
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Earth Sci. , 12 May 2022
Sec. Paleontology
Volume 10 - 2022 | https://doi.org/10.3389/feart.2022.875001
This article is part of the Research Topic Co-Evolution of Life and Environment During the Ediacaran-Cambrian Transition View all 6 articles
 Anton Kolesnikov1,2*
Anton Kolesnikov1,2*The siliciclastic succession of the Ediacaran Chernyi Kamen Formation in the Central Urals of Russia contains a rich complex of Ediacaran fossils that represent in situ benthic populations buried in life position. The present work reports Beltanelliformis konovalovi sp. nov. found as a paleocommunity of organisms and studied for their morphology and spatial distribution. Unlike the type species (B. brunsae), new fossils are preserved within fine-grained sandstone beds as three-dimensional pancake-like or flattened discoidal single, paired, or occasionally aggregated bodies with an ultra-thin rim. Statistical analysis shows significant predominance of small individuals in the paleocommunity, which in turn manifests different spatial distribution in comparison with larger ones. In addition, the studied paleocommunity exhibits patterns of self-organization in extremely shallow marine environment and adaption to periodic desiccation. These results provide quantitative support for the more complex ecology of Beltanelliformis and also expand the range of taphonomic possibilities of macroscopic life in the terminal Neoproterozoic.
Discoidal fossils are the most widespread and abundant in Ediacaran deposits. They are mostly characterized by rounded cyclic structures and known from both siliciclastic and carbonate sedimentary rocks (Fedonkin et al., 2007). Among them, Beltanelliformis represents one of the most revisited discoidal fossils. It includes taxa Beltanelloides and Nemiana, which were recently interpreted as variants of taphonomic forms (Ivantsov et al., 2014). Fossils of Nemiana are presented by accumulations of convex tubercles preserved on the sole of sandstone beds (positive hyporelief) (Narbonne and Hofmann, 1987), whereas the Beltanelloides are commonly defined by almost flattened imprints or organically preserved film-like specimens within clay (Leonov, 2007). These taxa have been previously interpreted as abiological structures or fossils of jellyfishes (Zaika-Novatsky and Palij, 1974), eukaryotic algae (Gnilovskaya et al., 1988), fungal colonies (Aseeva, 1988), corals (Seilacher, 1992), resting traces (Runnegar and Fedonkin, 1992), benthic demosponges (Leonov, 2007), or colonial prokaryotes (Ivantsov et al., 2014). However, it has been recently demonstrated that the organisms related to Beltanelliformis from the Ediacaran of the White Sea area may have had a cyanobacterial origin (Bobrovskiy et al., 2018a). In this paper, the first results of study of the new Ediacaran taxon Beltanelliformis konovalovi sp. nov. from the Chernyi Kamen Formation of the Central Urals are presented. New results show that organisms whose nature seemed relatively simple actually exhibit patterns of more complex ecology, such as self-organization in environment transitional from marginal marine to non-marine and possible adaption to periodic desiccation.
The Sylvitsa Group exposes in the western slope of the Ural mountains, a folded rim of the East European Platform (Figure 1A). The fossiliferous sites are located in the Sverdlovsk region of Russia along the Sylvitsa River valley, a right tributary of the Chusovaya River (Figure 1B). It represents ca. 1800-m-thick aluminosiliciclastic succession, erosionally overlaying mixed carbonate-siliciclastic Serebryanka Group and unconformably overlapped by Devonian deposits. Recent detailed lithological studies yielded that the Sylvitsa Group can be divided into four sedimentary systems comprising seven lithofacies and characterizing a wide spectrum (offshore muddy plains, prodelta, retreating fans, and delta plain) of sedimentary environments (Grazhdankin et al., 2010). Zircons from volcanic tuffs in the lower part of the Perevalok Formation have an U-Pb age 567.2 ± 3.9 Ma (Grazhdankin et al., 2011), marking the maximum age of the unit. The overlying ca. 1,300-m-thick Chernyi Kamen Formation has volcanic tuffs in which the U-Pb zircon age 557 ± 13 Ma has been obtained (Ronkin et al., 2006). The Late Neoproterozoic age for the Sylvitsa Group is also established by abundant and exceptionally well-preserved fossils of Ediacara-type soft-bodied biota, Miaohe-type carbonaceous compressions, concentric microbial colonies, and arumberiamorph structures (Grazhdankin et al., 2010; Kolesnikov et al., 2017).
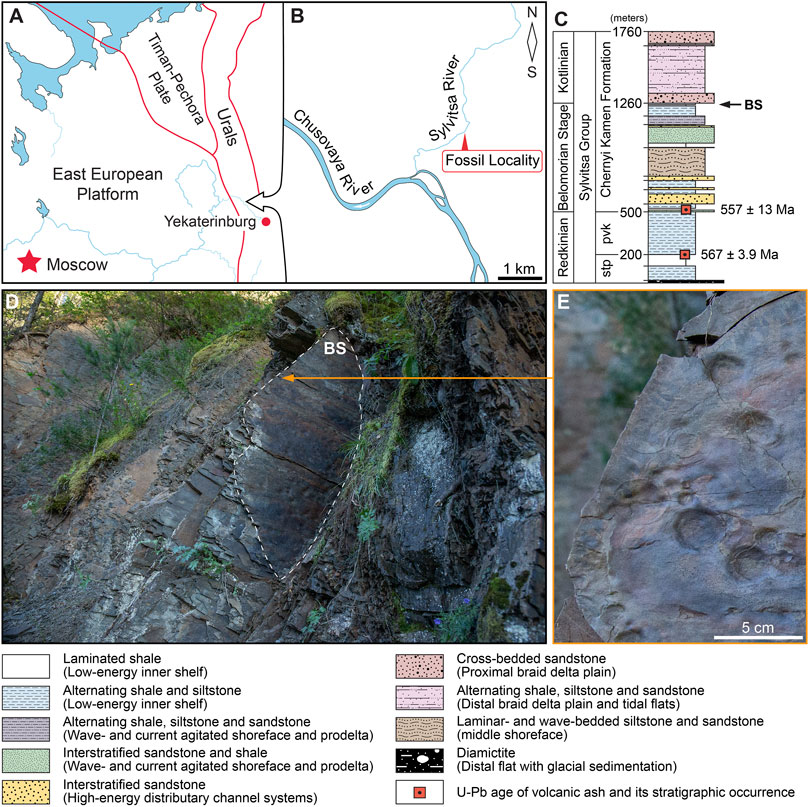
FIGURE 1. Fossil locality map and stratigraphic column: (A) Central Urals and its position on the East European Platform; (B) studied outcrop of the Chernyi Kamen Formation, lower reach of the Sylvitsa River, (Sverdlovsk region of Russia); (C) Sylvitsa Group section showing the position of the BS surface with Beltanelliformis konovalovi sp. nov. fossils; (D) field photograph of the BS surface brushed in the Konovalovka Member; (E) close-up field photograph of B. konovalovi sp. nov.
The Chernyi Kamen Formation consists of packages of interstratified sandstones and alternating shales and siltstones in the lower part (Figure 1C), thick packages of laminar- and wave-bedded fine-grained sandstones and interstratified sandstones and shales in the middle part, and upward the section grades into thick packages of cross-bedded sandstones and laminar alternating shales, siltstones, and sandstones. The fossiliferous stratum is a 10-mm-thick sandstone bed found in the uppermost part of the Konovalovka Member of the Chernyi Kamen Formation cropping out along the left bank of the lower reach of the Sylvitsa River. It lies within an interval of thin alternating laminar- and wave-rippled sandstones, siltstones, and shales ca. 750 m from the base of the formation. The sedimentary succession is considered transitional from marginal marine to extremely shallow as having been deposited in a tidal flat depositional system (Bobkov et al., 2019; Kolesnikov et al., 2012). U-Pb zircon dates in the Sylvitsa Group suggest that the Chernyi Kamen Formation may have been deposited coevally with the White Sea fossiliferous sedimentary succession (Fedonkin et al., 2007).
A total of 196 specimens were studied from the outcrop of the upper part of the Konovalovka Member (Figure 1E). The specimens were imaged using a Fujifilm GFX 50r digital mirrorless camera with a Fujinon GF 120 mm f/4.0 R LM OIS macro lens mounted on a Fujifilm MCEX-45G WR extension tube. A microscopic study involved observations of thin sections under an optical microscope Carl Zeiss Axiolab equipped with a digital camera AxioCam ERc5s. Thin sections were subjected also to analysis in a scanning electron microscope (SEM) Tescan VEGA3 to observe main taphonomic and morphological features in cross section with high resolution and energy-dispersive spectroscopy (EDS) to make elemental mapping, allowing for the characterization of the chemical composition of fossils.
To assess the interspecimen dynamics in paleocommunity, a detailed spatial point pattern analysis of the fossil-bearing surface is carried out. All specimens appear to be deformed in one direction as a result of tectonic activity and on the surface look like oval imprints. Assuming that the oval imprint before tectonic deformation may have had a circle outline, in the Chernyi Kamen Formation, the shortest diameters (Dmin) of oval-shaped specimens and the coordinates (X, Y) of their centers have been chosen. Spatial data were collected from the brushed BS surface using photo-mapping, with photographs captured under a light source at night. Measurements were obtained from digital photographs using Adobe Illustrator CC software. The spatial analysis was run in “R” software, version 4.0.3 (R Core Team 2020) and “RStudio,” version 1.4.1103 (RStudio, PBC 2021) using the packages “mclust” and “spatstat” (Fraley and Raftery, 2007; Baddeley et al., 2015).
Beltanelliformis konovalovi sp. nov. are found as an assemblage of macroscopic fossils preserved within the fine-grained sandstone bed and exposed on the BS surface (Figures 1D,E). Its brushed area is approximately 1.8 m2. The BS surface is 10 m higher Dickinsonia-bearing the DS surface, which was found earlier in this section (Bobkov et al., 2019; Mitchell et al., 2020) and 10 m lower base of the Krutikha Member of the Chernyi Kamen Formation. The BS surface is smooth, without any erosional or microbially induced sedimentary structures, and only specimens of B. konovalovi sp. nov. are found on it. This surface is sandwiched between alternating siltstone and sandstone beds with a thin biolaminated structure containing many casts of desiccation cracks, wrinkled textures, and halite pseudomorphs.
Petrographic sections (ca. 60 µm in thickness) demonstrate that the fossils and host rock both consist of fine sandstone grains. Furthermore, a thin section cut along the flattened fossil (Figure 2A) reveals a rim of thinly laminated sediment clearly visible around each specimen (Figure 2B). It displays that the fossils have three-dimensional style of preservation: the body has a pancake-like shape and its thickness can reach 1 mm in cross section. The thin section also shows that one body can overlap another: at the place where the overlapping occurs, the specimen with larger diameter presses through the smaller one (Figure 2B). In addition, inside the sandstone layer, another specimen can be seen in the thin section, which was not observed during the initial examination of the sample. That there is a rim in each specimen consisting of the thinly laminated finer material is also evident under the scanning electron microscope (Figure 2C). EDS mapping shows that the rim predominantly consists of ultra-thin laminas of Fe–Mn oxides, and its total thickness does not exceed 150 µm.
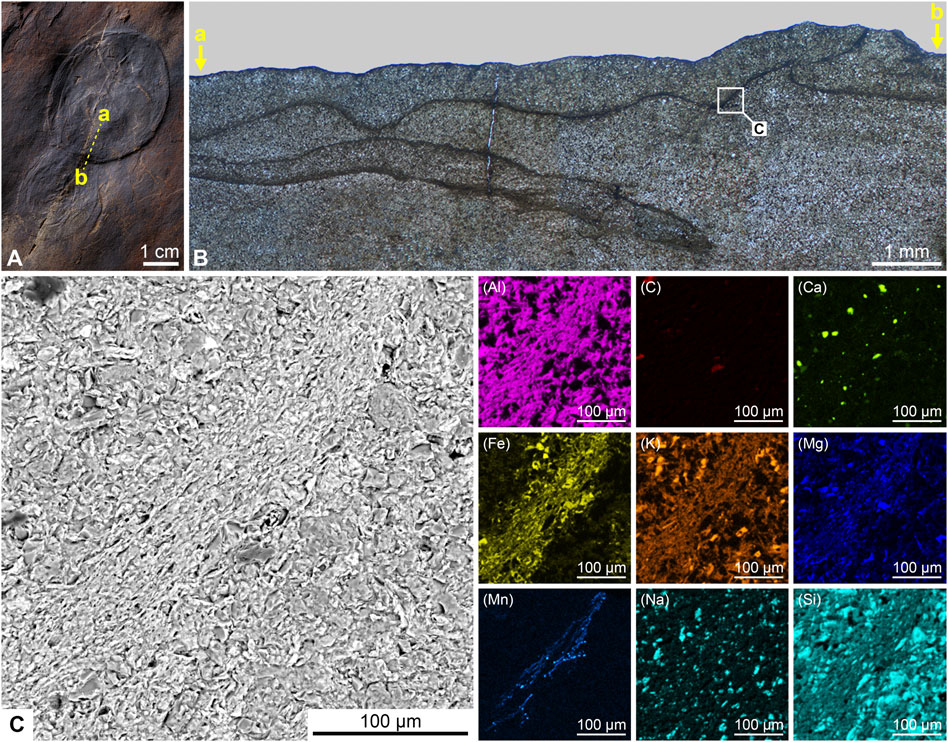
FIGURE 2. Three-dimensional preservation of Beltanelliformis konovalovi sp. nov. on the upper bedding surface in fine-grained sandstone: (A) CU2019/004, specimens adjoined to each other (dashed line marks a thin section surface); (B) photograph of thin section under optical microscope (white frame marks area studied in SEM); (C) back-scattered electron image of thin section and elemental mapping of the ultra-thin rim around specimen.
The age structure of the paleocommunity on the BS surface (Figure 3A) was estimated based on size frequency distribution. The plot (Figure 3B) shows positive skew distribution for the population (Shapiro–Wilk; W = 0.87, p = 0). The population can be divided into three cohorts by size (<5 mm, 5–8 mm, and >8 mm); for this purpose, a Bayesian Information Criterion (BIC) (Fraley and Raftery, 2007) has been used. As a result, there are three size groupings: small, medium, and large individuals of organisms. The all population from the BS surface exhibits spatial aggregation (Figure 3C), although selected analysis of spatial distribution shows different behavior between smaller, medium, and larger individuals. The larger individuals tend to form spatial segregation because a greater distance than expected by random distribution with Pair Correlation Function (PCF) values is placed below the bound for complete spatial randomness (CSR), whereas the smaller and medium individuals show relatively denser aggregations (Figure 3C).

FIGURE 3. Spatial distribution of Beltanelliformis konovalovi sp. nov. (A) Schematic map of the BS surface with positions of individuals and their diameters. (B) Plot of the estimated density of individuals and their diameters. (C) Plots of pair correlation functions describing the spatial distributions of all population and three distinct size groupings in comparison with the random distribution (999 simulations).
Genus Beltanelliformis Menner in Keller et al., 1974.
Beltanelliformis konovalovi sp. nov.
Etymology. From Russian “konovalovskaya podsvita,” in reference to the first finding in the Konovalovka Member of the Chernyi Kamen Formation in Central Urals.
Holotype. Specimen No. CU2019/001 (Figure 4B) stored in the Geological Institute of the Russian Academy of Sciences (GIN RAS), Moscow, Russia.

FIGURE 4. Beltanelliformis konovalovi sp. nov. from the BS surface: (A) CU 2019/002, single specimen with radial wrinkles; (B) CU2019/001, holotype, single specimen with radial and concentric wrinkles; (C) CU2019/003, two small specimens adjoined to each other; (D) CU2019/005, (E) CU2019/006, single, paired, and clustered specimens.
Type locality. Approximately 1.6 km upstream from the mouth of the Sylvitsa River, Central Urals, ca. 77 km west of Nizhniy Tagil city, Sverdlovsk region of Russia.
Diagnosis. Three-dimensional pancake-shaped to flattened discoidal fossils from 2 mm to 3 cm in diameter, consisting of thin rim and coarser homogenous filling matter.
Description and comparison. The specimens present single, paired, or occasionally aggregated bodies of prokaryotic macroscopic organisms lived on the floor or buried within the sediment. They can be preserved in fine-grained sandstone beds or on their top surfaces as low-relief (convex epirelief) discoidal or ellipsoidal structures (Figure 4). Their shortest diameter ranges between 2.7 and 29.5 mm. The specimens are flattened, with variously oriented wrinkle-like thin ridges in plain view: chaotic (Figure 4A); concentric (Figures 4B–D); and low-relief or smooth ones (Figure 4E). Along the edges, there are observable concentric folds (Figures 4B,D). Some of specimens have isometric depressions with thin concentric wrinkles and a relatively thick folded rim (Figures 4D,E). Frequently two or several specimens are adjoined or partially overlapped (Figures 4C,D) or organized into pair- or chain-like structures (Figure 4E). Homogenous filling sediment was possibly penetrated through the ultra-thin rim of the body. Unlike the type species (B. brunsae), B. konovalovi sp. nov. appear to be a flattened or disk-like nonspherical body with ultra-thin rim, frequently found as single and relatively large (up to 3 cm in diameter) fossil. Ivantsov et al. (2014) suggested that B. brunsae was a colonial spherical organism with a dense smooth envelope without any pores or openings, and its interior was filled with homogenous viscous material. In addition, in contrast to B. konovalovi sp. nov., B. brunsae in all cases represent large gatherings of organisms. Another taxon B. minutae (McIlroy et al., 2005) is characterized by significantly smaller size of specimens than in B. brunsae and B. konovalovi sp. nov.
Material. 196 specimens.
Occurrence. Central Urals, Sylvitsa and Kos’va (pers. obs.) rivers, Sverdlovsk and Perm’ regions, Russia; Konovalovka Member, Chernyi Kamen Formation, Sylvitsa Group, Upper Vendian, Ediacaran.
Remarks. Ivantsov et al. (2014) revised the preservation variants of Beltanelloides sorichevae, B. podolicus, Hagenetta aarensis, Medusinites palij, Nemiana simplex, and Namamedusium wendti and came to conclusion that all of them are synonyms of the type species B. brunsae Menner. In addition, they consider B. minutae as separate species. Thus, for the moment, genus Beltanelliformis may include three species B. brunsae, B. minutae, and B. konovalovi sp. nov.
Previous scholars have shown that fossils of Beltanelliformis have three main preservation variants: Nemiana-type variant, a relatively high convex mold on the sole of sandstone beds or lenses; Beltanelloides-type variant, a discoidal slightly convex imprint on the sole of fine-grained siltstones or organic films in clays; and concave smooth imprints (Ivantsov et al., 2014). This study demonstrates that the preservation variant of Beltanelliformis konovalovi sp. nov. from the Chernyi Kamen Formation is considerably different from the others. In contrast to them, all of the specimens from the Central Urals are preserved on the top surfaces of fine-grained sandstone beds, which is also not typical for many other Ediacaran macrofossils. Furthermore, thin sections through the specimens demonstrate more compelling evidence in favor of the three-dimensional variant of preservation of these fossils within sandstone beds such as in some representatives of vendobionts from the Ediacaran of the White Sea area and Namibia (Fedonkin et al., 2007). The presence of ultra-thin laminas of Fe–Mn oxides around each body allows interpreting it as three-dimensional envelopes or “death masks” (Gehling, 1999) produced by remains of bacterial material (Vasilatos and Economou-Eliopoulos, 2017) in a relatively anoxic setting after burial in sediment. In turn, the presence of variously oriented wrinkles on the top surfaces of the fossils can be a result of different stages of collapsing convex bubble-like bodies.
The size frequency distribution shows the predominance of smaller individuals in the studied population of B. konovalovi sp. nov. Assuming that the age of the organism is manifested in the body size, we can see a significant predominance of juvenile individuals in the population. Similar results were obtained a bit earlier in the analysis of Dickinsonia on the DS surface in the same fossil locality (10 m lower BS surface), where the specimens also demonstrate predominance of juveniles (Sozonov et al., 2019; Mitchell et al., 2020).
On the one hand, if Beltanelliformis was a metazoan organism, a possible scenario that can envisage the spatial and size frequency distribution in the population of B. konovalovi sp. nov. could be interpreted as “nesting grounds” fossilized in the intertidal zone or extremely shallow marine environment where the juveniles passed underwent maturation before migrating back to the deeper subtidal settings, as in case with Dickinsonia (Mitchell et al., 2020). It explains the high proportion of juveniles in a single population in the extremely shallow environment and their evidently low proportion in populations preserved in relatively deeper settings (Leonov, 2007; Leonov and Rudko, 2012). The studied population can be characterized as expanding, and the extremely shallow marine settings could have been favorable for these organisms.
On the other hand, Ivantsov et al. (2014) examined abundant fossil material and made assumption that these organisms were the remains of photosynthesizing bacterial colonies. Furthermore, recent results of the phylogenetic analysis provided a molecular-scale evidence for cyanobacterial origin of Beltanelliformis organically preserved in clays in the White Sea area (Bobrovskiy et al., 2018a). They also compared the fossils with modern spherical cyanobacterial colonies of the genus Nostoc and suggested that Beltanelliformis may be an ancient representative on nostocalean organisms. The results of spatial analysis of B. konovalovi sp. nov. (Figure 3) do not significantly contradict with the interpretation of cyanobacterial affinity for this fossil. For example, it was shown earlier that the size distribution of modern Nostoc spheres is also left-skewed (Steiner, 1997) and it is quite similar to the size distribution in case of Beltanelliformis fossils from the Central Urals. In addition to that, patterns of non-random spatial distribution in colonial organisms, including modern genus Nostoc, are inherent in self-organized bacterial communities (Marrocco et al., 2010; Xu et al., 2020). A possible explanation for such unusual behavior of B. konovalovi sp. nov. is that it was infaunal cyanobacterial self-organizing colony inhabited in extremely shallow marine environment and adapted to periodic desiccation. Thus, despite the fact that spatial behavior of both Beltanelliformis and Dickinsonia in the Chernyi Kamen Formation is quite similar, the phylogenetic relationships of these Ediacaran macroscopic organisms remain different: biomarkers obtained from organically preserved Dickinsonia in the White Sea indicate a metazoan affinity (Bobrovskiy et al., 2018b).
Relatively simple morphology of most representatives of the ancient macroscopic fossils and lack of evidence of recent analogs all combine to erect many speculations about the nature of the Ediacaran biota, starting from the most ancient animals and giant protozoans to lichens inhabiting on land (Xiao and Laflamme, 2009). Even if the principal outlines of these problematic organisms are becoming clearer, much of interest remains to be discovered. The results of studying Beltanelliformis konovalovi sp. nov. from the Central Urals add a new paleoecological view of these problematic organisms and also expand the range of taphonomic possibilities of prokaryotic life and self-organized systems in the terminal Neoproterozoic.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
The author confirms being the sole contributor of this work and has approved it for publication.
The core of the study was funded by the Russian Science Foundation (grant No. 21-77-10106). Thin-section processing and interpretation of photographs was supported by the Ministry of Education and Science of Russia (megagrant No. 075-15-2019-1883). Field research was carried out on the assignment to the Geological Institute of the Russian Academy of Sciences.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The author thanks Alexander Kiselev for providing SEM and EDS analysis in the GIN RAS.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2022.875001/full#supplementary-material
Aseeva, E. A. (1988). “Fossil Remains of Vendian Thallophytes,” in Biostratigraphy and Peleogeographic Reconstructions of the Precambrian of Ukraine. Editors V. A. Ryabenko, E. A. Aseeva, and V. V. Furtes (Kiev: Naukova Dumka), 81–92.
Baddeley, A., Rubak, E., and Turner, R. (2015). Spatial point Patterns. Methodology and Applications with R. Boca Raton, FL: Chapman and Hall/CRC.
Bobkov, N. I., Kolesnikov, A. V., Maslov, A. V., and Grazhdankin, D. V. (2019). The Occurrence of Dickinsonia in Non-marine Facies. Estudios Geologicos 75 (2), e096. doi:10.3989egeol.43587.55110.3989/egeol.43587.551
Bobrovskiy, I., Hope, J. M., Ivantsov, A., Nettersheim, B. J., Hallmann, C., and Brocks, J. J. (2018b). Ancient Steroids Establish the Ediacaran Fossil Dickinsonia as One of the Earliest Animals. Science 361, 1246–1249. doi:10.1126/science.aat7228
Bobrovskiy, I., Hope, J. M., Krasnova, A., Ivantsov, A., and Brocks, J. J. (2018a). Molecular Fossils from Organically Preserved Ediacara Biota Reveal Cyanobacterial Origin for Beltanelliformis. Nat. Ecol. Evol. 2, 437–440. doi:10.1038/s41559-017-0438-6
Fedonkin, M. A., Gehling, J. G., Grey, K., Narbonne, G. M., and Vickers-Rich, P. (2007). The Rise of Animals: Evolution and Diversification of the Kingdom Animalia. Baltimore: The Johns Hopkins University Press, 326.
Fraley, C., and Raftery, A. E. (2007). Bayesian Regularization for normal Mixture Estimation and Model-Based Clustering. J. Classification 24, 155–181. doi:10.1007/s00357-007-0004-5
Gehling, J. G. (1999). Microbial Mats in Terminal Proterozoic Siliciclastics: Ediacaran Death Masks. Palaios 14, 40–57. doi:10.2307/3515360
Gnilovskaya, M. B., Ishenko, A. A., Kolesnikov, Ch. M., Korenchuk, L. V., and Udal’tsov, A. P. (1988). Vendotenidy Vostochno-Evropeiskoi Platformy. Leningrad: Nauka, 143.
Grazhdankin, D. V., Marusin, V. V., Meert, J., Krupenin, M. T., and Maslov, A. V. (2011). Kotlin Regional Stage in the South Urals. Dokl. Earth Sc. 440 (1), 1222–1226. doi:10.1134/S1028334X11090170
Grazhdankin, D. V., Maslov, A. V., Krupenin, M. T., and Ronkin, Yu. L. (2010). Depositional Systems of the Sylvitsa Group (Upper Vendian of the Central Urals). Ekaterinburg: UrO RAN, 280.
Ivantsov, A. Y., Gritsenko, V. P., Konstantinenko, L. I., and Zakrevskaya, M. A. (2014). Revision of the Problematic Vendian Macrofossil Beltanelliformis (=Beltanelloides, Nemiana). Paleontol. J. 48 (13), 1415–1440. doi:10.1134/S0031030114130036
Keller, B. M., Menner, V. V., Stepanov, V. A., and Chumakov, N. M. (1974). New Finds of Metazoa in the Vendian of the Russian Platform. Izvestia Akademii Nauk SSSR Seriya Geologitcheskaya 12, 130–134.
Kolesnikov, A. V., Grazhdankin, D. V., and Maslov, A. V. (2012). Arumberia-Type Structures in the Upper Vendian of the Urals. Dokl. Earth Sci. 447 (1), 1233–1239. doi:10.1134/S1028334X12110013
Kolesnikov, A. V., Danelian, T., Gommeaux, M., Maslov, A. V., and Grazhdankin, D. V. (2017). Arumberiamorph Structure in Modern Microbial Mats: Implications for Ediacaran Palaeobiology. Bull. Soc. géol. Fr. 188 (5), 1–10. doi:10.1051/bsgf/2017006
Leonov, M. V. (2007). Comparative Taphonomy of Vendian Genera Beltanelloides and Nemiana: Taxonomy and Lifestyle. Geol. Soc. Lond. Spec. Publications 286, 259–267. doi:10.1144/SP286.18
Leonov, M. V., and Rud’ko, S. V. (2012). Finding of the Ediacaran-Vendian Fossils in the Far Taiga Deposits, Patom highlands. Stratigr. Geol. Correl. 20 (5), 497–500. doi:10.1134/S0869593812050061
Marrocco, A., Henry, H., Holland, I. B., Plapp, M., Séror, S. J., and Perthame, B. (2010). Models of Self-Organizing Bacterial Communities and Comparisons with Experimental Observations. Math. Model. Nat. Phenom. 5 (1), 148–162. doi:10.1051/mmnp/20105107
McIlroy, D., Crimes, T. P., and Pauley, J. C. (2005). Fossils and Matgrounds from the Neoproterozoic Longmyndian Supergroup, Shropshire, UK. Geol. Mag. 142 (4), 441–455. doi:10.1017/s0016756805000555
Mitchell, E. G., Bobkov, N., Bykova, N., Dhungana, A., Kolesnikov, A. V., Hogarth, I. R. P., et al. (2020). The Influence of Environmental Setting on the Community Ecology of Ediacaran Organisms. Interf. Focus. 10, 20190109. doi:10.1098/rsfs.2019.0109
Narbonne, G. M., and Hofmann, H. J. (1987). Ediacaran Biota of the Wernecke Mountains, Yukon, Canada. Palaeontology 30 (4), 647–676.
Ronkin, Y. L., Grazhdankin, D. V., Maslov, A. V., Mizens, G. A., Matukov, D. I., Krupenin, M. T., et al. (2006). U-pb (SHRIMP II) Age of Zircons from Ash Beds of the Chernokamen Formation, Vendian Sylvitsa Group (Central Urals). Dokl. Earth Sc. 411 (9), 1341–1345. doi:10.1134/S1028334X06090029
Runnegar, B. N., and Fedonkin, M. A. (1992). “Proterozoic Metazoan Trace Fossils,” in The Proterozoic Biosphere. A Multidisciplinary Study. Editors J. W. Schopf, and C. Klein (New York: Cambridge University Press), 369–388.
Seilacher, A. (1992). Vendobionta and Psammocorallia: Lost Constructions of Precambrian Evolution. J. Geol. Soc. 149, 607–613. doi:10.1144/gsjgs.149.4.0607
Sozonov, N. G., Bobkov, N. I., Mitchell, E. G., Kolesnikov, A. V., and Grazhdankin, D. V. (2019). Ecología de Dickinsonia en llanuras mareales. Estud. Geol. 75 (2), 116. doi:10.3989/egeol.43587.571
Steiner, M. (1997). Chuaria Circularis Walcott 1899 – “Megasphaeromorph Acritarch” or Prokaryotic Colony? Acta Universitatis Carolinae Geologica 40, 645–665.
Vasilatos, C., and Economou-Eliopoulos, M. (2017). Fossilized Bacteria in Fe-Mn-Mineralization: Evidence from the Legrena Valley, W. Lavrion Mine (Greece). Minerals 8, 107. doi:10.3390/min8030107
Xiao, S., and Laflamme, M. (2009). On the Eve of Animal Radiation: Phylogeny, Ecology and Evolution of the Ediacara Biota. Trends Ecol. Evol. 24 (1), 31–40. doi:10.1016/j.tree.2008.07.015
Xu, X., Risoul, V., Byrne, D., Champ, S., Douzi, B., and Latifi, A. (2020). HetL, HetR and PatS Form a Reaction-Diffusion System to Control Pattern Formation in the Cyanobacterium Nostoc PCC 7120. eLife 9, e59190. doi:10.7554/eLife.59190
Keywords: Beltanelliformis, Chernyi Kamen Formation, Ediacaran, Central Urals, taphonomy, spatial analysis, self-organization
Citation: Kolesnikov A (2022) Beltanelliformis konovalovi sp. nov. From the Terminal Neoproterozoic of Central Urals: Taphonomic and Ecological Implications. Front. Earth Sci. 10:875001. doi: 10.3389/feart.2022.875001
Received: 08 March 2022; Accepted: 12 April 2022;
Published: 12 May 2022.
Edited by:
Ben Yang, Chinese Academy of Geological Sciences (CAGS), ChinaReviewed by:
Olev Vinn, University of Tartu, EstoniaCopyright © 2022 Kolesnikov. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anton Kolesnikov, a29sZXNuaWtvdkBnaW5yYXMucnU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.