “The Kunga Group (Late Triassic - Early Jurassic), Queen Charlotte Islands, British Columbia: stratigraphic revisions and carbonate sedimentology,” in Evolution and hydrocarbon potential of the Queen Charlotte Basin, British Columbia. Editor G. W. Woodsworth (Ottawa, Ontario: Geological Survey of Canada), 163–172.
Golding, M. L., Orchard, M. J., and Zagorevski, A. (2016). Microfossils from the Cache Creek complex in northern British Columbia and southern Yukon. Geol. Surv. Can. Open File 8033, 25. doi:10.4095/298696
Haig, D. W., Rigaud, S., McCartain, E., Martini, R., Barros, I. S., Brisbout, L., et al. (2021). Upper Triassic carbonate platform facies, Timor-Leste: foraminiferal indices and regional tectonostratigraphic association. Palaeogeogr. Palaeoclimatol. Palaeoecol. 570, 110362. doi:10.1016/j.palaeo.2021.110362
Hayashi, S. (1968). The Permian conodonts in chert of the Adoyama Formation, Ashio Mountains, central Japan. J. Earth Sci. Jpn. 22 (2), 63–77.
Hounslow, M. W., Bachmann, G. H., Balini, M., Benton, M. J., Carter, E. S., Konstantinov, A. G., et al. (2021). The case for the global stratotype section and Point (GSSP) for the base of the Norian stage. Albertiana 46, 25–57.
Igo, H. (1989). Mixed conodont elements from Hachiman town, Mino terrane, Central Japan. Trans. Proc. Palaeontol. Soc. Jpn. New Ser. No. 156, 270–285.
Jeppsson, L., Anehus, R., and Fredholm, D. (1999). The optimal acetate buffered acetic acid technique for extracting phosphatic fossils acetic acid technique for extracting phosphatic fossils. J. Paleontology 73, 964–972. doi:10.1017/s0022336000040798
Karádi, V., Kozur, H. W., and Görög, A. (2013). “Stratigraphically important Lower Norian conodonts from the Csövar borehole (CSV-1), Hungary - comparison with the conodont succession of the Norian GSSP candidate Pizzo Mondello (Sicily, Italy),” in The Triassic System. Editors L. H. Tanner, J. A. Spielman, and S. G. Lucas (Albuquerque, NM, United States: New Mexico Museum of Natural History and Science Bulletin).
Katvala, E. C., and Stanley, G. D. (2008). “Conodont biostratigraphy and facies correlations in a Late Triassic island arc, Keku strait, southeast Alaska,” in The terrane puzzle: New perspectives on paleontology and stratigraphy from the North American Cordillera. Editors R. B. Blodgett, and G. D. Stanley Jr (Boulder, Colorado, USA: Geological Society of America Special Paper).
Kocsis, A. T., and Scotese, C. R. (2021). Mapping paleocoastlines and continental flooding during the Phanerozoic. Earth Sci. Rev. 213, 103463. doi:10.1016/j.earscirev.2020.103463
Mazza, M., Furin, S., Spötl, C., and Rigo, M. (2010). Generic turnovers of Carnian/Norian conodonts: climatic control of competition? Palaeogeogr. Palaeoclimatol. Palaeoecol. 290, 120–137. doi:10.1016/j.palaeo.2009.07.006
Mazza, M., and Martínez-Pérez, C. (2015). Unravelling conodont (Conodonta) ontogenetic processes in the Late Triassic through growth series reconstructions and X-ray microtomography. Boll. della Soc. Paleontol. Ital. 54 (3), 161–186. doi:10.4435/BSPI.2015.10
Mazza, M., Rigo, M., and Gullo, M. (2012). Taxonomy and stratigraphic record of the Upper Triassic conodonts of the Pizzo Mondello section (western Sicily, Italy), GSSP candidate for the base of the Norian. Riv. Ital. Paleontol. Stratigr. 118 (1), 85–130.
McCartain, E. (2014). Stratigraphic studies on Timor-Leste, School of Earth Sciences. Perth, Australia: The University of Western Australia, 505.
Metcalfe, I. (2021). Multiple Tethyan ocean basins and orogenic belts in Asia. Gondwana Res. 100, 87–130. doi:10.1016/j.gr.2021.01.012
Mosher, L. C. (1970). New conodont species as Triassic guide fossils. J. Paleontology 44 (4), 737–742.
Muffler, L. J. P. (1967). Stratigraphy of the Keku islets and neighboring parts of Kuiu and Kupreanof Islands southeastern Alaska. Denver, CO, USA: U.S. Geological Survey Bulletin.
Nicora, A., Balini, M., Bellanca, A., Bertinelli, A., Bowring, S. A., Di Stefano, P., et al. (2007). The Carnian/Norian boundary interval at Pizzo Mondello (Sicani Mountains, sicily) and its bearing for the definition of the GSSP of the Norian stage. Albertiana 36, 102–129.
Orchard, M. J. (1983). Epigondolella populations and their phylogeny and zonation in the Norian (Upper Triassic). Fossils Strata 15, 177–192.
Orchard, M. J. (1991a). “Upper Triassic conodont biochronology and new index species from the Canadian Cordillera,” in Ordovician to Triassic conodont paleontology of the Canadian Cordillera. Editors M. J. Orchard, and A. D. McCracken (Boulder, Colorado, USA: Geological Survey of Canada Bulletin).
Orchard, M. J. (1991b). “Late Triassic conodont biochronology and biostratigraphy of the Kunga Group, Queen Charlotte Islands, British Columbia,” in Evolution and hydrocarbon potential of the Queen Charlotte Basin, British Columbia. Editor G. W. Woodsworth (Boulder, Colorado, USA: Geological Survey of Canada Paper).
Orchard, M. J. (2013). “Five new genera of conodonts from the Carnian-Norian Boundary beds, northeast British Columbia, Canada,” in The Triassic system. Editors L. H. Tanner, J. A. Spielman, and S. G. Lucas (Albuquerque, NM, United States: New Mexico Museum of Natural History and Science).
Orchard, M. J. (2014). Conodonts from the Carnian-Norian boundary (Upper Triassic) of Black Bear Ridge, northeastern British Columbia, Canada. Albuquerque, NM, United States: New Mexico Museum of Natural History and Science.
Orchard, M. J. (2018). “The lower-middle Norian (Upper Triassic) boundary: new conodont taxa and a refined zonation,” in Conodont studies dedicated to the careers and contributions of Anita Harris, Glenn Merrill, Carl Rexroad, Walter Sweet, and Bruce Wardlaw. Editors D. Jeffrey Over, and Charles M. Henderson (Bulletins of American Paleontology).
Orchard, M. J. (2019). The Carnian-Norian boundary GSSP candidate at Black Bear Ridge, British Columbia, Canada: update, correlation, and conodont taxonomy. Albertiana 45, 50–68.
Orchard, M. J., Carter, E. S., Lucas, S. G., and Taylor, D. G. (2007a). Rhaetian (Upper Triassic) conodonts and radiolarians from New York Canyon, Nevada, USA. Albertiana 35, 59–65.
Orchard, M. J., Whalen, P. A., Carter, E. S., and Taylor, H. (2007b). Latest Triassic conodonts from radiolarian-bearing successions in Baja California sur. N. M. Mus. Nat. Hist. Sci. Bull. 41, 355–365.
Ramovš, A. (1994). Conodonten aus den obersten Amphiclinen-Schichten und die Karn/Nor-Grenze im voralpinen Raum der Julischen Alpen. Cl. 4, Hist. Nat. 35 (5), 101–109.
Rigo, M., Mazza, M., Karádi, V., and Nicora, A. (2018). “New Upper Triassic conodont biozonation of the Tethyan Realm,” in The Late Triassic world. Editor L. Tanner (Berlin, Germany: Springer).
Scotese, C. R. (1998). Paleogeographic map archive. PALEOMAP project. Dept. Of Geology, University of Texas. http://www.scotese.com.
Silberling, N. J. (1959). Pre-Tertiary stratigraphy and Upper Triassic paleontology of the Union District, Shoshone Mountains, Nevada. Denver, CO, USA: U. S. Geological Survey Professional Paper.
Stewart, J. H., Silberling, N. J., and Harwood, D. S. (1997). Triassic and Jurassic stratigraphy and paleogeography of west-central Nevada and eastern California, with a correlation diagram of Triassic and Jurassic rocks. Denver, CO, USA: U.S. Geological Survey Open-File Report.
Trop, J. M., Ridgway, K. D., Manuszak, J. D., and Layer, P. (2002). Mesozoic sedimentary-basin development on the allochthonous Wrangellia composite terrane, Wrangell Mountains basin, Alaska: A long-term record of terrane migration and arc construction. GSA Bull. 114 (6), 693–717. doi:10.1130/0016-7606(2002)114<0693:msbdot>2.0.co;2
Zhang, L., Orchard, M. J., Algeo, T. J., Chen, Z-Q., Lyu, Z., Zhao, L., et al. (2017). An intercalibrated Triassic conodont succession and carbonate carbon isotope profile, Kamura, Japan. Palaeogeogr. Palaeoclimatol. Palaeoecol. 519, 65–83. doi:10.1016/j.palaeo.2017.09.001
Keywords: Upper Triassic, conodont, Primatella, taxonomy, paleogeography
Citation: Orchard MJ (2023) The utility of the Upper Triassic conodont Primatella in Tethyan-Panthalassan correlation around the Carnian-Norian boundary. Front. Earth Sci. 11:1220590. doi: 10.3389/feart.2023.1220590
Received: 10 May 2023; Accepted: 01 August 2023;
Published: 24 August 2023.
Edited by:
Yanlong Chen, Northwest University, ChinaReviewed by:
Guichun Wu, Chinese Academy of Geological Sciences (CAGS), ChinaDmitry Ruban, Southern Federal University, Russia
Weiping Zeng, Huanggang Normal University, China
Copyright © 2023 Orchard. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael J. Orchard, bWlrZS5vcmNoYXJkQG5yY2FuLXJuY2FuLmdjLmNh