 Madani Benyoucef1*
Madani Benyoucef1* Adán Pérez-García2
Adán Pérez-García2 Mohamed Bendella3
Mohamed Bendella3 Francisco Ortega2
Francisco Ortega2 Romain Vullo4
Romain Vullo4 Imad Bouchemla1
Imad Bouchemla1 Bruno Ferré5
Bruno Ferré5- 1Faculty of Natural and Life Sciences, University Mustapha Stambouli of Mascara, Mascara, Algeria
- 2Grupo de Biología Evolutiva, Facultad de Ciencias, UNED, Madrid, Spain
- 3Mohamed Ben Ahmed Oran 2 University, Geodynamic of Basin and Sedimentary Balance Laboratory, Oran, Algeria
- 4Univ Rennes, CNRS, Géosciences Rennes, UMR, Rennes, France
- 5Saint-Étienne-du-Rouvray, Rennes, France
Excellent outcrops of the upper part of the “Continental Intercalaire” deposits occur in the Gara Samani area, southern Algeria. These strata are recognised herein as belonging to the Gara Samani Formation, which is composed of the Samani Vertebrate-rich Sand Member, the Samani Sandstone Member and the Méguidène Stratified Sand Member. The first two members are from a fluvial system and the last member corresponds to an aeolian (dune and interdune) system. They have preserved one of the most diverse continental vertebrate faunas of latest Albian–early Cenomanian age yet known from northern Africa. Most of the taxa identified there, cited for the first time for the “mid”-Cretaceous of Algeria, have not been analysed in detail so far; and many of them have never been figured. The faunal list of vertebrates from this locality was supplemented thanks to detailed analysis of new and relatively abundant finds. As a consequence, the Gara Samani vertebrate assemblage is recognised as composed of elasmobranchs (two Hybodontiformes, i.e., Tribodus sp. and Distobatidae indet.; and a Batoidea, Onchopristis numida), actinopterygians (Bawitius sp., Adrianaichthys sp., Obaichthys africanus, and Calamopleurus africanus), sarcopterygians (Mawsoniidae indet. and the Dipnoi, cf. Lavocatodus humei and Neoceratodus africanus), testudines (at least two Pelomedusoides, one of them being a representative of Araripemydidae), crocodyliforms (all remains being compatible with Neosuchia, represented at least by Elosuchus broinae), ornithocheirid pterosaurs, and non-avian dinosaurs (titanosauriform sauropods and at least two theropods, Spinosaurus aegyptiacus and Carcharodontosaurus saharicus). This fauna association displays close similarities to contemporaneous ones from North Africa, especially with that from the Kem Kem beds.
1 Introduction
During the first decades of the 19th century, field geologists had begun to explore the geology of North Africa. They subsequently discovered the widespread continental siliciclastic formations that build the slope and foot of the prominent limestone plateaus of the Tademaït and the Tinrhert hamadas. To describe them, they generally used different terms such as ‟argiles bariolées” (variegated clays), ‟grès à sphéroïdes” (spheroid sandstones), ‟terrains des gours” (gour areas; gour = plural form of gara, tabular hill, usually an erosional outlier), ‟grès à dragées” (pebbly sandstones), etc., all based on the lithological features commonly found within these numerous formations, scattered along the geological time scale and discontinuous in space.
In a short note entitled ‟Des principaux complexes continentaux du Sahara”, Kilian (1931) published a stratigraphic scheme of the successive continental series respectively found in the Sahara. He identified two large continental complexes, both under- and overlain by fossiliferous marine series: 1) the ‟Continental Intercalaire” (“intercalated continental series”), usually found over the last marine levels of the Palaeozoic (Namurian) strata and under the first marine level of the transgressive Upper Cretaceous strata (cf. Lefranc and Guiraud, 1990); 2) the ‟Continental terminal” (“topmost continental series”) overlying the marine upper Cretaceous strata and corresponding to the Cenozoic deposits. Moreover, subsequently Kilian (1937) precised the definition of the “Continental Intercalaire’’ and introduced the concept of the ‟Continental post-tassilien” (“post-tassilian continental series”), as a non-marine facies representing the upper part of the Palaeozoic deposits (Table 1). The lower boundary of the ‟Continental Intercalaire” complex is problematic. It does not correspond to any geological event in northern Africa. Kilian (1931) suggested the lower boundary should include all marine sediments of Carboniferous age. In contrast, the upper boundary of the “Continental Intercalaire” is defined at the base of a well-dated, fossiliferous late Cenomanian-Turonian limestone plateaus. The concept of “Continental Intercalaire” is applicable across all North Africa, but is obviously extremely vague from a stratigraphic point of view.

TABLE 1. Divisions of the northern African continental deposits after Kilian (1931), modified by Kilian (1937). Abbreviation: ICS, International Commission on Stratigraphy.
The stratotype for the “Continental Intercalaire” is located in the Djoua Valley, northern Hoggar, in the Algerian-Libyan bordering area. Coeval strata are located in continental basins: in Libya and Southern Egypt to the East, in Tunisia to the North, in Morocco to the West, and in Niger, Northwestern Nigeria, Northern Malawi, Southern Tanzania, and South Africa, all South of the Hoggar Mountains (Le Loeuff et al., 2012).
The occurrence of vertebrate fossil remains (bones and teeth) in formations assigned to the ‟Continental Intercalaire” sensu lato is reported in Algeria from several localities and has been the core topic of many papers (e.g., Haug, 1904; Haug, 1905; Lapparent, 1947; Lapparent, 1951; Lapparent, 1960; Tabaste, 1963; Bassoullet and Iliou, 1967; Broin et al., 1971; Lefranc and Toutin, 1971; Taquet, 1972; Taquet and Russell, 1998; Benyoucef et al., 2014; Benyoucef et al., 2015; Gabani et al., 2016), but these fossil deposits have been so far described and illustrated in a rather preliminary way. The most important ‟mid”-Cretaceous fossil deposits identified in Algeria are (Figure 1): Oued Boudjihane (Bassoullet and Iliou, 1967) in the Western Saharan Atlas, Menaguir and Kénadsa (Benyoucef et al., 2014; Benyoucef et al., 2015) in the Béchar area, Gara Samani in Timimoun, and Tab Tab and In Akhamil in the Djoua valley (Foureau, 1904; Haug, 1904; Haug, 1905; Tabaste, 1963). These are considered as Albian in age by former authors.
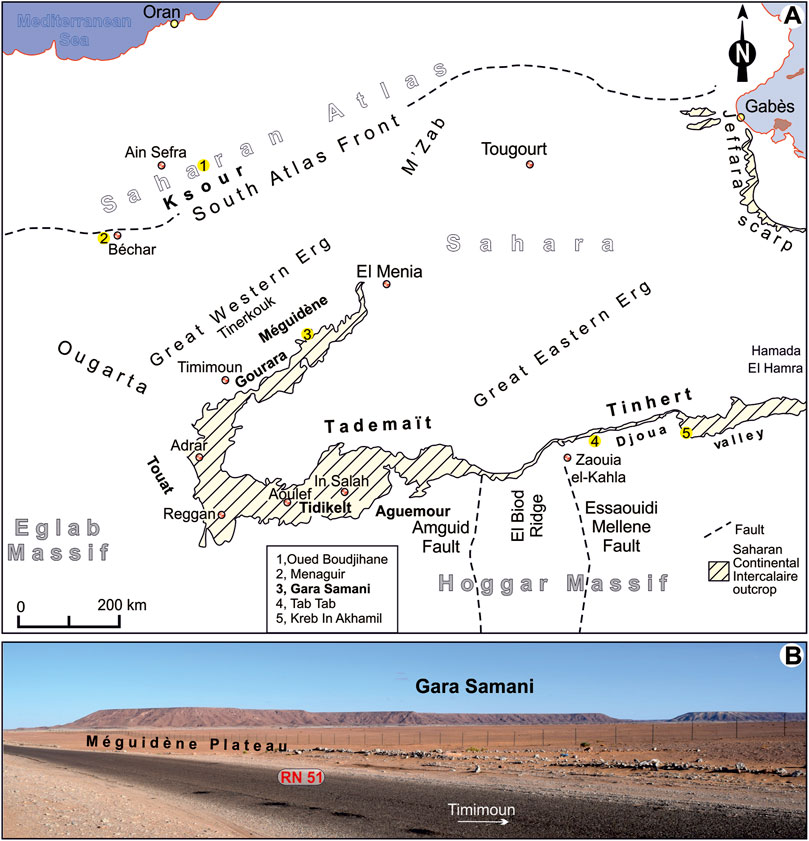
FIGURE 1. Location of the studied area. (A) Location of the studied section within the Saharan domain compared to the ‘‘Continental Intercalaire’’ outcrops and the main lower Cenomanian vertebrate sites (after: 1, Bassoullet and Iliou 1967; 2, Benyoucef et al., 2015; 3, present work, 4 and 5, Gabani et al., 2016); (B) Panoramic views of the studied succession (Western flank of Gara Samani).
The discovery of vertebrate remains on the paleontological site of the Gara Samani was reported by two herpetologists and behavioural specialists of varanids and sand vipers (C. Grenot and R. Vernet) during a monitoring mission called ‟Reptiles vivants du Sahara”, in February 1970. One year later, Broin et al. (1971) described these remains and proposed a preliminary faunal list, including, among others, Onchopristis numida, Spinosaurus maroccanus (cf. Taquet and Russell, 1998, but see also Buffetaut and Ouaja, 2002, Cavin et al., 2010, Dal Sasso et al., 2005 and Ibrahim et al., 2014 on the validity of the taxon), Carcharodontosaurus saharicus (Taquet, 1997) and a large crocodile skull later referred to Elosuchus cherifiensis, a species originally described by Lavocat (1955) from the Kem Kem beds of Morocco (cf. Lapparent de Broin, 2002). The vertebrate-rich deposits of Gara Samani are considered as of Albian age by Broin et al. (1971) and Lefranc and Toutin (1971).
In the present work, we review and revise the continental vertebrate taxa recovered from the Gara Samani during two field trips (November 2020 and March 2021). The collected fauna is briefly described and illustrated here. The lithostratigraphic and sedimentological framework of vertebrate-bearing strata is also provided.
2 Geological Setting
The geology of the Gourara-Méguidène area combines the geological history of an atypical but mobile, hercynian sag basin, and continental-dominant Mesozoic strata (Nedjari and Ait Ouali, 2018). Its singularity originates from its location on the suture zone between two cratons differing by their rigidity, age and, subsequently, respective behaviour through time.
The Devonian-Carboniferous strata had built an intensively folded range structured by a series of synclines and anticlines. Sedimentation took place in a distal-to-proximal delta setting controlled by both eustacy and tectonics during the Tournaisian and then the late Visean (Debachi, 2007).
The Lower Cretaceous deposits overlie a huge sedimentation gap spanning from the Late Carboniferous to the Late Jurassic, during which all the coeval-folded Palaeozoic strata had been peneplanated. They are lying in angular unconformity on the preceding strata (Palaeozoic) that had been eroded and altered during a long period of time. These are characterised by massive detrital inputs ( = ‟Continental Intercalaire” series sensu Kilian, 1931) that are exposed over almost the whole Sahara, including Precambrian-exposing areas. The ‟Continental Intercalaire” stretches from the edge of the Saharan platform where they build the continuous aureole from the El Menia (formerly named El Goléa) to the Southern edge of the El Hamra Hamada (Libya). In the Northwest, the ‟Continental Intercalaire” is exposed all over the Saharan Atlas and, in the Northeast, and at the foot of the Jeffara cliff in Tunisia (Figure 1A). In the South, the ‟Continental Intercalaire” is directly overlying in angular unconformity on the folded Palaeozoic marine formations.
In the Gara Samani, deposition of the ‟Continental Intercalaire” terminates with the gypsiferous clays (30–60 m-thick) of the EI Goléa Formation (‟Argiles d’El Goléa”; Lapparent, 1937). This lagoonal formation is always presumed under the upper Cenomanian fossiliferous (Neolobites vibrayeanus, bivalves, oysters, echinoids, and gastropods) limestone, indicating the huge Cenomanian marine transgression. Higher in the series, the deposits are marine and lagoonal: the Late Cretaceous marine settings display the hard limestone beds that build the current, vast Tademaït Plateau.
The geological map of Timimoun (1:50.000 scale, map NH-31-SO) sums up the key points of the geology of the Gourara-Méguidène area: According to this map, the ‟Continental Intercalaire” complex (‟Grès albien” of earlier authors) outcropping in the Gara Samani locality consists of four formal units which directly overlie the quartzite slab (‟dalle quartzitique” sensu Meyendorff, 1938 and unit B sensu Lapparent, 1947): 1) the Sand and Quartzite of Méguidène, 2) the Vertebrate-rich Sand of Samani (as the present investigated levels), 3) the Stratified Sand of Samani, and 4) the El Goléa Clays (Figure 2). The three first units correspond to Unit C of Lapparent (1947).
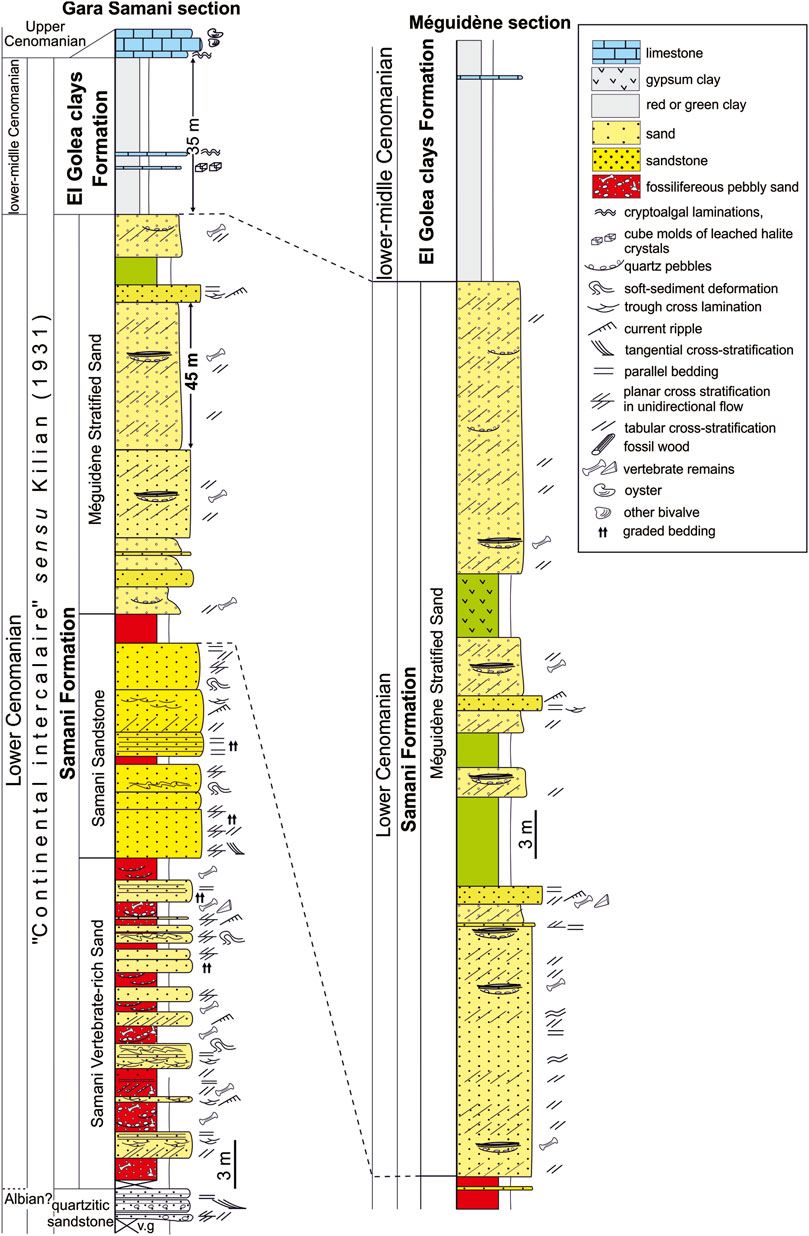
FIGURE 2. Lithostratigraphic log of the Samani Formation with respective stratigraphic members in the Gara Samani and Méguidène localities.
3 Materials and Methods
The Gara Samani (UTMS coordinates: 29°41′31.14″N; 1°56′20.62″E) is located about 40 km East from the Méguidène village, 2 km to the Southeast of the National Road RN 51 (Figure 1B), linking the towns of Timimoun and El Menia. Good exposures of the upper part of the “Continental Intercalaire” are at the foot of the Gara Samani within a 100–300 m-wide strip, the basal part of which is sand-dominated and particularly rich in vertebrate remains. The facies were identified based on lithofacies, bounding surfaces, internal sedimentary structures, and textures. The collected fossil vertebrate remains (fishes, turtles, crocodiles, pterosaurs, and non-avian dinosaurs) were cleaned, identified, described, discussed, and photographed using digital cameras. The respective figured specimens are housed in the collections of the Geomatics, Ecology and Environment Laboratory (University Mustapha Stambouli of Mascara, Algeria) under registration numbers LGS.
Institutional abbreviations: LGS, Laboratoire de Géomatique, Écologie et Environnement, Gara Samani section.
4 Description of the Gara Samani Formation
The ‟Continental Intercalaire” deposits outcropping around the Gara Samani (Northern Tademaït Plateau) is referred herein to the Gara Samani Formation, according to the eponymous tabular hill (see Figure 1B; Figures 2, 3). This formation is well exposed between the Méguidène village and the Gara Samani locality. It is correlated to the top of a package of continental sediments in basins scattered across Northern Africa, and referred by Kilian (1931) as the “Continental Intercalaire”, or the “intercalated continental (deposits)”.

FIGURE 3. Panoramic view of the northern side of Gara Samani showing the studied members.
The Gara Samani Formation would be the coeval deposits of the Djoua series on the Algerian-Libyan borders (Haug, 1904; Lapparent and Lelubre, 1948; Lefranc, 1983; Gabani et al., 2016), of the upper part of the Tiout Formation in the Ksour Mountains (Bassoullet and Iliou, 1967; Benyoucef et al., 2017), and to the ‟Grès rouges’’ Formation outcropping in the lowermost part of the Cretaceous succession of the Guir area (Benyoucef et al., 2014; Benyoucef et al., 2015; Benyoucef et al., 2016). In Northern Africa, outside Algeria, the Gara Samani Formation is correlated to the Ifezouane Formation (Dubar, 1948) in the Errachidia-Boudnib-Erfoud basin and to the Kem Kem beds in Morocco (Sereno et al., 1996; Cavin et al., 2010), to the Aïn el Guettar Formation in Tunisia (Buffetaut and Ouaja, 2002) to the Draa Ubari Formation in Libya (Rage and Cappetta, 2002), and to the Bahariya Formation in Egypt (Said, 1962).
In the studied area, the Gara Samani Formation reaches a thickness of ca 90 m, overlies the quartzitic sandstone slab that builds the Méguidène Plateau (Figure 1B), and underlies the El Goléa Clay Formation. It mainly consists of sand, sandstone and clay, with subordinate thin, interbedded conglomerate. It can be subdivided into three formal members (Figures 2, 3), based on their respective faunal, lithologic and sedimentologic features.
4.1 Samani Vertebrate-Rich Sand Member
This member is more widely exposed at Gara Samani than at any other area, and varies in thickness from 20 to 25 m. It is mostly composed of a fining-upward siliciclastic succession represented by fine to coarse-grained, moderately to poorly sorted, quartz sands, interbedded with sandstone, clay, chenalised quartz pebbles and conglomerate. The sand facies yields the main concentration of vertebrate remains (pterosaur teeth, theropod and crocodyliform teeth and bones, turtle plates and bones, elasmobranch, actinopterygian, and sarcopterygian fishes, and several morphotypes of coprolites; Figure 4A). The quartz grains are generally rounded to subrounded, brown, reddish brown or translucent, and have a characteristic dull or opaque (frosted) surface texture. The sandstone beds (0.40–1.5 m-thick) can be differentiated by an assemblage of sedimentary structures that includes trough cross lamination, planar cross lamination, planar horizontal laminations and fining up-ward trend (Figures 4B–E). Two sandstone beds are affected by soft-sediment deformation structures. The first deformed bed has a lateral continuity of about 20–25 m and a thickness of 0.8–1.2 cm, while the second deformed bed is 25–30 cm-thick with a lateral continuity of 8–10 m. These deformed beds show gentle deformation of laminae, with small load casts (10–20 cm-wide) and some dish and flame structures (Figure 4F). The clay facies is mainly red to greenish, and contains parallel lamination and rare fossiliferous wood remains. The thickness of this facies ranges from 0.10 to 1 m. The conglomerate lithofacies occurs in a minor proportion. It is generally composed of red-brown quartzite and grey to white-grey quartz pebbles (Figure 4G), sub-rounded, of 0.3–4 cm in diameter, with a fine-to medium-grained, poorly sorted, sand and sandstone matrix. It is commonly developed as thin lags (0.05–0.30 m-thick) on erosional planes. The conglomerate and pebbly sand lithofacies generally occur at the base of fining-upward cycle (Figure 4H).

FIGURE 4. Sedimentary facies from the Samani Vertebrate-rich Sand Member. (A) fossiliferous (vertebrates), poorly-sorted and medium-to coarse-grained sand including theropod teeth (arrow) and indeterminate bone; (B) Parallel-laminated sandstone interbedded within poorly sorted sand; (C,D), Parallel-laminated sandstones overlying trough cross-bedded medium-to coarse sandstone; (E) Parallel-laminated, poorly sorted sandstones showing fining-up ward trend; (F) Fine-to medium-grained, reddish sandstone showing soft sediment deformation (flame structures, black arrow); (G) Fossiliferous matrix-supported, massive conglomerate. Note that the quartz and quartzite pebbles are usually rounded, black arrows indicate an indeterminate bone; (H) Pebbly sandstone lag a few centimetres in thickness, including turtle plate (arrow) and other vertebrate remains. Hammer is 30 cm-long.
4.2 Samani Sandstone Member
This member extends over more than 15 m in outcrop and mostly consists of fine-to medium-grained, moderately sorted, reddish sandstone displaying trough and tabular cross-bedding, climbing ripples and parallel laminations (Figures 5A–C). These latter are slightly inclined (∼5°) in some cases. They exhibit, internally, normal grading, characterised by coarse-grained sandstones at the base that grade into fine-grained sandstones topwards. Lags of granules, vertebrate remains (turtles, crocodiles and fishes) and very coarse sand can sometimes be observed interlayered with horizontal laminae. The basal part of some sandstone beds contains intra-formational, green mud clasts. Two very well-exposed, deformed horizons are observed in the middle and upper parts of the member (Figure 5D). These deformed horizons are continuous and extend laterally for several 10 m, allowing the observation of structures both in cross-section and planar view.

FIGURE 5. Outcrop pictures from the Samani Sandstone Member showing the main facies observed around Gara Samani. (A) Fine-to medium-grained, tabular cross-laminated sandstone; (B) Graded lag deposit (1) overlain by tabular cross-laminated fine sandstone (2); (C) Trough cross-stratified, fine-to medium-sandstone; (D) Tabular cross-stratified sandstone (1) affected by decimetre-scale soft-sediment deformation structures in the mid-part (2). Hammer is 30 cm-long.
The sandstone facies are interlayered with lenticular, reddish-coloured siltstone and clay, of a few centimetres to 1.50 m thick, and laterally persistent on 5–30 m (most commonly less than 20 m). Small-scale cross-laminae may be observed within siltstone beds. Less common, thin, massive, matrix-supported, conglomerate beds (0.05–0.10 m-thick), with sharp bases, are interbedded within the red clays and the sandstone beds. Pebbles (quartz and quartzite) are usually rounded to subrounded and ranging between 0.5 and 2 cm in size. The conglomerate matrix is mainly upward-fining, formed by medium-to coarse-grained sand and/or occasionally with fine-grained gravel (Figure 5B).
4.3 Méguidène Stratified Sand Member
Outcrops of the Méguidène Stratified Sand Member were examined near the Méguidène village, around Gara Samani and in the Northern Aoulef town (Southern Tademait Plateau, Figure 1A). This member is approximately 55 m-thick and mostly consists of finely laminated and well cross-stratified, loose and uncemented sands. Cross-stratification is defined by grain-size and typically dips at 20 to 30 (Figure 6), and bounded by low-angle, planar to undulating surfaces. These sands are organised in tabular, metric intervals, with fine-to medium-grained, well sorted, well rounded, and highly spherical grains. They are generally pale to yellowish in colour; white and grey grains are also common. In places, pin-stripe lamination is apparent, highlighted by silt or very fine-grained sand accumulated on the concave portions of the low angle inclined laminae. Centimetre-scale, soft-sediment, deformation structures and millimetre-scale, wavy laminations, interpreted as adhesion structures, were also identified (Figure 6E). In addition to the sand facies, this member is locally interlayered with centimetric-thick intervals of grey-green clays and muddy silt, bounded at their base by erosional surfaces that may eventually include basal lags composed of vertebrate-rich (teeth of Spinosaurus aegyptiacus and O. numida, scales of Bawitius sp. and indeterminate bones and teeth) and gravelly sand (Figure 6C). The middle part of the member comprises tabular, fine-to medium-grained, moderately sorted, reddish sandstone bodies (0.4–1 m-thick) with horizontal to subhorizontal laminations and ripple marks, and occur interbedded with sets of cross-stratified sand. It is also intercalated by green clay (1.5–6 m-thick) and green gypsum clay (2.5–4 m -thick).

FIGURE 6. Outcrop pictures from the Méguidène Stratified Sand Member. (A) Well-rounded and sorted, fine-to medium-grained, sands organised as large-scale cross-bedded sets; (B–E) Spectacular example of low-angle cross-stratification featuring well-developed pinstripe texture which has likely formed due to differential cementation of translatent strata. Soft deformation structures (sds) can be observed in sets (see arrows in figure D). Note the intercalation of green clay layers (white arrows) and lag deposits (black arrows) in figure B (A, D-E, from the Méguidène section; B, from the southern foot of the Tademaït Plateau). Hammer is 30 cm long.
5 Sedimentological Interpretation
Two sedimentary systems are recognised in the Gara Samani Formation, based on the sedimentary structure of individual facies, texture, faunal content, bed geometry and stratigraphic context: a fluvial and an aeolian system.
5.1 Fluvial System
The lower two members are characterised by the occurrence of medium-to coarse-grained, siliciclastic bodies arranged into cycles, each with a basal erosion surface, fining-upward grain-size trend and internally comprising facies with aquatic, semi-aquatic and terrestrial fauna, and unidirectional palaeocurrent structures, comprising inner unidirectional tractive structures. These characteristics indicate accumulation within stream channels (Miall, 1996; Scherer et al., 2007; Ghazi and Mountney, 2009). Moderately to poorly sorted sand and sandstones displaying horizontally to low-angle cross-stratification, trough-cross stratification and massive structure suggest oscillation between high- and low-flow conditions during fluvial channel filling (Miall, 1996). Abundance of rounded, frosted quartz grains in the lower member indicate a significant input of wind-blown sand, although much of the primary aeolian deposits may have been reworked by fluvial processes. Occurrences of many internal erosional planes and fining-upward cycles in the sand and sandstones bodies suggest that channels were filled by multiple flood events (George and Berry, 1993; Herries, 1993). Mudstone clay and silty clay deposits are evidence of deposition from suspension under calm water conditions. They are common products of decreasing flow velocity or channel abandonment (Miall, 1996). Different possible driving forces of the deformed beds include gravity acting on slopes, unequal loading, gravitational instabilities due to a reverse density gradient, shear by aqueous or other currents, and biological and chemical agents. Sedimentary (flood, rapid sedimentation) and seismic processes have been identified as potential triggers causing the studied soft-sediment deformation structures.
5.2 Aeolian (Dune and Interdune) System
The sedimentary structures and features described from the Méguidène Stratified Sand Member compare favourably to those observed in modern and ancient aeolian systems: dune deposits (dry aeolian system) interbedded with the volumetrically less significant interdune facies and sheet sand strata (wet and damp aeolian system). The development of texturally mature (well-rounded and sorted), non-cohesive (high primary porosity and permeability), fine-to medium grained, sandy deposits, with highly spherical grains, organised as large-scale cross-bedded sets of variable thickness (0.3–1.5 m), suggests that they are residual deposits from aeolian dunes (dry aeolian system) (Hunter, 1977; Kocurek and Dott, 1981; Ulicný, 2004; Scherer et al., 2007). The highly friable nature suggests low cementation, which is responsible for high visible porosity. Pinstripe lamination is common in modern and ancient aeolian dune sediments (Hunter, 1977; Kocurek and Dott, 1981; Fryberger and Schenk, 1988) and is interpreted to record textural segregation of grain size, which leads to differential cementation within or adjacent to the finest sediments because of higher fluid capillarity (Fryberger and Schenk, 1988). The deformed laminae may have been generated by fluidization from water escape (Doe and Dott, 1980), due to rapid saturation of dry sands. Based on similar observations from another aeolian succession, Allen (1970) and Mountney and Thompson (2002) concluded that deformed laminae indicate the escape of water and air in areas of high interstitial water pressure.
The presence of tabular sandstone bodies showing horizontal laminations and ripple marks that interlayer with dune cross-strata suggests that these sandstones can be interpreted as aeolian sand sheets, formed as a consequence of limited wind energy or sand availability precluding aeolian dune formation (e.g., Trewin, 1993; Veiga et al., 2002; Biswas, 2005). The green clays, intercalated with aeolian dune strata, indicate deposition from suspension during periodic flooding of the interdune area. The occurrence of gypsum clay at the upper part of the third member indicates arid to semi-arid conditions. The most likely depositional environment is a saline sabkha. In many ergs (sand sea), areas of interdune sabkhas occur where the water table intersects the interdune troughs (Glennie, 1970).
Damp interdunes developed in areas where the capillary fringe of the water table intercepted the interdune region, or where the interdune area became wet as a consequence of fluvial inundation, responsible to the deposit of thin and discontinuous vertebrate-rich layers (lag deposits) and, more frequently, green clay, indicating relatively wetter periods. These latter are characterised by an increase in rainfall and runoff, which generated periods of fluvial incursions into the interdune areas. Damp interdunes are typically thin and discontinuous (Talbot, 1985; Sweet, 1999). The vertical interlayering of thin fluvial deposits with thick dry dune sands suggests that the fluvial flood events were episodic and of short duration. The rare presence of damp interdune deposits indicates that the system was partly a damp aeolian system (sensu Kocurek and Havholm, 1993), and water table rise was an important factor promoting its preservation (Stokes 1968).
6 Palaeontological Data
6.1 “Fishes”
The only information available so far on the “fishes” (i.e., non-tetrapod Gnathostomata) from Gara Sanami are those provided in the faunal list of Broin et al. (1971). However, none of these attributions were given with descriptions and comments, and no specimens were figured. Broin et al. (1971) indicated the presence of teeth of Ceratodus tiguidensis and Ceratodus africanus; dorsal spines of selachian fishes (Hybodus?); skull bone fragments of a coelacanthid fish (Mawsonia?); skull bone fragments, teeth, scales, and vertebrae of Lepidotes sp.; and rostral teeth and vertebrae of Onchopristis numidus.
Several taxa are recognised herein, including members of Chondrichthyes, Actinopterygii and Sarcopterigii. Representatives of two lineages of Elasmobranchii (Chondrichthyes) are also identified: Hybodontiformes and Batoidea. Oral teeth of the hybodont Tribodus sp. (Distobatidae) are recognised: they are small (less than 5 mm) and show a massive crown, subhexagonal (Figures 7A–C) to subtriangular (Figures 7D–F) in occlusal view. A faint, low cusp is present. The ornamentation consists of longitudinal ridges on the lingual face and on the labial margin of the occlusal face. No transverse ridges are present. The root shows numerous small pores and a few larger foramina, with no open canal or groove on the basal face (anaulacorhize vascularization). Some fragmentary dorsal spines with longitudinal ridges on the lateral faces (similar to the morphotype originally described as “Hybodus aschersoni”; Stromer, 1927; Werner, 1989) are also tentatively assigned to Tribodus sp. (Figure 8A). Originally described from the Albian of Brazil, this genus also occurs in Albian–Cenomanian deposits of North Africa and France (Vullo and Néraudeau, 2008). In Algeria, this genus was reported (as Protohypolophytes sp.) from the Albian of the Tiout Formation (Landemaine, 1991; Vullo and Néraudeau, 2008).

FIGURE 7. Selection of “fish” microremains from the Samani Vertebrate-rich Sand Member. (A–C) (LGS-45), tooth of Tribodus sp. in labial, lingual and occlusal views. (D–F) (LGS-46), tooth of Tribodus sp. in labial, lingual and occlusal views. (G,H) (LGS-47), pharyngeal tooth of Ginglymodii (?Adrianaichthys sp.) in lateral and distal views. (I–K) (LGS-48), tooth of Calamopleurus africanus in lingual, mesiodistal and occlusal views. Scale bars equal 500 µm.

FIGURE 8. Selection of “fish” macroremains from the Samani Vertebrate-rich Sand Member (A–F and J–R) and the Méguidène Stratified Sand Member (G–I). (A) (LGS-1), cf. Tribodus sp., incomplete dorsal fin spine (“Hybodus aschersoni” dorsal fin spine morphotype) in left lateral view. (B,C) (LGS-2 and LGS-3), Distobatidae indet., incomplete dorsal fin spine (“Hybodus aegyptiacus” dorsal fin spine morphotype) in right lateral and anterior views. (D–F). Onchopristis numida, rostral teeth (D, LGS-4; E, LGS-5) in dorsoventral view, vertebra (F, LGS-6) in anteroposterior view. (G–I) (LGS-7, LGS-8 and LGS-9), Bawitius sp., scales of in lateral view. (J–L) (LGS-10 and LGS-11), Adrianaichthys sp., scales in lateral view. (M) (LGS-12), Obaichthys africanus, scale in lateral view. (N,O), C. africanus jaw fragment (N, LGS-13) in occlusal view and vertebra (O, LGS-14) in posterior view. (P) (LGS-15), Mawsoniidae indet., opercle? fragment in lateral view. (Q) (LGS-16), cf. Lavocatodus humei, tooth plate in occlusal view. (R) (LGS-17), Neoceratodus africanus, tooth plate in occlusal view.
A second dorsal spine morphotype (Figures 8B,C), originally described as “Asteracanthus aegyptiacus” (Stromer, 1927; Werner, 1989), indicates the presence of another hybodont shark taxon at Gara Samani. It is characterised by longitudinal rows of small tubercles on the lateral faces. This form is herein referred to an indeterminate, second distobatid species (i.e., Distobatidae indet).
Rostral denticles of O. numida (Batoidea: Onchopristidae) have a characteristic morphology (Villalobos-Segura et al., 2021), with a cap bearing longitudinal folds and an apical barb on the posterior face (Figures 8D,E). The peduncle is grooved. The material of O. numida from Gara Samani also includes some pieces of calcified cartilage and isolated vertebrae (Figure 8F); such large, well-mineralized vertebrae are referred to this taxon because 1) they are identical to those described from the Moroccan Kem Kem beds (Villalobos-Segura et al., 2021: fig. 15), and 2) the other selachians taxa (i.e., Hybodontiformes) present in the Gara Samani assemblage lack calcified vertebrae (Maisey, 1982). Onchopristis numida, representing one of the largest known sclerorhynchoid species, is a common component of the Albian–Cenomanian vertebrate faunas from North Africa (e.g., Ibrahim et al., 2020), including Algeria (Benyoucef et al., 2015).
One of the clades of Actinopterygii recognised in Gara Samani is Cladistia. It is represented by a single form, Bawitius sp. (Polypteridae). Ganoid scales of Bawitius are distinguished by an incomplete ornamentation consisting of irregular ridges and patches of ganoin (Grandstaff et al., 2012; Cavin et al., 2015; Meunier et al., 2016) (Figures 8G–I). Scales referred to this polypteriform genus were reported from the early Cenomanian of Algeria (Benyoucef et al., 2015). Bawitius occurs in the Albian–Cenomanian of North Africa (Grandstaff et al., 2012; Cavin et al., 2015), and has also been recently found in coeval deposits from northeastern Brazil (Veiga et al., 2019).
The most diverse clade of Actinopterygii in Gara Samani is Holostei. Three representatives are identified: Adrianaichthys sp. (Lepidotidae), Obaichthys africanus (Obaiichthyidae) and Calamopleurus africanus (Amiidae). Ganoid scales identified as Adrianaichthys sp. are large (up to 4 cm in length) and devoid of ornamentation (Meunier et al., 2016) (Figures 8J–L). In addition, a single pharyngeal tooth with a slightly compressed crown and a recurved apex (Figures 7G,H) can be assigned tentatively to this genus because such teeth are present in lepidotid ginglymodian fishes (e.g., Pouech et al., 2015: fig. 4j). Adrianaichthys has been originally described from the Kem Kem beds of Morocco, and material (oral teeth and scales) tentatively referred to this genus is known from the early Cenomanian of Algeria (Benyoucef et al., 2015).
A ganoid scale ornamented with anteroposteriorly oriented ridges is attributed to O. africanus. Among these parallel ridges, the medial one is larger and more salient posteriorly. This specimen unambiguously belongs to the mid-Cretaceous gar O. africanus (Figure 8M) (Grande, 2010; Meunier et al., 2016). Known from Africa and Southwestern Europe (see Cavin et al., 2015), this species was previously described from the early Cenomanian of Algeria (Benyoucef et al., 2015).
A jaw fragment showing six tooth sockets with radiating striae (Figure 8N) and a vertebra with preserved neural and haemal arches (Figure 8O) can be assigned to C. africanus, a large amiid known from the late Albian-Cenomanian of Morocco and Algeria (Forey and Grande, 1998; Benyoucef et al., 2015; Cavin et al., 2015). A triangular, labiolingually compressed tooth, crown with sharp carinae is herein referred to this species (Figures 7I–K).
Sarcopterygii are represented by both coelacanths (Actinistia: Mawsoniidae indet.) and lungfishes (Dipnoi: cf. Lavocatodus humei and Neoceratodus africanus). Some fragments of large cranial bones ornamented with coarse ridges (e.g., opercle; Figure 8P) are typical of the family Mawsoniidae, a group especially well represented in the Albian-Cenomanian of North Africa and South America (Yabumoto and Uyeno, 2005; Benyoucef et al., 2015; Fragoso et al., 2019). This material probably belongs to Mawsonia and/or Axelrodichthys, but due to its fragmentary nature, we prefer to refer it as Mawsoniidae indet.
A worn tooth plate showing an almost flat occlusal surface and four poorly salient crests (Figure 8Q) is assigned to cf. L. humei (Ceratodontidae), a species well distributed in the Upper Cretaceous of North Africa (Churcher and De Iuliis, 2001; Claeson et al., 2014; Fanti et al., 2016). A few tooth plates, clearly longer than wide and with up to seven salient and sharp crests (Figure 8R), are similar to those of N. africanus (Neoceratodontidae). This species commonly occurs in the Albian–Cenomanian of North Africa (e.g., Cavin et al., 2015), including Algeria (Benyoucef et al., 2015).
6.2 Turtles
Broin et al. (1971) reported the presence of remains of undetermined turtles in Gara Samani. Subsequently, Broin (1977) alluded to one of the turtle taxa from this locality as a form of indeterminate precise systematic attribution but corresponding to a “primitive trionychoid”. Fragments of plates from its carapace were figured (see plate VII Figures 9, 10, 11 in Broin, 1977). That taxon was referred to Araripemys sp. by Fuente and Lapparent de Broin (1997) and Lapparent de Broin (2000). Lapparent de Broin (2000) indicated that, in addition to an araripemydid, other lineages of Pelomedusoides were present in Gara Samani, being recognised as indeterminate forms of Podocnemidoidea, belonging to Bothremydidae and, with doubt, to Podocnemididae. However, these attributions were not justified.

FIGURE 9. Selection of turtle remains from the Samani Vertebrate-rich Sand Member. (A–B) (LGS-28), left ilium of Pan-Pleurodira, in anterior (A) and posterior (B) views. (C–L), carapacial elements of Pelomedusoides. (C–D) LGS-29), neural, in dorsal (A) and ventral (B) views. (E–F) LGS-30), first right peripheral, in ventral (E) and dorsal (F) views. (G–H) LGS-31), proximal region of a costal, in dorsal (G) and ventral (H) views. (I–J) (LGS-32), costal, in dorsal (I) and ventral (J) views. (K–L) (LGS-33), partial right hypoplastron, in ventral (K) and dorsal (L) views. (M–R), carapacial elements of Araripemydidae. M-N (LGS-34), proximal region of a costal, in dorsal (M) and ventral (N) views. O-P (LGS-35), proximal region of a costal, in dorsal (O) and ventral (P) views. Q-R (LGS-36), distal region of a costal, in ventral (Q) and dorsal (R) views. Abbreviations (for the scutes): Ab, abdominal; Fe, femoral; M, marginal; Pl, pleural; V, vertebral.

FIGURE 10. Selection of Crocodyliformes remains (all of them attributed to Neosuchia indet.) from the Samani Vertebrate-rich Sand Member of the Samani Formation. (A) (LGS-18), posterior part of a left surangular and angular in lateral view. (B–C) (LGS-19), dentary and splenial fragment in lateral (B) and dorsal (C) views. (D) (LGS-20), fragment of a left angular in lateral view. (E–F) (LGS-21), osteoderm in dorsal (E) and (F) ventral views. (G–H) (LGS-22), osteoderm in dorsal (G) and (H) ventral views. (I-J) (LGS-23), osteoderm in dorsal (I) and (J) ventral views. (K–L) (LGS-24), osteoderm in dorsal (K) and (L) ventral views. (M) (LGS-25), osteoderm in dorsal view. (N) (LGS-26), osteoderm in dorsal view. (O–Q) (LGS-27), tooth in lingual (O), mesial or distal (P) and labial (Q) views.

FIGURE 11. Selection of pterosaur and dinosaur remains from the Samani Vertebrate-rich Sand Member of the Samani Formation. A-F, Pterosaur teeth, attributed to Ornithocheiridae indet. (A–C) (LGS-37), tooth in labial (A), lingual (B) and mesial (C) views. (D–F) (LGS-38), tooth in labial (A), lingual (B) and distal (C) views. G-W, Dinosaur cranial and postcranial remains. G-N, Theropod teeth. (G–H) (LGS-39), Spinosaurus aegyptiacus tooth in labial and lingual (G,I), and distal (H) views. (J–L) (LGS-40), Spinosaurus aegyptiacus tooth in labial and lingual (J, L), and distal (K) views. (M–N) (LGS-41), Carcharodontosaurus tooth in labial and distal views. (O,P) (LGS-42), Titanosauriformes indet. tooth in mesial or distal (O) and lingual (P) views. (Q–U) (LGS-43), centrum of a mid-cervical vertebra of Spinosaurus aegyptiacus in dorsal (Q), left lateral (R), ventral (S), anterior (T), and posterior (U) views. (V,W) (LGS-44), dentary and splenial fragment of Spinosaurus aegyptiacus in dorsal (V) and lateral (W) views.
All turtles so far identified in the African “Continental Intercalary” series are members of Pan-Pleurodira (Lapparent de Broin, 2000), and belong to Pelomedusoides (Gaffney et al., 2006; Gaffney et al., 2011). The greatest diversity of turtles reported for the ‘Continental Intercalary’ series, recognised in the Kem Kem Formation, is composed by representatives of Podocnemidinura (Hamadachelys escuilliei), Araripemydidae (lacking an infrafamiliary attribution), Euraxemydidae (Dirqadim schaefferi), and Bothremydidae (Galianemys whitei and Galianemys emringeri) (Gaffney et al., 2006; Gaffney et al., 2011; Ibrahim et al., 2020).
A partial illium with a well-developed dorsal margin, which was sutured to the carapace, is identified among the material recently found in Gara Samini (Figure 9A). These characters are compatible with those of the ilia of the pan-pleurodiran members, but not with those of Pan-Cryptodira.
Several partial carapacial plates with numerous small circular depressions on their outer surface are recognised among the new turtle material found at Gara Samani (Figures 9M–R). This allows its attribution to Araripemydidae, its presence in this locality being confirmed. Although the material from this lineage of turtles from Gara Samani was identified as attributable to an indeterminate form belonging to Araripemys (see Fuente and Lapparent de Broin, 1997; Lapparent de Broin, 2000), this genus is currently identified as exclusive to South America (Gaffney et al., 2006). A single form is currently identified for the African record, Taquetochelys decorata, from the Aptian (Lower Cretaceous) of Gadoufaoua (central Niger) (Broin, 1980; Pérez-García, 2019). The predominant presence of depressions instead of tubercles in the Gara Samani material, noticeably larger than the tubercles observed in the Gadoufaoua taxon, shows that it corresponds to a different form. Information currently available from the fragmentary remains of Gara Samani does not allow a more precise systematic attribution than Araripemydidae indet.
The most abundant plates correspond to a form that lacks such a marked ornamental pattern, the outer surface being recognised as slightly rough. Various elements of the carapace are identified, such as neurals (Figures 9C,D), peripherals (Figures 9E,F) or costals (Figures 9G–J), but also plastral remains (Figures 9K–L). Given the limitation of relevant characters from a systematic point of view in these remains, they are preliminarily recognised as Pelomedusoides indet. Its attribution to one or more forms cannot be determined.
6.3 Crocodyliforms
The mid-Cretaceous fauna of Crocodyliformes from North-Africa is represented by members of Notosuchia and Neosuchia (Sereno and Larsson, 2009; Ibrahim et al., 2020). The notosuchians diversity is composed by the peirosaurids Hamadasuchus rebouli Buffetaut, 1994 (Larsson and Sidor, 1999; Larsson and Sues, 2007) and Antaeusuchus taouzensis Nicholl et al., 2021; the uruguaysuchid Araripesuchus rattoides Sereno and Larsson, 2009 (Ibrahim et al., 2020); and the candidodontid Lavocatchampsa sigogneaurussellae Martin and Lapparent de Broin, 2016.
Neosuchians are represented by the pholidosaurs Elosuchus cherifiensis (Lavocat, 1955) (Lapparent de Broin, 2002; Meunier and Larsson, 2016) and Elosuchus broinae Meunier and Larsson, 2016 (Lapparent de Broin, 2002), the stomatosuchid Laganosuchus maghrebensis (Sereno and Larsson, 2009) and the giant Eusuchia Aegisuchus witmeri Holliday and Gardner, 2012.
The first crocodyliform remains found at Gara Samani were teeth, skin plates, skull and jaw fragments, vertebrae, a complete mandibular ramus, and a complete skull, which were attributed to, at least two different species (Broin et al., 1971). Among these earliest remains there is an almost complete skull of a longirrostral crocodyliform that is assigned to Elosuchus cherifiensis (Lapparent de Broin, 2002) and subsequently segregated to E. broinae (Meunier and Larsson, 2016).
The set of recently recovered material is made up of very fragmentary cranial fragments, some mandibular elements, teeth and osteoderms (Figure 10). Teeth attributed to crocodiles are conical, labially curved, with ornamented enamel and mesiodistal carinae (Figures 10O–Q). These teeth are similar to those attributed to Elosuchus in Gara Samani by de Lapparent de Broin (2002). Mandibular elements are represented by a small fragment of the central section of the mandible involving the dentary and splenial (Figures 10B). The lateral wall of the dentary is deep and presents an ornamentation composed of irregular pits. The fragment preserves two alveoli with sectioned teeth at the level of the base of the crown that present a circular section. The splenial is part of the lingual wall of the alveoli, so it must be representing a posterior section, but not so much so that the external mandibular fenestra is involved laterally. The contour of the alveoli is raised, generating a deep festooning between them. There are two sections of the posterior part of the mandible (Figures 10A,D) that allow recognition of an elongated external mandibular fenestra under which the dentary projects posteriorly. The surangular is part of the craniomandibular joint. The posterior extension of the surangular portion indicates that the retroarticular process may be relatively short and only slightly projected dorsally. Different morphologies of osteoderms (Figures 10E–N) have been identified that could belong to one or several forms. In general, they all present ornamentation made up of pits. Some osteoderms that are narrower than long, are attributed to the lateral ranges of the dermal armor. The osteoderms that could belong to the parasagittal plastron of the dorsal armor are subquadrate, with a smooth bar on its anterior edge and a marked longitudinal keel slightly displaced laterally. It is not possible to establish a precise specific attribution of these elements but, in general, they are compatible with the forms of Neosuchia described in the Kem-Kem Formation.
In conclusion, the Crocodyliformes recognised in Gara Samani belong to at least one form of Neosuchia, E. broinae. The fragmentary material recognized so far may be compatible with E. broinae or represent a broader diversity of neosuchian crocodilyforms yet to be defined.
6.4 Pterosaurs
So far, the presence of pterosaur remains had not been reported in Gara Samani. A few pterosaur teeth show a curved, slender crown with thin enamel covering the apical portion of the labial and lingual faces and descending along the mesial and distal carinae (Figures 11A–F). Some parallel longitudinal folds, more or less marked, are present on the lingual face. These teeth are indistinguishable from ornithocheirid specimens described from the Albian–Cenomanian deposits of North Africa (e.g., Wellnhofer and Buffetaut, 1999; Ibrahim et al., 2020), being identified as Ornithocheiridae indet.
6.5 Sauropod Dinosaurs
Sauropod remains from the “mid”-Cretaceous of Africa can be attributed to rebbachisaurid or titanosauriform sauropods (Mannion and Barrett, 2013; Holwerda et al., 2018). Only some of the teeth identified in Gara Samani can be attributed to sauropods (Broin et al., 1971). These teeth have parallel-sided crowns, subcylindrical cross section with slight labiolingual compression and presence of mesial and distal carinae can be preliminarly assigned to indeterminate Titanosauriformes (Figures 11O–P).
6.6 Theropod Dinosaurs
First remains attributed to theropod dinosaurs from Gara Samani were elements of the skull, jaw, teeth, vertebrae, fragments of limb bones (particularly metapods or unguals) attributed to a large form and a small one (Broin et al., 1971). Taquet and Russell (1998) assigned a rostrum with both premaxillae, maxillae, vomers, fragments of a right dentary, a fragment of premaxilla, two vertebral centra from cervical vertebrae, and a neural arch of a dorsal vertebra to Spinosaurus maroccanus. Taxonomy of “mid”-Cretaceous spinosaurs from North Africa is complex, with up to three species described: Spinosaurus aegyptiacus Stromer, 1915, Spinosaurus maroccanus Taquet and Russell, 1998 and Sigilmassasaurus brevicollis Russell, 1996. The presence of diagnostic characters in Spinosaurus maroccanus and Sigilmassasaurus brevicollis has been considered by different authors (McFeeters et al., 2013; Evers et al., 2015) and discussed by others (Ibrahim et al., 2014; Smyth et al., 2020) that conclude that Spinosaurus maroccanus and Sigilmassasaurus brevicollis both fall into the intraspecific variation of Spinosaurus aegyptiacus.
A centrum of a mid-cervical vertebra (Figures 11Q–U) from Gara Samani is herein attributed to Spinosaurus aegyptiacus. The centrum is strongly opisthocoelus. The attachment surface for the neural arch pedicles on the centrum has a marked hourglass outline. From the posteroventral surface of each parapophysis, a ridge extends posteriorly, converging toward the midline at the central part of the centrum. Posteriorly, the ventral surface is marked by two lateral ridges that define a flat platform, anteriorly these posterolateral ridges form two strong tubercles that converge towards the midline in the middle of the centrum as occurs in Spinosaurus aegyptiacus (Smyth et al., 2020). A posterior fragment of an articulated splenial and dentary (Figures 11V–W) can be also attributed to Spinosaurus aegyptiacus from the morphology of the remains of teeth preserved inside the alveoli.
Teeth relatively large, conical, with subcircular cross-sections, and with unserrated mesial and distal carinae are attributed to spinosaurine theropods (Figures 11G–L). The only spinosaurine that presents a similar dentition in the North-African context is Spinosaurus aegyptiacus, which is also consistent with the presence of axial elements attributable to this species in this locality. Teeth with these characteristics from Gara Samani present variations in the enamel texture and ornamentation, and in the density and extension of the longitudinal ridges. Without more information these differences are considered derived from the variability in the dentition of Spinosaurus aegyptiacus.
Some large and curved crowns (Figures 11M–N), with strong basal labiolingual compression and mesiodistal serrated carinae, are common in teeth in the Northern Saharan context and are generally attributed to the carcharodontosaurid C. saharicus. Usually, these teeth present enamel wrinkles flanking serrated carinae, but this feature was not recognised in any teeth of the Gara Samani locality. However, similar characteristics are observed in abelisaurid teeth (Fanti et al., 2014) and, subsequently, the presence of teeth attributable to the latter taxon cannot be ruled out.
7 Conclusion
Through the Late Cretaceous, the southern shore of the Tethys Ocean migrated north and south over short distances. These vicissitudes are documented in the ‘‘Continental Intercalaire”, a long series of mainly non-marine sediments deposited in which vertebrate fossils are common across southern Algeria (North Africa). The ‘‘Continental Intercalaire’’ deposits of Gara Samani (South Algeria) provided new insights into the litho- and biostratigraphy, as well as the palaeo-environmental evolution of the region.
The studied Gara Samani Formation is subdivided into three formal members: the Samani Vertebrate-rich Sand Member, the Samani Sandstone Member and the Méguidène Stratified Sand Member. The studied members show no real evidence of marine influence, and all indicate continental depositional systems. The two lower members are characterised by the occurrence of fining-upward, medium-to coarse-grained, siliciclastic bodies internally comprising facies with aquatic, semi-aquatic and terrestrial fauna, and indicate deposition within large stream channels. The sedimentary structures and features described from the third member compare favourably to those observed in modern and ancient aeolian systems: dune deposits (dry aeolian system) interbedded with interdune facies and sheet sand strata (wet and damp aeolian system).
The vertebrate fossil remains discovered at the Gara Samani site provide new data about the diversification of compositional vertebrate assemblage of the upper part of the ‘‘Continental Intercalaire’’ complex. The vertebrates recognised within are: elasmobranchs (Tribodus sp., Distobatidae indet. and O. numida), actinopterygians (Bawitius sp., Adrianaichthys sp., O. africanus and C. africanus), sarcopterygians (Mawsoniidae indet., cf. L. humei and N. africanus), turtles (at least two Pelomedusoides, one of them being a representative of Araripemydidae), crocodyliforms (E. broinae), ornithocheirid pterosaurs, and non-avian dinosaurs (titanosauriform sauropods, Spinosaurus aegyptiacus and C. saharicus). The similarity of the taxic composition of the vertebrate assemblage of the Gara Samani Formation with the “Kem Kem compound” assemblage and with the Bahariya assemblage led to consider the “Continental Intercalaire” in this part of Algeria to be early Cenomanian, and definitely not pre-Cenomanian an had been previously proposed.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Funding
This work was supported by the Scientific Research and Technological Development General Directorate “DGRSDT” (Algeria).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors are also grateful to Meksen Naima, Traoua Abdelhakiúand Rezkellah Walid for their valuable help in the field.
References
Allen, J. R. L. (1970). A Quantitative Model of Grain Size and Sedimentary Structures in Lateral Deposits. Geol. J. 7, 126–146. doi:10.1002/gj.3350070108
Bassoullet, J. P., and Iliou, J. (1967). Découverte de dinosaures associés à des crocodiliens et des poissons dans le Crétacé inférieur de l’Atlas saharien (Algérie). Comptes-rendus sommaires séances Société géologique Fr. 7, 294–295.
Benyoucef, M., Adaci, M., Meister, C., L Lang, E., Malti, F., Mebarki, K., et al. (2014). Le “Continental Intercalaire” dans la région du Guir (Algérie): nouvelles données sédimentologiques ichnologiques et paléontologiques. Rev. Paléobiologie 33, 281–297.
Benyoucef, M., Läng, E., Cavin, L., Mebarki, K., Adaci, M., and Bensalah, M. (2015). Overabundance of Piscivorous Dinosaurs (Theropoda: Spinosauridae) in the Mid-cretaceous of North Africa: The Algerian Dilemma. Cretac. Res. 55, 44–55. doi:10.1016/j.cretres.2015.02.002
Benyoucef, M., Mebarki, K., Ferré, B., Adaci, M., Bulot, L. G., Desmares, D., et al. (2017). Litho- and Biostratigraphy, Facies Patterns and Depositional Sequences of the Cenomanian-Turonian Deposits in the Ksour Mountains (Saharan Atlas, Algeria). Cretac. Res. 78, 34–55. doi:10.1016/j.cretres.2017.05.013
Benyoucef, M., Meister, C., Mebarki, K., Lang, E., Adaci, M., Cavin, L., et al. (2016). Évolution lithostratigraphique, paléoenvironnementale et séquentielle du Cénomano-Turonien dans la région du Guir (Ouest algéien). Carnets Géologie 16 (9), 271–295. doi:10.4267/2042/59926
Biswas, A. (2005). Coarse Aeolianites: Sand Sheets and Zibar-Interzibar Facies from the Mesoproterozoic Cuddapah Basin, India. Sediment. Geol. 174, 149–160. doi:10.1016/j.sedgeo.2004.11.005
Broin, F. de., Grenot, C., and Vernet, R. (1971). Sur la découverte d’un nouveau gisement de vertébrés dans le Continental Intercalaire Saharien: La Gara Samani (Algérie). Comptes Rendus l’Académie Sci. Paris272, 1219–1221.
Broin, F. de. (1977). Contribution á l’étude des Chéloniens. Chéloniens continentaux du Crétacé et du Tertiaire de France. Memoires Du. Muséum Natl. d’Histoire Nat. Paris 38 (1-IX), 1–366.
Broin, F. de. (1980). Les Tortues de Gadoufaoua (Aptien du Niger); aperçu sur la paléobiogéographie des Pelomedusidae (Pleurodira). Memoire Société Géologique Fr. 139, 39–46.
Buffetaut, É. (1994). A New Crocodilian from the Cretaceous of Southern Morocco. Comptes Rendus l’Académie Sci. Paris 319 (2), 1563–1568.
Buffetaut, E., and Ouaja, M. (2002). A New Specimen of Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with Remarks on the Evolutionary History of the Spinosauridae. Bull. Société Géologique Fr. 173 (5), 415–421. doi:10.2113/173.5.415
Candeiro, C. R. A., Fanti, F., Therrien, F., and Lamanna, M. C. (2011). Continental Fossil Vertebrates from the Mid-cretaceous (Albian-Cenomanian) Alcântara Formation, Brazil, and Their Relationship with Contemporaneous Faunas from North Africa. J. Afr. Earth Sci. 60, 79–92. doi:10.1016/j.jafrearsci.2011.02.004
Cavin, L., Boudad, L., Tong, H., Läng, E., Tabouelle, J., and Vullo, R. (2015). Taxonomic Composition and Trophic Structure of the Continental Bony Fish Assemblage from the Early Late Cretaceous of Southeastern Morocco. PLoS ONE 10, e0125786. doi:10.1371/journal.pone.0125786
Cavin, L., Tong, H., Boudad, L., Meister, C., Piuz, A., Tabouelle, J., et al. (2010). Vertebrate Assemblages from the Early Late Cretaceous of Southeastern Morocco: An Overview. J. Afr. Earth Sci. 57, 391–412. doi:10.1016/j.jafrearsci.2009.12.007
Churcher, C. S., and De Iuliis, G. (2001). A New Species of Protopterus and a Revision of Ceratodus Humei (Dipnoi: Ceratodontiformes) from the Late Cretaceous Mut Formation of Eastern Dakhleh Oasis, Western Desert of Egypt. Palaeontology 44 (2), 305–323. doi:10.1111/1475-4983.00181
Claeson, K. M., Sallam, H. M., O’Connor, P. M., and Sertich, J. J. W. (2014). A Revision of the Upper Cretaceous Lepidosirenid Lungfishes from the Quseir Formation, Western Desert, Central Egypt. J. Vertebrate Paleontology 34 (4), 760–766. doi:10.1080/02724634.2014.838574
Dal Sasso, C., Maganuco, S., Buffetaut, E., and Mendez, M. A. (2005). New Information on the Skull of the Enigmatic Theropod Spinosaurus, with Remarks on its Size and Affinities. J. Vertebrate Paleontology 25, 888–896. doi:10.1671/0272-4634(2005)025[0888:niotso]2.0.co;2
de Lapparent de Broin, F. (2002). Elosuchus, a New Genus of Crocodile from the Cretaceous of the North of Africa. Comptes Rendus Palevol 1 (5), 275–285. doi:10.1016/s1631-0683(02)00049-0
Debachi, F. (2007). Évolution géodynamique de la partie sud occidentale du bassin de Timimoun au cours du Carbonifère : Stratigraphie, Sédimentologie et Évolution Structurale. Thèse de Magister, FSTGAT, USTHB, Alger, 131.
Doe, T. W., and Dott, R. H. (1980). Genetic Significance of Deformed Cross-Bedding—With Examples from the Navajo and Webber Sandstones of Utah. J. Sediment. Petrology 50, 793–812. doi:10.1306/212f7aef-2b24-11d7-8648000102c1865d
Dubar, G. (1948). Carte géologique provisoire du Haut Atlas de Midelt au 1/200.000. Notice explicative. Notes et Mémoires du Service Géologique du Maroc, Rabat, 59 bis.
Evers, S. W., Rauhut, O. W. M., Milner, A. C., McFeeters, B., and Allain, R. (2015). A Reappraisal of the Morphology and Systematic Position of the Theropod dinosaurSigilmassasaurusfrom the "middle" Cretaceous of Morocco. PeerJ 3, e1323. doi:10.7717/peerj.1323
Fanti, F., Cau, A., Martinelli, A., and Contessi, M. (2014). Integrating Palaeoecology and Morphology in Theropod Diversity Estimation: A Case from the Aptian-Albian of Tunisia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 410, 39–57. doi:10.1016/j.palaeo.2014.05.033
Fanti, F., Larocca Conte, G., Angelicola, L., and Cau, A. (2016). Why So Many Dipnoans? A Multidisciplinary Approach on the Lower Cretaceous Lungfish Record from Tunisia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 449, 255–265. doi:10.1016/j.palaeo.2016.02.024
Forey, P. L., and Grande, L. (1998). An African Twin to the Brazilian Calamopleurus (Actinopterygii: Amiidae). Zoological J. Linn. Soc. 123 (2), 179–195. doi:10.1111/j.1096-3642.1998.tb01299.x
Foureau, F. (1904). Découverte de gîtes fossilifères dans le Djoua, à l'est de Timassânine (Sahara). Comptes rendus Hebd. séances l’Académie Sci. 138, 1527–1529.
Fragoso, L. G. C., Brito, P., and Yabumoto, Y. (2019). Axelrodichthys Araripensis Maisey, 1986 Revisited. Hist. Biol. 31 (10), 1350–1372. doi:10.1080/08912963.2018.1454443
Fryberger, S. G., and Schenk, C. J. (1988). Pin Stripe Lamination: A Distinctive Feature of Modern and Ancient Eolian Sediments. Sediment. Geol. 55, 1–15. doi:10.1016/0037-0738(88)90087-5
Fuente, M. d. l., and Lapparent de Broin, F. d. (1997). An Araripemys-like Decorated Pleurodire Turtle in the Paleocene of Northwestern Argentine. Geobios 30, 235–242. doi:10.1016/s0016-6995(97)80231-8
Gabani, A., Mammeri, C., Adaci, M., Bensalah, M., and Mahboubi, M. (2016). Le Créetacé continental à vertébrés de la bordure Sud du Plateau de Tinrhert : considéraions stratigraphiques et bilan paléontologique. Mémoire Serv. Géologique l’Algérie 19, 39–61.
Gaffney, E. S., Tong, H., and Meylan, P. A. (2006). Evolution of the Side-Necked Turtles: the Families Bothremydidae, Euraxemydidae, and Araripemydidae. Bull. Am. Mus. Nat. Hist. 300, 1–698. doi:10.1206/0003-0090(2006)300[1:eotstt]2.0.co;2
Gaffney, E. S., Meylan, P. A., Wood, R. C., Simons, E., and De Almeida Campos, D. (2011). Evolution of the Side-Necked Turtles: the Family Podocnemididae. Bull. Am. Mus. Nat. Hist. 350, 1–237. doi:10.1206/350.1
George, G. T., and Berry, J. K. (1993). “A New Lithostratigraphy and Depositional Model for the Upper Rotliegend of the UK Sector of the Southern North Sea,” in Characterization of Fluvial and Aeolian Reservoirs. Editors C. P. North, and D. J. Prosser (London: Geological Society, London, Special Publication), 73, 291–319. doi:10.1144/gsl.sp.1993.073.01.18
Ghazi, S., and Mountney, N. P. (2009). Facies and Architectural Element Analysis of a Meandering Fluvial Succession: The Permian Warchha Sandstone, Salt Range, Pakistan. Sediment. Geol. 221, 99–126. doi:10.1016/j.sedgeo.2009.08.002
Glennie, K. W. (1970). Desert Sedimentary Environments. Dev. Sedimentology 14, 222. doi:10.1016/s0070-4571(08)x7043-8
Grande, L. (2010). An Empirical Synthetic Pattern Study of Gars (Lepisosteiformes) and Closely Related Species, Based Mostly on Skeletal Anatomy. Resurrect. holostei 6, 1–871. American Society of Ichthyologists and Herpetologists, Special Publication.
Grandstaff, B. S., Smith, J. B., Lamanna, M. C., Lacovara, K. J., and Abdel-Ghani, M. S. (2012). Bawitius, Gen. nov., a Giant Polypterid (Osteichthyes, Actinopterygii) from the Upper Cretaceous Bahariya Formation of Egypt. J. Vertebrate Paleontology 32 (1), 17–26. doi:10.1080/02724634.2012.626823
Haug, E. (1904). Sur la faune des couches à Ceratodus crétacées du Djoua, près de Timassânine (Sahara). Comptes Rendus Hebd. Séances l’Académie Sci. Paris 138, 1529–1531. doi:10.5962/bhl.part.24288
Haug, E. (1905). “VIII Paléontologie,” in Documents scientifiques de la Mission Saharienne, Mission Foureau-Lamy d’Alger au Congo par le Tchad. Editor F. Foureau (Paris: Publication de la Société de Géographie), 139, 751–832.
Herries, R. D. (1993). “Contrasting Styles of Fluvial-Aeolian Interaction at a Downwind Erg Margin: Jurassic Kayenta-Navajo Transition, Northeastern Arizona, USA,” in Geological Society, London, Special Publications. Editors C. P. North, and J. D. Prosser, 73, 199–218. doi:10.1144/gsl.sp.1993.073.01.12
Holliday, C. M., and Gardner, N. M. (2012). A New Eusuchian Crocodyliform with Novel Cranial Integument and its Significance for the Origin and Evolution of Crocodylia. PLoS ONE 7 (1), e30471. doi:10.1371/journal.pone.0030471
Holwerda, F. M., Díez Díaz, V., Blanco, A., Montie, R., and Reumer, J. W. F. (2018). Late Cretaceous Sauropod Tooth Morphotypes May Provide Supporting Evidence for Faunal Connections between North Africa and Southern Europe. PeerJ 6, e5925. doi:10.7717/peerj.5925
Hunter, R. E. (1977). Basic Types of Stratification in Small Eolian Dunes. Sedimentology 24, 361–387. doi:10.1111/j.1365-3091.1977.tb00128.x
Ibrahim, N., Sereno, P. C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D. M., et al. (2014). Semiaquatic Adaptations in a Giant Predatory Dinosaur. Science 345, 1613–1616. doi:10.1126/science.1258750
Ibrahim, N., Sereno, P. C., Varricchio, D. J., Martill, D. M., Dutheil, D. B., Unwin, D. M., et al. (2020). Geology and Paleontology of the Upper Cretaceous Kem Kem Group of Eastern Morocco. Zookeys 928, 1–216. doi:10.3897/zookeys.928.47517
Kilian, C. (1931). Des principaux complexes continentaux du Sahara. Comptes-rendus sommaires séances Société Géologique Fr. 9, 109–111.
Kilian, C. (1937). Esquisse géologique du Sahara français à l’est du 6ème degré de longitude. Chronique des Mines et de la Colonization 58, 21–22.
Kocurek, G., and Dott, R. H. (1981). Distinctions and Uses of Stratification Types in the Interpretation of Eolian Sand. J. Sediment. Petro. 51, 79–95.
Kocurek, G., and Havholm, K. G. (1993). “Eolian Sequence Stratigraphy: A Conceptual Framework,” in Siliciclastic sequence stratigraphy. Editor P. Weimer, and H. W. Posamentier (American Association of Petroleum Geologists, Memoir) 58, 393–409.
Landemaine, O. (1991). Sélaciens nouveaux du Crétacé supérieur du Sud-Ouest de la France. Quelques apports à la systématique des élasmobranches. Saga 1, 1–45.
Lapparent, A. F. de., and Lelubre, M. (1948). Interprétation stratigraphique des séries continentales entre Ohanet et Bourarhet (Sahara Central). Comptes Rendus l’Académie Sci. Paris 227, 1106–1108.
Lapparent, J. de. (1937). Les Argiles d'El Goléa, Sahara Algérien : Étude Pétrographique. Bull. Serv. Carte Géogr. l'Algérie 8, 1–27.
Lapparent, A. F. de. (1947). Stratigraphie et âge du continental Intercalaire dans le Gourara, le Touat et le Tidikelt (Sahara algérien). Comptes Rendus Hebd. séances l’Académie Sci. Paris 225, 754–756.
Lapparent, A. F. de. (1951). Découverte de Dinosauriens, associés à une faune de Reptiles et de Poissons, dans le Crétacé inférieur de l’Extrême Sud tunisien. Comptes Rendus Hebd. Séances l’Académie Sci. Paris 232, 1430–1432.
Lapparent, A. F. de. (1960). Les Dinosauriens du “Continental Intercalaire” du Sahara central. Mémoires Société géologique Fr. Nouv. Série) 88, 1–56.
Lapparent de Broin, F. de. (2000). African Chelonians from the Jurassic to the Present: Phases of Development and Preliminary Catalogue of the Fossil Record. Palaeontol. Afr. 36, 43–82.
Larsson, H. C. E., and Sidor, C. A. (1999). Unusual Crocodyliform Teeth from the Late Cretaceous (Cenomanian) of Southeastern Morocco. J. Vertebrate Paleontology 19 (2), 398–401. doi:10.1080/02724634.1999.10011152
Larsson, H. C. E., and Sues, H.-D. (2007). Cranial Osteology and Phylogenetic Relationships of Hamadasuchus Rebouli (Crocodyliformes: Mesoeucrocodylia) from the Cretaceous of Morocco. Zoological J. Linn. Soc. 149 (4), 533–567. doi:10.1111/j.1096-3642.2007.00271.x
Lavocat, R. (1955). Découverte d'un crocodilien du genre Thoracosaurus dans le Crétacé supérieur d'Afrique. Bull. Muséum Natl. d'Histoire Nat. Paris 27, 338–340.
Le Loeuff, J., L., Lang, E., Cavin, L., and Buffetaut, E. (2012). Between Tendaguru and Bahariya: on the Age of the Early Cretaceous Dinosaurs Sites from the Continental Intercalaire and Other African Formations. J. Stratigr. 36, 1–18.
Lefranc, J. P. (1974). “Carte géologique de l'Algérie,” in Feuille Timimoun (NH 31 SO) au 1/500 000 Edit. Comptes Rendus de l'Académie des Sciences de Paris et Service Géologique de l’Algérie. Editors J. Conrad, and J. Ph. Lefanc (Alger.
Lefranc, J. P. (1983). Corrélations vers le nord et des cription stratigraphique détaillée du Continental Intercalaire (Mésozoïque Continental) de la Sebkha de Timimoun, Gourara, Sahara algérien. Comptes Rendus l'Académie Sci. Paris 269, 193–196.
Lefranc, J. P., and Guiraud, R. (1990). The Continental Intercalaire of Northwestern Sahara and its Equivalents in the Neighbouring Regions. J. Afr. Earth Sci. (and the Middle East) 10, 27–77. doi:10.1016/0899-5362(90)90047-i
Lefranc, J. P., and Toutin, N. (1971). Un gisement de Desertella foureaui (Lamellibranches préhétérodonte) dans le soubassement albien du Tademaït ouest (Sahara algérien). Bulletin de Société d'Histoire Naturelle de l'Afrique du Nord 62, 103–111.
Mannion, P. D., and Barrett, P. M. (2013). Additions to the Sauropod Dinosaur Fauna of the Cenomanian (Early Late Cretaceous) Kem Kem Beds of Morocco: Palaeobiogeographical Implications of the Mid-cretaceous African Sauropod Fossil Record. Cretaceous Research 45, 49–59. doi:10.1016/j.cretres.2013.07.007
Martin, J. E., and De Lapparent De Broin, F. (2016). A Miniature Notosuchian with Multicuspid Teeth from the Cretaceous of Morocco. Journal of Vertebrate Paleontology 36 (6), e1211534. doi:10.1080/02724634.2016.1211534
McFeeters, B., Ryan, M. J., Hinic-Frlog, S., Schröder-Adams, C., and Sues, H.-D. (2013). A Reevaluation of Sigilmassasaurus Brevicollis (Dinosauria) from the Cretaceous of Morocco. Can. J. Earth Sci. 50, 636–649. doi:10.1139/cjes-2012-0129
Meunier, F. J., Eustache, R.-P., Dutheil, D., and Cavin, L. (2016). Histology of Ganoid Scales from the Early Late Cretaceous of the Kem Kem Beds, SE Morocco: Systematic and Evolutionary Implications. Cybium 40 (2), 121–132.
Meunier, L. M. V., and Larsson, H. C. E. (2016). Revision and Phylogenetic Affinities of Elosuchus (Crocodyliformes). Zoological Journal of the Linnean Society 179, 169–200. doi:10.1111/zoj.12448
Meyendorff, A. (1938). Les couches de passage du Dévonien au Carbonifère dans le Gourara. (Sahara Occidental). Comptes Rendus de l'Académie des Sciences de Paris 209, 228–229.
Miall, A. D. (1996). The Geology of Fluvial Deposits (Sedimentrary Facies. Basin Analysis, and Petroleum Geology). Springer 565.
Mountney, N. P., and Thompson, D. B. (2002). Stratigraphic Evolution and Preservation of Aeolian Dune and Damp/Wet Interdune Strata: An Example From the Triassic Helsby Sandstone Formation, Cheshire Basin, UK. Sedimentology 49, 805–834.
Nedjari, A., and Ait Ouali, R. (2018). Le Gourara – Timimoun : de la synéclise hercynienne atypique aux continentaux. Mémoire du Service Géologique de l’Algerie 20, 3–49.
Nicholl, C. S. C., Hunt, E. S. E., Ouarhache, D., and Mannion, P. D. (2021). A Second Peirosaurid Crocodyliform from the Mid-cretaceous Kem Kem Group of Morocco and the Diversity of Gondwanan Notosuchians outside South America. Royal Society Open Science 8, 254. doi:10.1098/rsos.211254
Pérez-García, A. (2019). Identification of the Lower Cretaceous Pleurodiran Turtle Taquetochelys Decorata as the Only African Araripemydid Species. Comptes Rendus Palevol 18, 24–32. doi:10.1016/j.crpv.2018.04.004
Rage, J.-C., and Cappetta, H. (2002). Vertebrates from the Cenomanian, and the Geological Age of the Draa Ubari Fauna (Libya). Annales de Paléontologie 88, 79–84. doi:10.1016/s0753-3969(02)01040-6
Russell, D. A. (1996). Isolated Dinosaur Bones from the Middle Cretaceous of the Tafilalt, Morocco. Bulletin du Muséum National d’Histoire Naturelle, 4ème Série – Section C – Sciences de La Terre, Paléontologie, Géologie, Minéralogie 18 (2–3), 349–402.
Scherer, C. M. S., Lavina, E. L. C., Dias Filho, D. C., Oliveira, F. O., Bongiolo, D. E., and Aguiar, E. S. (2007). Stratigraphy and Fácies Architeture of the Fluvial-Aeolian-Lacustrine Sergi Formation (Upper Jurassic), Recôncavo Basin. Brazilian Journal of Geology 194, 169–193. doi:10.1016/j.sedgeo.2006.06.002
Sereno, P. C., Dutheil, D. B., Iarochene, M., Larsson, H. C. E., Lyon, G. H., Magwene, P. M., et al. (1996). Predatory Dinosaurs from the Sahara and Late Cretaceous Faunal Differentiation. Science 272, 986–991. doi:10.1126/science.272.5264.986
Sereno, P., and Larsson, H. C. E. (2009). Cretaceous Crocodyliforms from the Sahara. ZooKeys 28, 1–143. doi:10.3897/zookeys.28.325
Smyth, R. S. H., Ibrahim, N., and Martill, D. M. (2020). Sigilmassasaurus Is Spinosaurus: A Reappraisal of African Spinosaurines. Cretaceous Research 114, 104520. doi:10.1016/j.cretres.2020.104520
Stokes, W. M. L. (1968). Multiple Parallel-Truncation Bedding Planes – a Feature of Wind-Deposited Sandstone Formations. Journal of Sedimentary Petrology 38, 510–515. doi:10.1306/74D719D3-2B21-11D7-8648000102C1865D
Stromer, E. (1915). Ergebnisse der Forschungsreisen Prof. E. Stromer in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharîje-Stufe (unterstes Cenoman). 3. Das Original des Theropoden Spinosaurus aegyptiacus nov. gen., nov. spec. Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften. Mathematisch-Physikalische Klasse 28, 1–32.
Stromer, E. (1927). Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (Unterstes Cenoman). 9. Die Plagiostomen mit einem Anhang über Käno- und mesozoische Rückenflossenstacheln von Elasmobranchiern. Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften. Math. Naturwiss. Abt., N. F. 31 (5), 1–64.
Sweet, M. L. (1999). Interaction between Aeolian, Fluvial and Playa Environments in the Permian Upper Rotliegend Group, UK Southern North Sea. Sedimentology 46, 171–187. doi:10.1046/j.1365-3091.1999.00211.x
Tabaste, N. (1963). Étude de restes de poissons du Crétacé saharien. Mémoires de l'Institut Français d'Afrique Noire Mélanges ichthyologiques 68, 436–485.
Talbot, M. R. (1985). Major Bounding Surfaces in Aeolian Sandstones – a Climatic Model. Sedimentology 32, 257–265. doi:10.1111/j.1365-3091.1985.tb00508.x
Taquet, P. (1997). “Au temps des crocodiles mésozoïques sahariens,” in Hommage à Théodore Monod naturaliste d’exception. Editors R. Billard, and I. Jarry (Paris: Muséum National d’Histoire naturelle, Archives), 57–72. ISBN 2, 85653-509-7. doi:10.4000/books.mnhn.1593
Taquet, P., and Russell, D. A. (1998). New Data on Spinosaurid Dinosaur from the Early Cretaceous of the Sahara. Comptes Rendus de l'Académie des Sciences de Paris 327, 347–353. doi:10.1016/s1251-8050(98)80054-2
Taquet, P. (1972). Un exemple de datation et de corrélation stratigraphique basé sur les Captorhinomorphes (Reptiles Cotylosauriens). Bureau de Recherches Géologiques et Minières 77, 407–409.
Trewin, N. H. (1993). “Mixed Aeolian Sandsheet and Fluvial Deposits in the Tumblagooda Sandstone, Western Australia,” in Characterization of Fluvial and Aeolian Reservoirs. Editors C. P. North, and D. J. Prosser (London: Geological Society, London, Special Publication), 73, 219–230. doi:10.1144/gsl.sp.1993.073.01.13
Ulicný, D. (2004). A Drying-Upward Aeolian System of the Bohdasin Formation (Early Triassic), Sudetes of NE Czech Republic: Record of Seasonality and Longterm Paleoclimate Change. Sedimentary Geology 167, 17–39. doi:10.1016/j.sedgeo.2004.01.014
Veiga, G. D., Spalletti, L. A., and Flint, S. (2002). Aeolian-fluvial Interactions and Highresolution Sequence Stratigraphy of a Non-marine Lowstand Wedge: the Avilé Member of the Agrio Formation (Lower Cretaceous), Central Neuquén Basin, Argentina. Sedimentology 49 (5), 1001–1019. doi:10.1046/j.1365-3091.2002.00487.x
Villalobos-Segura, E., Kriwet, J., Vullo, R., Stumpf, S., Ward, D. J., and Underwood, C. J. (2021). The Skeletal Remains of the Euryhaline Sclerorhynchoid †Onchopristis (Elasmobranchii) from the ‘Mid’-Cretaceous and Their Palaeontological Implications. Zoological Journal of the Linnean Society 193 (2), 746–771. doi:10.1093/zoolinnean/zlaa166
Vullo, R., and Néraudeau, D. (2008). When the “Primitive” Shark Tribodus (Hybodontiformes) Meets the “Modern” Ray Pseudohypolophus (Rajiformes): the Unique Co-occurrence of These Two Durophagous Cretaceous Selachians in Charentes (SW France). Acta Geologica Polonica 58 (2), 249–255.
Wellnhofer, P., and Buffetaut, E. (1999). Pterosaur Remains from the Cretaceous of Morocco. Paläontologische Zeitschrift 73 (1), 133–142. doi:10.1007/bf02987987
Werner, C. (1989). Die Elasmobranchier-Fauna des Gebel Dist Member der Bahariya Formation (Obercenoman) der Oase Bahariya, Agypten. Palaeo Ichthyologica 5, 1–112.
Keywords: Africa, Algeria, Cenomanian, “Continental Intercalaire”, Gara Samani, vertebrates, diversity
Citation: Benyoucef M, Pérez-García A, Bendella M, Ortega F, Vullo R, Bouchemla I and Ferré B (2022) The “mid”-Cretaceous (Lower Cenomanian) Continental Vertebrates of Gara Samani, Algeria. Sedimentological Framework and Palaeodiversity. Front. Earth Sci. 10:927059. doi: 10.3389/feart.2022.927059
Received: 23 April 2022; Accepted: 10 June 2022;
Published: 06 July 2022.
Edited by:
Nour-Eddine Jalil, National Museum of Natural History (France), FranceReviewed by:
Brian Lee Beatty, New York Institute of Technology, United StatesMichael David Gottfried, Michigan State University, United States
Sebastian Apesteguia, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina
Copyright © 2022 Benyoucef, Pérez-García, Bendella, Ortega, Vullo, Bouchemla and Ferré. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Madani Benyoucef, YmVueW91Y2VmbWFkYUBnbWFpbC5jb20=