 Bian Wang1,2
Bian Wang1,2 Zhao-Qun Zhang1,2,3*
Zhao-Qun Zhang1,2,3* Yuan-Qing Wang1,2,3*
Yuan-Qing Wang1,2,3* Qian Li1,2
Qian Li1,2 Bin Bai1,2
Bin Bai1,2 Yan Liu1,2
Yan Liu1,2 Fang-Yuan Mao1,2Hai-Bing Wang1,2Jian Wang1,3
Fang-Yuan Mao1,2Hai-Bing Wang1,2Jian Wang1,3 Yan-Xin Gong1,3
Yan-Xin Gong1,3 Li-Ping Dong1,2
Li-Ping Dong1,2 Li-Hua Wang1,3Hai-Dan Ma1,3
Li-Hua Wang1,3Hai-Dan Ma1,3 Ran-Cheng Xu1,3Xiao-Yang Wang1,3
Ran-Cheng Xu1,3Xiao-Yang Wang1,3- 1Key Laboratory of Vertebrate Evolution and Human Origins, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences (CAS), Beijing, China
- 2CAS Center for Excellence in Life and Paleoenvironment, Beijing, China
- 3College of Earth and Planetary Sciences, University of Chinese Academy of Sciences, Beijing, China
For a hundred years the Saint Jacques area has been known to produce rich Oligocene vertebrate fossils, yet only a handful of previous studies have focused on this area. Since 2010, we have conducted 12 field expeditions to Saint Jacques, and here we report findings from our paleontological excavations and stratigraphical investigations. Twenty-two fossiliferous blocks across the area are recognized and a chronostratigraphic framework has been established to aid fossil collection. Fossil-mammal materials have been recovered in situ from 1635 localities and additionally from surface sediments. Fossiliferous blocks in the area are correlated by lithological similarity and lateral tracing. Lithologically, the area is mainly composed of reddish silty mudstone and muddy siltstone, with three distinctive layers of grayish white sandstone. The measured composite stratigraphic column spans 239 meters and are divided into 12 lithostratigraphic units. Contrary to previous knowledge that Saint Jacques contains two Oligocene mammalian assemblages, our preliminary biostratigraphic analysis of small mammals shows that the area documents successive faunal transition from the Eocene to possibly the early Miocene. The hyracodontid perissodactyl Ardynia, the ctenodactyloid rodent Gobiomys, and the basal Glires Gomphos from the bottom litho-units imply the presence of the Eocene–Oligocene boundary, while small mammal assemblage of the top units is similar to Miocene faunas in northern China and Mongolia. Thus, rock strata in Saint Jacques likely span the Eocene through the early Miocene, bracketing an entire Oligocene sequence within. In sum, our re-exploration of Saint Jacques has greatly expanded the chronostratigraphic and taxonomic coverage of the mammalian fossil collection from this area. This long, successive Oligocene sequence makes an important record for studying the Eocene–Oligocene Transition. Further study in this area will contribute to a range of paleontological and paleoenvironmental questions.
Introduction
The Oligocene epoch is a transitional time period marked by the most striking climate change in the Cenozoic from greenhouse to icehouse conditions, mainly evidenced by marine records (Zachos et al., 2001; Coxall et al., 2005; Eldrett et al., 2009; Westerhold et al., 2020) but less terrestrial records (Dupont-Nivet et al., 2007; Hren et al., 2013). This change in global climate is likely associated with the profound shifts observed in continental faunal and floral compositions (e.g., Zhang et al., 2012; Sun et al., 2014). The continental interior of Asia has been a focal region for studying faunal turnovers across the Eocene–Oligocene boundary (e.g., Meng and McKenna, 1998; Kraatz and Geisler, 2010). To better understand the response of land mammals to significant global and regional climatic changes, long sequences of terrestrial sediments with successive fossil records and precise stratigraphic calibration are pivotal.
Saint Jacques is one of the classic Oligocene fossil sites of Asia. Sitting on the right bank of the Yellow River near the town of Balagong, Hanggin Banner, Ordos City, Nei Mongol Autonomous Region, China, the namesake of the fossiliferous area is in fact a small town across the Yellow River in Dengkou County, Bayannur City (Figures 1A,B). Although known for its rich vertebrate fossils for nearly a century, detailed accounts on the lithology of the Saint Jacques area are still lacking, hindering finer divisions of the depositional sequence and fossil assemblages.

FIGURE 1. Location of Saint Jacques and its fossil localities. (A) Map of China, showing the location of Saint Jacques (red circle) in Nei Mongol (gray area). (B) The Saint Jacques fossiliferous area is near the town of Balagong, across the Yellow River from Dengkou County. Fossil localities are densely distributed in (C) Sanshenggongbei and (D) Langhaogou. White lines in (C) and (D): measured stratigraphic sections. Red lines: faults.
Geological setting
Located in the northwestern margin of the Ordos block, the study area is bounded by the Jilantai Basin to the west, Hetao Basin to the north, and Yinchuan Basin to the south. Cenozoic strata are well exposed in the gullies along the Yellow River, from Balagong area in the north to Qianlishan area in the south (Wang, 1987). Our preliminary exploration in the region finds no exposure of Cenozoic strata superimposed with Mesozoic strata. In the Qianlishan area, the Cenozoic strata overlie the Cambrian strata by faults. The earliest Cenozoic strata in this area is possibly Early–Middle Eocene, as evidenced by our findings of Gomphos fossil from Saint Jacques. Late Eocene to Oligocene strata are superimposed by Neogene strata. Quaternary terrace sediments of the Yellow River cover most of the highland. Major faults in the Saint Jacques area cut through the Eocene, Oligocene, and Neogene strata, revealing that tectonic movements were active during late Cenozoic period, as studied by Shi et al. (2020) in their review of the neotectonics around the Ordos block.
History of research
Vertebrate fossils were first discovered in the Saint Jacques area by French paleontologists Pierre Teilhard de Chardin and Emile Licent while exploring along the Yellow River in 1923 (Teilhard de Chardin and Licent, 1924a, 1924b). They initially considered the site Pliocene in age, but soon revised it to the Oligocene based on the occurrence of Paraceratherium (Teilhard de Chardin and Licent, 1924c). Later, the mammalian fossils from Saint Jacques were reported in detail by Teilhard de Chardin, (1926). In the decades to follow, this area was rarely investigated except for a short visit by the Sino-Soviet Paleontological Expedition in 1959 (Chow and Rozhdestvensky, 1960). In 1977 and 1978, more extensive explorations by the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) were carried out, revealing a greater exposure of fossiliferous area than previously recognized (Wang, 1987). However, further work on its geology was hampered by its complicated structure. Wang (1987) noted that “it [was] difficult to define a formal lithostratigraphic unit based on beds exposed in Saint-Jacques area” (p.45). Based on similarity in lithology and assemblage of mammal fossils, the beds in Saint Jacques were thought to be equivalent to the lower member of the Wulanbulage Formation in Qianlishan district (Wang, 1987). Further investigation on the mammal fossil have led subsequent authors to recognize two local faunas from different horizons at Saint Jacques, one from early Oligocene and the other possibly late Oligocene (Wang, 1987; Wang and Emry, 1991; Wang and Qiu, 2003). The faunas in Saint Jacques are primarily composed of small mammals, especially ctenodactyloid rodents, although several species of ungulates and carnivores are also present (Meng and McKenna, 1998; Wang and Qiu, 2003).
For over a decade now, a new team of IVPP researchers have been conducting field explorations back in the Saint Jacques area and have collected abundant fossil-mammal materials from different blocks and stratigraphic levels. New specimens of the carnivore Palaeogale sectoria, the hyracodontid Ardynia, and the lagomorph Ordolagus have been recently reported (Wang and Zhang, 2015; Bai et al., 2018; Angelone and Zhang, 2021), and studies on other groups are still underway.
In this study, we present the first detailed description of the lithology of the Saint-Jacques depositional sequence, along with a preliminary list of identified small mammals based on discoveries accumulated over 12 field seasons. Stratigraphical correlation of sections across the area provides a means for age comparison between fossils collected from different fault blocks. Our fine lithological divisions of this long sequence, coupled with abundant fossil material from extensive field surveys, allow for an analysis of faunal succession with improved temporal resolution. Although Saint Jacques deposits have long been considered to be exclusively Oligocene, we show that the area has well exposed sections with rich fossils that document successive faunal turnover from the Eocene to the Miocene, bracketing the entire Oligocene sequence within.
Methods
We carried out annual field expeditions to Saint Jacques in 2010 through 2022, with a gap year in 2016. Major faults in the area have previously hampered precise lithostratigraphical and biostratigraphical correlation in the past. Therefore, for the convenience of stratigraphical investigations and to avoid mixing faunal compositions across the faulted area, we divided the area into 22 fossiliferous blocks. Each block contains geographically and stratigraphically continuous exposure of beds bounded either by major faults or by topographic features. Blocks are divided into varied numbers of stratigraphic layers. Divisions are based on changes in particle size and sediment color, and they are ordered from bottom to top. For example, the strata exposed in block A are divided into eight layers, referred to as A1 (bottom) through A8 (top). We recorded fossil occurrences by their stratigraphic layers in their respective blocks, then we correlated facies across blocks by similarity in lithology.
Fossil collection started in 2011. Two collection methods are used: in situ collecting and surface collecting. When fossils are found in situ, we record the coordinates and elevation of the location and consider it a fossil locality. A locality is typically an area of a few meters across or less. Each locality is tied to a block and a stratigraphic level. Fossils that have been washed out from the place of burial and transported for a short distance were also collected from the surface. In these cases, we can usually still determine the lithological layer that the fossils are from, even though we cannot pinpoint an exact location. During each field season, we sorted through our findings daily and made field catalogue. Further identification and research were conducted after returning from the field. In order to minimize preservation bias, we made efforts to collect more thoroughly from blocks and layers in which fossils were not as abundant.
We measured five stratigraphic sections using a Brunton compass, a tape measure, and Jacob’s staff. Rock strata in Saint Jacques gently dip north-northeast at 5–20°. Because the dip of the beds is generally low, we measured sections primarily with a tape measure and then corrected for dip. Where the exposure is suitable, we used Jacob’s staff to measure the thicknesses of beds directly. The measured sections combined encompass nearly the entire stratigraphic sequence exposed in Saint Jacques. Three of the five measured sections are in Langhaogou, the E-W trending gully along which previous discoveries were made. The other two sections are located in what we refer to as Sanshenggongbei, approximately 4 km to the northeast of Langhaogou, a fossiliferous area that we newly discovered. The measured sections span about 100 m in elevation, close to the total topographic relief in Langhaogou. We mapped the locations of stratigraphic sections using a GPS device and described lithologies in detail as we measured sections. We recognized correlations of measured and unmeasured sections on the basis of the lateral and vertical relationships of facies sharing similar lithological properties and fossil evidence.
Results
Across the 22 fossiliferous blocks in Saint Jacques, we have documented 1635 fossil localities (Figure 1). The Sanshenggongbei area is divided into blocks Q, R, and T, which all have densely packed localities (Figure 1C). The other 19 blocks are distributed in Langhaogou, where more faults are present. Here, blocks A, C, D, J, and I have the most number of localities. Older strata in Langhaogou are generally in more southerly located blocks.
Lithostratigraphic correlation and classification
The five measured lithostratigraphic sequences are in blocks A, C, S–D, R, and T (Figure 2). Blocks S and D are stratigraphically continuous, only separated by a gully, and S3 partially overlaps with the lower part of D1. Fossiliferous layers are continuously numbered, except in two cases. D2 is a band of light colored siltstone sandwiched by reddish beds below and above. Vertebrate fossils are rare in D2; this layer is primarily used to separate D3 from D1, thereby achieving a higher stratigraphic resolution. The other exception is C8, a large lens of channel sandstone that cuts into the upper part of C7. Blocks A and C are largely comparable in lithology, only block C is thicker and includes more strata at the top. A5 and C5 have a distinctive brownish yellow color that differentiates this layer from strata below and above. This color is also observed in R8 in northern Saint Jacques. A7 and C7 are characterized by multiple bands of reddish mudstone (Figure 4). Additional fossiliferous areas that have been correlated to the measured sections similarity in lithology and fossil composition include blocks H, I, J, L, LD, and Q (Figure 3). Block E, representing the lower-most strata in this area, produced some remains of Gomphos but can not be correlated with any other blocks, and it is here excluded from the composite profile. Altogether, 12 lithostratigraphic units are recognized in ascending order:
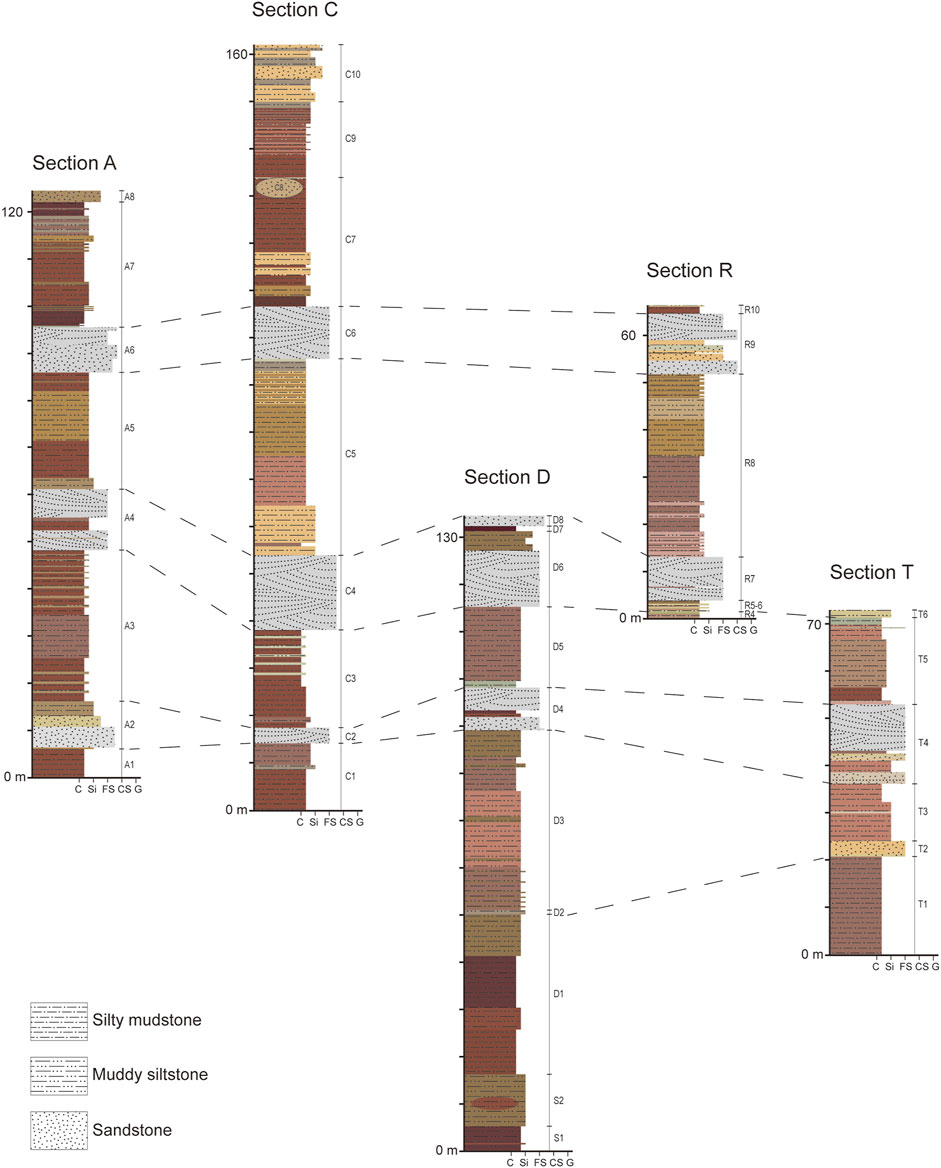
FIGURE 2. Stratigraphic columns of five measured sections at Saint Jacques, showing the correlation of units in different sections. Grain-size scale: C = clay; Si = silt; FS = fine sand; CS = coarse sand; G = gravel.
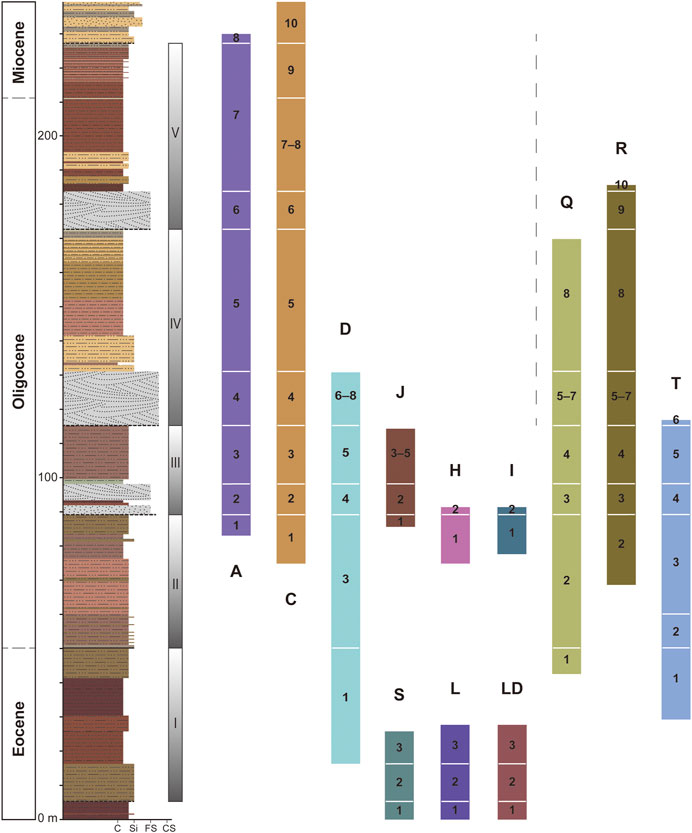
FIGURE 3. Composite stratigraphic profile for Saint Jacques (left column) and lithostratigraphic correlation of the main fossiliferous blocks (colored bars). Thicknesses of litho-units in individual blocks are not to scale. Color scheme of blocks matches legend in Figure 1. Grain-size scale: C = clay; Si = silt; FS = fine sand; CS = coarse sand.
Litho-unit 1 (S1, L1, LD1): Dark red muddy siltstone. The sedimentary structure is uniform with no visible lamination. This unit is rich in turtle fossils, while mammalian fossils are scarce. No underlying strata are exposed in block S or block L.
Litho-unit 2 (S2, L2, LD2): Brownish yellow fine sandstone interbedded with variably thick lenses of siltstone mixed with dark red muddy pebbles, carbonate nodules, and coarse sand. Cross bedding and lamination are developed in this unit. This unit includes the earliest occurrences of Leporidae, Ctenodactylidae, and Dipodidae in the sequence and the only occurrence of Gobiomys.
Litho-unit 3 (D1, S3, T1, L3, Q1): Thick layer of brownish red muddy siltstone to dark red silty mudstone. The color becomes lighter upwards with increasing amount of muddy siltstone. Massive, with no visible lamination. Fossils are sporadically discovered without any particularly rich locality. Karakoromys decessus and Ordolagus first occur in this unit.
Litho-unit 4 (A1, C1, D3, T2+3, R2, Q2, J1, H1, I1): The basal part of this unit is represented by thin layers of light brown siltstone with lamination; the upper part is composed of massive, structureless lighter brownish red muddy siltstone and darker brownish red silty mudstone. This unit contains well-preserved small mammal fossils, including relatively complete skulls and jaws. Species richness is markedly higher than in the underlying unit. Cricetops, Selenomys, Cyclomylus and other taxa begin to appear in the sequence.
Litho-unit 5 (A2, C2, D4, T4, R3, Q3, J2, H2, I2): Grayish white fine-to medium-grained sandstone with large-scale cross bedding, interbedded with lenses of coarser-grained sands with carbonate nodules or muddy pebbles. Vertebrate fossils are rich, including fish, frog, salamander, turtles, birds, and mammals. The mammalian fauna is similar to that of litho-unit 4.
Litho-unit 6 (A3, C3, D5, R4, T5, Q4, J3+4+5): Pale brownish red muddy siltstone or silty mudstone. Massive and structureless. Fossils are sporadically distributed. In block Q, a large lens of siltstone and fine sandstone is observed within the Q4; the lens is pinched off eastwards. Desmatolagus, Tataromys, and Karakoromys are abundant.
Litho-unit 7 (A4, C4, D6+7+8, R5+6+7, T6, Q5+6+7): Grayish white fine-to medium-grained sandstone with large-scale cross bedding. The reddish silty mudstone bed in between the sandstone varies in thickness from 0 to 2–3 m in different blocks. Fossils are rich in the sandstone. Karakoromys disappears from this unit.
Litho-unit 8 (A5, C5, Q8, R8): Thick layer of yellowish brown muddy siltstone and silty mudstone, massive and structureless, with manganese nodules. Small mammals are sporadically distributed in this unit. Sinolagomys first appear from the upper part of this unit.
Litho-unit 9 (A6, C6, R9): Grayish white fine-to coarse-grained sandstone with large-scale cross bedding. Large mammalian fossils are rich, including skulls and postcranial of giant rhinos. Tachyoryctoides are first documented from this unit.
Litho-unit 10 (A7, C7+8, R10): Massive brownish red silty mudstone interbedded with bands of reddish mudstone. C8 is composed of fine or coarse sandstone, pinched off westwards, thickening eastwards, and punctuated by a fault on the west side. Fossils of Sinolagomys and large-sized Yindirtemys are abundant in this unit.
Litho-unit 11 (C9): Alternating light brownish red muddy siltstone and dark brown silty mudstone. Sinolagomys remains abundant while Yindirtemys disappears in this unit.
Litho-unit 12 (C10): Pale orange fine-grained sandstone, with light brown muddy siltstone in the upper part. Small-sized Sinolagomys, Prodistylomys, and some insectivores are documented in this unit.
We created a composite stratigraphic profile of Saint Jacques using measurements from blocks S, D, and C, which combined encompass the longest temporal and lithostratigraphic records (Figure 4). The composite section spans 239 m. Particle size of the sediments is generally small. Aside from the three layers of fine- to medium-grained sandstone, the sequence is mainly composed of reddish silty mudstone and muddy siltstone. Rock color is generally darker towards the bottom and lighter towards the top. Section S through the bottom of section D (litho-units 1–3) is marked by dark red silty mudstone and moderate brown muddy siltstone. Several layers of pale orange yellow muddy siltstone appear in the upper half of the sequence.

FIGURE 4. Outcrop photographs of selected measured sections.
Small mammals in stratigraphic context
Fossil mammals are collected from litho-unit 1 through litho-unit 11. With a preliminary examination of the small mammals from these units, we have identified 65 taxa belonging in eleven families of small mammals, including seven families of rodents (Aplodontidae, Ctenodactylidae, Cricetidae, Cylindrodontidae, Dipodidae, Gobiomyidae, Tsaganomyidae), two families of lagomorphs (Leporidae and Ochotonidae), and two families of insectivores (Changlelestidae and Erinaceidae) (Table 1). Additionally, the basal Glires Gomphos has been found in block E, putting it to stratigraphically below litho-unit 1.

TABLE 1. Stratigraphic levels of selected small mammals identified from Saint Jacques.
Taxonomic richness of small mammals increases progressively from litho-unit 1 to litho-unit 8 and then sharply declines in higher units (Figure 5). The greatest increase occurs between litho-units 3 and 4. This pattern is a result of a step increase in the number of ochotonid taxa as well as the appearance of cricetids, tsaganomyids, and erinaceids in litho-unit 4. Ctenodactylid richness increases relatively steadily across these units. Among the rodents, the aplodontids appear the latest in the sequences and have the shortest duration—they are only found in litho-units 6 and 7. An abrupt drop in taxonomic richness is observed in litho-unit 9. The abundance of relatively complete large-mammal fossils in this unit suggests that flow velocity was high and not ideal for the deposition of small-mammal material. Therefore, the decline in the richness of small mammals may reflect, at least in part, the condition of preservation rather than a true ecological signal.
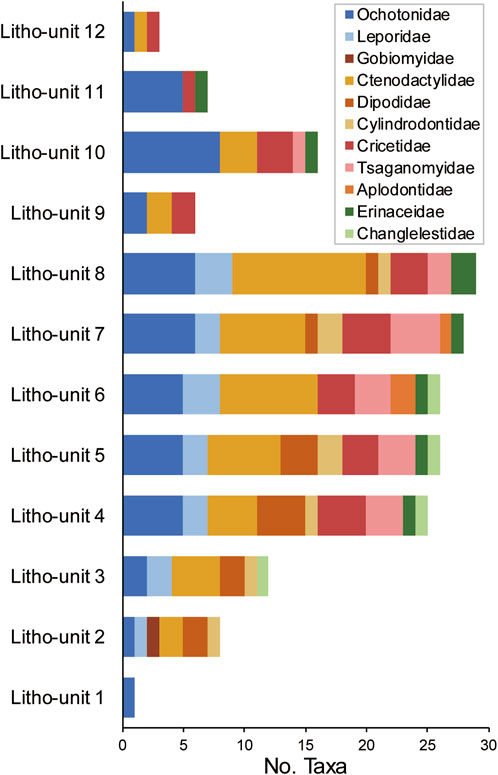
FIGURE 5. Number of small-mammal taxa in different lithostratigraphic units. Lagomorph families are in blue colors, rodent families in warm colors, and insectivore families in green colors.
Faunal turnover appears to be gradual across the sequence (Table 1). Even though the taxonomic richness remains relatively stable during the deposition of litho-unit 4 through litho-unit 8, the small-mammal fauna was not static. Rather, a stepwise transition in the taxonomic composition is documented, especially for the ochotonids, ctenodactylids, and cricetids.
Discussion
Our re-exploration of the Saint Jacques area has revealed developed terrestrial sediments with rich fossils. The significance of this work is several fold. First, we have recovered fossils from a larger area at Saint Jacques than previously known. In addition to the localities in Langhaogou (roughly equivalent to the former Saint Jacques site), we discovered a fossiliferous area to the northeast that was unknown before. These areas have been intensively surveyed over 12 field seasons, as shown by the high density of new fossil localities (Figure 1). Our continuous field expeditions have yielded thousands of pieces of fossil-mammal material, all tied to specific stratigraphic units. Comparing to the existing fauna lists of Saint Jacques (e.g., Meng and McKenna, 1998; Wang and Qiu, 2003), a number of taxa are recorded for the first time, including Sinolagomys ulunguensis, S. pachygnathus, Yindirtemys suni, Yindirtemys grangeri, Gobiomys, Tachyoryctoides kokonorensis, Eucricetodon jilantaiensis, Cyclomylus lohiculus, Allosminthus, Parasminthus, Prodistylomys, and Ayakozomys. Our thorough investigation of the field area thus reveals a fuller picture of the mammalian assemblages of Saint Jacques.
Second, we conducted the first detailed study on the lithological subdivision of Saint Jacques strata. This provided a lithological context for the fossiliferous sequence and allowed for the correlation of facies across blocks. The establishment of this chronostratigraphic framework formed the basis for documenting and understanding the deposition of fossil materials, with improved temporal resolution. We recognize 12 lithostratigraphic units in Saint Jacques, and mammalian fossils have been collected from all of them (Table 1; Figure 5). Additional material (i.e., Gomphos) has been found in even older strata (block E) in the area. This fine division of stratigraphic units allows for analyses of faunal transition in smaller time bins and with reduced errors introduced by time averaging.
Third, a preliminary examination of the rich small-mammal materials (especially rodents and lagomorphs) reveals that faunal assemblages in Saint Jacques show successive evolutionary stages. Most taxa occur in stratigraphically continuous litho-units (Table 1), documenting the pattern of faunal succession. No prominent hiatus in the depositional sequence has been observed. This record, therefore, provides an excellent opportunity for further studying the faunal composition of continental Asia from the late Eocene through the Oligocene. Wasiljeff et al. (2020) recently provided better age constraint on the Ulantatal sequence in Nei Mongol and demonstrated that major faunal turnover preceded (instead of precisely at) the Eocene–Oligocene (E–O) boundary. Research on the magnetostratigraphy of the Saint Jacques sequence is still underway. A deeper look into the timing and pattern of faunal transition in Saint Jacques will help elucidate the potential complexity in terrestrial faunal responses to broad-scale changes in climate and environment.
Prior to our revisit of Saint Jacques in 2010, the common understanding of its paleontology was that the area contained two local faunas, one of early Oligocene age and the other late Oligocene (e.g., Wang and Qiu, 2003). In contrast, our work now suggests that the sedimentary strata in Saint Jacques likely span the Eocene through the early Miocene, bracketing a long, successive Oligocene sequence within. Bai et al. (2018) reported A. praecox from L2 (litho-unit 2), correlating this unit to the Ergilian Asian Land Mammal Age (latest Eocene). In line with this result, we have identified Gobiomys in L2 (litho-unit 2). This rodent genus is known exclusively from the Eocene of Nei Mongol (Wang, 2001; Li et al., 2022). The presence of Ardynia praecox and Gobiomys shows that the lower Saint Jacques sequence extends into the Eocene. Another taxon that sheds light on the E–O boundary is Ardynomys, a primitive cylindrodontids previously known from the late Eocene of Nei Mongol (Wang and Wang, 1991; Wang and Meng, 2009; Gomes Rodrigues et al., 2014; Wasiljeff et al., 2020; Wasiljeff and Zhang, 2022), the late Eocene to the early Oligocene of Mongolia (Dashzeveg, 1996; Daxner-Höck et al., 2017), and the late Eocene of North America (Wood, 1970; 1974; Korth, 1992). In Saint Jacques, Ardynomys occurs in litho-units 3 and 4. Considering the occurrence of other Oligocene taxa (e.g., Desmatolagus gobiensis, Eucricetodon, Cricetops, Selenomys mimicus, Tsaganomys) in litho-unit 4, the epoch boundary is most likely within litho-unit 3 at Saint Jacques. Taxa in the top layers at Saint Jacques show similarity with late Oligocene to early Miocene faunas in northern China and Mongolia. For example, T. kokonorensis, Sinolagomys pachygnathus, Eucricetodon youngi, and Y. suni are representatives of the Xiejian faunas of China, considered to be the early Miocene in age (Li and Qiu, 1980; Qiu and Qiu, 1995; Qiu et al., 2001; Wang and Qiu, 2012; Qiu et al., 2013). In Saint Jacques, these species occur in litho-units 10 and 11. T. kokonorensis (litho-unit 10), Y. suni (litho-unit 11), Prodistylomys (litho-unit 12), and Ayakozomys (litho-unit 12) occur in the Valley of Lakes, Mongolia during the latest Oligocene to the early Miocene (Daxner-Höck et al., 2015; 2017; Harzhauser et al., 2017).
To conclude, our re-investigation of Saint Jacques has greatly expanded the chronostratigraphic and taxonomic coverage of the mammalian fossil collection from this area. Our preliminary biostratigraphic analysis of small mammals shows that the area preserves faunal succession from the late Eocene to the early Miocene, making it an important record for studying mammalian faunal turnovers during both the Eocene–Oligocene and the Oligocene–Miocene transitions.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Author contributions
Data synthesis and manuscript preparation: BW, Z-QZ, and Y-QW. All authors participated in field work and data collection.
Funding
Field work was supported by the Strategic Priority Research Program of Chinese Academy of Sciences (XDB26000000), the National Natural Science Foundation of China (41472003), and the Special Fund for Fossil Excavation and Preparation of the Chinese Academy of Sciences.
Acknowledgments
We are grateful to many individuals for their field assistance over the years: Wei Zhou, Shi-Jie Li, Qi Li, Yong-Xing Wang, Yong-Fu Wang, Xin-Yue Zhang, Ping Wang, and others. We would also like to thank the reviewers for their constructive feedback on the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Angelone, C., and Zhang, Z.-Q. (2021). Climate change and evolution of early lagomorphs (Mammalia): A study perspective based on new materials of Ordolagus from Nei Mongol (northern China). Vert. PalAsiat. 59, 138–168.
Bai, B., Wang, Y.-Q., and Zhang, Z.-Q. (2018). The late Eocene hyracodontid perissodactyl Ardynia from Saint Jacques, Inner Mongolia, China and its implications for the potential Eocene–Oligocene boundary. Palaeoworld 27, 247–257. doi:10.1016/j.palwor.2017.09.001
Chow, M. C., and Rozhdestvensky, A. K. (1960). Exploration in Inner Mongolia — A preliminary account of the 1959 field work of the Sino-Soviet paleontological expedition (SSPE). Vert. PalAsiat. 4, 1–10.
Coxall, H. K., Wilson, P. A., Palike, H., Lear, C. H., and Backman, J. (2005). Rapid stepwise onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean. Nature 433, 53–57. doi:10.1038/nature03135
Dashzeveg, D. (1996). A new Ardynomys (Rodentia, Cylindrodontidae) from the Eocene of the eastern Gobi desert, Mongolia. Palaeovertebrata 25, 339–348.
Daxner-Höck, G., Badamgarav, D., Barsbold, R., Bayarmaa, B., Erbajeva, M., Göhlich, U. B., et al. (2017). Oligocene stratigraphy across the Eocene and Miocene boundaries in the Valley of Lakes (Mongolia). Palaeobio. Palaeoenv. 97, 111–218. doi:10.1007/s12549-016-0257-9
Daxner-Höck, G., Badamgarav, D., and Maridet, O. (2015). Evolution of Tachyoryctoidinae (Rodentia, Mammalia): Evidences of the Oligocene and early Miocene of Mongolia. Ann. Naturhist. Mus. Wien, Ser. A 117, 161–195.
Dupont-Nivet, G., Krijgsman, W., Langereis, C., Abels, H. A., Dai, S., and Fang, X. (2007). Tibetan plateau aridification linked to global cooling at the Eocene–Oligocene transition. Nature 445, 635–638. doi:10.1038/nature05516
Eldrett, J., Greenwood, D., Harding, I., and Huber, M. (2009). Increased seasonality through the Eocene to Oligocene transition in northern high latitudes. Nature 459, 969–973. doi:10.1038/nature08069
Gomes Rodrigues, H., Marivaux, L., and Vianey-Liaud, M. (2014). Rodent paleocommunities from the Oligocene of Ulantatal (Inner Mongolia, China). Palaeovertebrata 38 (1–11), e3. doi:10.18563/pv.38.1.e3
Harzhauser, M., Daxner-Höck, G., Erbajeva, M. A., López-Guerrero, M., Maridet, O., Oliver, A., et al. (2017). Oligocene and early Miocene mammal biostratigraphy of the Valley of Lakes in Mongolia. Palaeobio. Palaeoenv. 97, 219–231. doi:10.1007/s12549-016-0264-x
Hren, M. T., Sheldon, N. D., Grimes, S. T., Collinson, M. E., Hooker, J. J., Bugler, M., et al. (2013). Terrestrial cooling in northern Europe during the Eocene–Oligocene transition. Proc. Natl. Acad. Sci. U.S.A. 110, 7562–7567. doi:10.1073/pnas.1210930110
Korth, W. W. (1992). Cylindrodonts (Cylindrodontidae, Rodentia) and a new genus of eomyid, Paranamatomys, (Eomyidae, Rodentia) from the Chadronian of Sioux County, Nebraska. Trans. Neb. Acad. Sci. 19, 75–82.
Kraatz, B. P., and Geisler, J. H. (2010). Eocene–Oligocene transition in Central Asia and its effects on mammalian evolution. Geology 38, 111–114. doi:10.1130/g30619.1
Li, C.-K., and Qiu, Z.-D. (1980). Early Miocene mammalian fossils of Xining basin, Qinghai. Vert. PalAsiat. 18, 198–214.
Li, Q., Li, Q., Xu, R., and Wang, Y. (2022). Rodent faunas, their paleogeographic pattern, and responses to climate changes from the early Eocene to the early Oligocene in Asia. Front. Ecol. Evol. 10, 955779. doi:10.3389/fevo.2022.955779
Lopatin, A. V. (2004). Early Miocene small mammals from the North Aral Region (Kazakhstan) with special reference to their biostratigraphic significance. Paleontol. J. 38, S217–S323.
Meng, J., and McKenna, M. (1998). Faunal turnovers of palaeogene mammals from the Mongolian plateau. Nature 394, 364–367. doi:10.1038/28603
Qiu, Z.-X., and Qiu, Z.-D. (1995). Chronological sequence and subdivision of Chinese Neogene mammalian faunas. Palaeogeogr. Palaeoclimatol. Palaeoecol. 116, 41–70. doi:10.1016/0031-0182(94)00095-p
Qiu, Z.-X., Qiu, Z.-D., Deng, T., Li, C.-K., Zhang, Z.-Q., Wang, B.-Y., et al. (2013). “Neogene land mammal stages/ages of China: Toward the goal to establish an Asian land mammal stage/age scheme,” in Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology, eds. X.-M. Wang, L. J. Flynn, and M. Fortelius (New York: Columbia University Press), 29–90.
Qiu, Z., Wang, B., Qiu, Z., Heller, F., Yue, L., Xie, G., et al. (2001). Land-mammal geochronology and magnetostratigraphy of mid-Tertiary deposits in the Lanzhou basin, Gansu Province, China. Eclogae. Geol. Helv. 94, 373–385.
Shi, W., Dong, S. W., and Hu, J. M. (2020). Neotectonics around the Ordos block, north China: A review and new insights. Earth Sci. Rev. 200, 102969. doi:10.1016/j.earscirev.2019.102969
Sun, J., Ni, X., Bi, S., Wu, W., Ye, J., Meng, J., et al. (2014). Synchronous turnover of flora, fauna, and climate at the Eocene–Oligocene boundary in Asia. Sci. Rep. 4, 7463. doi:10.1038/srep07463
Teilhard de Chardin, P. (1926). Description de mammifères Tertiaires de Chine et de Mongolie. Ann. Paleontol. 15, 1–52.
Teilhard de Chardin, P., and Licent, E. (1924a). On the geology of the northern, Western and southern borders of the Ordos, China. Geol. Bull. China 3, 37–44. doi:10.1111/j.1755-6724.1924.mp3001004.x
Teilhard de Chardin, P., and Licent, E. (1924b). Observations géologiques sur la bordure occidentale et méridional de l’Ordos. Bull. Soc. Géol. Fr. Ser. 4. 24, 29–91.
Teilhard de Chardin, P., and Licent, E. (1924c). Observations complémentaires sur la géologiques de l'Ordos. Bull. Soc. Géol. Fr. Ser. 4. 24, 462–464.
Wang, B.-Y. (1987). Discovery of Aplodontidae (Rodentia, Mammalia) from middle Oligocene of Nei Mongol, China. Vert. PalAsiat. 25, 32–45.
Wang, B.-Y., and Emry, R. J. (1991). Eomyidae (Rodentia: Mammalia) from the Oligocene of Nei Mongol, China. J. Vert. Paleontol. 11, 370–377. doi:10.1080/02724634.1991.10011404
Wang, B.-Y. (2001). Eocene ctenodactyloids (Rodentia, Mammalia) from Nei Mongol, China. Vert. PalAsiat. 39, 98–114.
Wang, B.-Y., and Meng, J. (2009). Ardynomys (Cylindrodontidae, Rodentia) from Nei Mongol, China. Vert. PalAsiat. 47, 240–244.
Wang, B.-Y., and Qiu, Z.-X. (2003). Note on early Oligocene ursids (Carnivora, Mammalia) from Saint Jacques, Nei Mongol, China. Bull. Am. Mus. Nat. Hist. 279, 116–124.
Wang, B.-Y., and Qiu, Z.-X. (2012). Tachyoryctoides (Muroidea, Rodentia) fossils from early Miocene of Lanzhou basin, Gansu Province, China. Swiss J. Palaeontol. 131, 107–126. doi:10.1007/s13358-011-0038-z
Wang, B.-Y., and Wang, P.-Y. (1991). Discovery of early medial Oligocene mammalian fauna from Kekeamu, Alxa left banner, Nei Mongol. Vert. PalAsiat. 29, 64–71.
Wang, J., and Zhang, Z.-Q. (2015). Phylogenetic analysis on Palaeogale (Palaeogalidae, Carnivora) based on specimens from Oligocene strata of Saint-Jacques, Nei Mongol. Vert. PalAsiat. 53, 310–334.
Wasiljeff, J., Kaakinen, A., Salminen, J. M., and Zhang, Z.-Q. (2020). Magnetostratigraphic constraints on the fossiliferous Ulantatal sequence in Inner Mongolia, China: Implications for Asian aridification and faunal turnover before the Eocene–Oligocene boundary. Earth Planet. Sci. Lett. 535, 116125. doi:10.1016/j.epsl.2020.116125
Wasiljeff, J., and Zhang, Z.-Q. (2022). Stratigraphical significance of Ulantatal sequence (Nei Mongol, China) in refining the latest Eocene and Oligocene terrestrial regional stages. Vert. PalAsiat. 60, 42–53.
Westerhold, T., Marwan, N., Drury, A. J., Liebrand, D., Agnini, C., Anagnostou, E., et al. (2020). An astronomically dated record of Earth’s climate and its predictability over the last 66 million years. Science 369, 1383–1387. doi:10.1126/science.aba6853
Wood, A. E. (1974). Early Tertiary vertebrate faunas Vieja group trans-pecos, Texas: Rodentia. Tex. Mem. Mus. Bull. 21, 1–112.
Wood, A. E. (1970). The early Oligocene rodent Ardynomys (family Cylindrodontidae) from Mongolia and Montana. Am. Mus. Novit. 2418, 1–18.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K. (2001). Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292, 686–693. doi:10.1126/science.1059412
Zhang, R., Kravchinsky, V. A., and Yue, L. (2012). Link between global cooling and mammalian transformation across the Eocene–Oligocene boundary in the continental interior of Asia. Int. J. Earth Sci. 101, 2193–2200. doi:10.1007/s00531-012-0776-1
Keywords: Nei Mongol, lithostratigaphy, Saint Jacques, Eocene-Oligocene boundary, Oligocene-Miocene boundary, mammals
Citation: Wang B, Zhang Z-Q, Wang Y-Q, Li Q, Bai B, Liu Y, Mao F-Y, Wang H-B, Wang J, Gong Y-X, Dong L-P, Wang L-H, Ma H-D, Xu R-C and Wang X-Y (2023) Lithostratigraphy of a long, fossiliferous Oligocene sequence: Revisiting Saint Jacques, Nei Mongol, China. Front. Earth Sci. 11:1127964. doi: 10.3389/feart.2023.1127964
Received: 20 December 2022; Accepted: 08 February 2023;
Published: 20 February 2023.
Edited by:
Shuhai Xiao, Virginia Tech, United StatesReviewed by:
Xiaoming Wang, Natural History Museum of Los Angeles County, United StatesYangfan Li, Northwest University, China
Copyright © 2023 Wang, Zhang, Wang, Li, Bai, Liu, Mao, Wang, Wang, Gong, Dong, Wang, Ma, Xu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuan-Qing Wang, d2FuZ3l1YW5xaW5nQGl2cHAuYWMuY24=; Zhao-Qun Zhang, emhhbmd6aGFvcXVuQGl2cHAuYWMuY24=