 Alison M. Murray
Alison M. Murray
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci. , 25 April 2022
Sec. Paleontology
Volume 10 - 2022 | https://doi.org/10.3389/feart.2022.892301
This article is part of the Research Topic African Vertebrates Through Times View all 5 articles
Cichlid fishes are an important component of the African freshwater ecosystem and the evolution of this group has captured the interest of ichthyologists for many decades. The distribution of cichlid fishes in both Africa and South America, and the apparent reciprocal monophyly of the cichlids in the two areas, has led to differences in opinion as to whether their modern distribution reflects a dispersal event from Africa to South America or was the result of a vicariant event–the break-up of the two continental masses in the mid-Cretaceous. If the vicariant event were the cause of the modern distribution, this would indicate an age for the family of at least 120 million years; however, the fossil record indicates a younger, Cenozoic, age for the family. The known fossil taxa in Africa often are difficult to assign to extant tribes which creates inaccuracies when they are used to date lineages within Cichlidae. The difficulty of assigning fossil cichlids to modern lineages is caused by most of these lineages being recognized based on DNA or soft-tissue characters, which are generally not preserved in fossils. This hampers our understanding of the early history of the family. Despite this, new African cichlid fossils that have been described in the past two decades and a new technique to determine relationships of these fossils are beginning to elucidate the early history of the family in Africa. Here the Palaeogene (Eocene and Oligocene) cichlids of Africa are reviewed and a new taxon from the Oligocene of Somalia, Somalichromis hadrocephalus gen. et sp. nov. is described.
Just over two decades ago, the oldest fossil cichlids were described from the Eocene of Africa (Murray, 2000). The Eocene African cichlids were joined a few years later by Eocene fossil cichlids from South America (Malabarba et al., 2006; Malabarba et al., 2010; Malabarba et al., 2014). These Eocene records remain the oldest known cichlids. The age of the oldest fossils, the distribution of cichlid fishes in both Africa and South America and the apparent reciprocal monophyly of the cichlids in the two areas, has led to disagreement among researchers as to whether the distribution of modern cichlids reflects a Cenozoic dispersal event from Africa to South America (e.g., Murray, 2001) or is the result of a vicariant event resulting from the break-up of the two continental masses in the mid-Cretaceous (e.g., Sparks, 2004; Sparks and Smith, 2004). If the vicariant event were the cause of the modern distribution, this would indicate the family must have arisen prior to the complete separation of the continents 119–105 million years ago (McLoughlin, 2001) and would give a minimum age of Aptian–Albian (late Early Cretaceous) for the family Cichlidae. However, the fossil record indicates a younger age for the family, with a suggested origin of cichlids in the early Cenozoic (Murray, 2001). This disparity in proposed ages of Cichlidae has been explored several times in the past two decades (e.g., Genner et al., 2007; Friedman et al., 2013; Schedel, 2020). In the more recent studies, the ages based on the fossil record have been supported by molecular phylogenetic analyses and an early Cenozoic age for the family is corroborated by nuclear DNA (Matschiner et al., 2017; Matschiner, 2019). This emphasizes the importance of understanding the earliest records of cichlid fishes, from the Eocene and Oligocene.
If the Cenozoic age for the family is correct, then the separation of the continents at that time indicates that the African cichlids evolved in Africa independently from their South American relatives, although both continental cichlid faunas were present at the latest by the Eocene (Murray 2000; Malabarba et al., 2006; Malabarba et al., 2010; Malabarba et al., 2014). Part of the difficulty in determining the origins of the Cichlidae are that the early fossil forms from Africa seem to be of generalized (primitive) form (pers. observ.) and have been difficult to assign to modern lineages below the level of subfamily. This contrasts with the South American fossils, which are considered to be relatively derived within the South American subfamily Cichlinae (Malabarba et al., 2014) with one of the Eocene forms having been placed in the extant genus Gymnogeophagus (Malabarba et al., 2010). Without a better understanding of the early African fossils, it remains difficult to elucidate the evolution and biogeography of the Cichlidae.
Since my review of African fossil cichlids (Murray, 2001), a number of important new African cichlid fossils have been described and named based on articulated material. Fossil cichlids can be difficult to assign to extant lineages because the extant forms are distinguished predominantly based on soft-tissue characters that do not preserve well (or at all) in the fossil record. A new method of assigning fossil cichlids to extant lineages, termed ‘the best fit approach’ was proposed by Penk et al. (2019) and has since been used by others (e.g., Altner et al., 2020; Přikryl et al., 2022) with some success. Here, a new species of cichlid is described from Oligocene deposits of the Daban Group of northern Somalia. This new species joins the Eocene Mahengechromis spp., and Oligocene Libyachromis fugacior and Macfadyena dabanensis as the only named Palaeogene African cichlids (Van Couvering, 1982; Murray, 2000; Přikryl et al., 2022). The new species is assessed using the methods of Penk et al. (2019) to determine the likely relationships.
The Daban Group was deposited during rifting and tectonic subsidence associated with the opening of the Gulf of Aden (Ali and Watts, 2015). The marine through continental deposits which filled the subsiding half graben are considered to be middle Eocene to Oligocene (Abbate et al., 1991) or Oligocene to Miocene (Ali and Watts, 2015) in age. The lower deposits represent deposition in a restricted marine lagoonal to deltaic environment, but these are overlaid by ephemeral to perennial lacustrine deposits (Abbate et al., 1991). It is from these latter that freshwater fossils were collected. Abbate et al. (1991) noted that the cichlids reported by Van Couvering (1982) came from the uppermost levels of the deposits, representing a perennial lake. The perennial lake deposits are reported as including green siltstones, sandstones, limestone and marlstone (Abbate et al., 1991) which matches the matrix of the cichlid Macfadyena dabanensis, described from the area (Van Couvering, 1982). These deposits grade into a thicker delta sequence which includes sandstones, siltstones and nodular fresh-water limestones (Abbate et al., 1991); these seem to better match the matrix of the new species described here. Although the Daban Group includes Eocene sediments, fossils recovered from the formation come from higher stratigraphic levels and are considered to be of Oligocene age (Ali and Watts, 2015). The sediment of the cichlids reported here matches that of the higher levels of the Daban Group, supporting their age as Oligocene, rather than Eocene.
The fossil specimens described here are in the collections of the Natural History Museum, London (NHM), catalogued under the prefix NHMUK P. Comparative material is stored in the collections of the University of Alberta Museum of Zoology (UAMZ). Comparative material is listed in Appendix A1. Photographs of the holotype material of Macfadyena dabanensis from presumed Oligocene deposits of Somalia were kindly provided by Matt Riley, Sedgwick Museum of Earth Sciences, University of Cambridge.
Following the methodology of Přikryl et al. (2022), and with the same expectation that a phylogenetic analysis would result in a very poorly resolved tree, the Somali cichlids were analysed using the “best-fit approach” of Penk et al. (2019) to determine the most-probable relationships of the Libyan fossil material. This method was devised specifically for fossil cichlids, to overcome issues that are associated with lack of soft-tissue preservation and consequent lack of data to include fossils in phylogenetic data sets of extant cichlids. Přikryl et al. (2022) used the meristic data of Penk et al. (2019) for the ‘haplotilapiines’ and added data for the Heterochromini, Tylochromini, Pelmatochromini, Etroplinae and Ptychochrominae, as well as named African Palaeogene (Eocene and Oligocene) cichlids (Přikryl et al., 2022; supplementary tables S3, S4). The “best-fit approach” uses features in combination to characterize tribes of cichlids, rather than relying on synapomorphies which are generally difficult to identify in fossil material. The new Somali species was compared to the tribes of African cichlids (Pseudocrenilabrinae), Madagascan cichlids (Ptychochrominae and Etroplinae), and African Palaeogene fossils in order to determine where it most likely belongs.
Subdivision Teleostei Müller, 1845 sensu Arratia, 1999
Order Cichliformes Betancur-R. et al., 2013
Family Cichlidae Bonaparte, 1835
Subfamily Pseudocrenilabrinae Fowler, 1934
Somalichromis gen. nov.
Type species: Somalichromis hadrocephalus sp. nov.
Etymology: The generic name is derived from the country of origin, Somalia, in combination with “chromis” the Greek root often used for cichlid fishes. Gender is masculine.
Diagnosis: as for type and only known species.
Somalichromis hadrocephalus sp. nov. (Figures 1–4).

FIGURE 1. Photograph of the holotype of Somalichromis hadrocephalus gen. et sp. nov., NHMUK P.29574. Scale bar equals 1 cm.
Holotype: A complete articulated fish preserved in left lateral view, specimen NHMUK P.29574 (Figure 1).
Paratypes: Several incomplete specimens are given paratype status. Specimen NHMUK P.29575: head and anterior portion of body preserved in left lateral view; NHMUK P.29579: portions of the head, dorsal fin, pectoral and pelvic girdles and body preserved on 4 pieces of matrix; NHMUK P29576: caudal skeleton and fin, pectoral and pelvic girdles on two pieces of rock; NHMUK P29577: a single piece of matrix preserving the ventral skull, pectoral and pelvic girdles; NHMUK P29578: the posteroventral part of the head and anterior portion of the body preserved on a single slab of matrix.
Type locality and age: The information on the catalogue labels indicates the specimens are from the Somaliland Protectorate (followed by the unknown designation M.739A.) and were given to the Natural History Museum by the Anglo-Saxon Petroleum Co., Ltd in September of 1952. The age and formation are given on the labels as “Eocene (?Middle), Middle Daban.” Based on this information, the cichlids are presumed to come from the upper (freshwater) sediments of the Daban Series and, therefore, are Oligocene in age (Abbate et al., 1991).
Etymology: The specific epithet is the combination of the Greek words “hadros” meaning bulky or stout, and “cephale” meaning head, to indicate the relatively large head compared to standard length of this taxon.
Diagnosis: Oligocene pseudocrenilabrine cichlid with three anal-fin spines, cycloid scales on body and head; head height equal to head length and both about 45% of standard length; distinguished from named tribes (some including fossil species) by having 14 dorsal-fin spines (unlike Etiini which has 15 dorsal-fin spines), two supraneurals (only a single supraneural or no supraneurals in Chromidotilapiini, Pelmatochromini, Coelotilapiini, Coptodonini, Heterotilapiini, Gobiocichlini, and Steatocranini; Tilapiini normally has one, rarely two supraneurals and Oreochromini has 1–2 supraneurals), unicuspid jaw and pharyngeal teeth (unlike the molariform teeth of Tylochromini or multi-cuspid teeth of many other cichlids).
The new species is also distinct from Palaeogene fossils left incertae sedis in the subfamily Pseudocrenilabrinae. It differs from the Eocene Mahengechromis by the lack of ctenoid scales (present on the body in Mahengechromis), from the Oligocene Libyachromis (which shares the above meristic features) by having a larger head in comparison to the body (head length 45% of standard length in the new species compared to just over 33% SL in Libyachromis); and from Macfadyena, which is also from the Daban Series of Somalia, by lacking granulations on the scale surface and lacking bicuspid teeth (both present in Macfadyena) and the presence of scales on the cheek (lacking in Macfadyena). It also differs from named Miocene cichlids in that bicuspid teeth are present in Palaeofulu, Kalyptochromis, Nderechromis and Tugenchromis, and tricuspid oral teeth are present in Rebekkachromis and Baringochromis; Warilochromis has unicuspid teeth but possesses enlarged canines in an outer row and smaller teeth in the inner row of the jaw bones (as well as a single supraneural and 33 vertebrae); in addition, Tugenchromis has three segments to the lateral line (only two in Somalichromis) and Baringochromis has only a single or no supraneurals (two in Somalichromis).
This is a fairly deep-bodied cichlid, with a body depth measured in front of the dorsal fin of 45% of standard length (SL). The head is round (i.e., the head height and length are roughly equal) and the head length (from the tip of the premaxilla to the posterior edge of the opercle) is about 45% SL. The holotype is the only complete specimen but the body depth has been distorted during preservation, with both pelvic girdles preserved in ventral view and body scales preserved ventral to the body limits between these fins and the anal fin (Figure 1). The amount of distortion indicates that the fish would not have been compressed in life, but would have had significantly more girth.
The skull of the holotype (Figure 2) is best preserved, but even in this specimen the bones have been displaced and not all are clear. The limits of the frontal, parietal, and pterotic cannot be determined. The lateral ethmoid is identifiable, but not well preserved, forming the anterior rim of the orbit. However, no details of any of these bones can be determined. The middle portion of the parasphenoid is also visible in the orbit; it is fairly deep and gently curves.
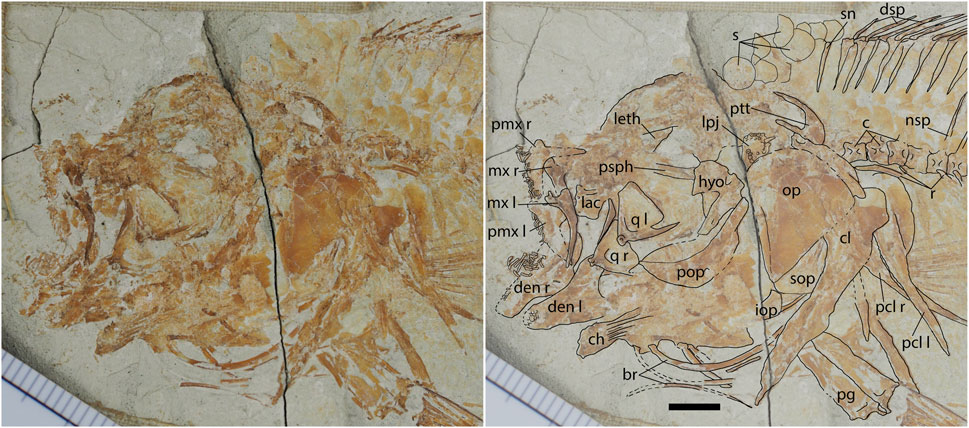
FIGURE 2. Photograph and interpretation of the head of the holotype of Somalichromis hadrocephalus gen. et sp. nov., NHMUK P.29574. Scale bar equals 5 mm.
The opercle is the largest bones of the cheek; it is rounded dorsally and comes to a point ventrally. The broad subopercle is visible along the posteroventral edge. Whether or not it has an anterodorsal projection as common in cichlids is not visible. Just anterior to the subopercle, the interopercle is partially visible but no details are clear.
The only element of the infraorbital series visible is the lacrimal. The number of pores on this bone cannot be determined. The bone is roughly square, being as deep as it is wide.
The left and right premaxillae are both visible, along with the left and right dentaries (Figure 2). All these bones bear small, unicuspid teeth of uniform size. The tips of at least some of these teeth are slightly flattened to be more spatulate than conical. The teeth form multiple rows on each bone, with sockets indicating at least four rows were present on the left dentary. The long ascending process of the premaxilla is not quite as long as the alveolar process; it is narrow and tapers gently. The alveolar process ends in a point. The maxilla is slender and curved. The dentary is deep posteriorly, and the length of the bone is only slightly shorter than the height. The anguloarticular cannot be distinguished.
The hyomandibula has a broad head with a slight notch in the dorsal edge, and an anterior flange. Both left and right quadrates are preserved. These are triangular in shape and have a deep cleft for articulation of the symplectic. The right ectopterygoid is preserved along the anterior edge of the right quadrate; the ectopterygoid is fairly narrow and angled in the middle. The left anterior ceratohyal is preserved; it narrows in the middle and becomes deeper posteriorly than it is anteriorly. Broad striations mark the middle, narrowed, portion of the bone. There are remains of five branchiostegal rays; these are fairly robust and long. The remains of the lower pharyngeal jaw (LPJ) are preserved, positioned at the anterodorsal corner of the opercle (Figure 2). The LPJ preserves several tooth sockets, indicating the bases of the teeth were round. It also preserves one unicuspid, conical tooth that seems slightly larger than the oral teeth.
There are 26 centra visible, including the terminal compound centrum, and an additional two centra are present based on the neural spines visible behind the skull bones at the back of the head. Therefore, there is a total of 28 vertebrae. Fifteen centra are anterior to the first anal-fin pterygiophore, and so are considered to be abdominal, although whether or not all of those centra bear ribs and not haemal spines cannot be determined. Thirteen centra are considered to be caudal. Only a single supraneural is clearly visible anterior to the dorsal-fin pterygiophores in the holotype (Figure 2), with bone fragments possibly of a second, but the paratype (NHMUK P.29575) clearly shows the presence of two supraneurals (Figure 3). The supraneurals are positioned upright and are slender with a flattened distal end tapering to a proximal point.

FIGURE 3. Photograph of the head of Somalichromis hadrocephalus gen. et sp. nov., paratype NHMUK P.29575. Scale bar equals 1 cm.
The preserved elements of the pectoral girdle are the cleithrum, supracleithrum, posttemporal, and postcleithra (Figure 2). The cleithrum has an expanded dorsal head which is blunt posteriorly. The supracleithrum is elongate, with a thicker anterior edge (where the sensory canal would be positioned) and thinner posterior flange along the entire length. The posttemporal has a long, narrow ventral process and an even longer dorsal process. The upper postcleithrum bears an enlarged dorsal plate, and it appears that there are two postcleithra on each side. The scapula and coracoid are not distinguishable. There are 10 pectoral-fin rays.
The pelvic girdle meets the pectoral girdle well below the level of the pectoral fin. The left and right pelvic bones meet along the midline. There is a small median projection on each half of the girdle in the midline. Remains of five rays and one spine are visible.
The dorsal fin is made up of 14 spines and ten rays (XIV, 10) and the anal fin has three spines and eight (holotype) or nine (NHMUK P.29575) rays (III, 8–9). The last two rays in both fins are counted separately although they share a single pterygiophore. There are 22 dorsal-fin pterygiophores and eight anal-fin pterygiophores. The caudal fin, as is normal in cichlids, has 16 branched rays for a fin-ray formula of I,8,8,I. All five hypurals appear to be separate elements, with no fusion in either fin lobe (Figure 4). The hypurapophysis is not distinguishable.

FIGURE 4. Photograph and interpretation of the caudal fin of Somalichromis hadrocephalus gen. et sp. nov. (A), holotype NHMUK P.29574; (B), paratype NHMUK P.29576.
Large cycloid scales cover the body (Figure 1); there are no sign of any ctenoid scales. Small cycloid scales also cover the cheek region, particularly clear overlying the quadrate in the holotype, but there are none visible on the opercle (Figure 2). Scales on the chest, ventral to the pectoral girdle and just above the pelvic girdle, are about two-thirds the size of the flank scales (NHMUK P. 29577, P.29575) There are 20 pored scales in the upper lateral line and perhaps eight in the lower lateral line. In total, there are about 30 scales in a single line along the flank from the opercle to the end of the hypural plates. Small scales continue onto the caudal fin with four in each row overlapping the proximal bases of the caudal-fin rays.
Macfadyena dabanensis was described from the Daban Series, from which Somalichromis hadrocephalus also comes. Macfadyena comes from lacustrine paper shales (Van Couvering, 1982) and the holotype is preserved in very fine fragile layers (M. Riley, pers. comm. 2018). This is quite unlike the preservation of the Somalichromis, which is preserved in much more consolidated sandstone. This may indicate that the two cichlids inhabited different environments. As noted in the diagnosis above, Somalichromis can be clearly distinguished from Macfadyena.
Somalichromis hadrocephalus exhibits a number of meristic features that allow its placement within Pseudocrenilabrinae to be assessed using the ‘best-fit approach’ of Penk et al. (2019). These are a dorsal-fin formula of XIV, 10 (which excludes it from Heterochromini, Coelotilapiini, Heterotilapiini, Pelmatotilapiini, Steatocranini, as well as Etroplinae); anal-fin formula of III, 8–9 (which again excludes it from Etroplinae, and Heterotilapiini, but also from Chromidotilapiini); 2 supraneural bones (which excludes it from Chromidotilapiini, Pelmatochromini, Coelotilapiini, Coptodonini, Heterotilapiini, Gobiocichlini, Pelmatotilapiini, Steatocranini; most Tilapiini also have a single supraneural, but occasionally some have two), and 28 (15 abdominal and 13 caudal) vertebral centra (which excludes it from subfamilies Etroplinae and Ptychochrominae, as well as the African tribes Heterochromini, Tylochromini, Etiini, Coelotilapiini, Coptodonini, Heterotilapiini, and Steatocranini. Taken in combination, these features exclude Somalichromis from belonging to any lineage except Oreochromini and Tilapiini, which are arguably more variable (in terms of meristic ranges) than any of the other lineages (features for each lineage are summarized in Přikryl et al., 2022; supplementary table S3).
Exclusion from Oreochromini or Tilapiini cannot be confirmed based on the meristic features above, because the greater range of counts in these two tribes encompasses the counts for Somalichromis, as they also do for Libyachromis (Přikryl et al., 2022). However, it is rare to have two supraneurals in these tribes (Dunz and Schliewen, 2013; Penk et al., 2019). Based on possession of bi- and tri-cuspid teeth in these groups, Somalichromis can be excluded from both. Oreochromini may be closely related to Tilapiini, or not (e.g., see Dunz and Schliewen, 2013:Figure 2), which indicates that more work is needed to understand these lineages.
Among named African fossil cichlids (see Přikryl et al., 2022; supplementary table S3), Somalichromis shares the presence of two supraneurals with Libyachromis, Macfadynena, Palaeofulu, Kalyptochromis and Rebekkachromis (this latter varies from 1–2 supraneurals present); the rest of the fossil taxa have a single supraneural (unknown in Tugenchromis). Rebekkachromis has higher caudal and total vertebral counts, Kalyptochromis has higher dorsal and anal fin-spine counts, and Tugenchromis has lower dorsal fin counts. In terms of these meristic features, Libyachromis, Macfadyena, and Palaeofulu are most similar to Somalichromis. Libyachromis has only recently been described, but Macfadyena and Palaeofulu were named over 40 years ago (Van Couvering, 1982) and should be reassessed in light of the vast amount of research that has been done on cichlids in the past several decades. (Planned work on Macfadyena was prevented by the global pandemic). As noted by Přikryl et al. (2022) for Libyachromis, features these fossil cichlids share (counts of vertebrae, fin spines, fin rays, and supraneurals), or features shared only by Somalichromis and Libyachromis (unicuspid teeth), are considered primitive for cichlids. Somalichromis therefore joins these other fossil forms in being a relatively basal form, probably placed close to the base of the Pseudocrenilabrinae.
Přikryl et al. (2022) recently assessed a number of named African cichlid fossils. They noted that among the Miocene taxa, Oreochromimos and Rebekkachromis represent Oreochromini, Tugenchromis is assigned to the base of the “East African Radiation” and Warilochromis and Baringochromis are members of the haplochromiine group (see also original information on these taxa from Altner et al., 2017, Altner et al., 2020; Altner and Reichenbacher 2020). Therefore, all of these lineages have a documented presence in the Miocene. Another Miocene fossil must also be mentioned, Oreochromis lorenzoi. This fossil is not from Africa, but instead was recovered from Messinian (latest Miocene) deposits of Italy (Carnevale et al., 2003). Oreochromis lorenzoi documents that the Oreochromini lineage had not only evolved, but had invaded other land masses by the late Miocene. Oreochromis is also represented in Africa in the late Miocene and Pliocene (Murray and Stewart, 1999).
None of the Oligocene African cichlids can be readily assigned to extant tribes, although all have been included in Pseudocrenilabrinae (as are all extant African cichlids). Libyachromis was noted as possibly belonging to Heterochromini or being a basal Pseudocrenilabrinae of uncertain position (Přikryl et al., 2022). Heterochromini is probably the most basal member of the Pseudocrenilabrinae (e.g., Farias et al., 2000; Keck and Hulsey, 2014). These Oligocene cichlids therefore provide little in the way of information on the age of the subfamily, but do indicate that the distribution of cichlids in Africa during the Oligocene was already quite extensive. Somalichromis joins Macfadyena in documenting cichlids in Somalia, East Africa, and Libyachromis documents cichlids in North Africa (Libya) in the Oligocene.
The oldest known African cichlids, five species of Mahengechromis, are more problematic in terms of relationships. These species were described at a time when our conceptions of cichlid relationships were quite different from now. Based on scale morphology, Mahengechromis was placed in an existing cladogram in which “tilapiines” (including oreochromines and others) were considered relatively primitive (Murray, 2001). Our understanding of “tilapiines” has changed greatly since then (Dunz and Schliewen, 2013). Additionally, the presence of ctenoid scales, considered to have been derived only once in the cladogram used for Mahengechromis, is now better documented as variable throughout Pseudocrenilabrinae. Mahengechromis is in need of re-examination and re-assessement, and this is a planned future project of another researcher (X. Kevrekidis pers. comm. January 2022).
The fossil record of cichlids in Africa includes articulated, reasonably well-preserved, skeletons from Eocene, Oligocene and Miocene deposits. The Miocene specimens have allowed some lineages of the subfamily Pseudocrenilabrinae to be confidently dated (Altner and Reichenbacher, 2015; Kevrekidis et al., 2019; Penk et al., 2019). In contrast, the Oligocene forms, Libyachromis and Somalichromis, appear to be more generalized and are placed towards the base of the African cichlid tree, as early members of the subfamily Pseudocrenilabrinae. The Eocene African cichlids, five species in the genus Mahengechromis, bear a mosaic of features, some that are considered more primitive (unicuspid teeth, general body form, vertebral counts, and fin-spine and ray counts) and others that may or may not be primitive for the family (ctenoid scales) or may be more derived features (single supraneural). In the past two decades, our knowledge of the fossil record of this evolutionarily very interesting family has increased, but the relationships among all these forms remain to be elucidated. The presence of generalized basal pseudocrenilabrines in the Oligocene suggests that during this time, all the different lineages within the subfamily had not yet diversified. This supports the idea of a Cenozoic, rather than mid-Cretaceous, age for cichlids with evolution of the African tribes in the later Palaeogene and Neogene.
The original contributions presented in the study are included in the article/supplementary materials, further inquiries can be directed to the corresponding author.
The author confirms being the sole contributor of this work and has approved it for publication.
This research was supported by the National Science and Engineering Research Council of Canada Discovery grant 327448.
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
My thanks to Emma Bernard, NHM London for lending specimens, and to Matt Riley, Sedgwick Museum of Earth Sciences, University of Cambridge for providing photographs of the holotype of Macfadyena dabensis. Thanks also to the reviewers, Giorgio Carnevale and Bouziane Khalloufi, and guest editor Nour-Eddine Jalil for helpful comments on the manuscript.
br, branchiostegal rays; c, centrum; cc, compound centrum; ch, anterior ceratohyal; cl, cleithrum; den, dentary; dsp, dorsal-fin spines; ep, epural; hs, haemal spine; hy (number), hypural (numbered); hyo, hyomandibula; iop, interopercle; l, left; lac, lacrimal; leth, lateral ethmoid; lpj, lower pharyngeal jaw (fused fifth ceratobranchials); mx, maxilla; nsp, neural spine; op, opercle; pcl, postcleithrum; pg, pelvic girdle; phy, parhypural; pmx, premaxilla; pop, preopercle; psph, parasphenoid; ptt, posttemporal; pu2 (pu3), second (third) preural centrum; q, quadrate; r, right; s, scales; sn, supraneural (predorsal); sop, subopercle; un, uroneural.
Abbate, E., Bruni, P., and Sagri, M. (1991). Development of a Lacustrine Terminal Fan and a Coarse Grained delta in the Tertiary Daban Basin (Northern Somalia). Cuadernos de Geología Ibérica 15, 279–298.
Ali, M. Y., and Watts, A. B. (2015). Tectonic Evolution of Sedimentary Basins of Northern Somalia. Basin Res. 28, 340–364. doi:10.1111/bre.12113
Altner, M., and Reichenbacher, B. (2015). A New Fossil Cichlid from the Middle Miocene in the East African Rift Valley (Tugen Hills, Central Kenya): First Record of a Putative Ectodini. Front. Mar. Sci. 2. doi:10.3389/conf.FMARS.2015.03.00145
Altner, M., and Reichenbacher, B. (2020). A Small Cichlid Species Flock from the Upper Miocene (9-10 MYA) of Central Kenya). Hydrobiologia, 848 3613–3637. doi:10.1007/s10750-020-04358-z
Altner, M., Ruthensteiner, B., and Reichenbacher, B. (2020). New Haplochromine Cichlid from the Upper Miocene (9-10 MYA) of Central Kenya. BMC Evol. Biol. 20 65 doi:10.1186/s12862-020-01602-x
Altner, M., Schliewen, U. K., Penk, S. B. R., and Reichenbacher, B. (2017). †Tugenchromis pickfordi, gen. et sp. nov., from the upper Miocene-a stem-group cichlid of the 'East African Radiation'. J. Vertebr. Paleontol. 37, e1297819. doi:10.1080/02724634.2017.1297819
Arratia, G. (1999). “The Monophyly of Teleostei and Stem-Group Teleosts. Consensus and Disagreements,” in Mesozoic Fishes 2 - Systematics and Fossil Record. Editors G. Arratia, and H.-P. Schultze (München: München Verlag, Dr. Friedrich Pfeil), 265–334.
Betancur-R, R., Broughton, R. E., Wiley, E. O., Carpenter, K., López, J. A., Li, C., et al. (2013). The Tree of Life and a New Classification of Bony Fishes. Plos Curr. 5, 1–45. doi:10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288
Bonaparte, C. L. (1835). Prodromus Systematis Ichthyologiae. Nuovi Annali delle Scienze naturali Bologna 2, 181–277.
Carnevale, G., Sorbini, C., and Landini, W. (2003). Oreochromis lorenzoi, a New Species of Tilapiine Cichlid from the Late Miocene of central Italy. J. Vertebr. Paleontol. 23, 508–516. doi:10.1671/1858
Dunz, A. R., and Schliewen, U. K. (2013). Molecular Phylogeny and Revised Classification of the Haplotilapiine Cichlid Fishes Formerly Referred to as “Tilapia”. Mol. Phylogenet. Evol. 68, 64–80. doi:10.1016/j.ympev.2013.03.015
Farias, I. P., Ortí, G., and Meyer, A. (2000). Total Evidence: Molecules, Morphology, and the Phylogenetics of Cichlid Fishes. J. Exp. Zool. 288, 76–92. doi:10.1002/(sici)1097-010x(20000415)288:1<76::aid-jez8>3.0.co;2-p
Fowler, H. W. (1934). Fishes Obtained by Mr. H.W. Bell-Marley Chiefly in Natal and Zululand in 1929 to 1932. Proc. Acad. Nat. Sci. Philadelphia 86, 405–514.
Friedman, M., Keck, B. P., Dornburg, A., Eytan, R. I., Martin, C. H., Hulsey, C. D., et al. (2013). Molecular and Fossil Evidence Place the Origin of Cichlid Fishes Long after Gondwanan Rifting. Proc. R. Soc. B. 280, 20131733. doi:10.1098/rspb.2013.1733
Genner, M. J., Seehausen, O., Lunt, D. H., Joyce, D. A., Shaw, P. W., Carvalho, G. R., et al. (2007). Age of Cichlids: New Dates for Ancient Lake Fish Radiations. Mol. Biol. Evol. 24, 1269–1282. doi:10.1093/molbev/msm050
Keck, B. P., and Hulsey, C. D. (2014). Continental Monophyly of Cichlid Fishes and the Phylogenetic Position of Heterochromis multidens. Mol. Phylogenet. Evol. 73, 53–59. doi:10.1016/j.ympev.2014.01.011
Kevrekidis, C., Valtl, M., Penk, S. B. R., Altner, M., and Reichenbacher, B. (2019). Rebekkachromis Nov. Gen. From the Middle-Upper Miocene (11 MYA) of Central Kenya: the Oldest Record of a Haplotilapiine Cichlid Fish. Hydrobiologia 832, 39–64. doi:10.1007/s10750-018-3715-8
Malabarba, M. C., Malabarba, L. R., and López-Fernández, H. (2014). On the Eocene Cichlids from the Lumbrera Formation: Additions and Implications for the Neotropical Ichthyofauna. J. Vertebr. Paleontol. 34, 49–58. doi:10.1080/02724634.2013.830021
Malabarba, M. C., Malabarba, L. R., and Papa, C. D. (2010). Gymnogeophagus eocenicus, N. Sp. (Perciformes: Cichlidae), an Eocene Cichlid from the Lumbrera Formation in Argentina. J. Vertebr. Paleontol. 30, 341–350. doi:10.1080/02724631003618348
Malabarba, M. C., Zuleta, O., and Papa, C. D. (2006). Proterocara argentina, a New Fossil Cichlid from the Lumbrera Formation, Eocene of Argentina. J. Vertebr. Paleontol. 26, 267–275. doi:10.1671/0272-4634(2006)26[267:paanfc]2.0.co;2
Matschiner, M. (2019). Gondwanan Vicariance or Trans-Atlantic Dispersal of Cichlid Fishes: a Review of the Molecular Evidence. Hydrobiologia 832, 9–37. doi:10.1007/s10750-018-3686-9
Matschiner, M., Musilová, Z., Barth, J. M. I., Starostová, Z., Salzburger, W., Steel, M., et al. (2017). Bayesian Phylogenetic Estimation of Clade Ages Supports Trans-Atlantic Dispersal of Cichlid Fishes. Syst. Biol. 66, syw076–22. doi:10.1093/sysbio/syw076
McLoughlin, S. (2001). The Breakup History of Gondwana and its Impact on Pre-cenozoic Floristic Provincialism. Aust. J. Bot. 49, 271–300. doi:10.1071/bt00023
Müller, J. (1845). Über den Bau und die Grenzen der Ganoiden, und über das natürliche System der Fische. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1845, 117–216.
Murray, A. M. (2000). Eocene Cichlid Fishes from Tanzania, East Africa. J. Vertebr. Paleontol. 20, 651–664. doi:10.1671/0272-4634(2000)020[0651:ECFFTE]2.0.CO;2
Murray, A. M., and Stewart, K. M. (1999). A New Species of Tilapiine Cichlid from the Pliocene, Middle Awash, Ethiopia. J. Vertebr. Paleontol. 19, 293–301. doi:10.1080/02724634.1999.10011142
Murray, A. (2001). The Fossil Record and Biogeography of the Cichlidae (Actinopterygii: Labroidei). Biol. J. Linn. Soc. 74, 517–532. doi:10.1006/bijl.2001.0599
Penk, S. B. R., Altner, M., F. Cerwenka, A., Schliewen, U. K., and Reichenbacher, B. (2019). New Fossil Cichlid from the Middle Miocene of East Africa Revealed as Oldest Known Member of the Oreochromini. Sci. Rep. 9, 10198. doi:10.1038/s41598-019-46392-5
Přikryl, T., Kaur, J., and Murray, A. M. (2022). New Oligocene Pseudocrenilabrinae Cichlid Fishes (Teleostei, Cichlidae) from Freshwater Deposits of Libya. J. Syst. Palaeontology 19, 1–24. doi:10.1080/14772019.2022.2033861
Schedel, F. D. B. (2020). Phylogeny, Divergence Time Estimates and Systematics of African Cichlids (Cichlidae: Pseudocrenilabrinae), with a Focus on the Rheophilic Cichlids of East and Central Africa (München: Faculty of Biology of the Ludwig-Maximilian University of Munich), 240. PhD dissertation.
Sparks, J. S. (2004). Molecular Phylogeny and Biogeography of the Malagasy and South Asian Cichlids (Teleostei: Perciformes: Cichlidae). Mol. Phylogenet. Evol. 30, 599–614. doi:10.1016/S1055-7903(03)00225-2
Sparks, J. S., and Smith, W. L. (2004). Phylogeny and Biogeography of Cichlid Fishes (Teleostei: Perciformes: Cichlidae). Cladistics 20, 501–517. doi:10.1111/j.1096-0031.2004.00038.x
Ptychochrominae
Paratilapia polleni UAMZ F8934, UAMZ F9052
Ptychochromis oligacanthus UAMZ F uncatalogued
Etroplinae
Paretroplus menarambo UAMZ F9051
Pseudocrenilabrinae
Heterochromini
Heterochromis multidens UAMZ F 8917
Tylochromini
Tylochromis sudanensis UAMZ F8937 and UAMZ F uncatalogued
Pelmatochromini
Pelvicachromis pulcher UAMZ F8915
Hemichromini
Hemichromis sp. UAMZ F8916
Hemichromis elongatus UAMZ F uncatalogued.
Keywords: Cichlidae, Daban series, East Africa, Oligocene, Somalia
Citation: Murray AM (2022) Cenozoic Cichlids of Africa (Cichlidae: Pseudocrenilabrinae) With the Description of a New Species From the Oligocene of Somalia. Front. Earth Sci. 10:892301. doi: 10.3389/feart.2022.892301
Received: 08 March 2022; Accepted: 31 March 2022;
Published: 25 April 2022.
Edited by:
Nour-Eddine Jalil, National Museum of Natural History, FranceReviewed by:
Giorgio Carnevale, University of Turin, ItalyCopyright © 2022 Murray. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alison M. Murray, YW1tdXJyYXlAdWFsYmVydGEuY2E=, b3JjaWQub3JnLzAwMDAtMDAwMS05NjQ4LTI5MDI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.