 Juliana M. Leme
Juliana M. Leme Heyo Van Iten2,3
Heyo Van Iten2,3 Marcello G. Simões
Marcello G. Simões- 1Department of Sedimentary and Environmental Geology, Geosciences Institute, Universidade de São Paulo, São Paulo, Brazil
- 2Department of Geology, Hanover College, Hanover, IN, United States
- 3Department of Invertebrate Paleontology, Research Associate, Cincinnati Museum Center, Cincinnati, OH, United States
- 4Sector of Zoology, Universidade Estadual Paulista, IB, UNESP, Botucatu, Brazil
Paraconularia ediacara n. sp., the oldest documented conulariid cnidarian, is described based on a compressed thin specimen from the terminal Ediacaran Tamengo Formation near Corumbá, Mato Grosso do Sul State, Brazil. The conulariid was collected from a laminated silty shale bed also containing Corumbella werneri and vendotaenid algae. The specimen consists of four partial faces, two of which are mostly covered, and one exposed corner sulcus. The two exposed faces exhibit 32 bell-curve-shaped, nodose transverse ribs, with some nodes preserving a short, adaperturally directed interspace ridge (spine). The transverse ribs bend adapertureward on the shoulders of the corner sulcus, within which the ribs terminate, with the end portions of the ribs from one face alternating with and slightly overlapping those from the adjoining face. This is the first Ediacaran body fossil showing compelling evidence of homology with a particular conulariid genus. However, unlike the periderm of Phanerozoic conulariids, the periderm of P. ediacara lacks calcium phosphate, a difference which may be original or an artifact of diagenesis or weathering. The discovery of P. ediacara in the Tamengo Formation corroborates the hypothesis, based in part on molecular clock studies, that cnidarians originated during mid-late Proterozoic times, and serves as a new internal calibration point, dating the split between scyphozoan and cubozoan cnidarians at no later than 542 Ma. Furthermore, P. ediacara reinforces the argument that the final phase of Ediacaran biotic evolution featured the advent of large-bodied eumetazoans, including, possibly, predators.
Introduction
One of the fundamental problems in the study of the history of life is the timing of the origins of the major groups of metazoans. Molecular clock studies (Runnegar, 1982; Hedges et al., 2004; Peterson et al., 1979; Erwin et al., 2011; dos Reis et al., 2015; Dohrmann and Wörheide, 2017) have placed the origins of the metazoan phyla within the Tonian (max.) to Ediacaran (min.) interval, dating key branching events, including the protostome-deuterostome split and the split between cnidarians and other eumetazoans, at various points within this broad time span. Standing in contrast to the results of molecular clock studies is present understanding of the Neoproterozoic fossil record. Specifically, Ediacaran body fossils currently interpreted as skeletonized or soft-bodied eumetazoans, for example Cloudina, Corumbella, and Kimberella (Fedonkin and Waggoner, 1997; Fedonkin et al., 2007; Bobrovskiy et al., 2018; Dunn et al., 2021), are less than 600 million years old, and thus at present there is a substantial gap between the ages of the apparent first appearances of eumetazoans in the body fossil record and the oldest molecular clock estimates of their times of origin.
Described in this article is the first documented Neoproterozoic conulariid, Paraconularia ediacara n. sp. from the terminal Ediacaran Tamengo Formation (upper Corumbá Group) of west-central Brazil. This conulariid, currently represented by a flattened partial periderm preserving such anatomical features as transverse ribs, nodes, and microlamellae, was originally identified (Van Iten et al., 2014a; Van Iten et al., 2016) as Paraconularia sp. Previously, the oldest known occurrences of Paraconularia were in strata of Middle Devonian age (Hergarten, 1985; Babcock and Feldmann, 1986), and therefore the presence of this genus in the topmost part of the Ediacaran System is truly noteworthy.
Conulariids in general are an extinct order (Conulariida) of marine cnidarians, last occurring in the topmost Triassic (Norian) Stage (Lucas, 2012; Barth et al., 2013), that may have been most closely related to scyphozoan cnidarians of the extant order Coronata (Van Iten et al., 2006a). Conulariids and coronates are united by the possession of a prominent, sessile polyp stage that produced/produces a multi-lamellar, steeply pyramidal or conical periderm bearing (in some species) internal projections along the perradii and interradii (Werner, 1966; Van Iten et al., 1996). Conulariids also exhibit similarities to staurozoans, including tetramerous radial symmetry and prominent gastric septa (Van Iten, 1992; Jerre, 1994; Marques and Collins, 2004; Van Iten et al., 2006a), and thus the hypothesis of a sister group relationship between conulariids and staurozoans (Marques and Collins, 2004) may merit further investigation. The oldest previously known conulariids are Baccaconularia meyeri and B. robinsoni from the Furongian (late Cambrian) Saint Lawrence Formation of southwestern Wisconsin and southeastern Minnesota, United States (Hughes et al., 2000; Van Iten et al., 2006b). Two other Ediacaran fossils, Vendoconularia triradiata (late Ediacaran, White Sea Coast, Russia) (Ivantsov and Fedonkin, 2002; Van Iten et al., 2005; Ivantsov et al., 2019), preserved as molds and casts, and conulariid-like carbonaceous compression fossils from the early Ediacaran Lantian Formation of South China (Yuan et al., 2011; Van Iten et al., 2013), have been interpreted as conulariids or have been compared with this group (Ivantsov and Fedonkin, 2000; Van Iten et al., 2005; Ivantsov et al., 2019). However, hypotheses of homology between these Ediacaran fossils and conulariids have been challenged (Grazhdankin, 2014; Wan et al., 2016; Dzik et al., 2017). In contrast, conulariids exhibit detailed similarities in gross morphology to carinachitiids and hexangulaconulariids, two families of small shelly fossils (SSFs) from the basal (Fortunian) stage of the Cambrian System (Morris and Menge, 1992), and at present there seems to be no better candidate for the nearest relatives of these SSF taxa than conulariids (Guo et al., 2020a; Guo et al., 2020b; Guo et al., 2021). Importantly, the possible presence of Cambrian conulariids or closely related forms immediately above the Ediacaran-Cambrian boundary itself suggests that conulariids may have originated during Neoproterozoic times.
The terminal Ediacaran genus Corumbella, currently known from localities in North and South America and Iran (Pacheco et al., 2015; Walde et al., 2015; Vaziri et al., 2018; Walde et al., 2019; Amorim et al., 2020), has also been interpreted as a polypoid scyphozoan closely related to conulariids (Van Iten et al., 2014a; Van Iten et al., 2016; Pacheco et al., 2015). It should be noted, however, that Walde et al. (2019, p. 335) hypothesized that Corumbella was a worm-like bilaterian.
The discovery of Paraconularia in strata of latest Ediacaran age not only demonstrates that conulariids crossed the crucial Ediacaran-Cambrian boundary, but it also corroborates the hypothesis that phylum Cnidaria originated during the Neoproterozoic. Additionally, the existence of this ancient scyphozoan, extant species of which engage in predation (Pearse et al., 1987), may provide additional support for the hypothesis (e.g., Hua et al., 2003; Schiffbauer et al., 2016) that the origin of predation and complex food webs predated the beginning of the Phanerozoic Eon and the Cambrian Explosion.
Geological Setting
The late Ediacaran to earliest Cambrian (ca. 565-539 Ma; see Linnemann et al., 2019) Corumbá Group, named after the city of Corumbá in Mato Grosso do Sul State (west-central Brazil), crops out at the junction of the Amazon Craton, the northern Rio Apa Block, and the folded southern Paraguay Belt (Amorim et al., 2020) (Figure 1). It was initially deposited in an elongate rift basin that evolved into a passive continental margin hosting shallow to deep marine environments. The basin was deformed during the Brazilian Orogeny, which resulted in the formation of the southern part of the Paraguay Belt in southwestern Brazil (Almeida, 1968; Gaucher et al., 2003; Alvarenga et al., 2009; Boggiani et al., 2010; Warren et al., 2014; Amorim et al., 2020). The Corumbá Group exhibits a maximum thickness of about 400 m and is subdivided into five formations (Figure 2). The lowermost, or Cadiueus Formation, consists of conglomerate, sandstone, and shale, while the overlying Cerradinho Formation is composed of sandstone, shale, and carbonates (limestone and dolostone). Above this unit, the Bocaina Formation, composed of dolomite and subordinate shale, directly underlies the Tamengo Formation, which ranges from 80 to 100 m thick and consists predominantly of dark gray carbonaceous limestone and subordinate silty shale (Amorim et al., 2020). Both lithologies yield macroscopic body fossils, the most conspicuous of which are the skeletonized eumetazoans Cloudina lucianoi (in limestone) and Corumbella werneri (in silty shale). The Corumbá Group terminates with the Guaicurus Formation, a thick package of uniform shale which has yielded trace fossils of meiofaunal bilaterians (Parry et al., 2017).
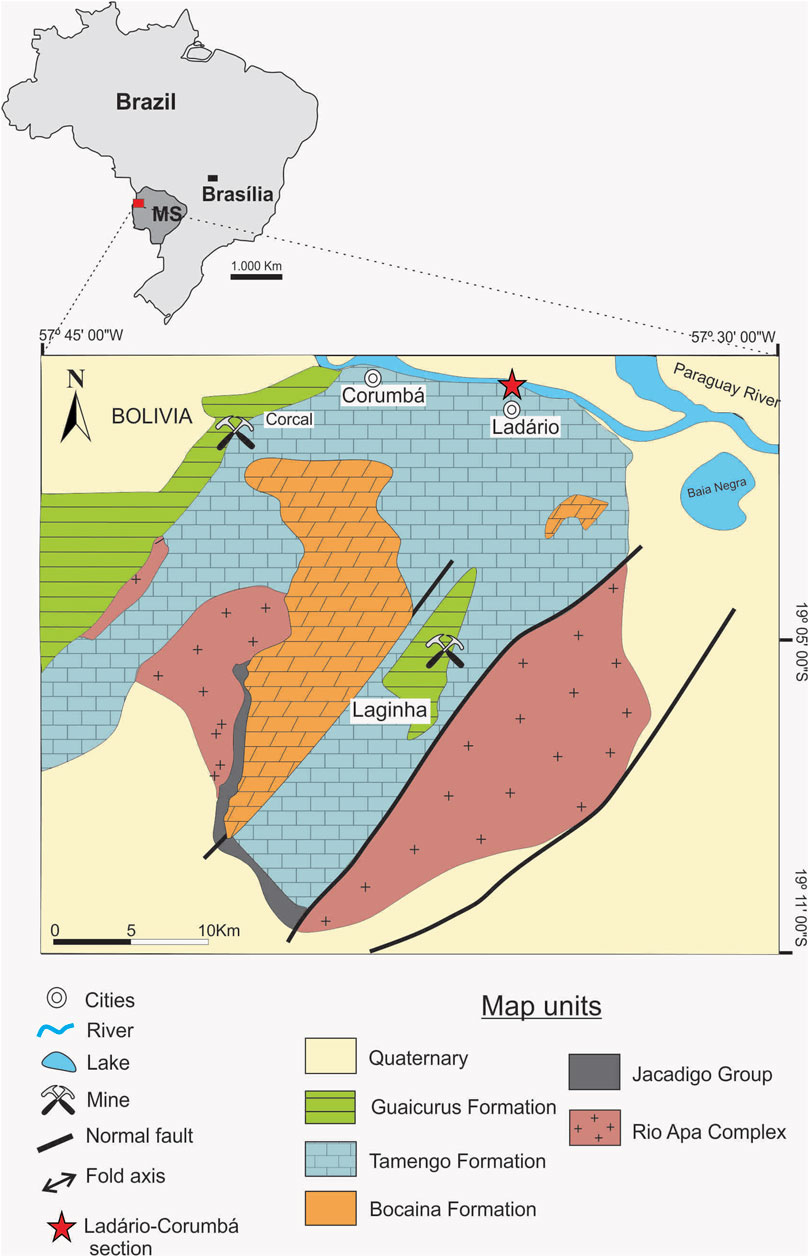
FIGURE 1. Geological map of the Paraguay Belt in South America, showing the location of the Corumbá (Brazil) area, the Ladário-Corumbá section (red star), and the geology of the Corumbá Group.
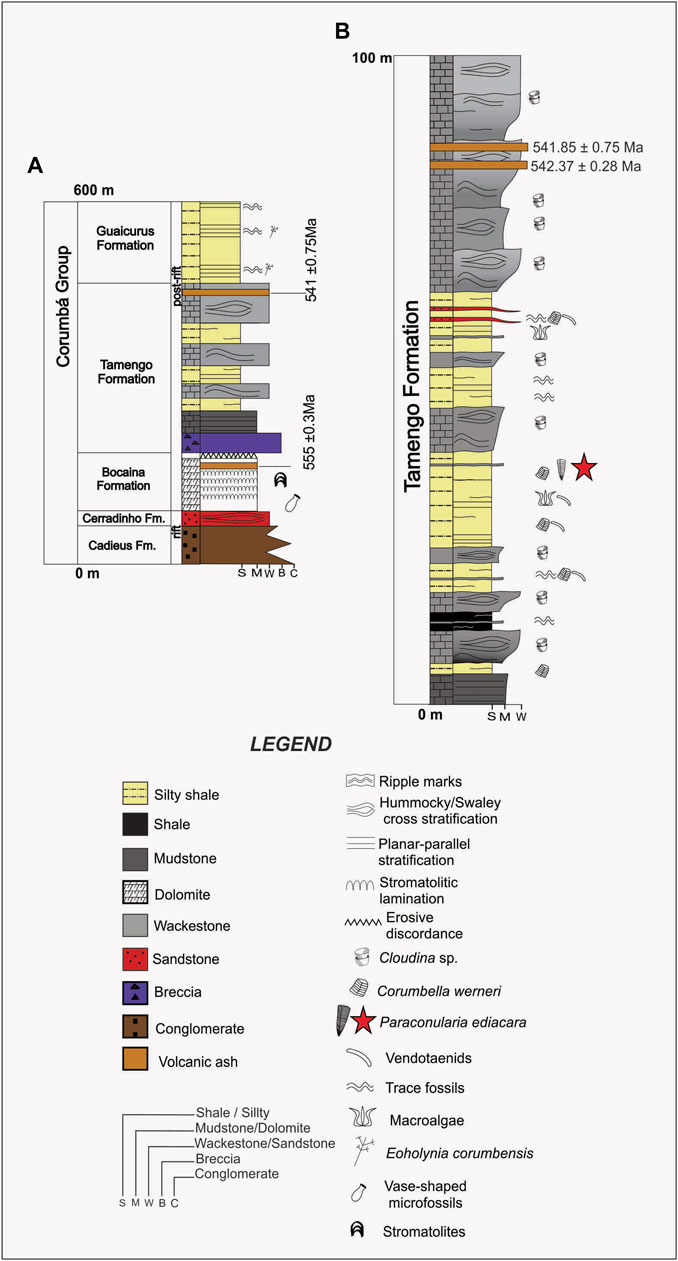
FIGURE 2. Measured stratigraphic columns for (A) the Corumbá Group and (B) the Tamengo Formation, showing in (B) the levels of occurrence of Cloudina, Corumbella, vendotaenids, macroalgae, and Paraconularia ediacara n. sp. (red star). Modified from Amorim et al. (2020).
High-precision dating of two volcanic tuffs situated a few meters below the top of the Tamengo Formation yielded mean U-Pb ages of 541.85 ± 0.75 Ma and 542.27 ± 0.38 Ma, respectively (Parry et al., 2017). Combined with an age of 555.18 ± 0.30 Ma for a tuff bed near the top of the underlying Bocaina Formation (Parry et al., 2017), these two dates indicate that the entire Tamengo Formation is latest Ediacaran in age. This conclusion is corroborated by the presence throughout the Tamengo Formation of Cloudina (Figure 2), a likely index fossil for the latest Ediacaran (Xiao et al., 2016). Importantly, Paraconularia ediacara occurred at a level situated well below the top of the Tamengo Formation, and therefore its age may be several million years older than 542 Ma, the approximate age of the second oldest tuff layer mentioned above.
Materials and Methods
The Tamengo Formation conulariid, formally diagnosed and described below, was collected from an outcrop designated as ELC (Ladário/Corumbá Escarpment) IV (Amorim et al., 2020) and located on the south bank of the Paraguay River near the village of Ladário (Figures 1, 2). The conulariid occurred within a 1.5-m-thick, non-bioturbated, finely laminated silty shale bed, at a level approximately 45 m below the top of the Tamengo Formation (Figure 2). The same bed, including the bedding plane on which the conulariid occurred, also yields Corumbella, vendotaenids, and macroalgae. The conulariid was revealed by splitting a hand-sized slab into two pieces, one bearing the part and the other the counterpart. Both are housed in the paleontological collections of the Department of Sedimentary and Environmental Geology of the Geosciences Institute of the University of São Paulo, under collection number 1T/2301a, b.
Assignment of the conulariid to a new species of Paraconularia is based on two lines of evidence: (1) recognition of a set of gross morphological features uniquely exhibited by conulariids in general (e.g., Hughes et al., 2000; Van Iten et al., 2008); and (2) comparisons with illustrations and reposited specimens of previously described Paraconularia from Devonian through Triassic rocks in North and South America (Driscoll, 1963; Babcock and Feldmann, 1986; Babcock et al., 1987; Babcock, 1993; Leme et al., 2004; Escalante-Ruiz et al., 2014), Africa (Van Iten et al., 2008), Europe (de Koninck, 1883; Slater, 1907; Hergarten, 1985; Barth et al., 2013; Lukševičs, 2020), Asia (Xu and Li, 1979; Zhu, 1985; Swami et al., 2017; Min et al., 2021), and Australia/New Zealand/Tasmania (Thomas, 1969; Waterhouse, 1979; Parfrey, 1982; Waterhouse, 1986). Imaging and Raman compositional analysis of the new conulariid were carried out in the Department of Sedimentary and Environmental Geology of the Geosciences Institute of the University of São Paulo, São Paulo State, Brazil. Low magnification optical examination and light photography were conducted using an OLYMPUS DSX stereomicroscope under low angle illumination. A small fragment of the periderm was coated with gold and examined using a LEO 440 scanning electron microscope (SEM).
SYSTEMATIC PALEONTOLOGY

Diagnosis: Large Paraconularia with circular nodes that are moderately coarse, widely spaced (10-11 per 5 mm), and extended adapertureward as a short, adaperturally tapered interspace ridge. Faces approximately equal in width. Transverse ribs in the apertural/upper middle region of the periderm low bell-curve-shaped and uninterrupted at the facial midline, numbering 10-11 per 10 mm. Apical angle approximately 10°. Corner sulcus subangular; corners and facial midline without internal thickening or carina.
Description: Part and counterpart of a thin (∼ 0.2–0.5 mm), strongly compressed (transversely) fragment of a steeply pyramidal, four-sided periderm measuring approximately 34 mm long and 24 mm wide and lying parallel to bedding. Exposed portion of the periderm consists primarily of two mutually adjacent, very gently tapered partial faces and the corner sulcus between them (Plate I, Figures 1, 2). Faces originally about equal in width. Nearly smooth inner surface of small portions of the two mostly covered faces visible at the broken apical end of the part (Plate I, Figure 1). Apical region entirely missing. Apertural margin may be partially preserved. Apical angles ∼10°. Length of the complete periderm exceeded 100 mm (as indicated by adapical extension of the truncated facial midlines). External surface of the two exposed faces exhibits 32 trochoidal, thickened, node-bearing transverse ribs separated by broad interspaces and numbering 10-11 per 10 mm. Transverse ribs adaperturally arcuate, approximately bell-curve-shaped (inflected circular curve geometry; Babcock and Feldmann, 1986), crossing both faces without interruption or diminution at the facial midline (Plate I, Figures 1, 2, 5, 7). Transverse ribs bent adapertureward on the shoulders of the subangular corner sulcus, within which they terminate, with the end portions of the transverse ribs from one face alternating with and slightly overlapping those from the adjoining face (Plate I, Figures 1, 3–6). Nodes moderately coarse, separated from each other by a gap that measures approximately 0.5–1.5 (rarely 2.0) node diameters in length, numbering from 10 to 11 per 5 mm (Plate I, Figures 5, 8). Many nodes extended adapertureward as a short, spine-like interspace ridge (adapertural spine; Babcock and Feldmann, 1986) (Plate I, Figure 5). Nodes of every other transverse rib form rectilinear files that are nearly parallel to the facial midline or the nearest corner. Corners and facial midlines without internal thickening or carina. Schott (apical wall) absent. Periderm very finely lamellar (Plate I, Figures 9, 10), apparently organic, though possibly with originally phosphatic microlamellae lost secondarily.
Derivation of name: ediacara, from Ediacaran, the age of the conulariid occurrence.
Type material: The holotype, reposited in the palaeontological collections of the Department of Sedimentary and Environmental Geology, University of São Paulo, São Paulo State, Brazil (1T/2301 a, b).
Occurrence: Thick silty shale bed in the middle part of the Tamengo Formation (upper Corumbá Group) at Locality ELC (Ladário/Corumbá Escarpment) IV (Amorim et al., 2020) on the south bank of the Paraguay River near the village of Ladário in Mato Grosso do Sul State, southwestern Brazil.
Age and horizon: Latest Ediacaran (no younger than 542 Ma), approximately 45 m below the top of the Tamengo Formation (upper Corumbá Group) (Parry et al., 2017).
Remarks and comparisons: Together with previously published anatomical data and illustrations (e.g., Babcock and Feldmann, 1986), the photographic illustrations here presented (Plate I, Figures 1, 2, 4, 5, 8–10; Figure 3) show that P. ediacara exhibits a complex suite of gross anatomical features that is shared only with Devonian-Triassic conulariids that have been placed in the genus Paraconularia. To be sure, there has been some divergence of opinion regarding the diagnostic characters of Paraconularia (e.g., Babcock and Feldmann, 1986; Van Iten et al., 2008; Min et al., 2021), and there is also some uncertainty surrounding the diagnostic characters and taxonomic composition of Conulariida itself (e.g., Hughes et al., 2000). Be that as it may, in recent years it has generally been assumed (e.g., Van Iten et al., 2006b; 2014b) that Paraconularia and similar genera such as Conularia are members of a single, monophyletic group that excludes medusozoan taxa lacking a finely lamellar, steeply pyramidal, organo-phosphatic or (possibly) organic periderm with (usually) sulcate corners bounding four faces bearing regularly arrayed small nodes and/or transverse ribs. Among previously known genera from the terminal Ediacaran and basal Cambrian, only Corumbella (terminal Ediacaran) and Carinachites (basal Cambrian, ca. 535 Ma, Han et al., 2017) are similar to P. ediacara and many other conulariids in having an originally quadrate skeleton showing regular corrugation of the gently tapered sides or faces. Absent in these two genera, however, are the fine details and pattern of arrangement of the transverse ribs of Paraconularia. As discussed in part by Van Iten et al. (2016), the faces of Paraconularia, including the type species, are crossed by adaperturally arcuate or angulated, generally node-bearing transverse ribs which terminate within the corner sulcus. There, the end portions of the transverse ribs trend obliquely adapertureward, with the end portions of the transverse ribs from one face alternating with and partially overlapping those from the adjoining face. Adapertural bending of the transverse ribs on the shoulders of the corner sulcus can be subtle, and it has been reported (Min et al., 2021) that in some species such bending is absent. Along the facial midline, the transverse ribs may be interrupted, with the ends of the ribs on one half face arranged in alternation with those on the other half face, or they may be continuous, as in both P. ediacara and the type species (see de Koninck, 1883, pl. LIV, Figure 9). As in most conulariids having trochoidal (longitudinally) transverse ribs, the periderm thickens from the center line of the interspaces to the axial plane of the fold-like transverse ribs, in such a way as to reduce the relief of the transverse ribs on the inner surface of the periderm (see for example Ford et al., 2016, Figure 3E). In many of the species bearing nodes, including P. ediacara, the adapertural half of each node is developed into a short, spine-like ridge (adapertural spine; Babcock and Feldmann, 1986) that extends part way across the interspaces. Finally, the pattern of arrangement of the nodes on the faces is such that the nodes of every other transverse rib are collinear, forming longitudinal series that are approximately parallel to the facial midline or nearest corner.

FIGURE 3. Paraconularia sp. (Mississippian, Kentucky, United States; GP-1E 11672). (A) compressed partial specimen displaying two faces and the corner sulcus between them (photograph oriented with the apertural end of the fossil at the top). Yellow open rectangles highlight stretches of the corner sulcus (C) in which adapertural bending and alternation of the ends of the transverse ribs, which alternate as well along the facial midline (ML), are best displayed. (B) detail of the right face just below the apertural end of the fossil. Yellow arrows immediately to the right of the facial midline highlight some of the minute nodes. Scale bar: 10 mm.
In addition to being far older geologically than other described species in Paraconularia, P. ediacara is also distinguishable from them morphologically, being characterized by the following unique set of gross anatomical features: (1) transverse ribs in the apertural/upper middle region low bell-curve-shaped (angulated circular curve geometry; Babcock and Feldmann, 1986), continuous across the facial midline, numbering from 10 to 11 per 10 mm; and (2) rib nodes moderately large, clearly separated from each other and developed adaperturally into a short, spine-like interspace ridge. It is perhaps the second item that is most important, though, as in nearly all other known species the transverse ribs are either smooth (nodes are absent) or the nodes are very small and mutually contiguous or nearly so (Figure 3; see also illustrations in Babcock and Feldmann, 1986). Further grounds for assigning the Tamengo Formation Paraconularia to a new species are provided by the fact that the difference in absolute age between it and Devonian Paraconularia is roughly 130 million years, which is about an order of magnitude greater than the estimated life spans of the longest-lived invertebrate/cnidarian species (Valentine, 1970; Raup, 1978, 1991; Sepkoski, 1992; Lawton and May, 1995).
Preservation and taphonomy: The holotype and only known specimen of P. ediacara n. sp. is a fragment of an elongate pyramidal periderm, the faces of which lie parallel to bedding and probably measured at least 100 mm long when complete. It consists of a combination of skeletal material, possibly altered, and external molds of the same. The periderm does not react with dilute HCl and therefore is not calcareous, as expected given that the periderm of Phanerozoic conulariids is organo-phosphatic (Ford et al., 2016). It should be noted here that Van Iten et al. (2016) incorrectly reported the length of the fragment as 26 mm, which is close to its maximum width. The incompleteness of the fossil, which lacks the apical third and probably part of the middle region, is not an artifact of collecting, as the margins of the specimen are fully bordered by silty shale, and the specimen itself was revealed by splitting the host rock slab. The present distorted state of the specimen, with the two exposed faces and corner sulcus nearly touching the relatively smooth, mostly hidden facial pair/corner beneath them (Plate I, Figure 1), is typical of thin-walled conulariids preserved parallel to bedding in shale (see for example Babcock and Feldmann, 1986, figures. 18.5 and 26.4). The smoothness of the hidden faces, the broken edges of which are nevertheless sharp, also is typical of conulariids and reflects inward thickening and consequent diminution of the relief of the transverse ribs on the inner surface of the periderm. Owing possibly to having been restricted to the exteriormost levels in the periderm, which can undergo exfoliation during splitting of the host rock, the short interspace ridges are not evident at all places on the fossil and are easiest to discern on external molds. Finally, the presence of imbricated fragments of Corumbella in the same silty shale bed that yielded P. ediacara (Rodrigues et al., 2003; Amorim et al., 2020) suggests that the specimen was transported and fragmented prior to final burial.
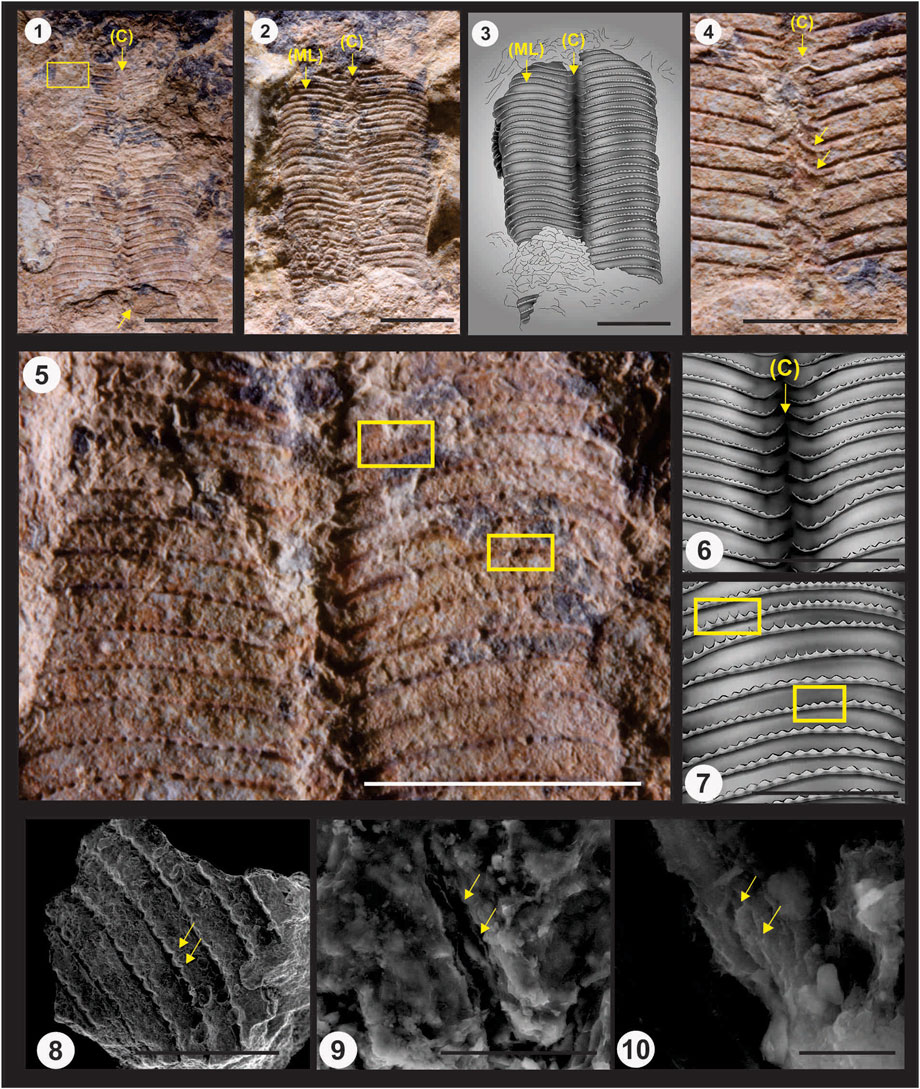
FIGURE 4. Paraconularia ediacara n. sp. (terminal Ediacaran, middle Tamengo Formation, upper Corumbá Group, Mato Grosso do Sul, Brazil; specimen GP-IT 2301, Geosciences Institute, University of São Paulo). 1, 2, color light photographs (both oriented with the apertural end of the fossil at the top); 1, the part, showing the two exposed partial faces and corner sulcus (indicated by the arrow labelled C) between them. The truncated apical ends of the two mostly covered faces project slightly from underneath the truncated apical ends of the two exposed faces (bottom arrow). Open yellow rectangle outlines the area from which the small fragment of periderm for SEM imaging (Figures 8–10 below) was extracted; 2, the counterpart (again with the corner indicated by an arrow labelled C, and with one of the facial midlines indicated by an arrow labelled ML); 3, schematic drawing of the part, highlighting the alternation of the nodose transverse ribs in the corner sulcus (C) and the continuation of the transverse ribs across the facial midline (ML). 4, 5, color light photographs (both oriented with the apertural end of the fossil at the top); 4, detail of the exposed corner sulcus of the counterpart. Note the pronounced adapertural deflection and alternation of the end portions of the transverse ribs within the corner sulcus (yellow arrows); 5, detail of the lower (apical) portion of the two exposed faces of the part, showing the widely spaced nodes. Upper rectangle highlights several nodes preserving the short interspace ridges in positive relief, while the lower rectangle highlights several nodes showing much shorter interspace ridges; 6, schematic drawing of a portion of the corner sulcus (C) shown in 5; 7, schematic drawing of the facial area with rectangles shown in 5; 8-10, SEM photomicrographs (secondary electron mode) of a small fragment of the periderm; 8, exterior surface of the periderm, showing several transverse ribs, widely spaced nodes, and very short, spine-like interspace ridges (pointing toward the apertural end of the periderm, yellow arrows); 9, detail of the fragment shown in 8, with canyon-like fractures exposing the edges of several microlamellae (yellow arrows); 10, detail of one of the fractures shown in 9 and exposing microlamellae (yellow arrows). Scale bar: 5 to 8 mm (Figures 1–3, 5); 7 mm (Figure 4); 5 mm (Figures 6, 7); 3 mm (Figure 8); 40 μm (Figure 9); 5 μm (Figure 10).
The high magnification SEM images (Plate I, Figures 9, 10) reveal that the periderm of P. ediacara is composed of extremely thin (< 5 μm), mutually parallel microlamellae. This is the basic microstructure of Phanerozoic conulariids preserving the periderm, which is a bi-composite material consisting of apatitic and organic microlamellae arranged in alternation (Ford et al., 2016). Color images (Plate I, Figures 1, 2, 4, 5) of P. ediacara show that it is variegated, with irregularly bounded areas of pale tan-brown periderm occurring alongside patches of darker-colored periderm. Similar color variation is exhibited by the host silty shale and may reflect post-burial diagenesis and/or chemical weathering. In other words, the original periderm probably was uniform in color, as is the periderm of Phanerozoic conulariids, which however may bear pigmented lines or bands corresponding to internal carinae at the corners and/or facial midlines (Van Iten, 1992). Lastly, and as reported previously by Van Iten et al. (2016), Raman analysis of portions of the periderm failed to yield any lines diagnostic of phosphorous, one of the elemental constituents of apatite; instead, it appears that the periderm is now, and may originally have been, entirely or predominantly organic in composition. This hypothesis can potentially be tested by imaging broken edges of the periderm at very high magnifications. If in fact originally apatitic microlamellae have been lost secondarily, then there might still be an extremely narrow gap between the preserved, organic microlamellae. Otherwise, all microlamellae should be in direct contact with each other. Our photomicrographs (Plate I, Figures 9, 10) show no clear evidence of microscopic gaps, and thus, at this point, we are inclined to think that originally apatitic microlamellae were not present.
Discussion
Significance
The discovery of Paraconularia in strata of terminal Ediacaran age is significant for several reasons. First, the new conulariid expands the list of skeletonized or tubular Ediacaran genera, including Cloudina, Namacalathus, Namapoikia, and Sinotubulites, which are generally regarded as eumetazoans (e.g., Fedonkin, 1992; Fedonkin and Waggoner, 1997; Grazhdankin, 2014; Grazhdankin, 2016; Becker-Kerber et al., 2017).
Second, and as noted above, conulariids were scyphozoan cnidarians or close medusozoan relatives of this group. Moreover, even though other Ediacaran taxa, most notably Corumbella and Vendoconularia, have been interpreted as scyphopolyps, the hypothesis of a scyphozoan affinity for P. ediacara enjoys a substantially stronger basis in comparative anatomy. It should be noted here that Bjarmia cycloplerusa, described by Grazhdankin, (2016) from the late Ediacaran Erga Formation (southeastern White Sea area, Russia), was classified by this author as a coronate scyphomedusa (jellyfish). However, this occurrence was not included by Young and Hagadorn, (2020, p. 185) among their “thirteen confirmed medusa-bearing deposits”, the oldest of which (the Chengjiang Lagerstätte) is early Cambrian (Stage 3) in age. Thus, either singly or together with certain other body fossils, P. ediacara now constitutes the strongest paleontological evidence of the presence of scyphozoans or any cnidarians during Neoproterozoic times.
Third, Paraconularia is now one of the longest lived eumetazoan genera, ranging downward from the Upper Triassic into the topmost Ediacaran, or through about 340 million years of geological time. Conulariids and Paraconularia are established (respectively) as a eumetazoan order and genus that survived the end-Ediacaran extinction event, a status currently shared with the agglutinated foraminiferan Platysolenites (Kontorovich et al., 2009), certain microfossils (Anderson et al., 2017; Grazhdankin et al., 2020), and three skeletonized metazoan genera, namely Anabarites, Cambrotubulus, and Cloudina (Zhu et al., 2017). Furthermore, cladistic analyses of phylogenetic relationships among genera within Conulariida (De Moraes Leme et al., 2008; Van Iten et al., 2014b) suggest that Paraconularia is a relatively apical branch, thus pushing the origin of conulariids even farther into the deep past.
Fourth, conulariids are now known from a level below that of the first occurrence of most SFFs (Zhu et al., 2017), in other words before the putative onset of the Cambrian Explosion of eumetazoan (mainly bilaterian) diversity. If in fact there was such an event (Blair and Hedges, 2005; Wood et al., 2019), then conulariids were around well before it started, and they were an order of magnitude larger than early Cambrian small shelly fossils. Put another way, the discovery of conulariids in the Tamengo Formation adds to a growing body of evidence (e.g., Darroch et al., 2015; Darroch et al., 2016; Zhu et al., 2017; Darroch et al., 2018b) of some degree of phylogenetic and ecological continuity across the Ediacaran-Cambrian boundary.
Fifth, the apparent organic composition of the Tamengo Formation periderm raises the intriguing possibility that the earliest conulariids were non-mineralizing, with production of phosphatic micro-lamellae within the clade having originated after the end of the Ediacaran but before the end of the Cambrian, as indicated by the presence of mineralized Baccaconularia in the Furongian of the north-central United States (Hughes et al., 2000; Van Iten et al., 2005). Conulariids in certain Phanerozoic strata, for example Anaconularia anomala from the Upper Ordovician Letná and Zahořany formations of Bohemia (Bruthansová and Van Iten, 2020), are known only from molds and casts, and thus post-mortem loss of the entire periderm has occurred in some cases, though the causes of this phenomenon in conulariids have yet to be determined. In the present case, the hypothesis of complete or partial loss of an apatitic skeletal component can potentially be tested through the discovery and microstructural analysis of additional conulariid specimens from the Tamengo and other Ediacaran formations.
Sixth, by analogy with extant scyphozoans, P. ediacara was a predator. Using elongate tentacles armed with nematocysts, modern scyphozoans prey primarily on meso-zooplankton (Grazhdankin, 2016). To date, no such soft-part structures have been detected in conulariids. Nevertheless, if P. ediacara was a predator, it seems likely that its prey were microscopic, possibly single-celled organisms living in the water column or even on the seafloor (see also discussions in the sections below). Again, by analogy with modern scyphozoans, P. ediacara may also have had the ability to assimilate dissolved organic matter (Arai, 1997).
Finally, the presence of conulariids in latest Ediacaran strata implies that Cnidaria has an even deeper Proterozoic evolutionary history. According to a previous cladistic analysis of the phylogenetic relationships among major groups within the phylum (Van Iten et al., 2006a; 2014a), conulariids (Conulariida) originated after Scyphozoa and Cubozoa split from each other, and after the most recent common ancestor of these two classes split from its most recent common ancestor with Hydrozoa. Still earlier, medusozoans (Scyphozoa, Cubozoa, Hydrozoa, and Staurozoa) split from their most recent common ancestor with Anthozoa. It should be noted, however, that certain more recent studies of cnidarian phylogeny (e.g., Zhao et al., 2019) have concluded that Scyphozoa is paraphyletic. Be that is it may, the origin of Conulariida, now placed on the basis of body fossil evidence within the Neoproterozoic, was preceded by multiple branching events in the evolutionary history of Cnidaria. Moreover, and in accordance with the phylogenetic trees of Van Iten et al. (2006a) and Van Iten et al. (2014a), Cubozoa and Scyphozoa diverged from their most recent common ancestor no later than 542 Ma, the minimum absolute age of P. ediacara, which may therefore serve as a new internal calibration point for molecular clock studies of the evolution of Cnidaria.
Ediacaran Marine Paleoecology
Together with the presence of the putative cnidarian polyps Corumbella and Cloudina in late Ediacaran rocks in Africa and North and South America (e.g., Kouchinsky et al., 2012; Pacheco et al., 2015; see however Yang et al., 2020 for a discussion of the possible annelid affinities of Cloudina), the presence of P. ediacara in the Tamengo Formation of Brazil may lend additional weight to the hypothesis (e.g., Hua et al., 2003; Schiffbauer et al., 2016) that the feeding strategy of predation, exhibited by extant medusozoans in general (Arai, 1997), originated before the close of the Neoproterozoic. Additionally, the existence of other late Ediacaran organisms capable of skeletogenesis or tube construction, including Namacalathus, Namapoikia, and Sinotubulites (e.g., Fedonkin, 1992; Fedonkin and Waggoner, 1997; Grotzinger et al., 2000; Hofmann and Mountjoy, 2001; Wood et al., 2002; Hua et al., 2003; Grazhdankin, 2014; Grazhdankin, 2016), further signals an increase in ecological complexity near the end of the Proterozoic. Indeed, some authors (e.g., Narbonne, 2005) have argued that terminal Ediacaran marine communities were comparable in productivity and trophic structure to modern marine ecosystems, and it is becoming increasingly likely that terminal Ediacaran ecosystems featured complex food chains composed of herbivores, filter feeders, and predators (Vermeij, 1989; Lipps and Culver, 2002; Babcock et al., 2005; Xiao and Laflamme, 2009; Laflamme et al., 2013; Droser and Gehling, 2015; Schiffbauer, et al., 2016; Gibson et al., 2019; Muscente et al., 2019; Wood, et al., 2019; Cracknell et al., 2021). Animals in these ecosystems developed a variety of foraging strategies based on microorganisms (picoplankton, microplankton, and microbial mats) as the primary producers (Vidal and Moczydłowska-Vidal, 1997; Gehling, 1999; Seilacher, 1999; Lipps and Culver, 2002; Darroch et al., 2016; Darroch et al., 2018a; Darroch et al., 2018b; Darroch et al., 2020), though with certain modern elements such as bioturbating infauna also present. In short, then, the occurrence of P. ediacara and other, possible predators (Corumbella and Cloudina) in the latest Ediacaran Tamengo Formation lends additional plausibility to the hypothesis of increasing complexity of ecosystems just prior to the beginning of the Phanerozoic.
Taphonomy and Epifaunal Tiering of the Tamengo Formation Biota
Paraconularia ediacara, Corumbella, and Cloudina in the Tamengo Formation were components of a marine macrobenthic ecosystem developed in a mixed carbonate-siliciclastic ramp setting (Amorim et al., 2020). Together with vendotaenids (Vendotaenia sp.), macroalgae, and the ichnofossil Multina minima (Parry et al., 2017), P. ediacara and Corumbella occur on bedding planes in silty shales deposited in the outer to distal mid-ramp facies (Amorim et al., 2020). Both taxa are represented by comminuted, flattened, loosely packed remains that are randomly arranged in the sedimentary matrix. Therefore, these fossils probably are parautochthonous to allochthonous elements that settled from suspension on a deep (below storm wave base), low-energy fine-grained bottom. Shallower, more proximal parts of the ramp were frequently affected by storms, and the skeletons of P. ediacara and Corumbella may have been transported basinward over hundreds of meters, a taphonomic condition well exemplified by Devonian conulariids preserved in distal shale facies (Rodrigues et al., 2003). The occurrence of Corumbella as imbricated bioclasts at the base of lens-shaped deposits of very fine sand (Amorim et al., 2020) suggests further that these skeletons were sturdy enough to survive tractive transport. Importantly, in situ (i.e., preserved in life position) occurrences of P. ediacara and Corumbella have not been found in the Tamengo Formation. However, dense accumulations of in situ Cloudina shells are known from the shallow water, microbially-induced carbonates making up the bulk of the formation. Although single bedding planes bearing in situ associations of all three benthic invertebrates are unknown, their frequent co-occurrence (particularly Corumbella and Cloudina) in polytypical assemblages suggest that they may have colonized the same bottoms or at least mutually adjacent ones. Indeed, in situ Cloudina–Corumbella–Namacalathus associations are known from inner ramp carbonate deposits of the Ediacaran Itapucumi Group in Paraguay (Warren et al., 2012; Warren et al., 2017).
The three macrobenthic invertebrate taxa present in the Tamengo Formation seem to have been sessile epifaunal members of a low-to high-density, tiered community that flourished in a ramp setting subjected to intermittent burial events. The occurrence of these fossils together with meiofaunal ichnofossils (Parry et al., 2017) suggests a relatively complex tiering structure with distinct epifaunal guilds and even a shallow infaunal one. Based on estimates of the maximum original height of the best-preserved fossil specimens, at least two and possibly three nonoverlapping tiers extended from 0 to 10 cm above the seafloor. The lowest level from 0 to 1 cm was dominated by Cloudina, which grew at a subhorizontal to oblique (occasionally vertical) attitude relative to the seafloor (Becker-Kerber et al., 2017). This animal was largely prone, having only one or two of its nested funnel-like segments oriented slightly oblique to bedding (Becker-Kerber et al., 2017) to benefit from bottom currents. Corumbella werneri had a resistant but flexible carapace (Pacheco et al., 2015) which grew to an estimated 80 mm in length (Babcock et al., 2005). This species lived with the apical end embedded in muddy sediment (Pacheco et al., 2011; Pacheco et al., 2015), a condition also observed in specimens from the Ediacaran Itapucumi Group of Paraguay (Warren et al., 2012; Warren et al., 2017). Thus, C. werneri occupied a substantially higher tiering level than that of Cloudina, while P. ediacara, which appears to have been somewhat longer than C. werneri, may have occupied an even higher tiering level. As was the case for other Ediacaran assemblages (see Clapham and Narbonne, 2002, p. 630), the biomass of the Tamengo Formation community was concentrated in the basal 10 cm above the seafloor. The tiering structure of this community appears to have been controlled both by constructional differences between taxa and by feeding behavior, possibly with Cloudina being a passive filter feeder (Becker-Kerber et al., 2017) and both C. werneri and P. ediacara engaging, possibly, in active predation.
Concluding Statement
The first known Ediacaran conulariid, Paraconularia ediacara n. sp., is diagnosed and described based on an incomplete but otherwise well-preserved specimen from the terminal Ediacaran Tamengo Formation of southern Brazil. The discovery of this body fossil has important implications for studies of the origins of the major groups of animal phyla and the early evolution of marine ecosystems. It is hoped that further collecting at the Ladário localities near Corumbá (Mato Grosso do Sul State) will yield additional material of this conulariid, which provides further support for the hypothesis of a relatively deep Neoproterozoic origin for phylum Cnidaria and therefore, possibly, of predation as well.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
JL and HV performed laboratory work and taxonomical identification. All three co-authors worked together in the field to search for additional specimens of P. ediacara, participated in the formulation of interpretations and hypotheses in the discussion section of the paper, and approved the submitted version.
Funding
JL was supported by the FAPESP (proc. 13/17835-8; 16/06114-6) and HV was supported in part by a research grant from the Hanover College Faculty Development Committee.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
T. R. Fairchild (University of São Paulo, Brazil) is gratefully acknowledged as the collector of the Tamengo Formation Paraconularia specimen. Engineer I. J. Sayeg is thanked for assistance with scanning electron microscopy, and L. E. Anelli is thanked for assistance with light photography. Permission to examine reposited specimens of Phanerozoic species of Paraconularia was granted by M. Coyne (Geological Survey of Canada, Ottawa), A. Howell (Redpath Museum, McGill University, Montreal, Canada), J. Miller-Camp (Indiana University, Bloomington, United States), J. Darrell and C. Sendino (Natural History Museum, London, United Kingdom), Lisa Amati (New York State Museum, Albany, United States), D. Erwin (United States National Museum, Washington, D. C.), and T. Adrain (University of Iowa, Iowa City, United States). The comparison specimen of Paraconularia sp. from the Mississippian of Kentucky (United States) was photographed by T. V. Van Iten. Finally, the thoughtful and constructive reviews of the three referees are greatly appreciated.
References
Almeida, F. F. (1968). Evolução Tectônica Do Centro Oeste Brasileiro No Proterozoico Superior. An. Acad. Bras. ciência Supl. simpósio manto Super. 40, 285–296.
Alvarenga, C. J. S., Boggiani, P. C., Babinski, M., Dardenne, M. A., Figueiredo, M. F., Santos, R. V., et al. (2009). “The Amazonian Paleocontinent,” in Neoproterozoic-Cambrian Tectonics, Global Change and Evolution: A Focus on Southwest Gondwana. Editors C. Gaucher, A. N. Sial, G. P. Halverson, and H. E. Frimmel (Amsterdam: Elsevier), 498.
Amorim, K. B., Afonso, J. W. L., Leme, J. d. M., Diniz, C. Q. C., Rivera, L. C. M., Gómez‐Gutiérrez, J. C., et al. (2020). Sedimentary Facies, Fossil Distribution and Depositional Setting of the Late Ediacaran Tamengo Formation (Brazil). Sedimentology 67 (7), 3422–3450. doi:10.1111/sed.12749
Anderson, R. P., Macdonald, F. A., Jones, D. S., McMahon, S., and Briggs, D. E. G. (2017). Doushantuo-type Microfossils from Latest Ediacaran Phosphorites of Northern Mongolia. Geology 45 (12), 1079–1082. doi:10.1130/G39576.1
Babcock, L. E. (1993). “Exceptionally Preserved Conulariids from the Conularienschichten, Fossil-Lagerstätten in the Devonian of Brazil,” in Fosiles y Facies de Bolivia – Vol. II Invertebrados y Paleobotanica. Editor R. Suarez-Soruco (Revista Tecnica de YPFB), 13-14, 77–91.
Babcock, L. E., and Feldmann, R. M. (1986). Devonian and Mississippian Conulariids of North America. Part B. Paraconularia, Reticulaconularia, New Genus and Organisms Rejected from Conulariida. Ann. Carnegie Mus. 55, 411–479.
Babcock, L. E., Feldmann, R. M., Wilson, M. T., and Suárez-Riglos, M. (1987). Devonian Conulariids of Bolivia. Natl. Geogr. Res. 3, 210–231.
Babcock, L. E., Grunow, A. M., Sadowski, G. R., and Leslie, S. A. (2005). Corumbella, an Ediacaran-Grade Organism from the Late Neoproterozoic of Brazil. Palaeogeogr. Palaeoclimatol. Palaeoecol. 220 (1-2), 7–18. doi:10.1016/j.palaeo.2003.01.001
Barth, G., Kozur, H. W., Franz, M., and Weems, R. E. (2013). “Paraconularia Bachmanni N. Sp. From the Germanic Upper Triassic, the First Fresh-Water Occurrence of a Conulariid,” in The Triassic System. Editors L. H. Tanner, J. A. Spielmann, and S. G. Lucas (New Mexico Museum of Natural History and Science, Bulletin), 61, 42–47.
Becker-Kerber, B., Pacheco, M. L. A. F., Rudnitzki, I. D., Galante, D., Rodrigues, F., and Leme, J. d. M. (2017). Ecological Interactions in Cloudina from the Ediacaran of Brazil: Implications for the Rise of Animal Biomineralization. Sci. Rep. 7, 5482. doi:10.1038/s41598-017-05753-8
Blair, J. E., and Hedges, S. B. (2005). Molecular Clocks Do Not Support the Cambrian Explosion. Mol. Biol. Evol. 22 (3), 387–390. doi:10.1093/molbev/msi039
Bobrovskiy, I., Hope, J. M., Ivantsov, A., Nettersheim, B. J., Hallmann, C., and Brocks, J. J. (2018). Ancient Steroids Establish the Ediacaran Fossil Dickinsonia as One of the Earliest Animals. Science 361 (6408), 1246–1249. doi:10.1126/science.aat7228
Boero, F., Bouillon, J., and Piraino, S. (2005). The Role of Cnidaria in Evolution and Ecology. Italian J. Zoology 72, 65–71. doi:10.1080/11250000509356654
Boggiani, P. C., Gaucher, C., Sial, A. N., Babinski, M., Simon, C. M., Riccomini, C., et al. (2010). Chemostratigraphy of the Tamengo Formation (Corumbá Group, Brazil): A Contribution to the Calibration of the Ediacaran Carbon-Isotope Curve. Precambrian Res. 182, 382–401. doi:10.1016/j.precamres.2010.06.003
Bruthansová, J., and Van Iten, H. (2020). Invertebrate Epibionts on Ordovician Conulariids from the Prague Basin (Czech Republic, Bohemia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 558, 109963. doi:10.1016/j.palaeo.2020.109963
Clapham, M. E., and Narbonne, G. M. (2002). Ediacaran Epifaunal Tiering. Geol 30 (7), 627–630. doi:10.1130/0091-7613(2002)030<0627:eet>2.0.co;2
Cracknell, K., Garcia-Bellido, J. G., Gehling, M. J., Ankor, S. A. F., García-Bellido, D. C., Gehling, J. G., et al. (2021). Pentaradial Eukaryote Suggests Expansion of Suspension Feeding in White Sea-Aged Ediacaran Communities. Sci. Rep. 11, 4121. doi:10.1038/s41598-021-83452-1
Darroch, S. A. F., Boag, T. H., Racicot, R. A., Tweedt, S., Mason, S. J., Erwin, D. H., et al. (2016). A Mixed Ediacaran-Metazoan Assemblage from the Zaris Sub-Basin, Namibia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 459, 198–208. doi:10.1016/j.palaeo.2016.07.003
Darroch, S. A. F., Cribb, A. T., Buatois, L. A., Germs, G. J. B., Kenchington, C. G., Smith, E. F., et al. (2020). The Trace Fossil Record of the Nama Group, Namibia: Exploring the Terminal Ediacaran Roots of the Cambrian Explosion. Earth-Science Rev. 212, 103435. doi:10.1016/j.earscirev.2020.103435
Darroch, S. A. F., Smith, E. F., Laflamme, M., and Erwin, D. H. (2018a). Ediacaran Extinction and Cambrian Explosion. Trends Ecol. Evol. 33, 653–663. doi:10.1016/j.tree.2018.06.003
Darroch, S. A. F., Laflamme, M., and Wagner, P. J. (2018b). High Ecological Complexity in Benthic Ediacaran Communities. Nat. Ecol. Evol. 2, 1541–1547. doi:10.1038/s41559-018-0663-7
Darroch, S. A. F., Sperling, E. A., Boag, T. H., Racicot, R. A., Mason, S. J., Morgan, A. S., et al. (2015). Biotic Replacement and Mass Extinction of the Ediacara Biota. Proc. R. Soc. B 282 (1814), 20151003. doi:10.1098/rspb.2015.1003
de Koninck, L. G. (1883). Faune du calcaires carbonifère de la Belgique, Partie 4, Gasteropodes. Ann. Musée R. d’Histoire Nat. Belg. Ser. Paleontol. 8, 193–213.
Dohrmann, M., and Wörheide, G. (2017). Dating Early Animal Evolution Using Phylogenomic Data. Sci. Rep. 7, 3599. doi:10.1038/s41598-017-03791-w
De Moraes Leme, J., Guimarães Simões, M., Carlos Marques, A., Van Iten, H., and Marques, A. C. (2008). Cladistic Analysis of the Suborder Conulariina Miller and Gurley, 1896 (Cnidaria, Scyphozoa; Vendian-Triassic). Palaeontology 51 (3), 649–662. doi:10.1111/j.1475-4983.2008.00775.x
dos Reis, M., Thawornwattana, Y., Angelis, K., Telford, M. J., Donoghue, P. C. J., and Yang, Z. (2015). Uncertainty in the Timing of Origin of Animals and the Limits of Precision in Molecular Timescales. Curr. Biol. 25 (22), 2939–2950. doi:10.1016/j.cub.2015.09.066
Driscoll, E. G. (1963). Paraconularia Newberryi (Winchell) and Other Lower Mississippian Conulariids from Michigan, Ohio, Indiana, and Iowa, 18. Michigan: Contributions from the Museum of Paleontology of the University of Michigan, 33–46.
Droser, M. L., and Gehling, J. G. (2015). The Advent of Animals: The View from the Ediacaran. Proc. Natl. Acad. Sci. U.S.A. 112, 4865–4870. doi:10.1073/pnas.1403669112
Dunn, F. S., Liu, A. G., Grazhdankin, D. V., Vixseboxse, P., Flannery-Sutherland, J., Green, E., et al. (2021). The Developmental Biology of Charnia and the Eumetazoan Affinity of the Ediacaran Rangeomorphs. Sci. Adv. 7 (30), 1–13 13. doi:10.1126/sciadv.abe0291
Dzik, J., Baliński, A., and Sun, Y. (2017). The Origin of Tetraradial Symmetry in Cnidarians. Lethaia 50, 306–321. doi:10.1111/let.12199
Erwin, D. H., Laflamme, M., Tweedt, S. M., Sperling, E. A., Pisani, D., and Peterson, K. J. (2011). The Cambrian Conundrum: Early Divergence and Later Ecological Success in the Early History of Animals. Science 334, 1091–1097. doi:10.1126/science.1206375
Escalante-Ruiz, A. R., Quiroz-Barroso, S. A., and Tovar, F. S. (2014). Conuláridos misisípicos de Nochixtlán, Oaxaca, México: consideraciones sistemáticas, estratigráficas y paleobiogeográficas. Rev. Bras. Paleontol. 17 (2), 195–206. doi:10.4072/rbp.2014.2.07
Fedonkin, M. A., Gehling, J. G., Grey, K., Narbonne, G. M., and Vickers Rich, P. (2007). The Rise of Animals: Evolution and Diversification of the Kingdom Animalia. Baltimore: The Johns Hopkins University Press, 326.
Fedonkin, M. A. (1992). “Vendian Faunas and the Early Evolution of Metazoa,” in Origin and Early Evolution of the Metazoa. Topics in Geobiology. Editors J. H. Lipps, and P. W. Signor (Boston, MA: Springer), 10. doi:10.1007/978-1-4899-2427-8-4
Fedonkin, M. A., and Waggoner, B. M. (1997). The Late Precambrian Fossil Kimberella Is a Mollusc-like Bilaterian Organism. Nature 388, 868–871. doi:10.1038/42242
Ford, R. C., Van Iten, H., and Clark, G. R. (2016). Microstructure and Composition of the Periderm of Conulariids. J. Paleontol. 90 (3), 389–399. doi:10.1017/jpa.2016.63
Gaucher, C., Boggiani, P. C., Sprechmann, P., Sial, A. N., and Fairchild, T. R. (2003). Integrated correlation of the Vendian to Cambrian Arroyo del Soldado and Corumbá Groups (Uruguay and Brazil): palaeogeographic, palaeoclimatic and palaeobiologic implications. Precambrian Res. 120, 241–278. doi:10.1016/s0301-9268(02)00140-7
Gehling, J. G. (1999). Microbial Mats in Terminal Proterozoic Siliciclastics: Ediacaran Death Masks. Palaios 14 (1), 40–57. doi:10.2307/3515360
Gibson, B. M., Rahman, I. A., Maloney, K. M., Racicot, R. A., Mocke, H., LaflammeDarroch, M. S. A. F., et al. (2019). Gregarious Suspension Feeding in a Modular Ediacaran Organism. Sci. Adv. 5, eaaw0260. doi:10.1126/sciadv.aaw0260
Götte, A. (1887). Entwicklungs Geschichte der Aurelia aurita und Cotylorhiza tuberculata. Abh. Entwicklungsgeschichte Tiere 4, 1–79.
Grazhdankin, D. (2016). Forbidden Fruits in the Garden of Ediacara. Paläontologische Z. 90, 649–657. doi:10.1007/s12542-016-0327-3
Grazhdankin, D., Nagovitsin, K., Golubkova, E., Karlova, G., Kochnev, B., Rogov, V., et al. (2020). Doushantuo-Pertatataka-type Acanthomorphs and Ediacaran Ecosystem Stability. Geology 48 (7), 708–712. doi:10.1130/G47467.1
Grazhdankin, D. (2014). Patterns of Evolution of the Ediacaran Soft-Bodied Biota. J. Paleontol. 88, 269–283. doi:10.1666/13-072
Grotzinger, J. P., Watters, W. A., and Knoll, A. H. (2000). Calcified Metazoans in Thrombolite-Stromatolite Reefs of the Terminal Proterozoic Nama Group, Namibia. Paleobiology 26 (3), 334–359. doi:10.1666/0094-8373(2000)026<0334:cmitsr>2.0.co;2
Guo, J., Han, J., Iten, H. V., Wang, X., Qiang, Y., Song, Z., et al. (2020a). A Fourteen-Faced Hexangulaconulariid from the Early Cambrian (Stage 2) Yanjiahe Formation, South China. J. Paleontol. 94 (1), 45–55. doi:10.1017/jpa.2019.56
Guo, J., Han, J., Van Iten, H., Song, Z., Qiang, Y., Wang, W., et al. (2020b). A New Tetraradial Olivooid (Medusozoa) from the Lower Cambrian (Stage 2) Yanjiahe Formation, South China. J. Paleontol. 94 (3), 457–466. doi:10.1017/jpa.2019.101
Guo, J., Han, J., Van Iten, H., Song, Z., Qiang, Y., Wang, W., et al. (2021). A Ten-Faced Hexangulaconulariid from Cambrian Stage 2 of South China. J. Paleontol. 95 (5), 957–964. doi:10.1017/jpa.2021.25
Han, J., Li, G., Wang, X., Yang, X., Guo, J., Sasaki, O., et al. (2017). Olivooides-like Tube Aperture in Early Cambrian Carinachitids (Medusozoa, Cnidaria). J. Paleontol. 92, 3–13. doi:10.1017/jpa.2017.10
Hedges, S., Blair, J. E., Venturi, M. L., and Shoe, J. L. (2004). A Molecular Timescale of Eukaryote Evolution and the Rise of Complex Multicellular Life. BMC Evol. Biol. 4, 2. doi:10.1186/1471-2148-4-2
Hofmann, H. J., and Mountjoy, E. W. (2001). Namacalathus-Cloudina Assemblage in Neoproterozoic Miette Group (Byng Formation), British Columbia: Canada's Oldest Shelly Fossils. Geology 29 (12), 1091–1094. doi:10.1130/0091-761310.1130/0091-7613(2001)029<1091:ncainm>2.0.co;2
Hua, H., Pratt, B. R., and Zhang, L.-Y. (2003). Borings in Cloudina Shells: Complex Predator-Prey Dynamics in the Terminal Neoproterozoic. Palaios 18, 454–459. doi:10.1669/0883-1351(2003)018<0454:bicscp>2.0.co;2
Hughes, N. C., Gunderson, G. O., and Weedon, M. J. (2000). Late Cambrian Conulariids from Wisconsin and Minnesota. J. Paleontology 74, 828–838. doi:10.1666/0022-3360(2000)074<0828:lccfwa>2.0.co;2
Ivantsov, A. Y., and Fedonkin, M. A. (2002). Conulariid-like Fossil from the Vendian of Russia: a Metazoan Clade across the Proterozoic/Palaeozoic Boundary. Palaeontology 45, 1219–1229. doi:10.1111/1475-4983.00283
Ivantsov, A. Y., Vickers-Rich, P., Zakrevskaya, M. A., and Hall, M. (2019). Conical Thecae of Precambrian Macroorganisms. Paleontol. J. 53, 1134–1146. doi:10.1134/s0031030119110054
Jerre, F. (1994). Anatomy and Phylogenetic Significance of Eoconularia Loculata, a Conulariid from the Silurian of Gotland. Lethaia 27, 97–109. doi:10.1111/j.1502-3931.1994.tb01562.x
Kontorovich, A. E., Sokolov, B. S., Kontorovich, V. A., Varlamov, A. I., GrazhdankinEfimov, D. V. A. S., Efimov, A. S., et al. (2009). The First Section of Vendian Deposits in the Basement Complex of the West Siberian Petroleum Megabasin (Resulting from the Drilling of the Vostok-3 Parametric Borehole in the Eastern Tomsk Region). Dokl. Earth Sc. 425 (1), 219–222. doi:10.1134/s1028334x09020093
Kouchinsky, A., Bengtson, S., Runnegar, B., Skovsted, C., Steiner, M., and Vendrasco, M. (2012). Chronology of Early Cambrian Biomineralization. Geol. Mag. 149, 221–251. doi:10.1017/s0016756811000720
Laflamme, M., Darroch, S. A. F., Tweedt, S. M., Peterson, K. J., and Erwin, D. H. (2013). The End of the Ediacara Biota: Extinction, Biotic Replacement, or Cheshire Cat? Gondwana Res. 23, 558–573. doi:10.1016/j.gr.2012.11.004
Leme, J., Rodrigues, S. C., Simões, M. G., and Van Iten, H. (2004). Sistemática dos conulários (Cnidaria) da Formação Ponta Grossa (Devoniano), Estado Do Paraná, bacia Do Paraná, Brasil. Rev. Bras. Paleontol. 7 (2), 213–222. doi:10.4072/rbp.2004.2.14
Linnemann, U., Ovtcharova, M., Schaltegger, U., Gärtner, A., Hautmann, M., Geyer, G., et al. (2019). New High-Resolution Age Data from the Ediacaran-Cambrian Boundary Indicate Rapid, Ecologically Driven Onset of the Cambrian Explosion. Terra nova. 31 (1), 49–58. doi:10.1111/ter.12368
Lipps, J. H., and Culver, S. J. (2002). The Trophic Role of Marine Microorganisms Through Time. Paleontol. Soc. Pap. 8, 69–92. doi:10.1017/S1089332600001066
Lucas, S. G. (2012). The Extinction of the Conulariids. Geosciences 2, 1–10. doi:10.3390/geosciences2010001
Lukševičs, E. (2020). First Record of Conulariids from the Tērvete Formation, Upper Devonian of Latvia. Est. J. Earth Sci. 69, 262–268. doi:10.3176/earth.2020.21
Marques, A. C., and Collins, A. G. (2004). Cladistic Analysis of Medusozoa and Cnidarian Evolution. Invertebr. Biol. 123, 23–42. doi:10.1111/j.1744-7410.2004.tb00139.x
Miller, S. A., and Gurley, W. F. E. (1896). New Species of Paleozoic Invertebrates from Illinois and Other States. Ill. State Mus. Nat. Hist. 11, 1–50.
Min, H., Wang, X.-L., and Wang, H. (2021). A New Permian Species of Paraconularia from Abaga Banner, Inner Mongolia. Palaeoworld. doi:10.1016/j.palwor.2021.10.002
Morris, S. C., and Menge, C. (1992). Carinachitiids, Hexangulaconulariids, and Punctatus: Problematic Metazoans from the Early Cambrian of South China. J. Paleontol. 66 (3), 384–406. doi:10.1017/s0022336000033953
Muscente, A. D., Bykova, N., Boag, T. H., Buatois, L. A., Mángano, M. G., Eleish, A., et al. (2019). Ediacaran Biozones Identified with Network Analysis Provide Evidence for Pulsed Extinctions of Early Complex Life. Nat. Commun. 10, 911. doi:10.1038/s41467-019-08837-3
Narbonne, G. M. (2005). The Ediacara Biota: Neoproterozoic Origin of Animals and Their Ecosystems. Annu. Rev. Earth Planet. Sci. 33 (1), 421–442. doi:10.1146/annurev.earth.33.092203.122519
Pacheco, M. L. A. F., Leme, J. M., and Machado, A. F. (2011). Taphonomic analysis and geometric modelling for the reconstitution of the Ediacaran metazoan Corumbella werneri Hahn et al. 1982 (Tamengo Formation, Corumbá Basin, Brazil). Journal of Taphonomy 9 (4), 269–283.
Pacheco, M. L. A. F., Galante, D., Rodrigues, F., de M. Leme, J., Bidola, P., Hagadorn, W., et al. (2015). Insights into the Skeletonization, Lifestyle, and Affinity of the Unusual Ediacaran Fossil Corumbella. Plos One 10, e0114219. doi:10.1371/journal.pone.0114219
Parfrey, S. M. (1982). Palaeozoic Conulariids from Tasmania. Alcheringa An Australas. J. Palaeontol. 6, 69–77. doi:10.1080/03115518208565421
Parry, L. A., Boggiani, P. C., Condon, D. J., Garwood, R. J., Leme, J. d. M., McIlroy, D., et al. (2017). Ichnological Evidence for Meiofaunal Bilaterians from the Terminal Ediacaran and Earliest Cambrian of Brazil. Nat. Ecol. Evol. 1, 1455–1464. doi:10.1038/s41559-017-0301-9
Pearse, V., Pearse, J., Buchsbaum, M., and Buchsbaum, R. (1987). Living Invertebrates. Pacific Grove, California: Boxwood Press, 848.
Peterson, K. W. (1979). “Development of Coloniality in Hydrozoa,” in Biology and Systematics of Colonial Organism. Editors G. Larwood, and B. R. Rosen (New York: Academic Press).
Rodrigues, S., Simões, M., and Leme, J. M. (2003). Tafonomia Comparada Dos Conulatae (Cnidaria), Formação Ponta Grossa (Devoniano), Bacia Do Paraná, Estado Do Paraná. Rev. Bras. Geociências 33, 165–186. doi:10.25249/0375-7536.2003334379388
Runnegar, B. (1982). The Cambrian Explosion: Animals or Fossils? J. Geol. Soc. Aust. 29 (3-4), 395–411. doi:10.1080/00167618208729222
Schiffbauer, J. D., Huntley, J. W., O’Neil, G. R., Darroch, S. A. F., Laflamme, M., and Cai, Y. (2016). The Latest Ediacaran Wormworld Fauna: Setting the Ecological Stage for the Cambrian Explosion. GSA Today 26 (11), 4–11. doi:10.1130/gsatg265a.1
Seilacher, A. (1999). Biomat-related Lifestyles in the Precambrian. Palaios 14, 86–93. doi:10.2307/3515363
Swami, N., Satish, S. C., Laishram, R., and Dharwadkar, A. (2017). First Record of Extinct Paraconularia (Cnidaria, Scyphozoa) from Tethyan Sequence (Upper Permian) of Spiti Valley, Himachal Himalaya, India. Palaeontol. Electron. 2042A (3), 1–6. doi:10.26879/736
Thomas, G. A. (1969). Notoconularia, a New Conularid Genus from the Permian of Eastern Australia. J. Paleontology 43 (5), 1283–1290.
Van Iten, H., Leme, J. M., Marques, A. C., and Simões, M. G. (2013). Alternative interpretations of some earliest Ediacaran fossils from China. Acta Palaeontologica Polonica 51 (1), 111–113. doi:10.4202/app.2011.0096
Van Iten, H., Marques, A. C., Leme, J. d. M., Pacheco, M. L. A. F., and Simões, M. G. (2014a). Origin and Early Diversification of the Phylum Cnidaria Verrill: Major Developments in the Analysis of the Taxon's Proterozoic-Cambrian History. Palaeontology 57, 677–690. doi:10.1111/pala.12116
Van Iten, H., Burkey, M. H., Leme, J. M., and Marques, A. C. (2014b). Cladistics and Mass Extinctions: the Example of Conulariids (Scyphozoa, Cnidaria) and the End Ordovician Extinction Event. GFF 136, 275–280. doi:10.1080/11035897.2014.880506
Van Iten, H., Fitzke, J. A., and Cox, R. S. (1996). Problematical Fossil Cnidarians from the Upper Ordovician of the North-Central USA. Palaeontology 39, 1037–1064.
Van Iten, H., Leme, J. M., Simões, M. G., Marques, A. C., and Collins, A. G. (2006a). Reassessment of the Phylogenetic Position of Conulariids (?Vendian-Triassic) within the Subphylum Medusozoa (Phylum Cnidaria). J. Syst. Palaeontol. 4 (1), 109–118. doi:10.1017/s1477201905001793
Van Iten, H., Leme, J. d. M., and Simões, M. G. (2006b). Additional observations on the gross morphology and microstructure of Baccaconularia Hughes, Gunderson et Weedon, 2000, a Cambrian (Furongian) conulariid from the north-central USA. Palaeoworld 15, 294–306. doi:10.1016/j.palwor.2006.10.006
Van Iten, H., Leme, J. M., De Moraes Leme, J., Coelho Rodrigues, S., and Guimaraes Simoes, M. (2005). Reinterpretation of a Conulariid-like Fossil from the Vendian of Russia. Palaeontology 48 (3), 619–622. doi:10.1111/j.1475-4983.2005.00471.x
Van Iten, H., Leme, J. M., Pacheco, M. L. A. F., Simões, M. G., Fairchild, T. R., Rodrigues, F., et al. (2016). “Origin and Early Diversification of Phylum Cnidaria: Key Macrofossils from the Ediacaran System of North and South America,” in The Cnidaria, Past, Present and Future. Editors S. Goffredo, and Z. Dubinsky (Cham: Springer International Publishing), 31–40. doi:10.1007/978-3-319-31305-4_3
Van Iten, H. (1992). Morphology and Phylogenetic Significance of the Corners and Midlines of the Conulariid Test. Paleontology 35, 335–358.
Van Iten, H., Moussa, K., and Yahaya, M. (2008). Conulariids of the Upper Talak Formation (Mississippian, Visean) of Northern Niger (West Africa). J. Paleontology 82, 178–182. doi:10.1666/06-083.1
Vaziri, S. H., Majidifard, M. R., and Laflamme, M. (2018). Diverse Assemblage of Ediacaran Fossils from Central Iran. Sci. Rep. 8, 5060. doi:10.1038/s41598-018-23442-y
Verrill, A. E. (1865). Classification of Polyps (Extract Condensed from Synopsis of the Polyps and Corals of the North Pacific Exploring Expedition under Commodore C. Ringgold and Captain John Rogers, U.S.N.). Commun. Essex Inst. 4, 145–152.
Vidal, G., and Moczydłowska-Vidal, M. (1997). Biodiversity, Speciation, and Extinction Trends of Proterozoic and Cambrian Phytoplankton. Paleobiology 23 (2), 230–246. doi:10.1017/s0094837300016808
Walde, D. H.-G., Weber, B., Erdtmann, B. D., and Steiner, M. (2019). Taphonomy ofCorumbella Wernerifrom the Ediacaran of Brazil: Sinotubulitid Tube or Conulariid Test? Alcheringa An Australas. J. Palaeontol. 43, 335–350. doi:10.1080/03115518.2019.1615551
Walde, D. H. G., do Carmo, D. A., Guimarães, E. M., Vieira, L. C., Erdtmann, B.-D., Sanchez, E. A. M., et al. (2015). New Aspects of Neoproterozoic-Cambrian Transition in the Corumbá Region (State of Mato Grosso Do Sul, Brazil). Ann. Paléontologie 101, 213–224. doi:10.1016/j.annpal.2015.07.002
Wan, B., Yuan, X., Chen, Z., Guan, C., Pang, K., Tang, Q., et al. (2016). Systematic Description of Putative Animal Fossils from the Early E Diacaran L Antian F Ormation of S Outh C Hina. Palaeontology 59, 515–532. doi:10.1111/pala.12242
Warren, L. V., Pacheco, M. L. A. F., Fairchild, T. R., Simões, M. G., Riccomini, C., Boggiani, P. C., et al. (2012). The Dawn of Animal Skeletogenesis: Ultrastructural Analysis of the Ediacaran Metazoan Corumbella Werneri. Geology 40, 691–694. doi:10.1130/g33005.1
Warren, L. V., Quaglio, F., Riccomini, C., Simões, M. G., Poiré, D. G., Strikis, N. M., et al. (2014). The Puzzle Assembled: Ediacaran Guide Fossil Cloudina Reveals an Old Proto-Gondwana Seaway. Geology 42, 391–394. doi:10.1130/g35304.1
Warren, L. V., QuaglioSimões, F. M. G., SimõesRiccomini, M. G. C., Gaucher, C., Riccomini, C., Poiré, D. G., et al. (2017). Cloudina-Corumbella-Namacalathus Association from the Itapucumi Group, Paraguay: Increasing Ecosystem Complexity and Tiering at the End of the Ediacaran. Precambrian Res. 298, 79–87. doi:10.1016/j.precamres.2017.05.003
Waterhouse, J. B. (1986). Late Palaeozoic Scyphozoa and Brachiopoda (Inarticulata, Strophomenida, Productida and Rhynchonellida) from the Southeast Bower Basin, Australia. Palaeontographica 193, 1–77.
Waterhouse, J. B. (1979). Permian and Triassic Conulariid Species from New Zealand. J. R. Soc. N. Z. 9 (4), 475–489. doi:10.1080/03036758.1979.10421833
Werner, B. (1966). Stephanoscyphus (Scyphozoa, Coronatae) und seine direkte Abstammung von den fossilen Conulata. Helgol. Wiss. Meeresunters 13, 317–347. doi:10.1007/bf01611953
Wood, R. A., Grotzinger, J. P., and Dickson, J. A. D. (2002). Proterozoic Modular Biomineralized Metazoan from the Nama Group, Namibia. Science 296, 2383–2386. doi:10.1126/science.1071599
Wood, R., Liu, A. G., Bowyer, F., Wilby, P. R., Dunn, F. S., Kenchington, C. G., et al. (2019). Integrated Records of Environmental Change and Evolution Challenge the Cambrian Explosion. Nat. Ecol. Evol. 3, 528–538. doi:10.1038/s41559-019-0821-6
Xiao, S., and Laflamme, M. (2009). On the Eve of Animal Radiation: Phylogeny, Ecology and Evolution of the Ediacara Biota. Trends Ecol. Evol. 24 (1), 31–40. doi:10.1016/j.tree.2008.07.015
Xiao, S., Narbonne, G. M., Zhou, C., Laflamme, M., Grazhdankin, D. V., Moczydlowska-Vidal, M., et al. (2016). Towards an Ediacaran Time Scale: Problems, Protocols, and Prospects. Episodes 39 (4), 540–555. doi:10.18814/epiiugs/2016/v39i4/103886
Xu, G., and Li, F. (1979). New Conulariid Genera and Species in China. Acta Geol. Sin. 53 (2), 91–98.
Young, G. A., and Hagadorn, J. W. (2020). Evolving Preservation and Facies Distribution of Fossil Jellyfish: a Slowly Closing Taphonomic Window. Boll. della Soc. Paleontol. Ital. 59, 185–203. doi:10.4435/BSPI.2020.22
Yuan, X., Chen, Z., Xiao, S., Zhou, C., and Hua, H. (2011). An Early Ediacaran Assemblage of Macroscopic and Morphologically Differentiated Eukaryotes. Nature 470, 390–393. doi:10.1038/nature09810
Zhao, Y., Vinther, J., Parry, L. A., Wei, F., Green, E., Pisani, D., et al. (2019). Cambrian Sessile, Suspension Feeding Stem-Group Ctenophores and Evolution of the Comb Jelly Body Plan. Curr. Biol. 29, 1112–1125. doi:10.1016/j.cub.2019.02.036
Zhu, M., Zhuravlev, A. Y., Wood, R. A., Zhao, F., and Sukhov, S. S. (2017). A Deep Root for the Cambrian Explosion: Implications of New Bio- and Chemostratigraphy from the Siberian Platform. Geology 45, 459–462. doi:10.1130/g38865.1
Keywords: conulariids, systematics, Ediacaran, Tamengo Formation, Paleoecology
Citation: Leme JM, Van Iten H and Simões MG (2022) A New Conulariid (Cnidaria, Scyphozoa) From the Terminal Ediacaran of Brazil. Front. Earth Sci. 10:777746. doi: 10.3389/feart.2022.777746
Received: 15 September 2021; Accepted: 18 March 2022;
Published: 08 June 2022.
Edited by:
Simon Darroch, Vanderbilt University, United StatesReviewed by:
Marc Laflamme, University of Toronto Mississauga, CanadaRoss Anderson, University of Oxford, United Kingdom
Frankie Dunn, University of Oxford, United Kingdom
Copyright © 2022 Leme, Van Iten and Simões. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juliana M. Leme, bGVtZUB1c3AuYnI=