
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci., 12 January 2023
Sec. Paleontology
Volume 10 - 2022 | https://doi.org/10.3389/feart.2022.1058104
This article is part of the Research TopicComparative Anatomy and Evolution of Aquatic MammalsView all 3 articles
 Romala Govender1,2*
Romala Govender1,2* Felix G. Marx3,4
Felix G. Marx3,4Marine sediments from the western coast of southern Africa record the origin of the Benguela Upwelling System, one of the most productive in the world. High productivity, in turn, is reflected in a diverse marine mammal fossil assemblage, comprising whales, dolphins and a phocid seal. Here, we describe new records of baleen whale (mysticete) fossils from the early Pliocene localities of Saldanha Steel, Milnerton and Langebaanweg, as well as several potentially younger specimens trawled from offshore sediments. The presence of the extinct rorquals Diunatans and Fragilicetus suggests biogeographical links with the eastern North Atlantic and, thus, potentially antitropical population structuring. The trawled specimens also include rorquals (e.g., the blue whale, Balaenoptera cf. musculus), as well as a right whale (Eubalaena) and a pygmy right whale (Caperea). The latter is the first fossil of this family every discovered in Africa, and only the seventh specimen to be reported worldwide.
The Benguela Upwelling System (BUS) along Africa’s southwest coast supports one of the world’s most productive marine ecosystems (Ansorge and Lutjeharms, 2007). After originating during the Late Miocene, the BUS intensified during the early Pliocene (ca 5 million years, Ma), creating a nutrient-rich environment with a diverse marine mammal assemblage (Hendey, 1976; Hendey, 1981; Tankard and Rogers, 1978; Siesser, 1980; Siesser and Dingle, 1981; Govender et al., 2016; Govender, 2019; 2021). The latter inhabited a cold temperate coast featuring scattered islands during times of high sea level (Hendey, 1981; Erasmus, 2005; Roberts et al., 2011). Embayments and lagoons provided shelter, feeding opportunities and likely also breeding grounds, as suggested by the presence of pups and calves (Hendey, 1981; Govender et al., 2012; Govender et al., 2016; Govender, 2019; Govender, 2021).
Previous studies on the Mio-Pliocene marine mammals from western South Africa have revealed a phocid seal (Homiphoca capensis), sperm whales, beaked whales, dolphins, porpoises, right whales, rorquals, and even a grey whale (Hendey and Repenning, 1972; Muizon and Hendey, 1980; Govender et al., 2011; Govender et al., 2012; Govender et al., 2016; Govender, 2015a; Govender, 2015b; Govender, 2018; Govender, 2019; Govender, 2021). Together, the latter suggest links with the North Atlantic, the Mediterranean, and the eastern North and South Pacific (Govender, 2019; Govender, 2021). In addition, offshore finds hint at a diverse beaked whale assemblage, again reflecting high primary productivity (Bianucci et al., 2007).
Here, we further expand the regional Pliocene marine mammal record via new fossils from the localities of Saldanha Steel, Milnerton and Langebaanweg (“E” Quarry). In addition, we report specimens trawled from the seafloor off the Cape Peninsula and south coast (Figure 1A).
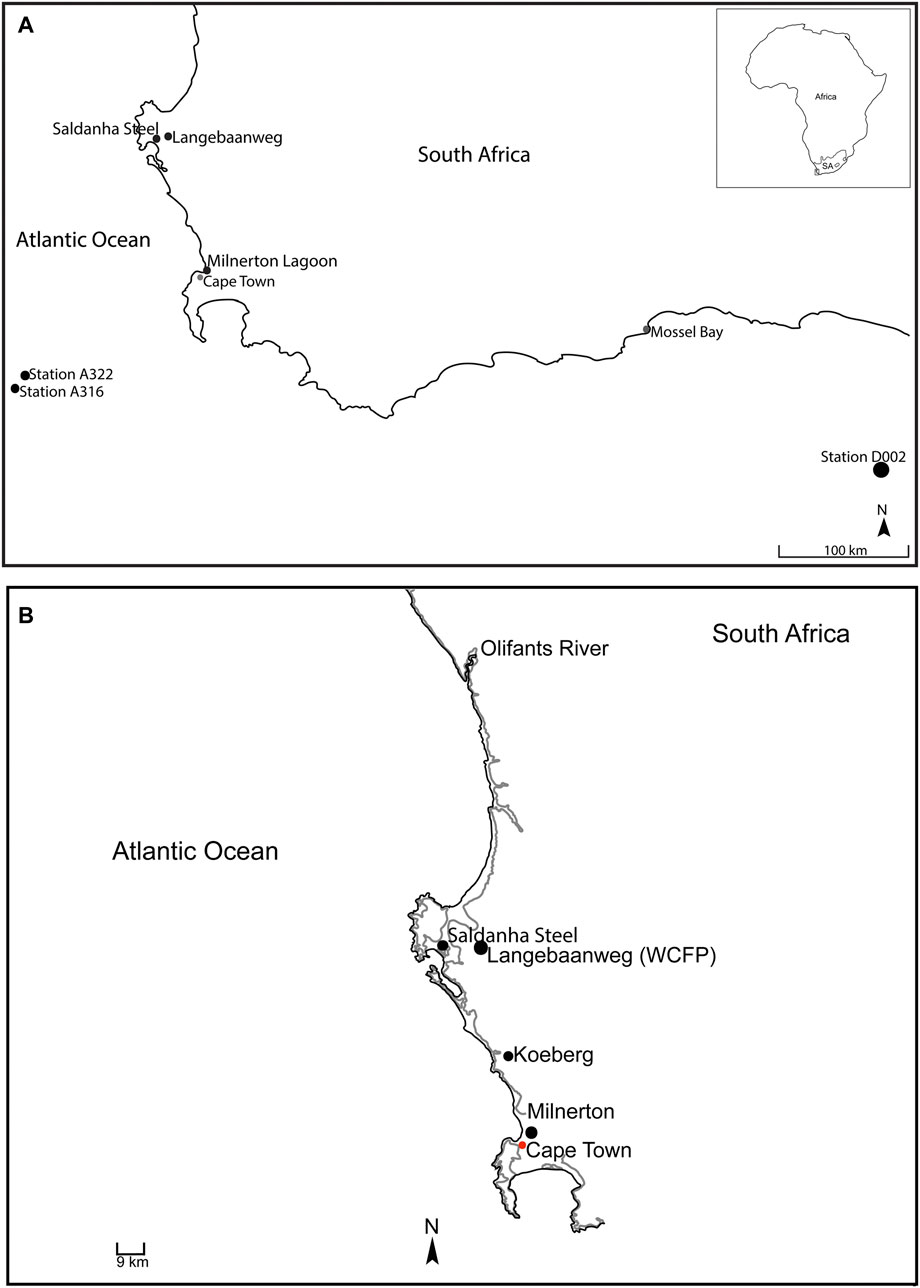
FIGURE 1. (A) Fossil localities on the west coast where specimens were collected. (B) Coastline during the 30 m sea level high when sediments were deposited along the west coast (grey early Pliocene coastline).
During the Mio-Pliocene, sea surface temperatures off the South African west coast changed from warm tropical to cold temperate (Siesser and Dingle, 1981; Pether et al., 2000). Intensification of upwelling created a nutrient-rich coastal environment characterised by moist onshore conditions, warmer offshore sea surface temperatures, and phosphorite formation (Siesser and Dingle, 1981; Tankard and Rogers, 1978).
Marine deposits recording this development occur at the fossil-rich Langebaanweg “E” Quarry, which straddles the Middle Miocene to early Pliocene (Hendey, 1981; Hendey, 1989; Roberts et al., 2011). The specimens described here were collected from the upper part of the Varswater Formation, which records a 30 m transgression and comprises the Langeberg Quartzose Sand (LQSM) and the Muishond Fontein Pelletal Phosphorite Members (MPPM) (Roberts et al., 2011). The latter was deposited in an embayment with a barrier bar between “E” Quarry and granitic offshore islands (Figure 1B; Tankard, 1974; Hendey, 1976; Smith and Haarhoff, 2006), and includes estuarine, marine and some river facies. Fossils from the LQSM and MPPM are mainly concentrated in lag deposits likely dating to the earliest Pliocene, 5.15 ± .1 Ma (Roberts et al., 2011).
In the vicinity of Saldanha Steel, fossiliferous shallow marine deposits occur in channel-like depressions possibly formed by the proto-Berg River (Rogers, 1980; Roberts, 1997). Pristine teeth belonging to coastal sharks suggest a low energy, possibly estuarine environment, but some specimens also show abrasion due to tidal reworking or secondary derivation from older deposits (Roberts 1997). The occurrence of phosphatised shark teeth plausibly locates Saldanha Steel in the upper Varswater Formation, tentatively implying an earliest Pliocene age (see above; Rogers, 1980; Roberts, 1997; Roberts et al., 2011). Warm-water species like Galeocerdo cf. cuvier, Carcharhinus leucas, C. obscurus and Carcharias cf. taurus suggest tropical conditions still notably warmer than today (Tankard, 1975; Hendey, 1981; Roberts, 1997; Kock, 2002). The presence of Isurus and Carcharhinus altimus (including some juveniles) furthermore indicates an inshore environment crossed by deep channels (Kock, 2002).
At Milnerton, the MPPM extends to the mouth of the Diep River (Rogers, 1980; Rogers, 1982; Roberts, 2006; Avery and Klein, 2011). Cetacean fragments and a small terrestrial component (Gomphotherium, equid and rhinoceros remains) have been found at the Milnerton Lagoon mouth, with additional material of uncertain provenance occurring up to 200 m offshore (Avery, 1995). Like Koeberg, this area may have been the site of a temporary lagoon delimited by a barrier spit (Rogers, 1979).
Besides these onshore locations, Neogene offshore deposits occur along the entire coast except for a small gap west of the Agulhas Bank (Rogers, 1980; Dingle et al., 1983; Avery, 1995; Roberts et al., 2006). The overall sediment cover is thin and somewhat erratic, however, and seafloor exposures are limited (Wigley and Compton 2006). Phosphatised cetacean remains trawled from this area could date to the Mio-Pliocene, but plausibly also be as young as Pleistocene or even Holocene (Compton and Wiltshire 2009).
Specimens from Saldanha Steel come from the first and second excavations in 1997 and were retrieved via sieving (Roberts, 1997). They are relatively well preserved, but all suffered some damage. At Milnerton, material was found either at the lagoon mouth or picked up from the beach in 1974 and again in late 1993 to early 1994 (from catalogues). Fossils from this area often show signs of rolling, abrasion, and breakage. Two periotics from Langebaanweg (“E” Quarry) were collected in 1968 (SAM-PQL-11680) and 1977 (SAM-PQL-54708), respectively; they are abraded and fragmentary, and only preserve the pars cochlearis.
Finally, all but one of trawled specimens were collected using a 15-foot beam trawl at stations A316 (34° 42S 16° 54′E; 1,500–1,580 fathoms = 2743.2–2889.5 m) and A322 (34° 36S 17° 00′E; 2743 m) of the RV Africana II routine cruise off the Cape Peninsula in December 1959 (Thandar, 1999). One specimen was collected from Station D002 (35° 01S 24° 60′E; 914 m) of the F. R. S. Africana Voyager III demersal cruise, which began at Cape Agulhas (20°E) in 1993 (Barange et al., 1998). All specimens are now housed in the Cenozoic Palaeontology Collections at Iziko South African Museum (SAM). Anatomical terminology follows Mead and Fordyce (2009) and Ichishima et al. (2021).
Cetacea Brisson, 1762
Mysticeti Gray, 1864
Balaenopteridae Gray, 1864
Balaenoptera Lacépède, 1804
Balaenoptera cf. musculus (Linnaeus, 1758)
Referred material. SAM-PQ-3082—right bulla lacking the posterior portion of the outer lip (Figures 2A–D; Table 1).

FIGURE 2. Balaenoptera cf. musculus SAM-PQ-3082 in (A) ventral, (B) dorsal, (C) medial, and (D) lateral view, scale=5 cm. Balaenoptera sp. SAM-PQ-3086 in (E) posteromedial, (F) posterolateral, (G) posteroventral, and (H) dorsal view, scale=2 cm. Diunatans luctoretemergo SAM-PQSS-223 in (I) medial, (J) lateral, (K) posterior, and (L) dorsal view, scale=2 cm.
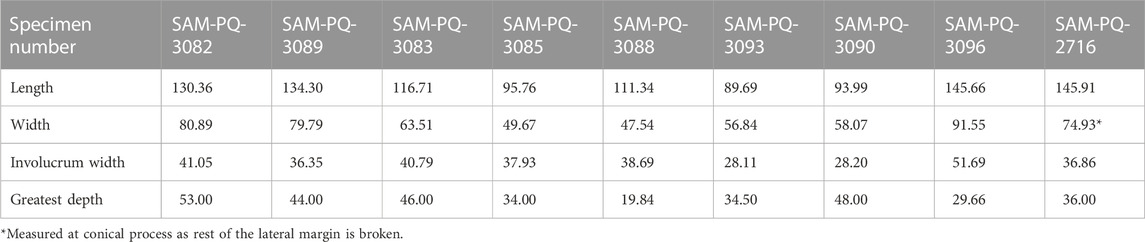
TABLE 1. Measurements (in mm) of isolated tympanic bullae from Saldanha Steel, Langebaanweg, Milnerton, and offshore trawls.
Locality and Horizon. Trawled, RV Africana II, 8 December 1959, Station A316.
Age. Unknown.
Description. In ventral view, the bulla is relatively long (130 mm; Table 1) and marked by a somewhat squared anterior border with a weakly developed anterolateral shelf. In dorsal view, the main ridge is moderately convex. The involucral ridge is notably retracted from both the medial and posterior margins, and anteriorly does not extend beyond the main ridge. The Eustachian outlet is relatively narrow. As far as can be told, the sigmoid process would have been positioned at the midline. In lateral view, the posterior rim of the tympanic cavity lies far forward of the posterior border of the bulla. In medial view, there is a relatively well developed inner posterior prominence which imparts a concave outline to the dorsal surface of the involucrum. The main ridge is prominent and elevated.
Comments. SAM-PQ-3082 closely matches B. musculus in its overall shape, retracted involucral ridge, weakly developed anterolateral shelf, centrally positioned sigmoid process, well developed posterior extension, and notable inner posterior prominence. It is, however, somewhat smaller than extant blue whales, with reported bulla lengths for the latter ranging from 134 to 146 mm (Ekdale et al., 2011). Among extant rorquals, a length of 130 mm is consistent with the upper size range of B. physalus and B. borealis. SAM-PQ-3082 differs from both, however, in having a marked inner posterior prominence, a less developed anterolateral shelf, an anteriorly retracted involucral ridge, and a more centrally positioned sigmoid process.
cf. Balaenoptera sp.
Referred material. SAM-PQ-3086—small left periotic showing signs of weathering (Figures 2E–H; Table 2).
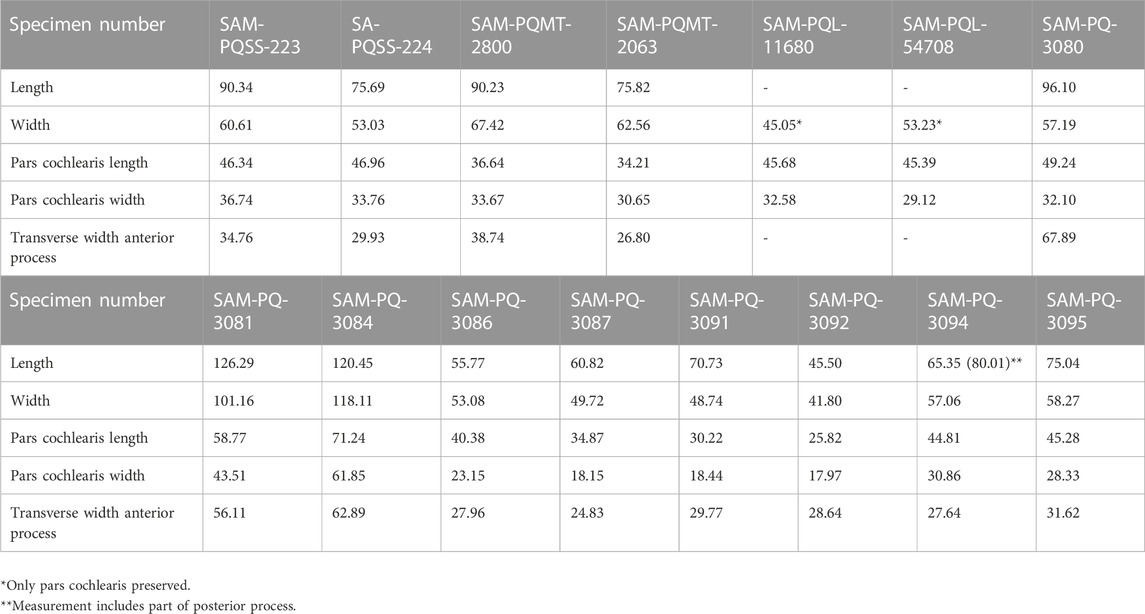
TABLE 2. Measurements (in mm) of isolated periotics from Saldanha Steel, Langebaanweg, Milnerton, and offshore trawls.
Locality and Horizon. Trawled, RV Africana II, 10 December 1959, Station A322.
Age. Unknown
Description. In ventral view, the anterior process is bluntly triangular and somewhat thickened transversely. In medial view, the anterior process is triangular, pointed, and separated from the pars cochlearis by a broad embayment. The lateral tuberosity appears elongate, low, and rounded, but this may at least partially be an artefact of weathering. The pars cochlearis is elongated cranially, and dorsally bears a shallow promontorial groove. The posteromedial border of the pars cochlearis bulges posteriorly and partially obscures the cochlear window. The posterior cochlear crest appears indistinct, but this again may reflect weathering. In dorsal view, the proximal opening of the facial canal is approximately as large as the remainder of the internal acoustic meatus. The perpendicular crest is thin and deeply recessed. The apertures for the cochlear and vestibular aqueducts are slit-like and arranged en echelon. In posterior view, the cochlear window is separated from the aperture for the cochlear aqueduct.
Comments. SAM-PQ-3086 resembles Balaenoptera in having a thin and deeply recessed perpendicular crest, a notable degree of cranial elongation of the pars cochlearis, a broad-based lateral tuberosity, a proximal opening of the facial canal that is roughly as large as the remainder of the internal acoustic meatus, and a cochlear window that is distinct from the aperture for the cochlear aqueduct. It differs from other members of Balaenoptera and Megaptera in retaining a promontorial groove and in having a less pronounced lateral tuberosity; from Diunatans, Fragilicetus and Plesiobalaenoptera in having a thinner, recessed perpendicular crest; from Diunatans, Eschrichtius, Fragilicetus, Gricetoides, Marzanoptera, Plesiobalaenoptera and Protororqualus in having the cochlear window separated from the aperture for the cochlear aqueduct; from Archaebalaenoptera, Eschrichtioides, Eschrichtius, Gricetoides, Incakujira, Miobalaenoptera, Norrisanima and Parabalaenoptera in having a cranially less elongated pars cochlearis; and from Nehalaennia in having a shorter and blunter anterior process and lateral tuberosity in ventral view.
Diunatans Bosselaers and Post, 2010
Diunatans luctoretemergo Bosselaers and Post, 2010
Referred specimen. SAM-PQSS-223 (SAM-PQY-112)—incomplete right periotic (Figures 2I–L; Table 2).
Locality and horizon. Saldanha Steel, excavation, Dump 2/7.
Age. Early Pliocene (Zanclean).
Description. In ventral view, the anterior process is elongate and relatively thick with nearly parallel medial and lateral borders. In medial view, the anterior process is triangular, pointed, and bears a marked fossa for the attachment of the tensor tympani. The lateral tuberosity is bluntly triangular. The pars cochlearis is somewhat elongated cranially but remains longer than tall; its anterior portion remains attached to the anterior process. The posteromedial border of the pars cochlearis bulges posteriorly and partially obscures the cochlear window. The posterior cochlear crest is broadly triangular and points posteriorly. In dorsal view, the proximal opening of the facial canal is separated from the remainder of the internal acoustic meatus by an extremely robust perpendicular crest. The promontorial groove is shallow (partially due to weathering) but broad. In posterior view, the cochlear window is confluent with the aperture for the cochlear aqueduct. In lateral view, there is a cavity of unknown function or origin that may be related to the anteroexternal sulcus.
Comments. SAM-PQSS-223 resembles Diunatans luctoretemergo in having a robust and sharply triangular anterior process bearing a fossa for the tensor tympani, a confluent cochlear window and aperture for the cochlear aqueduct, a thick perpendicular crest, and a broad promontorial groove. It differs from Fragilicetus in having a more pointed anterior process in medial view and a more robust perpendicular crest; from Archaebalaenoptera, Balaenoptera, Eschrichtioides, Eschrichtius, Gricetoides, Incakujira, Marzanoptera, Megaptera, Miobalaenoptera, Norrisanima and Parabalaenoptera in having a cranially less elongated pars cochlearis; from Balaenoptera, Incakujira, Megaptera and Norrisanima in having a confluent cochlear window and aperture for the cochlear aqueduct; from Plesiobalaenoptera in having a more pointed anterior process and a shallower promontorial groove, and in lacking a clearly defined superior process and a sulcus for the facial nerve extending cranially from the proximal opening of the facial canal; from Nehalaennia in having a smaller lateral tuberosity in medial view; from Marzanoptera and Protororqualus in having a more pointed anterior process in medial view; and from Protororqualus in having a thicker perpendicular crest.
Fragilicetus Bisconti and Bosselaers, 2016
Fragilicetus sp.
Referred specimens. SAM-PQSS-224, PQMT-2800, SAM-PQL-54708—incomplete right periotics; SAM-PQMT-2063, SAM-PQ-3084—incomplete left periotics (Figures 3, 4A–H; Table 2).

FIGURE 3. Fragilicetus sp. SAM-PQSS-224 in (A) medial, (B) lateral, (C) posterior, and (D) dorsal view, scale=2 cm. PQMT-2800 in (E) medial, (F) lateral, (G) posterior, and (H) dorsal view, scale=2 cm. SAM-PQL-54708 in (I) medial, (J) lateral, (K) posterior, and (L) dorsal view, scale=2 cm.

FIGURE 4. Fragilicetus sp. SAM-PQMT-2063 in (A) ventromedial, (B) lateral, (C) posterior, and (D) dorsal view, scale=2 cm. SAM-PQ-3084 in (E) medial, (F) lateral view, (G) posterior, and (H) dorsal view, scale=2 cm. Balaenopteridae indet. A, SAM-PQ-3094 in (I) ventromedial, (J) lateral, (K) posterodorsal, and (L) dorsal view, scale=2 cm.
Locality and horizon. SAM-PQSS-224—Saldanha Steel, Dump 1; SAM-PQMT-2063—Milnerton Beach; SAM-PQMT-2800—Milnerton Lagoon, south of the river mouth; SAM-PQ-3084—trawled, RV Africana II, 8 December 1959, Station A316; SAM-PQL-54708—Bed 3aN, Dump 8, MPPM, Langebaanweg.
Age. SAM-PQ-3084—unknown; the remainder all date to the early Pliocene (Zanclean).
Description. In ventral view, the anterior process is elongate and pointed, and bears a distinct ridge for the insertion of the tensor tympani. The body of the periotic is slightly hypertrophied. In medial view, the anterior process is sharply pointed and bordered by a well-developed triangular lateral tuberosity. The pars cochlearis is somewhat elongated cranially but remains longer than tall; its anterior portion is detached from the anterior process. The posteromedial border of the pars cochlearis bulges posteriorly and partially obscures the cochlear window. The posterior cochlear crest is triangular and points directly posteriorly. In dorsal view, the proximal opening of the facial canal is separated from the remainder of the internal acoustic meatus by a robust perpendicular crest. In posterior view, the cochlear window is confluent with the aperture for the cochlear aqueduct.
Comments. The new specimens resemble Fragilicetus in having a confluent cochlear window and aperture for the cochlear aqueduct (in SAM-PQSS-224, SAM-PQMT-2063, SAM-PQMT-2800 and SAM-PQL-54708), a robust perpendicular crest, and an anterior process that is elongate, transversely compressed and bordered by a blunt lateral tuberosity in ventral view. They differ from F. velponi in having a more pointed anterior process in medial view and a triangular, rather than finger-like, posterior cochlear crest; from Archaebalaenoptera, Balaenoptera, Eschrichtioides, Eschrichtius, Gricetoides, Incakujira, Marzanoptera, Megaptera, Miobalaenoptera, Norrisanima, and Parabalaenoptera in having a cranially less elongated pars cochlearis; from Balaenoptera, Incakujira, Megaptera, and Norrisanima in having a confluent cochlear window and aperture for the cochlear aqueduct; from Archaebalaenoptera in having the anterior portion of the pars cochlearis clearly separated from the anterior process; from Diunatans and Plesiobalaenoptera in having a less globose pars cochlearis in ventral view, and in lacking a deeply excavated promontorial groove; from Nehalaennia in having a less globose pars cochlearis and blunter lateral tuberosity; from Parabalaenoptera in having a cranially less elongated pars cochlearis and a medially less excavated anterior process; from Marzanoptera, Miobalaenoptera and Protoroqualus in having a more pointed anterior process in medial view; and from Protororqualus in having a thicker perpendicular crest.
Balaenopteridae indet. A
Referred material. SAM-PQ-3094—left periotic; SAM-PQ-3095—right periotic (Figures 4I–L, 5A–D; Table 2).

FIGURE 5. Balaenopteridae indet. A SAM-PQ-3095 in (A) ventromedial, (B) lateral, (C) posterior, and (D) dorsal view, scale=2 cm. Balaenopteridae indet. SAM-PQ-3081 in (E) medial, (F) lateral, (G) posterior, and (H) dorsal view, scale=2 cm. SAM-PQL-11680, (I) ventromedial, (J) lateral, (K) posterior, and (L) dorsal view, scale=2 cm.
Locality and Horizon. Trawled, RV Africana II, 10 December 1959, Station A322.
Age. Unknown.
Description. In ventral view, the anterior process is short and thick, with a blunt and somewhat squared tip. The body of the periotic shows no sign of hypertrophy. In medial view, the anterior process is likewise somewhat squared and flanked by a short, knob-like lateral tuberosity. The pars cochlearis is slightly elongated cranially, largely thanks to the border of the internal acoustic meatus being raised into a sharp rim. There is a well-marked promontorial groove. The posterior cochlear crest is triangular and points directly posteriorly. In dorsal view, the perpendicular crest is relatively thin and slightly recessed into the internal acoustic meatus. The proximal opening of the facial canal is circular and approximately half the size of the remainder of the internal acoustic meatus. In posterior view, the cochlear window is circular and well separated from the aperture for the cochlear aqueduct.
Comments. SAM-PQ-3094 and 3095 resemble Balaenoptera in having a relatively thin perpendicular crest that is recessed into the internal acoustic meatus, and a cochlear window that is distinct from the aperture for the cochlear aqueduct. They differ from Balaenoptera in having a blunter anterior process, a pars cochlearis showing relatively little cranial elongation, a smaller lateral tuberosity, and a proximal opening of the facial canal that is notably smaller than the remainder of the internal acoustic meatus; from Diunatans, Fragilicetus, and Plesiobalaenoptera in having a thinner, recessed perpendicular crest; from Diunatans, Eschrichtius, Fragilicetus, Gricetoides, Marzanoptera, Plesiobalaenoptera, and Protororqualus in having the cochlear window separated from the aperture for the cochlear aqueduct; from Archaebalaenoptera, Balaenoptera, Eschrichtioides, Eschrichtius, Gricetoides, Incakujira, Marzanoptera, Miobalaenoptera, Norrisanima, Megaptera, and Parabalaenoptera in having a cranially less elongated pars cochlearis; from Nehalaennia in having a blunter anterior process and smaller lateral tuberosity in medial view; and from Protororqualus in having a circular cochlear window and a more crest-like rim of the internal acoustic meatus.
Balaenopteridae indet.
Referred material. SAM-PQ-3092—incomplete left periotic; SAM-PQ-3081, 3087, SAM-PQL-11680—incomplete right periotics; SAM-PQ-2716, 3083, 3089—incomplete left tympanic bullae; SAM-PQ-3085, 3088, 3093—incomplete right tympanic bullae (Figures 5E–L, 6, 7, 8A–H; Tables 1, 2).

FIGURE 6. Balaenopteridae indet. SAM-PQ-3092 in (A) medial, (B) lateral view, (C) posterior, and (D) dorsal view, scale=2 cm. SAM-PQ-3087 in (E) medial, (F) lateral, (G) posterior, and (H) dorsal view, scale=2 cm. SAM-PQ-2716 in (I) ventral, (J) dorsal, (K) lateral, and (L) medial view, scale=5 cm.

FIGURE 7. Balaenopteridae indet., SAM-PQ-3083 in (A) ventral, (B) dorsal, (C) lateral, and (D) medial view, scale=5 cm. SAM-PQ-3089 in (E) ventral, (F) dorsal, (G) lateral, and (H) medial view, scale=5 cm. SAM-PQ-3085 in (I) ventral, (J) dorsal, (K) lateral, and (L) medial view, scale=5 cm.

FIGURE 8. Balaenoptera indet., SAM-PQ-3088, (A) ventral, (B) dorsal, (C) lateral, and (D) medial view, scale=5 cm. SAM-PQ-3093 in (E) ventral, (F) dorsal, (G) lateral, and (H) medial view, scale=5 cm. Caperea sp., SAM-PQ-3090 in (I) ventral, (J) dorsal, (K) lateral, and (L) medial view, scale=5 cm.
Locality and Horizon. SAM-PQ-3081, 3083, 3085—trawled, RV Africana II, 8 December 1959, Station A316; SAM-PQ-3087–3089, 3092 and 3093—trawled, RV Africana II, 10 December 1959, Station A322; SAM-PQ-2716—trawled, F. R. S. Africana, May 1993, Station D002, SAM-PQL-11680—“E” Quarry, Langebaanweg.
Age. SAM-PQL-11680—early Pliocene (Zanclean); the age of the remainder is unknown.
Description. The state of preservation of this material is variable, but all of the periotics except SAM-PQ-3081 have damaged anterior processes and lack the posterior processes, and the tympanic bullae lack the outer lip. In medial view, the pars cochlearis is somewhat cranially elongated but longer than tall and detached from the anterior process. In dorsal view, the proximal opening of the facial canal appears separated from the remainder of the internal acoustic meatus by a robust perpendicular crest, but details are obscured by the poor preservation of SAM-PQ-3092. In SAM-PQ-3087, the cochlear window is confluent with the aperture for the cochlear aqueduct. In ventral view, the tympanic bullae are broadly reniform and rounded transversely. In dorsal view, the main ridge is moderately convex and detached from the medially retracted involucral ridge. The involucral incisure is poorly developed. In medial view, the inner posterior prominence is low to indistinct, with no sign of a medial furrow.
Comments. The periotics resemble balaenopterids in having a cranially somewhat elongated pars cochlearis and differ from extant rorquals in having a robust perpendicular crest. The tympanic bullae resemble balaenopterids in their reniform shape, poorly developed inner posterior prominence, and medially retracted involucral ridge.
Cetotheriidae Brandt, 1872
Neobalaeninae Gray, 1873
Caperea Gray, 1864
Caperea sp.
Referred material. SAM-PQ-3090—incomplete left tympanic bulla (Figures 8I–L; Table 1).
Locality and Horizon. Trawled, RV Africana II, 10 December 1959, Station A322.
Age. Unknown.
Description. In ventral view, the anteromedial surface of the bulla is rounded transversely and lacks an anterolateral shelf. In dorsal view, the anterior margin of the bulla is squared, which in turn reflects the broad, rectangular Eustachian outlet. The main ridge is strongly convex. The involucral ridge extends to the posterior edge of the bulla but is retracted medially. The surface of the involucrum appears smooth with no obvious transverse creases. The medial margin of the involucrum gradually curves posterolaterally. In medial view, the anterior border of the bulla is notably squared and the dorsal surface of the involucrum appears flat with no inner posterior prominence.
Comments. SAM-PQ-3090 resembles Caperea in having squared anterior margins in dorsal and medial view, a broad Eustachian outlet, and a flattened dorsal margin of the involucrum with no inner posterior prominence. It differs from C. marginata in having a medially retracted involucral ridge and a gradually curving, rather than rectangular, medial margin of the involucrum in dorsal view. The absence of transverse creases on the involucrum also differs but could be a juvenile feature given the comparatively small size of this specimen (Ekdale et al., 2011).
Balaenidae Gray, 1821
Eubalaena Gray, 1864
Eubalaena sp.
Referred specimen. SAM-PQ-3096—incomplete right tympanic bulla (Figures 9A–D; Table 1).

FIGURE 9. Eubalaena sp. SAM-PQ-3096 in (A) ventral, (B) dorsal, (C) lateral, and (D) medial view, scale=5 cm. Mysticeti indet., SAM-PQ-3091 in (E) ventromedial, (F) lateral, (G) posterior, and (H) dorsal view, scale=2 cm. SAM_PQ-3080 in (I) ventromedial, (J) lateral, (K) posterior, and (L) dorsal view, scale=2 cm.
Locality and horizon. Trawled off southwest Cape coast.
Age. Unknown.
Description. In ventral view, the anteromedial surface of the bulla is notably concave transversely. There is no anterolateral shelf. In dorsal view, the anterior margin of the bulla is squared and almost perpendicular to the weakly convex main ridge. The Eustachian outlet is relatively narrow, whereas the involucrum is relatively broad throughout. The involucral ridge is neither medially nor anteriorly retracted. The dorsal surface of the involucrum bears numerous well-defined transverse creases. In medial view, the anterior border of the bulla is pointed. From its anterior margin, the involucrum gradually rises dorsally, levels out, and then steeply descends towards the posterior border of the bulla. There is no inner posterior prominence.
Comments. SAM-PQ-3096 resembles Balaena and Eubalaena in being relatively large (146 mm; Table 1), with the bullae of extinct balaenid genera like Antwerpibalaena, Archaeobalaena, Balaenella, Balaenula, Peripolocetus being ≥40 mm shorter. Differs from Balaena in having a less acutely pointed anteromedial corner in ventral view, a somewhat narrower anterior portion of the involucrum in dorsal view, and a more pointed anterior margin in medial view.
Mysticeti indet.
Referred material. SAM-PQ-3091, 3080—right periotics (Figures 9E–L; Table 2).
Locality and horizon. SAM-PQ-3080—trawled, RV Africana II, 8 December 1959, Station A316; SAM-PQ-3091—trawled, RV Africana II, 10 December 1959, Station A322.
Age. Unknown.
Description. In ventral view, the anterior process is slightly thickened and flanked by a blunt lateral tuberosity. In medial view, the anterior process is triangular and relatively smooth, with no depression or ridge for the attachment of the tensor tympani. The pars cochlearis is not cranially elongated and bears a well-marked promontorial groove. The cochlear window is flush with the remainder of the pars cochlearis. In dorsal view, the proximal opening of the facial canal is compressed transversely, smaller than the remainder of the internal acoustic meatus, and separated from the latter by a robust perpendicular crest. In SAM-PQ-3080, a circular hiatus Fallopii opens just anterior to the proximal opening of the facial canal; no such opening is evident in SAM-PQ-3091, but the anterior border of proximal opening of the facial canal is somewhat teardrop-shaped. The suprameatal area is broad and bordered by a low superior process. The aperture for the cochlear aqueduct is circular and neither arranged en echelon with that for the vestibular aqueduct, nor aligned with the proximal opening of the facial canal and the remainder of the internal acoustic meatus. In posterior view, the cochlear window is separated from the aperture for the cochlear aqueduct.
Comments. SAM-PQ-3080 and 3091 differ from balaenids in having a longer, triangular anterior process in medial view, and in lacking marked hypertrophy of the lateral tuberosity and body of the periotic; from balaenopteroids in having an aperture for the cochlear aqueduct that is circular and not arranged en echelon with that for the vestibular aqueduct; and from balaenopterids in lacking a cranially elongated pars cochlearis and recessed cochlear window in medial view. The broad suprameatal area, robust perpendicular crest, and transversely compressed proximal opening of the facial canal are reminiscent of cetotheriids, but there is no obvious alignment of the aperture of the cochlear aqueduct with the proximal opening of the facial canal and the remainder of the internal acoustic meatus.
Biogeography. Modern marine mammals often either have large geographic ranges with pronounced population structuring, or form clades of closely related species separated by a geographic barrier (Yonezawa et al., 2009; Churchill et al., 2014; do Amaral et al., 2017; Holt et al., 2020). The warm waters of the tropics are perhaps the most notable driver of such patterns, with several temperate or cool-water species showing marked antitropicality (Davies, 1963). The biogeography of extinct marine mammals is usually hard to ascertain, owing to the scarcity of described material, time-averaging of fossil assemblages, and strong regional differences in research effort and rock availability (Uhen and Pyenson, 2007; Pyenson et al., 2009).
Notwithstanding these caveats, it seems noteworthy that the extinct rorquals Diunatans and Fragilicetus were first described from the Pliocene of the eastern North Atlantic (Bosselaers and Post, 2010; Bisconti and Bosselaers, 2016). Their presence in South Africa i) suggests a relatively large geographic range, consistent with that of most living rorquals; and ii) necessitates dispersal across the equator prior to the onset of Northern Hemisphere glaciation. Comparatively warm Pliocene conditions would likely have made the tropics hard to cross for many temperate animals. If so, both Diunatans and Fragilicetus could plausibly have exhibited some degree of antitropicality, as previously suggested for certain other extinct cetacean populations and/or species (Bianucci et al., 2016; Govender et al., 2016; Tsai et al., 2017; Bisconti et al., 2021).
Palaeoenvironment. Strengthening of the BUS during the Late Miocene to early Pliocene resulted in cooler temperatures and increased marine primary production along the coast (Dupont et al., 2009). The western margin of South Africa changed from sandy and rocky beaches to sheltered lagoons and embayments (Kensley, 1972; 1977; Pether, 1994; Siesser and Dingle, 1981; Smith and Haarhoff, 2006). This contrasts with the situation today, where the coast is open to the prevailing westerly winds and swells (Barendse and Best, 2014).
Deposition of the MPPM and equivalent strata took place during the 30 m high of the early Pliocene transgression (Hendey, 1981; Roberts et al., 2011). During this time, Saldanha Steel would have been in a shallow marine or estuarine environment (Roberts, 1997), whereas Milnerton was probably located further offshore with the occasional lagoon forming during regressions (Rogers, 1979; 1980). Similar conditions were likely encountered by Diunatans and Fragilicetus in the Pliocene North Sea, where fossils were deposited in a cool-temperate nearshore environment that was 30–50 m deep (see summary in Lambert and Gigase, 2007).
Fossil record of pygmy right whales. Neobalaenines have a largely obscure fossil record that has only recently been somewhat elucidated by finds from Peru, Japan, Italy, Australia and, perhaps, Argentina (Bisconti, 2012; Fitzgerald, 2012; Buono et al., 2014; Tsai et al., 2017; Marx et al., 2018). The tympanic bulla described here (SAM-PQ-3090) is the first pygmy right whale fossil ever reported from Africa, and only the seventh fossil described worldwide. Notably, five of these seven fossils come from the Southern Hemisphere, matching the exclusively southern distribution of living Caperea marginata (Kemper, 2014). Unfortunately, the uncertain age of SAM-PQ-3090 makes it impossible to say whether its presence reflects an ancient geographic pattern, or merely an old record of modern Caperea.
We report new records of the extinct Pliocene rorquals Diunatans and Fragilicetus from the west coast of South Africa. Both genera also once occurred in the Pliocene North Sea, suggesting large geographical ranges and, plausibly, antitropicality. Trawled specimens of uncertain age include further rorquals, as well as a right whale and the first pygmy right whale fossil from Africa.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
This research was funded by an NRF/African Origins Platform grant (grant numbers UID117782 and UID136514).
We thank the Cenozoic collections staff, Sarena Govender (Assistant Collections Manager: Cenozoic Palaeontology) and Dimakatso Mokgoshi (AOP Technical Assistant: Cenozoic Palaeontology) at Iziko South African Museum, Cape Town, for their assistance.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ansorge, I. J., and Lutjeharms, J. R. E. (2007). “The cetacean environment off southern Africa,” in Whales and dolphins of the southern African subregion. Editors P. B. Best (Cape Town: Oxford University PressCambridge University Press), 5–13.
Avery, G., and Klein, R. G. (2011). Review of fossil phocid and otariid seals from the southern western coasts of South Africa. Trans. Roy. Soc. S. Afr. 66 (1), 14–24. doi:10.1080/0035919X.2011.564490
Barange, M., Pillar, S. C., and Hampton, I. (1998). Distribution patterns, stock size and life history strategies of Cape horse mackerel Trachurus trachurus capensis, based on bottom trawl and acoustic surveys. S. Afr. J. Mar. Sci. 19 (1), 433–447. doi:10.2989/025776198784126917
Barendse, J., and Best, P. B. (2014). Shore-based observations of seasonality, movements, and group behavior of southern right whales in a non-nursery area on the South African west coast. Mar. Mamm. Sci. 30, 1358–1382. doi:10.1111/mms.12116
Bianucci, G., Collareta, A., Post, K., Varola, A., and Lambert, O. (2016). A new record of messapicetus from the pietra leccese (late Miocene, southern Italy): Antitropical distribution in a fossil beaked whale (cetacea, ziphiidae). Riv. Ital. Paleontol. Stratigr. 122, 63–74. doi:10.13130/2039-4942/6930
Bianucci, G., Lambert, O., and Post, K. (2007). A high diversity in fossil beaked whales (Mammalia, Odontoceti, Ziphiidae) recovered by trawling from the sea floor off South Africa. Geodiversitas 29, 561–618.
Bisconti, M., and Bosselaers, M. (2016). Fragilicetus velponi: A new mysticete genus and species and its implications for the origin of balaenopteridae (mammalia, cetacea, mysticeti). Zool. J. Linn. Soc. 177 (2), 450–474. doi:10.1111/zoj.12370
Bisconti, M. (2012). Comparative osteology and phylogenetic relationships of Miocaperea pulchra, the first fossil pygmy right whale genus and species (Cetacea, Mysticeti, Neobalaenidae). Zool. J. Linn. Soc. 166 (4), 876–911. doi:10.1111/j.1096-3642.2012.00862.x
Bisconti, M., Ochoa, D., Urbina, M., and Salas-Gismondi, R. (2021). Archaebalaenoptera eusebioi, a new rorqual from the late Miocene of Peru (Cetacea, Mysticeti, Balaenopteridae) and its impact in reconstructing body size evolution, ecomorphology and palaeobiogeography of Balaenopteridae. J. Syst. Palaeontol. 19, 1129–1160. doi:10.1080/14772019.2021.2017363
Bosselaers, M., and Post, K. (2010). A new fossil rorqual (mammalia, cetacea, balaenopteridae) from the early Pliocene of the North Sea, with a review of the rorqual species described by owen and van beneden. Geodiversitas 32 (2), 331–363. doi:10.5252/g2010n2a6
Buono, M. R., Dozo, M. T., Marx, F. G., and Fordyce, R. E. (2014). A Late Miocene potential neobalaenine mandible from Argentina sheds light on the origins of the living pygmy right whale. Acta Palaeontol. Pol. 59, 787–793. doi:10.4202/app.2012.0122
Churchill, M., Boessenecker, R. W., and Clementz, M. T. (2014). Colonization of the Southern Hemisphere by Fur seals and sea lions (Carnivora: Otariidae) revealed by combined evidence phylogenetic and Bayesian biogeographical analysis. Zool. J. Linn. Soc. 172, 200–225. doi:10.1111/zoj.12163
Compton, J. S., and Wiltshire, J. G. (2009). Terrigenous sediment export from the Western margin of South Africa on glacial to interglacial cycles. Mar. Geol. 266, 212–222. doi:10.1016/j.margeo.2009.08.013
Davies, J. L. (1963). The antitropical factor in cetacean speciation. Evolution 17 (1), 107–116. doi:10.2307/2406339
Dingle, R. V., Siesser, W. G., Newton, A. R., Balkema, A. A., do, Amaral, maral, A. R., et al. (1983). Mesozoic and Tertiary Geology of Southern Africa, 25. Rotterdam, 241–259. doi:10.1007/s10914-016-9376-3, and Moreno, I. B. (2017). Historical biogeography of Delphininae dolphins and related taxa (Artiodactyla: Delphinidae)J. Mamm. Evol.
Ekdale, E. G., Berta, A., and Deméré, T. A. (2011). The comparative osteology of the petrotympanic complex (ear region) of extant baleen whales (Cetacea: Mysticeti). PLOS ONE 6, e21311. doi:10.1371/journal.pone.0021311
do Amaral, K. B., Amaral, A. R., Ewan Fordyce, R., and Moreno, I. B. (2017). Historical biogeography of Delphininae dolphins and related taxa (Artiodactyla: Delphinidae). J. Mamm. Evol. 25, 241–259. doi:10.1007/s10914-016-9376-3
Dupont, L. M., Rommerskirchen, F., Condon, T., Mollenhauer, G., and Schefuss, E. (2009). “Historical biogeography of Delphininae dolphins and related taxa (Artiodactyla: Delphinidae),” in American geophysical union, fall meeting abstracts 1, 1396.
Erasmus, L. (2005). Virtual reconstruction of stratigraphy and past landscapes in the West Coast Fossil Park region. Stellenbosch, South Africa: Unpublished MSc ThesisUniversity of Stellenbosch. http://hdl.handle.net/10019.1/2530.
Fitzgerald, E. M. G. (2012). Possible neobalaenid from the Miocene of Australia implies a long evolutionary history for the pygmy right whale Caperea marginata (Cetacea, Mysticeti). J. Vertebr. Paleontol. 32 (4), 976–980. doi:10.1080/02724634.2012.669803
Govender, R., Chinsamy, A., and Ackermann, R. R. (2012). Anatomical and landmark morphometric analysis of fossil phocid seal remains from Langebaanweg, West Coast of South Africa. Trans. Roy. Soc. S. Afr. 67, 135–149. doi:10.1080/0035919X.2012.724471
Govender, R., Avery, G., and Chinsamy, A. (2011). Pathologies in the early Pliocene phocid seals from Langebaanweg, south Africa. S. Afr. J. Sci. 107, 72–77. doi:10.4102/sajs.v107i1/2.230
Govender, R., Bisconti, M., and Chinsamy, A. (2016). A late Miocene–early Pliocene baleen whale assemblage from Langebaanweg, west coast of South Africa (Mammalia, Cetacea, Mysticeti). Alcheringa 40, 542–555. doi:10.1080/03115518.2016.1159413
Govender, R. (2021). Early Pliocene fossil cetaceans from Hondeklip Bay, Namaqualand, south Africa. Hist. Biol. 33 (4), 574–593. doi:10.1080/08912963.2019.1650273
Govender, R. (2018). Extension of the geographic range of Homiphoca into Northern Cape Province, South Africa, with comments on the possible feeding strategies used by Homiphoca. J. Vert. Paleontol. 38 (3), e1463536. doi:10.1080/02724634.2018.1463536
Govender, R. (2019). Fossil cetaceans from Duinefontein (Koeberg) an early Pliocene site on the southwestern Cape, south Africa. Palaeontol. Electron. 22 (1), 1–21. doi:10.26879/673
Govender, R. (2015a). Preliminary phylogenetics and biogeographic history of the Pliocene seal, Homiphoca capensis from Langebaanweg, South Africa. Trans. Roy. Soc. S. Afr. 70 (1), 25–39. doi:10.1080/0035919X.2014.984258
Govender, R. (2015b). Shark–cetacean trophic interaction, duinefontein, koeberg, (5 Ma), south Africa. S. Afr. J. Sci. 111 (11-12), 7. doi:10.17159/sajs.2015/20140453
Hendey, Q. B. (1981). Palaeoecology of the Late Tertiary fossil occurrences in ‘E’ Quarry, Langebaanweg, South Africa, and a reinterpretation of their geological context. Ann. S. Afr. Mus. 84 (1), 1–104.
Hendey, Q. B., and Repenning, C. A. (1972). A Pliocene phocid from South Africa. Ann. S. Afr. Mus. 59, 71–89.
Hendey, Q. B. (1976). The Pliocene fossil occurrences in ‘E’ Quarry, LBW, south Africa. Ann. S. Afr. Mus. 69, 215–247.
Holt, B., Marx, F. G., Fritz, S., Lessard, J.-P., and Rahbek, C. (2020). Evolutionary diversification in the marine realm: A global case study with marine mammals. Front. Biogeogr. 12, e45184. doi:10.21425/F5FBG45184
Ichishima, H., Kawabe, S., and Sawamura, H. (2021). The so-called foramen singulare in cetacean periotics is actually the superior vestibular area. Anat. Rec. 304, 1792–1799. doi:10.1002/ar.24585
Kemper, C. M. (2014). “Family Neobalaenidae (pygmy right whale),” in Handbook of the mammals of the world - volume 4, sea mammals. Editors D. E. Wilson, and R. A. Mittermeier (Barcelona: Lynx Edicions), 214–220.
Kensley, B. F. (1977). A second assemblage of Pliocene invertebrate fossils from Langebaanweg, Cape. Ann. S. Afr. Mus. 72, 189–210.
Kensley, B. F. (1972). Pliocene marine invertebrates from Langebaanweg, Cape province. Ann. S. Afr. Mus. 60, 173–190.
Kock, A. (2002). Preliminary identification of Neogene fossil shark teeth from theVarswater Formation of the west coast of South Africa. [honour’s dissertation]. Cape Town, SA: University of Cape Town.
Lambert, O., and Gigase, P. (2007). A monodontid cetacean from the early Pliocene of the North Sea. Bull. l'Institut R. Sci. Nat. Belg. Sci. Terre 77, 197–210.
Marx, F. G., Park, T., Fitzgerald, E. M. G., and Evans, A. R. (2018). A Miocene pygmy right whale fossil from Australia. PeerJ 6, e5025. doi:10.7717/peerj.5025
Mead, J. G., and Fordyce, R. E. (2009). The therian skull: A lexicon with emphasis on the odontocetes. Smithson. Contrib. Zool. 627, 1–249. doi:10.5479/si.00810282.627
Muizon, C. d., and Hendey, Q. B. (1980). Late Tertiary seals of the South Atlantic Ocean. Ann. S. Afr. Mus. 82, 91–128.
Pether, J. (1994). The sedimentology, palaeontology and stratigraphy of coastal-plain deposits at Hondeklip Bay, Namaqualand, South Africa. [master’s thesis]. Cape Town, South Africa: University of Cape Town.
Pether, J., Roberts, D. L., and Ward, J. D. (2000). “Coastal cenozoic deposits,” in The Cenozoic of Southern Africa. Editors T. C. Partridge (Oxford: U.K Maud Oxford University Press), 33–54.
Pyenson, N. D., Irmis, R. B., Lipps, J. H., Barnes, L. G., Mitchell, E. D., and McLeod, S. A. (2009). Origin of a widespread marine bonebed deposited during the middle Miocene Climatic Optimum. Geology 37 (6), 519–522. doi:10.1130/g25509a.1
Roberts, D. L. (2006). “Lithostratigraphy of the sandveld group,” in South African committee for stratigraphy lithostratigraphic series 9, 25–26.
Roberts, D. L., Botha, G. A., Maud, R. R., and Pether, J. (2006). “Coastal cenozoic deposits,” in The geology of South Africa. Editors M. R. Johnson, C. R. Anhaeusser, and R. J. Thomas (Pretoria: Geological Society of South Africa, Johannesburg/Council for Geoscience), 605–628.
Roberts, D. L. (1997). Report prepared for Saldanha Steel project. Pretoria: Pty) LtdCouncil for Geoscience Geological Survey.Fossil occurrence at the Saldanha Steel site
Roberts, D. L., Matthews, T., Herries, A. I. R., Boulter, C., Scott, L., Dondo, C., et al. (2011). Regional and global context of the late cenozoic Langebaanweg (LBW) palaeontological site: West coast of South Africa. Earth-Sci Rev. 106, 191–214. doi:10.1016/j.earscirev.2011.02.002
Rogers, J. (1979). The sedimentary succession at the Koeberg nuclear power station, Melkbosstrand. 18th Congress of Geological Society of South Africa, 310–322.
Rogers, J. (1980). First report on the cenozoic sediments between Cape Town and eland's bay. Geol. Surv. S. Afr. Open File 165. (unpublished).
Rogers, J. (1982). Lithostratigraphy of Cenozoic sediments between Cape Town and Eland’s Bay. Palaeoecol. Afr. 15, 121–137.
Siesser, W. G., and Dingle, R. V. (1981). Tertiary Sea-level movements around southern Africa. J. Geol. 89 (4), 523–536. doi:10.1086/628618
Siesser, W. G. (1980). Late Miocene origin of the Benguela upswelling System off northern Namibia. Sci. Ser. 208 (4441), 283–285. doi:10.1126/science.208.4441.283
Smith, R. M. H., and Haarhoff, P. (2006). Sedimentology and taphonomy of an early Pliocene sivathere bonebed at Langebaanweg, western Cape province, south Africa. Afr. Nat. Hist. 2, 197.
Tankard, A. J., and Rogers, J. (1978). Late cenozoic palaeoenvironments on the west coast of southern Africa. J. Biogeog. 5 (4), 319–337. doi:10.2307/3038026
Tankard, A. J. (1975). The late cenozoic history and palaeoenvironments of the coastal margin of the South-western Cape province, South Africa. Unpublished PhD thesis: South Africa. Rhodes University.
Tankard, A. J. (1974). Varswater Formation of the langebaanweg-saldanha area, Cape province. Trans. Geol. Soc. S. Afr. S. Afr. J. Geol.) 77, 263–283.
Thandar, A. S. (1999). Deep-sea holothuroids taken by the R.V. Africana II in 1959, from off the west coast of the Cape Peninsula, South Africa. Ann. S. Afr. Mus. 105 (9), 363–409.
Tsai, C.-H., Collareta, A., Fitzgerald, E. M. G., Marx, F. G., Kohno, N., Bosselaers, M., et al. (2017). Northern pygmy right whales highlight Quaternary marine mammal interchange. Curr. Biol. 27, R1058–R1059. doi:10.1016/j.cub.2017.08.056
Uhen, M. D., and Pyenson, N. D. (2007). Diversity estimates, biases and historiographic effects: Resolving cetacean diversity in the tertiary. Palaeontol. Electron. 10, 10–12. http://palaeo-electronica.org/paleo/2007_2/00123/index.html.
Wigley, R. A., and Compton, J. S. (2006). Late Cenozoic evolution of the outer continental shelf at the head of the Cape Canyon, South Africa. Mar. Geol. 22, 1–23. doi:10.1016/j.margeo.2005.09.015
Keywords: Neogene, Quaternary, west coast, Benguela Upwelling System, Diunatans, Fragilicetus, Eubalaena, Caperea
Citation: Govender R and Marx FG (2023) New cetacean fossils from the late Cenozoic of South Africa. Front. Earth Sci. 10:1058104. doi: 10.3389/feart.2022.1058104
Received: 30 September 2022; Accepted: 28 December 2022;
Published: 12 January 2023.
Edited by:
Borja Figueirido, University of Malaga, SpainReviewed by:
Michelangelo Bisconti, University of Turin, ItalyCopyright © 2023 Govender and Marx. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Romala Govender, cmdvdmVuZGVyQGl6aWtvLm9yZy56YQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.