 Robert R. Reisz
Robert R. Reisz Diane Scott1
Diane Scott1
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci., 07 March 2022
Sec. Paleontology
Volume 10 - 2022 | https://doi.org/10.3389/feart.2022.847560
This article is part of the Research TopicOrigin and Early Evolution of AmniotesView all 16 articles
The detailed description of the cranial anatomy of Cotylorhynchus romeri, a very large caseid synapsid from the lower Permian Hennessey Formation of Oklahoma, uncovered several potential autapomorphies, including parietal-postorbital contact greatly reduced by broad anterior process of supratemporal; the postparietals are transversely broad and contact the supratemporals laterally; the quadratojugal has a pennant-like occipital process; the stapes has a short shaft and a ventral process that abuts against the palate; the bulbous marginal dentition narrows distally and carries three small denticles; the vomer has three large teeth along its medial edge; parasphenoidal dentition is present; and the surangular overlaps the posterodorsal tip of the dentary and excludes it from the coronoid eminence. Owing to lack of comparative cranial material in most large caseids, the evolutionary history of these autapomorphies remains ambiguous because they cannot be determined in the closest relatives of Cotylorhynchus romeri. Our description of the skull of Cotylorhynchus romeri is consonant with the hypothesis that this caseid was a high-fibre terrestrial herbivore. The recent hypothesis that Cotylorhynchus romeri was primarily aquatic was proposed on a paleobiological basis that ignored paleontological and taphonomic evidence from the Hennessey Formation. Autochthonous preservation of several articulated skeletons of Cotylorhynchus romeri in subaerially deposited sediments that also preserve “swarms” of an aestivating fossil amphibian (Brachydectes) indicate that this caseid was the largest tetrapod of a terrestrial fauna that lived in a monsoonal climate.
Caseid synapsids have been known since early in the last century, when Paul C. Miller discovered the Cacops bonebed at the lower Permian Indian Creek locality, near the Big Wichita River, Baylor County, Texas, and Williston (1910) described Casea broilii, one of the three important components of that unusual assemblage. The first large caseid Cotylorhynchus romeri was described by Stovall (1937) from the lower Permian Hennessey Formation of Oklahoma. Romer and Price (1940) gave a rather brief description of this species. Numerous specimens were subsequently collected, but very little additional information was published until the description of the postcranial skeleton by Stovall et al. (1966). The bulk of subsequent work on caseids was conducted and published by Olson and colleagues (Olson and Beerbower, 1953; Olson, 1954; Olson, 1962; Olson and Barghusen, 1962) culminating in a review of the family (Olson, 1968). Despite this concerted effort, the cranial anatomy of caseids remains poorly documented. Subsequent phylogenetic analyses of synapsid relationships (Reisz, 1980; Brinkman and Eberth, 1983; Reisz, 1986) have recognized the significance of caseids as basal synapsids, but the lack of detailed anatomical features of the caseid skull hampered any attempts to resolve the interrelationships of basal synapsids, and the relative positions of caseid, varanopid, and eothyridid synapsids are still controversial (MacDougall et al., 2018; Ford and Benson, 2019).
The family Caseidae is composed of 13 species that exhibit a striking diversity in size, but all apparently conform to a relatively narrow morphological skull pattern. Most of the taxa are based on poorly preserved materials, making phylogenetic analyses difficult. However, four caseids have sufficiently well-preserved skeletons to be useful in providing detailed comparative anatomical information about the anatomy of this important group. These are Cotylorhynchus romeri from the lower Permian of Oklahoma, Ennatosaurus tecton from the middle Permian of Northern Russia, Euromycter rutenus (formerly Casea rutena; for nomenclatural emendation, see Reisz et al, (2011)) from the Autunian (lower Permian) of France, and Martensius bromackerensis from the lower Permian of Germany (Berman et al., 2020). Although the genotype of the family, Casea broilii, is known from three skulls as well as postcranial materials, all are badly preserved and prepared, making detailed anatomical comparisons difficult. Maddin et al. (2008) redescribed Ennatosaurus tecton on the basis of one of the skulls from the Pinega locality that has been carefully prepared and a new skull collected from the Mezen River Basin. The skull of Euromycter rutenus was described in detail by Sigogneau-Russell and Russell (1974), but critical areas of its anatomy, like the skull table, and occipital regions, have remained uncertain. The discovery and description of Eocasea (Reisz and Fröbisch, 2014) the oldest known member of the clade, has added limited but valuable information about the posterior part of the skull. However, most detailed anatomical data is offered by the known specimens of Cotylorhynchus romeri, the basis of this work.
The holotype of Cotylorhynchus romeri, consisting of a partial right side of a skull, part of an interclavicle, and left and right manus (OMNH 00637), was discovered in 1937 by William Strain and described by Stovall (1937). Although numerous other specimens were collected and the postcrania was described in detail by Stovall et al. (1966), only brief, partial descriptions of the cranial anatomy were ever published (Romer and Price, 1940; Olson, 1968). The most complete skull (OMNH 04329) from the Norman area, Cleveland County, Oklahoma, together with the holotype are used for the following description and reconstruction.
Synapsida Osborn, 1903
Caseasauria Williston, 1912
Caseidae Williston, 1911
Cotylorhynchus romeri Stovall, 1937
Holotype: OMNH 00637, partial right side of skull (Figure 1), right lower jaw, partial interclavicle, and left manus.
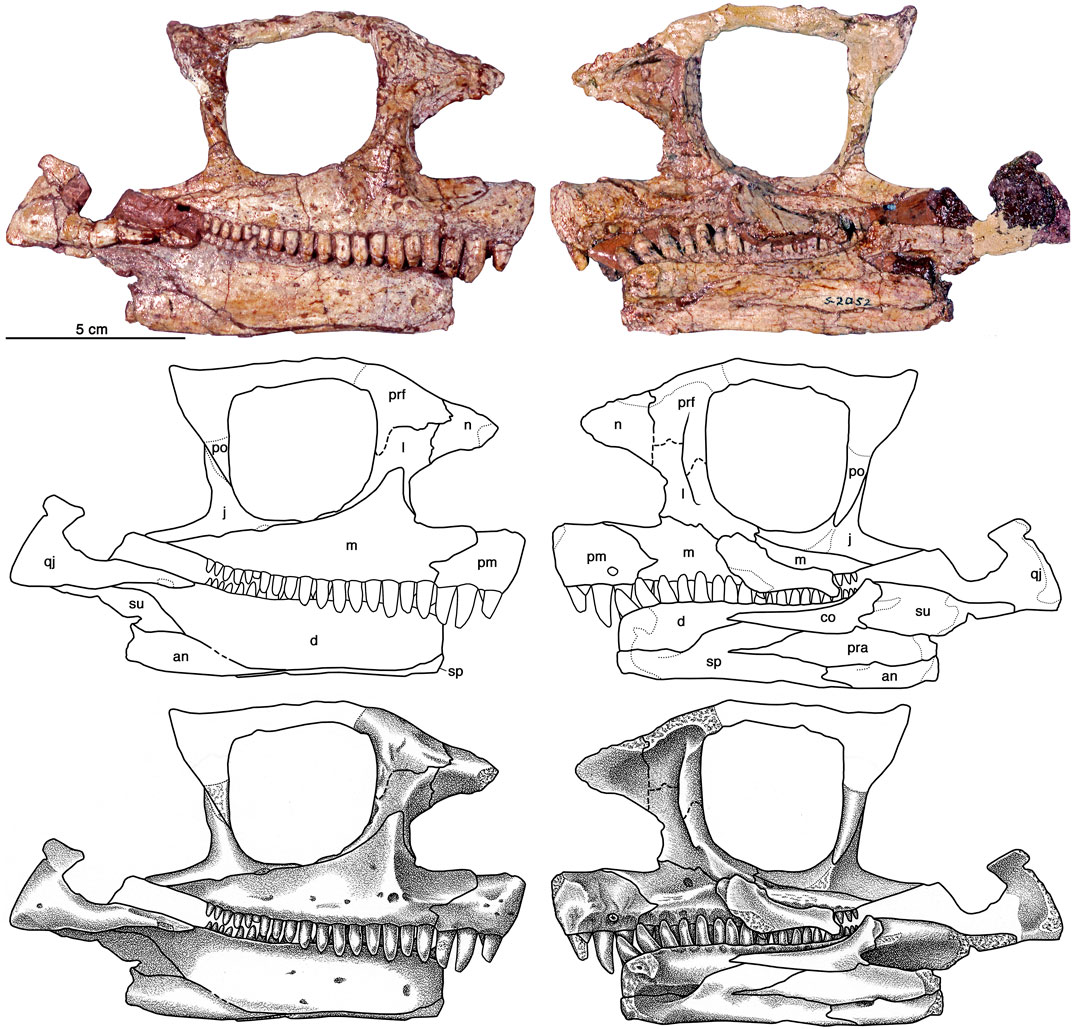
FIGURE 1. Cotylorhynchus romeri, OMNH 00637, holotype. Right lateral and medial views of skull. Abbreviations: an, angular; co, coronoid; d, dentary; j, jugal; l, lacrimal; m, maxilla; n, nasal; pm, premaxilla; po, postorbital; pra, prearticular; prf, prefrontal; qj, quadratojugal; sp, splenial; su, surangular.
Referred Specimens: OMNH 00605, tail, pelvis, and hind limb; OMNH 00630, various bones and bone fragments; OMNH 00655, complete skeleton; OMNH 01673, complete mounted skeleton; OMNH 01704, skull and jaws; OMNH 04329, skull and jaws (Figures 2, 3).

FIGURE 2. Cotylorhynchus romeri, OMNH 04329, referred specimen. Photographs in lateral and occipital views.
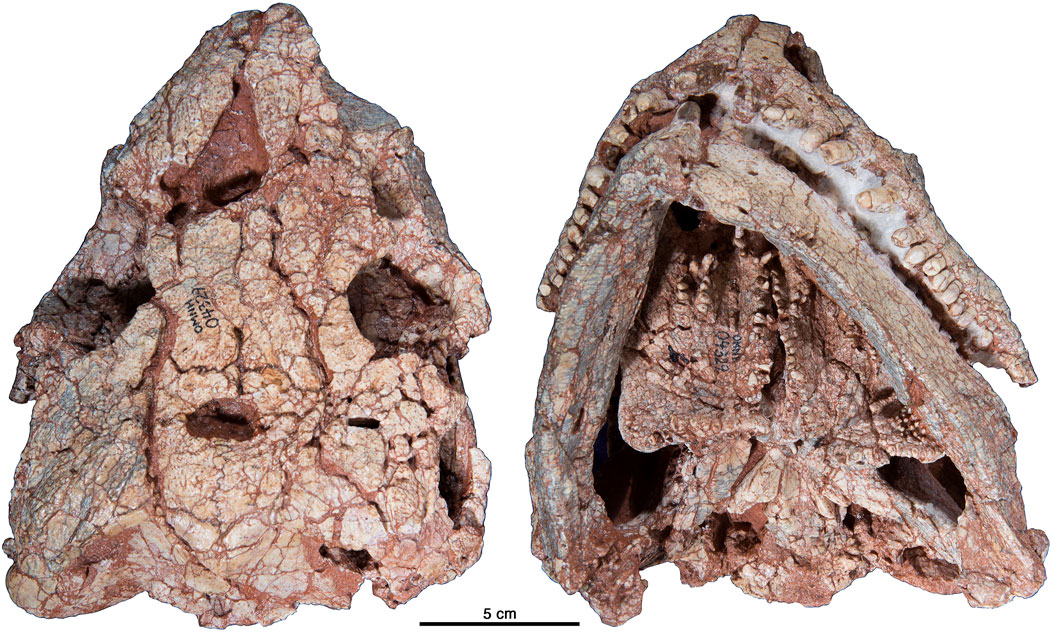
FIGURE 3. Cotylorhynchus romeri, OMNH 04329, referred specimen. Photographs in dorsal and palatal views.
Diagnosis: A large caseid distinguished by transversely broad postparietals that contact the supratemporals laterally, large supratemporal that restricts contact between the parietal and postorbital, stapes that has short massive distal shaft and a ventral process that is braced against the quadrate ramus of the pterygoid, vomerine dentition accentuated by three large teeth along the medial edge of the bone; parasphenoidal dentition present, and large surangular that overlaps the posterodorsal tip of the dentary and excludes it from the coronoid eminence. Maxillary tooth crowns are broad basally and narrow towards the tip and culminate with three small cuspules.
Locality and Horizon: Holotype from OMNH locality V134, Logan County, Oklahoma, United States OMNH 04329 from OMNH locality V381, Cleveland County, Oklahoma, United States. See Czaplewski et al. (1994) for provenance of other referred specimens listed above. All specimens are from the Hennessey Formation, lower Permian (Kungurian).
Stratigraphic Note: The Hennessey Formation was revised and raised to group status and divided into the Fairmont Shale, the Kingman Sandstone, and the Salt Plains Formation (Wood and Burton, 1968; Bingham and Moore, 1975), but this arrangement has been disputed by N. Suneson and colleagues (Suneson and Hemish, 1998; Suneson et al., 1999; Lucas and Suneson, 2002). In light of this controversy, we follow the traditional usage of the Hennessey Formation for convenience because the majority of the paleontological field work is conceived in the framework of the traditional view of this geological unit (Vaughn, 1958; Olson, 1967; Olson, 1970).
The skull conforms to the general pattern of caseids, with an anteriorly tilted snout and a strikingly broad skull roof which has a low lateral profile (Figures 4, 5). Except for the slender anterior tip of the snout, every other region of the skull has a broad cross-sectional outline, with the lateral width far exceeding its height. Total skull length is ca. 20 cm, which is ca. 10% of the snout-vent length using the skeletal reconstruction of Stovall et al. (1966, fig. 17).
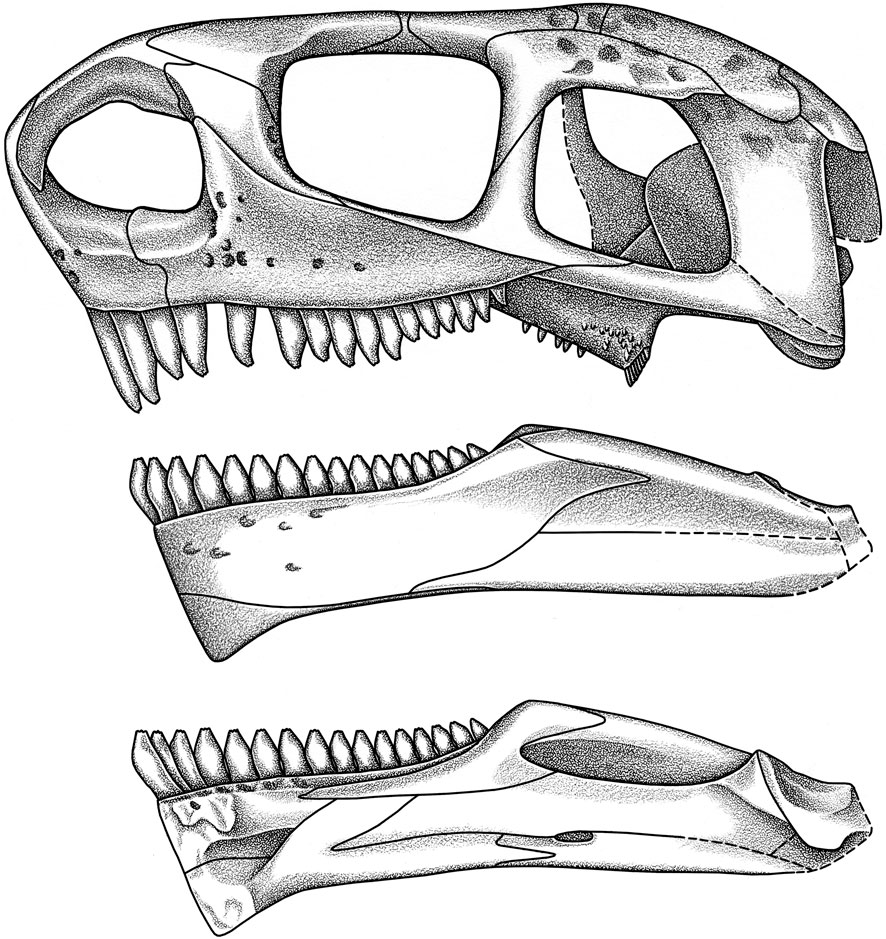
FIGURE 4. Cotylorhynchus romeri. Skull reconstruction in left lateral view, and medial and lateral views of mandible.

FIGURE 5. Cotylorhynchus romeri. Skull reconstruction in dorsal, ventral, posterior and anterior views.
As in other caseids, the external naris is unusually large and its outer margins extend onto the external surface of the surrounding bones. This results in the formation of large shelves on the premaxilla, the maxilla, the lacrimal, and a smaller shelf on the nasal. The combined narial shelves of the nasal, the lacrimal, and the maxilla form a massive posteromedial shelf of the naris (Figures 4, 5), a feature that is also present in Ennatosaurus tecton and Euromycter rutenus, and is probably also present in other large caseids, but are not well preserved. In the small, more basal taxa like Oromycter (Reisz, 2005), the maxilla lacks the distinct narial shelf seen in the larger more derived taxa, as is the case with the maxilla of Martensius (Berman et al., 2020). It is probable that this greatly enlarged narial opening may be related to the forward tilting of the snout, resulting in the formation of a slight rostrum. It is clear, however, that the great narial enlargement is exaggerated in the more derived caseid taxa, and that the narial shelf evolved within the clade. The overall result of the greatly enlarged external naris has also resulted in the shortening of the lacrimal bone in derived caseids and of the antorbital region of the cheek. In Cotylorhynchus romeri the external edges of this naris form a surface area that is nearly equal to that of the orbit, and approximately equal to that of the temporal fenestra, a feature which also characterizes the other, more derived caseids. In contrast to the very large external naris, the orbit is relatively small, and is underlain by a deep surborbital ramus. The temporal fenestra is relatively larger than in the smaller caseids, and this size is mainly at the expense of the squamosal bone posteriorly, and the subtemporal bar ventrally, both being reduced in size. The broad skull table is pierced by a very large pineal foramen, just posterior to the deep orbital emarginations.
One of the most striking features of the skull is related to ornamentation of the outer surface of the dermal skull roof (Figures 3, 5). In contrast to the rather smooth lateral surfaces of the skull roof and the mandible, the dorsal surface of the skull table is deeply pitted by strongly developed pattern of ornamentation. This pattern is characterized by the presence of distinct, round pits and short grooves that cover the preserved surfaces of the skull table and dorsal surface of the snout. This ornamentation extends on to the dorsal surface of the postorbital, prefrontal, and nasal, but not to their lateral surfaces. The squamosal seems to represent an exception to this pattern, in that its dorsal portion, close to its suture to the supratemporal and the postorbital is gently ornamented, but this is clearly part of the lateral surface of the skull roof. Compared to Euromycter rutenus, and similar to the skull roof of Ennatosaurus tecton (q.v. Maddin et al., 2008), the interorbital region of the skull roof is distinctly waisted by the medial protrusion of the orbits in Cotylorhynchus romeri; however, the condition in Ennatosaurus tecton is more pronounced because of the wide separation of the prefrontal and the postfrontal, which results in the frontal making a greater contribution to the orbital margin than in either Cotylorhynchus romeri and Euromycter rutenus (Sigogneau-Russell and Russell, 1974).
The occipital region is characterized by the wide occipital plate and similarly broad exoccipitals on either side of the foramen magnum (Figures 2, 5). The supraccipital-opisthotic complex is framed above and laterally by the slender, delicate postparietal, and the hatchet-shaped tabular. Both the paroccipital plate and the posttemporal fenestra are unexpectedly large for a synapsid, but comparison with other caseids reveals that this is a characteristic feature of this clade. The massive, relatively stubby appearance of the stapes is another unusual feature of the occiput, as is the relatively small, ventrally facing occipital condyle.
The palatal surface of the skull is broad (Figures 3, 5), as indicated by the large mediolateral dimensions of all palatal bones. This surface is also characterized by the presence of unexpectedly large, slender teeth that are arranged in rows across the posterior edge of the transverse flange of the pterygoid, along the medial edges of the pterygoid and the vomer. The teeth not only form discrete fields on the body of the transverse flange and along the pterygoid-palatine suture, but are also present on the parasphenoid, and many of these teeth are tall and relatively slender. A few of the palatal teeth are preserved at the tip, and they have the same distal morphology as the marginal teeth, showing three small cuspules.
The mandible is short and massive, with a large ventrally expanded mandibular symphysis, and a well-developed coronoid eminence that rises above the level of the tooth row, and well above the level of the jaw articulation (Figures 1, 2, 4). There is only a single coronoid bone that extends far anteriorly, nearly to the level of the symphysis. Although there is no evidence of dental occlusion, the articular surface of the mandible is broader and much longer than the corresponding quadrate condyles, raising the possibility of propalinal movement of the mandible. The pterygoideus process, formed by the prearticular and articular bones, is massive and extends far medially.
The premaxilla is a relatively large element that forms the anteroventral portion of the snout (Figures 2–8). It consists of a ventral alveolar region that supports three teeth and an anterodorsally extending ramus that forms the anterior margin of the external naris. The ventral portion is the thickest region of the element. In ventral aspect the ventral portion arches slightly anterolaterally to form the anterior-most part of the upper dental arcade. Posteriorly this portion of the premaxilla forms an anteroposteriorly short but dorsoventrally deep scarf contact with the maxilla; the anterodorsal corner of the ventral portion of the premaxilla was slightly overlapped by the septomaxilla. The lateral surface of the bone is bifaceted and consists of a ventral, slightly laterally leaning wall and a dorsal narial shelf that curves anterodorsally to form the deep portion of the anterodorsal ramus. The lateral surface proper is largely flat and featureless apart from the presence of several small foramina, a couple of which appear to be supralabial foramina. Terminating along a broadly sigmoidal suture with the maxilla just posterior to the midpoint of the external naris, the ventral ramus is intermediate in length between the short ramus described for Ennatosaurus tecton and the relative long one documented in Euromycter rutenus. The lateral (or facial) surface of the ventral portion of the bone merges seamlessly with the anterior surface of the anterodorsal ramus, which curves posterodorsally at the approximate mid-height of the external naris to deeply overlap the anterodorsal surface of the nasal. Together the paired anterodorsal processes of the premaxillae form a narrow internarial bar. Relative to the ventral portion of the bone, the internarial bar of Cotylorhynchus romeri is approximately as narrow as the internarial bars in Ennatosaurus tecton and Euromycter rutenus. A substantial part of the premaxilla is deep to the superfical, facial part of the bone, forming a shelf that extends medially from the ridge that marks the narial border. The medial surface of the ventral portion of the premaxilla is exposed in the holotype, revealing that the premaxilla is thickest along the dorsal margin of the alveolar portion of the bone. The latter leans strongly medially (mesially) and is sculpted with shallow, arch-like fossae that are associated with each tooth position, and the fossa associated with the second tooth position also features a circular resorption pit. Anteriorly the medial surface contacting the contralateral element conforms to the roughly lenticular cross section of the ventral portion of the premaxilla. The palatal ramus of the premaxilla is little more than a slight swelling immediately lateral to the symphysis that is marked with a roughened surface for contacting the vomer. The first premaxillary tooth is damaged in both specimens, but the second and third are well preserved and reveal that the crown of each is largely conical in structure, twice the height of basal (mesial-distal) diameter, with a tip that curves slightly lingually and bears three small cuspules aligned mesiodistally. Judging from what remains of its base, the first premaxillary tooth was subcircular in cross section, as is the second premaxillary tooth. The base of the third tooth is, however, oval in cross section, being slightly compressed mesiodistally compared to the preceeding teeth. All teeth are protothecodont in implantation, and judging by the position of the resorption pit for the second premaxillary tooth in the holotype (Figure 4), the root is no more than half the height of the crown.
The maxilla of Cotylorhynchus romeri (Figures 4, 6, 7) is triangular in lateral aspect and closely resembles the low, triangular elements illustrated for Ennatosaurus tecton and Euromycter rutenus (Sigogneau-Russell and Russell, 1974; Maddin et al., 2008). The maxilla forms an acute triangular dorsal process in the narrow facial region between the external naris and the orbit, and reaches its greatest height along the posterior margin of the external naris. Judging from the medial and lateral views of the anterior facial region of OMNH 00637 (Figure 1), the dorsal process of the maxilla extends sightly dorsal to the midpoint of the height of the antorbital portion of the lacrimal, and so is roughly comparable with the conditions seen both in Ennatosaurus tecton (q.v. Maddin et al., 2008) and Euromycter rutenus (Sigogneau-Russell and Russell, 1974). Posteriorly the maxilla tapers to an acute tip that ends just anterior to the level of the midpoint of the lateral temporal opening. The ventral margin is very shallowly convex throughout its length, contrasting with the faintly sigmoidal ventral margin in Euromycter rutenus and the more conspicuous anterior convexity that forms the anterior two thirds of the ventral margin of this bone in Ennatosaurus tecton. As documented in all caseids in which this element is preserved, the lateral surface of the maxilla is perforated by numerous foramina. Most of these small openings are aligned along a line that parallels the ventral margin at the height of the ventral margin of the external naris, and presumably are supralabial foramina. In medial aspect the maxilla features a ridge that arises anteriorly along its contact with the premaxilla and extends posteriorly and dorsally along the alveolar portion of the element. Judging from the extent of roughened bone, this ridge appears to have contacted the palatine at a point directly ventral to the posterior-most extent of the external naris, posteriorly along the medial ridge to a point dorsal to the 10th or 11th maxillary tooth (approximately midway between the orbital mid-point and the postorbital process). The alveolar ridge supports 17 teeth in OMNH 00637 (Figure 1) and 13 teeth in (the left maxilla) of OMNH 04329 (Figures 6, 7). This range in the number of teeth in Cotylorhynchus romeri is slightly more than that in Casea rutena (11 teeth) and substantially more than in Casea broilii (9 teeth) and Ennatosaurus (8 teeth). The maxillary teeth are very similar to the premaxillary teeth in morphology. The anterior-most maxillary tooth is subequal to the posterior-most premaxillary tooth, and the maxillary teeth decrease progressively in size posteriorly. In contrast to the posterior-most premaxillary tooth, the crowns of the maxillary teeth are more compressed mesiodistally. The tips of the well preserved crowns exhibit three closely grouped cuspules that are aligned mesiodistally.
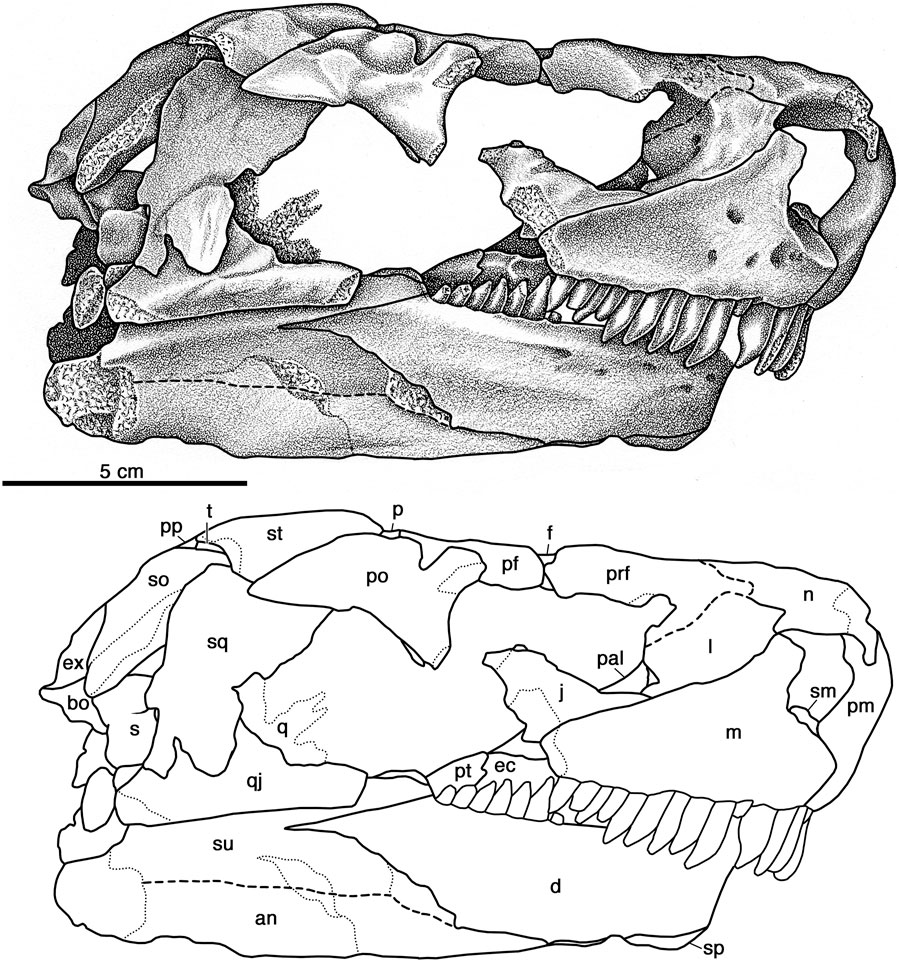
FIGURE 6. Cotylorhynchus romeri, OMNH 04329, referred specimen. Skull in right lateral view. Abbreviations used in Panel 6, Figures 7–9: an, angular; ar, articular; bo, basioccipital; co, coronoid; d, dentary; ec, ectopterygoid; ex, exoccipital; f, frontal; j, jugal; l, lacrimal; lj, lower jaw; m, maxilla; n, nasal; o, opisthotic; p, parietal; pp, postparietal; pal, palatine; pf, postfrontal; pm, premaxilla; po, postorbital; pp, postparietal; pra, prearticular; prf, prefrontal; ps, parasphenoid; pt, pterygoid; q, quadrate; qj, quadratojugal; s, stapes; sm, septomaxilla; so, supraoccipital; sp, splenial; sph, sphenethmoid; sq, squamosal; st, supratemporal; su, surangular; t, tabular; v, vomer.
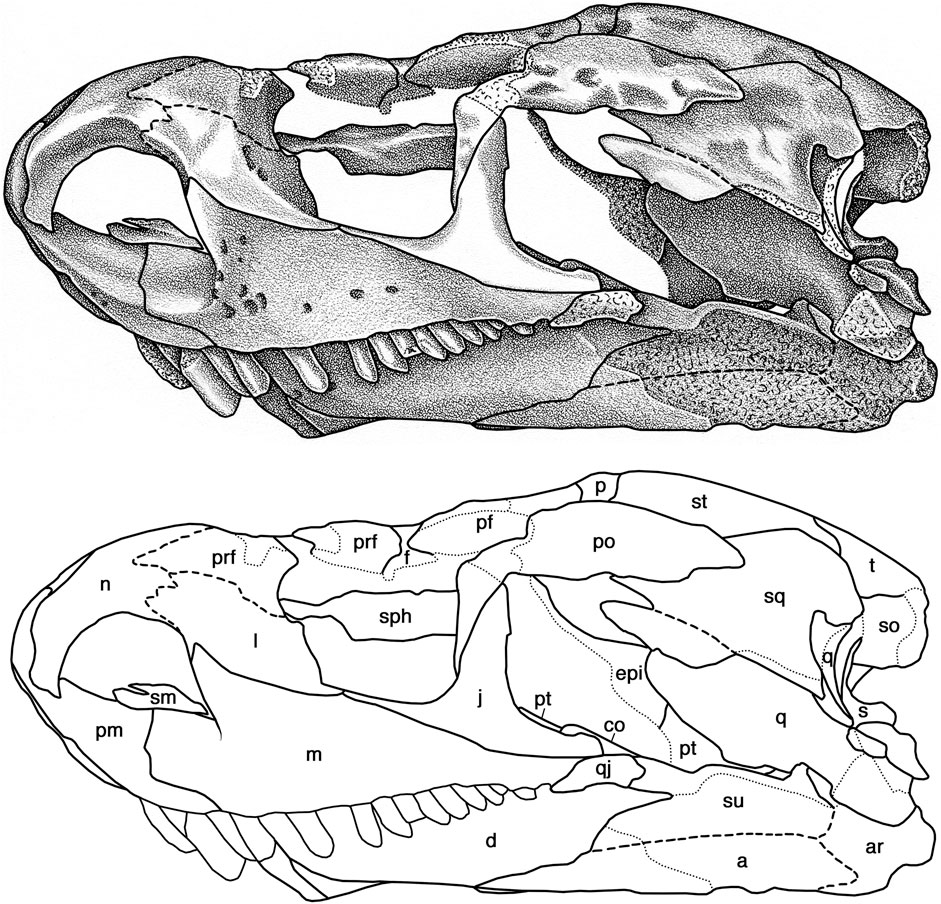
FIGURE 7. Cotylorhynchus romeri, OMNH 04329, referred specimen. Skull in left lateral view. Abbreviations as in Figure 6.
Nothing more than fragments of the septomaxilla are preserved in OMNH 04329 (Figures 6, 7). The left element is marginally better preserved and shows that it is comprised of an expanded ventral base that contacted the narial shelves of the premaxilla and the maxilla, and supported what appears to be a slightly narrower dorsal component. The latter is damaged on both sides and the full extent and the nature of the contacts with the circum-narial bones, if any, cannot be determined.
The nasals of OMNH 04329 (Figures 6–8) are better preserved than the partial element in OMNH 00637 (Figure 1) and together afford a complete view of this bone. The greater part of the nasal is an anterodorsally arching, triangular plate of bone that roofs nearly the entire rostral region. In dorsal aspect the nasal forms a slightly rounded internarial bar with the premaxilla. It expands in transverse breadth posteriorly, reaching its broadest extent where it makes a narrow contact with the lacrimal. Judging from the sutures with the prefrontal and the lacrimal in OMNH 00637, the nasal is overlain by the anterior tip of the prefrontal and the posterolateral margin of the nasal most likely continues posteromedially from the external suture with the lacrimal to the posterolateral corner of the bone where it makes contact with the frontal just medial to the prefrontal. The posterior portion of the nasal is damaged but the lateral suture with the prefrontal was roughly straight whereas the posterior suture with the frontal appears to have been coarsely serrate. This is roughly comparable to the organization of the nasal for Euromycter rutenus in one of the two interpretations of Sigogneau-Russell and Russell (1974); the sutural relationships of these elements in Ennatosaurus tecton remain conjectural (Maddin et al., 2008). Broad, shallow dimples and shallow, short furrows sculpt the dorsal surface of the bone. The anteriorly narrowing portion of the nasal arches anteroventrally, forming an extensive scarf joint with the dorsal process of the premaxilla. Deep to the free edge of the bone, the nasal forms a curving, medially sloping narial emargination that forms most of the dorsal margin of the external naris (Figure 7).
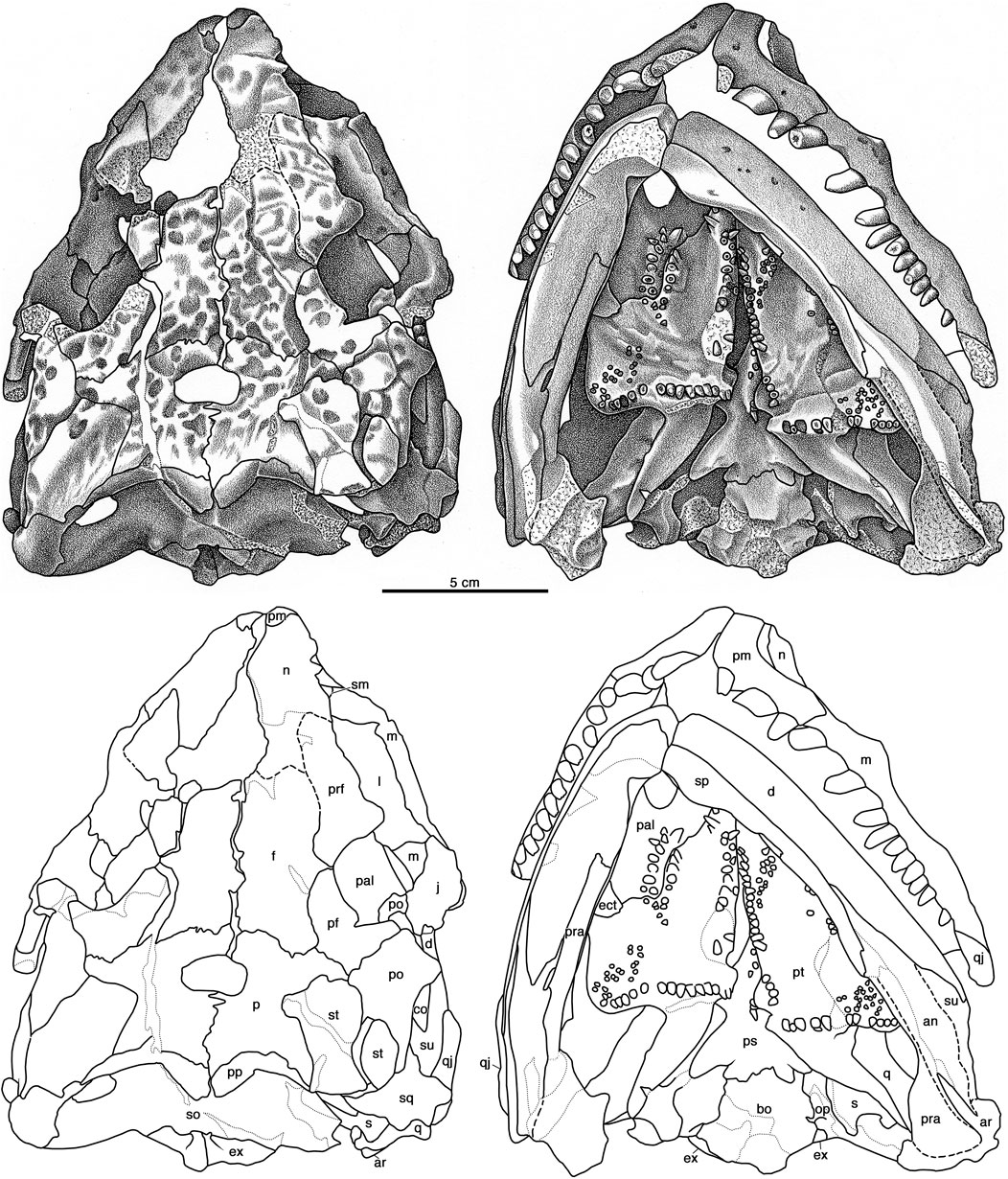
FIGURE 8. Cotylorhynchus romeri, OMNH 04329, referred specimen. Dorsal and ventral views of skull. Abbreviations as in Figure 6.
Between both OMNH 00637 and OMNH 04329 the morphology of the lacrimal of Cotylorhynchus romeri can be fully documented (Figures 1, 6, 7). The lacrimal spans the anterior cheek between the external naris and the orbit, forming the greater part of the “orbitonarial bar” sensu Maddin et al. (2008). The lacrimal is bounded anterodorsally by the nasal, directly dorsally by the prefrontal, directly ventrally by the maxilla, and posteroventrally by the jugal. The contact with the nasal in OMNH 04329 is conspicuously greater than that seen in OMNH 00637, suggestive of either individual variability in the extent of this suture or, given the greater size of the latter specimen, an ontogenetic difference. The suture with the prefrontal is a coarsely meandering, overlapping contact. The medial aspect of the lacrimal in OMNH 00637 indicates that the lateral surface is deeply overlain by the dorsal process of the maxilla. The facial surface of the lacrimal has an irregular, hourglass-shaped outline. The anterior margin of the bone thins medially to form a crescentic narial shelf that is continuous with those of the nasal and the maxilla. The posterior region of the lacrimal thickens medially to create a transversely deep, posterolaterally facing antorbital buttress. Two lacrimal puncti are present here, aligned with and positioned immediately medial (deep) to the ridge that marks the transition from the facial portion of the bone to the sloping, posterior surface of the antorbital margin. In lateral view the facial portion of the lacrimal thins posteroventrally and makes a point contact with the jugal. The medial aspect of OMNH 00637 reveals that the antorbital portion of the lacrimal transitions to a thick, medial ridge with a slightly deeper contact with the jugal medially than is evident laterally, and it extensively overlaps the medial surface of the maxilla ventral to the anteroventral corner of the orbit (Figure 1). In overall morphology, the lacrimal of Cotylorhynchus romeri is closely comparable to those of Ennatosaurus tecton and Euromycter rutenus, except that the lacrimal has a greater facial extent in Ennatosaurus tecton (q.v. Maddin et al., 2008) and the narial shelf in Euromycter rutenus is more developed ventrally (Sigogneau-Russell and Russell, 1974).
The prefrontal of Cotylorhynchus romeri conforms with the general morphology of this bone among other basal synapsids (Figures 1, 6–8). In lateral aspect it forms the anterodorsal corner of the orbit. The facial portion of the bone can be divided into a dorsal contribution to the skull table that contacts the frontal and the nasal, and an anterior portion that contacts the lacrimal. The entire skull table portion of the prefrontal is ornamented with the same pattern of broad, shallow divots and sulci seen on the nasal and the other skull table elements. This contrasts with the reduced sculpturing seen in Euromycter rutenus (Sigogneau-Russell and Russell, 1974, Figure 1) and Ennatosaurus tecton (q.v. Maddin et al., 2008). The skull table portion of the prefrontal features a relatively small lappet that extends posteromedially into an embayment in the anterolateral margin of the frontal. In this regard the prefrontal of Cotylorhynchus romeri is similar to that of Euromycter rutenus; Ennatosaurus tecton appears to lack this extension of the prefrontal and this condition is presumably an autapomorphy of that species. Deep to the facial portion of the bone, the prefrontal forms an extensive, sloping orbital shelf that merges smoothly with the antorbital buttress of the lacrimal ventrally. The medial aspect of OMNH 00637 clarifies that the antorbital portion of the prefrontal is a thin, transversely deep process that extends ventrally to a point that is midway between the dorsal surface of the prefrontal and the ventral margin of the skull.
OMNH 04329 preserves nearly complete frontals in almost perfect articulation with each other and with neighboring elements (Figures 6–8). Each frontal is, like that of Ennatosaurus tecton, a distinctly rectangular structure in dorsal aspect. Unlike the frontal of Ennatosaurus tecton, with its broad contribution to the orbital margin, and similar to the frontal of Euromycter rutenus, that of Cotylorhynchus romeri exhibits a tiny lateral lappet that makes a commensurately tiny contribution to the orbital margin. The lateral lappet of Cotylorhynchus romeri is, however, smaller than that of Euromycter rutenus relative to the rest of the bone. With respect to the lateral lappet, the anterior portion of the frontal is roughly equal in anteroposterior extent to the posterior portion of the bone. The significance of these proportions are difficult to compare with the frontal of Ennatosaurus tecton, which is autapomorphic in the absence of the lateral lappet and its conspicuous contribution to the dorsal margin of the orbit (Maddin et al., 2008), as well as to the frontal of Euromycter rutenus, in which the anterior extent of the bone is uncertain (Sigogneau-Russell and Russell, 1974). The suture with the nasal is damaged but what is present suggests that it was broadly interdigitated. Anterolaterally the frontal met the prefrontal along a slightly sigmoidal contact. Posteriorly, the suture with the parietal is weakly interdigitated, and the frontal received the postfrontal in a broadly curving posterolateral embayment. The dorsal surface of the frontal is richly ornamented with the same system of shallow dimples and sulci present on the surrounding skull-roof elements. What little is visible of the ventral surface of the frontals of OMNH 04329 indicates that they are smoothly finished (Figures 6, 7).
The jugal of Cotylorhynchus romeri is a slender triradiate element (Figures 1, 6–8). The dorsal ramus of the jugal is an acute triangle of bone that contacts the ventral process of the postorbital and with that element forms the relatively slender postorbital bar. The anterior suborbital process has a narrowly acute profile in lateral aspect and a slightly deeper profile in medial view (Figure 4). The jugal makes a narrow contact with the lacrimal and extends farther anteriorly than that in Euromycter rutenus (Sigogneau-Russell and Russell, 1974), but not as far anteriorly as the jugal of Ennatosaurus tecton (q.v. Maddin et al., 2008). The suborbital process of the jugal thickens ventromedially (Figure 1) and protrudes a thin medial process that contacts the palatine anteriorly and the ectopterygoid posteriorly (Figure 8). The subtemporal process of the jugal extends posteriorly and slightly ventrally as a narrow, tongue-like process that overlies the posterior tip of the maxilla and makes a narrow overlapping suture with the subtemporal process of the quadratojugal, which in turn contacts the maxilla to exclude the jugal from the ventral margin of the skull. As in other caseids in which the temporal region is well preserved (Sigogneau-Russell and Russell, 1974; Maddin et al., 2008), the jugal does not contact the squamosal.
The parietal is a broad element in Cotylorhynchus romeri that forms the greater part of the posterior skull table (Figure 8). In dorsal aspect it is more similar to that of Ennatosaurus tecton (q.v. Maddin et al., 2008) than that of Euromycter rutenus (Sigogneau-Russell and Russell, 1974), primarily because the parietals of the two larger caseids exhibit a distinct lateral lappet that is interposed between the postfrontal, postorbital, and supratemporal bones; the lateral lappet of the parietal of Cotylorhynchus romeri is, however, anteroposteriorly narrower than that of Ennatosaurus tecton. The anterior margin of the bone forms a tightly serrate sutures with the frontal and the postfrontal. Laterally the parietal meets the postorbital via a butt-like contact, and posterolaterally it receives the supratemporal in a deep and broad embayment. The posterior margin of the parietal is shallowly to moderately concave and receives the dorsal margin of the postparietal; the parietal does not contact the tabular as in Euromycter rutenus or as inferred for Ennatosaurus tecton. The pineal foramen is positioned anteriorly along the interparietal suture. It is transversely broader than it is anteroposteriorly long, as in Ennatosaurus tecton, but in contrast to the subcircular opening in Euromycter rutenus. The dorsal surface is ornamented with the pits and furrows seen on most of the other roofing elements.
The postfrontal of Cotylorhynchus romeri is a broadly crescentic plate of bone that forms the posterodorsal corner of the orbit (Figures 6–8). Compared to the surrounding roofing elements, the postfrontal of Cotylorhynchus romeri is intermediate in relative size compared to the relatively larger postfrontal of Euromycter rutenus and the reduced postfrontal of Ennatosaurus tecton. As in the last caseid, the postfrontal of Cotylorhynchus romeri is almost entirely restricted to the dorsal skull table apart from a ventrally extending process that contributes to the postorbital bar. Although slightly separated from the prefrontal by the frontal dorsally, the anterior tip of the postfrontal closely approaches the prefrontal ventral to the frontal, and on the right side of OMNH 04329 makes a point contact with the prefrontal. The dorsal surface of the postfrontal is entirely ornamented with subcircular pits and furrows.
None of the available postorbitals are complete but together they afford a full appreciation of the morphology of this element (Figures 1, 6–8). The postorbital can be subdivided into an anterior portion that lies ventral to the postfrontal and forms the greater part of the posterior margin of the orbit and the upper half of the postorbital bar, and a larger, tongue-like temporal portion that forms the upper margin of the temporal opening, contacts the parietal and the supratemporal along an irregularly sigmoidal medial margin, and overlaps the anterodorsal corner of the squamosal. In these general respects it resembles the postorbitals of Euromycter rutenus and Ennatosaurus tecton. The dorsal surface of the postorbital is sculpted with the same system of pits and furrows seen on the surrounding skull table elements.
As in Euromycter rutenus and Ennatosaurus tecton, the supratemporal of Cotylorhynchus romeri is a relatively large, subrectangular plate of bone that is aligned anteromedially in the posterolateral corner of the skull roof (Figures 6–9). It is primarily a skull roof element, with a minor contribution to the posterodorsal rim of the occiput. It has its most extensive contacts with the parietal anteromedially and the postorbital laterally. Posterolaterally it overlaps the dorsal margin of the squamosal, and directly posteriorly it overlaps the tabular laterally and forms a short, slightly curving suture with the postparietal medially. Its dorsal surface bears a sparser complement of the shallow pits and furrows that ornament most of the other skull roof elements.
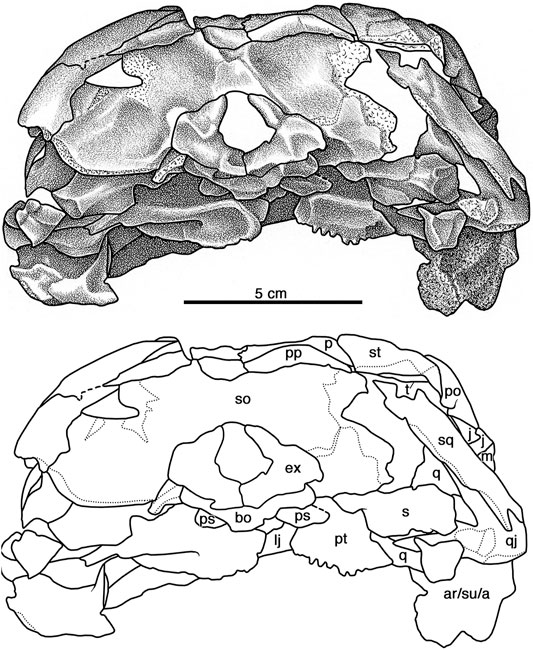
FIGURE 9. Cotylorhynchus romeri, OMNH 04329, referred specimen. Occipital view of skull. Abbreviations as in Figure 6.
OMNH 04329 preserves both postparietals (Figures 8, 9). It is a distinct, paired element, as in Ennatosaurus tecton (q.v. Maddin et al., 2008), versus the apparently single, median postparietal that is interpreted for Euromycter rutenus (Sigogneau-Russell and Russell, 1974). The postparietal of Cotylorhynchus romeri is autapomorphic among caseids in being transversely broad: it extends laterally from the midline, making an extensive suture with the posterior margin of the parietal and extending beyond the parietal to make a rather extensive contact with the supratemporal that aligns with the occipital rim. In both dorsal or occipital views the postparietal has a pennant-like profile, narrowing progressively laterally from its greatest height at the midline. It is entirely an occipital element. The smooth occipital surface of the postparietal parallels the curvature of the occipital rim and slopes posteroventrally from the rim to overlap the dorsal margin of the supraoccipital from the midline to the post-temporal opening. Directly dorsal to this opening, the postparietal shares a short, straight suture with the tabular (Figure 9).
The squamosal is a tall, quadrangular element that forms the posterodorsal corner of the temporal region of the skull (Figures 6–9). Whereas Maddin et al. (2008) ascribed a short anterior ramus and a long ventral ramus to the squamosal of Ennatosaurus tecton, the squamosal of Cotylorhynchus romeri narrows slightly from its maximum breadth dorsally along its contacts with the postorbital, the supratemporal, and the tabular. Its shallowly concave anterior margin froms the posterior border of the temporal fenestra. The posterior margin of the squamosal drops vertically from its contact with the tabular at the posterodorsal corner of the temporal region and extends ventrally down to the condylar region of the skull. Ventrally the squamosal overlaps the posterodorsal margin of the quadratojugal, but neither element in OMNH 04329 is well preserved in this area and we have tentatively reconstructed the ventral margin of the squamosal as a faintly convex, obliquely aligned edge of bone. Whereas the dorsal third of the lateral surface of the squamosal bears the dermal sculpturing seen on most skull-roofing elements, the remainder of the lateral surface of the bone is smooth.
The morphology of the tabular of Cotylorhynchus romeri is quite distinct from that of other caseids in which this element is known. It is tall, curved, crescent of flat bone that forms the posterodorsal rim of the occiput (Figures 8, 9). Dorsally it is a narrow, attenuating triangle of bone that underlies the supratemporal and the postparietal and forms the dorsal margin of the post-temporal opening. The bone widens as it curves posteroventrally, completing the occipital rim laterally and making extensive overlapping contacts with the squamosal laterally and the supraoccipital medially. Thus, the tabular of Cotylorhynchus romeri is more extensive than this element in either Ennatosaurus tecton or Euromycter rutenus, which has been interpreted as relatively small occipital elements that lie entirely dorsal to the post-temporal opening (Ennatosaurus tecton) or slightly broader and contacting the supratemporal and the paraoccipital process of the supratemporal (Euromycter rutenus). However, neither makes a conspicuous lateral contact with the squamosal as the tabular does in Cotylorhynchus romeri.
The quadratojugal of Cotylorhynchus romeri is triangular element (Figures 1, 6–8) that closely resembles that of Euromycter rutenus. It forms the posteroventral corner of the skull roof and the greater part of the subtemporal bar. The posterior portion of the quadratojugal is an extensive, slightly curving plate of bone that contacts the quadrate medially and is extensively overlapped by the ventral tip of the squamosal. A distinctive feature of the quadratojugal is an acuminate occipital process that extends medially from the ventrolateral corner of the facial part of the bone to overlie the posterodorsal surface of the condylar portion of the quadrate. Such an occipital process of the quadratojugal is not present in Ennatosaurus tecton, and although its presence is indeterminate in Euromycter rutenus, no other early amniote exhibits this quadratojugal morphology and thus we interpret this feature as an autapomorphy of Cotylorhynchus romeri. The right elements of OMNH 00637 (Figure 1) and OMNH 04329 (Figures 6, 7) together indicate that the quadratojugal extended dorsally under the squamosal at least as far as the midpoint of the temporal fenestra, a feature that is also noted to be present in Euromycter rutenus and in Eocasea martini. Anterior to its contact with the squamosal, the quadratojugal narrows progressively anteriorly, forming the posterior portion of the subtemporal bar and farther anteriorly to form scarf joints with the jugal and the maxilla. Medial to the maxilla, the quadratojugal extends anteriorly almost as far as the base of the postorbital bar. Together with the maxilla, the quadratojugal excludes the jugal from the ventral margin of the skull.
The vomer is preserved only in OMNH 04329. Although it is not completely exposed in palatal aspect (Figure 8) because of the occluded lower jaws, an oblique view of the palate reveals that the vomer is a paired bone. Each bears a short row of three large gently recurved teeth, taller than any other teeth on the palatal surface.
Both palatines are preserved in articulation with surrounding palatal and skull-roof elements in OMNH 04329. The holotype OMNH 00637 preserves a smoothly finished bone fragment that may be the remains of the right palatine. The right palatine of OMNH 04329 is well exposed in palatal aspect (Figures 3, 8), and both palatines can be viewed through the orbits in dorsal aspect (Figures 3, 8). In palatal aspect the palatine is a flat element that is deeply incised anteriorly by the internal naris and features an anteroposteriorly aligned cluster of palatal teeth on a slightly swollen mound of bone. As in other early synapsids, the palatine contacts the vomer anteriorly, the pterygoid posteromedially, the ectopterygoid posterolaterally, and both the maxilla (ventrally) and the lacrimal (dorsally) laterally. The anterior end of the palatine forms a thin, tongue-like process that overlaps the posterior end of the vomer. The suture with the pterygoid is broadly sigmoidal (Figures 3, 8). Immediately lateral to the suture with the pterygoid the posterolateral margin of the palatine angles anterolaterally and narrowly overlaps the anteromedial edge of the ectopterygoid along a relatively straight line of contact (Figures 3, 8). The palatine contacts the skull roof along a relatively straight anterolateral margin, which sutures to a ventromedial process of the lacrimal and, directly ventrally, shares a more extensive anteroposterior contact with the medial surface of the maxilla (Figure 5). Ten subconical teeth are present on the slightly thickened region of bone adjacent to the middle part of the suture shared with the pterygoid, the palatine’s contribution to the pterygo-palatine tooth cluster that is usually present on the palate of most early amniotes.
The ectopterygoid is a flat tongue of bone positioned at the anterior end of the subtemporal fossa. Its anteromedial margin is entirely sutured to the palatine anteriorly (Figures 3, 8), and the medial end of the ectopterygoid forms an overlapping contact with the pterygoid (Figure 8). The approximate lateral two-thirds of the posterior margin of the ectopterygoid contributes to the palatal margin of the subtemporal fossa. Laterally the ectopterygoid has straight sutures with the jugal (dorsally) and the maxilla (ventrally). The dorsal surface of the ectopterygoid is flat and featureless, and the ventral surface is devoid of teeth (Figures 3, 8).
OMNH 04329 preserves both pterygoids in complete articulation with the surrounding palatal, skull roof, and palatoquadrate regions (Figures 3, 8). In ventral aspect, each pterygoid is a triradiate element with a broadly triangular anterior portion, an acutely triangular quadrate process that extends posterolaterally, and between them a quadrangular mass of bone that forms the basicranial recess medially and transverse process laterally. The anterior portion has an extensive contact laterally with the palatine and makes smaller contacts with the vomer directly anteriorly and the contralateral bone anteromedially. Posterior to the last contact, the medial margin of the pterygoid extends posteriorly and slightly laterally to the basicranial region, and together the paired pterygoids form a narrow interpterygoid vacuity. The palatal surface of the pterygoid bears teeth in four identifiable groupings: a medial cluster that borders the interpterygoid vacuity, a smaller grouping of teeth that contributes to the pterygo-palatine tooth cluster, a posterolateral cluster of small teeth on the transverse flange, and a discernible row of large teeth that borders the posterior margin of the transverse flange and extends medially to the basicranial region. The medial tooth cluster is similar to those of Euromycter rutenus and Ennatosaurus tecton, but features some of the largest palatal teeth, positioned irregularly up to three teeth across anteriorly, with the cluster narrowing posteriorly to teeth aligned in a single row posterior to the approximate midpoint of the interpterygoid vacuity. The teeth of the pterygo-palatine cluster are largest anteriorly and progressively diminish in size posteriorly to the degree that the smallest teeth are roughly one-third the basal diameter of the largest teeth anteriorly; Ennatosaurus tecton differs in that teeth of varying sizes are implanted randomly throughout the cluster. Approximately 20 small teeth form an anteromedially aligned, elliptical grouping on the transverse flange, akin to a similar cluster of small teeth in Euromycter rutenus (Sigogneau-Russell and Russell, 1974, Figure 5); Ennatosaurus tecton does not exhibit this cluster of small teeth (Maddin et al., 2008). A row of 14 teeth (plus one alveolus) extends along the posterior margin on the transverse flange on the well exposed right pterygoid (Figure 6). The largest teeth in this row are implanted opposite the base of the quadrate flange, and the teeth progressively diminish in size (basal diameter) both medially at the basicranial recess and laterally towards the apex of the transverse flange. This condition contrasts with the double row of transverse flange teeth documented for Euromycter rutenus, and the narrow cluster of teeth of varying size described for Ennatosaurus tecton (q.v. Maddin et al., 2008).
Although technically not an element of the palate, the quadrate is described here because it bridges the palate proper to the posterior of the skull roof. The left quadrate of OMNH 04329 is reasonably well exposed because of the loss of most of the quadratojugal and the squamosal on that side (Figures 7, 8). Here the quadrate is seen to be normally developed for an early synapsid in being comprised of a compact condylar region and a plate-like lamella. The condylar region is broader transversely than it is anteroposteriorly long. Although the mandible is tightly occluded and as a result the articulating surface for the jaw joint is not exposed, the contact with the articular suggests that the articulating surface of the quadrate is approximately 33% broader transversely than long anteroventrally. In this respect the dimensions of the articulating surface of the quadrate of Cotylorhynchus romeri is notably narrower (in anteroposterior dimension) than the articulating surface of the quadrate of Ennatosaurus tecton (q.v. Maddin et al., 2008, Figure 1C). In posterior aspect the condylar portion of the quadrate is overlain by the ventral part of the quadratojugal, which extends a pennant-like process directly medially across the posterodorsal surface of the condylar portion; immediately dorsal to the base of this medial process of the quadratojugal is the posterior opening of the quadrate foramen. Immediately anterodorsal to the condylar region the quadrate narrows transversely and expands dorsoventrally as it transitions to the plate-like dorsal lamella. In ventral aspect the dorsal lamella is aligned anteromedially, contacts the quadrate flange of the pterygoid for most of its length, and extends anteriorly beyond the transverse flange of the palate to make a slight overlapping contact with the epipterygoid (Figure 7). The posterior margin of the dorsal lamella is aligned subvertically and forms narrow contacts with the quadratojugal and the squamosal.
The following description of the braincase elements of Cotylorhynchus romeri is based on that of OMNH 04329, which preserves an articulated and nearly complete braincase. As in most early amniotes, the parasphenoid and the basisphenoid are fused together indistinguishably and is here referred to as the parabasisphenoid. Both prootics are presumably preserved in OMNH 04329, but neither is exposed and are not described in this work. Nothing of the braincase of OMNH 00637 is known.
The parabasisphenoid of OMNH 04329 is well preserved but is accessible primarily in ventral aspect (Figure 8). Its ventral profile resembles more closely that of Euromycter rutenus than that of Ennatosaurus tecton and this is largely due to the wing-like cristae ventrolaterale. Anteriorly the parabasisphenoid projects a broad-based cultriform process of indeterminate length. The ventral surface of the cultriform process houses several teeth that form an irregular row along the midline of the process and extend posteriorly on a ventral swelling of bone to a point level with the neck of the parabasisphenoid. Cotylorhynchus romeri appears to differ from other caseids, including C. hancocki and Angelosaurus romeri, in having few teeth on the parasphenoid. In particular, this feature appears to distinguish the two large species of Cotylorhynchus (Olson, 1968).
Whereas the left basipterygoid process is totally obscured by the left pterygoid, the right basipterygoid process is partly exposed in ventral aspect, but little can be said about this process beyond the observation that is relatively broad in anteroposterior dimension. Measured slightly obliquely from the posterior margin of the basipterygoid process to the point at which the parasphenoid emerges anterior to the basipterygoid recess of the pterygoid, the anteroposterior breadth of right basipterygoid process is about one-third the maximum width of the parabasisphenoid, as measured across the cristae ventrolaterale. Immediately posterior to the basipterygoid processes the bone narrows to a distinctive neck and then expands laterally posteriorly to produce the paired, wing-like cristae ventrolaterales. In ventral aspect, each crista ventrolateralis forms a low, rounded ridge that gradually swells in size posterolaterally, and between the paired cristae lies a shallow, triangular fossae. The posterior margin of each crista is a slightly sigmoidal free edge of bone that merges medially with the main body of the element. Unlike the conditions in either Euromycter rutenus or Ennatosaurus tecton, the main body of the parabasisphenoid extends slightly posterior to the cristae ventrolaterale to form a coarsely sigmoidal contact with the basisphenoid.
In contrast to the conditions in both Euromycter rutenus and Ennatosaurus tecton, the basioccipital of Cotylorhynchus romeri is a relatively broad element, being 25% greater in transverse dimension than anteroposteriorly long (Figure 8). Anteriorly the basioccipital contacts the parabasisphenoid across a slightly swollen contact, narrows slightly posteriorly, and then progressively extends laterally towards the posterolateral corner of the bone where the basioccipital contributes to the jugular foramen. In ventral aspect, the posterior margins of the bone form a very obtuse angle that is interrupted medially by the lozenge-shaped occipital condyle, which extends anteriorly on the ventral surface of the basioccipital as a triangular swelling that merges with the surrounding ventral surface about half way to the anterior margin. The basioccipital forms a butt contacts with the opisthotics laterally, and posteriorly it underlies the paired exoccipitals, which form the dorsal rim of the occipital condyle (Figures 8, 9). The openings for the hypoglossal nerves (cranial nerve XII) are not discernible.
The opisthotic is an irregular bone with a large medial portion that contacts the basioccipital, the exoccipital, the supraoccipital, and the stapes, and a lateral, plate-like paroccipital process (Figures 8, 9). The ventral surface of the medial portion exhibits a thick ridge that receives the medial rim of the stapedial head; little else can be said of the ventral surface of the bone in this region because of overlying elements and surrounding matrix. Directly medially the opisthotic is bounded by the exoccipital and dorsomedially is fused indistinguishably to the supraoccipital. The greater part of the opisthotic exposed in posterior (occipital) aspect is the paroccipital process. Dorsally, this process forms the ventral margin of the posttemporal opening, and dorsolaterally its forms an extensive contact with the tabular. At the ventrolateral tip of the contact with the tabular, the lateral margin of the paroccipital process extends directly ventrally to the level of the condyle, and then the ventral margin of the process curves slightly dorsally to end medially where the opisthotic is notched at the point this element contributes to the jugular foramen. The occipital surface of the paroccipital process is shallowly scooped out (Figure 9), presumably for the former attachment of rectus capitis muscles.
The paired exoccipitals form a robust horseshoe-shaped structure in occipital aspect that forms the greater part of the foramen magnum (Figures 8, 9). The exoccipitals sit atop the posterior quarter or so of the basioccipital and contact each other ventrally along the midline and form the dorsal portions of the occipital condyle. The exoccipitals floor at least 1 cm of the cavum cranii judging from what can be seen of the contributions of the exoccipitals through the foramen magnum in dorsal aspect (Figures 3, 8). Lateral to the condylar portion the ventral portion each exoccipital extends dorsolaterally and expands in height. Ventrally the exoccipital extends laterally slightly beyond the contact with the basioccipital to make a small contribution to the rim of the jugular foramen and contact the base of the paroccipital process of the paroccipital process. The exoccipital exhibits its greatest transverse breadth between the floor of the braincase and the ventral-most point of contact with the opisthotic. The exoccipital gradually narrows dorsally and then, at a point level roughly level to the midpoint of the foramen magnum, the bone narrows progressively to an acuminate tip. The exoccipital of Ennatosaurus tecton is not known. The exoccipitals of Euromycter rutenus are fused to each other and to the basioccipital, and the exoccipitals appear to be autapomorphic in being widely separated dorsally across the foramen magnum (Sigogneau-Russell and Russell, 1974, Figure 6).
The supraoccipital is a low, wide plate of bone that is bounded dorsally by the postparietals, laterally by the posttemporal openings, and ventrally by the opisthotics, the exoccipitals, and the foramen magnum (Figures 8, 9). The occipital surface is excavated by a pair of fossae, presumably for the former attachment of rectus capitis muscles, that are separated at the midline by a swelling of bone that attenuates dorsally. The supraoccipital of Ennatosaurus tecton is not known well enough for comparison. That of Euromycter rutenus is relatively taller and slightly narrower in occipital view and appears to have a relatively greater contribution to the dorsal margin of the foramen magnum that does the supraoccipital of Cotylorhynchus romeri.
Stapedial morphology is poorly documented for caseids. Fortunately, both stapes are preserved in OMNH 04329 (Figures 8, 9). As in most early amniotes, the stapes of Cotylorhynchus romeri consists medially of an expanded footplate and laterally of a posteroventrally-directed shaft. The footplate contacts the parabasisphenoid, the basioccipital, and the opisthotic; we cannot determine if the footplate also contacts the prootic because both the latter bone and the dorsal part of the footplate are inaccessible. The shaft of the stapes is lenticular to quadrangular in cross section, with a thickened, middle portion that thins more dorsally than ventrally. The dorsal process arises from the thinning dorsal surface of the shaft of the left stapes, but because of the overlying paroccipital process of the left opisthotic, nothing else of the dorsal process can be described; the dorsal process of the right stapes is not accessible. Opposite the dorsal process, a low, keel-like flange extends along the ventral margin of the shaft. This keel-like flange has not been described heretofore among caseids, and may represent an attachment point for connective tissue that, in life, extended to the hyoid apparatus.
The dentary is a low tooth-bearing element that occupies approximately two-thirds of the length of the lateral surface of the lower jaw (Figures 1, 6–8). It is roughly rectangular anteriorly, gradually expanding in height posterior to the symphysial region and reaching its greatest height immediately anterior to the contact with the angular. Posterior to the 11th tooth position, and concomitant with the contact with the angular, the bone curves posterodorsally, reaching its highest point at the point where it contacts the coronoid and the surangular, and posterior to which the dentary attenuates conspicuously to a sharp posterior tip that overlaps the lateral surface of the surangular. The lateral surface is smooth and featureless apart from the presence of a few labial foramina. The ventral margin of the element is entirely excluded from the ventral margin of the mandible by the splenial. The dentary forms the greater part of the mandibular symphysis. The symphysial pad of the dentary has a robustly lenticular or pyriform profile in medial aspect (Figure 4). Immediately posterior to the symphysis, the dentary, together with the splenial, is excavated medially for the foramen intermandibularis medius. Ventral to the fifth tooth position, the splenial closes off the foramen intermandibularis medius and overlaps the alveolar portion of the dentary. The medial exposure of the dentary is progressively diminished posteriorly as the splenial increases in height, and then is reduced to a narrow strip of bone dorsal to the elongate coronoid. The right dentary of OMNH 00637 exhibits 19 tooth positions, and we estimate that the dentaries of OMNH 04329 each supported approximately 16 teeth. These teeth exhibit the same morphology as the teeth in the upper jaw and, like those teeth, exhibit a progressive decrease in size posteriorly.
Between OMNH 00637 and OMNH 04329 there are three splenials that together afford a nearly complete understanding of this element (Figures 1, 8, 9). In lateral aspect the symphysial portion of the splenial forms a distinct chin, as in Ennatosaurus tecton, and the splenial narrowly attenuates posteriorly to a point contact with the angular. In medial aspect, the symphysial pad of the splenial has a comma-shaped outline. The medial surface of the splenial curves laterally and then posteriorly, forming the ventral half of the foramen intermandibularis medius. At the level of the third tooth position, the splenial thickens to give rise to dorsally directed flange that forms the posterior margin of the foramen intermandibularis medius and gradually rises in height to make contact with and underlie the coronoid. Ventrally the splenial continues posteriorly to form the ventral margin of the mandible and the bone pinches out ventral to the level of the foramen intermandibularis caudalis (“Meckelian foramen” of Reisz and Scott, 2002; Maddin et al., 2008). The splenial is deeply bisected by the prearticular in medial aspect, and presumably the former element is deeply overlain by the latter.
As noted by previous workers (Romer and Price, 1940; Laurin and Reisz, 1995; Maddin et al., 2008), Cotylorhynchus romeri exhibits a single coronoid in each mandibular ramus (Figures 1, 8). The coronoid is a crescentic blade of bone that caps the coronoid eminence and extends anteriorly as far as the seventh mandibular tooth position. The dorsal margin descends from the coronoid eminence to border the ventral margins of the resorption pits of the posterior-most 13 dentary teeth. From the acuminate anterior end of the bone, the ventral margin of the coronoid extends posteriorly over a small portion of dentary, the splenial, and the approximate middle third of the prearticular. The posterior margin of the coronoid is conspicuously notched where it forms the anterior end of the adductor fossa, and the remainder of the posterior margin meanders dorsally along the contact with the surangular to finally join the free margin of the bone at the coronoid eminence. The coronoid has a very narrow lateral exposure at the apex of the coronoid eminence. In contrast to Casea broilii and Ennatosaurus tecton, which exhibit coronoid dentition to various degrees (Williston, 1910; Romer and Price, 1940; Maddin et al., 2008), the medial surface of the coronoid of Cotylorhynchus romeri is smooth and featureless.
The surangular of Cotylorhynchus romeri is distinctive among caseids in overlapping the posterodorsal tip of the dentary and excluding the latter bone from the coronoid eminence (Figures 1, 6, 7). The anterodorsal corner of the surangular forms the base of the coronoid eminence in lateral aspect. Anteroventrally the surangular extends slightly farther anteriorly than anterodorsally, and shares an overlapping contact with the dentary along a slightly irregular, weakly convex suture. In lateral aspect, the surangular of Cotylorhynchus romeri appears to extend as far ventrally as the surangular of Euromycter rutenus, in contrast the relatively low surangular of Ennatosaurus tecton (q.v. Maddin et al., 2008). In neither specimen is the suture between the surangular and the angular clear, but what is preserved suggests that laterally the contact was roughly straight and parallel to the ventral margin of the mandible. Posteriorly the surangular sheathed the lateral surface of the articular. The surangular is thickest dorsally where it forms the dorsal margin of the adductor fossa. The dorsal margin descends and thins gradually posteriorly from a high point at the coronoid eminence. At the three-quarter mark, the dorsal margin of the surangular expands medially to abut the anterior end of the articular; in lateral aspect this manifests as a small, rounded protuberance immediately anterior to the jaw articulating surface (Figure 7).
The angular forms the posterventral portion of each lower jaw and is the second largest element of the mandible after the dentary (Figures 1, 8). Posteriorly it overlaps the lateral surface of the articular and from there extends anteriorly as a trough-like structure. The lateral portion of the trough extends dorsally as a low flange that contacts the surangular posteriorly along a roughly straight, horizontal contact, and the dentary anteriorly via an overlapping contact that results in the angular attenuating in height and pinching out in lateral aspect where it contacts the splenial. In ventral and medial views (Figures 1, 8) the medial portion of the angular trough extends dorsally to form the base of the medial wall of the adductor fossa and its contacts the prearticular along almost its entire dorsal margin. The contact between the angular and the prearticular is interrupted by a low, elliptical foramen intermandibularis caudalis. The formation of the foramen intermandibularis caudalis by only the angular and the prearticular is seen also in Ennatosaurus tecton (q.v. Maddin et al., 2008) among caseids, and in Aerosaurus wellesi (q.v. Langston and Reisz, 1981) and Ophiacodon (Romer and Price, 1940) among other early synapsids. Ventral to the contact with the prearticular anterior to the foramen, the angular forms a widely sigmoidal suture with the splenial in medial aspect.
The prearticular is a long, low, plate of bone on the medial surface of the lower jaw (Figure 1). In ventral aspect the bone has a subtriangular profile where it sheathes most of the ventral surface of the articular. Anterior to the contact with the articular, the prearticular transitions to a vertically aligned plate of bone that, in medial aspect, forms the dorsal margin of the medial wall of the adductor fossa and overlies the angular. Anterior to the midpoint of adductor fossa, the prearticular gradually increases in height, makes contact with the coronoid at the anterior end of the adductor fossa, and underlies the latter bone as it continues anteriorly for a couple of centimeters until it reaches the dorsal part of the splenial. Ventrally, the prearticular forms a relatively small foramen intermandibularis caudalis with the angular and continues anteriorly beyond that to finally make contact with the ventral portion of the splenial. Anteriorly the prearticular sheathes the medial surface of the splenial and progressively attenuates to an acuminate tip almost directly ventral to the anterior-most point of the coronoid.
Both articulars are present in OMNH 04329, but only the left element is preserved well enough for description (Figures 6–9). It is a low, blocky structure sandwiched into its position at the posterior end of the mandibular ramus by the prearticular, the surangular, and the angular. The dorsal surface, although not fully exposed because of the articulated quadrates, is clearly dominated by the articulating surface for the jaw joint. In posterior view the articulating surface is a rounded W-shape, forming rounded fossae for the condyles of the quadrate and bounded by a sharp lip of bone. In the left lateral view the intercondylar ridge can be seen to rise anterodorsally to meet the medial protuberance of the surangular. Compact bone has been eroded from the posterior surfaces of both articulars, but what remains suggests that the articular supported a modest retroarticular process.
As one of the first described and best represented large caseids, Cotylorhynchus romeri has factored into every consideration of the evolutionary history of the family Caseidae (Stovall et al., 1966; Olson, 1968; Maddin et al., 2008; Reisz and Fröbisch, 2014; Romano and Nicosia, 2015; Romano et al., 2017). In the strict consensus trees of the most inclusive phylogenetic systematic investigations of Caseidae (e.g., Maddin et al., 2008; Reisz and Fröbisch, 2014; Romano and Nicosia, 2015; Romano et al., 2017), Cotylorhynchus romeri forms polytomies with its congeners Cotylorhynchus bransoni and Cotylorhynchus hancocki, with Angelosaurus romeri (and Angelosaurus dolani when included), and with Alierasaurus ronchii (when included).
Upon undertaking an anatomical description of the cranial anatomy of Cotylorhynchus romeri, we anticipated the discovery of additional characters of phylogenetic usefulness that might help to resolve the uncertainties that continue to dog the interrelationships of the largest caseids. We were gratified to discover several apomorphic features of the skull of Cotylorhynchus romeri and we collated them as autapomorphies for this species (see Diagnosis). However, we recognize that these characters are ambiguous autapomorphies for Cotylorhynchus romeri because cranial materials are sparse or totally unknown for Cotylorhynchus bransoni, Cotylorhynchus hancocki, Angelosaurus dolani, Angelosaurus romeri, and Alierasaurus ronchii. For this reason, we feel that the new anatomical information gleaned in our description of the cranial skeleton of Cotylorhynchus romeri does not warrant a new phylogenetic analysis of Caseidae at this time beyond what has been recently published (Berman et al., 2020). However, going forward the new data will be very useful for considerations of amniote and synapsid phylogenetic analyses. For example, the presence of a well developed dorsal process of the quadratojugal extending beneath the squamosal is present in Cotylorhynchus romeri, a feature that is absent in non-synapsid amniotes and stem amniotes. This potential synapsid character is also known to be present in Ophiacodon, Edaphosaurus, and Dimetrodon. The distribution of this character among caseids is currently unknown and merits investigation.
Cotylorhynchus romeri long has been regarded as one of the largest terrestrial herbivores of the early Permian (e.g., Romer and Price, 1940). This animal has been estimated to have reached a maximum length of about 4.5 m and a mass of 330 kg (Stovall et al., 1966). Whereas Romer and Price (1940, p. 377) described the marginal teeth as blunt, the tips of the crowns bear several cuspules, as do the marginal teeth of its more recently described relatives Euromycter rutenus and Ennatosaurus tecton (Sigogneau-Russell and Russell, 1974; Maddin et al., 2008). The observation that the marginal teeth of Cotylorhynchus romeri each bear only a few cuspules suggests that this species was adapted to a different fodder (or range of fodder) than Euromycter rutenus and Ennatosaurus tecton (Sigogneau-Russell and Russell, 1974; Maddin et al., 2008).
An intriguing feature of the dentition of the known Cotylorhynchus romeri specimens is that the tooth rows lack replacement gaps. Typically, Permo-Carboniferous amniotes have multiple gaps in their dentition as a result of polyphyodonty, their continual, relatively rapid tooth replacement pattern. The presence of gaps in the tooth row is in part a reflection of the nature of fossil preservation, with fully ankylosed marginal teeth being preserved whereas replacement teeth are frequently lost because they were not yet ankylosed to the jaw elements. Cotylorhynchus romeri appears not to show this pattern, with no discernable gaps in the marginal dentition, a pattern that appears to be also present in other caseids (Olson, 1968; Sigogneau-Russell and Russell, 1974; Maddin et al., 2008). This may be a reflection of two aspects of caseid paleobiology: reduced rates of tooth replacement and increased longevity of functional teeth. Histological sections of caseid marginal dentition may help determine the reason for this potential adaptation to herbivory.
The discrepancy between the articulating surface on the quadrate condyles and the receiving cotyles on the articular hints at the possibility of fore-and-aft translation of the mandible, but the lack of conspicuous wear on the marginal teeth and the absence of tooth plates argues against the kind of oral processing inferred for edaphosaurid synapsids (Modesto, 1995) and moradisaurine reptiles (Dodick and Modesto, 1995). Moreover, the presence of tall, relatively slender palatal teeth in Cotylorhynchus romeri is puzzling. These teeth must have worked against a presumably tough tongue. We tentatively propose that Cotylorhynchus romeri used its marginal teeth to crop mouthfuls of plant matter, and then used the tongue to press mouthfuls of food against the palate in order to perforate the food with the tall palatal teeth, an action that may have served to enhance cellulolytic fermentation of the food in the gut.
Cotylorhynchus romeri is the largest member of a predominantly terrestrial fauna of the Hennessey Formation of Oklahoma. Specimens of the lysorophid Brachydectes elongatus (Lysorophus tricarinatus of Olson, 1967), the captorhinid reptile Captorhinikos parvus, and nectridean Peronedon primus are abundant in this formation (Olson, 1967; Olson, 1970; Haglund, 1977). The captorhinid reptile Captorhinikos chozaensis and the microsaurs Rhynchonkos stovalli, Aletrimyti gaskillae, and Dvellecanus carrolli are known from single specimens (Vaughn, 1958; Szostakiwskyj et al., 2015). Finally, a partial skull, a partial tooth plate, and indeterminate cranial remains of the lungfish genus Gnathorhiza were listed by Olson (1967). Specimens of the captorhinid reptile Labidosaurikos meachami and the sphenacodontid Dimetrodon giganhomogenes have been reported from the Hennessey Formation (e.g., Seltin, 1959) but according to Olson (1967) these specimens were collected from what he termed a 15-m-thick “transitional zone” that conformably overlies the Garber Formation and is not part of the “Hennessey proper” (Olson, 1967, p. 84). Thus, Cotylorhynchus romeri is the largest tetrapod of a terrestrial fauna that, in the words of Olson (1967), is “odd” because of the total absence of large carnivorous vertebrates.
Recently, Lambertz et al. (2016) challenged the view that Cotylorhynchus romeri and other large caseids were terrestrial. On the basis of postcranial histology (“osteoporotic” cancellous bone structure), they proposed that Cotylorhynchus romeri and other large caseids were aquatic. A critical appraisal of Lambertz et al.’s (2016) osteohistology is beyond the scope of this work (a description of the cranial skeleton of Cotylorhynchus romeri), but we find it remarkable that the gross anatomy of the entire skeleton of Cotylorhynchus romeri exhibits not a single unequivocal adaptation to an aquatic lifestyle.
In support of their argument that large caseids were aquatic, Lambertz et al. (2016) argued that, despite their remains being preserved in strata that have traditionally been interpreted as recording an arid upland habitat, large caseid burials were entirely allochthonous, i.e. they did not live where they were buried. However, Lambertz et al. (2016) argument here is not circumspect. Stovall et al. (1966, p. 3) remarked that the preservation of specimens of Cotylorhynchus romeri in the Hennessey Formation was quite different to that of Texas redbeds localities, where tetrapods are usually found disarticulated and when articulated remains are found, they “appear to have undergone decomposition in a variety of poses” that those authors attribute to rivers or streams transporting the remains and depositing them in back eddies. In contrast, several specimens of Cotylorhynchus romeri were found in the Hennessey Formation dorsal up with their limbs spread outwards as though the animals “were bogged down and entombed in swamps” (Stovall et al., 1966, p. 3), burials that are clearly autochthonous. This preservational mode accords well with the geology and the taphonomy of the formation if Cotylorhynchus romeri is a terrestrial tetrapod that lived in a hot, dry environment that was punctuated periodically by monsoons and subjected to occasional massive flooding events. The red mudrocks (“shales”) of the Hennessey Formation have traditionally been interpreted as having been deposited in hot, dry conditions (Olson, 1967). A monsoon climate is suggested by the abundant skeletons of the aestivator Brachydectes, which Olson (1967) noted is preserved in nodules in such great numbers that Olson (1967) described them as “swarms,” and by the scarce remains of the lungfish Gnathorhiza, another well-known aestivator. Together, the presence of terrestrial reptiles and microsaurs, the presence of aestivators (Gnathorhiza, Brachydectes) and the absence of obligatorily aquatic vertebrates strongly suggests that the Hennessey fauna lived in a dry habitat that was periodically (presumably seasonally) punctuated by monsoons. It seems highly improbable that evolution would produce a large caseid, one which exhibits no gross anatomical aquatic adaptations for an aquatic lifestyle, that was a primarily aquatic, very large tetrapod in a monsoonal habitat. It is much more likely that Cotylorhynchus romeri was a terrestrial animal that had to endure monsoonal rains and the concomitant flooding of low-lying areas, with some individuals occasionally succumbing to major flooding events (to be preserved as articulated skeletal remains).
The “osteoporotic” cancellous bone structure of caseids described by Lambertz et al. (2016) is intriguing, but we feel that further comparative osteohistology among early tetrapods is needed before a thorough understanding of the differences in cancellous bone structure can be assessed for paleobiological implications.
OMNH, Sam Noble Museum of Natural History, University of Oklahoma, Norman, Oklahoma, United States.
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Project conceived by RR, specimen preparation, illustration, and reconstructions by DS, manuscript preparation and editing by RR and SM.
RR is supported by NSERC Discovery Grant from the Natural Sciences and Engineering Research Council (NSERC) of Canada and by Jilin University, China. SM is supported by a New Opportunities Fund award from the Canadian Foundation for Innovation and a grant from the Nova Scotia Research and Innovation Trust.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank R. Cifelli of the Sam Noble Museum of Natural History, University of Oklahoma, for the loan of specimens.
Berman, D. S., Maddin, H. C., Henrici, A. C., Sumida, S. S., Scott, D., and Reisz, R. R. (2020). New Primitive Caseid (Synapsida, Caseasauria) from the Early Permian of Germany. Ann. Carnegie Mus. 86, 43–75. doi:10.2992/007.086.0103
Bingham, R. H., and Moore, R. L. (1975). Reconnaissance of the Water Resources of Oklahoma City Quadrangle. Central Oklahoma: Oklahoma Geological Survey Hydrologic Atlas 4, scale 1:250,000, 4.
Czaplewski, N. J., Cifelli, R. L., and Langston, W. (1994). Catalog of Type and Figured Vertebrates, Oklahoma Museum of Natural History. Norman, Okla: Oklahoma Geological Survey, University of Oklahoma, 35.
Dodick, J. T., and Modesto, S. P. (1995). The Cranial Anatomy of the Captorhinid Reptile Labidosaurikos meachami from the Lower Permian of Oklahoma. Palaeontology 38, 687–711.
Ford, D. P., and Benson, R. B. J. (1995). The Phylogeny of Early Amniotes and the Affinities of Parareptilia and Varanopidae. Nat. Ecol. Evol. 4, 57–65. doi:10.1038/s41559-019-1047-3
Haglund, T. R. (1977). New Occurrences and Paleoecology of Peronedon primus Olson (Nectridea). J. Paleont. 51, 982–985.
Lambertz, M., Shelton, C. D., Spindler, F., and Perry, S. F. (2016). A Caseian point for the Evolution of a Diaphragm Homologue Among the Earliest Synapsids. Ann. N.Y. Acad. Sci. 1385, 3–20. doi:10.1111/nyas.13264
Langston, W., and Reisz, R. R. (1981). Aerosaurus wellesi, New Species, a Varanopseid Mammal-like Reptile (Synapsida: Pelycosauria) from the Lower Permian of New Mexico. J. Vertebr. Paleontol. 1, 73–96. doi:10.1080/02724634.1981.10011881
Laurin, M., and Reisz, R. R. (1995). A Reevaluation of Early Amniote Phylogeny. Zool. J. Linn. Soc. 113, 165–223. doi:10.1111/j.1096-3642.1995.tb00932.x
Lucas, S. G., and Suneson, N. H. (2002). Amphibian and Reptile Tracks from the Hennessey Formation (Leonardian, Permian), Oklahoma County, Oklahoma. Okla. Geol. Notes 62, 56–62.
MacDougall, M. J., Modesto, S. P., Brocklehurst, N., Verrière, A., Reisz, R. R., and Fröbisch, J. (2018). Commentary: A Reassessment of the Taxonomic Position of Mesosaurs, and a Surprising Phylogeny of Early Amniotes. Front. Earth Sci. 6, 99. doi:10.3389/feart.2018.00099
Maddin, H. C., Sidor, C. A., and Reisz, R. R. (2008). Cranial Anatomy of Ennatosaurus Tecton (Synapsida: Caseidae) from the Middle Permian of Russia and the Evolutionary Relationships of Caseidae. J. Vertebr. Paleontol. 28, 160–180. doi:10.1671/0272-4634(2008)28[160:caoets]2.0.co;2
Modesto, S. P. (1995). The Skull of the Herbivorous Synapsid Edaphosaurus boanerges from the Lower Permian of Texas. Palaeontology 38, 213–239.
Olson, E. C., and Barghusen, H. (1962). Late Permian Terrestrial Vertebrates, U. S. A. And U. S. S. R. Trans. Am. Philos. Soc. 52, 1–68. doi:10.2307/1005904
Olson, E. C., and Beerbower, J. R. (1953). The San Angelo Formation, Permian of Texas and its Vertebrates. J. Geol. 61, 384–423. doi:10.1086/626109
Olson, E. C. (1954). Fauna of the Vale and Choza: 7. Pelycosauria: Family Caseidae. Fieldiana, Geology. 10, 193–204.
Olson, E. C. (1962). Late Permian Terrestrial Vertebrates, U. S. A. And U. S. S. R. Trans. Am. Philos. Soc. 52, 1–223. doi:10.2307/1005904
Olson, E. C. (1970). New and Little Known Genera and Species of Vertebrates from the Lower Permian of Oklahoma. Fieldiana, Geol. 18, 359–434.
Osborn, H. F. (1903). On the Primary Division of the Reptilia into Two Sub-Classes, Synapsida and Diapsida. Science 17, 275–276. doi:10.1126/science.17.424.275-c
Reisz, R. R., and Fröbisch, J. (2014). The Oldest Caseid Synapsid from the Late Pennsylvanian of Kansas, and the Evolution of Herbivory in Terrestrial Vertebrates. PLoS ONE 9, e94518. doi:10.1371/journal.pone.0094518
Reisz, R. R. (1986). “Pelycosauria,” in Handbuch der paläoherpetologie. Editor P. Wellenhofer (Stuttgart: Gustav Fischer Verlag), 17A, 1–102.
Reisz, R. R., and Scott, D. (2002). Owenetta kitchingorum, sp. nov., a Small Parareptile (Procolophonia: Owenettidae) from the Lower Triassic of South Africa. J. Vertebr. Paleontol. 22, 244–256. doi:10.1671/0272-4634(2002)022[0244:oksnas]2.0.co;2
Reisz, R. R. (2005). Oromycter, a New Caseid from the Lower Permian of Oklahoma. J. Vertebr. Paleontol. 25, 905–910.
Reisz, R. R., Maddin, H. C., Fröbisch, J., and Falconnet, J. (2011). A New Large Caseid (Synapsida, Caseasauria) from the Permian of Rodez (France), including a reappraisal of “Casea” rutena Sigogneau-Russell and Russell. Geodiversitas 33, 227–246. doi:10.5252/g2011n2a2
Reisz, R. R. (1980). “The Pelycosauria: a Review of Phylogenetic Relationships,” in The Terrestrial Environment and the Origin of Land Vertebrates. Editor A. L. Panchen (New York: Academic Press), 553–592.
Romano, M., and Nicosia, U. (2015). Cladistic Analysis of Caseidae (Caseasauria, Synapsida): Using the Gap-Weighting Method to Include Taxa Based on Incomplete Specimens. Palaeontology 58, 1109–1130. doi:10.1111/pala.12197
Romano, M., Ronchi, A., Maganuco, S., and Nicosia, U. (2017). New Material of Alierasaurus Ronchii (Synapsida, Caseidae) from the Permian of Sardinia (Italy), and its Phylogenetic Affinities. Palaeont. Electr. 20 (2), 1–27. Available at: http://www.palaeo-electronica.org/content/2017/1889-new-material-of-alierasaurus. doi:10.26879/684
Romer, A. S., and Price, L. W. (1940). Review of the Pelycosauria. Geol. Soc. Am. Sp. Pap. 28, 1–534. doi:10.1130/spe28-p1
Sigogneau-Russell, D., and Russell, D. E. (1974). Étude du premier caséidé (Reptilia, Pelycosauria) d’Europe occidentale. Bull. Mus. Nat. D’hist. Natur. Série 3 (230), 145–215.
Stovall, J. W. (1937). Cotylorhynchus romeri, a New Genus and Species of Pelycosaurian Reptile from Oklahoma. Am. J. Sci. Ser. 5 (34), 308–313.
Stovall, J. W., Price, L. I., and Romer, A. S. (1966). The Postcranial Skeleton of the Giant Permian Pelycosaur Cotylorhynchus romeri. Bull. Mus. Comp. Zool. Harv. Univ. 135, 1–30.
Suneson, N. H., and Hemish, L. A. (1998). Geologic Map of the Piedmont and Bethany NE 7.5’ Quadrangles, Kingfisher, Logan, Canadian, and Oklahoma Counties, Oklahoma. Okla. Geol. Surv. Open-File Rep. 2-98, 1 sheet, scale 1:24,000.
Suneson, N., Stanley, T. M., and Price, J. D. (1999). Geologic Map of the Bethany and Britton Quadrangles, Canadian and Oklahoma Counties, Oklahoma. Okla. Geol. Surv. Open-File Rep. 2-99, 1 sheet, scale 1:24,000.
Szostakiwskyj, M., Pardo, J. D., and Anderson, J. S. (2015). Micro-CT Study of Rhynchonkos stovalli (Lepospondyli, Recumbirostra), with Description of Two New Genera. PLoS One 10 (6), e0127307. doi:10.1371/journal.pone.0127307
Vaughn, P. P. (1958). A Specimen of the Captorhinid Reptile Captorhinikos chozaensis Olson, 1954, from the Hennessey Formation, Lower Permian of Oklahoma. J. Geology. 66, 327–332. doi:10.1086/626510
Williston, S. W. (1910). New Permian Reptiles: Rhachitomous Vertebrae. J. Geology. 18, 585–600. doi:10.1086/621786
Williston, S. W. (1911). American Permian Vertebrates. Chicago: University of Chicago Press, 145. doi:10.1086/621786
Williston, S. W. (1912). Primitive Reptiles. A Review. J. Morphol. 23, 637–666. doi:10.1002/jmor.1050230404
Keywords: Amniota, Caseidae, Paleozoic, Permian, large herbivore, terrestrial, Synapsida
Citation: Reisz RR, Scott D and Modesto SP (2022) Cranial Anatomy of the Caseid Synapsid Cotylorhynchus romeri, a Large Terrestrial Herbivore From the Lower Permian of Oklahoma, U.S.A. Front. Earth Sci. 10:847560. doi: 10.3389/feart.2022.847560
Received: 02 January 2022; Accepted: 11 February 2022;
Published: 07 March 2022.
Edited by:
Martin Daniel Ezcurra, Museo Argentino de Ciencias Naturales Bernardino Rivadavia, ArgentinaReviewed by:
Agustin G. Martinelli, Museo Argentino de Ciencias Naturales Bernardino Rivadavia, ArgentinaCopyright © 2022 Reisz, Scott and Modesto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Robert R. Reisz, cm9iZXJ0LnJlaXN6QHV0b3JvbnRvLmNhJiN4MDIwMGE7
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.