
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci., 13 January 2023
Sec. Paleontology
Volume 10 - 2022 | https://doi.org/10.3389/feart.2022.1048800
This article is part of the Research TopicCo-Evolution of Life and Environment During the Ediacaran-Cambrian TransitionView all 6 articles
 Zuchen Song1
Zuchen Song1 Junfeng Guo1,2*
Junfeng Guo1,2* Jian Han2
Jian Han2 Heyo Van Iten3,4*
Heyo Van Iten3,4* Yaqin Qiang1
Yaqin Qiang1 Jiaxin Peng1Jie Sun1,2Yajuan Zheng1
Jiaxin Peng1Jie Sun1,2Yajuan Zheng1 Xinyao Huang1
Xinyao Huang1 Zhifei Zhang2
Zhifei Zhang2Hexangulaconulariids, a family of biradially symmetrical medusozoan cnidarians, have been widely reported from the Lower Cambrian of South China. The four currently recognized genera of hexangulaconulariids differ from each other mainly in the number of faces in the abapical region of the periderm. However, previously published illustrations of the monospecific type genus, Hexangulaconularia, clearly show two distinct morphotypes, one with six faces and the other with 10. Specimens with 10 faces are herein reassigned to the genus Decimoconularia. In addition, the new species D. anisfacialis is described from the Kuanchuanpu Formation (Cambrian Fortunian Stage) in the Kuanchuanpu and Shizhonggou sections in Ningqiang County, southern Shaanxi Province, China. Also described are additional specimens of H. formosa from the same formation in the Zhangjiagou section in Xixiang County, southern Shaanxi Province, and from Member 2 of the Yanjiahe Formation (Cambrian Fortunian Stage) in western Hubei Province. The discovery of D. anisfacialis extends the known stratigraphical range of Decimoconularia, now composed of two species, downward from Cambrian Stage 2 into the Fortunian Stage. Additionally, certain specimens previously assigned to H. formosa are reassigned to D. anisfacialis. The diagnoses of Hexangulaconularia, Decimoconularia, and Hexangulaconulariidae are emended accordingly. In accordance with the rule of time priority, the previously designated type genus and species, Hexaconularia He and Yang, 1986 and Hexaconularia sichuanensis He and Yang, 1986, are replaced herein by Hexangulaconularia He, in Xing et al., 1983.
Small shelly fossils (SSFs), the dominant faunal element of the initial stages of the Cambrian Explosion, preserve a wealth of information concerning the early diversification of the metazoan phyla (e.g., Qian, 1989; Bengtson et al., 1990; Li et al., 2007; Shu, 2008; Shu et al., 2014; Zhu et al., 2019; Guo et al., 2020b). SSFs have been studied intensively for several decades and additional material continues to emerge, permitting in some cases the erection of new species and the revision of existing supraspecific taxa (e.g., Guo et al., 2020a; 2021a). One such taxon, the Family Hexangulaconulariidae (He, 1987), is thought to be an extinct group of polypoid medusozoan cnidarians with a biradially symmetrical, bipartite periderm having a variable number of faces in the periderm’s fan-shaped abapical region. In specimens where the faces are clearly delineated, these features are corrugated planes forming obtuse and/or acute interior angles. In addition to the variable number of faces, hexangulaconulariids also exhibit variation in the relative widths of the faces, the anatomy of the boundaries between them, the rate of adapertural expansion of the adapical region, and the degree of differentiation of this region from the apical region. The family currently comprises four genera: Arthrochites Chen, 1982 and Hexangulaconularia He in Xing et al., 1983, previously described from the Cambrian Fortunian Stage (Xing et al., 1983; Qian, 1989; Conway Morris and Chen 1992), and Decimoconularia (Guo et al., 2021; Guo et al., 2021a) and Septuconularia (Guo et al., 2020; Guo et al., 2020a; Song et al., 2022), previously described from Cambrian Stage 2. All four currently known genera are from South China.
Decimoconularia differs from other hexangulaconulariids in having a total of 10 faces in the abapical region (Guo et al., 2021a). In the type species, D. isofacialis, the 10 faces are essentially equal in width (Guo et al., 2021a). The present report describes new Decimoconularia specimens from the Fortunian Kuanchuanpu Formation in southern Shaanxi Province. Also described are additional Hexaconularia specimens from the same rock unit and locality and from Member 2 of the Yanjiahe Formation (Cambrian Fortunian Stage). The new Decimoconularia specimens differ from D. isofacialis in several characteristics, including the relative widths of the faces. We believe that these differences necessitate the erection of a new species, D. anisfacialis, to increase the total number of species in Decimoconularia to two and extend the stratigraphical range of this genus downward into the Fortunian Stage. Additionally, specimens placed by previous authors (He and Yang, 1986; Conway Morris and Chen, 1992; Van Iten et al., 2010; Zheng, 2012; Steiner et al., 2014; Duan et al., 2017; Xian et al., 2018; Zhang et al., 2018; Han et al., 2020) in Hexangulaconularia are reassigned to D. anisfacialis. We also emend the diagnoses of both Hexangulaconularia and Decimoconularia, as well as the diagnosis of the family Hexangulaconulariidae.
Thirteen Decimoconularia anisfacialis n. sp. specimens were extracted from phosphatic limestone samples collected from the 1) Kuanchuanpu Formation (Cambrian Fortunian Stage) in the Shizhonggou section (SN021-K29-14, SN023-K30-189, SN047-K30-76, SN096-K44-131, SN096-K44-295, SN114-K32-79, and SN116-K52-53) and Kuanchuanpu section (KP-45-1, KP-45-2, KP-45-3, KP-45-4, and KP-45-5) (Ningqiang County, southern Shaanxi Province) and the 2) Kuanchuanpu Formation in the Zhangjiagou section (XX-99-284) in Xixiang County (Steiner et al., 2007, Figure 1). Nine Hexangulaconularia formosa specimens were collected from the Kuanchuanpu Formation in the Zhangjiagou section (XX33-52, XX62-238, XX72-47, XX78-452, XX79-65, XX91-610, XX94-79, and XX96-250) in Xixiang County, southern Shaanxi Province, and from Member 2 of the Yanjiahe Formation (Cambrian Fortunian Stage) in the Gunziao section (CUBar66-2) in Yichang city, western Hubei Province (Guo et al., 2020a, Figures 1.1–1.4).
All 22 fossil specimens were hand-picked under a binocular microscope from insoluble residues produced by the digestion of the rock samples in 10% acetic acid. Gold-coated specimens were imaged using an FEI Quanta 400 FEG scanning electron microscope (SEM). Sixteen specimens (SN, XX) are deposited in the Early Life Institute (ELI), Northwest University, Xi’an, China; the other six specimens (KP, CU) are housed at Chang’an University, Xi’an, China.
Terminology—The anatomical terminology used herein mostly follows the precedents in Conway Morris and Chen (1992), Van Iten et al. (2010), Han et al. (2018), and Guo et al. (2020a, 2021a).
Phylum Cnidaria Verrill, 1865.
Subphylum Medusozoa Petersen, 1979.
Subclass ?Conulata Moore and Harrington, 1956.
Family Hexangulaconulariidae He, 1987.
Type genus—Hexangulaconularia He, in Xing et al., 1983 from the Hongcunping Formation (Cambrian Fortunian Stage) in the Maidiping section, Emei County, Sichuan Province, China.
Emended diagnosis—Laterally compressed, adapically tapered, biradially symmetrical periderm with variably differentiated apical and abapical regions (portions). The abapical region comprises six to 14 more or less well-defined, adapically narrowing faces with transverse ribbing and bounded by shallow narrow furrows and/or ridges. The wide oral end of the periderm is open, with the apical end originally being rounded. The relatively small apical region is weakly to strongly differentiated from the rest of the periderm, with or without somewhat irregular, transverse fine wrinkles. The interspaces between the transverse ribs of the abapical region may be crossed by closely spaced, irregular wrinkles.
Remarks—The emended diagnosis above is consistent with that of Guo et al. (2021a), differing mainly in explicitly noting the presence of more or less distinct, apical and adapical regions.
| Genus Hexangulaconularia He, in Xing et al., 1983, emend.
1983 ?Conulariella (nomen nudum) Yang et al., p. 108
1983 Hexaconularia (nomen nudum) Yang et al., p. 108
1983 Hexangulaconularia He, in Xing et al., p.171
1986 Hexaconularia He and Yang, p. 35
1987 Hexangulaconularia He; He, p. 11
1987 Hexaconularia He and Yang; He, p. 12
1987 Hexaconularia He and Yang; Braier and Singh, p. 337
1989 Hexangulaconularia He; Qian, p. 160
1992 Hexaconularia He and Yang; Conway Morris and Chen, pp. 400–401.
Type species—Hexangulaconularia formosa He, in Xing et al., 1983 from the Hongcunping Formation (Cambrian Fortunian Stage) in the Maidiping section, Emei County, Sichuan Province, China.
Emended diagnosis—The periderm consists of clearly differentiated apical and adapical regions, with a total of six faces in the adapical region, three on each of the two major sides of the periderm.
Stratigraphical range—Cambrian Fortunian Stage.
Remarks—The genera Hexangulaconularia and Hexaconularia were formally established in 1983 (Xing et al., 1983) and 1986 (He and Yang, 1986), respectively. Thus, according to the rule of time priority in the International Code of Zoological Nomenclature (ICZN, 23) regulations, the genus Hexaconularia is invalid. While the genus Hexangulaconularia is currently monotypic, this may change. The emended diagnosis herein presented is based on characteristics appropriate to distinguish among hexangulaconulariid genera, with other previously included characteristics (Van Iten et al., 2010) best serving to distinguish among species.
| Hexangulaconularia formosa He, in Xing et al., 1983.
1983 ?Conulariella quadratas (nomen nudum) Yang et al., p. 108, pl. III, Figure 9
1983 Hexaconularia multicostata (nomen nudum) Yang et al., p. 108, pl. III, Figures 6, 12
1983 Hexaconularia nanjiangensis (nomen nudum) Yang et al., p. 108, pl. III, Figure 8
1983 Hexaconularia sp. (nomen nudum) Yang et al., p. 108, pl. III, Figure 7
1983 Hexaconularia sichuanensis (nomen nudum) Yang et al., p. 108, pl. III, Figure 10
1983 Hexaconularia xinliensis (nomen nudum) Yang et al., p. 108, pl. III, Figure 11
1983 Hexangulaconularia formosa He, in Xing et al., p. 172, pl. 15, Figures 1–3
1986 ?Conulariella quadratas He and Yang, p. 38–39, pl. IV, Figure 1
1986 Hexaconularia breviculus He and Yang, p. 36, pl. IV, Figures 7–9
1986 Hexaconularia sichuanensis He and Yang, pp. 35–36, pl. IV, Figures 5, 6
1986 Hexaconularia xinliensis He and Yang, pp. 36–37, pl. IV, Figures 3, 4
1986 Hexaconularia xinliensis compressiformis He and Yang, p. 37, pl. V, Figure 1
1986 Hexaconularia xinliensis crassiformis He and Yang, p. 37, pl. V, Figures 3, 4
1986 Hexaconularia bellalus He and Yang, p. 37, pl. V, Figures 8–10
1986 Hexaconularia bellalus compressiformis He and Yang, p. 37, pl. V, Figure 2
1986 Hexaconularia multicostata He and Yang, p. 38, pl. V, Figures 5, 6
1986 Hexaconularia longelus He and Yang, p. 38, pl. V, Figures 11–12
1987 ?Barbitositheca ansata Brasier and Singh, pp. 335, 337, Figure 7.3
1987 Hexaconularia cf. formosa Brasier and Singh, p. 337, Figure 7.4–7.6
1987 Hexaconularia xinliensis He and Yang; He, p. 12, pl. I, Figure 8
1987 Hexangulaconularia formosa He; He, p. 12, pl. I, Figures 9–10
1989 Hexangulaconularia formosa He; Qian, pp.161–162, pl. 28, Figures 7–9
1989 Hexangulaconularia xinliensis He and Yang; Qian, pp. 162–163, pl. 10, figs. 12–13, pl. 28, Figure 6
1992 Hexaconularia sichuanensis He and Yang; Conway Morris and Chen, pp. 400–401, Figures 12.8, 12.11–12.14
2004 Hexaconularia sichuanensis He and Yang; Steiner et al., p. 270, Figures 8.1–8.2
2010 Hexaconularia sichuanensis He and Yang; Van Iten et al., p. 193, 195, Text-Figures 1–2
2012 Hexaconularia sichuanensis He and Yang; Zheng, pp. 70–72, Figures 4.17A–F
2014 Hexaconularia sichuanensis He and Yang; Steiner et al., p. 104, Figures 7.19–7.20
2014 Hexaconularia sichuanensis He and Yang; Yang et al., p. 39, Figure 12N
2016 Hexaconularia sichuanensis He and Yang; Liu et al., p. 1045, Figures 1.9
2017 Pseudooides prima Qian; Duan et al., pp. 7–9, Figures 5A–G, 5J–5K
2017 Hexaconularia sichuanensis He and Yang; Zhang et al., pp. 271–272, Figures 1, 2
2018 Hexaconularia sichuanensis He and Yang; Shao et al., p. 7, Figures 5.4–5.5
Holotype—10134 from the Maidiping section in Emei County.
Paratypes—10135 from the same stratigraphic level and locality as the holotype.
New material—Nine specimens from the Kuanchuanpu Formation (Cambrian Fortunian Stage) in the Zhangjiagou section in Xixiang County, southern Shaanxi Province (XX33-52, XX62-238, XX72-47, XX78-452, XX79-65, XX91-610, XX94-79, XX96-250) and from Bed 2 of the Yanjiahe Formation (Cambrian Fortunian Stage) in the Gunziao section near the city of Yichang, western Hubei Province (CUBar66-2), China.
Description—Incomplete (broken) specimens ranging from 1.1 to 1.5 mm in length along the apical-abapical axis and from 1.4 to 1.8 mm in maximum width (Table 1). The apical region is missing or very poorly preserved. The original apertural margin was not preserved. The periderm is very thin and apparently non-laminar. The angle of adapertural expansion of the abapical region (abr) ranges from approximately 46° to 82° (Table 1) in the plane of its greatest width, with the two major sides of the abapical region meeting along a pair of marginal longitudinal ridges (mlr) (Figure 1D). The apertural margin was not preserved. Each of the two major sides of the adapical region consists of three well-defined, adapically tapering faces. The boundaries between the broad central face and the two relatively narrow faces bordering it are marked by more or less well-defined, shallow narrow furrows along which the transverse ribs terminate (Figure 1). The transverse ribs on the four narrow faces are straight to gently arched abapically and variably spaced (0.04–0.08 mm) (Table 1) in most specimens, more closely so near the apical end of the faces. The transverse ribs on the broad central face are variable in course; they are rectilinear in some cases but with a sharp bend near the interfacial boundaries (Figures 1G,H,I). In other cases, they are slightly undulatory, with or without a sharp bend near the interfacial boundaries.
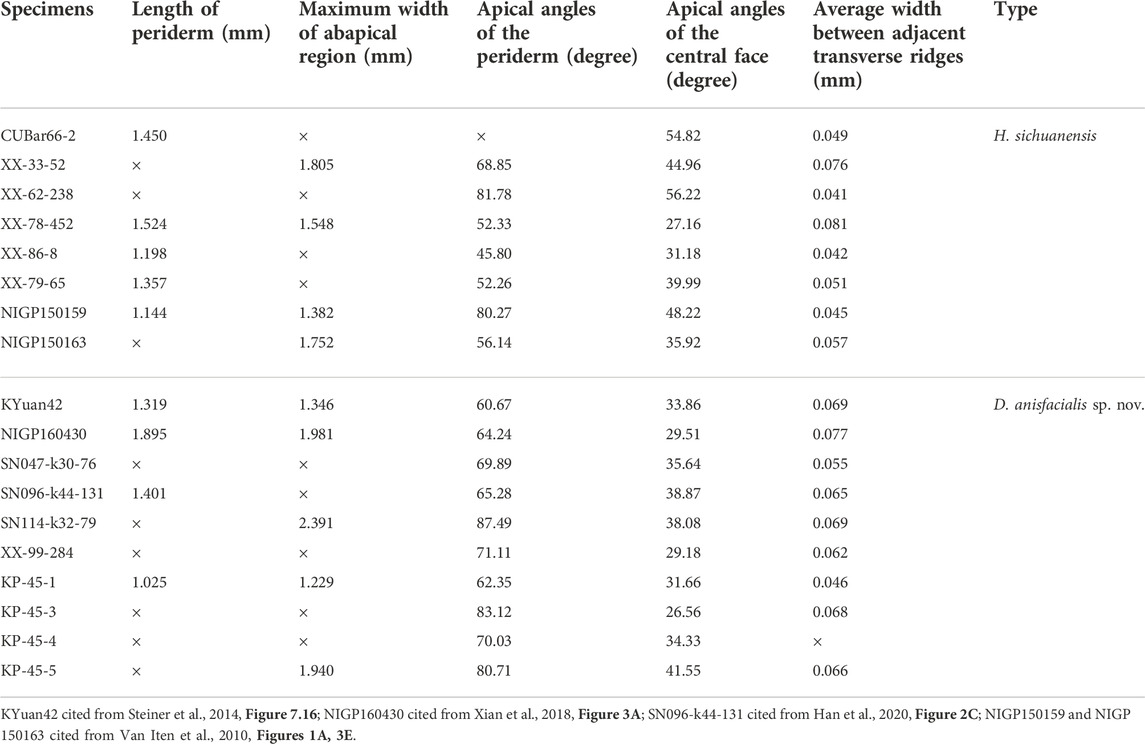
TABLE 1. Measurements of eight well-preserved Hexaconularia sichuanensis and 10 well-preserved Decimoconularia anisfacialis sp. nov specimens.

FIGURE 1. Hexangulaconularia formosa from the Kuanchuanpu Formation (Cambrian Fortunian Stage) of southern Shaanxi Province and Bed 2 of the Yanjiahe Formation (Cambrian Fortunian Stage) of western Hubei Province, China. (A–G,I) XX78-452, XX79-65, XX91-610, XX62-238, XX33-52, CUBar66-2, XX94-79, and XX96-250, respectively. (H) Magnified view of an interfacial (longitudinal) furrow and transverse ribs (G) showing sharp, abapical bending of the transverse ribs near the interfacial furrow and the absence of such a furrow along the locus of the points of inflection of the transverse ribs. Abbreviations: abr=abapical region; apr=apical region; cf=central face; if=interfacial (longitudinal) furrow; lfs=lateral faces; mlr=marginal longitudinal ridge; tr=transverse rib.
Remarks—The genera Hexangulaconularia and Hexaconularia were both first reported in 1983 (Xing et al., 1983; Yang et al., 1983). However, Yang et al. (1983) did not provide a description (only illustrations), and it was not until 1986 that Hexaconularia, with type species Hexaconularia sichuanensis, was officially established (He and Yang, 1986). Hence, Hexaconularia and Hexaconularia sichuanensis are nomen nuda and are replaced herein by Hexangulaconularia and H. formosa, respectively. The Kuanchuanpu and Yanjiahe formation specimens described above are similar to each other and to specimens of Hexangulaconularia formosa previously described from the Kuanchuanpu Formation (e.g., Van Iten et al., 2010). The course of the transverse ribs on the broad central face is variable, with some ribs consisting of three distinct, rectilinear segments (namely, a long central segment and two shorter lateral segments, each deflected toward the apical end of the periderm) and others being slightly undulatory but lacking well-defined bending near their two ends. The geometry of the transverse ribs and the angle of expansion (apical angle) of the central faces of Hexangulaconularia are similar to the two broad faces of the conulariid Conulariella Bouček, 1939 (Ordovician, western Europe and South China; Van Iten et al., 2013); thus, it is not surprising that some authors (Yang et al., 1983; He and Yang, 1986) (provisionally) assigned Hexaconularia specimens to Conulariella.
| Stratigraphical range—Cambrian Fortunian Stage.
Genus Decimoconularia (Guo et al., 2021), emend.
Type species—Decimoconularia isofacialis (Guo et al., 2021) from the Watsonella crosbyi Assemblage Zone of the Yanjiahe Formation (Member 5, Cambrian Stage 2) in the Yanjiahe section near the city of Yichang, Hubei Province, China (Guo et al., 2021b).
Emended diagnosis—The apical region of the periderm is weakly to strongly differentiated. The adapical region has a total of 10 faces, with five on each of the two major sides.
Remarks—As in the case of the emended diagnosis of Hexangulaconularia, the diagnosis of Decimoconularia herein omits those characteristics which do or may potentially differentiate species within the genus.
| Decimoconularia anisfacialis new species.
1986 Hexaconularia nanjiangensis He and Yang, p. 36, pl. IV, Figure 2
1986 Hexaconularia multicostata He and Yang, p. 38, pl. V, Figure 7
1992 Hexaconularia sichuanensis He and Yang; Conway Morris and Chen, pp. 400–401, Figures 12.9, 12.10?
2012 Hexaconularia ningqiangensis Zheng, pp. 70–72, Figures 4.17 G–K
2014 Hexaconularia sichuanensis He and Yang; Steiner et al., p. 104, Figures 7.13–7.16
2017 ?Pseudooides prima Qian; Duan et al., pp. 7–9, Figures 5H–I
2018 Hexaconularia sichuanensis He and Yang; Xian et al., p. 228, Figure 3A
2018 Hexaconularia sichuanensis He and Yang; Zhang et al., p. 16, Figure 2.9
2020 Hexaconularia sichuanensis He and Yang; Han et al., p. 70, Figure 2C.
Holotype—KP-45-1 from the Kuanchuanpu Formation (Lower Cambrian, Fortunian Stage) in the Kuanchuanpu section in Ningqiang County, southern Shaanxi Province.
Paratypes—Eleven nearly complete and two partial specimens from the Kuanchuanpu Formation (Lower Cambrian, Fortunian Stage) in the Shizhonggou and Kuanchuanpu sections in Ningqiang County (SN021-K29-14, SN023-K30-189, SN047-K30-76, SN096-K44-131, SN096-K44-295, SN114-K32-79, SN116-K52-53, KP-45-1, KP-45-2, KP-45-3, KP-45-4, and KP-45-5) and the Zhangjiagou section in Xixiang County (XX99-284).
Diagnosis—Periderm with distinct apical and abapical regions. The abapical region has 10 faces, with five on each of the two major sides.
Occurrence—Cambrian Fortunian Stage; Kuanchuanpu Formation, northern Sichuan and southern Shaanxi provinces, China.
Description—Incomplete (broken) specimens ranging from 1.0 to 1.9 mm in length along the apical-abapical axis and from 1.2 to 2.0 mm in maximum width. The periderm is very thin and apparently non-laminar. The angle of adapertural expansion of the abapical region (abr) ranges from approximately 60° to 87° in the plane of its greatest width, with the two major sides of the abapical region meeting along a pair of marginal longitudinal ridges (mlr). The transverse cross-section of the aperture (ap) is narrowly lenticular (Figure 2B). The original apertural margin was not preserved.

FIGURE 2. Decimoconularia anisfacialis n. sp. from the Kuanchuanpu Formation (Cambrian Fortunian Stage) of southern Shaanxi Province, China. (A,F,L) KP-45-1, SN114-k32-79, and SN047-K30-76, respectively. (B–C,G–H) Apertural and apical views of (A,F), respectively. (D,E) Magnified views of (A), showing the trochoidal transverse ribs, adjacent faces separated by interfacial (longitudinal) furrows, and transverse ribs puckered along the interfacial (longitudinal) furrow. (I,J,K) Magnified views of (F,G,H) respectively, showing adjacent faces separated by interfacial (longitudinal) furrows, two sides separated by marginal longitudinal ridges, and the very narrow aperture. (M,N) Magnified views of (L), showing the concave interspace between adjacent transverse ribs, fine irregular wrinkles within the interfacial (longitudinal) furrow, and puckering of the transverse ribs within and along the interfacial (longitudinal) furrow. Abbreviations: abr=abapical region; ap=aperture; apr=apical region; cf=central face; if= interfacial (longitudinal) furrow; lfs=lateral faces; mf=marginal furrow; mfs=marginal faces; mlr=marginal longitudinal ridge; tr=transverse rib.
Each major side of the abapical region (abr) has one central face (cf), two lateral faces (lfs), and two marginal faces (mfs). The central face is approximately 2–3 times wider than the other four faces (Figure 2A, Figure 4), with the lateral faces being slightly wider than the marginal faces and the angle of adapertural expansion of the central face ranging from approximately 27° to 42° (Table 1). Shallow interfacial (longitudinal) furrows (if) separate the faces. All faces exhibit coarse, trochoidal (long.), nearly evenly spaced transverse ribs (tr) that terminate along the interfacial boundaries and the boundaries between the two major sides. The crests of the transverse ribs are sharp, smooth, and spaced 0.4–0.8 mm apart. The transverse ribs of the lateral and marginal faces are straight, while the transverse ribs of the central face are straight or bent slightly adapicalward near the interfacial furrows. The ends of the transverse ribs on a given face abut or are very slightly offset from those of the neighboring faces and puckered along the two interfacial (longitudinal) furrows between the central face and the two lateral faces. The concave interspace between adjacent transverse ribs is smooth or gently wrinkled.
The apical region (apr) is strongly compressed, with a deep terminal concavity and divided by a partly marginal furrow (mf). The external ornament of the apical region consists of irregular, fine transverse ribs or wrinkles, in some cases showing bifurcation.
Internal transverse septa were not observed, and there is no evidence of relic soft tissue.
Etymology—From the Latin roots anis (unequal) and facialis (faces), highlighting the characteristic differences in width between the faces of the adapical region of the periderm.
Comparisons—Unlike D. isofacialis, the 10 faces of which are essentially equal in width, each major side of D. anisfacialis n. sp. exhibits a broad central face flanked by two pairs of narrower, lateral and marginal faces (Figures 2A,F,L, 3, 4). Additionally, whereas the interfacial boundaries of D. isofacialis consist of a furrow (groove) in approximately the apertural half of the abapical region and a collinear ridge in approximately the apical half (Guo et al., 2021a), those of D. anisfacialis consist entirely of a furrow (Figures 2D,I,M). The transverse ribs of D. anisfacialis are mostly straight (Figures 2A,F,L), while those of D. isofacialis arch gently toward the aperture. The average distance between adjacent transverse ribs is also greater in D. anisfacialis than in D. isofacialis (Figures 2D,I,M). In both species, the apical angle of the major faces is approximately 75°; however, the apical angle of the central face is 19° in D. isofacialis and 34° in D. anisfacialis. The apical region of D. isofacialis is incomplete (Guo et al., 2021a); thus, it is not yet possible to make detailed comparisons of this portion of the periderm with that of D. anisfacialis.

FIGURE 3. Possible growth series of Decimoconularia anisfacialis n. sp. from the Kuanchuanpu Formation (Cambrian Fortunian Stage) of southern Shaanxi Province, China. (A–G) SN096-K44-295, KP-45-3, SN023-K30-131, KP-45-5, SN096-k44-131 (taken from Han et al., 2020, Figure 2C), KP-45-4, and KP-45-2.
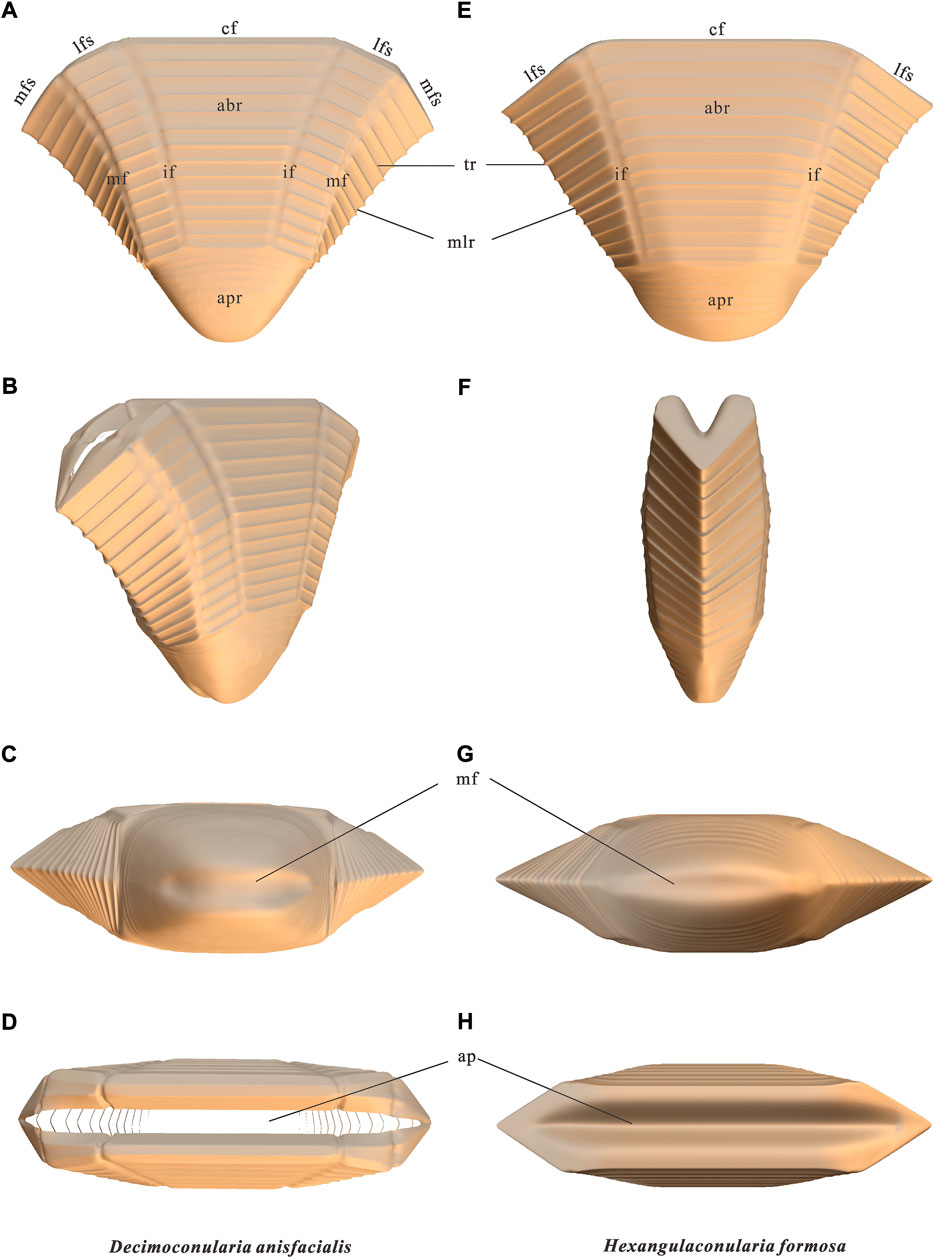
FIGURE 4. Three-dimensional reconstructions of Decimoconularia anisfacialis n. sp. and Hexangulaconularia formosa. (A,E,B,F,C,G,D,H) Side, oblique lateral, apical, and apertural views, respectively. The color of the reconstructions does not represent the actual color of the animals. Abbreviations: abr=abapical region; ap=aperture; apr=apical region; cf=central face; if=interfacial (longitudinal) furrow; lfs=lateral faces; mf=marginal furrow; mfs=marginal faces; mlr=marginal longitudinal ridge; tr=transverse rib.
Additional remarks—Some specimens previously identified as Hexangulaconularia formosa exhibit 10 distinct, mutually inclined, unequal faces and are similar to D. anisfacialis in all other characteristics described above (Table 2). These specimens, assigned herein to D. anisfacialis, are listed above in the synonymy.

TABLE 2. Principal morphological differences among Hexangulaconularia formosa, Decimoconularia anisfacialis, and Decimoconularia isofacialis.
The new Hexangulaconularia formosa and Decimoconularia anisfacialis sp. nov. material expands the known diversity and disparity of hexangulaconulariids and extends the stratigraphical range of Decimoconularia downward from Cambrian Stage 2 into the Fortunian Stage. Although beyond the scope of the present investigation, analysis of the phylogenetic relationships among the known species of hexangulaconulariids will be crucial in addressing the problem of the evolutionary origins of genera having six, 10, or 14 total faces (Figure 5). Finally, the functional morphology of hexangulaconulariids is poorly understood, owing in large part to the absence of soft tissue preservation. The slit-like narrow aperture of Septuconularia yanjiaheensis may have reduced the risk of sediment grains entering the gastrovascular cavity and spaces between the soft polyp and the inner surface of the periderm (Guo et al., 2020a; Song et al., 2022). Additionally, the puckering and wrinkling observed in Decimoconularia anisfacialis sp. nov. suggests that the periderm of this species may have been somewhat flexible.
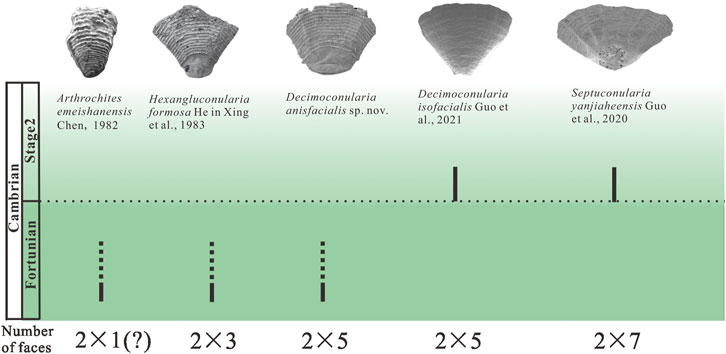
FIGURE 5. Stratigraphical distribution and face variation of Hexangulaconulariids in South China. The image of Arthrochites emeishanensis is modified from Figure 12 in Conway Morris and Chen (1992); Hexangluconularia formosa is modified from Figure 1 in Van Iten et al. (2010); Decimoconularia anisfacialis sp. nov. is specimen KP-45-1; Decimoconularia isofacialis and Septuconularia yanjiaheensis are modified from Figure 1in Guo et al. (2021a) and Figure 2 in Guo et al. (2020a), respectively.
Decimoconularia anisfacialis exhibits 10 faces rather than six. Thus, Decimoconularia can be interpreted as a minor but relatively long-ranging clade within the medusozoan family Hexangulaconulariidae. Moreover, following the ICZN, Hexangluaconularia He in Xing et al., 1983 has time priority over Hexaconularia in He and Yang 1986; consequently, the type genus of family Hexangulaconulariidae is changed herein from Hexaconularia to Hexangluaconularia, and the species name Hexaconularia sichuanensis is replaced by Hexangulaconularia formosa.
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding authors.
Conceptualization: JG, JH, and ZS; writing-original draft: ZS; writing-review and editing: HV, JG, JH, and ZZ; sample collection: YQ, JP, JS, YZ, and XH.
This research was funded by grants from the Natural Science Foundation of China (Nos. 42172016 to JG; 41890844 to ZZ, and JG; 41621003 to JH), the Strategic Priority Research Program of the Chinese Academy of Sciences (No. XDB26000000 to JH and JG), the Key Scientific and Technological Innovation Team Project in Shaanxi Province, the State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (Nos. 203106 to JG and 163107 to JH), and the National College Students’ innovation and entrepreneurship training program (No. S202210710197 to XH).
The authors of this paper thank the two reviewers (Peiyun Cong and Kun Liang) and the editor for their constructive comments and suggestions.
The authors declare that their research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Bengtson, S., Conway Morris, S., Cooper, B. J., Jell, P. A., and Runnegar, B. N. (1990). Early cambrian fossils from South Australia. Mem. Ass. Australas. Palaeontols. 9, 1–364.
Bouček, B. (1939). Conularida, in Handbuch der Paläozoologie 2A. Editor O. H. Schindewolf (Berlin: Gebruder Borntrager). A113–A131.
Brasier, M. D., and Singh, P. (1987). Microfossils and precambrian–cambrian boundary stratigraphy at maldeota, lesser himalaya. Geol. Mag. 124, 323–345. doi:10.1017/s0016756800016666
Chen, M. (1982). New knowledge of the fossil assemblages from maidiping section, Emei county, sichuan with reference to the sinian–cambrian boundary. Sci. Geol. Sin. 3, 253–263. (in Chinese with English abstract). doi:10.1007/BF01033890
Conway Morris, S., and Chen, M. (1992). Carinachitiids, hexaconulariids, and punctatus: Problematic metazoans from the early cambrian of south China. J. Paleontol. 66, 384–406. doi:10.1017/s0022336000033953
Duan, B., Dong, X., Porras, L., Vargas, K., Cunningham, J. A., and Donoghue, P. C. J. (2017). The early Cambrian fossil embryo Pseudooides is a direct-developing cnidarian, not an early ecdysozoan. Proc. R. Soc. B 284, 20172188. doi:10.1098/rspb.2017.2188
Guo, J., Han, J., Van Iten, H., Song, Z., Qiang, Y., Wang, W., et al. (2021a). A ten-faced hexangulaconulariid from cambrian stage 2 of south China. J. Paleontol. 95, 957–964. doi:10.1017/jpa.2021.25
Guo, J., Han, J., Van Iten, H., Wang, X., Qiang, Y., Song, Z., et al. (2020a). A fourteen-faced hexangulaconulariid from the early cambrian (stage 2) Yanjiahe formation, South China. J. Paleontol. 94, 45–55. doi:10.1017/jpa.2019.56
Guo, J., Li, G., Qiang, Y., Song, Z., Zhang, Z., Han, J., et al. (2021b). Watsonella crosbyi from the lower cambrian (terreneuvian, stage 2) Yanjiahe formation in three gorges area, South China. Palaeoworld 30, 1–19. doi:10.1016/j.palwor.2020.04.006
Guo, J., Qiang, Y., Han, J., Song, Z., Wang, W., and Zhang, Z. (2020b). Recent progress on small shelly fossils from the cambrian (terreneuvian) Yanjiahe formation in three gorges. Earth Sci. Front. 27, 104–115. (in Chinese with English abstract). doi:10.13745/j.esf.sf.2020.6.5
Han, J., Li, G., Wang, X., Yang, X., Guo, J., Sasaki, O., et al. (2018). Olivooides-like tube aperture in early Cambrian carinachitids (Medusozoa, Cnidaria). J. Paleontol. 92, 3–13. doi:10.1017/jpa.2017.10
He, T. (1987). Early Cambrian conulariids from the Yangtze platform and their early evolution. J. Chengdu Coll. Geol. 14, 7–18. (in Chinese with English abstract).
He, Y., and Yang, X. (1986). Early cambrian coelenterates from nanjiang, sichuan. Bull. Chengdu Inst. Geol. M. R. Chin. Acad. Geol. Sci. 7, 31–43. (in Chinese with English abstract).
Li, G., Steiner, M., Zhu, X., Yang, A., Wang, H., and Erdtmann, B. D. (2007). Early Cambrian metazoan fossil record of South China: Generic diversity and radiation patterns. Palaeogeogr. Palaeoclimatol. Palaeoecol. 254, 229–249. doi:10.1016/j.palaeo.2007.03.017
Liu, Y., Wang, Q., Shao, T., Dong, J., Tang, H., Zhang, Y., et al. (2016). Xixiang BiotaAnother rare scientific window of the Cambrian explosion. Acta Geol. Sin. 90, 1045–1046.
Moore, R. C., and Harrington, H. (1956). “Conulata,” in Treatise on Invertebrate Paleontology, Coelenterata. F54–F66.
Petersen, K. W. (1979). Development of coloniality in Hydrozoa, 11, 105–139. Biol. Syst. Colon. Org.,
Qian, Y. (1989). Early Cambrian small shelly fossils of China with special reference to the Precambrian–Cambrian boundary,” in Stratigraphy and Palaeontology of systematic boundaries in China, precambrian-cambrian boundary 2Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (Nanjing: Nanjing University Press), 1–341.
Shao, T., Tang, H., Liu, Y., Waloszek, D., Maas, A., and Zhang, H. (2018). Diversity of cnidarians and cycloneuralians in the fortunian (early cambrian) Kuanchuanpu Formation at Zhangjiagou, South China. J. Paleontol. 92, 115–129. doi:10.1017/jpa.2017.94
Shu, D. (2008). Cambrian explosion: Birth of tree of animals. Gondwana Res. 14, 219–240. doi:10.1016/j.gr.2007.08.004
Shu, D., Isozaki, Y., Zhang, X., Han, J., and Maruyama, S. (2014). Birth and early evolution of metazoans. Gondwana Res. 25, 884–895. doi:10.1016/j.gr.2013.09.001
Song, Z., Guo, J., Han, J., Van Iten, H., Qiang, Y., Peng, J., et al. (2022). A new species of Septuconularia (Hexangulaconulariidae, Cnidaria) from cambrian stage 2, South China. Acta Geol. Sin. 96, 757–765. doi:10.1111/1755-6724.14917
Steiner, M., Li, G., Qian, Y., Zhu, M., and Erdtmann, B. D. (2007). Neoproterozoic to early Cambrian small shelly fossil assemblages and a revised biostratigraphic correlation of the Yangtze Platform (China). Palaeogeogr. Palaeoclimatol. Palaeoecol. 254, 67–99. doi:10.1016/j.palaeo.2007.03.046
Steiner, M., Li, G., Qian, Y., and Zhu, M. (2004). Lower cambrian small shelly fossils of northern sichuan and southern Shaanxi (China), and their biostratigraphic importance. Geobios 37, 259–275. doi:10.1016/j.geobios.2003.08.001
Van Iten, H., Muir, L. A., Botting, J. P., Zhang, Y. D., and Li, J. P. (2013). Conulariids and sphenothallus (Cnidaria, Medusozoa) from the tonggao formation (lower ordovician, China). Bull. Geosci. 88, 713–722. doi:10.3140/bull.geosci.1400
Van Iten, H., Zhu, M., and Li, G. (2010). Redescription of Hexaconularia He and Yang, 1986 (lower Cambrian, South China): Implications for the affinities of conulariid-like small shelly fossils. Palaeontology 53, 191–199. doi:10.1111/j.1475-4983.2009.00925.x
Verrill, A. E. (1865). Classification of polyps extract condensed from synopsis of the polyps and corals of the north pacific exploring expedition under commodore C. Ringgold and captain john rogers, U. S. N. Commun. Essex Inst. 4, 145–152, doi:10.1080/00222936508679407
Xian, X., Zhang, H., Liu, Y., and Zhang, Y. (2019). Diverse radial symmetry among the Cambrian Fortunian fossil embryos from northern Sichuan and southern Shaanxi Provinces, South China. Palaeoworld 28, 225–233. doi:10.1016/j.palwor.2018.10.006
Xing, Y., Ding, Q., Luo, H., He, T., and Wang, Y. (1983). The Sinian–Cambrian boundary of China. Beijing: Bulletin of the Institute of Geology, Chinese Academy of Geological Sciences, 1–262. (in Chinese).
Yang, B., Steiner, M., Li, G., and Keupp, H. (2014). Terreneuvian small shelly faunas of East Yunnan (South China) and their biostratigraphic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 398, 28–58. doi:10.1016/j.palaeo.2013.07.003
Yang, X., He, Y., and Deng, S. (1983). On the sinian–cambrian boundary and the small shelly fossil assemblages in nanjiang area, sichuan. Bull. Chengdu Inst. Geol. Mineral Resour. Chin. Acad. Geol. Sci. 4, 91–110. (in Chinese with English abstract).
Zhang, H., Liu, Y., Qin, J., Zhang, Y., Shao, T., Zhang, T., et al. (2017). New materials of lower cambrian microfossils Hexaconularia sichuanensis from Zhangjiagou section in southern Shaanxi. Acta Palaeontol. Sin. 56, 1‒9. (in Chinese with English abstract). doi:10.19800/j.cnki.aps.2017.04.004
Zhang, H., Maas, A., and Waloszek, D. (2018). New material of scalidophoran worms in orsten-type preservation from the cambrian fortunian stage of south China. J. Paleontol. 92, 14–25. doi:10.1017/jpa.2017.39
Zheng, Y. (2012). The study of microfossils in the lower cambrian from the southern Shaanxi province and northern sichuan province. in Doctoral dissertation. Xi’an: Chang’an University, 1–117. (in Chinese with English abstract).
Keywords: small shelly fossils, Medusozoa, Hexangulaconulariids, Lower Cambrian, South China
Citation: Song Z, Guo J, Han J, Van Iten H, Qiang Y, Peng J, Sun J, Zheng Y, Huang X and Zhang Z (2023) A new species of Decimoconularia (Cnidaria, Medusozoa) from the Lower Cambrian of South China. Front. Earth Sci. 10:1048800. doi: 10.3389/feart.2022.1048800
Received: 20 September 2022; Accepted: 07 November 2022;
Published: 13 January 2023.
Edited by:
Ben Yang, Chinese Academy of Geological Sciences (CAGS), ChinaReviewed by:
Peiyun Cong, Yunnan University, ChinaCopyright © 2023 Song, Guo, Han, Van Iten, Qiang, Peng, Sun, Zheng, Huang and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Junfeng Guo, anVuZmVuZ2dAY2hkLmVkdS5jbg==; Heyo Van Iten, dmFuaXRlbkBoYW5vdmVyLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.