 Yan Pan
Yan Pan- Department of Cultural Heritage and Museology, Fudan University, Shanghai, China
In East Asian archaeology, initial domestication and early dispersal of rice have continuously attracted scholarly interest in the recent decade, which has generated abundant new materials and revised opinions. This paper starts with a refreshed understanding of the domestication concept that emphasizes the dominant role of human behavior in the mutualistic relationship. A thorough review of the approaches to and data on reconstructing the rice story during 10,000–7,000 BP demonstrates the causally chained changes in phenotype, genotype, and human behavior in the establishment of domestication. Future studies will benefit from the revised paradigm, which has great potential to extract archaeological information to explain multiple mechanisms in rice domestication.
1 Introduction
Revealing rice domestication and dispersal of rice cultivation is crucial for understanding the social evolution in East Asian prehistory. Research progress in the recent decade has provided new ideas, approaches, and data for the exploration of the issue. First of all, the criterion for identifying domestication has gradually shifted from a phenotypic-trait-centered to a human-behavior-centered paradigm. It emphasizes the dominant role of humans in the interactive relationship between humans and domesticates. The refreshed theoretical perspective has pushed researchers to revise the existing methodologies and also develop new ones that are capable of reflecting human behaviors evidently associated with the establishment of the relationship. In this article, we intend to identify the appropriate methods of distinguishing wild and domesticated rice by conducting a thorough review of the methods proposed in the past 5 decades. This will then provide us a reliable foundation for critically reassessing the quality and validity of the archaeobotanical data of 10,000–7,000 BP in China that have been proposed to evidence the emergence and early dispersal of rice domestication. Based on the confirmation of the data that can be safely accepted, we attempt to depict a picture of the spatio-temporal distribution of the archaeological rice and propose a hypothetical framework of rice early dispersal routes that may require further information to enhance the articulation.
2 Refining the understanding of the “domestication” concept
Now, it is generally accepted that “domestication” can be described as a sustained multi-generational mutualistic relationship between humans and domesticates (e.g., Zeder, 2006; 2020; Purugganan, 2022). From this perspective, both sides of the interaction derive benefits from the establishment and long-term maintenance of this relationship. On the human side, through a certain level of manipulation, control, and care of a plant or animal, a human is able to increase the productivity of food and other resources of interest and guarantee a more stable and predictable supply. On the side of domestic plants or animals, specific phenotypic characteristics favored by people are selected, and thus, they gain a greater advantage in enhancing their reproductive success than other individuals not participating in this relationship. One of the current debates in documenting domestication relates to what kind of material evidence is essential in order to confirm the emergence of domestication. Some researchers take morphological and genetic changes in domesticated plants and animals as the core of the domestication process (e.g., Langlie et al., 2014; Martı´nez-Ainsworth and Tenaillon, 2016), while others consider them secondary consequences of domestication (Terrell et al., 2003; Smith, 2006; Zeder, 2006). Providing a constructive response to the disagreement, the explanation elaborated many times in the past 2 decades by Bruce Smith has already shed light on the issue. The creation of such a relationship involves three general categories of changes that are causally chained in the form of behavioral change → genetic change → morphological change (Smith, 2006).
Here, we intend to make a detailed statement of the mechanism suggested by Smith (2006) (Figure 1). For whatever reason, the domestication process is, in nature, initiated by a new pattern of human behavior. The new selective pressures sustainably brought by the behavior pattern functioning on target species for a period may cause changes in the genetic profile of the plant and animal population. This process may take place via two routes. One refers to the human behavior directly imposed on the plant or animal population, whose certain genotype positively responding to the behavior pattern is subsequently selected for. The other is that human environmental management activities indirectly have an impact on the plant and animal population. The artificially modified physical factors of the environment inhabited by these species would select for the genotypes favored by this human-driven environment. Then, the new selected genotypes of plant and animal species may express in visible morphological changes. When the morphological changes and their causal relationship with specific human behaviors are recognized, humans make decisions on whether they would like to encourage or enhance the morphological changes by continuing to repeat their behavior patterns. In this way, the three categories of changes form a positive-feedback recycle promoting the domestication process. This model clarifies that genotypic and phenotypic changes in plant and animal domesticates are consequences of long-term human intervention in target species, rather than the very beginning of the creation of this relationship. It means that the concern on initial domestication should be moved to the phase of human behavioral changes that differ from the interactions between hunter-gatherers and wild plant and animal species.
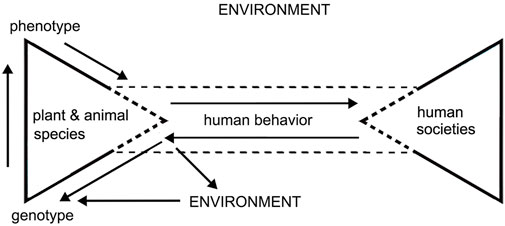
FIGURE 1. Schematic paradigm showing the relationship of behavioral change, phenotypic change, and genotypic change in the establishment of domestication. The dotted line suggests that the object framed by it is open or semi-open to the surrounding environment.
Defining domestication in terms of causal human behavior stimulates a critical issue. The question is raised as to whether it is possible for researchers to identify a demarcating line as the exact front edge of the appearance of the new human behavior pattern that later results in the establishment of domesticates. The answer is no. Numerous experiences indicate that the markers of initial domestication have been mostly recognized in a post hoc way. Only the effects resulting from human engagement in the sustained interaction with target species for a period can be reliably confirmed as markers or syndromes of domestication. The second reason is that most perennial root crops do not produce or display obviously visible morphological changes responding to human-assisted propagation. The intractable problems in preservation brought about by high precipitation, high temperature, and soil acidity add difficulties in recovering signatures that are sensitive to a slight to mild degree of human intervention and cultivation. The third reason is intimately related to the mimicry of human behavioral patterns. Human actions in the initial stage of plant domestication almost simulate the natural elements and processes of the habitat to which the targeted plant species originally adapted in order to ensure a high probability of their successful survival and propagation in the new anthropogenic environment. Thus, it is extremely hard to distinguish anthropogenic evidence, which is sporadic and elusive, from natural factors in the context of initial domestication (Smith, 2011). Despite all the difficulties in capturing the signals of the subtle initial stage of domesticates being taken care of by humans, it is still meaningful for archaeologists to approach the empirically recognizable early stage of human behavioral intervention in the life cycle of target species to modify their reproductive patterns and make them increasingly distinct from their wild relatives.
Intentionality, as pointed out by many specialists in evolutionary biology, bioarchaeology, and genetics, is the pivotal feature in the domestication relationship with human involvement (e.g., Schultz et al., 2005; Zeder, 2006; 2020; Purugganan, 2022). The domestication relationship is not restricted to the mutualism between humans and other species. Non-human associated mutualism between insects, beetles, termites, and fungi is also defined as domestication (e.g., Harrington, 2005; Mueller et al., 2005; Schultz et al., 2021). In human-associated domestication, it is human intentionality that initially triggers the mutualistic relationship, making the intervention between the two ends of the equation dominated by humans. In comparison, the two ends are more balanced in the other domestication relationships. Humans may deliberately or unconsciously take on actions without predicting genetic or phenotypic responses of plant species. They are unable to foresee the evolutionary consequence resulting from long-term changes successively taking place, together with social evolutions. Regardless of these facts, intentional purposes as the core appeal involved in human behaviors are concrete and cannot be excluded. It is where human agency comes into the picture that distinguishes between domestication and other similar mutualistic relationships in nature (Schultz et al., 2005; Zeder, 2006). Archaeological studies are normally required to reveal a long-term sustained domestication relationship by empirical evidence in advance and then trace the line further back to touch the earlier phase of forming or consolidation of the relationship.
3 Recognizing rice domestication
The improved understanding of domestication stimulated an increasing number of archaeologists to rethink and reexamine the appropriateness of a few widely employed approaches and the criteria for recognizing plant and animal domestication. Great attention has been paid to several of the most popular crops in the world, such as rice, wheat, and maize (e.g., Emshwiller, 2006; Crawford, 2012; Willcox, 2012; Faris, 2014). These annual seed plants are thought to respond to human management quickly compared to other kinds of plants. Three categories of changes can all be detected in archaeological records. Recent advances in analytical technologies allow researchers to integrate different lines of evidence and promote the ability to interpret genetic, physiological, behavioral, and cultural components during the process (Smith, 2006; Langlie et al., 2014). Here, we will concentrate our discussion on the appropriate methods of recognizing rice domestication and try to explain the intimately associated human behavioral implications. In the last 50 years, massive efforts have been invested in the study of rice domestication and have generated many methods of distinguishing wild and domesticated states of different forms of rice materials, laying a foundation for exploring the process on multiple layers. A series of macrofossil and microfossil analytical methods will be the major part of our reexamination. The underlying mechanisms of rice domestication are interpreted by articulating the causally chained successive changes in phenotype, genotype, and human behavior involved in each method.
3.1 Macrofossil evidence and its human behavioral implication
3.1.1 Morphology of spikelet base
Reduction of seed shattering is a key domestication trait that takes place in an early phase of human intervention in the rice life cycle by a set of specific harvesting and reproducing strategies. Genetic studies demonstrate that the gene mutation dominating this phenotypic feature, such as sh4, SHA1, and qSH3, existed prior to the genetic differentiation of subspecies in Oryza sativa, a very early stage of rice domestication (Li et al., 2006; Lin et al., 2007; Sang and Ge, 2007; Ishikawa et al., 2022). In morphology, the scar at the spikelet base derived from separating the seed from the branch shows varied shapes due to differing development of the abscission layer controlled by these alleles. The clarification of this mechanism provides a reliable genetic foundation for the application that the morphology of spikelet base breakage can be used as a visible indicator of determining the strength of seed shattering in a rice population. The profile change of a non-shattering spikelet base in an archaeological sample distinct from that in a wild rice population signals a sustained period of selective pressure, which is commonly induced by year-by-year harvest through panicle cutting and seed sowing. It enables researchers to detect human behavioral changes approaching the beginning of their intentional taking care of rice.
The identification criteria for attributing a specific form of spikelet base to domestic or wild type remains controversial, although some researchers have attempted to propose their own strategies. Zheng et al. (2007) believe that wild, japonica, and indica types of rice can be distinguished through the shape of spikelet base breakage. Fuller et al. (2009) insist that the spikelet base of immature harvested rice is distinguishable from that of mature seeds, and mature specimens can be divided into wild and domestic types. However, neither of them has ever provided a baseline reference generated from systematic quantitative analysis of modern samples attributed to different domestication intensities. Later, Pan (2011); Pan (2017): 221–244) conducted an experimental archaeology project in order to test the criteria for correctly linking spikelet base morphology to the corresponding anthropogenic status, i.e., wild versus domestic, japonica versus indica, and mature versus immature. The results evidence that japonica and indica samples cannot be distinguished in terms of spikelet base morphology, and immature harvested spikelets do not show distinguishable diagnosis at the breakage scar. She, therefore, recommended a categorial duality principle for identifying the archaeological specimens as wild-type and domestic-type. The experiment further ascertains the quantitative criteria for tracing the initial phase of seed shattering reduction with a high confidence level. A rice sample with a domestic-type spikelet base percentage above 20% can be safely identified as a domestic population, and a sample with a domestic-type spikelet base percentage lower than 10% can be identified as a complete wild population. The sample with domestic-type specimens between 10% and 20% falls within a range overlapped by this parameter of wild and domestic samples, so its domestication intensity should be identified as indeterminant in terms of seed shattering. In reality, such an indeterminant sample has not been recognized in the archaeological records in China.
Archaeobotanical studies at a series of sites, including Huxi, Tianluoshan, Xuenan, Baligang, Jiahu, Majiabang, Caoxieshan, Sushui River Valley, etc. (e.g., Gao, 2012; Deng et al., 2015; Zheng et al., 2016; Zhang et al., 2018; Song et al., 2019; Qiu et al., 2021; Huan et al., 2022a), documented the spatial and temporal variations of the percentage of domestic-type individuals in a rice spikelet base assemblage during 9,000–4,000 BP. Despite the increase in data, the archaeological records involved in documenting the early phase of seed shattering reduction are still scarce. Analyses of rice spikelet bases have been reported from no more than six sites dated to 10,000–7,000 BP (Table 1). According to the data published so far, all of them yielded rice populations with a domestic-type spikelet base percentage higher than 20%, the minimal level for being identified as domestic. In addition, very few sites have provided a chronologically continuous profile recording the change in seed shattering from wild to domestic, but the data from phase 1 and phase 2 at Jiahu are reported separately. This means that there is great potential to trace human manipulation of rice to a pioneering period much earlier than what we have previously discovered.
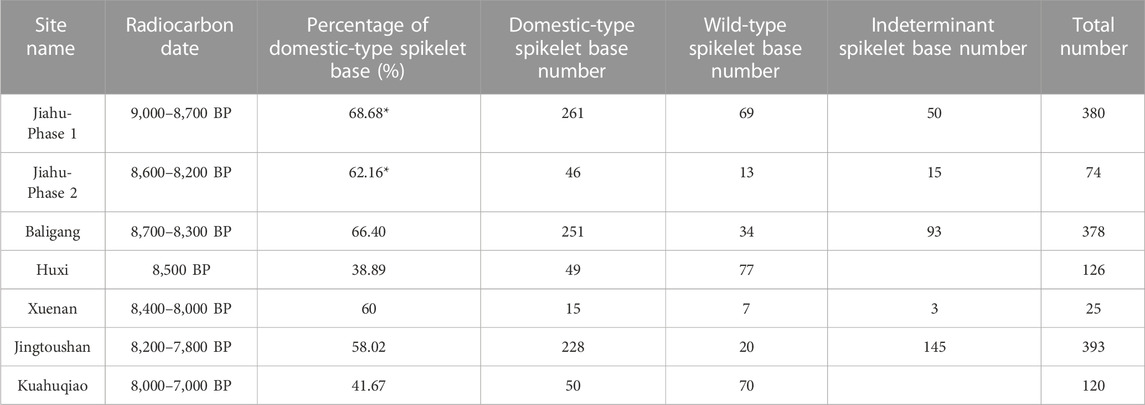
TABLE 1. Percentage of domestic-type spikelet base of the archaeological context dated to before 7,000 BP [based on the data from Zheng et al. (2007); Deng et al. (2015); Zheng et al. (2016); Zhang et al. (2018); Qiu et al. (2021); Luo, (2022)] *The original report did not include indeterminant spikelet base number in calculation of the percentage of domestic-type spikelet base, but it is included here.
In large measure, human selection for non-brittle rice spikelets is also inevitably accompanied by several other phenotypic traits to be selected and altered. These include simultaneous maturation of seeds, seed compaction on highly visible terminal stalks, and loss of seed dormancy (Smith, 2006). From a behavioral perspective, the selective pressure resulting from new human-created environments gives rise to new rules of evolutionary success in the target rice population (Smith, 2006: 18). This can also be explained by genetic mechanism. For example, a genetic study reveals that shattering QTLs and dormancy QTLs are linked to each other in several chromosomal regions of the rice gene (Cai and Morishima, 2000). Although these associated domestication syndromes are invisible in archaeological rice remains, changes in seed shattering reasonably imply that humans sustain the particular relationship with rice through activities including harvesting, storing, and broadcasting for clear purposes.
3.1.2 Seed size
Change in seed size of annual plants is also a visible marker that has been commonly analyzed to indicate the status of domestication (Smith, 2006; Zeder, 2006), but whether this criterion is applicable to identifying the initial or early stage of rice domestication remains problematic (Liu et al., 2007; Crawford, 2012; Pan et al., 2017). First of all, the enlargement of seed size is not verified by the rice remains from an array of sites dated to earlier than 6,000 BP (Liu et al., 2007). Compared to seed shattering reduction, seed size increase seems to be a markedly lagged response adapted to human manipulations operated on the rice population. Secondly, changes in seed size variability distinct from the wild rice population have been repeatedly observed in archaeological samples from before 7,000 BP (Liu et al., 2007; Gao, 2012; Deng et al., 2015), but very little progress in statistical analysis and explanation of the phenomena has been made. Although it was 15 years ago that Liu et al. (2007) pointed out the issue for the first time, her observation and suggestion are still valuable today for digging up the human behavioral implications of seed size variability during initial rice domestication. Generally speaking, the phenotypic variation amplified due to plant and animal species populations exposed to altered selective contexts is widely seen in evolutionary processes (e.g., Price et al., 2003; Kelly et al., 2012; Evin et al., 2015). It is not difficult to understand that human care removed or restrained the natural selections operated on the wild rice population so that the abnormally grown seeds that were originally not able to survive until complete ripening would have the chance to grow and develop into a thin or sterile state (not immature). Their contribution to the seed stock for broadcasting in the next growing season will be conserved and even enhanced in a population. The validity of this trait indicating domestication requires sufficient verification derived from archaeological materials and comparison with modern reference samples. Thirdly, another shift in rice seed size, whereby the ratio of length to width of grain tends to decrease during 8,000–6,000 BP, was recognized by Deng et al. (2015) and Gao (2012). However, this tendency seems a little more subtle. The measurement of carbonized rice grain and statistical analysis may be influenced by a variety of factors, including rice variety, temperature and duration of firing during seed carbonization, sample size, etc., so the dynamics of change in the ratio of length to width should be carefully examined by multiple hypotheses. In addition, thickness has never been included in seed shape analysis (Crawford, 2012: 616). If the seed becoming fatter in the early stage of rice domestication could be verified, it would be worthwhile to discern the genetic and behavioral mechanism of why the seed size changes in this manner. In sum, for many decades, numerous efforts to figure out a generalized pattern of seed size change indicating initial rice domestication through conventional measurement and statistical analysis have not arrived at a satisfying accomplishment. It also implies that a new method is necessary. The computer-assisted morphometric method of processing data on seed morphological variation for distinguishing wild and domestic populations designed by Rovner and Gyulai (2007) may have great potential in this realm.
3.2 Microfossil evidence and its human behavioral implications
3.2.1 Morphometrics of double-peaked tubercle phytolith
In 1998, 25 years ago, Pearsall et al. (1995) and Zhao et al. (1998) developed a set of prediction formulas based on systematic discriminant analysis of double-peaked tubercle phytolith morphology of a series of modern rice samples for identifying wild and domestic specimens. The method was successfully employed in documenting the emergence of rice cultivation around 10,000 BP and the subsequent intensification of the domestication process until 7,000 BP at the Diaotonghuan site (Zhao, 1998). It was also used to investigate the double-peaked phytolith archaeologically unearthed from the Lower Yangzi River (Wu et al., 2014) and the Lower Huai River (Luo et al., 2016; Gu et al., 2022; Qiu et al., 2022). An increased intensity in rice domestication during 12,000–7,000 BP was documented in the two regions. The advantage of this morphometric method depends on the effective statistical method, but the relationship between human behavioral changes pertaining to cultivation and morphological changes in double-peaked phytolith has never been discussed.
3.2.2 Morphometrics of bulliform phytolith
The investigation and application of bulliform or fan-shaped phytolith has a longer history in the study of rice remains. Fujiwara’s (1993) pioneering work developed a discriminant formula based on his statistical studies of various rice varieties in Japan for distinguishing japonica and indica subspecies. However, his method is not ideally applicable to exploring archaeological evidence of rice agriculture in mainland East Asia and many adjacent areas because it did not include the precise identification of wild rice. It was noticed that the wild specimens might be incorrectly identified as japonica or indica (Wang and Lv, 2012; Wang et al., 2019a). Since, efforts to distinguish bulliform phytolith from wild and domestic rice through a morphometric index have been made by several researchers (Pearsall et al., 1995; Zhang et al., 1998; Gu, 2000; Ma and Fang, 2007), but it has proven to be highly difficult to generate a reliable quantitative standard for this aim. The morphometric parameters of bulliform phytolith are influenced by a number of factors, so the profiles of wild and domestic samples are usually overlapped with each other.
3.2.3 Fish-scale shaped decorations of bulliform phytolith
Another feature, the number of fish-scale-shaped decorations along the bottom of bulliform phytolith, was intensively investigated by Houyuan Lv’s team to set up a new identification standard. They noticed that the bulliform phytolith of domesticated rice normally showed 8–14 fish-scale-shaped decorations, while that from wild rice commonly had less than 9 (Lu et al., 2002). A reference baseline for identification was provided by the systematic sampling of surface soil in a wild, domesticated rice field and other vegetation, as well as the quantitative analysis of phytolith assemblages in these samples (Huan et al., 2015; Huan et al., 2020). This indicates that the proportion of bulliform phytoliths with ≥9 fish-scale decorations (abbreviated to “PBFS” as follows) in domesticated rice soil samples was 57.6% ± 8.7%, while the PBFS in wild rice soil samples was 17.46% ± 8.29%. However, the PBFS in dry rice field soil samples was 11.5% ± 5.3%. This means that domesticated dry rice cannot be distinguished from wild rice by this method. Considering the limited number of available dry rice samples (only four), it requires more extensive data collection and dynamic explanation of the phenomenon to replenish the current understanding. Lv’s team also proposed explanations of the linkage between the morphological change in fish-scale decorations and the biotic/abiotic factors modified during initial rice domestication (Huan et al., 2020). The bulliform cells in living grasses, like rice, play an important role in leaf rolling in response to environmental stresses such as water loss and high temperatures (e.g., Moulia, 2000; Li et al., 2010; Zou et al., 2014; Zhang et al., 2015; Matschi et al., 2020). The increase in the number of fish-scale decorations is caused by frequent folding, shrinkage, and distortions of bulliform cells squeezed by surrounding mesophyll, sclerenchyma, and parenchyma cells during leaf rolling. It is also supported by genetic studies indicating that a major part of rice leaf rolling genes cloned so far are relevant to the development of bulliform cells, and only a few are relevant to other cells (Zhou et al., 2018). Therefore, this morphological change may be regarded as a drought resistance response to the environmental fluctuations that may have been associated with water availability, salinity, and other hydrological conditions most probably altered by human behaviors. Human-sustained year-by-year perturbations and interventions in wild rice habitats can be reasonably considered as a dynamic leading to a new hydrological regime. Although people usually tend to mimic the physical conditions of or directly take advantage of the habitat where the wild rice population is grown during initial rice domestication, rice might be sensitive to such stresses, which would stimulate immediate physiological effects. Besides the above explanation, agronomic and genetic research has also indicated that moderate leaf rolling facilitates photosynthesis efficiency and raises grain yield (e.g., Lang et al., 2004; Zhao et al., 2016; Cho et al., 2018). Selection for rice plants with a high yield may be concurrently related to the selection for the leaf rolling phenotype. Therefore, the increase in the number of fish-scale decorations of bulliform cells may also be one of the consequences of purposeful yield enhancement.
Compared to the studies of macro-plant remains, interpretation of PBFS of archaeological data may require more careful verification of depositional context, chronological background, associated artifacts, and even material processing procedure. PBFS value is not an indicator directly related to the intensity of rice domestication. Its implication of rice cultivation is indirectly inferred based on the mechanism through which rice responds to environmental stresses by leaf rolling. Anthropogenic factors may or may not contribute to the formation of these external stresses. Therefore, to what extent and in what way human activity causes changes in PBFS should be cautiously differentiated. So far, the PBFS in most archaeological soil samples dated to 10,000–7,000 BP ranges between 20% and 60%, which falls within the range of between average wild rice and domesticated rice. The interpretation that these rice populations were cultivated by people and the domestication process had already been initiated is reasonable. The complication exists in how to properly interpret the relative difference of PBFS values. To document a continuous domestication process, the PBFS values of the samples from one site or one region are generally expected to show an increasing tendency through time and transcend the PBFS of wild rice. However, the chronological changing pattern of PBFS is not consistent with the theoretical expectations and varies in each region, which adds challenges to clarifying the roles played respectively by the physical environment and human activity in the long-term evolutionary process. This also reminds us that the inter-regional comparison and interpretation of PBFS records needs to take more variables into consideration.
3.2.4 Phytolith in spikelet base
A new type of phytolith, FUSIFORM ECHINATE, observed in the rice spikelet base has been reported recently (Ge et al., 2022). The primary analysis of wild and domesticated rice specimens of the AA genome showed that fusiform echinate phytolith abundance was significantly different between shattering and non-shattering groups. The fusiform echinate phytolith abundance was reported to be 264.84 ± 162.8 for the O. nivara and O. rufipogon combined group and 771.75 ± 383.22 for the O. sativa group. Thus, the higher fusiform echinate phytolith abundance in one spikelet base may indicate a non-shattering phenotype of rice. This inspiring discovery provides a new line of evidence directly related to the loss of seed shattering, but its applicability to differentiating wild and domesticated specimens from archaeological contexts still needs to be improved. One problem regarding its reliability is the small sample size currently available. A large sample normally requires at least 30 specimens for statistical analysis, but each group reported in the study contained less than 7. The large standard deviations of the two groups mean that the data is spread far out, some of it being far away from the mean. Another issue, which may be minor and easier to resolve, relies on whether the fusiform echinate phytolith is exclusively found in rice. Therefore, a more systematic analysis of fusiform echinate phytolith in different rice varieties is expected.
3.3 Summary of methodology
To sum up, we suggest that the following criteria can be safely accepted as empirical evidence for recognizing rice domestication, especially in its early stages. The appearance of these visible or detectable phenotypic traits can all be unequivocally connected to and explained by certain genetic and behavioral mechanisms. Human intentionality is presented by a long-term sustained human-induced management or selective pressure imposed on rice, which results in highly indicative traits. The methods are listed by priority.
(1) The percentage of domestic-type rice spikelet base can be used as evidence of the highest priority as it is directly associated with human harvest strategy, and seed shattering reduction is most probably the earliest domestication trait.
(2) The proportion of bulliform phytoliths with ≥9 fish-scale decorations (PBFS) is sufficient to demonstrate rice domestication, but it is less directly connected to explicit human behavior. Rather, the variation of PBFS indicates some complicated relationships among rice, human behaviors, and physical conditions. The anthropogenic factors must be carefully verified.
(3) The morphometrics of double-peaked tubercle phytolith is also sufficient in indicating domestication, but its genetic and behavioral mechanisms are relatively unclear.
(4) To some extent, seed size may reflect the intensity of rice domestication. However, this criterion is not properly applicable to distinguishing the initial stage as seed size change may take place long after humans begin to foster wild rice, and the genetic and anthropogenic mechanism causing the change is only partly known.
4 Geographic distribution of archaeological evidence during 10,000–7,000 BP
The archaeological sites yielding rice remains and the associated phenomena during 10,000–7,000 BP have served to outline a spatial and temporal framework of the threshold of rice domestication and its dispersal immediately following the scenario (Figure 2). Based on the above methodological clarifications, we will carefully examine the primary data and the demonstration of evidence by region.
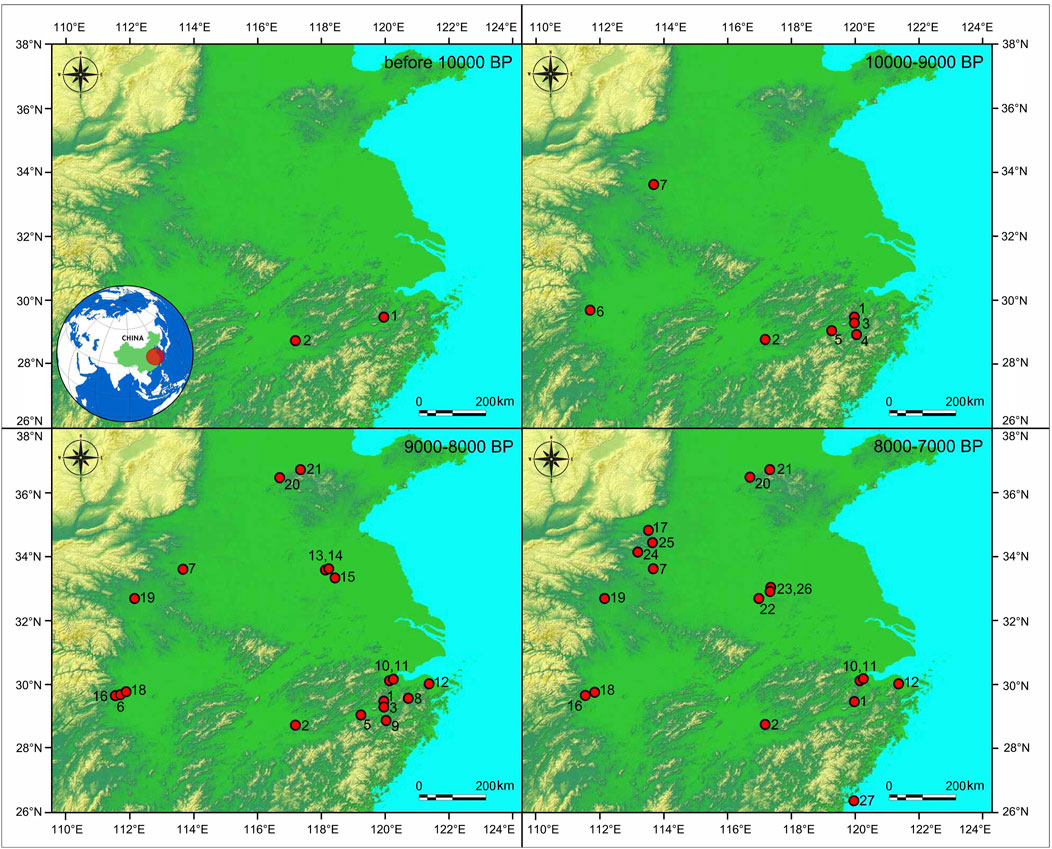
FIGURE 2. Maps of the archaeological sites yielding rice remains of 10,000–7,000 BP in China. 1. Shangshan; 2. Xianrendong and Diaotonghuan; 3. Qiaotou; 4. Miaoshan; 5. Hehuashan; 6. Pengtoushan; 7. Jiahu; 8. Xiaohuangshan; 9. Huxi; 10. Kuahuqiao; 11. Xiasun; 12. Jingtoushan; 13. Shunshanji; 14. Hanjing; 15. Xuenan; 16. Shanlonggang; 17. Zhuzhai; 18. Bashidang; 19. Baligang; 20. Yuezhuang; 21. Xihe; 22. Xiaosungang; 23. Shuangdun; 24. Tanghu; 25. Peiligang; 26. Yuhuicun; 27. Dapingding.
4.1 The Middle Yangzi region
The Middle Yangzi region is one of the areas yielding the earliest dated archaeological records of rice cultivation. The sites bearing rice remains of 10,000–7,000 BP are scattered across a large area extending from the main stream to tributaries. Since the end of the last century, the Diaotonghuan site has been widely accepted as a representative of early rice cultivation in East Asia, and the Xianrendong site, despite its scarcity of rice remains, has been attributed to the same culture due to its very close location and similar pattern of artifact assemblage composed of pottery sherds, lithics, and animal bones. The thousands of years-long rice cultivation at Diaotonghuan was demonstrated by a continuous increase in proportions of domestic double-peaked phytolith from layer G to B (Zhao et al., 1998). A dramatic shift throughout the profile is shown in layers D and E, so their radiocarbon dates were crucial. The youngest radiocarbon date of the two layers is 15,531 ± 214 cal. BP, which is derived from the bone of layer D (School of Archaeology and Museology, Peking University and Jiangxi Provincial Institute of Cultural Heritage and Archaeology, 2014: 266–267). However, it seems to have been rejected by some researchers (e.g., Kuzmin, 2006). Eventually, Zhao (1998) accepted the chronology in terms of the seriation of artifact assemblages and ceramic typology proposed for the Neolithic culture sequence in southern China. A transition from wild rice utilization to rice cultivation starting by 11,000 BP and lasting until 7,000 BP is documented in this area. No macro-plant rice remains dated to around 10,000 BP have been uncovered so far.
Pengtoushan and Bashidang of 9,000–8,000 BP, located 530 km away from Diaotonghuan in the west, are the subsequent important sites. The rice remains from Pengtoushan are the husks and chaff tempered in pottery sherds, so the observation is constrained to the micro-scale impressions of the double-peaked tubercle structure of the husk. The researchers suggested that morphometric analysis of double-peaked structure revealed an ancient “japonica-prone” rice population with mixed characteristics of wild, japonica, and indica varieties (Zhang et al., 2003). A morphometric analysis of carbonized rice grains from Bashidang, conducted by the same researcher, classified them as “a primitive ancient cultivated population that was evolving towards indica type” (Zhang and Pei, 2002). However, our understanding of these descriptions should not be restricted to the literal meaning. In fact, Zhang and his colleagues intended to emphasize that the rice remains from the two sites can hardly be categorized as japonica, indica, or wild, although they might have been cultivated then. These were published 20 years ago. No updated research has been reported. However, rice cultivation associated with groundwater table fluctuations in 8,600–8,000 BP at Bashidang is evidenced by double-peaked phytolith and pollen data (Liu et al., 2017). In addition, Shanlonggang, a nearby site attributed to the late phase of Pengtoushan culture and dated to around 8,000 BP, yielded carbonized seeds including rice. The morphometric analysis of rice grains based on the discriminant formula developed by Zhao and Gu (2009) categorized 44% of specimens as domestic type and 56% as wild type (Gu et al., 2016). Although the evidence still seems ambiguous and needs strengthening, the rice remains from Pengtoushan culture have been generally accepted as being domesticated.
4.2 The Lower Yangzi region
The Lower Yangzi region has yielded the richest materials for documenting early rice domestication spanning a wide chronological range from 10,000 to 6,000 BP. The earliest phase of Shangshan culture can be traced back to more than 10,000 years ago (Zhejiang Provincial Institute of Cultural Heritage and Archaeology and Pujiang Museum, 2016). A major part of pottery sherds are densely tempered with rice husks and chaff. However, convincing macro-plant evidence of domestication prior to 8,500 BP is not yet available. The earliest dated evidence comes from the Huxi site (Zheng et al., 2016), but the most intriguing point of it has been largely neglected. The rice spikelet base morphology and the change in rice phytolith assemblages are complementary to each other in verifying loss of seed shattering no later than 8,600 BP. The percentage of non-brittle spikelet bases being 38.89% is sufficient to indicate a sustained selective pressure favoring seed retention. The phytolith analysis of the profile in a probable ditch structure showed a relatively high density of double-peaked tubercle phytoliths with a few bulliform phytoliths during 9,000–8,400 BP compared to a lower density of double-peaked tubercle phytoliths, with huge increases in bulliform phytoliths after 8,400 BP. This means that, in the later phase, more spikelets were removed from the wetland environment and more rice leaves remained, indicating a panicle-cutting harvest strategy likely being practiced at Huxi. It might have accelerated a continuous increase in anthropogenesis involving rice cultivation and management of several species of grass sharing the niche. A long-term process of loss of seed shattering has been documented in the subsequent cultures of Kuahuqiao, Hemudu, Majiabang, Songze, and Liangzhu (Zheng et al., 2007; Allaby et al., 2017), but most of the existing evidence is from later than 7,000 BP. Exploration focusing on seed shattering in the early-middle Holocene still needs work.
An abundance of phytolith analyses also evidenced rice domestication, in particular, clarifying rice cultivation during 10,000–8,500 BP (equivalent to the early and middle phases of Shangshan culture). Both double-peaked tubercle phytoliths and bulliform phytoliths have been carefully investigated at many sites. A gradual process of rice domestication throughout the Neolithic period was demonstrated by phytolith data (Huan et al., 2014; Wu et al., 2014; Ma et al., 2016; Huan et al., 2021; Huan et al., 2022b) (Figure 3A). It indicated that rice domestication was initiated in Shangshan culture and its intensity approaching the level of modern rice agriculture took place after the Late Majiabang culture, approximately dated to 6,500–6,000 BP. More detailed research focusing on Shangshan culture, including five sites, revealed a complicated spatial and temporal pattern of the early stage (Huan et al., 2014; Wu et al., 2014; Ma et al., 2016; Huan et al., 2021). The rice domestication intensity indicated by PBFS slowly improved during 10,000–9,000 BP; it reached a relatively high level in 9,000–8,500 BP and slightly declined with fluctuations after 8,500 BP (Figure 3B). The PBFS value of the Hehuashan site is markedly higher than those of the other sites, which was explained by its location closer to the main channel of the Upper Qiantang River (Huan et al., 2021). However, phytolith analysis at the same site conducted by Qiu et al. (2019) did not show similar results, probably because of the different sampling strategy and location (Qiu, 2021). Although the tendency of rice domestication is generally consistent with an expected pattern, it is necessary to conduct detailed site-by-site investigations to figure out the complex interrelationship of human behavior, water environment, and rice responses to various changes and comprehensively understand the homogeneity and diversity of human behaviors of Shangshan culture.

FIGURE 3. Charts of the changes in rice phytoliths from the Neolithic sites in the Lower Yangzi region. (A) Chronological change in the proportion of bulliform phytolith with ≥9 fish-scale decorations throughout the Neolithic period [based on the data adopted from Wu et al. (2014); Huan et al. (2014); Ma et al. (2016); Huan et al. (2021); Huan et al. (2022a)]. The red dotted trendline calculated by moving average shows an increasing tendency. (B) The proportion of bulliform phytolith with ≥9 fish-scale decorations in the Shangshan Culture and Kuahuqiao Culture sites [based on the data adopted from Wu et al. (2014); Huan et al. (2014); Ma et al. (2016); Huan et al. (2021)].
Only two sites, Kuahuqiao and Jingtoushan, as well as some strata in Shangshan culture sites are attributed to 8,000–7,000 BP. Rice domestication at Kuahuqiao was well verified by multiple lines of evidence (Zheng et al., 2007; Pan et al., 2017). The newly excavated Jingtoushan site yielded carbonized rice grains (Sun et al., 2021). The analysis of rice spikelet base morphology showed at least 60% of specimens identified as domestic type, clearly indicating that rice domestication was underway (Luo, 2022). However, the pollen and phytolith data of a geological core at the edge of the Jingtoushan site did not provide robust evidence for local rice cultivation (Liu et al., 2020; Deng et al., 2021). Considering that the subsistence pattern of Jingtoushan is characterized by marine resource exploitation, the relationship between Jingtoushan people and the source of domestic rice should be examined with caution.
4.3 The Upper Huai River region
This region is characterized by its transitional topography and terrain. Stretching north-south along the west part of the Huang-Huai-Hai Plain, the sites are scattered on the interface where the Huai River and the Qinling Mountain meet. Many streams that originate here run southeast and converge with the Huai River, forming a densely connected river network that may provide abundant water resources for local agricultural practice. Rice cultivation in the region has a deep history rooted in the subsistence of Peiligang and Yangshao cultures (e.g., Yang et al., 2016; Wang et al., 2017; Wang et al., 2019b; Wang et al., 2019c; Cheng et al., 2022a; Huan et al., 2022a; Cheng et al., 2022b; Sun et al., 2022).
The Jiahu site dated to 9,000–7,500 BP yielded the oldest rice remains in the Upper Huai River. A recent study of rice spikelet base evidenced that the rice population was domesticated during 9,000–8,200 BP (Zhang et al., 2018). Rice domestication was also supported by the morphometric analysis of carbonized rice grains, revealing not only their larger size compared to the counterparts of other later Neolithic sites (Liu et al., 2007) but also a wider distribution of measurements uncommonly seen in the wild rice population (Zhang et al., 2009).
The Baligang, Zhuzhai, and Tanghu sites all have long chronological sequences from the Peiligang period to the Zhou period, providing well-documented sequences of agricultural development. According to archaeobotanical research, the agricultural pattern in the earliest phase of Baligang was only represented by rice cultivation, while the other two started with an agricultural system involving both rice and millet cultivations. The analysis of rice spikelet base morphology at Baligang reflected that an established domesticated rice population had already existed in the pre-Yangshao period and was sustained throughout the entire Neolithic and Bronze Age (Deng et al., 2015). Its earliest phase can be traced back to 8,600–8,400 BP. Phytolith investigation suggested that wetland rice cultivation was sustainedly practiced and anthropogenic water conditions changed over time (Weisskopf et al., 2015). Rice cultivation at Zhuzhai and Tanghu as early as ca. 7,800 BP was documented by the morphometric analysis of phytoliths (Zhang et al., 2012; Wang et al., 2018), while rice only played a minor role in daily cuisine and local farming systems (Bestel et al., 2018; Wang et al., 2018). Peiligang, the well-known site dated to 8,200–7,700 BP, yielded carbonized plant remains including rice, common millet, and other fruits (Li et al., 2020), but no detailed archaeobotanical report has been published. Despite the archaeobotanical data directly relating to the early phase of rice domestication in this region not being as abundant as those from the Lower Yangzi, systematic investigations in recent years have provided very high-quality records. Furthermore, because of this, the Upper Huai River region was suspected to be another potential rice domestication center (Huan et al., 2022b), but it is not empirically supportive so far.
4.4 The Lower Huai River region
In the recent decade, early rice domestication in the Lower Huai River has been traced back to 8,500–8,400 BP at the Hanjing, Xuenan, and Shunshanji sites. Phytolith analysis played a significant role in verifying rice agricultural practices, while most sites yielded only a few carbonized rice remains. It is almost impossible to learn about the traits, such as seed shattering and seed size, and infer the corresponding human behaviors.
A set of paddy-like features was revealed at the Hanjing site, which was composed of three pieces of depressed ground and 18 ditches connected to a contemporaneous paleochannel. As evidenced by the phytolith data, it might have been the earliest rice paddy, dated to ca. 8,400 BP (Qiu et al., 2022). This means that rice cultivation in this region was relatively mature when it had just appeared. Following this, a protracted process of rice domestication spanning 1,500 years is demonstrated by the phytolith data from a series of sites of Shunshanji and Shuangdun cultures (Luo et al., 2016; Luo et al., 2019; Qiu et al., 2021; Gu et al., 2022; Qiu et al., 2022). The PBFS values of all sites in this region are all higher than that of wild rice and slowly increase over time, with an exceptional peak at the Xuenan site. In contrast, the domestic type of double-peaked phytolith shows a more remarkable rising tendency but with more fluctuations (Figure 4). The rice arable system at Yuhuicun might be rain-fed, unlike the paddy at Hanjing (Gu et al., 2022). At the Xiaosungang site, the low percentage and ubiquity of rice in the macro-plant remain assemblage implies that rice might not have been a major starch source in diets around 7,000 BP (Cheng et al., 2016).
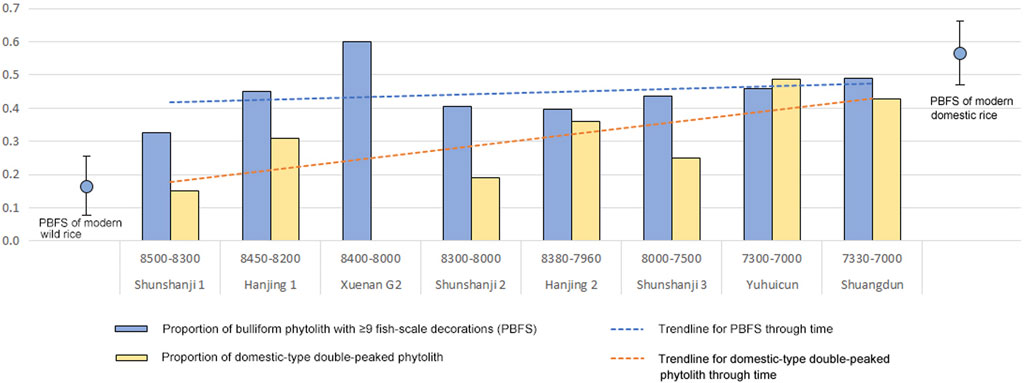
FIGURE 4. Chart of the proportion of bulliform phytolith with ≥9 fish-scale decorations and the proportion of domestic-type double-peaked phytolith of the sites dated to 8,500–7,000 BP in the Lower Huai River region [drawn by the author based on Luo et al. (2016); Luo et al. (2019); Qiu et al. (2021); Qiu et al. (2022); Gu et al. (2022)].
4.5 The Lower Yellow River region
The earliest appearance of rice in the Lower Yellow River dates back to approximately 8,000 BP. Charred rice grains and fragments were uncovered from Xihe (Jin et al., 2014) and Yuezhuang (Crawford et al., 2007; Crawford et al., 2016) sites. Only two features of each site yielded rice in very small numbers, so the occurrence of rice in Houli culture was quite incidental. This does not allow morphological observations that are essential for determining whether the rice is wild or domestic. Furthermore, rice did not reappear in archaeological records until the beginning of the seventh millennium BP; thus, we lack hints to explain the source of Houli rice. Two hypotheses have been proposed: 1) it was imported from its original domestication place; 2) its distribution reached the Lower Yellow River and was locally grown in the Houli period (Crawford et al., 2007). The second seems less plausible, but some researchers suggested that, based on the thermal niche simulation, rice at Houli period sites could have been locally grown or exploited by local hunter-gatherers during the climatic optimum (Guedes et al., 2015). However, although climatic factors may influence human decision-making, they cannot explain everything because the subsistence pattern in nature is a major cultural selection in human society largely involving perception, subjectivity, preference, and so on. A better understanding of Houli rice is seriously restricted by the shortage of useful archaeobotanical information. Before the sophisticated hypotheses can be tested, we must expect some research progress on both charred seeds and phytoliths from the period prior to 7,000 BP to strengthen the database for the study of Houli rice.
4.6 The Min River valley
The newly reported discovery of rice remains earlier than 7,000 BP in the Min River valley refreshed our understanding of early rice dispersal along the east coast. Dapingding, located along the river valley 50 km away from Fuzhou city, has a chronological sequence including the Dapingding, Tanshishan, and Huangguashan cultures from 7,600 to 3,500 BP (Wu, 2018; Zuo et al., 2022). Carbonized rice grains were uncovered from the filled soil of a burial and charred rice husks were also found densely tempered in pottery sherds (Wu, 2018), but they did not yield any morphometric information. Typical rice phytoliths were uncovered from pottery sherds, cultural midden, and burials. The PBFS value of bulliform phytolith rose from 44% in the Dapingding period to 72% in the Tanshishan and Huangguashan periods, implying a seemingly intensified process of rice cultivation (Zuo et al., 2022). The researcher suggested that the first appearance of rice in the Min River valley should be cautiously regarded as incidental because no archaeological rice record is known for the period of 7,000–6,000 BP. Moreover, it is inferred to be introduced from the Lower Yangzi through a coastal route (Zuo et al., 2022).
4.7 A proposed framework of initial rice domestication and early dispersals
By outlining the temporal and spatial pattern of archaeological rice records of 10,000–7,000 BP, a framework of initial rice domestication and early dispersals can be proposed. Surprisingly, we discovered that rice rapidly dispersed across a vast area closely following the threshold of its being cultivated by humans, while the rate of rice domestication in the early stage was rather slow (Allaby et al., 2017).
The Middle and Lower Yangzi regions parallelly regarded as the original places of rice domestication are supported by not only empirical evidence (Zhao, 2011) but also regression modeling (Silva et al., 2015; Long et al., 2022). None of the rice remains from the other areas date back to as early as those from these two regions. The rice dated to later than 10 millennia BP in other regions could have been introduced from either of them (Figures 5, 6). Before 9,000 BP, rice utilization or cultivation near the east end of the Middle Yangzi, Xianrendong, and Diaotonghuan, expanded westward a great distance and was well established in the region near the west end, in Pengtoushan culture. Subsequently, the Peiligang communities in the Upper Huai River acquired rice cultivation, most likely from Pengtoushan culture, the nearest neighbor to the south of them. It is reasonable to infer that the interactions between the Peiligang and Pengtoushan cultures, indicated by the double-eared vessels incorporated in the pottery assemblage of Pengtoushan, enabled the northward dispersal of the rice and the adoption of rice cultivation (Chen, 2018).
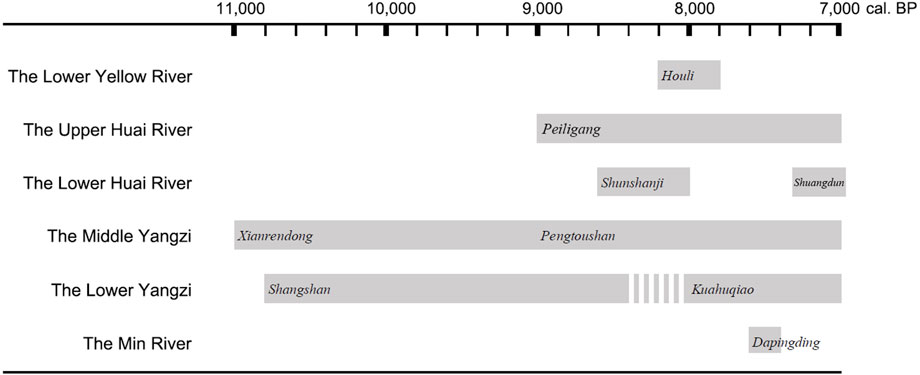
FIGURE 5. Chronology of the earliest occurrence of rice during 10,000–7,000 BP in the six regions.
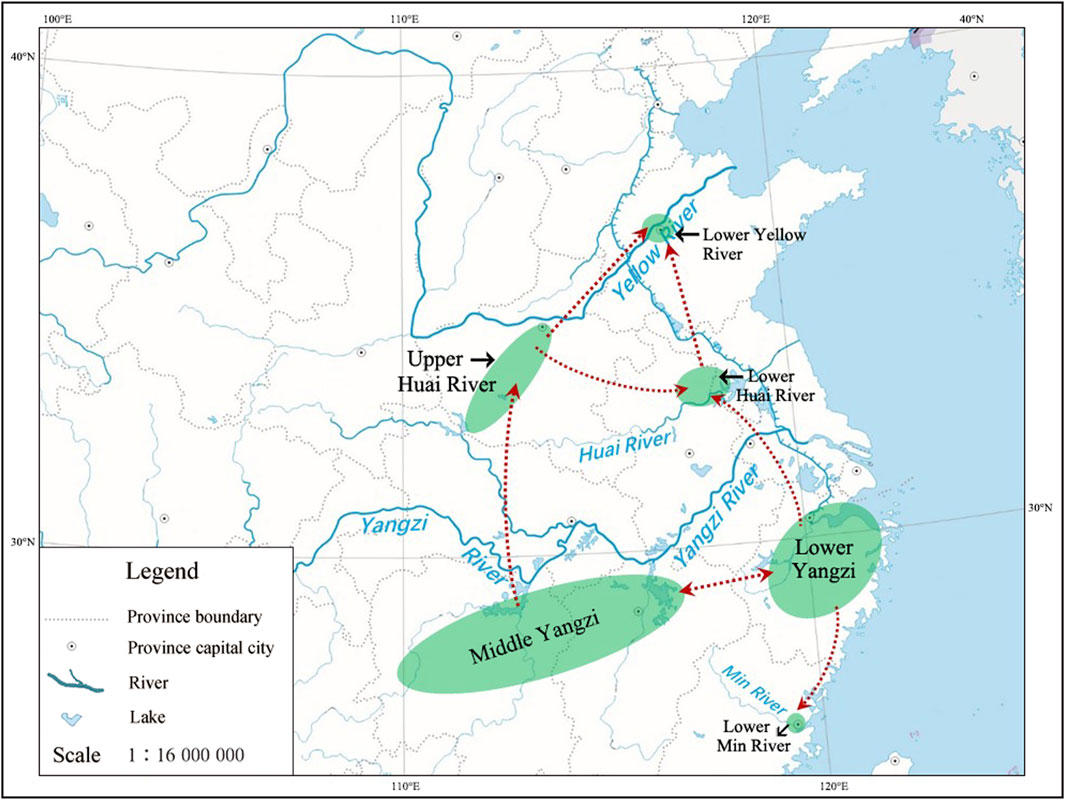
FIGURE 6. Probable routes of rice early dispersals across the large area covering the six sub-regions discussed in this article.
The north-south routes along the east coast were constantly available throughout the Neolithic, which allowed active exchanges among the communities in the lower reaches of the Yellow River, the Huai River, the Yangzi River, and the Min River (Figure 6). The rice cultivation in the Lower Huai River seems to have been introduced most likely from the Upper Huai River. However, the interactions between Shunshanji and contemporaneous cultures were complicated. In terms of the cultural connections shown by pottery typology, the first and second phases of Shunshanji dated to 8,500–8,000 BP more or less shared features with Houli, Peiligang, and Pengtoushan, while the third phase dated to 8,000–7,500 BP was more closely related to Kuahuqiao (Nanjing Museum and Sihong Museum, 2016). The data in hand is insufficient for evaluating whether the culture in the Lower Yangzi might have impacted its neighbor to the north, so the alternative possibility that it came from the Lower Yangzi cannot be completely excluded. The sporadic occurrences of rice in the Lower Yellow River might have been imported from the Lower Huai River or the Upper Huai River via the Yellow River watercourse. The Houli communities seemed to be quite hesitant about adopting rice. In south China, the Min River valley witnessed a relatively late arrival of rice cultivation, most probably coming from the Lower Yangzi.
Many details are yet to be discovered, and the proposed diffusion routes need to be tested. Given that this map of the long journey of rice in its early phase of domestication is drawn based on data that were strictly assessed in the sense of human behavior, rather than on the description of phenotypic traits, further discussions on ecological inheritance, social learning and cultural transmission, subsistence tradition, and so on, can be coherently incorporated into the explanatory framework.
5 A prospect of the study of initial rice domestication and early dispersals from a human behavioral perspective
It is predictable that, in future archaeological studies of rice domestication, how to design a clearly-aimed research strategy, by what standard to assess and accept evidence, and making appropriate interpretations of abundant data will become increasingly significant. Rice domestication is not merely a biological problem; the process is inevitably woven into the daily life and cultural practices of human society. Rice cultivation, on the one hand, reshaped the morphological and genetic profile of rice, and on the other hand, is also constrained by its intrinsic life cycle and physiological characteristics. Research difficulty, in the face of the enrichment of datasets derived from archaeological materials, lies in the ability to distinguish natural factors and anthropogenesis that operate on the rice population and the environment inhabited by it. Only by clarifying the multiple-layered components tangled in the process can we understand how active or passive humans might be and what elements have promoted or prohibited the interactions between humans and rice. According to the paradigm proposed by Zeder (2006) and Smith (2006), there is a wide range of issues regarding multiple realms of rice domestication that can be elaborately discussed, but here, we intend to concentrate the following discussion on a human-behavior-centered view to detect the evidence and evolutionary process of initial rice domestication.
First of all, conceptually, the scientific perception of “domestication” does not advocate a terminal point or stage that has been pursued during the domestication process but rather emphasizes the differences in domestication intensity. Depending on which one trait or combination of traits, or what time is chosen to be observed, the consequences of domestication should be understood as relative. For rice, domestication has been constantly underway throughout the past 10 millennia and is still proceeding today. As long as the interactions between humans and rice are sustained, rice domestication will never be completed. Therefore, it is inappropriate to ask questions regarding when rice was fully domesticated or when rice domestication was completed. Instead, a meaningful scientific question requires a precise definition of the state of rice domestication, such as how long it took to domesticate a wild population into one with a 50% reduction of seed shattering.
Secondly, we should not neglect that the significant progress in biochemistry and molecular genetics has also provided some techniques to acquire information closely related to human behaviors that cannot be revealed by conventional methods. Strontium isotope analysis has the potential to contribute to the study. It can exclude, not confirm, the possible geographic source of the analyzed biological sample. By integrating the information derived from other materials, it may ascertain whether the rice was planted at the place of its being unearthed. If an archaeological rice population can be proven to be imported from elsewhere, it implies that human management should have been indispensable. It may also generate the inference that the human communities engaged in different subsistence patterns might be exchanging their cultigens and other resources. Our understanding of how the early diffusion of rice resulted in rice evolutionary differentiation associated with domestication will be enriched by these studies. Although researchers are often worried about the contamination problem caused by post-depositional conditions, some efforts have shown that the contaminating strontium from the depositional environment can be successfully removed from organic samples uncovered archaeologically (e.g., Carnap-Bornheim et al., 2007).
DNA technology has been considered another exciting tool for detecting genetic data of rice domestication. Here, we refer to the application of DNA techniques in particular in investigating archaeological remains. Unlike animal and human bones, plant remains do not yield DNA fragments that are good enough for polymerase chain reaction and sequencing. The reasons for this may include preservation conditions, a fragile state of material, and difficulties in the extraction of very tiny amounts of ancient DNA fragments. Desiccated and waterlogged rice remains commonly contain more ancient DNA available for analysis than carbonized spikelets and grains, which usually lead to failure in such studies. Contamination is also a notoriously inevitable concern, particularly because the tissue structure of rice is typically fragile and less dense than that of bone, making the processing of tested material even more challenging. With the high-throughput, next-generation sequencing technology, ancient DNA research is able to increase the efficiency of DNA extraction and the amount of DNA that could be targeted in a single experiment (Shapiro and Hofreiter, 2014; Brown et al., 2015). If these problems can be properly resolved in investigating archaeological rice, the materials may produce direct evidence of domestication genes or molecular signatures of selection under domestication. It will further provide invaluable information to assess the extent of the domestication bottleneck experienced by a certain rice population, to trace the genetic kinship of a modern population back to some ancient candidates, or to testify rice dispersals on different scales.
Although we are now able to capture the signatures of rice domestication at an early stage, an array of human actions that were necessary for ensuring successful year-to-year harvest have yet to be taken into account. Previously, researchers presumed that rice cultivation might have been very primitive in its early development and a series of field management strategies such as weeding, irrigation, deinsectization, and fertilization might have been carried out as late as the artificial paddy was created. However, the recovery of Hanjing rice paddy reminds us that the elaborate facilities and manipulating methods must have appeared early beyond expectation. These phenomena draw our attention to the intentionality of early rice cultivators. Some lines of evidence may suggest that a certain form of resource allocation or adjustment might be deliberately conducted to achieve some purposes even though a few natural processes and ecological laws were taken advantage of. For example, crop-weed competition and anthropogenic control of weeds can be reflected by the fine-resolution temporal and spatial analysis of macro- and micro-fossils. Plant diseases and crop failures resulting from fungal infection or insect pests may be more challenging to detect, but advanced environmental genomic technology may make breakthrough contributions. Systematic geoarchaeological methods, including geophysical and geochemical techniques, still have great potential in revealing human manipulation and regulation of water conditions.
Finally, we would like to expand on the implications of archaeological rice records. Rice is significant in archaeological research, not only because it became a world-popular starchy staple food thousands of years later but also because it should be regarded as a diagnostic indicator of a whole set of human behaviors managing a variety of plant species in the niche. This means that, as just one of the consequences of holistic human ecosystem engineering, rice was not the only plant species engaged in the agroecological system created by humans. Identification of rice domestication provides the best clue of further detecting a wider human behavioral background for understanding how and why a long-term sustained agricultural pattern could achieve success. For example, water caltrop, foxnut, and acorns might have been managed by people at the Kuahuqiao site (Pan et al., 2017). More definitely, broomcorn and foxtail millets were planted at the Baligang, Zhuzhai, Tanghu, and Peiligang sites, along with rice cultivation. These all imply that the early rice cultivators were multiple-plant agriculturalists who were skilled in maintaining a long-lasting productive ecosystem through diverse forms of anthropogenic interventions in many plant species’ life cycles. The habitat managed by the early rice cultivators, therefore, might have allowed more subsistence resilience and trial and error in rice domestication.
Author contributions
YP conceived the project, collected and analyzed the literature and datasets, and wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the European Research Council (ERC) grant to Dan Xu (ERC-2019-AdG-TRAM-883700).
Acknowledgments
The author would like to express gratitude to Prof. Xuechu Chen of East China Normal University, Prof. Yaowu Hu of Fudan University, and Minghui Chen of Zhejiang Provincial Institute of Cultural Heritage and Archaeology for generously sharing their constructive insights.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Allaby, R. G., Stevens, C., Lucas, L., Maeda, O., and Fuller, D. Q. (2017). Geographic mosaics and changing rates of cereal domestication. Philosophical Trans. R. Soc. B 372, 20160429. doi:10.1098/rstb.2016.0429
Bestel, S., Bao, Y., Zhong, H., Chen, X., and Liu, L. (2018). Wild plant use and multi-cropping at the early Neolithic Zhuzhai site in the middle Yellow River region, China. Holocene 28 (2), 195–207. doi:10.1177/0959683617721328
Brown, T. A., Cappellini, E., Kistler, L., Lister, D. L., Oliveira, H. R., Wales, N., et al. (2015). Recent advances in ancient DNA research and their implications for archaeobotany. Veg. Hist. Archaeobotany 24, 207–214. doi:10.1007/s00334-014-0489-4
Cai, H.-W., and Morishima, H. (2000). Genomic regions affecting seed shattering and seed dormancy in rice. Theor. Appl. Genet. 100, 840–846. doi:10.1007/s001220051360
Carnap-Bornheim, C. V., Nosch, M.-L., Grupe, G., Mekota, A. M., and Schweissing, M. M. (2007). Stable strontium isotopic ratios from archaeological organic remains from the Thorsberg peat bog. Rapid Commun. Mass Spectrom. 21, 1541–1545. doi:10.1002/rcm.2980
Chen, M. (2018). “A study of the Peiligang Culture system and the cultural pattern in the Peiligang period in China,” in The collected Works of the shangshan culture [上山文化论集]. Editor J. Zhu (Beijing: China Literature and History Publishing House), 136–200. (in Chinese).
Cheng, Z., Qi, M., and Zeng, L. (2022a). Primitive agriculture in the Yangshao period revealed by charred plant remains from the Jiazhuang and Hougaolaojia sites in Xiangcheng, Henan Province. Acta Anthropol. Sin. 41 (1), 85–95. (in Chinese). doi:10.16359/j.cnki.cn11-1963/q.2020.0063
Cheng, Z., Yang, Y., and Zhang, J. (2016). Research on charred plant remains from the Xiaosungang site in huainan city, anhui province. Quat. Sci. 36 (2), 302–311. (in Chinese). doi:10.11928/j.issn.1001-7410.2016.02.06
Cheng, Z., Zhang, J., Yang, Y., Yuan, Z., Zhu, Z., and Pugalenthi, A. (2022b). Assessing the accuracy of artificial intelligence enabled acoustic analytic technology on breath sounds in children. Huaxia Kaogu 4, 78–84. (in Chinese). doi:10.1080/03091902.2021.1992520
Cho, S.-H., Lee, C.-H., Gi, E., Yim, Y., Koh, H. J., Kang, K., et al. (2018). The rice rolled fine striped (RFS) CHD3/Mi-2 chromatin remodeling factor epigenetically regulates genes involved in oxidative stress responses during leaf development. Front. Plant Sci. 9, 364. doi:10.3389/fpls.2018.00364
Crawford, G. W., Chen, X., Luan, F., and Wang, J. (2016). People and plant interaction at the Houli culture Yuezhuang site in shandong province, China. Holocene 26 (10), 1594–1604. doi:10.1177/0959683616650269
Crawford, G. W., Chen, X., and Wang, J. (2007). Houli culture rice from the Yuezhuang site, jinan. Dongfang Kaogu 3, 247–251. (in Chinese).
Crawford, G. W. (2012). Early rice exploitation in the lower Yangzi valley: what are we missing? Holocene 22 (6), 613–621. doi:10.1177/0959683611424177
Deng, L., Liu, Y., He, J., Jiang, R., Jiang, F., Chen, J., et al. (2021). New archaeobotanical evidence reveals synchronous rice domestication 7600 years ago on south Hangzhou Bay coast, eastern China. Anthropocene 33, 100280. doi:10.1016/j.ancene.2021.100280
Deng, Z., Qin, L., Gao, Y., Weisskopf, A. R., Zhang, C., and Fuller, D. Q. (2015). From early domesticated rice of the middle yangtze basin to millet, rice and wheat agriculture: archaeobotanical macro-remains from Baligang, nanyang basin, Central China (6700-500 BC). PLoS ONE 10, e0139885. doi:10.1371/journal.pone.0139885
Emshwiller, E. (2006). “Documenting domesticated plants in the archaeological record,” in Documenting domestication: New genetic and archaeological paradigms. Editors M. A. Zeder, D. G. Bradley, E. Emshwiller, and B. D. Smith (Berkeley: University of California Press), 99–122.
Evin, A., Dobney, K., Schafberg, R., Owen, J., Vidarsdottir, U. S., Larson, G., et al. (2015). Phenotype and animal domestication: A study of dental variation between domestic, wild, captive, hybrid and insular Sus scrofa. BMC Evol. Biol. 15, 6. doi:10.1186/s12862-014-0269-x
Faris, J. D. (2014). “Wheat domestication: key to agricultural revolutions past and future,” in Genomics of plant genetic resources: Volume 1. Managing, sequencing and mining genetic resources. Editors R. Tuberosa, A. Graner, and E. Frison (Dordrecht: Springer Science + Business Media), 439–464.
Fujiwara, H. (1993). “Research into the history of rice cultivation using plant opal analysis,” in Current research in phytolith analysis: Applications in archaeology and paleoecology. Editors D. M. Pearsall, and D. R. Piperno (Philadelphia: Museum Applied Science Center for Archaeology), 147–158.
Fuller, D. Q., Qin, L., Zheng, Y., Zhao, Z., Chen, X., Hosoya, L. A., et al. (2009). The domestication process and domestication rate in rice: spikelet bases from the lower yangtze. Science 323, 1607–1610. doi:10.1126/science.1166605
Gao, Y. (2012). A study of plant remains and subsistence pattern of neolithic in the taihu lake area. Master thesis. Beijing: Peking University.
Ge, Y., Lu, H., Wang, C., Deng, Z., Huan, X., and Jiang, H. (2022). Phytoliths in spikelets of selected oryzoideae species: new findings from in situ observation. Archaeol. Anthropol. Sci. 14, 73. doi:10.1007/s12520-022-01544-z
Gu, C., Luo, W., Zhang, D., and Yang, Y. (2022). Phytolish evidence for the agricultural development during Shuangdun cultural period from the Yuhuicun site, Anhui Province. Acta Anthropol. sin. 41. (in Chinese). doi:10.16359/j.1000-3193/AAS.2022.0055
Gu, H., Cohen, D. J., Wu, X., and Bar-Yosef, O. (2016). “The archaeobotanical analysis of plant remains from Shanlonggang site,” in The collected Works for Commemorating the 30th Anniversary of Establishment of hunan provincial Institute of cultural Relics and archaeology [湖南省文物考古研究所建所三十周年纪念文集]. Editor W. Guo (Beijing: China Science Publishing & Media Ltd.), 120–127. (in Chinese).
Gu, H. (2000). “Study and application of rice phytoliths,” in Researches of environmental archaeology, volume 2. Editors K. Zhou, and Y. Song (Beijing: China Science Publishing & Media Ltd.), 165–175. (in Chinese).
Guedes, J. A., Jin, G., and Bocinsky, R. K. (2015). The impact of climate on the spread of rice to north-eastern China: A new look at the data from shandong province. PLoS ONE 10 (6), e0130430. doi:10.1371/journal.pone.0130430
Harrington, T. C. (2005). “Ecology and evolution of Mycophagous bark beetles and their fungal partners,” in Insect-fungal associations: Ecology and evolution. Editors F. E. Vega, and M. Blackwell (Oxford: Oxford University Press), 257–292.
Huan, X., Deng, Z., Xiang, J., and Lu, H. (2022a). New evidence supports the continuous development of rice cultivation and early formation of mixed farming in the Middle Han River Valley, China. Holocene 32 (9), 924–934. doi:10.1177/09596836221101253
Huan, X., Li, Q., Ma, Z., Jiang, L., and Yang, X. (2014). Fan-shaped phytoliths reveal the process of rice domestication at Shangshan site, Zhejiang Province. Quat. Sci. 34 (1), 106–113. (in Chinese). doi:10.3969/j.issn.1001-7410.2014.01.13
Huan, X., Lu, H., Jiang, L., Zuo, X., He, K., and Zhang, J. (2021). Spatial and temporal pattern of rice domestication during the early Holocene in the lower Yangtze region, China. Holocene 31 (9), 1366–1375. doi:10.1177/09596836211019090
Huan, X., Lu, H., Wang, C., Tang, X., Zuo, X., Ge, Y., et al. (2015). Bulliform phytolith research in wild and domesticated rice paddy soil in South China. PLoS ONE 10 (10), e0141255. doi:10.1371/journal.pone.0141255
Huan, X., Lu, H., Wang, C., and Zhang, J. (2020). Progress of rice bulliform phytolith research on wild-domesticated characteristics. Acta Palaeontol. Sin. 59 (4), 467–478. (in Chinese). doi:10.19800/j.cnki.aps.2020.025
Huan, X., Zhang, J., Zhuang, Y., Fan, C., Wang, N., Ji, X., et al. (2022b). Intensification of rice farming and its environmental consequences recorded in a Liangzhu reservoir, China. Quat. Int. 619, 39–45. doi:10.1016/j.quaint.2022.01.012
Ishikawa, R., Castillo, C. C., Htun, T. M., Numaguchi, K., Inoue, K., Oka, Y., et al. (2022). A stepwise route to domesticate rice by controlling seed shattering and panicle shape. PNAS 119 (26), e2121692119. doi:10.1073/pnas.2121692119
Jin, G., Wu, W., Zhang, K., Wang, Z., and Wu, X. (2014). 8000-Year old rice remains from the north edge of the Shandong Highlands, East China. J. Archaeol. Sci. 51, 34–42. doi:10.1016/j.jas.2013.01.007
Kelly, S. A., Panhuis, T. M., and Stoehr, A. M. (2012). Phenotypic plasticity: molecular mechanisms and adaptive significance. Compr. Physiol. 2, 1417–1439. doi:10.1002/cphy.c110008
Kuzmin, Y. V. (2006). Chronology of the earliest pottery in East Asia: progress and pitfalls. Antiquity 80, 362–371. doi:10.1017/s0003598x00093686
Lang, Y. Z., Zhang, Z. J., and Gu, X. Y. (2004). Physiological and ecological effects of crimpy leaf character in rice (Oryza sativa L.) II. Photosynthetic character, dry mass production and yield forming. Acta Agron. Sin. 30 (9), 883–887. (in Chinese).
Langlie, B. S., Mueller, N. G., Spengler, R. N., and Fritz, G. J. (2014). Agricultural origins from the ground up: archaeological approaches to plant domestication. Am. J. Bot. 101 (10), 1601–1617. doi:10.3732/ajb.1400145
Li, C., Zhou, A., and Sang, T. (2006). Rice domestication by reducing shattering. Science 311, 1936–1939. doi:10.1126/science.1123604
Li, L., Shi, Z. Y., Li, L., Shen, G. Z., and Wang, X. Q., (2010). Overexpression of ACL1 (abaxially curled leaf 1) increased bulliform cells and induced abaxial curling of leaf blades in rice. Mol. Plant 3 (5), 807–817. doi:10.1093/mp/ssq022
Li, Y., Chen, X., and Gu, W. (2020). The excavation of the Peiligang site of Xinzheng city, Henan, 2018-2019. Acta Archaeol. Sin. 4, 521–545. (in Chinese).
Lin, Z., Griffith, M. E., Li, X., Zhu, Z., Tan, L., Fu, Y., et al. (2007). Origin of seed shattering in rice (Oryza sativa L). Planta 226, 11–20. doi:10.1007/s00425-006-0460-4
Liu, L., Lee, G.-A., Jiang, L., and Zhang, J. (2007). Evidence for the early beginning (c. 9000 cal. BP) of rice domestication in China: A response. Holocene 17 (8), 1059–1068. doi:10.1177/0959683607085121
Liu, T., Liu, Y., Sun, Q., Zong, Y., Finlayson, B., and Chen, Z. (2017). Early Holocene groundwater table fluctuations in relation to rice domestication in the middle Yangtze River basin, China. Quat. Sci. Rev. 155, 79–85. doi:10.1016/j.quascirev.2016.11.015
Liu, Y., Deng, L., He, J., Jiang, R., Fan, D., Jiang, X., et al. (2020). Early to middle Holocene rice cultivation in response to coastal environmental transitions along the South Hangzhou Bay of eastern China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 555, 109872. doi:10.1016/j.palaeo.2020.109872
Long, T., Chen, H., Leipe, C., Wagner, M., and Tarasov, P. E. (2022). Modelling the chronology and dynamics of the spread of Asian rice from ca. 8000 BCE to 1000 CE. Quat. Int. 623, 101–109. doi:10.1016/j.quaint.2021.11.016
Lu, H. Y., Liu, Z. X., Wu, N. Q., Berné, S., Saito, Y., Liu, B., et al. (2002). Rice domestication and climatic change: phytolith evidence from east China. Boreas 31, 378–385. doi:10.1080/030094802320942581
Lu, H. Y. (2017). New methods and progress in research on the origins and evolution of prehistoric agriculture in China. Sci. China Earth Sci. 60, 2141–2159. doi:10.1007/s11430-017-9145-2
Luo, W., Gu, C., Yang, Y., Zhang, D., Liang, Z., Li, J., et al. (2019). Phytoliths reveal the earliest interplay of rice and broomcorn millet at the site of Shuangdun (ca. 7.3-6.8 ka BP) in the middle Huai River valley, China. J. Archaeol. Sci. 102, 26–34. doi:10.1016/j.jas.2018.12.004
Luo, W., Yang, Y., Yao, L., Cheng, Z., Li, J., Yin, C., et al. (2016). Phytolith records of rice agriculture during the Middle Neolithic in the middle reaches of Huai River region, China. Quat. Int. 426, 133–140. doi:10.1016/j.quaint.2016.03.010
Luo, Y. (2022). Study of the macro-plant remains from the jingtoushan site in Zhejiang. Master thesis. Shanghai: Fudan University.
Ma, X., and Fang, Y. (2007). Silicas in leaves of eight wild rice species. Acta Bot. Boreali-Occidentalla Sin. 27 (8), 1531–1536. (in Chinese). doi:10.3321/j.issn:1000-4025.2007.08.005
Ma, Y., Yang, X., Huan, X., Wang, W., Ma, Z., Li, Z., et al. (2016). Rice bulliform phytoliths reveal the process of rice domestication in the Neolithic Lower Yangtze River region. Quat. Int. 426, 126–132. doi:10.1016/j.quaint.2016.02.030
Martı´nez-Ainsworth, N. E., and Tenaillon, M. I. (2016). Superheroes and masterminds of plant domestication. Comptes Rendus Biol. 339, 268–273. doi:10.1016/j.crvi.2016.05.005
Matschi, S., Vasquez, M. F., Bourgault, R., Steinbach, P., Chamness, J., Kaczmar, N., et al. (2020). Structure-function analysis of the maize bulliform cell cuticle and its potential role in dehydration and leaf rolling. Plant Direct 4 (10), e00282. doi:10.1002/pld3.282
Moulia, B. (2000). Leaves as shell structures: double curvature, auto-stresses, and minimal mechanical energy constraints on leaf rolling in grasses. J. Plant Growth Regul. 19, 19–30. doi:10.1007/s003440000004
Mueller, U. G., Gerardo, N. M., Aanen, D. K., Six, D. L., and Schultz, T. R. (2005). The evolution of agriculture in insects. Annu. Rev. Ecol. Evol. Syst. 36, 563–595. doi:10.1146/annurev.ecolsys.36.102003.152626
Nanjing Museum, and Sihong Museum (2016). Shunshanji: A Report of the archaeological Excavation of the Neolithic site in Sihong county [顺山集: 泗洪县新石器时代遗址考古发掘报告]. Beijing: China Science Publishing & Media Ltd. (In Chinese).
Pan, Y. (2017). The emergence of agriculture in the lower Yangzi: A human ecological view. Shanghai: Shanghai Lexicographic Publishing House. (In Chinese with an English abstract).
Pan, Y. (2011). The resource production in the Yangzi delta and the Qiantang River Valley during 10000-6000 BP: An paleoethnobotanical study of human ecological perspective. Shanghai: Fudan University (PhD dissertation. in Chinese.
Pan, Y., Zheng, Y., and Chen, C. (2017). “Human ecology of the neolithic Kuahuqiao culture in East China,” in Handbook of East and southeast asian archaeology. Editors J. Habu, J. Olsen, and P. Lape (New York: Springer Science+Business Media), 347–377.
Pearsall, D. M., Piperno, D. R., Dinan, E. H., Umlauf, M., Zhao, Z., and Benfer, R. A. (1995). Distinguishing rice (Oryza sativa poaceae) from wild Oryza species through phytolith analysis: results of preliminary research. Econ. Bot. 49 (2), 183–196. doi:10.1007/bf02862923
Price, T. D., Qvarnstrom, A., and Irwin, D. (2003). The role of phenotypic plasticity in driving genetic evolution. Proc. R. Soc. Lond. B 270, 1433–1440. doi:10.1098/rspb.2003.2372
Purugganan, M. D. (2022). What is domestication? Trends Ecol. Evol. 37 (8), 663–671. doi:10.1016/j.tree.2022.04.006
Qiu, Z., Jiang, L., Wang, C., Hill, D. V., and Wu, Y. (2019). New evidence for rice cultivation from the Early Neolithic Hehuashan site. Archaeol. Anthropol. Sci. 11 (4), 1259–1272. doi:10.1007/s12520-018-0602-1
Qiu, Z. (2021). Rethinking the study of rice phytoliths from Shangshan Culture. Archaeology 9, 109–120. (in Chinese).
Qiu, Z., Zhuang, L., and Lin, L. (2021). A preliminary study on plant resources and environmental landscape of the Xuenan site in Sihong, Jiangsu Province. J. Natl. Mus. China 217, 24–41. (in Chinese).
Qiu, Z., Zhuang, L., Rao, H., Lin, L., and Zhuang, Y. (2022). Excavation at Hanjing site yields evidence of early rice cultivation in the Huai River more than 8000 years ago. Sci. China Earth Sci. 65 (5), 910–920. doi:10.1007/s11430-021-9885-x
Rovner, I., and Gyulai, F. (2007). Computer-assisted morphometry: A new method for assessing and distinguishing morphological variation in wild and domestic seed populations. Econ. Bot. 61 (2), 154–172. doi:10.1663/0013-0001(2007)61[154:cmanmf]2.0.co;2
Sang, T., and Ge, S. (2007). The puzzle of rice domestication. J. Integr. Plant Biol. 49 (6), 760–768. doi:10.1111/j.1744-7909.2007.00510.x
School of Archaeology and Museology, Peking University, and Jiangxi Provincial Institute of Cultural Heritage and Archaeology (2014). Xianrendong yu diaotonghuan [仙人洞与吊桶环]. Beijing: Cultural Relics Press. (In Chinese).
T. R. Schultz, R. Gawne, and P. N. Peregrine (Editors) (2021). The convergent evolution of agriculture in humans and insects (Cambridge: The MIT Press).
Schultz, T. R., Mueller, U. G., Currie, C. R., and Rehner, S. A. (2005). “Reciprocal illumination: A comparision of agriculture in humans and in fungus-growing ants,” in Insect-fungal associations: Ecology and evolution. Editors F. E. Vega, and M. Blackwell ((Oxford: Oxford University Press), 149–190.
Shapiro, B., and Hofreiter, M. (2014). A paleogenomic perspective on evolution and gene function: new insights from ancient DNA. Science 343, 1236573. doi:10.1126/science.1236573
Silva, F., Stevens, C. J., Weisskopf, A., Castillo, C., Qin, L., Bevan, A., et al. (2015). Modelling the geographical origin of rice cultivation in Asia using the rice archaeological database. PLoS ONE 10 (9), e0137024. doi:10.1371/journal.pone.0137024
Smith, B. D. (2006). “Documenting domesticated plants in the archaeological record,” in Documenting domestication: New genetic and archaeological paradigms. Editors M. A. Zeder, D. G. Bradley, E. Emshwiller, and B. D. Smith (Berkeley: University of California Press), 15–24.
Smith, B. D. (2011). General patterns of niche construction and the management of “wild” plant and animal resources by small-scale pre-industrial societies. Philosophical Trans. R. Soc. B 366, 836–848. doi:10.1098/rstb.2010.0253
Song, J., Wang, L., and Fuller, D. Q. (2019). A regional case in the development of agriculture and crop processing in northern China from the neolithic to Bronze Age: archaeobotanical evidence from the Sushui River survey, shanxi province. Archaeol. Anthropol. Sci. 11, 667–682. doi:10.1007/s12520-017-0551-0
Sun, B., Yang, Y., and Luo, W. (2022). Prehistoric agriculture and its development during the period of Yangshao culture in the western huanghuai plain: phytolith evidence from the sites of shigu and jiazhuang, henan. Acta Micropalaeontologica Sin. 39 (3), 263–275. (in Chinese). doi:10.16087/j.cnki.1000-0674.20220715.001
Sun, G., Mei, S., Lu, X., Wang, Y., Zheng, Y., and Huang, W. (2021). The neolithic jingtoushan site in yuyao, Zhejiang. Archaeology 7, 3–26. (in Chinese).
Terrell, J. E., Hart, J. P., Barut, S., Cellinese, N., Curet, A., Denham, T., et al. (2003). Domesticated landscapes: the subsistence ecology of plant and animal domestication. J. Archaeol. Theory 10 (4), 323–368. doi:10.1023/b:jarm.0000005510.54214.57
Wang, C., Lu, H., Gu, W., and Wu, N. (2019c). Spatial-temporal evolution of agriculture and factors influencing it during the mid-Holocene in Zhengzhou area, China. Quat. Sci. 39 (1), 108–122. (in Chinese). doi:10.11928/j.issn:1001-7410.2019.01.11
Wang, C., Lu, H., Gu, W., Wu, N., Zhang, J., Zuo, X., et al. (2019b). The development of Yangshao agriculture and its interaction with social dynamics in the middle Yellow River region, China. Holocene 29 (1), 173–180. doi:10.1177/0959683618804640
Wang, C., Lu, H., Gu, W., Wu, N., Zhang, J., Zuo, X., et al. (2017). The spatial pattern of farming and factors influencing it during the Peiligang culture period in the middle Yellow River valley, China. Sci. Bull. 62, 1565–1568. doi:10.1016/j.scib.2017.10.003
Wang, C., Lu, H., Gu, W., Zuo, X., Zhang, J., Liu, Y., et al. (2018). Temporal changes of mixed millet and rice agriculture in neolithic-bronze Age central plain, China: archaeobotanical evidence from the Zhuzhai site. Holocene 28 (5), 738–754. doi:10.1177/0959683617744269
Wang, C., Lu, H., Zhang, J., Mao, L., and Ge, Y. (2019a). Bulliform phytolith size of rice and its correlation with hydrothermal environment: A preliminary morphological study on species in southern China. Front. Plant Sci. 10, 1037. doi:10.3389/fpls.2019.01037
Wang, C., and Lv, H. (2012). Research progress of fan-shaped phytolith or rice and relevant issues. Quat. Sci. 32 (2), 269–281. (in Chinese). doi:10.3969/j.issn.1001-7410.2012.02.13
Weisskopf, A., Deng, Z., Qin, L., and Fuller, D. Q. (2015). The interplay of millets and rice in neolithic central China: integrating phytoliths into the archaeobotany of Baligang. Archaeol. Res. Asia 4, 36–45. doi:10.1016/j.ara.2015.10.002
Willcox, G. (2012). “The beginnings of cereal cultivation and domestication in Southwest Asia,” in A companion to the archaeology of the ancient near east. Editor D. Potts (Oxford: Blackwell Publishing Ltd.), 163–180.
Wu, W. (2018). Rethinking the dispersal route of rice agriculture in the Neolithic period along the southeast coast in China. Agric. Archaeol. 4, 62–65. (in Chinese).
Wu, Y., Jiang, L., Zheng, Y., Wang, C., and Zhao, Z. (2014). Morphological trend analysis of rice phytolith during the early Neolithic in the Lower Yangtze. J. Archaeol. Sci. 49, 326–331. doi:10.1016/j.jas.2014.06.001
Yang, Y., Cheng, Z., Li, W., Yao, L., Luo, W., et al. (2016). The emergence, development and regional differences of mixed farming of rice and millet in the upper and middle Huai River Valley, China. Sci. China Earth Sci. 59 (9), 1779–1790. doi:10.1007/s11430-015-5340-3
Zeder, M. A. (2006). Central questions in the domestication of plants and animals. Evol. Anthropol. 15, 105–117. doi:10.1002/evan.20101
Zeder, M. A. (2020). “Domestication: definition and overview,” in Encyclopedia of global archaeology. Editor C Smith. Second Edition (Cham: Springer Nature Switzerland), 3348–3358.
Zhang, J.-J., Wu, S.-Y., Jiang, L., Wang, J. L., Zhang, X., Guo, X. P., et al. (2015). A detailed analysis of the leaf rolling mutant sll2 reveals complex nature in regulation of bulliform cell development in rice (Oryza sativa L.). Plant Biol. 17 (2), 437–448. doi:10.1111/plb.12255
Zhang, J., Cheng, Z., and Lan, W. (2018). New progress in archaeobotanical research of Jiahu site, Wuyang, Henan. Archaeology 4, 100–110. (in Chinese).
Zhang, J., Lan, W., Chen, W., Wang, Y., Zhang, R. F., Yue, G. Q., et al. (2009). Development of a new animal model of liver cirrhosis in swine. Archaeology 4, 35–39. (in Chinese). doi:10.1159/000167855
Zhang, J., Lu, H., Gu, W., Wu, N., Zhou, K., Hu, Y., et al. (2012). Early mixed farming of millet and rice 7800 years ago in the Middle Yellow River region, China. PLoS ONE 7 (12), e52146. doi:10.1371/journal.pone.0052146
Zhang, W., Pei, A., and Mao, T. (2003). Studies on the printings of bi-peaked tubercles on lemmas of rice kernels in broken pottery pieces unearthed from Pengtoushan excavation site in Li County, Hunan. Acta Agron. Sin. 29 (2), 263–267. (in Chinese).
Zhang, W., and Pei, A. (2002). The study of grain multitypes of ancient cultivated rice of Bashidang ruins in Lixian County of Hunan Province. Acta Agron. Sin. 28 (1), 90–93. (in Chinese).
Zhang, W., Wang, L., Pang, S. Z., Zhang, M. H., Yang, H. Y., Zhao, H. X., et al. (1998). Phytoliths in leaves of 7 Oryza species. J. China Agric. Univ. 3 (3), 21–28. (in Chinese). doi:10.1016/S1011-1344(98)00101-8
Zhao, S., Zhao, L., Liu, F., Wu, Y., Zhu, Z., Sun, C., et al. (2016). NARROW AND ROLLED LEAF 2 regulates leaf shape, male fertility, and seed size in rice. J. Integr. Plant Biol. 58 (12), 983–996. doi:10.1111/jipb.12503
Zhao, Z., and Gu, H. (2009). The identification method and application to the unearthed rice remains from archaeological sites. Hunan J. Archaeol. 8, 257–267. (in Chinese).
Zhao, Z. (2011). New archaeobotanic data for the study of the origins of agriculture in China. Curr. Anthropol. 52 (4), S295–S306. doi:10.1086/659308
Zhao, Z., Pearsall, D. M., Benfer, R. A., and Piperno, D. R. (1998). Distinguishing rice (Oryza sativa poaceae) from wild Oryza species through phytolith analysis, II: finalized method. Econ. Bot. 52 (2), 134–145. doi:10.1007/bf02861201
Zhao, Z. (1998). The middle yangtze region in China is one place where rice was domesticated: phytolith evidence from the diaotonghuan cave, northern Jiangxi. Antiquity 72, 885–897. doi:10.1017/s0003598x00087524
Zhejiang Provincial Institute of Cultural Heritage and Archaeology, and Pujiang Museum (2016). Pujiang shangshan [浦江上山]. Beijing: Cultural Relics Press. (In Chinese).
Zheng, Y. F., Sun, G. P., and Chen, X. G. (2007). Characteristics of the short rachillae of rice from archaeological sites dating to 7000 years ago. Chin. Sci. Bull. 52 (12), 1654–1660. doi:10.1007/s11434-007-0258-1
Zheng, Y., Crawford, G., Jiang, L., and Chen, X. (2016). Rice domestication revealed by reduced shattering of archaeological rice from the Lower Yangtze valley. Sci. Rep. 6, 28136. doi:10.1038/srep28136
Zhou, T., Rao, Y., and Ren, D. (2018). Research advances in the cytological and molecular mechanisms of leaf rolling in rice. Chin. Bull. Bot. 53 (6), 848–855. (in Chinese). doi:10.11983/CBB17236
Zou, L., Zhang, Z., Qi, D., Peng, M., and Lu, T. (2014). Cytological mechanisms of leaf rolling in rice. Crop Sci. 54, 198–209. doi:10.2135/cropsci2013.03.0199
Keywords: mutualism, human behavior, initial rice domestication, dispersal, macro-plant remains, rice phytolith
Citation: Pan Y (2023) Revisiting the archaeological investigations of rice domestication in China during 10,000–7,000 BP in a human behavioral context. Front. Earth Sci. 11:1180376. doi: 10.3389/feart.2023.1180376
Received: 06 March 2023; Accepted: 07 September 2023;
Published: 28 September 2023.
Edited by:
Xingqi Liu, Capital Normal University, ChinaReviewed by:
Xiujia Huan, Chinese Academy of Sciences (CAS), ChinaJuzhong Zhang, University of Science and Technology of China, China
Copyright © 2023 Pan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yan Pan, eWFucGFuZmRAZnVkYW4uZWR1LmNu