
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci. , 08 June 2022
Sec. Paleontology
Volume 10 - 2022 | https://doi.org/10.3389/feart.2022.757629
This article is part of the Research Topic Yedoma Permafrost Landscapes as Past Archives, Present and Future Change Areas View all 27 articles
 Tatiana V. Kuznetsova1,2*
Tatiana V. Kuznetsova1,2* Sebastian Wetterich3,4
Sebastian Wetterich3,4 Heidrun Matthes5
Heidrun Matthes5 Vladimir E. Tumskoy6,7
Vladimir E. Tumskoy6,7 Lutz Schirrmeister3*
Lutz Schirrmeister3*The Yedoma Ice Complex in northern Yakutia provides perfect preservation conditions for frozen remains of vertebrate animals. Even complete mummified specimens of the late Pleistocene Beringian Mammoth fauna such as woolly mammoth, woolly rhinoceros, horse, and bison are occasionally found in permafrost deposits across eastern Siberia, i.e., in West Beringia, although bones are much more commonly found. The present study characterizes mammal bones from late Pleistocene and Holocene permafrost deposits exposed on the Oyogos Yar coast, part of the southern shore of the Dmitry Laptev Strait that connects the Laptev and East Siberian seas. The study applies a method to characterize fossil bone samples by the location of their discovery and by the accuracy of their relation to a depositional horizon. We analyzed a total of 38 finite radiocarbon ages of bone material from mammoth, horse, and musk ox, spanning from about 48.8 to 4.5 ka BP and including both our own data and data from the literature, in addition to previous publications that reported numerous bones with infinite ages from the Oyogos Yar coast. The distribution of bones and tooth along the coastal permafrost exposure is not uniform; it depends upon whether the material was found in situ, on thermo-terraces, or on the shore. The overall bone collection consists of 13 species of which Mammuthus primigenius (woolly mammoth, 41%), Bison priscus (bison, 19%), Equus ex gr., caballus (horse, 19%), and Rangifer tarandus (reindeer, 16%) predominate. The fossil bone species distribution is similar to those of other prominent Yedoma outcrops in the region, i.e., on Bykovsky Peninsula and on Bol’shoy Lyakhovsky Island. Correlation analysis shows that the Oyogos Yar bone sampling sites of different geomorphological settings are similar to each other but not to all sampling sites within the other two locations on Bykovsky Peninsula and on Bol’shoy Lykahovsky Island. High similarities in terms of correlation coefficients between specific sampling sites are often not represented in the cluster analysis.
Numerous finds of bones and soft tissues of large mammals have been made on the southern shore of the Dmitry Laptev Strait on the Oyogos Yar coast (Figure 1) since the 19th century. The first detailed description and topographic survey of the Oyogos Yar coast was carried out in 1909 by two members of an expedition of the Ministry of Trade and Industry of Russia, E.F. Skvortsov and N.A. Iyudin (Skvortsov, 1914; Skvortsov, 1930). They noted abundant bone fossils on the beach, especially those of woolly mammoths. Here, bones and tooth accumulate occasionally due to thermo-denudation and thermo-erosion of the permafrost coast and subsequent slumping. The Yana-Indigirka Lowland, including the Oyogos Yar coast, has been actively studied since the end of the 1920’s. Mammal bones from this area were sent to the zoological, geological, and paleontological institutes in Moscow, where they have been stored until now (Kuznetsova et al., 2004; Kuznetsova and Starodubtseva, 2009).
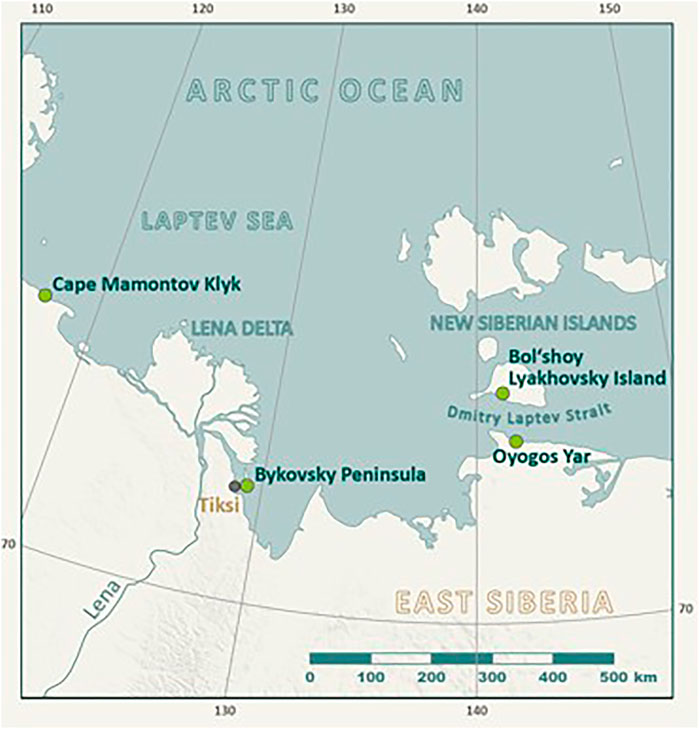
FIGURE 1. Study region in the East Siberian Arctic showing the Oyogos Yar study site and other relevant study sites on the New Siberian Archipelago (Bol’shoy Lyakhovsky Island) and at the shore of the Laptev Sea (Cape Mamontov Klyk, Bykovsky Peninsula, and Lena Delta) mentioned in the paper.
In the 20th century, Grigoriev (1932), Spizharskii (1940), Gusev (1958), Romanovskii (1958), Romanovskii (1961a), Romanovskii (1961b), and Kuznetsova (1965) actively studied the geological structure of this region and developed stratigraphic schemes for the late Quaternary deposits exposed here. The stratigraphic scheme for the Yana-Indigirka coastal plain by Gusev (1958) defined for the first time a “Mammoth horizon” occupying a distinct stratigraphic position. Later stratigraphic schemes for the region by Ivanov (1972) and Prokhorova and Ivanov (1973) characterized several formations of the middle and late Pleistocene by the presence of Mammoth fauna remains. Active exploration for fossil mammoth tusks resumed at the end of the 20th century, even in previously inaccessible regions. Consequently, the number of harvested tusks sharply increased. Furthermore, carcasses and soft tissue material of large Mammoth fauna animals became available for paleontological research (e.g., Boeskorov et al., 2013; Boeskorov et al., 2014).
According to Sher et al. (2005) the taxonomic composition of mammal bone assemblages can be affected by external factors, especially local geological processes and taphonomic conditions. Therefore, such collections may provide a modified picture of the relative abundances of species in the past. Furthermore, the typical patterns of taphonomy (Efremov, 1950) suggest that fossil bones represent only a small fraction of previously existing animals.
The Beringia Land Bridge connected Eurasia and North America during the sea-level low stand of the last glacial period. Therefore, vast shelf and lowland areas between the Scandinavian-Kara ice sheets and the Laurentide ice sheet remained unglaciated and provided space for extensive permafrost formation in polygonal tundra landscapes. Here, abundant tundra-steppe vegetation sustained the Mammoth fauna, which was characterized by large grazers such as woolly mammoth, bison, musk ox, horse, reindeer, saiga, and woolly rhinoceros. Fossil evidence of this unique ecosystem of the last ice age still attracts public interest, while paleo-environmental research that employs floral and faunal fossils enables insights into ecosystem functioning as well as mechanisms of adaptation to changing climatic conditions (Sher et al., 2005; Andreev et al., 2011; Nikolskiy et al., 2011; Pitulko et al., 2017; Pavlova and Pitulko, 2020). In this context, the present study contributes to paleo-environmental research in West Beringia and aims 1) to describe and document the paleozoological characteristics of the late Pleistocene and Holocene deposits of the Oyogos Yar coast, 2) to assign whenever possible the mammal bone findings to cryostratigraphic units either by dating or by detailed documentation of the sites of finds in a systematic manner, and 3) to deduce the local distribution pattern of mammal bones onthe Oyogos Yar coast and to compare the composition of the West Beringian Mammoth fauna to that of mammal bone collections from neighboring prominent localities on the New Siberian Islands and on the Bykovsky Peninsula.
Oyogos Yar is a stretch of the Arctic Ocean coastline located in the southwesternmost part of the East Siberian Sea, and forming part of the southern coast of the Dmitry Laptev Strait (Figure 1). Our study was performed at the Kondrat’eva Yedoma and the surrounding thermokarst basins (alases in Russian) in the eastern part of the Oyogos Yar coast. The Kondrat’eva Yedoma is the largest permafrost outcrop along this part of the coastline and stretches for about 5 km, from the mouth of the Kondrat’eva River to the west (between about 72.683°N, 143.475°E and 72.672°N, 143.635°E). The height of the Kondrat’eva Yedoma reaches 41 m above sea level (asl). The paleontological material discussed in this study was collected in 2002 and 2007 over 10 km of coastline, on the exposures as well as on the beach (Kuznetsova, 2003; Kuznetsova, 2008).
The modern periglacial landscape of the Oyogos Yar hinterland is characterized by Yedoma uplands intersected by alases and thermo-erosional gullies and valleys. Based on the cryostratigraphy exposed on the coast and its geocryological properties, such as ice content, the interplay of thermo-dedunation and thermo-abrasion erosional processes has shaped the coastal topography into steep bluffs and thaw slumps (thermo-cirques in Russian), with thermo-terraces topped by thermokarst mounds (baidzherakhs in Russian) that represent sedimentary centers of ice-wedge polygons that remained after the wedge ice melted (Figure 2). The Oyogos Yar coast erodes at an overall long-term rate of up to −6.5 ± 0.2 ma−1 (Günther et al., 2013). Such rapid permafrost erosion provides annually renewed access to formerly frozen material, including the fossil mammal bones which are the focus of our study.

FIGURE 2. Coastal exposures at the Kondrat’eva Yedoma (Dmitry Laptev Strait) showing (A) the Yedoma Ice Complex headwall (note climber dressed in blue at the top of the frozen ground deposit for scale), (B) the thermo-terrace below the headwall, (C) an ice wedge pseudomorph in Krest Yuryakh deposits, and (D) an ice wedge pseudomorph in lateglacial to Holocene alas deposits. Note in situ tusk findings in (C) and (D). Photographs in August 2007 by T. Opel, AWI (A), F. Kienast, Senckenberg Weimar (B), L. Schirrmeister, AWI (C), and V.V. Kunitsky, MPI Yakutsk (D).
In recent years, geocryological investigations of the Oyogos Yar coastal exposures have been undertaken during Russian, Russian-Japanese, and Russian-German efforts that provided a general understanding of the local cryostratigraphic horizons (Figure 3) and their formation (Konishchev and Kolesnikov, 1981; Tomidiaro et al., 1982; Nagaoka et al., 1995; Tumskoy, 2012), as well as insights into paleo-ecology and paleo-climate (Wetterich et al., 2009; Andreev et al., 2011; Kienast et al., 2011; Opel et al., 2011; Boeskorov et al., 2013; Rudaya et al., 2015; Wetterich et al., 2016; Opel et al., 2017a; Opel et al., 2017b; Neretina et al., 2020). Based on the stratigraphic chart proposed by Tumskoy and Kuznetsova (2022) for northern Yakutia, and follow-up permafrost studies that included dating results (Schirrmeister et al., 2002a; Wetterich et al., 2014; Wetterich et al., 2016; Wetterich et al., 2019; Opel et al., 2017a; Zimmermann et al., 2017; Wetterich et al., 2021a), the permafrost deposits exposed on both coasts of the Dmitry Laptev Strait discontinuously cover the last about 200,000 years before present and span from Marine Isotope Stage (MIS) 7 to MIS 1. At the study site, a Yedoma Ice Complex of MIS 3 age is preserved in the Yedoma upland, while adjacent thermokarst basins of lateglacial as in Figure 2 and MIS 1 age are composed of lacustrine and palustrine deposits and reach up to 15 m asl (Figure 3). Deposits underlying these two main sequences belong to the lacustrine Krest-Yuryakh stratum of MIS 5 age, which is commonly considered to represent the last Interglacial (e.g., Andreev et al., 2009; Wetterich et al., 2009; Kienast et al., 2011). Further MIS 5 deposits belong to the Buchchagy Ice Complex, while the chronostratigraphic position of the Kuchchugui stratum is still in question due to scarcity of geochronologic data (MIS 5 vs. MIS 6; Andreev et al., 2004; Tumskoy, 2012). After their formation, ice-rich permafrost deposits such as the Buchchagy Ice Complex or the Yedoma Ice Complex might have been subjected to thaw during successive warm periods such as the Last Interglacial or the Holocene. If thawing occurred, the remaining and refrozen mineral and organic depositional components are defined as taberal deposits (Kaplina, 2009). Taberal deposits are present in the cryostratigraphic record of the Oyogos Yar coast and are therefore considered in the study (Figure 3).
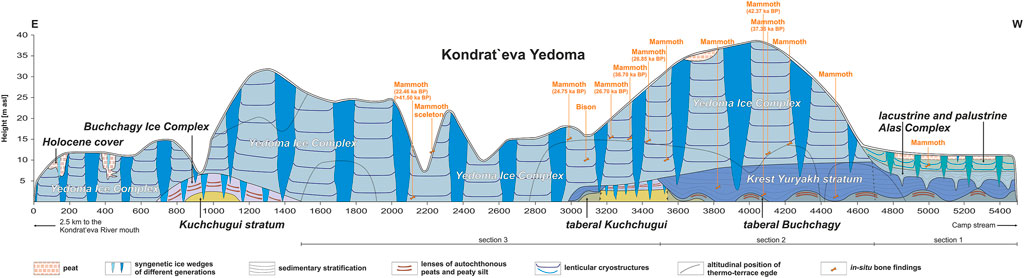
FIGURE 3. Schematic structure of the Kondrat’eva Yedoma exposure on the Oyogos Yar coast from 2.5 to 8 km west of the Kondrat’eva River mouth [redrawn with changes from Tumskoy and Kuznetsova (2022).
It is well known that well-preserved Mammoth fauna fossils can be found on the Oyogos Yar coast and at the Kondrat’eva Yedoma in particular (Smirnov, 2003). However, only two skeletal elements from the eastern part of the Oyogos Yar coast are stored in the Zoological Institute of the Russian Academy of Sciences (St. Petersburg, Russia); the upper left tooth of a woolly mammoth, transferred to the institute in 1972, and a horse skull without a lower jaw, found at the mouth of the Rebrova River and transferred to the collection in 1991 by M.V. Sablin and O.R. Potapova. Single finds of the remains of fossil mammals from the area of the Kondrat’eva Yedoma are stored in the Diamond and Precious Metals Geology Institute, Siberian Branch, RAS and the Mammoth Museum of the North-eastern Federal University (NEFU, Yakutsk, Russia).
In 2009 and 2010, two outstanding discoveries were made on the Oyogos Yar coast. In 2009, 30 km west of the mouth of the Kondrat’eva River, a part of the carcass of a woolly mammoth was found; this specimen is known as mammoth Yuka and was radiocarbon-dated to 34,300 +260/−240 a BP (GrA 53289; Boeskorov et al., 2013). The remains of the Yuka woolly mammoth include by the skull with cheek teeth and tusks, the lower jaw with cheek teeth, the lower parts of the legs with soft tissues, the skin, some soft tissues of the body, and part of the axial skeleton. The left ear, trunk, and lips are preserved on the head. There are three finger-like processes at the end of the trunk. The hair is gray-brown and dark brown on the body and red on the legs. The maximum length of the hair on the hips is 40–42 cm (Boeskorov et al., 2013). A fragment of a horse carcass was found in the same area in 2010, and radiocarbon-dated to 4,630 ± 30 a BP (GrA 54209; Boeskorov et al., 2013). Today, this partial horse carcass is named the Yukagir horse, of which the head and neck, part of the torso containing some internal organs, the hind legs, and the tail have been preserved. The dark brown mummified skin was preserved in fragments. Short hair, 4.5–7 cm long and so dark brown as to be almost black, is preserved on the distal parts of the legs only (Boeskorov et al., 2013).
A total of 43 radiocarbon dates for mammoth bones and tooth, of which 20 are finite dates for specimens from the mainland shore of the Dmitry Laptev Strait, was first presented by Nikolskiy and Basilyan (2004) and Nikolskiy et al. (2011). These age determinations were obtained for bones from three locations, namely Svyatoy Nos, Ulakhan-Taala, and Kondrat’eva Yedoma, and are discussed in the context of the present study.
Although scientific sampling of large mammal bone remains on the Oyogos Yar coast (Figure 1) has been taking place for more than 100 years, the paleontological collection of the Russian-German SYSTEM LAPTEV SEA collaboration (since 1998) is the most complete and representative for this coastal region. Mammal bones and bone fragments were collected on the Oyogos Yar coast during fieldwork in 2002 and 2007, and identified afterwards (Kuznetsova, 2003; Kuznetsova, 2008). For a comprehensive and complete study of the paleozoological material, all mammalian remains regardless of their state of preservation, including small bone fragments, were collected and identified. This allowed the most accurate possible estimation of the relative abundances of primary large mammal species of the late Pleistocene Mammoth fauna that inhabited this area. The overwhelming majority of the finds were identified down to species level using the collection of the Department of Osteology of the Zoological Museum of Lomonosov Moscow State University (Russia) as comparative material. Thus, we determined the paleozoological characteristics of the late Pleistocene and Holocene deposits of the eastern part of the Oyogos Yar coast. In 2002 and 2007, we collected 1977 samples of fossil bones and tooth (Table 1).
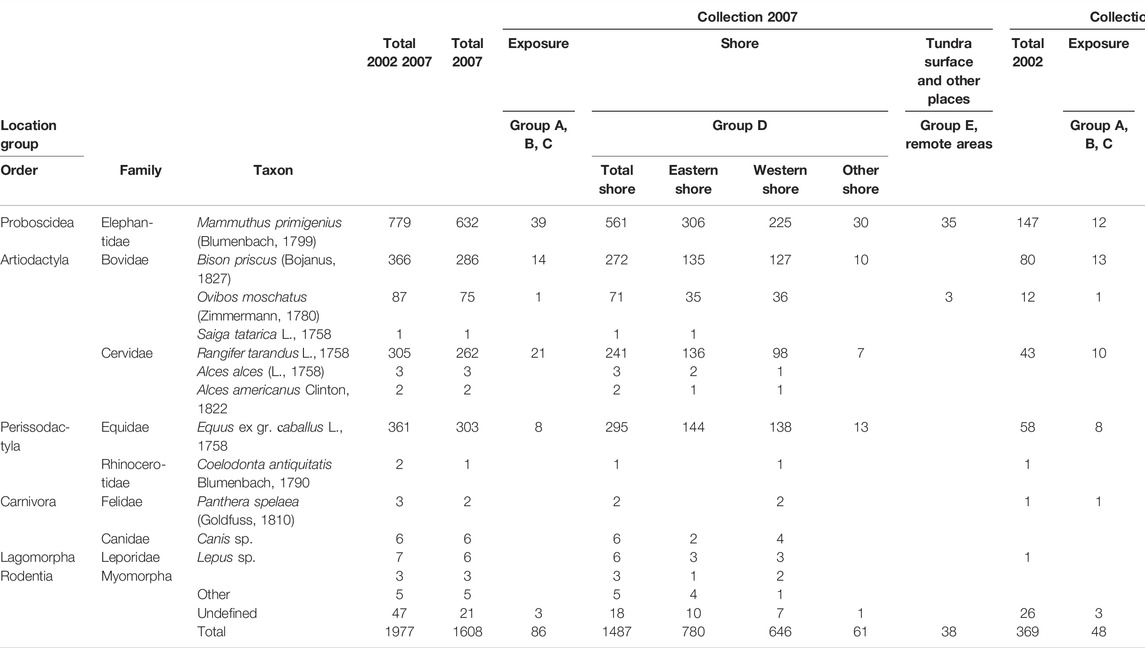
TABLE 1. Collection of mammalian remains sampled at Oyogos Yar in 2002 and 2007.
Statistical analyses were conducted in order to evaluate the degree of similarity among the species distributions for the Mammoth fauna at the different sampling sites associated with the three areas discussed: the Oyogos Yar coast at the Kondrat’eva Yedoma west of the mouth of the Kondrat’eva River, the southern coast of Bol’shoy Lyakhovsky Island near the mouth of the Zimov’e River, and on the Bykovsky Peninsula at the Mamontova Khayata Yedoma (Figure 1). First, Pearson correlation coefficients were calculated between the species distributions of all sampling sites to assess whether the level of uniformity among sites within one location characterized by different geomorphological settings is comparable to the level of uniformity among sites in different locations. Second, to further explore those differences and similarities, agglomerative hierarchical cluster analysis was then performed on the species counts for each sample site Given that the numbers are actual counts of Mammoth fauna bones, chi-squared distances were used in the analysis; clustering method was “average.” The statistical significance of the clusters recovered by the analysis was assessed using a bootstrapping approach following Suzuki and Shimodeira (2006). The analysis was performed using R’s Pvclust Package (Suzuki et al., 2019; R Core Team, 2021).
While collecting bones, each sample was marked immediately upon discovery and the location of the find was mapped (Kuznetsova, 2008). Group A bones were obtained in situ, i.e., within the frozen sediment, so that the location and vertical position in the section are known. Group A contains the smallest number of bone specimens due to the specifics of collecting paleontological material from ice-rich permafrost deposits that thaw intensively during the summer season, and in which freshly exposed boness are rather quickly relocated and buried in slump debris. Group B bones were obtained from thermo-terraces of thaw slumps, having been previously thawed and relocated, but were found in positions that indicated roughly which part of the bluff above they had come from. Therefore, the upper and lower boundaries of their possible occurrence in the section could be determined, and aligned to a specific cryostratigraphic unit. Group C bones also occurred within the thawed debris at the base of the exposure, but in locations that provided no indication of their original positions. Group D bones, which were found on the beach, represent the most abundant category. This group is further divided into three subgroups: those from the west part of the coast, those from the east part of the coast, and those from the mouth of the stream. Group E bones were found on the surface of the tundra in the hinterland. specimens of the A and B groups have direct stratigraphic importance for the interpretation of the permafrost sequences. For in situ group A specimens, the stratigraphic position is known, or the minimum height in section can at least be estimated. For group B specimens, approximate upper and lower bounds can be placed on their original stratigraphic positions.
To document the sites of finds on the beach with greater precision, the coast at Kondrat’eva Yedoma west was divided during field work into three parts (Figure 3). Section 1 was the beach under the Alas basin lying between 7.2 and 8.0 km from the mouth of the Kondrat’eva River (4,700–5,500 m of the Kondrat’eva Yedoma profile); Section 2 spanned the beach below the cliff of the high Yedoma upland, between 6 and 7.2 km from the river mouth (3,500–4,700 m of the Kondrat’eva Yedoma profile); and Section 3 spanned the beach between 4 and 6 km from the river mouth (1,500—3,500 m of the Kondrat’eva Yedoma profile), which is the coast of a less elevated part of the Yedoma upland. However, when the sea is rough, bones can be transported along the beach for considerable distances, as we observed during fieldwork. Therefore, the division into Sections 1–3 was not strict, and was used only with reference to the numerous bones found on the east part of the coast. All bones from the western shore (west from the Camp stream, area is not shown in Figure 3) are considered to belong to one group.
In total, 21 bone fragments representing mammoth, horse, bison, and musk oxen were selected for radiocarbon dating, which was done either by acceleration mass spectrometry (AMS) or conventional 14C age determination. The latter required sample weights of about 1 kg. Dating was performed at the Laboratory of Isotope Geochemistry and Geochronology of the Geological Institute of the Russian Academy of Sciences Moscow, Russia (GIN), at the Leibniz Laboratory for Radiometric Dating and Stable Isotope Research, University of Kiel, Germany (KIA), and at the Center of Isotope Research, University of Groningen, the Netherlands (GrA). Dating was initially possible only using the conventional radiocarbon method, in the laboratory of the Geological Institute RAS in Moscow. Large, well-preserved samples were selected for this. After fieldwork in 2002 a single in situ sample from the outcrop was dated, as were four large mammoth and horse samples from the thermo-terrace and six samples from the shore. The largest musk ox bone collected on the coast was also dated, because this species is rare. After fieldwork in 2007, five horse bones were given to the same laboratory, two from the thermo-terrace and three from the shore. Four samples (three mammoth and one musk ox) were dated by the AMS radiocarbon method due to the small size of the bone fragment (Table 2). Ages are reported as uncalibrated years before present (a BP).
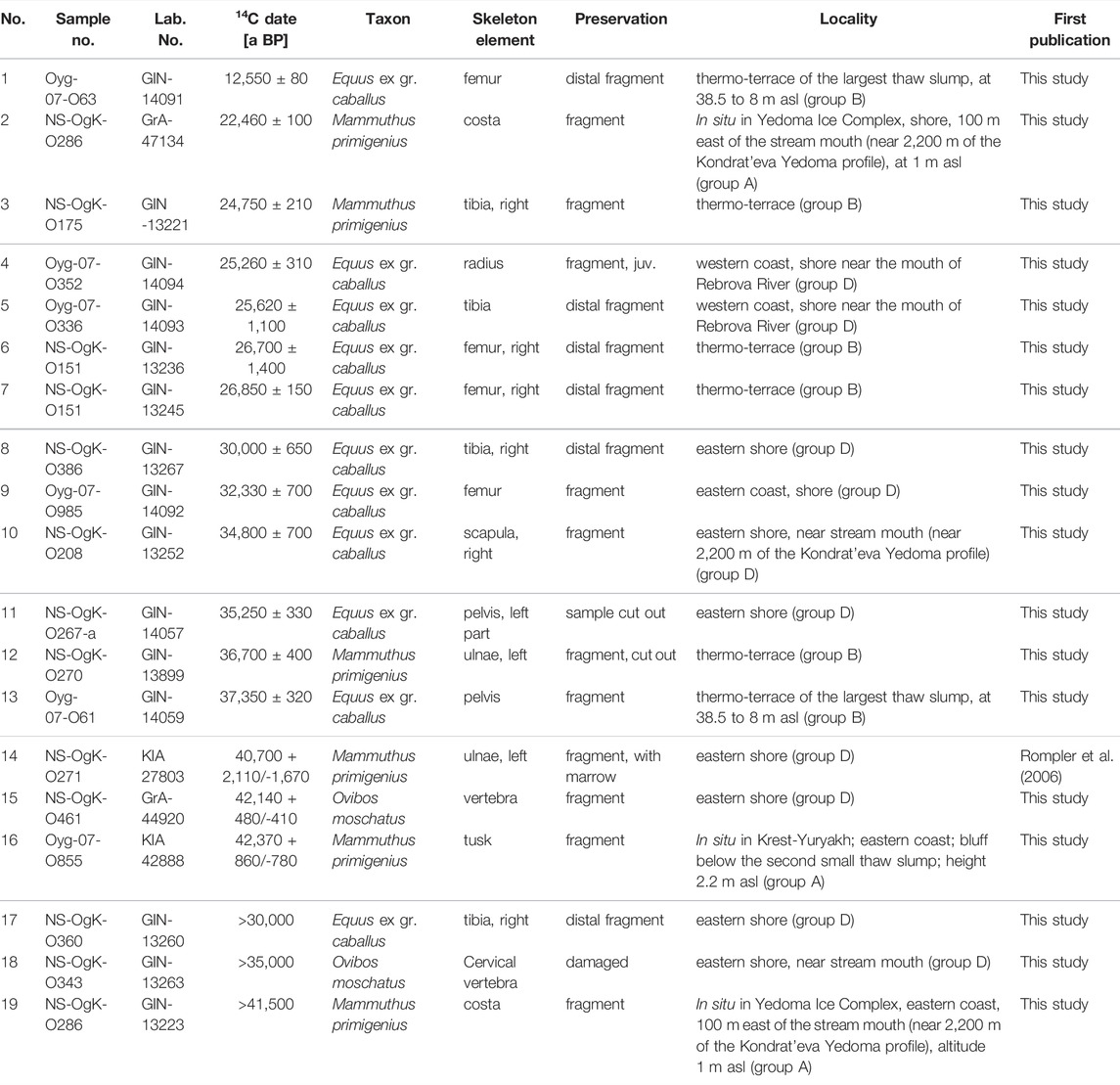
TABLE 2. List of radiocarbon dates derived from collagen of Mammoth fauna bones collected at Oyogos Yar shown in order of increasing age. GIN - Geological Institute of the Russian Academy of Sciences, Moscow (Russia); GrA—Center of Isotope Research University of Groningen (Netherlands); KIA—Leibniz Laboratory for Radiometric Dating and Stable Isotope Research University of Kiel (Germany). Samples 2 and 19 were taken from the same costa of Mammuthus primigenius.
From the entire mammal bone collection sampled in 2002 and 2007 (1977 samples in total), 1925 bones were identified and analyzed (Figure 4). Most of the collection was systematically obtained in August 2007 near the Kondrat’eva Yedoma and adjacent alases. Over 4 weeks of fieldwork it was possible to study the host deposits, and to carefully differentiate the sites where bones were found. In contrast, the samples from 2002 derived from a limited 1 day reconnaissance; consequently, the 2002 and 2007 collections are considered separately. The 2007 samples include more in situ finds and more small bones and small fragments of large bones, which commonly require more time and effort to locate (Figure 5). The 2002 samples, by contrast, include more large bones and more finds from the beach (Figure 6).
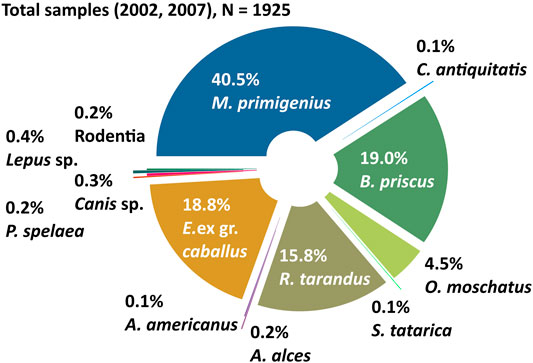
FIGURE 4. Composition of the mammal bone collections from Oyogos Yar showing the total collection from 2002 to 2007 (N = 1925). The percentages are calculated from the total number of identified samples (Table 1).
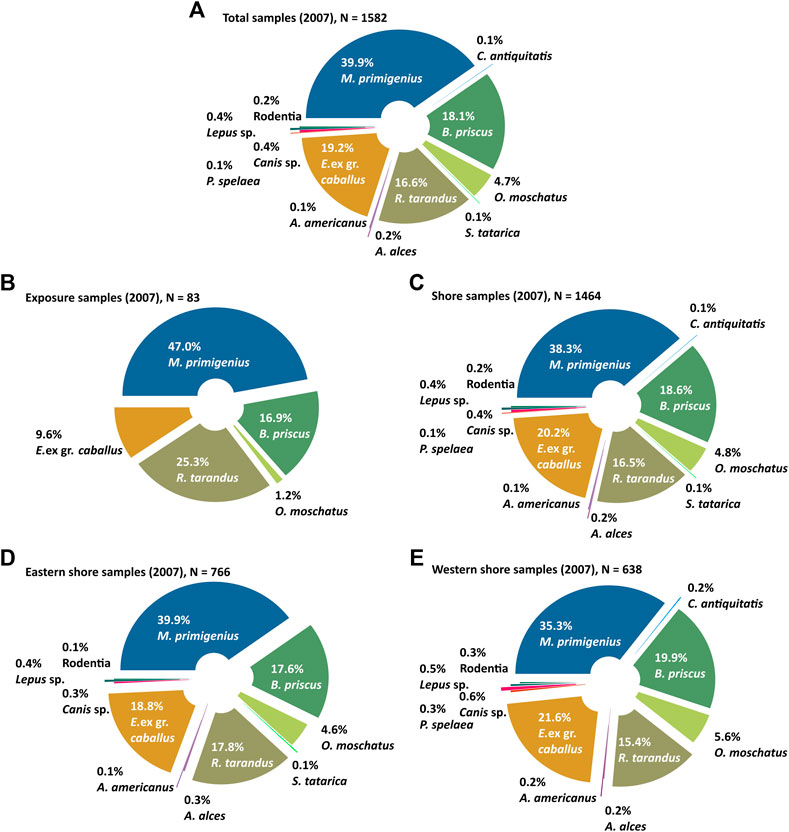
FIGURE 5. Composition of the mammal bone collections from Oyogos Yar showing (A) the total collection from 2007 (N = 1582) differentiated into (B) the exposure collection (N = 83) and (C) the shore collection (N = 1464), which is further differentiated into (D) the eastern shore collection (N = 766) and (E) the western shore collection (N = 638). The percentages are calculated from the total number of identified samples (Table 1). Note, that 35 bones found at other places and 61 bones found at other shore locations than (D) or (E) are summarized in (A), but not differentiated in this figure. The full record is given in Table 1.
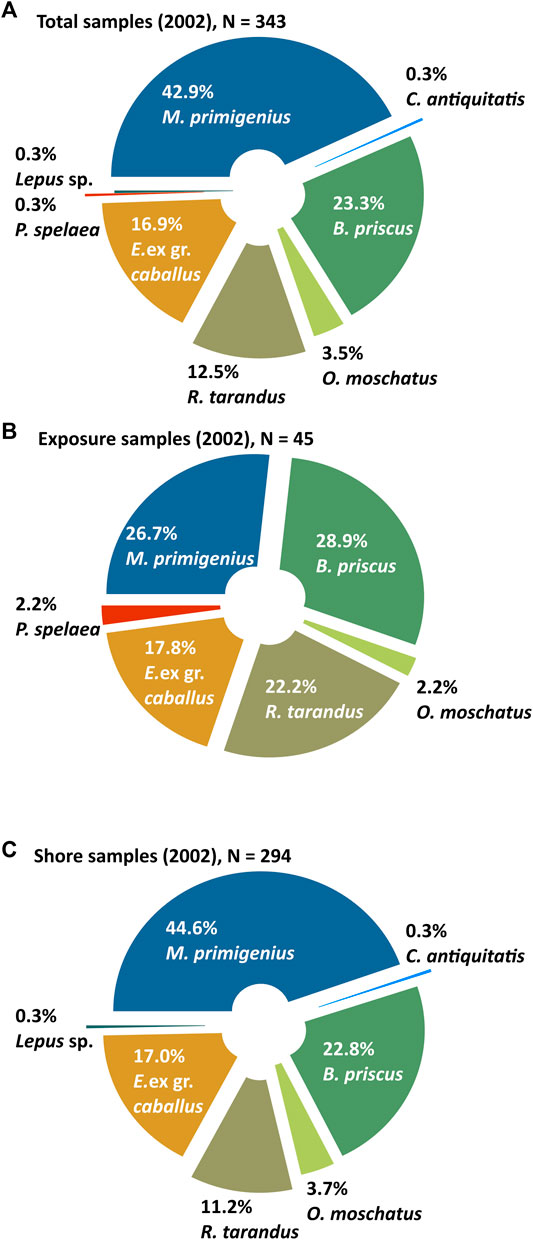
FIGURE 6. Composition of the mammal bone collections from Oyogos Yar showing (A) the total collection from 2002 (N = 343) differentiated into (B) the exposure collection (N = 45) and (C) the shore collection (N = 294). The percentages are calculated from the total number of identified samples (Table 1).
In total, the collection from 2007 comprises 1,608 bones and fragments, of which 21 could not be identified and five belong to birds or to arctic ringed seals. Only the 1582 identified terrestrial mammal bones (Figure 5A) are considered further. Among this total, woolly mammoth [Mammuthus primigenius (Blumenbach, 1799)] bones and fragments prevail (39.9%), while horse [Equus ex gr. caballus L., 1758] (19.2%), Pleistocene bison [Bison priscus (Bojanus, 1827)] (18.1%), reindeer [Rangifer tarandus L., 1758] (16.6%), and musk ox [Ovibos moschatus (Zimmermann, 1780)] (4.7%) make up lesser shares of the collection. Bones of other mammals such as woolly rhinoceros [Coeleodonta antiquitatis (Blumenbach, 1790)], elk [Alces alces L., 1758], moose [A. americanus (Clinton, 1822)], saiga antelope [Saiga tatarica L., 1758], rodents [Rodentia], lagomorphs [Lepus sp.], and predators such as wolves [Canis sp.] and cave lions [Panthera spelaea (Goldfuss, 1810)], are rare and each represent far less than 1% of the collection (Figure 5A; Table 1).
The 2007 bone collection is differentiated into exposure samples, including groups A, B, and C (Figure 5B) and shore samples corresponding to group D (Figures 5C–E). A few samples were collected on the surface of the tundra (group E), and at other outcrops far west of Kondrat’eva Yedoma. Samples collected from the outcrops (exposure samples defined as groups A, B, and C) sum up to 83 identified fossil bones, representing only about 5.2% of the entire collection. Such a proportion is typical for Yedoma Ice Complex localities. From the Kondrat’eva exposures, remains of woolly mammoth predominate (47.0%), followed by reindeer (25.3%), bison (16.9%), and horse (9.6%) (Figure 5B).
Only six bones were found in situ in the exposure (group A, Figure 3). In total, 78 samples were collected from the thermo-terraces (group B) of Kondrat’eva Yedoma, and 2 samples (bison and reindeer) were found within the thawed debris at the base of the exposure (group C). The Group A specimens include one damaged radius of a bison, and five fragments of woolly mammoth tusks. The in situ findings were confined to the sediments of the Yedoma Ice Complex and to taberal Kuchchugui, Krest-Yuryakh, and Alas deposits (Figure 3).
The samples collected directly from the thermo-terrace (group B), were found mainly at altitudes from 8 to 16 m asl. Of the 78 fossil bones or fragments, 75 have been identified, while three remain unidentified. Remains of M. primigenius (45.3%) predominate, followed by R. tarandus (26.7%), B. priscus (16.0%), and E. ex gr. caballus (10.7%), related to the identified 75 bones.
The majority of the bone material considered in this study, totaling 1464 identified samples, was obtained from the shore and constitutes group D. There are 1404 samples from the eastern and western shores (Figure 5C), and 60 bones collected from the shore in remote areas (“other shore” in Table 1; not shown in Figure 5).
From the beach at the Kondrat’eva Yedoma cliff and the eastern part of the alas (eastern shore, Table 1), 766 samples were collected and identified (Figure 5D). Woolly mammoth predominates (39.9%), followed by horse (18.8%), reindeer (17.8%), bison (17.6%), and musk ox (4.6%) (Figure 5D). Among rare species, saiga antelope were represented by a distal fragment of a humerus, elk by thoracic and lumbar vertebrae and a second phalanx, wolves by a pelvic fragment and second upper molar (identified by M.V. Sotnikova, Geological Institute RAS Moscow, Russia), hares by skull, pelvic, and femoral fragments, and lemmings by a skull (Dicrostonyx sp., determined by A.S. Tesakov, Geological Institute RAS Moscow, Russia).
West of the Camp stream at the beach below the alas outcrop (western shore, Table 1; not shown in Figure 3), 646 samples were collected, of which 638 were identified (Figure 5E). Here, woolly mammoth (35.3%), horse (21.6%), bison (19.9%), reindeer (15.4%), and musk ox (5.6%) were the most common species. Woolly rhinoceros were represented by the proximal fragment of the third metacarpal bone (Mc III), cave lions by a tooth and epistrophy of, Canis lupus by three teeth and a femur fragment (identified by M.V. Sotnikova), hares by three limb bones, lemmings by a pelvis and a right mandibular ramus containing two teeth (Dicrostonyx cf. torquatus, identified by A.S. Tesakov), and Alces sp. by a distal femur fragment, a damaged lumbar vertebra, and a fragment of a thoracic vertebra. Moose were represented by a right lower third molar (M3) and a second phalanx, identified as A. americanus (identified by P.A. Nikolskiy, Geological Institute RAS Moscow, Russia).
With respect to species that were part of the Mammoth fauna and still inhabit the territory, it is difficult to distinguish between fossil and modern material collected on the shore, which are often in a similar state of preservation. Therefore, it is impossible to say for sure whether fossil or modern remains of reindeer, wolf, and hare were found. The last of these, at least, is present in the fossil bone collection from Bykovsky Peninsula, where fossil hare remains were collected in situ (Kuznetsova et al., 2019).
In terms of bone fragmentation and quality of bone preservation, specimens from the western shore (Figure 5E) differ from those from the eastern part (Figure 5D). On the western shore, there were fewer unbroken large limb bones, but many small limb bones, small fragments of various bones, and small fragments of mammoth tusks. This indicates that the bones were repeatedly redeposited from older to younger deposits and sorted by waves on the beach. This hypothesis is also confirmed by the collection of samples from a small part of the western coast, which we identified as “coast with small bones” (2.6–3.1 km west of the Camp stream). This area was almost completely covered with small tusk and bone fragments, and small bones. On this small part of the beach, 8.4% of the total western coast samples were collected; about 80% of the samples from this area are small fragments of mammoth tusks.
Interesting paleontological material includes a slightly damaged skull of a male musk ox with the lower jaw, and a fragment of a humerus with soft tissues and the ulna (Tumskoy and Dobrynin, 2008). Both were found at the mouth of a stream located 2.3 km southeast of the mouth of the Krest-Yuryakh River, in slumping debris at an altitude of 6–7 m asl. This fragment of the musk ox skeleton probably thawed out of Bychchagy Ice Complex sediments. We can assume that these are fragments of the skeleton of one individual, because they were found close to one another and in an identical state of preservation.
A fragmentary woolly mammoth skeleton was found on the surface of the tundra near a stream at a distance of 4.7 km from the mouth of the Kondrat’eva River (at 2,200 m along the Kondrat’eva Yedoma profile) (Figure 3), at an altitude of 12 m asl. This skeleton thawed in a thermokarst mound of the Yedoma Ice Complex and was first found in 1994 by local people (personal communication). The skull was broken into many small pieces, probably when tusks and teeth were knocked out of it. We collected 32 pieces of this skeleton: a tooth fragment, six fragments of the cervical vertebrae, four fragments of the thoracic vertebrae, five fragments of the lumbar vertebrae, some ribs, a scapula, a fragment of the pelvis, a metapodium, and an os carpale.
We conducted a reconnaissance trip to the Kondrat’eva Yedoma on 30 August 2002 (Schirrmeister et al., 2003a) and collected 369 bones and fragments, of which 343 were identified (Table 1; Figure 6A). We consider the 2002 collection separately as it was collected in 1 day, which is reflected in an even greater predominance of bones collected on the shore over material collected at the outcrop. The 2002 collection contains many large limb bones and few small bone fragments and small mammal bones and teeth. The 2002 landing site was located at the mouth of a creek situated 4.7 km west of the Kondrat’eva River mouth, at a horizontal distance of 2,200 m on the Kondrat’eva Yedoma profile shown in Figure 3. As in the 2007 collection, mammoth (42.9%), bison (23.3%), horse (16.9%), and reindeer (12.5%) bones predominate, while musk ox remains (3.5%) make up a smaller share (Figure 6A).
A total of 48 specimens (45 identified) representing groups A and B were collected from the exposure and the thermo-terrace (Figure 6B), but only one bone (a large fragment of a mammoth rib) was found in situ. Next to this rib in the coastal outcrop, at a height of 1 m asl, several more ribs were observed, but they could not be collected due to inaccessibility. The ribs protruded from the roof of a wave-cut notch right above the beach. Most likely a part of this skeleton, or the complete mammoth skeleton, which had been buried in the exposed Yedoma Ice Complex was destroyed. It is possible that the bones found directly next to the described site of the possible burial of this mammoth skeleton, including a pelvis (right and left halves), a damaged left femur, and a fragment of the left humerus, were part of the destroyed skeleton. The bones belong to a young individual and bone marrow is preserved in the limb bones. The rib, sampled in situ, was dated twice (Table 2) to >41,500 a BP (GIN-13223) and 22,460 ± 100 a BP (GrA-47134). It is not clear how such considerable age differences between samples from one bone can be explained. A fragment of the left half of the pelvis was also dated, to 35,250 ± 330 a BP (GIN-14057).
Forty-seven specimens were collected from the thermo-terrace, including three indeterminate bone fragments. Among the 44 identified specimens found on the thermo-terrace (Figure 6B), bison bones (28.9%) predominate slightly over mammoth bones (26.7%), whereas reindeer bones make up 22.2% of the total, and horses 17.8%. The 44 specimens also include a single musk ox horn sheath, and a single fragment of a cave lion femur.
Some 317 specimens collected from the shore represent 86% of the entire 2002 collection, but 23 could not be identified due to poor preservation. The species present among the 294 identified bone remains include woolly mammoth (44.6%), bison (22.8%), horse (17.0%), reindeer (11.2%), and musk ox (3.7%) (Figure 6C). In addition to bones of these five most common Mammoth fauna species, a damaged cervical vertebra of woolly rhinoceros and a tibial fragment of hare were found. The entire 2002 collection differentiates into 245 samples from the shore below the highest outcrop of the Kondrat’eva Yedoma (between 2,200 m and 5,500 m on the Kondrat’eva Yedoma profile shown in Figure 3) and 68 samples from the shore at the mouth of the creek at the landing site (at 2,200 m on the Kondrat’eva Yedoma profile shown in Figure 3). The last group of samples includes four bones, which probably belong to the mammoth skeleton described above and were collected at a distance of 150 m east of the landing site. No specimens were collected to the east of the landing site due to lack of time. Two mammoth partial limb bones (ulnare and radius fragments), and two tusk fragments, were found on the tundra surface (group E) near the landing site.
Permafrost is very favorable for the preservation of bone material, but the bodies of dead animals in permafrost pass through different taphonomic filters, e.g., animal feeding, frost weathering, chemical alteration, and abrasion by running water (Sher et al., 2005). A single bone indicates the prior existence of not only hundreds of missing bones of the same animal, but perhaps thousands of non-preserved bones of other animals. A further argument for high abundance of animals in the Mammoth fauna is that mammoth, horse, bison, and reindeer are all herd species; they never live alone. Therefore, the large quantities of bones clearly indicate that a relatively large number of these grazing animals existed in the past, and suggest the existence of appropriate pastures and a tolerable climate.
The total collection of paleontological material can be divided into two unequal parts according to quality of preservation. The smaller part consists of the few samples that provide a basis for partially reconstructing the conditions of their burial, which in turn must have influenced the process of fossilization (taphonomy). The larger part consists of bones, in varying states of preservation, that provide no evidence from which the conditions of their burial might be reconstructed. In some cases, mode of preservation makes it possible to partly reconstruct the depositional or redepositional processes to which a given sample was subjected. Furthermore, some bones show signs of having been gnawed by predators. Large limb bones retain bone marrow preserved inside.
The first way bones can be preserved in permafrost is by being covered in concretions of hydrated iron phosphate (vivianite), indicating diagenetic mineralization and waterlogged storage conditions. The presence of vivianite on bones indicates the conditions under which they were buried, or potentially redeposited. Bones covered in vivianite were buried either in swampy conditions or in deposits with high ice content, which melted and re-froze to form taberal deposits. In either case, the bones were preserved under relatively anoxic conditions and the decomposition of organic matter led to the formation of authigenic aggregates of phosphates, present assoil forms of vivianite (Rothe et al., 2016). Most of the bones in the collection that are covered in vivianite belong to mammoths (16 samples). Far fewer represent other animals: bison (five), horse (two), musk ox (one), and reindeer (one). Three out of five mammoth tusk fragments found in situ exhibit vivianite concretions. These tusks were obtained from taberal Kuchchugui deposits, from Krest-Yuryakh deposits in the eastern part of the section (Figure 3), and from taberal Yedoma Ice Complex deposits in the western part of the section. Most of the bones and bone fragments covered in vivianite were collected from the western and eastern parts of the coast. The preservation of vivianite, however, indicates that the bone material was not transported over a large distance, as such transport would have destroyed the crystal concretions.
The second way bones can be preserved is by being covered with iron oxides. A total of ten specimens covered with amorphous iron oxides were collected: three mammoth samples (tusk, tooth, and rib fragments), four bison samples (femur and humerus fragments, astragalus, tooth), and three reindeer bones (damaged femur and os centrotarsale, distal fragment of metatarsale). Three of these samples were found on the outcrop, and the other seven on the east shore. The three skeletal elements found on the outcrop include the tusk fragment, which was discovered in situ, as well as the mammoth rib fragment and damaged reindeer femur. The rib fragment is from the first rib, and was found on a thermo-terrace beneath the Yedoma Ice Complex at a height of 10 m in the eastern part of the section, whereas the damaged femur was collected on a 6–8 m high thermo-terrace which was also located beneath the Yedoma Ice Complex. The presence of amorphous iron oxides on the bones indicates a long residence time in well-aerated low-moisture sediments, or on the surface of the ground, and limited relocation.
Three samples are coated with both vivianite and iron oxides. These are fragments of the tusk (in situ) from the taberal Kuchchugui deposits, and fragments of mammoth tooth and bison femur from the beach. The presence of both vivianite and iron oxides indicates that a given bone was alternately under conditions conducive to the formation of vivianite and conditions conducive to the formation of iron oxides. For example, a mammoth tusk fragment was syncryogenetically buried in Kuchchugui deposits and covered in oxides. Subsequently, the syncryogenic Kuchuguy sediments were thawed, so that the tusk fragment was then subjected to conditions of excess water and lack of oxygen, and became overgrown with vivianite.
The third state in which bones can be found is showing evidence of having been gnawed by predators; samples showing evidence of such damage include fragments of large limb bones, a pelvic fragment, heel bones, and astragali. In our opinion, the bones were gnawed before they were buried, but unequivocal confirmation of this interpretation will require examination of the tooth marks left by the predators. Bones in this condition include four mammoth bones, three horse bones, and one bone each from musk ox and bison. The gnawed mammoth and horse bones were collected from both the western and eastern parts of the shore; the single musk ox and bison bones were collected from one part of the shore only. As mentioned earlier, the musk ox specimen, a distal fragment of the humerus with associated soft tissue, was found in slumped debris on the outcrop located 2.3 km southeast of the mouth of the Krest-Yuryakh River.
Eight fragments of large limb bones are probably the best preserved because they contain bone marrow; these represent the fourth preservation type. The presence of bone marrow is an indicator of rapid bone burial. Four mammoth bone fragments, two musk ox bone fragments, and one each from bison and reindeer, all containing marrow, were found. All were found on the shore. It is possible that a much greater number of bones contain bone marrow; many of the whole large limb bones of horses, bison, reindeer, and musk ox probably contain bone marrow inside, but we did not carry out any special research in this direction.
There are also two interesting samples in the collection. One is a fragment of horse humerus which is gnawed by predators and covered with vivianite. This combination suggests that the bone most likely lay on the surface for some time, and then was redeposited in marshy conditions or it was buried in Ice Complex deposits which melted and re-froze to form taberal deposits. Another single specimen is a fragment of a bison femur with bone marrow that is covered with vivianite. This combination indicates that the bone was probably quickly buried in swampy conditions, The largest number of bones and their fragments were well preserved but show different stages of alteration. In this group we noted bones with varying degrees of roundness indicating transportation. A total of 135 rounded bones and fragments were identified, which is 7.0% of the entire collection (7.9% of the 2007 collection) They are visually differentiated into weakly-rounded, medium-rounded, and strongly-rounded bones, and bones worn by sea-ice. Weakly-rounded bones include carpal and tarsal bones and fragments of the second phalanges of horses, bison, and reindeer totaling 13 samples collected on the beach west of the Camp stream. The medium-rounded bones comprise 107 samples and consist of bones and fragments from 39 horse specimens, 33 bison specimens, 23 reindeer specimens, six mammoth bone fragments, five musk ox bone fragments, and one wolf bone fragment. The small number of rounded mammoth bones, which otherwise dominate the shore collection, indicates a significant resistance of mammoth bones to mechanical abrasion and rounding. Among the rounded bones, tarsal and carpal bones predominate, due to their shape. Sesamoid bones, phalanges, and fragments of large limb bones are present significantly less frequently. Bones of medium roundness were collected both on the eastern and the western parts of the shore. The strongly-rounded bones include nine samples. Five bone fragments belong to bison limb bones and two to horse limb bones; one bone each belongs to mammoth and reindeer. The rounded bones also include bones worn by sea-ice. There are only six such bones: three horse bones (two astrogalus and a tibia fragment), a bison carpal bone, a musk ox astrogalus, and a reindeer astrogalus. This form of preservation is characterized by a completely flat surface on one side of the bones; all morphological structures of the bone (outgrowths, depressions, grooves, broadness, etc.) have been erased. This level surface could have been formed by ice rubbing against the bone while the bone was frozen in beach sediments. A strong degree of such wear is very clearly visible on the bones. Such bones are found on both sides of the coast. The small number of samples preserved in this state does not mean that the other bones, especially the rounded and strongly-rounded ones, did not experience ice friction, but ice rubbing against bones frozen into the sediments of the beach can be clearly established for a few bones only.
Unfortunately, radiocarbon dates of bones from the Oyogos Yar coast, in contrast to bone dates from the Bykovsky Peninsula, the New Siberian Islands, and the Lena River Delta, are scarce. We submitted 21 samples from our collection for dating; 19 dates have been received (Table 2). In a fragment of a left mammoth humerus and in a fragment of a left mammoth ulna, collected on the shore at a relative distance of 2,350 m and between 2,200 and 5,500 m (Figure 3), collagen was absent. The humerus fragment was collected on the shore under the outcrop at a relative distance of 2,350 m (Figure 3), and the ulna fragment was collected on the shore (Figure 3). Of the 19 radiocarbon dates obtained, three are infinite. Two dates were determined from one sample, a Mammuthus primigenius rib. The youngest date is 12,550 ± 80 a BP; the oldest is 42,370 + 860/-780 a BP. Of the 18 bones examined, ten specimens are from horses, six specimens are from mammoths with two dates from one bone, and two specimens are from musk ox (Table 2). If we consider where samples chosen for dating were collected, two bones (which yielded three dates), a fragment of a rib and a fragment of a mammoth tusk, were collected in situ. Six samples from thermo-terraces were dated: two fragments of mammoth bones (ulnare, tibia) and four fragments of horse bones (three femur fragments and a pelvis fragment). Ten specimens were dated from the shore, including six horse bone fragments, two specimens belonging to mammoths, and two specimens from musk ox (Table 2). Specimens from the shore were collected near the mouth of the Rebrova River (horse radius and ulna fragments), from the shore below the Kondrat’eva Yedoma and an alas at a relative distance of 5,000 m shown in Figure 3 (fragments of two tibia and a femur fragment from a horse, a fragment of a mammoth pelvis, and a musk ox vertebra fragment), at the mouth of the stream at 2,200 m relative distance shown in Figure 3 (horse shovel fragment and damaged musk ox vertebra), and 150 m from the 2002 drop-off site (mammoth pelvis).
The fragment of a mammoth rib collected in situ has been dated twice. The time scatter turned out to be significant: >41,500 a BP (GIN-13223) and 22,460 ± 100 a BP (GrA-47134). We cannot explain this significant time discrepancy, because the samples given for dating were both from one bone. The left branch of pelvis that probably belongs to the same mammoth individual as the fragment of rib was dated to 35,250 ± 330 a BP (GIN-14057). The AMS date of 42,370 + 860/-780 a BP (KIA-42888) obtained from the tusk which was found in situ is also unexpected. It does not correspond to the stratigraphic position of the host sediments as it is too young. We attribute this discrepancy to the contamination of the sample with modern organic matter.
We cannot clearly confirm the regularity of the distribution of new radiocarbon dates and radiocarbon dates from the Oyogos Yar published earlier (Rompler et al., 2006; Nikolskiy et al., 2011; Boeskorov et al., 2013). The total of 38 radiocarbon final dates is a small database. Most of the dates fall in the time interval from 42.5 to 22.5 ka BP, of which seven dates in the period from 27.5 to 25 ka BP. In the period from 40 to 37.5 ka BP dates are absent (Figure 7).
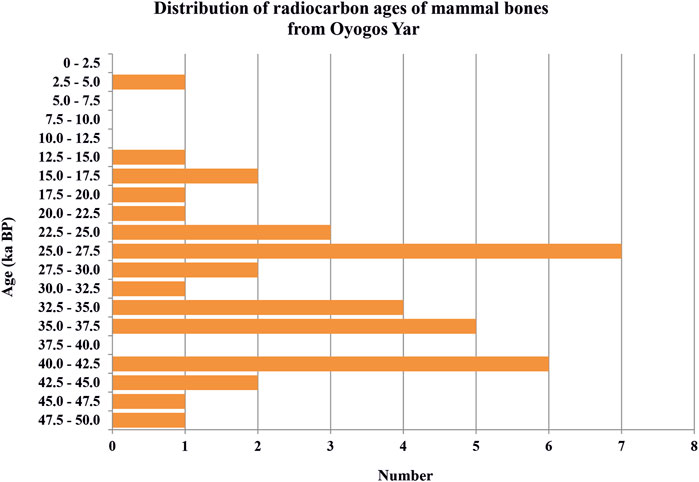
FIGURE 7. Distribution of radiocarbon ages of mammal bones (38) from the Oyogos Yar coast including data from Nikolskiy et al., 2011 (N = 20), Boeskorov et al., 2013 (N = 2), Rompler et al., 2006 (N = 1), and this study (N = 15). Only finite dates are given as uncalibrated ages.
In general, the ratio of bone remains from the main large grazer species of the Mammoth fauna that we found is close to the known percentage of animals in the past fauna. However, the predominance of woolly mammoth remains in the collection can be explained not only by a significant mammoth predominance in the fauna, but also due to the better preservation of large and thick mammoth bones. The large percentage of reindeer bones may be due to bones of modern reindeer being included in the collection. Reindeer is the only large mammalian species of Mammoth fauna that still inhabits the described territory at the present time.
The bones of small adult mammoths (vertebral, carpal, and tarsal bones and others) were collected from both the eastern and western parts of the Oyogos Yar coast. These bones are much smaller in size than similar bones of other mammoths of the same age. Single finds of bones of small adult mammoths were previously described from Bol’shoy Lyakhovsky Island, Bykovsky Peninsula, and the interfluve of the Olenek and Anabar rivers (Kuzmina et al., 1999; Kuznetsova and Kuzmina, 2000; Sher et al., 2000; Kuznetsova et al., 2004). These findings make it possible to put forward a hypothesis about the co-existence of larger and smaller woolly mammoths during some time intervals (Nikolaev et al., 2011) which was further confirmed by Boeskorov et al. (2017) and the present study.
Age information from mammoth bones of group A and group B was obtained from nine samples (Table 2; Figure 3), which range from >41.5 to 12.5 ka BP. This aligns the MIS 3 to 2 period of Yedoma Ice Complex formation with the presence of Mammoth fauna, although in situ findings in deposits predating the Yedoma Ice Complex complicate the picture. For example, the age of a mammoth tusk dated to 42.37 ka BP (Figure 3; KIA 42888) and the age of wood fragments dated to 47.7 ka BP (KIA 25730; Opel et al., 2017a), both found in ice-wedge casts attributed to the MIS 5 Krest-Yuryakh stratum, call into question the in situ preservation of the organic material or, in turn, the age of the Krest-Yuryakh stratum that has previously been IRSL-dated to 102.4 ± 9.7 kyr ka (Opel et al., 2017a). Here, permafrost degradation, material mobilization, and re-freezing might explain the discordance of older host deposits and the clearly younger bone ages. If this explanation is correct, the present Krest-Yuryakh host deposits must have thawed to incorporate re-located bone material from stratigraphically younger strata, and subsequently have re-frozen in situ. This would further imply a warm period sometime between MIS 5 (Krest-Yuryakh) and MIS 3 (Yedoma) or during MIS 3. The regional MIS climate optimum as recorded on Bol’shoy Lyakhovsky Island took place between 48 and 38 ka BP (Andreev et al., 2009; Wetterich et al., 2014) while, in the broader regional context, warmer-than-today summers have been reconstructed on Bykovsky Peninsula at about 48 and 35 ka BP based on pollen, plant macrofossils, and insects (Schirrmeister et al., 2002b; Kienast et al., 2005) and in the Lena Delta at about 48 ka BP, 43.5 to 41 ka BP, and 36 ka BP based on chironomids (Wetterich et al., 2021b). If such a warming episode was able to melt surface wedge ice and create initial thermokarst with a high-center polygonal surface in places, the finding of bones and other organic material post-dating the Krest-Yuryakh host deposits could make sense. However, the intact, thus undisturbed, sedimentary structures of lacustrine laminated Krest-Yuryakh ice-wedge casts call this interpretation into question. The fact that some bones, including this tusk, were for a long time in thawed sediments without access to oxygen under the water of a lake is also indicated by the vivianite covering them.
The 28 radiocarbon dates are known from the Kondrat’eva Yedoma outcrop and the coast below it. Seventeen dates were obtained from bones from this collection (Table 2) and 11 dates were published earlier (Nikolskiy et al., 2011). The date range is from >50.0 to 12.5 ka BP. Of these, eight (28.6%) are infinite dates, which may indicate a significant amount of bone remains from deposits older than 47.4 thousand years and a wide distribution of these deposits. Many infinite dates (more than 42%) were also obtained from the bones from the Zimov’e outcrop of the Bolshoy Lyakhovsky Island (the northern coast of the Dmitry Laptev Strait). This also seems to be due to the good exposure of ancient Ice Complexes (Tumskoy, 2012).
The range of finite mammal bone ages at the Kondrat’eva Yedoma exposure covers the MIS 3 to 2 period of Yedoma Ice Complex formation almost completely (Figure 8), while host deposits are radiocarbon-dated from 48.5 to 32.2 ka BP (Schirrmeister et al., 2011). The 12 radiocarbon dates belong to the period of Kondrat’eva Yedoma Ice Complex deposition. A fragment of a mammoth ulna with bone marrow dated to 40.7 + 2.1/−1.7 ka BP indicates very rapid sedimentation of some layers of the Ice Complex (Table 2). It should be noted that eight dates are younger than 32.2 ka BP, which is the upper age limit obtained from Yedoma Ice Complex deposits (Figure 8). The MIS 2 Yedoma Ice Complex, including the sedimentary legacy of the Last Glacial Maximum, has not yet been identified on the Oyogos Yar coast. We noted a similar situation when studying the southern coast of B.L. Island and explained it by fragmentation of the MIS 2 aged deposits (Andreev et al., 2009). Further research on the Zimov’e section on Bol’shoy Lyakhovsky Island indicated MIS 2 deposits dated from 29.3 to 21.7 ka BP (Wetterich et al., 2011; Wetterich et al., 2021a). Apparently on the Kondrat’eva Yedoma outcrop deposits younger than 32.2 ka BP were fragmented and melted.
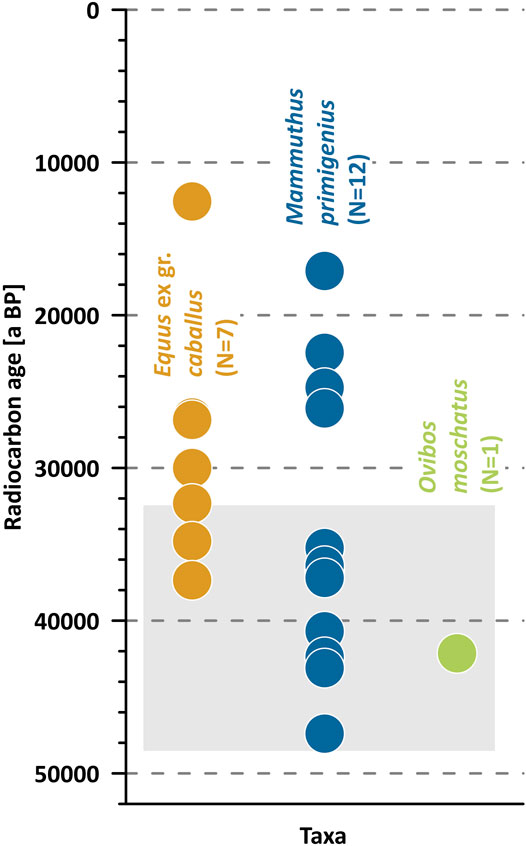
FIGURE 8. Distribution of radiocarbon ages of mammal bones differentiated by taxa from the Kondrat’eva Yedoma exposure including data from Nikolskiy et al., 2011 (N = 6), Rompler et al., 2006 (N = 1), and this study (N = 13). Only finite dates are given as uncalibrated ages. The gray rectangle highlighted the age range of Kondrat’eva Yedoma Ice Complex host deposits, which was radiocarbon-dated from 48.5 to 32.2 ka a BP (Schirrmeister et al., 2011)
One bone was found on the thermo-terrace; it was a distal fragment of horse femur, dated 12.55 ± 0.08 ka BP. It was buried in deposits younger than the Yedoma Ice Complex, but older than the Holocene deposits. Such deposits have not been found either on the Kondrat’eva Yedoma or on the Zimov’e outcrop, although there are bones of this age from both outcrops (Andreev et al., 2009). Thus, dating of bone material often gives dates different from those of the sediments indicating that sediments that were deposited on these outcrops are currently eroded, extending the geological history of the area.
The distribution of the radiocarbon dates of the present study and of studies published earlier (Rompler et al., 2006; Nikolskiy et al., 2011; Boeskorov et al., 2013) from Oyogos Yar coast, a total 64 dates from >51.0 to 4.63 ka BP, is available. The 38 finite dates from 48.8 to 12.55 ka BP and the Holocene date (4.63 ka BP) have been analyzed (Figure 7). The largest number of dates refers to the time interval from 45 to 22.5 ka BP (MIS 3-2 Yedoma Ice Complex) with three time-peaks: 45–40, 37.5–32.5, and 30–22.5 ka BP. The age of the most famous find from Oyogos Yar, the Yuka mammoth (34.3 ka BP; Boeskorov et al., 2013; Lopatin, 2021), also fits well into the upper part of the MIS 3 Yedoma Ice Complex of Oyogos Yar. The ages of E. ex gr. caballus bones in the present study are restricted to the MIS 3-2 period and range from 37.4 to 12.55 ka BP. The discovery of the Yukagir horse (Equus sp.) dated to 4.63 ka BP (Boeskorov et al., 2013) confirms a horse presence in the Holocene on Arctic coasts from the Taimyr Peninsula to Oyogos Yar and maybe even further east, as well as on the New Siberian Islands (Lazarev, 1980; Kuznetsova et al., 2001; Kuznetsova and van der Plicht, 2009).
The observed pattern, covering the vast area of Beringia during the late Pleistocene with different local geologies and collection histories, cannot be explained by a random interaction of different local factors. We believe that it is related to a general influence of environmental changes that affected northeastern Siberia during this period.
Bone collections of a similar size as collections from the Oyogos Yar coast are known from Bol’shoy Lyakhovsky Island (Kuznetsova et al., 2015; Wetterich et al., 2011; Wetterich et al., 2021a) and the Bykovsky Peninsula (Sher et al., 2005; Kuznetsova et al., 2019). Other collections from Cape Mamontov Klyk (western Laptev Sea shore, Schirrmeister et al., 2008), from different locations in the Lena Delta (Schirrmeister et al., 2003a; Wetterich et al., 2008; Wetterich et al., 2020; Wetterich et al., 2021b), from the Yana Lowland (Yana RHS site, e.g., Basilyan et al., 2011), the Indigirka Lowland (site, e.g., Pitulko, 2011; Pitulko et al., 2014), the Kolyma Lowland (e.g., Sher, 1971), and Wrangel Island (e.g., Vartanyan et al., 1993) complement the regional picture of the West Beringian mammoth fauna.
In detail, the bone collection from the southern coast of Bol’shoy Lyakhovsky Island comprises 1,026 bones, including one bird bone and 14 unidentified bones (Kuznetsova and Kuzmina, 2000; Kuznetsova, 2007). The resulting 1011 bones of the Mammoth fauna almost equally represent mammoth (257) and horse bones (253), as well as bison (200) and reindeer (185). Hare (24), cave lion (4), and woolly rhinoceros (6) are more abundant in this collection than in collections from Oyogos Yar and Bykovsky.
The bone collection obtained at different locations on Bykovsky Peninsula, but mainly at the Mamontova Khayata, comprises a total of 1,192 bones, including five bird bones and 71 unidentified bones (Kuznetsova et al., 2019). The resulting total of 1,116 identified mammal bones lacks findings of rare species found in the Oyogos Yar record such as woolly rhinoceros and saiga antelope.
Regional statistical analyses (Figure 9) were done for the Oyogos Yar collection in comparison to the Bol’shoy Lyakhovsky Island collection from 1999 (Kuznetsova and Kuzmina, 2000; Kuznetsova, 2007) and the Bykovsky Peninsula from 1998 (Kuznetsova et al., 2019). Consideration of the three collections shows similar proportions for mammoth at Oyogos Yar (39.1%) and Bykovsky Peninsula (38.0%), while Lyakhovsky has fewer mammoth bones (26.5%). The proportion of bison bones in the Oyogos Yar collection (19.5%) is similar to that at Bykovsky (20.8%). Reindeer bones are similar in abundance in all three collections (Oyogos Yar 16.3%, Bykovsky 14.4%, Lyakhovsky 18.5%). Horse bones are least frequent from Oyogos Yar (19.0%) while they are similar for Bykovsky (24.5%) and Lyakhovsky (23.3%). Musk ox finds differ by a few percent (Oyogos Yar 4.6%, Bykovsky 1.9%, Lyakhovsky 7.3%).
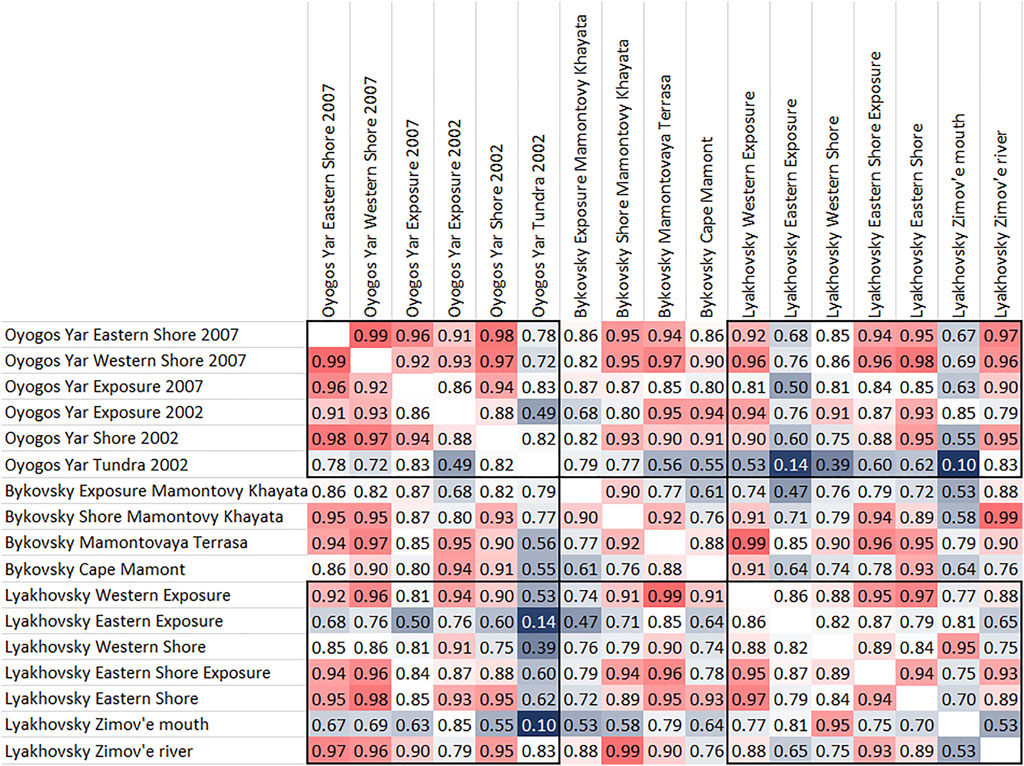
FIGURE 9. The Pearson correlation coefficients for the different sampling sites of the collections from Oyogos Yar 2007 + 2002, Bykovsky Peninsula 1998, and Bol’shoy Lyakhovsky Island 1999.
The two Oyogos Yar Shore 2007 sampling sites and the Oyogos Yar Exposure 2007 sample site have high correlation coefficients among each other, with highest correlation coefficients between the shore sites, where Oyogos Yar Western and Eastern Shore 2007 have a correlation coefficient of 0.99 (Figure 9). Similarly, high correlation is found between those two sampling sites and Oyogos Yar Shore 2002 (0.98 and 0.97).
The Bykovsky Peninsula sampling sites show weaker correlations among each other. The Bykovsky Exposure Mamontovy Khayata site and the Bykovsky Shore Mamontovy Khayata site show a correlation coefficient of 0.9; the Bykovsky Shore Mamontovy Khayata and the Bykovsky Mamontovaya Terrasa (alas) site show a correlation coefficient of 0.92 (Figure 8).
The samples from Bol’shoy Lyakhovsky Island show high similarity between the Eastern Exposure, the Eastern Shore Exposure, and the Eastern Shore sites (Figure 9). In addition, the Eastern Shore Exposure is similar to the Western Exposure site with a correlation coefficient of 0.95. The Western Shore site is similar to the Zimov’e mouth site, with a correlation coefficient of 0.95.
The Lyakhovsky Zimov’e mouth sample site is not highly correlated to any other site at any location. The Lyakhovsky Eastern Exposure also does not exhibit much similarity to any other sample site, like the Oyogos Yar Tundra 2002 sample, which is the most weakly correlated to all other samples, with correlation coefficients between 0.49 and 0.83. The Bykovsky Mamontovy Khayata Exposure is somewhat similar to the Bykovsky Mamontovy Khayata Shore site (correlation coefficient 0.90), but different from all other sampling sites. Bykovsky Cape Mamont is also somewhat different than all other sites; its highest similarity is to the Oyogos Yar Exposure site with a correlation coefficient of 0.94.
The Oyogos Yar Exposure 2002 sample site shows high correlation with Bykovsky Mamontovaya Terrasa and Bykovsky Cape Mamont (0.95 and 0.94, respectively). The samples from the Bykovsky Mamontovy Khayata Shore site also show high correlation coefficients with the Oyogos Yar Western and Eastern Shore 2007 sites (both 0.95) and the Oyogos Yar Shore 2002 site (0.93). The Lyakhovsky Eastern Shore site also shows high correlation coefficients with the Oyogos Yar Shore 2002 site and the Oyogos Yar Eastern and Western Shore 2007 sites, while the correlation of the Lyakhovsky Eastern Shore site with the Bykovsky Mamontovy Khayata Shore site is only 0.89 (Figure 9).
Contrary to the correlation analysis, similarity analysis via agglomerative hierarchical cluster analysis (Figure 10) groups the Lyakhovsky Western Shore together with the Lyakhovsky Zimov’e Mouth (high similarity, height 0.078) and the Lyakhovsky Eastern Exposure (lower similarity, cluster height 0.190) sites. Furthermore, high similarity is found between the Oyogos Yar Shore 2002, Oyogos Yar Eastern Shore 2007, and Oyogos Yar Western Shore 2007 sites. Those three sampling sites are grouped together with the Lyakhovsky Eastern Shore Exposure and the Lykhovsky Eastern Shore sites (high similarity, cluster height 0.096). The similarity of the Oygos Yar Western and Eastern Shore 2007 sites to the Bykovsky Mamontovy Khayata Shore and the Lyakhovsky Zimov’e River sites shown from the correlation analysis is not reflected in the clustering. Furthermore, the Oyogos Yar Exposure 2002, Bykovsky Mamontovaya Terrasa, and Lyakhovsky Western Exposure sites are grouped together (high similarity, cluster height 0.079), forming a cluster of exposure-like sites from all three locations. The Bykovsky Cape Mamont, Bykovsky Mamontovy Khayata Exposure, and Oyogos Yar Exposure 2007 sites cannot be associated directly with any cluster. The Lyakhovsky Zimov’e River and Bykovsky Mamontovy Khayata Shore sites form a fourth cluster (high similarity, cluster height 0.076). Again, the similarity of those sites to the Oyogos Yar Western and Eastern Shore 2007 sites demonstrated in the correlation analysis is not reflected in the clustering. The Oyogos Yar Tundra 2002 site is distinctly different from all other sampling sites.
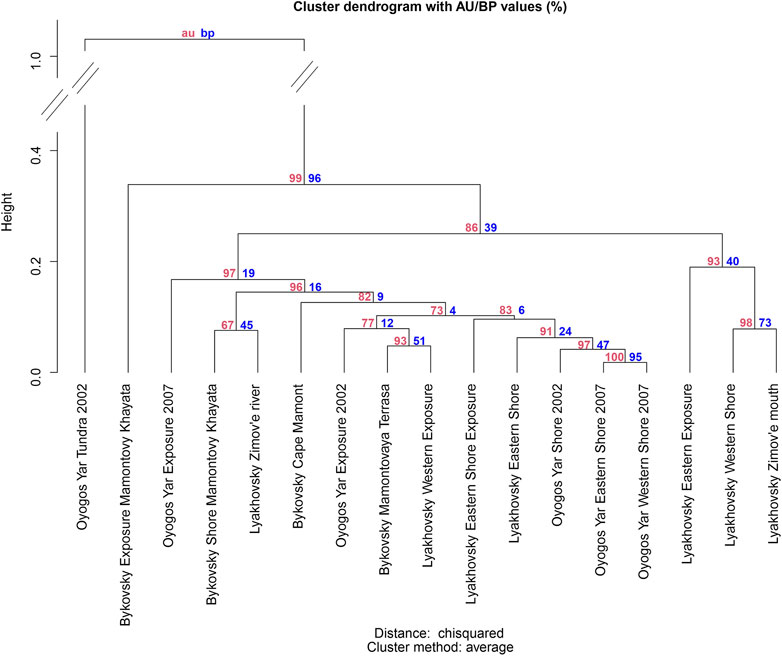
FIGURE 10. Cluster dendrogram for the hierarchical clustering of mammoth fauna species bone counts for all sampling sites. Distances are assessed using the chi-squared method. Clustering method is “average.” Numbers at the dendrogram edges are for basic bootstrapping probability significance values (red) and corrected approximately unbiased significance values (green), statistical significances according to the bootstrapping approach used in pvclust.
In summary, the correlation analysis shows that the Oyogos Yar sampling sites are rather similar to each other, with the exception of the Tundra sample site. Cluster analysis, however, indicates that the Oyogos Yar Shore sites are more similar to shore sites from Bol’shoy Lyakhovsky Island and less similar to the other Oyogos Yar sites. Correlation analysis does not show overall similarities of all sampling sites within the other two locations. High similarities in terms of correlation coefficients between specific sampling sites are often not represented in the cluster analysis. In addition to the “shore cluster,” there is one cluster containing exposure-like sites from all locations (Oyogos Yar Exposure 2002, Bykovsky Mamontovy Terrasa, and Lyakhovsky Western Exposure), where cluster analysis favors the similarities between those samples over similarities to other sites shown in the correlation analysis, e.g., between the Oyogos Yar Exposure 2002 and the Bykovsky Cape Mamont sites and between the Bykovsky Mamontovy Terrasa and the Oyogos Yar Shore sites. Both cluster analysis and correlation analysis show a high similarity between the Bykovsky Mamontovy Khayata Shore and Lyakhovsky Zimov’e River sites. While the correlation analysis clearly indicates low similarities for the Lyakhovsky Eastern Exposure site, cluster analysis places this sample site together with the Lyakhovsky Western Shore and the Lyakhovsky Zimov’e River sites with a statistical significance of 0.93. However, cluster analysis also suggests that this cluster is distinctly different from the other clusters.
The Oyogos Yar coast is known for finds of late Pleistocene Mammoth fauna remains. Exceptional recent findings of frozen mammoth and horse carcasses in permafrost deposits exposed on the Oyogos Yar coast have drawn attention to this paleo-environmental archive. However, to date the paleontological characteristics of the deposits in this area have not been described in detail. There are also few mammal bone remains from this region kept in museums. Our studies in 2002 and 2007 allow us to give a paleozoological description of the Kondrat’eva Yedoma outcrop, which is one of the Oyogos Yar locations that is richest in paleontological remains. The newly presented collection of mammal bones from the Oyogos Yar coast, sampled in 2002 and 2007, provides well-based insights into species composition and prevalence of the regional Mammoth fauna due to its large total size of 1925 bone specimens. The approach of collecting 100% of bone remains both at the outcrop and on the shore allowed for the most complete restoration to date of the percentage of large mammal late Pleistocene Mammoth fauna that inhabited this area.
The collection from Oyogos Yar consists of 13 mammal species, of which woolly mammoth (40.5%), bison (19%), horse (18.8%), and reindeer (15.8%) predominate. Rare findings of woolly rhinoceros, saiga antelope, elk, and moose as well as of cave lion and wolf are each below 1% of the entire collection. This is fairly comparable to findings from other prominent Yedoma outcrops in the Laptev Sea region on Bykovsky Peninsula and on Bol’shoy Lyakhovsky Island.
The identification of various forms of bone material preservation made it possible to identify different groups of bones, indicating their burial conditions. Limb bones containing bone marrow can serve as indicators of rapid burial in permafrost sediments, without subsequent thawing or redeposition. Bones covered with vivianite indicate that the decomposition of organic matter took place under anoxic conditions, which led to the formation of vivianite crusts on the bone surface. This could happen either when fragments of the skeleton were deposited under bog conditions and were subsequently rapidly buried, or as a result of thawing of ancient ice complexes and the formation of taberal deposits.
Parallel radiocarbon dating of Yedoma Ice Complex sediments and mammal bones has produced interesting results. About half of the finite radiocarbon bone dates are younger than the dates of the sediments in which they were found. This indicates that the Yedoma formation occurred later than 32.3 ka BP, and these deposits were subsequently eroded. The formation of the Oyogos Yar Yedoma Ice Complex ceased at about 32 ka BP. Younger deposits are only small local bodies containing a large amount of mammalian bone remains; we were unable to identify these remains during our work on the section. The discovery of a Holocene horse mummy once again confirms our assumptions about the distribution of horses from Taimyr Peninsula to the Kolyma River during the Holocene.
Considering the locations where the bones were found within the modern coastal morphology enhances estimates of the cryostratigraphic and paleontological implications of the bone findings.
Correlation analysis of three bone collections shows that the Oyogos Yar sampling sites are rather similar to each other but not to all sampling sites at two other locations on Bykovsky Peninsula and on Bol’shoy Lykahovsky Island sampled in 1998 and 1999, respectively. The cluster analysis does not reflect the high similarities in terms of correlation coefficients between specific sampling sites.
Publcly available datasets were analyzed in this study. This data can be found here: The datasets analyzed for this study can be found in the PANGAEA database (https://www.pangaea.de/) as Kuzmina (2007; https://doi.org/10.1594/PANGAEA.615803), Kuznetsova (2007; https://doi.org/10.1594/PANGAEA.615995), Kuznetsova (2007; https://doi.org/10.1594/PANGAEA.619071), and Kuznetsova (2009; https://doi.org/10.1594/PANGAEA.728108).
TK designed the study, collected and identified the bone samples. VT, LS, and SW took part in the fieldwork and contributed cryostratigraphic data and interpretations. HM provided statistical analyses. TK, VT, LS, HM, and SW wrote the manuscript and contributed to the final submitted version.
This work was supported by the Laptev Sea System program and the RFBR grant No. 16-05-01116. For financial support of the research TK is grateful to the scholarship program of the Otto Schmidt Laboratory St. Petersburg (OSL Fellowship 2002–2009, 2015 programs). The research VT was supported by the RSF grant No. 21-77-30001.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors gratefully cordially thank their expedition team members in 2002 and 2007, Alexander Derevyagin and Dmitry Dobrynin (both MSU), Viktor Kunitsky (Mel’nikov Permafrost Institute Yakutsk SB RAS), Thomas Opel (AWI), Frank Kienast (Senckenberg Weimar), Lyubov Kudrevataya (Lena Delta Reserve Tiksi), and Andrey Gorodinsky (AARI St. Petersburg). Logistical support was provided by the LENA expeditions 2002 and 2007. Waldemar Schneider (AWI) as well as Dmitry Mel’nichenko and colleagues (Hydrobase Tiksi) are acknowledged. We are grateful to Sebastian Laboor (AWI) who assembled Figure 1 and to Candace O’Connor who helped with native English corrections and editing. N.E. Zaretskaya (Laboratory of Paleogeography, Institute of Geography RAS Moscow, Russia) and J. van der Plicht (Center for Isotope Research, University of Groningen, The Netherlands) are acknowledged for the radiocarbon dating. We are grateful to our colleagues A.S. Tesakov, M.V. Sotnikova and P.A. Nikolskyi (Quaternary Stratigraphy Laboratory, Institute of Geology RAS Moscow, Russia), and A.N. Kalyakin (MSU) for support identifying material from rodents and lagomorphs, predatory mammals, elk, moose, and birds.
Andreev, A. A., Grosse, G., Schirrmeister, L., Kuznetsova, T. V., Kuzmina, S. A., and Bobrov, A. A. (2009). Weichselian and Holocene Palaeoenvironmental History of the Bol'shoy Lyakhovsky Island, New Siberian, Arctic Siberia. Boreas 38, 72–110. doi:10.1111/j.1502-3885.2008.00039.x
Andreev, A. A., Schirrmeister, L., Tarasov, P. E., Ganopolski, A., Brovkin, V., Siegert, C., et al. (2011). Vegetation and Climate History in the Laptev Sea Region (Arctic Siberia) during Late Quaternary Inferred from Pollen Records. Quat. Sci. Rev. 30, 2182–2199. doi:10.1016/j.quascirev.2010.12.026
Andreev, A., Grosse, G., Schirrmeister, L., Kuzmina, S., Novenko, E., Bobrov, A., et al. (2004). Late Saalian and Eemian Palaeoenvironmental History of the Bol'shoy Lyakhovsky Island (Laptev Sea Region, Arctic Siberia). Boreas 33, 319–348. doi:10.1080/03009480410001974
Basilyan, A. E., Anisimov, M. A., Nikolskiy, P. A., and Pitulko, V. V. (2011). Wooly Mammoth Mass Accumulation Next to the Paleolithic Yana RHS Site, Arctic Siberia: its Geology, Age, and Relation to Past Human Activity. J. Archaeol. Sci. 38 (9), 2461–2474. doi:10.1016/j.jas.2011.05.017
Boeskorov, G. G., Potapova, O. R., Mashchenko, E. N., Protopopov, A. V., Kuznetsova, T. V., Agenbroad, L., et al. (2014). Preliminary Analyses of the Frozen Mummies of Mammoth (Mammuthus primigenius), Bison (Bison priscus) and Horse (Equus sp.) from the Yana-Indigirka Lowland, Yakutia, Russia. Integr. Zool. 9, 471–480. doi:10.1111/1749-4877.12079
Boeskorov, G. G., Protopopov, A. V., Mashchenko, E. N., Potapova, O. R., Kuznetsova, T. V., Plotnikov, V. V., et al. (2013). New Findings of Unique Preserved Fossil Mammals in the Permafrost of Yakutia. Dokl. Biol. Sci. 452, 291–295. doi:10.1134/s0012496613050116
Boeskorov, G., Zhegallo, V., Kuznetsova, T., and Tikhonov, A. (2017). “The Possible Cases of “dwarfism” in Woolly Mammoths from Eastern Siberia,” in Abstracts of the VII International Conference Mammoths and their relatives-ICMR-2017 (Taiwan: Taichung), P-02.
Efremov, I. A. (1950). Taphonomy i Geologicheskaya letopis' (Taphonomy and Geological Record). Trudy Paleontologicheskogo Instituta SSSR 24, 1–178. (in Russian).
Grigoriev, A. A. (1932). “Ob Oledenenii Territorii Yakutii v Chetvertichnyi Period (To the Glaciation of Yakutia at the Quaternary),,” in Proceedings of the Quaternary Research Commission (Leningrad: Soviet Academy of Sciences Publishers), 31–42. (in Russian).
Günther, F., Overduin, P. P., Sandakov, A. V., Grosse, G., and Grigoriev, M. N. (2013). Short- and Long-Term Thermo-Erosion of Ice-Rich Permafrost Coasts in the Laptev Sea Region. Biogeosciences 10, 4297–4318. doi:10.5194/bg-10-4297-2013
Gusev, A. I. (1958). K Stratigrafii Chetvertichnykh Otlozhenii Zapadnoi Chasti Primorskoi Ravniny (Towards Stratigraphy of Quaternary Deposits of the Western Part of the Coastal Lowland). Arct. Geol. 80 (5), 79–86. Article Collection on (in Russian).
Ivanov, O. A. (1972). “Stratigrafiya i Korrelyatsiya Neogenovykh i Chetvertichnykh Otlozhenii Subarkticheskikh Ravnin Vostochnoi Yakutii (Stratigraphy and Correlation of Neogene and Quaternary Deposits on the Subarctic Lowlands of the Eastern Yakutia),” in Problemy Izucheniya Chetvertichnogo Perioda (Problems of Investigation of the Quaternary) (Moscow: Nauka), 202–211. (in Russian).
Kaplina, T. N. (2009). Alasnye Kompleksy Severnoi Yakutii (Alas Complexes of Northern Yakutia). Earth’s Cryosphere XIII (4), 3–17. (in Russian).
Kienast, F., Schirrmeister, L., Siegert, C., and Tarasov, P. (2005). Palaeobotanical Evidence for Warm Summers in the East Siberian Arctic during the Last Cold Stage. Quat. Res. 63 (3), 283–300. doi:10.1016/j.yqres.2005.01.003
Kienast, F., Wetterich, S., Kuzmina, S., Schirrmeister, L., Andreev, A. A., Tarasov, P., et al. (2011). Paleontological Records Indicate the Occurrence of Open Woodlands in a Dry Inland Climate at the Present-Day Arctic Coast in Western Beringia during the Last Interglacial. Quat. Sci. Rev. 30 (17-18), 2134–2159. doi:10.1016/j.quascirev.2010.11.024
Konishchev, V. N., and Kolesnikov, S. F. (1981). “Osobennosti Stroeniya i Sostava Pozdnekaynozoiskikh Otlozhenii v Obnazhenii Oyogosskii Yar (Peculiarities of the Structure and Composition of Late Cenozoic Deposits at the Oyogos Yar Exposure)”. Problems Cryolithology IX, 107–117. (in Russian).
Kuzmina, S., Kuznetsova, T., and Sher, A. (1999). Paleontological Research on the Bykovsky Peninsula. Rep. Polar Res. 315, 179227–187257. doi:10.2312/BzP_0315_1_1999
Kuznetsova, T. (2008). Fossils of the Mammoth Fauna. Rep. Polar Mar. Res. 584 (139–140), 215–248. doi:10.2312/BzPM_0584_2008
Kuznetsova, T., and Kuzmina, S. (2000). Paleontological Research at the Southern Coast of Bol'shoy Lyakhovsky Lsland. Rep. Polar Mar. Res. 354 (151–161), 223–253. doi:10.2312/BzP_0354_1_2000
Kuznetsova, T. P. (1965). “Chetvertichnykh Otlozheniyakh s Podzemnym L’dom na Yano-Indigirskoy Nizmennosti i o-ve Bol’shom Lyakhovskom (About Quaternary Deposits with Ground Ice on the Yana-Indigirka lowland and Bol’shoy Lyakhovsky Island)” in Podzemnyi Led (Underground Ice) (Moscow: MSU Publishers), 120–132. (in Russian).
Kuznetsova, T. V. (2007). Paleontological Collection of the “Mammoth” Fauna From the Museum of the Lena Delta Reserve. Rep. Polar Mar. Res. 550 (135–138), 173–195.
Kuznetsova, T. V. (2009). Bones Collection on New Siberian Islands from the Expedition LENA 2007, Appendix 4.2. PANGAEA. doi:10.1594/PANGAEA.728108
Kuznetsova, T. (2003). List of Bone Samples of the New Siberian Islands. Rep. Polar Mar. Res. 466, 289–313. doi:10.1594/PANGAEA.619071
Kuznetsova, T. V., Schirrmeister, L., and Noskova, N. G. (2004). “Kollektsii “Mammontovoy fauny” iz raiona morya Laptevykh v muzeyakh i institutakh Rossiyskoy Arfdemii nauk (Colldection of “Mammoth Fauna” from the Laptev Sea Region in Museums and Institutes of the Russian Academy of Sciences),” in Abstracts, of the conference “Regional Geology Problems, Museum Perspective” (Moscow, 45–50. (in Russian).
Kuznetsova, T. V., Schirrmeister, L., and Tumskoy, V. E. (2015). “Pleistocene - Holocene Terrestrial Palaeoenvironmental Changes at the New Siberian Islands and Adjacent Areas (Arctic Siberia),” in Abstracts of the Third International Conference and Workshop Past Gateways-2015, May 18–22, 2015 (Potsdam, Germany: Terra Nostra), 51.
Kuznetsova, T. V., and Starodubtseva, I. A. (2009). “Mamonty i istoriya geologicheskogo izucheniy poberezhiya morya Laptevikh i Novosibirskikh ostrovov (Mammoths and the History of Geological Research of the Laptev Sea Region and the New Siberian Islands)” in System Laptev Sea and Adjacent Arctic Seas: State of the Art and History of Development. Editors H. Kassens, A. P. Lisitsyn, J. Thiede, E. I. Polyakova, L. A. Timokhov, and I. E. Frolov (Moscow: MSU Publishers), 481–500. (in Russian).
Kuznetsova, T. V., Sulerzhitsky, L. D., and Siegert, C. (2001). “New Data on the “Mammoth” Fauna of the Laptev Shelf Land (Arctic Siberia)”, in Proceedings of First International Congress – The World of Elephants (Rome), 289–292.
Kuznetsova, T. V., Tumskoy, V. E., Schirrmeister, L., and Wetterich, S. (2019). Paleozoologitcheskaya kharakteristika pozdneneopleistotsen - golotsenovykh otlozheniy Bikovskogo poluostrova (Severnaya Yakutiya) (Paleozoological Characteristics of Late Neopleistocene–Holocene Deposits of the Bykovsky Peninsula, Northern Yakutia). Zoological J. 98 (11), 1268–1290. (in Russian). doi:10.1134/S0044513419110102
Kuznetsova, T. V., and van der Plicht, H. (2009). “Pozdnepleistocenovie i golocenovie loshadi Severnoy Yakutii (Late Pleistocene and Holocene Horses of Northern Yakutia)” in Proceedings of the All-Russian Conference “200th Anniversary of Patriotic Paleotology” (Moscow, Russia, 65–66. (in Russian).
Lazarev, P. A. (1980). Antropogenovie loshadi Yakutii. (Anthropogenic Horses in Yakutia). Moscow: Nauka, 1–190. (in Russian).
Lopatin, A. V. (2021). Yuka the Mammoth, a Frozen Mummy of a Young Female Woolly Mammoth from Oyogos. Paleontol. J. 55 (11), 1270–1274. doi:10.1134/S0031030121110046
Nagaoka, D., Saijo, K., and Fukuda, M. (1995). Sedimental Environment of the Edoma in High Arctic Eastern Siberia. Proceedings of the Third Symposium on the joint Siberian permafrost Studies between Japan and Russia in 1994 (Tsukuba, Japan), 8–13.
Neretina, A. N., Gololobova, M. A., Neplyukhina, A. A., Zharov, A. A., Rogers, C. D., Horne, D. J., et al. (2020). Crustacean Remains from the Yuka Mammoth Raise Questions about Non-analogue Freshwater Communities in the Beringian Region during the Pleistocene. Sci. Rep. 10, 859. doi:10.1038/s41598-020-57604-8
Nikolaev, V. I., Kuznetsova, T. V., and Alekseev, O. A. (2011). “Predvaritel’nie rezultati izotopnikh i geokhimicheskikh issledovaniy pozdnepleistotsenovikh mamontov Severnoi Yakutii (Preliminary Results of Isotopic and Geochemical Studies of Late Pleistocene Mammoths of Northern Yakutia)” Izv. Ross. Akad. Nauk. Seriya Geogr. 2, 78–88. (in Russian).
Nikolskiy, P. A., and Basilyan, A. E. (2004). “Mys Svytoi Nos – opornyi razrez chetvertichnykh otlozhenii severa Yano-Indigirskoy nizmennosti (Svyatoi Nos Cape – the Main Section of Quaternary Deposits at the North of Yana-Indigirka Lowland),” in Estestvennaya Istoriya Rossiyskoi Vostochnoi Arktiki v Pleistotsene i Golotsene (The Natural History of the Russian Eastern Arctic in Pleistocene and Holocene) (Moscow: GEOS), 5–13. (in Russian).
Nikolskiy, P. A., Sulerzhitsky, L. D., and Pitulko, V. V. (2011). Last Straw versus Blitzkrieg Overkill: Climate-Driven Changes in the Arctic Siberian Mammoth Population and the Late Pleistocene Extinction Problem. Quat. Sci. Rev. 30, 2309–2328. doi:10.1016/j.quascirev.2010.10.017
Opel, T., Dereviagin, A. Y., Meyer, H., Schirrmeister, L., and Wetterich, S. (2011). Palaeoclimatic Information from Stable Water Isotopes of Holocene Ice Wedges on the Dmitrii Laptev Strait, Northeast Siberia, Russia. Permafr. Periglac. Process. 22, 84–100. doi:10.1002/ppp.667
Opel, T., Wetterich, S., Meyer, H., Dereviagin, A. Y., Fuchs, M. C., and Schirrmeister, L. (2017a). Ground-ice Stable Isotopes and Cryostratigraphy Reflect Late Quaternary Palaeoclimate in the Northeast Siberian Arctic (Oyogos Yar Coast, Dmitry Laptev Strait). Clim. Past. 13, 587–611. doi:10.5194/cp-13-587-2017
Opel, T., Laepple, T., Meyer, H., Dereviagin, A. Y., and Wetterich, S. (2017b). Northeast Siberian Ice Wedges Confirm Arctic Winter Warming over the Past Two Millennia. Holocene 27, 1789–1796. doi:10.1177/0959683617702229
Pavlova, E. Y., and Pitulko, V. V. (2020). Late Pleistocene and Early Holocene Climate Changes and Human Habitation in the Arctic Western Beringia Based on Revision of Palaeobotanical Data. Quat. Int. 549, 5–25. doi:10.1016/j.quaint.2020.04.015
Pitulko, V., Pavlova, E., and Nikolskiy, P. (2017). Revising the Archaeological Record of the Upper Pleistocene Arctic Siberia: Human Dispersal and Adaptations in MIS 3 and 2. Quat. Sci. Rev. 165, 127–148. doi:10.1016/j.quascirev.2017.04.004
Pitulko, V. V., Basilyan, A. E., and Pavlova, E. Y. (2014). The Berelekh Mammoth “Graveyard”: New Chronological and Stratigraphical Data from the 2009 Field Season. Geoarchaeology 29, 277–299. doi:10.1002/gea.21483
Pitulko, V. V. (2011). The Berelekh Quest: A Review of Forty Years of Research in the Mammoth Graveyard in Northeast Siberia. Geoarchaeology 26, 5–32. doi:10.1002/gea.20342
Prokhorova, S. M., and Ivanov, O. A. (1973). Olovonosnye Granitoidy Yano-Indigirskoi Nizmennosti i Svyazannye s Nimi Rossypi (Tin-Bearing Granitoids of the Yana-Indigirka Lowland and Associated with Them Placers). Leningrad: Nedra, 229. (in Russian).
R Core Team (2021). R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing.
Romanovskii, N. N. (1961a). Erozionno-termokarstovye Kotloviny na Severe Primorskikh Nizmennostey Yakutii i Novosibirskikh Ostrovakh (Thrermokarst-Erosional Depressions in the Northern Coastal Lowlands of Yakutia and the New Siberian Islands). Merzlotnye issledovaniya (Permafrost Res.) I, 124–144. (in Russian).
Romanovskii, N. N. (1961b). O Stroenii Yano-Indigirskoi Primorskoi Allyuvialnoi Ravniny i Usloviyakh ee Formirovaniya (About the Structure of the Yana-Indigirka Coastal Alluvial plain and Conditions of its Formation). Merzlotnye issledovaniya (Permafrost Res.) II, 129–138. (in Russian).
Romanovskii, N. N. (1958). Permafrost Envelopment Structures in Quaternary Sediments. Sci. Rep. High. Sch. Geol. Geogr. Sci. 3, 185–188. (in Russian).
Römpler, H., Rohland, N., Lalueza-Fox, C., Willerslev, E., Kuznetsova, T., Rabeder, G., et al. (2006). Nuclear Gene Indicates Coat-Color Polymorphism in Mammoths. Science 313 (5783), 62. doi:10.1126/science.1128994
Rothe, M., Kleeberg, A., and Hupfer, M. (2016). The Occurrence, Identification and Environmental Relevance of Vivianite in Waterlogged Soils and Aquatic Sediments. Earth-Science Rev. 158, 51–64. doi:10.1016/j.earscirev.2016.04.008
Rudaya, N., Protopopov, A., Trofimova, S., Plotnikov, V., and Zhilich, S. (2015). Landscapes of the 'Yuka' Mammoth Habitat: A Palaeobotanical Approach. Rev. Palaeobot. Palynology 214, 1–8. doi:10.1016/j.revpalbo.2014.12.003
Schirrmeister, L., Grosse, G., Kunitsky, V., Magens, D., Meyer, H., Dereviagin, A., et al. (2008). Periglacial Landscape Evolution and Environmental Changes of Arctic Lowland Areas for the Last 60 000 Years (Western Laptev Sea Coast, Cape Mamontov Klyk). Polar Res. 27, 249–272. doi:10.1111/j.1751-8369.2008.00067.x
Schirrmeister, L., Grosse, G., KunitskyMayer, V. H., Derevyagin, A., and Kuznetsova, T. (2003a). Permafrost, periglacial and paleo-environmental studies on New Siberian Islands. Reports on Polar and Marine Research. Russian-German Cooperation System Laptev Sea. The Expeditions Lena 466, 195–314.
Schirrmeister, L., Grosse, G., Schwamborn, G., Andreev, A. A., Meyer, H., Kunitsky, V. V., et al. (2003b). Late Quaternary History of the Accumulation Plain North of the Chekanovsky Ridge (Lena Delta, Russia): A Multidisciplinary Approach. Polar Geogr. 27 (4), 277–319. doi:10.1080/789610225
Schirrmeister, L., Kunitsky, V., Grosse, G., Wetterich, S., Meyer, H., Schwamborn, G., et al. (2011). Sedimentary Characteristics and Origin of the Late Pleistocene Ice Complex on North-East Siberian Arctic Coastal Lowlands and Islands - A Review. Quat. Int. 241, 3–25. doi:10.1016/j.quaint.2010.04.004
Schirrmeister, L., Oezen, D., and Geyh, M. A. (2002a). 230Th/U Dating of Frozen Peat, Bol'shoy Lyakhovsky Island (Northern Siberia). Quat. Res. 57, 253–258. doi:10.1006/qres.2001.2306
Schirrmeister, L., Siegert, C., Siegert, C., Kuznetsova, T., Kuzmina, S., Andreev, A., et al. (2002b). Paleoenvironmental and Paleoclimatic Records from Permafrost Deposits in the Arctic Region of Northern Siberia. Quat. Int. 89, 97–118. doi:10.1016/s1040-6182(01)00083-0
Sher, A., Parmuzin, I., and Bortsov, A. (2000). Mamma1 Bone Collecting. Rep. Polar Res. 354, 181268–182269. doi:10.2312/BzP_0354_1_2000
Sher, A. V., Kuzmina, S. A., Kuznetsova, T. V., and Sulerzhitsky, L. D. (2005). New Insights into the Weichselian Environment and Climate of the East Siberian Arctic, Derived from Fossil Insects, Plants, and Mammals. Quat. Sci. Rev. 24, 533–569. doi:10.1016/j.quascirev.2004.09.007
Sher, A. V. (1971). Säugetierfunde und Pleistozänstratigraphie in der Kolyma Niederung. Berichte Dtsch. Ges. für Geol. Wiss. A 16 (2), 113–125. (in German).
Skvortsov, E. F. (1914). Lensko-Kolymskaya Ekspeditsiya 1909 g. (Lena-Kolyma Expedition, 1909). Izvestiya Imperatorskogo Russkogo Geograficheskogo Obshchestva (Trans. Imperial Russ. Geogr. Soc.) 50 (7), 401–428. (in Russian).
Skvortsov, E. F. (1930). ℌV Pribrezhnykh Tundrakh Yakutii (Dnevnik Astronoma Lensko-Kolymskoy Ekspeditsii 1909 g. (On the Coastal Tundra of Yakutia. (Diary of the Astronomer of the Leno-Kolyma Expedition 1909)),” in Izvestiya Komiteta po Izucheniyu Yakutskoy SSR (Proceedings of the Committee for Yakutian SSR Investigation), XV. Leningrad: Academy of Sciences Publishers, 1–244. (in Russian).
Smirnov, A. N. (2003). Iscopaemaya mamontovaya kost’ (Fossil Mammoth Bone). Proc. NIIGA-VNIIOkeangeologia. Saint-Petersburg. 172. (in Russian).
Spizharskii, T. N. (1940). Quaternary Glaciation of the Lena-Indigirka Region. Problems Arct. 11, 70–81. (in Russian).
Suzuki, R., and Shimodaira, H. (2006). Pvclust: an R Package for Assessing the Uncertainty in Hierarchical Clustering. Bioinformatics 22 (12), 1540–1542. doi:10.1093/bioinformatics/btl117
Suzuki, R., Terada, Y., and Shimodaira, H. (2019). Pvclust: Hierarchical Clustering with P-Values via Multiscale Bootstrap Resampling. R Package Version 2.2-0. Available at: https://CRAN.R-project.org/package=pvclust.
Tomidiaro, S. V., Chernen’kiy, B. I., and Bashlavin, D. K. (1982). “Loess-ice Formation of the Shelf Type and the Outcrop Oyogos Yar,” in Permafrost and Geological Processes and the Palaeogeography of the Northeast Asian Lowlands. Editor Yu. V. Shumilov (Magadan: Soviet Academy of Sciences Publishers, Far-east Scientific Center), 30–53. (in Russian).
Tumskoy, V. E., and Dobrynin, D. (2008). Stratigraphical and Geomorphological Studies along the South Coast of Bol’shoy Lyakhovsky Island and along the Oyogos Yar Coast. Rep. Polar Mar. Res. 584, 41–50. doi:10.2312/BzPM_0584_2008
Tumskoy, V. E., and Kuznetsova, T. V. (2022). Geological Structure and Cryostratigraphy of the Quaternary Deposits of Oyogos Yar (South Coast of the Dmitry Laptev Strait, Northern Yakutia). Front. Earth Sci. 10, 789421 doi:10.3389/feart.2022.789421
Tumskoy, V. E. (2012). Osobennosti Kriolitogeneza Otlozheniy Severnoi Yakutii v Srednem Pleistotsene – Golotsene (Peculiarities of Cryolithogenesis of Deposits in Northern Yakutia in the Middle Neopleistocene-Holocene). Earth’s Cryosphere XVI (1), 12–21. (in Russian).
Vartanyan, S. L., Garutt, V. E., and Sher, A. V. (1993). Holocene Dwarf Mammoths from Wrangel Island in the Siberian Arctic. Nature 362 (6418), 337–340. doi:10.1038/362337a0
Wetterich, S., Kizyakov, A., Fritz, M., Wolter, J., Mollenhauer, G., Meyer, H., et al. (2020). The Cryostratigraphy of the Yedoma Cliff of Sobo-Sise Island (Lena Delta) Reveals Permafrost Dynamics in the Central Laptev Sea Coastal Region during the Last 52 Kyr. Cryosphere 14, 4525–4551. doi:10.5194/tc-14-4525-2020
Wetterich, S., Kuzmina, S., Andreev, A. A., Kienast, F., Meyer, H., Schirrmeister, L., et al. (2008). Palaeoenvironmental Dynamics Inferred from Late Quaternary Permafrost Deposits on Kurungnakh Island, Lena Delta, Northeast Siberia, Russia. Quat. Sci. Rev. 27 (15), 1523–1540. doi:10.1016/j.quascirev.2008.04.007
Wetterich, S., Meyer, H., Fritz, M., Mollenhauer, G., Rethemeyer, J., Kizyakov, A., et al. (2021a). Northeast Siberian Permafrost Ice-Wedge Stable Isotopes Depict Pronounced Last Glacial Maximum Winter Cooling. Geophys. Res. Lett. 48, e2020GL092087. doi:10.1029/2020gl092087
Wetterich, S., Rudaya, N., Nazarova, L., Syrykh, L., Pavlova, M., Palagushkina, O., et al. (2021b). Paleo-Ecology of the Yedoma Ice Complex on Sobo-Sise Island (EasternLena Delta, Siberian Arctic). Front. Earth Sci. 9, 681511. doi:10.3389/feart.2021.681511
Wetterich, S., Rudaya, N., Kuznetsov, V., Maksimov, F., Opel, T., Meyer, H., et al. (2019). Ice Complex Formation on Bol'shoy Lyakhovsky Island (New Siberian Archipelago, East Siberian Arctic) Since about 200 Ka. Quat. Res. 92 (2), 530–548. doi:10.1017/qua.2019.6
Wetterich, S., Rudaya, N., Tumskoy, V., Andreev, A. A., Opel, T., Schirrmeister, L., et al. (2011). Last Glacial Maximum Records in Permafrost of the East Siberian Arctic. Quat. Sci. Rev. 30, 3139–3151. doi:10.1016/j.quascirev.2011.07.020
Wetterich, S., Schirrmeister, L., Andreev, A. A., Pudenz, M., Plessen, B., Meyer, H., et al. (2009). Eemian and Late Glacial/Holocene Palaeoenvironmental Records from Permafrost Sequences at the Dmitry Laptev Strait (NE Siberia, Russia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 279, 73–95. doi:10.1016/j.palaeo.2009.05.002
Wetterich, S., Tumskoy, V., Rudaya, N., Andreev, A. A., Opel, T., Meyer, H., et al. (2014). Ice Complex Formation in Arctic East Siberia during the MIS3 Interstadial. Quat. Sci. Rev. 84, 39–55. doi:10.1016/j.quascirev.2013.11.009
Wetterich, S., Tumskoy, V., Rudaya, N., Kuznetsov, V., Maksimov, F., Opel, T., et al. (2016). Ice Complex Permafrost of MIS5 Age in the Dmitry Laptev Strait Coastal Region (East Siberian Arctic). Quat. Sci. Rev. 147, 298–311. doi:10.1016/j.quascirev.2015.11.016
Zimmermann, H., Raschke, E., Epp, L., Stoof-Leichsenring, K., Schirrmeister, L., Schwamborn, G., et al. (2017). The History of Tree and Shrub Taxa on Bol'shoy Lyakhovsky Island (New Siberian Archipelago) since the Last Interglacial Uncovered by Sedimentary Ancient DNA and Pollen Data. Genes 8, 273. doi:10.3390/genes8100273
Keywords: paleozoology, Mammoth fauna, permafrost, Yedoma Ice Complex, late Pleistocene, west Beringia, Oyogos Yar coast
Citation: Kuznetsova TV, Wetterich S, Matthes H, Tumskoy VE and Schirrmeister L (2022) Mammoth Fauna Remains From Late Pleistocene Deposits of the Dmitry Laptev Strait South Coast (Northern Yakutia, Russia). Front. Earth Sci. 10:757629. doi: 10.3389/feart.2022.757629
Received: 12 August 2021; Accepted: 21 April 2022;
Published: 08 June 2022.
Edited by:
Bruce S. Lieberman, University of Kansas, United StatesReviewed by:
Ryan T. Tucker, Stellenbosch University, South AfricaCopyright © 2022 Kuznetsova, Wetterich, Matthes, Tumskoy and Schirrmeister. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tatiana V. Kuznetsova, dGF0a3V6MjAxMkBtYWlsLnJ1; Lutz Schirrmeister, bHV0ei5zY2hpcnJtZWlzdGVyQGF3aS5kZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.