 Xiang Li
Xiang Li Ying Zhang2
Ying Zhang2 Yuewu Sun
Yuewu Sun- 1College of Earth Sciences, Jilin University, Changchun, China
- 2International Centre for Geoscience Research and Education in Northeast Asia, Jilin University, Changchun, China
- 3Research Center of Palaeontology and Stratigraphy, Jilin University, Changchun, China
- 4Key-Lab for Oil Shale and Paragenetic Minerals of Jilin Province, Changchun, China
The Middle–Late Triassic climates have attracted the attention of paleontological and geological scientists for the Carnian pluvial event in the early Late Triassic. The event is well-documented in the pelagic and epi-continental marine deposits of the Tethys, Gondwana, and Laurasia. However, inland terrestrial deposits are less frequently depicted, with high-resolution palynological biostratigraphy constraints. In this study, we report the palynological records from the YC8-1 and YC7-3 sub-members of the Yunmeng profile in the Ordos Basin, China, where the YC7-3 was dated at 236.0–234.1 Ma. Two palynological assemblages were recognized and named the Lundbladispora watangensis–Taeniaesporites combinatus assemblage and Lundbladispora communis–Discisporites granulus assemblage for the YC8-1 and YC7-3 sub-members, respectively. Their ages were determined to be the latest Ladinian and early Carnian, respectively, for their stratigraphic correlations with the global boundary stratotype section and point (GSSP) of the base of Carnian in Europe and additional co-occurring floras with condonts in the Upper Triassic of South China. The coexistence of Cyathidites minor (Couper, 1953), Dictyophyllidites harrisii (Couper, 1958), Apiculatisporis bulliensis (Helby ex De Jersey, 1972), Aratrisporites xiangxiensis (Li and Shang, 2011), Piceaepollenites omoriciformis (Bolkh.) (Xu and Zhang, 1984), Podocarpidites ornatus (Pocock, 1962), Discisporites granulus (Zhang, 1984), and Classopollis (Pflug, 1953) is equivalent to the Carnian palynostratigraphic criterion reported in the North China palynofloristic realm. Vegetational changes, especially those occurring at the boundary between Ladinian and Carnian, account for approximately 70% of ferns and over 30% of gymnosperms lost. These were discovered and attributed to the strong seasonal arid climate, indicated by the emergence of Cheirolepidiaceae and Pinuspollenites. We thus know that the climate during the latest Ladinian and early Carnian was “hot house” with seasonal aridity. In addition, three strong monsoonal pluvial pulses were signaled by the humidity index of lowland plants. The present study will enable a better understanding the Carnian pluvial event in the Late Triassic inland basin.
1 Introduction
While the Triassic period (201.3–252 Ma; Benton and Newell, 2014) might be “hot house” period, as evidenced by ice-free poles covered by green forests (Harris, 1937; Vasilevskaya, 1972; Smith, 1974; Harland, 1997; Cuneo et al., 2003; Strullu-Derrien et al., 2012), eolian deserts were widespread in tropical and subtropical areas during the Early and Middle Triassic, with green vegetation present only along permanent rivers (termed “gallery forest”; Zhu et al., 2020; Shi et al., 2021). The most important plate tectonic event occurred in the tropical Tethys realm, represented by the closure of the Paleo-Tethys and opening of the Neo-Tethys during the Triassic period (Boucot et al., 2009; Domeier and Torsvic, 2014). The climate of the supercontinent Pangea was characterized by a mega-monsoonal circulation which reached its maximum volume in the Triassic (Kutzbach and Gallimore, 1989; Parrish, 1993).
Meanwhile, in the circum-Paleo-Tethys area during the Early and Middle Triassic, the climate might have been like the Mediterranean of today. Hot and dry summers might have eliminated most plant life from the winter ponds, which were inhabited by small hydrophytic quillworts and surrounded by lycopod shrubs and small conifers (Retallack, 1997). Thence, the Late Triassic climate changed to pluvial, with green vegetation and coal deposits overlying the former red continental sediments (Boucot et al., 2009). Frequent floods may have led to the formation of black organic-rich shale in lakes and the Panthalassa seacoast (Hornung and Brandner, 2005; Liu et al., 2022)—a notable characteristic of the Carnian Pluvial Event (CPE) (Simms and Ruffell, 1990). The palynoflora, both in the tropical Tethys (Dubiel et al., 1991; Shi et al., 2009; Preto et al., 2010; Wang et al., 2012; Mueller et al., 2016) and boreal areas (Mueller et al., 2015), and even in the middle latitudes of Gondwana (Césari and Colombi, 2016; Colombi et al., 2021), recorded vegetation and climate changes in the seasonal paleoclimate’s rainfall during the CPE. As it coincided with negative carbon isotopic excursion (Dal Corso et al., 2012; Dal Corso et al., 2015), the CPE has been attributed to the release of CO2 from large volcanic eruptions (Dal Corso et al., 2012; Dal Corso et al., 2015; Mueller et al., 2016).
This study presents the results of a palynological investigation of lacustrine deposits that were calibrated as early Carnian by U-Pb isotopic dating of bentonites in the Ordos Basin, China (Figure 1), in order to shed new light on our understanding of the CPE in the Late Triassic inland basin.
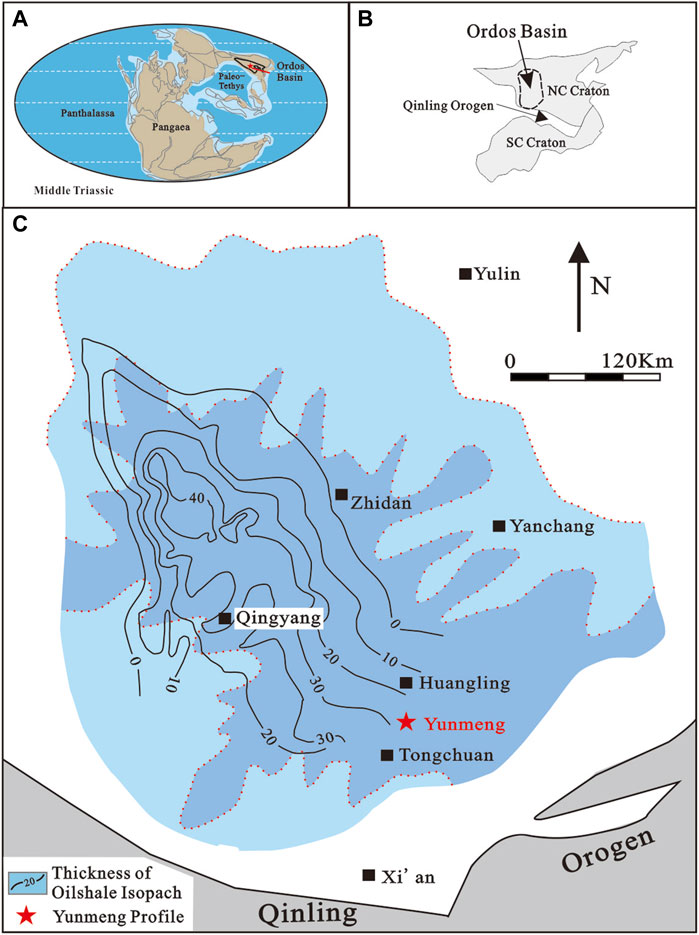
FIGURE 1. Geographical and geological location of the Yunmeng Profile in the Ordos Basin (after Sun et al., 2020). (A) Reconstructed topography in late Middle Triassic (after Zhang et al., 2019). (B) Geographic map of China showing location of the Ordos Basin; NC Craton, North China Craton; SC Craton, South China Craton. (C) Paleogeographic map of YC7 showing the isopach of oil-shale.
2 Geological setting
Located in the eastern realm of the Paleo-Tethys, the Triassic Ordos Basin was situated at 18.3–25.4°N (Ma et al., 1993) in the North China Craton. The basin was derived from an epi-continental sea that developed during the Late Carboniferous to Middle Triassic. The Ordos became a foreland basin due to the collision of the South and the North China Cratons along the Mianlue suture from approximately 250 Ma; however, recent detrital zircon U-Pb isotope dating of the early Late Triassic Yanchang Formation shows that the protoliths were sourced only from the North China Craton and not the South China Craton and the Qinling Orogenic Belt (Xie and Heller, 2013). The deposition of Triassic sediments in the Ordos Basin can be divided into two cycles (IGCAGS, 1980; PCOC, 1992): 1) the Lower and Middle Triassic cycles, including the Liujiagou, Heshanggou, and Ermaying formations, which are widespread throughout the entire North China Craton; 2) the Upper Triassic cycle composed of the Yanchang Formation (sensu lato) comprising alluvial, fluvial, deltaic, and lacustrine sedimentary rocks, approximately 1,000–1,300 m in thickness. The Yanchang Formation (Pan, 1934) or Group (ECSDC, 2000) can be divided into five members (Y1–Y5) (IGCAGS, 1980) and 10 oil layers or members (YC1–YC10), based on the depositional sequences (S1–S5), regional and local indicator beds (K0–K9), and bentonite beds (B0–B6) (PCOC, 1992; Pang et al., 2010; Yang et al., 2017; Deng et al., 2018; Zhang et al., 2019; Sun et al., 2020). The Chang 7 oil layer (member) of the Yanchang Formation (YC7) is one of the best layers of source rocks in the Ordos Basin (PCOC, 1992), shale oil being the most important fossil energy source in China and elsewhere (Jin et al., 2019; Liu et al., 2022). YC7 shale oil is deposited in deep or semi-deep lacustrine environments when the basin extends to its maximum; known as “Zhangjiatan Shale,” it is also a regional marker for stratigraphic correlation. The shale has also been proposed for the division of Triassic strata and their geological age. Its presence was assigned to the uppermost Tongchuan Formation (IGCAGS, 1980; Tong et al., 2019) or the lowest Yongping Formation (Li et al., 2016), while its age was assigned to either the Middle (IGCAGS, 1980; Deng et al., 2018) or Late Triassic (Li et al., 2016; Tong et al., 2019; Sun et al., 2020). However, the Yanchang Formation (sensu lato) is widely used (Bureau of Geology and Mineral Resources of Shaanxi Province, 1998) in geological references (Table 1). The formation conformably overlies the Middle Triassic Ermaying Formation (or Zhifang Formation), which is characterized by purple or grayish-red siltstones and mudstones intercalated within grayish and yellowish-green sandstones.
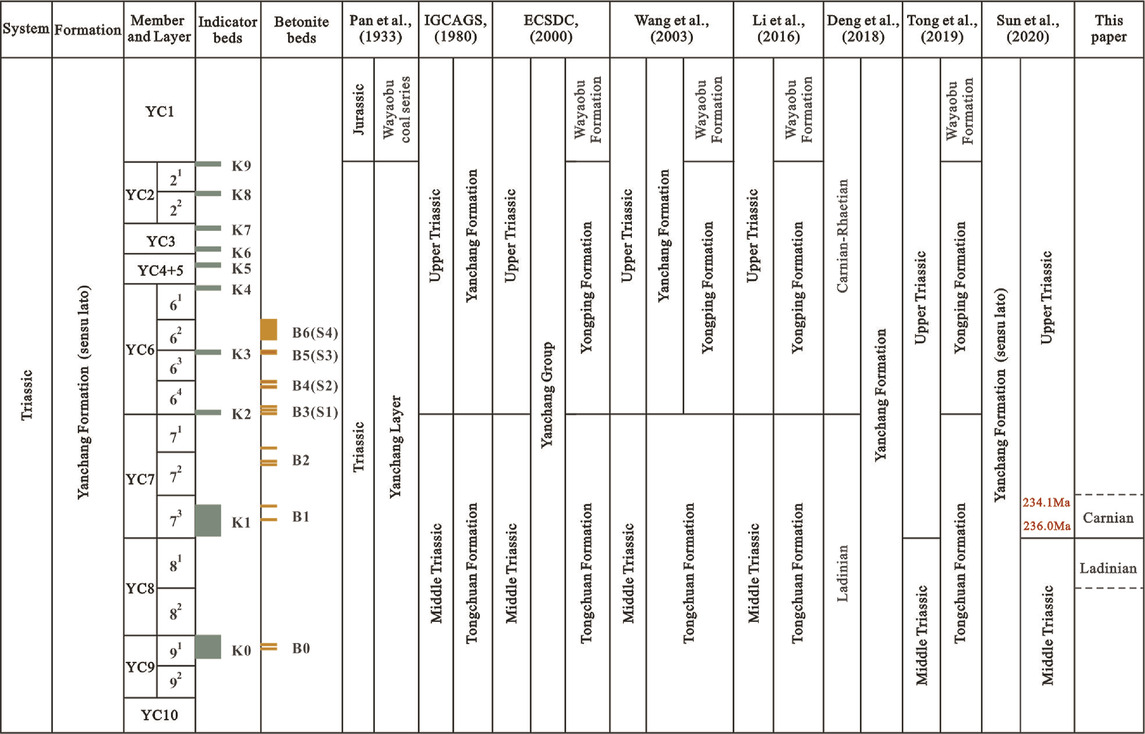
TABLE 1. Historical division of the Yanchang Formation (sensu lato) in Ordos Basin, China.
3 Materials and methods
Palynological samples were collected from the Yunmeng profile (GPS: N 35°15′58.87″, E 109°13′50.7″, H 1,298.45 m) which was manually outcropped at Ruzhihe Village in Yunmeng Town, Yijun County, Tongchuan City, Shaanxi Province, northwestern China (Figure 1). YC8 and YC7 on the profile are construed as Middle and Late Triassic in age, respectively (Ji and Meng, 2006; Sun et al., 2020). The total thickness of the measured Yunmeng profile is >30 m (Supplementary Figure S1). Some 27 palynological samples were collected from the black shale layers of YC8-1 (9.01 m thick and dominated by gray fine-grained sandstone and siltstone), and 60 samples were collected from YC7-3 (21.29 m thick and mainly composed of dark gray, brown oil-shale, shale, and thin-layered bentonite).
Each 50 g sample was broken into pieces <1.0 mm in diameter and treated with HCl (30%) for 24 h and HF (36%) for 2 days. ZnCl-mixed KI heavy liquid (2.2 g/cm3) and an 8 µm sieve were used to separate the organic residue from minerals and gather the palynomorphs. All samples, slides, and stubs numbered with the prefix STYM were housed in the Research Center of Paleontology and Stratigraphy, Jilin University, China. Details of the samples are provided in Supplementary Appendix S1.
4 Results
4.1 Palynological assemblages
Among the 87 palynological samples collected from the Yunmeng profile, only 15 samples containing over 100 grains of pollen and spores were quantitatively analyzed. A total of 45 species assigned to 24 genera (Figures 2–4) were described; their botanical affinity is shown in Supplementary Appendix S2. Their relative abundance was calculated using TiLIA software (2.1.1). Two palynological assemblages were recognized using CONISS within TiLIA (Grimm, 1991–2016) in ascending order (Figure 5).
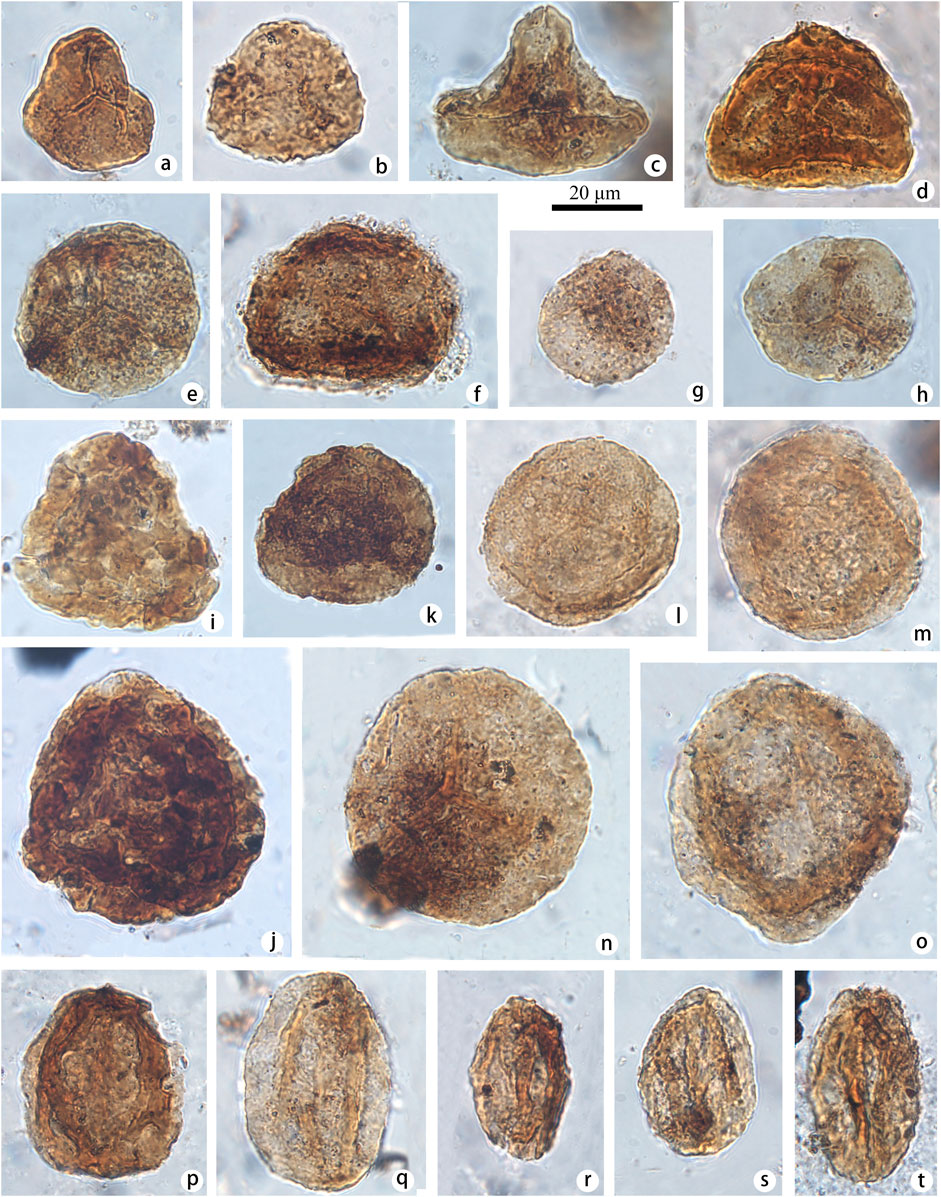
FIGURE 2. Selected spores and pollen identified at the Yunmeng profile (taxon name followed by sample number and position in England finder in parentheses). (A) Cyathidites minor (Couper, 1953) (a-STYM94, J11-2); (B) Converrucosisporites decoratus (Shang, 2011) (b-STYM48, R17-4); (C) Dictyophyllidites harrisii (Couper, 1958) (c-STYM48, N34-3); (D) Duplexisporites rotundatus (Shugaevskaya, 1969) (d-STYM01, N41-4); (E) Verrucosisporites rotundus (Singh, 1964) (e-STYM03, N23-3); (F) Kraeuselisporites spinosus (Jansonius, 1962) (f-STYM16, F20-1); (G) Apiculatisporites bulliensis (Helby et De Jersey, 1972) (g-STYM94, R8-3); (H) Lundbladispora playfordi (Balme, 1963) (h-STYM12, I26-1); (I) Asseretospora gyrata (Playford et Dettmann) (Schuurman, 1977) (i-STYM16, B21-4); (J) Asseretospora curvata (Qu, 1980) (j-STYM28, J12-2); (K) Lundbladispora watangensis (Qu, 1984) (k-STYM12, R36-2); (L, M) Lundbladispora subornata (Ouyang and Li, 1980) (l-STYM01, O45-2; m-STYM01, U25-4); (N) Lundbladispora communis (Ouyang and Li, 1980) (n-STYM94, K17-3); (O) Lundbladispora sp. (o-STYM01, M26-4); (P) Aratrisporites exiguous (Qu, 1984) (p-STYM28, M31-2); (Q) Aratrisporites tenuispinosus (Playford, 1965) (q-STYM01, N37-1); (R) Aratrisporites coryliseminis (Klaus, 1960) (r-STYM01, N37-1); (S, T) Aratrisporites granulates (Klaus) (Playford and Dettmann, 1965) (s-STYM21, R45-4, t-STYM01, T16-2).
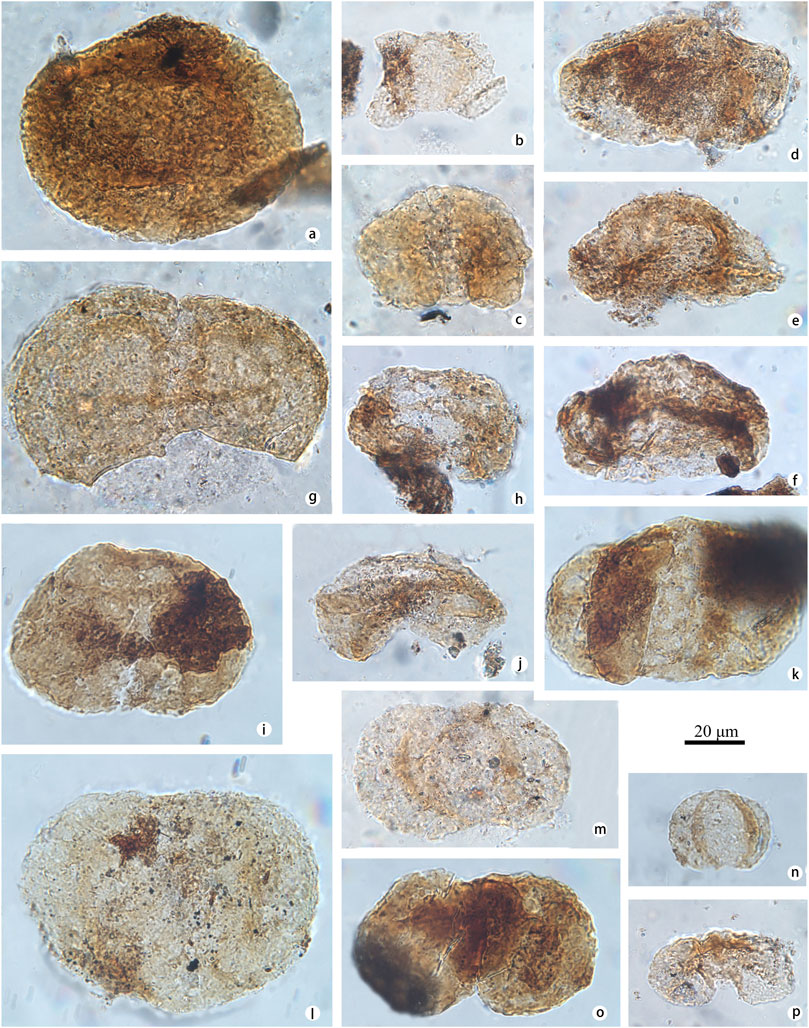
FIGURE 3. Selected pollen identified at the Yunmeng profile (taxon name followed by sample number and position in England finder in parentheses). (A) Plicatipollenites indicus (Lele, 1964) (a-STYM01, J46-3); (B) Striatoabieites sp. 1 (b-STYM21, L47-4); (C) Striatoabieites brickii (Sedova, 1956) (c-STYM01, L19-3); (D–F) Striatoabieites duivenii (Jansonius) (Hart, 1964), (d-STYM21, N41-3; e-STYM16, S14-1; f-STYM28, S23-1); (G) Taeniaesporites combinatus (Qu and Wang, 1990) (g-STYM01, M18-4); (H) Taeniaesporites albertae (Jansonius, 1962) (h-STYM28, F39-1); (I) Taeniaesporites junior (Klaus, 1960) Wu, 1982 (i-STYM88, A42-1); (J) Taeniaesporites leptocorpus (Qu, 1984) (j-STYM21, L45-1); (K) Protohaploxypinus samoilovichii (Jansonius) (Hart, 1964) (k-STYM48, T13-3); (L) Protopinus subluteus Bolkh, 1956 (l-STYM21, O34-4); (M) Alisporites nuthallensis (Clarke, 1965) (m-STYM01, K28-4); (N) Alisporites parvus (De Jersey, 1962) (n-STYM82, E21-4); (O) Platysaccus luteus (Bolkh.) (Li and Shang, 1980) (o-STYM88, I41-3); (P) Platysaccus queenslundi (De Jersey, 1962) (p-STYM21, L44-2).
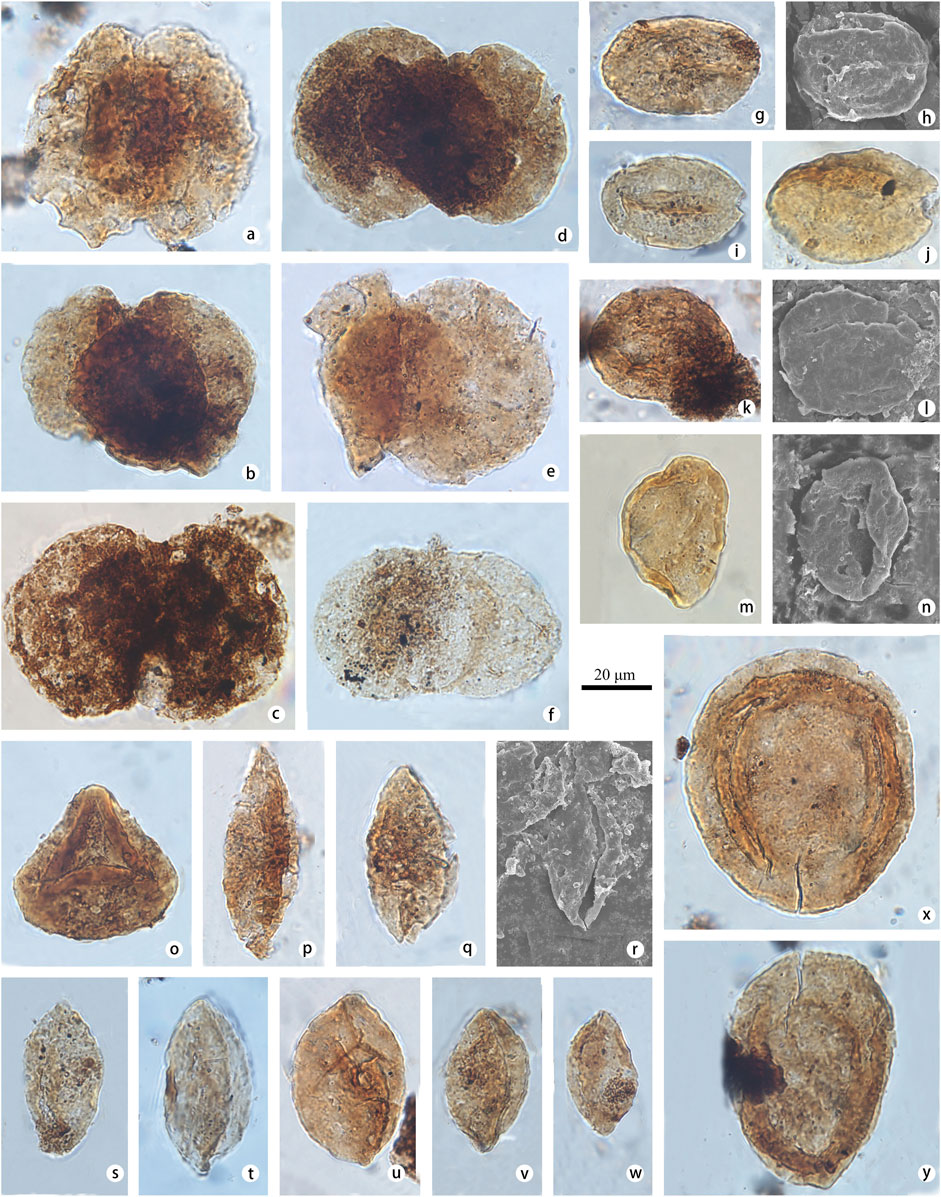
FIGURE 4. Selected pollen identified at the Yunmeng profile (taxon name is followed by sample number and position in England finder or stub number in parentheses). (A) Podocarpidites paulus (Bolkh.) (Xu and Zhang, 1980) (a-STYM16, P7-2); (B, C) Podocarpidites ornatus (Pocock, 1962) (b-STYM94, M34-4; c-STYM97, M34-1); (D, E) Podocarpidites unicus (Bolkh.) (Pocock, 1970) (d-STYM-88, N35-2; e-STYM16, R42-1); (F) Podocarpidites multicinus (Bolkh.) (Pocock, 1970) (f-STYM30, I29-4); (G–J) Ovalipollis ovalis (Krutzsch, 1955) (g, h-STYM01, O23-1, Stub-01; i-STYM26, P36-1; j-STYM01, N33-1); (K, L) Ovalipollis breviformis (Krutzsch, 1955) (k, l-STYM21, Q37-3, Stub-21); (M, N) Chasmatosporites apertus (Rogalska) (Nilsson, 1958) (m, n-STYM12, T20-3, Stub-12); (O) Duplicisporites granulatus (Leschik, 1955) (o-STYM94, L37-4); (P–R) Cycadopites typicus (Maljavkina) (Pocock, 1970) (p-STYM48, D46-1; q, r-STYM16, T23-2, Stub-16); (S) Cycadopites adjectus (De Jersey, 1964) (s-STYM27, I34-2); (T) Cycadopites tivoliensis (De Jersey, 1971) (t-STYM28, M47-4); (U–W) Ginkgocycadophytus spp. (u-STYM74, R34-1; v-STYM97, J31-2; w-STYM90, P16-2); (X, Y) Discisporites spp. (x-STYM74, D44-2; y-STYM94, G38-2).
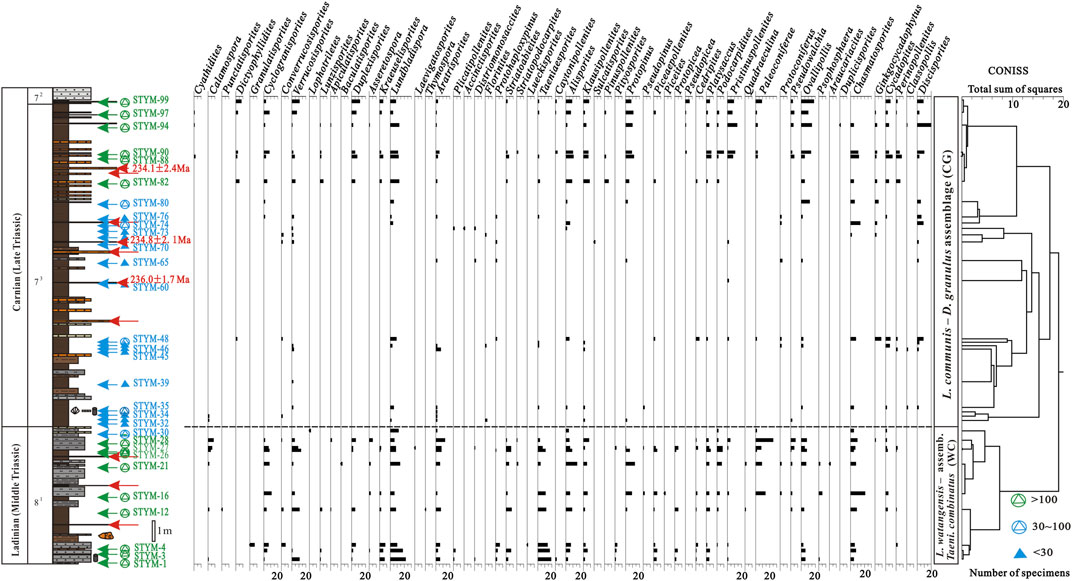
FIGURE 5. Stratigraphy, lithology, productive sample position, and relative sporo-pollen abundances at the Yunmeng profile, Ordos Basin, China.
4.1.1 Lundbladispora watangensis–Taeniaesporites combinatus assemblage (WC)
This palynological assemblage comprised 10 samples collected from the YC8-1 sub-member in the Yunmeng profile, from which 1,092 specimens were identified. The taxa in this assemblage comprised 121 species of 47 genera, including spores of 54 species belonging to 17 genera and pollens of 67 species belonging to 30 genera, respectively (Supplementary Appendix S1).
The fern spores (21%–48%) in the WC palynological assemblage mainly belonged to Lundbladispora (5%–20%), Aratrisporites (2%–10%), Verrucosisporites (0%–11%), Cyclogranisporites (1%–8%), Duplexisporites (0%–6%), and Calamospora (0%–6%). The dominant species were L. watangensis (0%–6%), L. playfordi (0%–5%), Aratrisporites tenuispinosus (0%–6%), A. granulates (0%–3%), Verrucosisporites krempii (0%–6%), V. scitulus (0%–5%), Duplexisporites rotundatus (0%–6%), Kraeuselisporites apiculatus (0%–5%), and Asseretospora gyrata (0%–2%). The pollen (52%–79%) in the WC assemblage mainly comprised Taeniaesporites (2%–18%), Chasmatosporites (4%–16%), and Alisporites (3%–13%), as well as Platysaccus (1%–5%), Striatoabieites (0%–7%), Podocarpidites (0%–7%), Ovalipollis (0%–7%), Klausipollenites (0%–6%), Piceaepollenites (0%–4%), Cycadopites (0%–3%), and Cedripites (0%–3%). The most important species of pollen in the WC assemblage were Taeniaesporites combinatus (0%–9%), T. watangensis (0%–1%), Alisporites aequalis (0%–5%), Platysaccus proximus (0%–3%), P. queenslundi (0%–2%), Striatoabieites duivenii (0%–2%), Ovalipollis ovalis (0%–4%), O. breviformis (0%–3%), Podocarpidites paulus (0%–2%), and Cycadopites typicus (0%–2%).
4.1.2 Lundbladispora communis–Discisporites granulus assemblage (CG)
This palynological assemblage comprised 20 samples collected from the YC7-3 sub-member in the Yunmeng profile, from which 1,083 specimens were identified. The taxa in this assemblage were less diversified, with only 76 species of 45 genera in total, including spores of 22 species belonging to 13 genera and pollen of 54 species belonging to 32 genera (Supplementary Appendix S1).
Fern spores (20%–39%) in the CG palynological assemblage mainly consisted of Lundbladispora (4%–12%), Cyclogranisporites (2%–8%), Dictyophyllidites (0%–5%), Aratrisporites (1%–3%), Kraeuselisporites (0%–3%), Lunzisporites (0%–5%), and Cyathidites (0%–1%). Common species identified were Lundbladispora communis (0%–4%), L. playfordi (1%–4%), Dictyophyllidites harrisii (0%–5%), Duplexisporites rotundatus (0%–6%), Lunzisporites sparsus (0%–5%), and Kraeuselisporites spinosus (0%–3%). Pollen (52%–79%) in the CG assemblage mainly comprised Ovalipollis (3%–13%), Discisporites (0%–17%), Protopinus (5%–11%), and Pristinuspollenites (0%–12%). Alisporites (2%–8%), Klausipollenites (1%–8%), Podocarpidites (0%–7%), Platysaccus (2%–5%), and Cycadopites (3%–7%) were common. The most important pollens (with stratigraphic significance) in the CG assemblage were Discisporites granulus (0%–4%), Cycadopites typicus (1%–3%), Ovalipollis ovalis (1%–3%), Klausipollenites schaubergeri (0%–3%), and P. queenslundi (0%–2%).
4.2 Comparison of the palynological assemblages in the Ordos Basin
Qu (1980) and Liu et al. (1981) reported palynological assemblages from the upper member of the Tongchuan Formation on the Qishuihe profile near the Yunmeng profile. These were characterized by abundant fern spores (66.8%–77%) represented by Verrucosisporites (8.3%–16.7%), Punctatisporites (33.2%), and Todisporites (46.5%) (Qu, 1980; Liu et al., 1981). Aratrisporites are normally rare in the Qishuihe profile, whereas they are abundant (13%–29%) in the Qingyang profile (Liu et al., 1981). Miao et al. (1984) also recorded the palynological assemblage from the upper member of the Tongchuan Formation in the Guluo and Kongyu outcrops in Hongdong, Shanxi Province, North China, located in the southeastern Ordos Basin. The assemblage is dominated by fern spores (60.4%–78.0%) including Punctatisporites (42.8%–65.1%), Calamospora (4.0%–6.3%), and Verrucosisporites (2.0%–7.0%). The pollen (22.0%–39.6%) in the assemblage mainly comprises Piceaepollenites (7.0%–11.6%), Chordasporites (1.6%–8.7%), and Psophosphaera (2.5%–8.0%) (Miao et al., 1984). As the upper member of the Tongchuan Formation includes YC8 and YC7, researchers in the 1980s did not realize the palynological difference between the two members. Sun et al. (1995) reported that the Anisian-Ladinian palynological assemblage in the northern China palynofloristic realm could not be divided into two sub-assemblages according to their sporo-pollen sampling horizons and named the Punctatisporites–Aratrisporites–Taeniaesporites–Parataeniaesporites assemblages for both the Ermaying and Tongchuan formations in the Ordos Basin (Sun et al., 1995). Furthermore, the spores in the Yongping and Wayaobu formations (equivalent to members YC7 to YC1) on the Tuweihe profile in the northern Ordos Basin accounted for 56.2%–61.5% of the total grains (Wang et al., 2003). Therefore, it seems difficult to determine whether the palynological characteristics of the YC7 member are related to the lower layers (from YC8 to YC10) or upper member (from YC6 to YC1).
Finally, based on the samples from 12 boreholes (including X17, X30, X36, X40, X43, and X44) in the Ordos Basin, located 136 km westward of the Yunmeng profile, the palynological assemblages for the YC8 and YC7 members were separated and named Aratrisporites–Punctatisporites and Asseretospora–Walchiites, respectively (Ji and Meng, 2006). The Aratrisporites–Punctatisporites assemblage is characterized by abundant fern spores (52.5%–88.7%), including Aratrisporites (6.8%–66.2%), Punctatisporites (4.0%–50.4%), Osmundacidites (3.5%–25.6%), Verrucosisporites (2.3%–18.0%), and gymnosperm pollen (11.3%–45.7%) represented by Alisporites (4.0%–14.1%), Abietineaepollenites (4.8%–10.2%), Pinuspollenites + Piceaepollenites (2.2%–13.3%), Walchiites (2.2%–12.0%), and Cycadopites (1.0%–6.0%). The Asseretospora–Walchiites assemblage comprises fern spores (15.9%–44.7%) represented by Asseretospora (5.2%–20.2%) and Aratrisporites (4.0%–15.8%), and gymnosperm pollen (53.3%–84.1%) including Abietineaepollenites + Pinuspollenites (8.4%–34.0%), Walchiites (4.0%–22.2%), and Alisporites (4.7%–15.6%).
The present WC and CG assemblages are comparable to the Aratrisporites–Punctatisporites and Asseretospora–Walchiites assemblages, respectively. The WC assemblage from the YC8-1 sub-member (i.e., the upper part of YC8 member) is part of the Aratrisporites–Punctatisporites assemblage from the whole YC8 member, while the CG assemblage from the YC7-3 sub-member (the lower part of the YC7 member) is part of the Asseretospora–Walchiites assemblage. This enables researchers to see all characteristics and the details of the palynological transition from the uppermost Ladinian to the lowest Carnian. The percentage of spores decreases from the YC8 to YC7, with an obvious excursion at the boundary between YC8 and YC7. While this might result from the withdrawal of lake shore vegetation accompanying the lake’s widening and deepening, the number of spore species decreased greatly, especially Lundbladispora and Aratrisporites. Deng et al. (2018) summarized the palynological assemblages of the Yanchang Formation (sensu lato) in the Ordos Basin, named the Asseretospora–Apiculatisporis–Chordasporites (YC1–YC6) assemblage and Punctatisporites–Aratrisporites–Verrucosisporites (YC7–YC10) assemblage. Spore abundance varied between 30% and 70% in each, and, in many outcrops, the Aratrisporites content was as high Punctatisporites and Verrucosisporites in the Punctatisporites–Aratrisporites–Verrucosisporites assemblage, including the Qishuihe profile (Deng et al., 2018).
Three palynological assemblage zones were recently recognized based on 11 samples from the Triassic Tanzhuang and Anyao formations in the Jiyuan Basin (which might comprise the Ordos Basin), Henan Province, China—named the Paleoconiferus–Cyclogranisporites–Rotundipollis, Cyclogranisporites–Osmundacidites–Punctatisporites, and Pseudopicea–Paleoconiferus–Protoconiferus assemblage zones (in ascending order) (Lu et al., 2021). The first and last assemblages were dominated by gymnosperm pollen (52.3% and 47.6%, respectively) represented by Paleoconiferus and Pseudopicea, with fern spores mainly including Cyclogranisporites, Osmundacidites, and Punctatisporites. However, the second assemblage was dominated by algae (56.5%), fern spores (33.3%), and gymnosperm pollen (10.2%). This was also observed in the present WC and CG assemblages, which were dominated by gymnosperm pollen, along with algae at the horizon of the CG assemblage.
5 Discussion
5.1 Geological age
5.1.1 Isotopic dating
Although the geological age of YC7-3 on the Yunmeng profile is assigned to the Carnian based on U-Pb isotopic dating of three continuous rhyolitic tuff layers and ages 236.0 ± 1.7 Ma, 234.8 ± 2.1 Ma, and 234.1 ± 2.4 Ma have been obtained in ascending order (Sun et al., 2020), the latest isotopic dating near the Yunmeng profile showed the ages to be much older (240.9 ± 0.9 Ma for the detrital zircons from the tuffaceous sandstone in Mazhuang; Zhao et al., 2020). The old group of detrital zircons could not be regarded as of eruptive and depositional ages (Fedo et al., 2003), and they were also detected as inherited zircons in sample STYMD-62 (Sun et al., 2020). Some inherited zircons were dated older in the Yunmeng profile; however, considering the number of concording zircons and the stratigraphic sequence, the isotopic dating results indicated that YC7 in Ordos Basin was deposited at 234.1–236.0 Ma (Sun et al., 2020).
5.1.2 Palynological biostratigraphy
Unlike in South China and Europe, the Triassic successions in the Ordos Basin are terrestrial without any marine interlayers; therefore, palynology is one of the major fossils indexes used for stratigraphic correlation (Liu et al., 1981; Shang, 2011). At the GSSP section of the Carnian, the palynological criteria of the basal Carnian are the first observations of Vallasporites ignacii, Patinasporites densus, and Aulisporites cf. A. astigmosus, together with Duplicisporites verrucosus and Camerosporites secatus, “Lueckisporites” cf. singhii, which are auxiliary indicators of the Carnian (Mietto et al., 2012). Because the GSSP of the Carnian is indicated by Daxatina canadensis, it is lower than the traditional base of the Carnian (including Cordevolian, Julian, and Tuvalian) in the Alps and circum-Tethys area, which was marked by the Trachyceras aon zone (Visscher and Brugman, 1981). Meanwhile, C. secatus was recorded in Yunnan and Guizhou, China, where the bases of the palynomorph-bearing strata are Carnian-aged marine limestones, as evidenced by the presence of conodonts (Neogondolella polygnathiformis zone; Yang et al., 1995). Thus, this praecolpate pollen, together with other palynomorphs in the Banan and Sanqiao formations in Guizhou Province, China, are late Tuvalian in age (Shang, 2011). The present WC and CG assemblages in the Yunmeng profile in the Ordos Basin shared 17 known species with those in the Banan and Sanqiao formations, Guizhou Province, China, including Calamospora nathorstii, C. padata, Punctatisporites crassexinis, Dictyophyllidites harrisii, Verrucosisporites congestus, V. morulae, Lundbladispora playfordi, Aratrisporites scabratus, Lueckisporites triassicus, Taeniaesporites albertae, T. junior, T. leptocorpus, Klausipollenites decipiens, Platysaccus proximus, P. queenslandi, Ovalipollis ovalis, and Cycadopites tivoliensis. In addition to these species, 22 have been recorded in Late Triassic deposits in Eurasia, South America, North America, Australia, and Antarctica (Halle, 1908; Couper, 1953; Couper, 1958; Leschik, 1955; Nilsson, 1958; Klaus, 1960; De Jersey, 1962; De Jersey, 1964; De Jersey, 1972; Bharadwaj et Singh, 1964; Mädler, 1964; Clarke, 1965; Gair et al., 1965; Playford, 1965; Playford and Dettmann, 1965; Scheuring, 1970; Arjang, 1975; Fisher and Bujak, 1975; Schuurman, 1977): Cyathidites minor, Verrucosisporites krempii, V. remyanus, Lunzisporites lunzensis, L. sparsus, Asseretospora gyrate, Aratisporites coryliseminis, A. granulates, A. coryliseminis, A. strigosus, A. tenuispinosus, A. aequalis, A. circulicorpus, A. nuthallensis, A. parvus, Ovalipollis grebeae, O. minimus, Duplicisporites granulatus, Chasmatosporites apertus, Cycadopites adjectus, C. reticulatus, and Perinopollenites elatoides. However, six species—including Cyclogranisporites multigranus, Apiculatisporis bulliensis, Kraeuselisporites apiculatus, K. spinosus, Plicatipollenites indicus, and K. schaubergeri—from the Permian or even the Carboniferous of France, England, Canada, Australia, and India (Potonie and Klaus, 1954; Jansonius, 1962; Lele, 1964; Smith and Butterworth, 1967) have also been recorded in the Triassic of South and North China (Song et al., 2000; Ouyang et al., 2011; Ji et al., 2015; Ji et al., 2017; Liu et al., 2015). Moreover, 18 species from the Jurassic–Cretaceous or Paleogene from Canada and Siberia—Converrucosisporites saskatchewanensis, Verrucosisporites asymmetricus, V. rotundus, Duplexisporites rotundatus, Alisporites bilateralis, A. rotundus, Protopinus subluteus, Piceaepollenites omoriciformis, Platysaccus luteus, Podocarpidites multicinus, P. ornatus, P. paulus, P. unicus, Pseudowalchia crocea, Ovalipollis breviformis, Psophosphaera cognatus, Araucariacites australis, and Cycadopites typicus (Cookson, 1947; Malyavkina, 1949; Krutzsch, 1955; Bolkhovitina, 1956; Rouse, 1959; Pocock, 1962; Pocock, 1970; Singh, 1964; Shugaevskaya, 1969)—were found in the present assemblages; however, most of them have also been reported in other Triassic palynological assemblages worldwide (Varyukhina, 1961; Clarke, 1965; Scheuring, 1966; Schulz, 1967; Pocock, 1970; Warrington, 1970; Gradstein, 1971; Planderova, 1972; Horowitz, 1973; Tuzhikova, 1975; 1979; Kimyai, 1977; Tuzhikova, 1980; Semenova, 1987; Song et al., 2000). Another 20 local species, including Cyclogranisporites callosus, Converrucosisporites decorates, Verrucosisporites mimicus, V. scitulus, Apiculatisporis pilosus, Baculatisporites versiformis, Asseretospora curvata, L. communis, L. microreticuIata, L. minuta, L. sinica, L. subornata, L. watangensis, Aratrisporites exiguous, Arat. xiangxiensis, Taeniaesporites combinatus, T. divisus, T. watangensis, Podocarpidites transverses, and D. granulus, have been recorded in the Triassic or early Jurassic in China, especially in the Ordos Basin (Ouyang, 1982; Miao et al., 1984; Zhang, 1984; Song et al., 2000). Therefore, the overall geological age of the present palynoflora is around the Carnian.
The CG assemblage from YC7-3 was considered to be Late Triassic Carnian in age because, around the world, the species Cyathidites minor (Couper, 1953), Dictyophyllidites harrisii (Couper, 1958), Apiculatisporis bulliensis (Helby ex De Jersey, 1971), Aratrisporites xiangxiensis (Li and Shang, 1980), Piceaepollenites omoriciformis (Bolkh.) (Xu and Zhang, 1980), Podocarpidites ornatus (Pocock, 1962), and Discisporites granulus (Zhang, 1984), and genus Classopollis (Pflug, 1953) were only found in the CG assemblage and were never aged older than the Carnian of the Late Triassic (Figure 6).
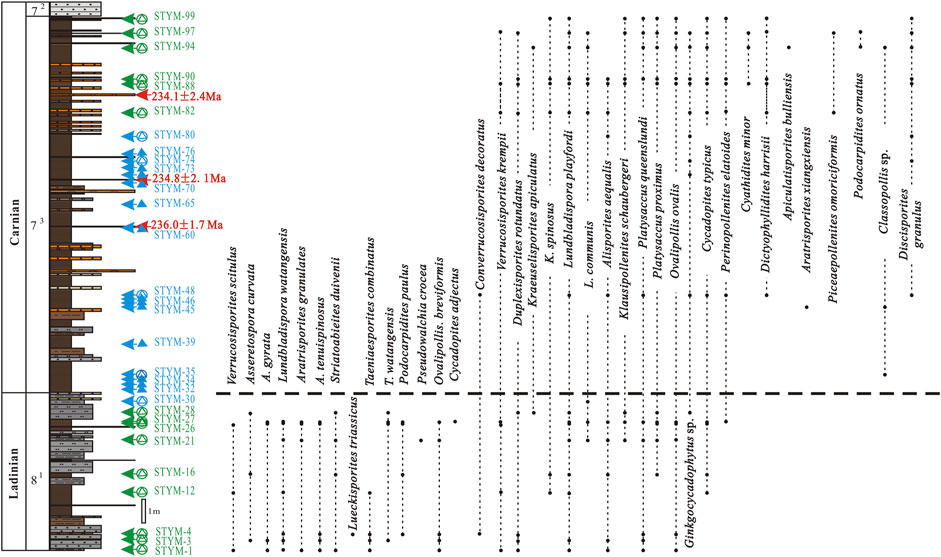
FIGURE 6. Distribution of selected palynological taxa during the Ladinian–Carnian transition in the Yunmeng profile, Ordos Basin, China.
Meanwhile, some of the species in the WC assemblage, including Lundbladispora nejburgii (Schulz, 1964), Plicatipollenites indicus (Lele, 1964), Striatoabieites bricki (Sedova, 1956), S. duivenii (Jansonius) (Hart, 1964), and Klausipollenites schaubergeri (Potonie et Klaus) (Jansonius, 1962) are normally found in the Lower Triassic and the Anisian in Eurasia (Geiger and Hopping 1968; Sakulina, 1973; Vinogradova, 1974; Tuzhikova, 1975; Warrington, 1979). Some local species, such as Cyclogranisporites callosus (Du, 1985), Verrucosisporites mimicus (Qu and Wang, 1986), Lundbladispora communis (Ouyang and Li, 1980), L. microreticulata (Qu, 1982), L. minuta (Qu, 1984), Lundbladispora sinica (Ouyang and Li, 1980), Lundbladispora subornata (Ouyang and Li, 1980), L. watangensis (Qu, 1984), Aratrisporites exiguous (Qu, 1984), Taeniaesporites combinatus (Qu and Wang, 1990), Taeniaesporites divisus (Qu, 1982), and Taeniaesporites watangensis (Qu, 1984) were reported in the Lower Triassic and never in the Upper Triassic in China. Thus, the age of the WC assemblage from YC8-1 was assigned to the Middle Triassic Ladinian (Figure 6).
5.2 Reconstructed palynofloras
Based on the compositions, habitats, and vertical distributions of the mother plants of pollen and spores, three palynofloras were reconstructed on the Yunmeng profile in Ordos Basin, China (Figure 7).
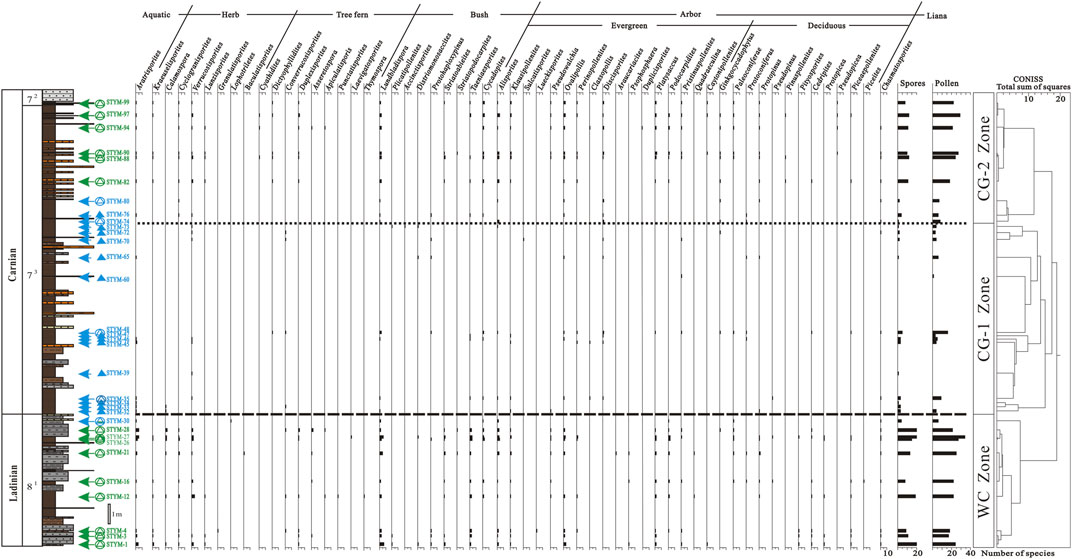
FIGURE 7. Diversity and reconstructed living habit of palynofloras at the Yunmeng profile, Ordos Basin, China.
5.2.1 Latest Ladinian palynoflora
The latest Ladinian palynoflora reconstructed from the YC8-1 sub-member on the Yunmeng profile in the Ordos Basin can be represented by the L. watangensis–T. combinatus assemblage (WC). It is quite diverse in its composition, comprising 121 species of 47 genera (Supplementary Appendix S1). The spores in this palynoflora include 54 species belonging to 17 genera. The most diverse genera were Verrucosisporites (10 species), Lundbladispora (10 ), and Aratrisporites (8 ), which accounted for half of the spore species. The botanical affinity of Verrucosisporites spores might be with those of lycophytes (Isoetales) or Pteridophytes (Zygopteridales, Botryopteridales, and Marattiales) (Mamay, 1950; Balme, 1995; Taylor et al., 2009); however, Bromsgrovia with in situ Verrucosisporites spores from the Anisian in the United Kingdom are considered horsetails (Seyfullah et al., 2013). Therefore, the Verrucosisporites in the present flora are considered sphenophytes, an emerging plant on the Triassic Ordos Lake shore. Lundbladispora spores belong to Pleuromeiales, lycophytes (Balme, 1995), and are recorded in situ in the Devonian lycopod strobill Bisporangiostrobus (Chitaley and McGregor, 1988) and Triassic Pleuromeia sternbergii (Grauvogel-Stamm and Lugardon, 2004). As the remarkable characteristic of three rounded papillae between the trilete rays on the proximal surface is found in the Triassic Pleuromeia cone, we agree that the mature microspores of P. sternbergii are Lundbladispora-type (Balme, 1995). Aratrisporites spores are additional lycophytic microspores that have been reported in situ in Annalepis zeilleri, Araucarites tomiensis, Cyclostrobus sydneyensis, Lycostrobus scottii, Tomiostrobus radiates, and Isoetes ermayingense (Wang, 1991; Balme, 1995) and are considered quillwort and an aquatic lycopod (Retallack, 1997). Sphenophytes include Calamospora, Cyclogranisporites; lycophytes, Kraeuselisporites; and pteridophytes, Lunzisporites, Punctatisporites, Granulatisporites, Lophotriletes, Baculatisporites, Apiculatisporis, Duplexisporites, Asseretospora, Laevigatosporites, and Thymospora. These are herbs or fern trees surrounding the lake and riverside or distributed on the wet lowland.
The pollen of the latest Ladinian palynoflora mainly comprises Pteridospermophytes: Taeniaesporites (seven species), Alisporites (seven), coniferophytes: Podocarpidites (five), Ovalipollis (five), and cycadophytes: Cycadopites (five species). Taeniaesporites and Alisporites have bisaccate pollen with or without striae on the corpus and are considered Permotheca disparis-type pollen because of their morphological similarity to in situ P. disparis (Zalessky) Naugolnykh pollen (Krassilov et al., 1999; Zhang J. G. et al., 2021). They might have been evergreen peltaspermale shrubs. Ovalipollis (Krutzsch) Klaus is an elliptical protobisaccate pollen with anasulcus being regarded as Majonica alpina-type (Zhang J. G. et al., 2021) and might be an evergreen tree of Voltziales. The mother plants yielding Podocarpidites pollen should be Podocarpaceae arbors in the dry lowlands and uplands as canopy forests (Li et al., 2020). Cycadopites are heliophyte shrubs present in the lowlands (Mander et al., 2010; Paterson et al., 2016). These five gymnosperm pollen genera might represent the leading taxa in lowland and upland vegetation. Other diverse pollen species included coniferophytes: Platysaccus (four species) and Pteridospermophytes: Klausipollenites (three) and Striatoabieites (three), which are evergreen trees and shrubs. Compared to evergreen plants, deciduous arbor conifer trees are diverse at the genus level but impoverished at the species level: Protopinus, Pseudopinus, Cedripites, Piceaepollenites, Piceites, Protopicea, and Pseudopicea. They may have been present in the upland or mountainous areas.
In summary, the latest Ladinian palynoflora are diverse in every niche, including lakes, bogs, rivers, wet and dry lowlands, uplands, and mountainous peaks. The lacustrine vegetation is characterized by aquatic Aratrisporites quillworts, Verrucosisporites, and Calamospora sphenopsids as emerging plants on the shore. The wet lowland vegetation mainly comprises Lundbladispora lycopods on the riverbank, Verrucosisporites (Kraeuselisporites sphenopsids), and some herbaceous ferns such as Lunzisporites, Apiculatisporis, and Asseretospora. The dry lowland vegetation is characterized by evergreen Cycadopites, Taeniaesporites bushes, and Duplexisporites–Asseretospora fern trees. The upland vegetation consists of diverse evergreen arbor forests represented by Alisporites, Ovalipollis, Platysaccus, and Podocarpidites mingled with deciduous conifer trees such as Cedripites and Piceaepollenites on the high mountains.
5.2.2 Earliest Carnian palynoflora
The earliest Carnian palynoflora reconstructed from the lower part of the YC7-3 sub-member on the Yunmeng profile in Ordos Basin can be represented by the early sub-assemblage of the L. communis–D. granulus assemblage (CG-1). This comprises 35 species of 29 genera, which is small not only in the number of species but also in the quantity of specimens compared to the latest Ladinian palynoflora (Supplementary Appendix S1; Figure 7). The spores in this small palynoflora include only nine species of six genera: Lundbladispora (three species), Converrucosisporites (two), Aratrisporites, Calamospora, Verrucosisporites, and Dictyophyllidites. Gymnosperm pollen (26 species of 23 genera) are relatively diverse at the genus level, including Pteridospermophytes (Protohaploxypinus, Taeniaesporites, Alisporites, Klausipollenites, Sulcatisporites), Coniferophytes (Plicatipollenites, Accinctisporites, Protohaploxypinus, Pseudowalchia, Ovalipollis, Paleoconiferae, Protoconiferus, Pristinuspollenites, Protopinus, Pseudopinus, Cedripites, and Piceaepollenites), Cycadophytes (Cycadopites), and Ginkgophytes (Ginkgocycadophytus).
Although it is small, the earliest Carnian palynoflora covers most niches, including lakes, rivers, wet and dry lowlands, uplands, and mountains. The lacustrine vegetation comprises aquatic Aratrisporites quillworts and Verrucosisporites–Calamospora sphenopsids as emerging plants on the shore. The wet lowland vegetation consists of Lundbladispora lycopods on the riverbank and some fern trees represented by Dictyophyllidites and Converrucosisporites. The dry lowland vegetation is characterized by evergreen Cycadopites bushes and Pteridospermophytes shrubs such as Taeniaesporites and Protohaploxypinus, as well as evergreen voltzialean shrubs such as Plicatipollenites and Accinctisporites. The upland vegetation consists of evergreen arbor forests represented by Alisporites, Klausipollenites, Pseudowalchia, Ovalipollis, Platysaccus, and Pristinuspollenites mingled with deciduous conifer trees such as Cedripites and Piceaepollenites on the high mountains. The most conspicuous characteristic of upland vegetation is the acceding of Cheirolepidiaceae and Cupressaceae evergreen scaly leaf trees represented by Discisporites and Perinopollenites, respectively. This indicates that the two coniferous families most likely originated in the early Carnian (Taylor et al., 2009).
5.2.3 Early Carnian palynoflora
The early Carnian palynoflora reconstructed from the upper part of the YC7-3 sub-member on the Yunmeng profile in Ordos Basin can be represented by the late sub-assemblage of the L. communis–D. granulus assemblage (CG-2). It comprises 61 species belonging to 37 genera, which is much more diverse than the earliest Carnian palynoflora (Figure 7). The spores in this palynoflora comprise 16 species belonging to 11 genera, including Lundbladispora (three species), Verrucosisporites (two), Duplexisporites (two), Aratrisporites, Kraeuselisporites, Cyclogranisporites, Lunzisporites, Cyathidites, Dictyophyllidites, Asseretospora, and Apiculatisporis. The gymnosperm pollen (45 species in 26 genera) is quite diverse, especially comprising Cycadopites (five species), Alisporites (five), Taeniaesporites (three), Ovalipollis (three), Platysaccus (three), and Podocarpidites (three species).
The early Carnian palynoflora shows characteristics of recovery from the earliest Carnian palynoflora and adaptation to seasonal climate. Almost all niches are occupied by different kinds of plants. The lacustrine vegetation is represented by Verrucosisporites–Cyclogranisporites sphenopsids as emerging plants on the shore, as well as some aquatic Aratrisporites quillworts. The wet lowland vegetation includes abundant Lundbladispora lycopods on the riverbank and some fern trees, such as Cyathidites, Dictyophyllidites, Duplexisporites, Asseretospora, and Apiculatisporis. Dry lowland vegetation is characterized by evergreen Cycadopites bushes and Pteridospermophytes shrubs, such as Taeniaesporites and Striatoabieites. The upland vegetation consists of evergreen arbor forests represented by Alisporites, Klausipollenites, Pseudowalchia, Ovalipollis, Discisporites, Platysaccus, Podocarpidites, and Pristinuspollenites mingled with deciduous Ginkgocycadophytus and conifer trees such as Protopinus and Pseudopicea on the high mountains. The most conspicuous characteristic of the dry lowland vegetation is the disappearance of Utrechtiaceae evergreen conical leaf shrubs, which are seen in the earliest Carnian flora represented by Plicatipollenites and Accinctisporites; however, they are neither the main components in the latest Ladinian nor in the earliest Carnian palynofloras.
5.3 Climatic changes
5.3.1 Early and Middle Triassic climate
The Early and Middle Triassic climate in the Tethys area was arid in the tropical and subtropical areas (Parrish, 1993; Wilson et al., 1994; Kent and Olsen, 2000), with green vegetation present only along the permanent rivers (the gallery forests) (Shi et al., 2021). However, the arid climate periodically experienced humid pulses during the Early and Middle Triassic (Preto et al., 2010), and the middle and late Pelsonian humid events of the Anisian are well-documented by macroflora and palynoflora fossils Kustatscher and Van Konijnenburg-van Cittert, 2010). These humid pulses might have been important for climate change in the Ordos Basin as the changes caused the transition in the deposits from the magenta-red deposit of the Heshanggou Formation to the green and gray deposits with red interlayers of the Zhifang Formation (Qu, 1980). The disappearance of the reddish layers and the appearance of the coal seams indicate the beginning of the Yanchang Formation (sensu lato) (IGCAGS, 1980), which might be attributed to the Ladinian humid event marked by paleokarst breccia in the Dolomite Alps (Mutti and Weissert, 1995).
5.3.2 Carnian pluvial event
The Carnian pluvial event (CPE) is recognized as a carbonate crisis (Schlager and Schollnberger, 1974), reef crisis (Brandner, 1984; Donofrio., 1991; Ruttner et al., 1991), and black shale events (Hornung and Brandner, 2005) that occurred in the early Carnian epoch, in both the tropical Tethys and boreal areas (Dubiel et al., 1991; Shi et al., 2009; Preto et al., 2010; Wang et al., 2012; Mueller et al., 2015; Mueller et al., 2016). Since the CPE is also marked by negative carbon isotopic excursion (Dal Corso et al., 2012; Dal Corso et al., 2015), it is regarded as a global event caused by mega-monsoons (Dubiel et al., 1991; Mutti and Weissert, 1995) or increased atmospheric CO2 from volcanic eruptions (Dal Corso et al., 2012; 2015; Mueller et al., 2016). The CPE is marked by a short-lived increase in rainfall (Preto and Hinnov, 2003; Hornung and Brandner, 2005; Rigo et al., 2007) in the Carnian. The vegetation changes in the western Tethys during the CPE were characterized by a decrease in pollen grains normally attributed to xerophytic upland vegetation and an increase in hygrophytic associations attributable to herbaceous Filicopsida (ferns), Lycopodiales (clubmosses), Equisetopsida (horsetails), and Cycadeoidales (Roghi et al., 2010). However, palynological data from the YC7 and YC8 members in the Xifeng area of Qingyang, Shaanxi Province (Ji and Meng, 2006; Ji and Zhu, 2013) might reveal another scenario: the climate in the early Late Triassic might have been as warm and humid as that in the late Middle Triassic. Recent studies in the Jiyuan Basin, Henan Province, China, showed a depositional thickness from the CPE of approximately 40 m in the ZJ-1 borehole based on four of eleven palynological samples scattered in a 140-m thick core (Lu et al., 2021).
Based on previous research (Balme, 1995; Taylor et al., 2009; Zhang J. G. et al., 2021), the botanical affinities, habits, eco-groups, and environments of the palynological taxa were reconstructed for the palynoflora on the Yunmeng profile in the Ordos Basin (Table 2). To better understand climate change, both the abundance of palynomorphs and the diversity at the species level were statistically analyzed (Figure 8). Not included in the analysis were 16 productive samples (STYM-30, -32, 33, -34, -39, -45, -46, -47, -48, -60, -65, -70, -72, -73, -74, and -80) because they included <10 taxa. The other 16 samples were distributed in two groups: the uppermost part of the YC8-1 sub-member and the upper part of the YC7-3 sub-member and were subjected to quantitative analysis (Visscher and Van der Zwan, 1981). The results show that the climates revealed by the two palynofloras were similar, with both vegetation types including hydro-, meso-, and xerophytic elements (Figure 8). The xerophytic plants comprised 11%–38% of the vegetation at the species level and 14%–35% of the total sporo-pollen specimens. Evergreen shrubs, trees, and deciduous arbors were present in the latest Ladinian and early Carnian palynofloras. The hydrophytic plants, including lycophytes, sphenophytes, and pteridophytes, comprised 25%–50% and 20%–51% of the total species and specimens, respectively. The mesophytic plants included Pteridospermophytes, Ullmanniaceae, Majonicaceae, Araucariaceae, Podocarpaceae, Caytoniales, and Cycadopites, and comprised 35%–54% and 32%–49% of the total species and specimens, respectively. Therefore, the phytoclimatic compositions of the two palynofloras were generally quite stable. This was also evidenced by the coexistence of gnetalean liana plants (Chasmatosporites) in the two palynofloras (Supplementary Appendix S1; Table 2).
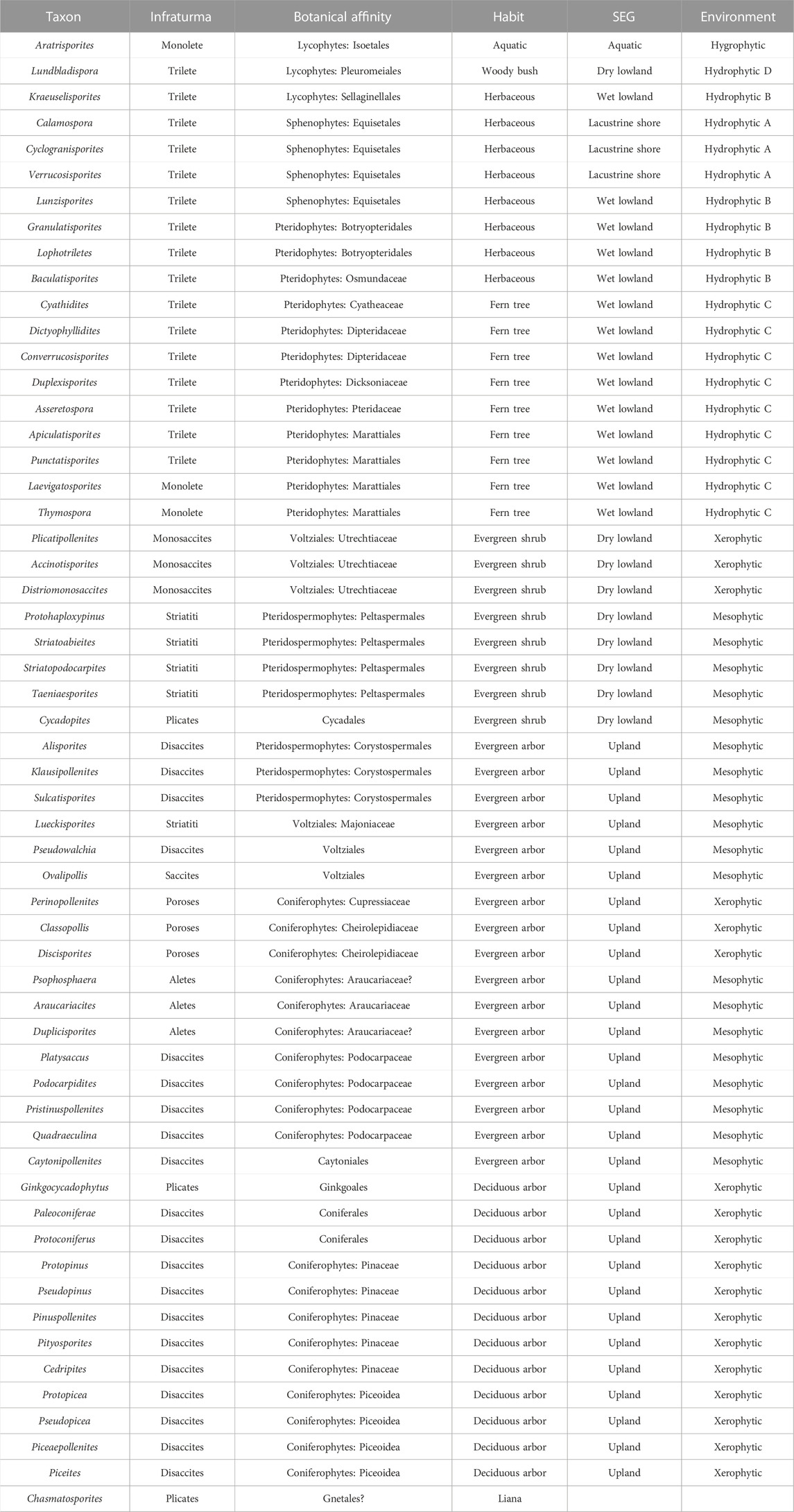
TABLE 2. Botanical affinities, habits, eco-groups, and environments of dispersed palynofloras in Ordos Basin, China.
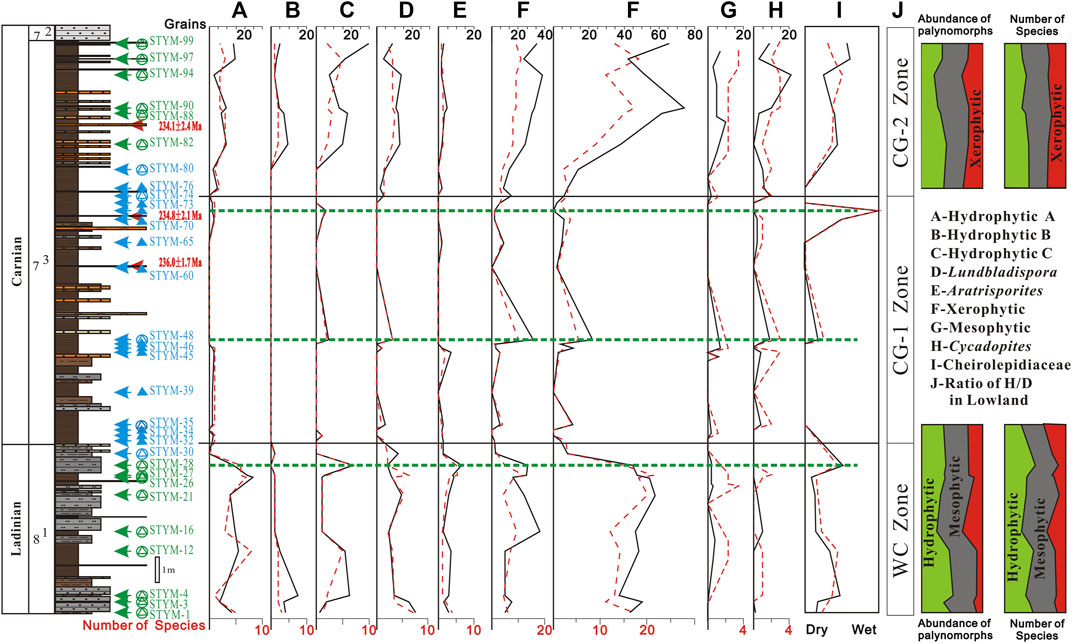
FIGURE 8. Reconstructed humidity based on diversity and abundance of palynofloras at Yunmeng profile, Ordos Basin, China.
However, the compositions of the flora in the two assemblages differed greatly; the WC assemblage in the YC8-1 sub-member included 121 species in 47 genera, while the CG assemblage in the YC7-3 sub-member included only 76 species in 45 genera. The diversity lost at the species level was 38%. Furthermore, examination of the spores showed that 38 species—70% of the spores in the WC assemblage—were lost, including many species from Aratrisporites, Lundbladispora, Calamospora, Verrucosisporites, Cyclogranisporites, Asseretospora, and Apiculatisporites. Although the deepening and enlargement of the Triassic Ordos Lake might have led to the mother plants being far from the deep and semi-deep lake deposits, the aquatic Aratrisporites also greatly declined, with only one species remaining from the former eight. This might imply that the lake was eutrophic, as also evidenced by the burst of algae and organic-rich oil shales in the YC7-3 sub-member (Liu et al., 2022). Regarding eco-groups (Figure 8), the hydrophytic A and B types are composed of horsetails and herbaceous ferns; they were quite abundant and diverse in the WC zone but rare or absent in the CG zone, especially in the CG-1 sub-zone. Dendroid ferns also exhibited a similar pattern. Therefore, the lake was eutrophic, and the climate was warm but likely much more arid than before.
Approximately a third of the pollen (26 species of gymnosperm pollen) in the WC assemblage was lost in the CG assemblage. The lost gymnosperms mainly included Peltaspermales, Corystospermales, Voltziales, Araucariaceae, and some old conifers living upland and in dry lowlands as evergreen shrubs and deciduous and evergreen trees, which included the mesophytic and xerophytic plants. Therefore, this decline is less affected by eco-groups, habitats, and climate type, which must be an overall loss. Meanwhile, Cheirolepidiaceae and Pinuspollenites emerged, and Utrechtiaceae conical leaf shrubs survived. This also indicates a drier climate. In summary, the loss of spores and pollen in every niche and the emergence of most xerophytic Cheirolepidiaceae plants indicate that the climate became drier when the lake became deeper, broader, and euphytic. This coincided with a strong seasonal monsoon climate under greenhouse conditions.
5.3.3 Monsoon arid climate
The monsoon arid climate might have begun in the latest Ladinian when the flora began to fade at the top of the YC8-1 sub-member (Figure 8). Most hydrophytic and mesophytic plants recovered until the upper part of the YC7-3 sub-member, and even xerophytic plants recovered by adding more xerophytic plants. However, hydrophytic C (dendroid ferns), Lundbladispora, Cycadopites, and Cheirolepidiaceae exhibited one or two peaks in abundance and diversity in the CG-1 sub-zone. These eco-group peaks in the CG-1 sub-zone appear contradictory; based on these peaks, hydrophytic C, hydrophytic D, and Cycadopites favor humid climate, while others such as xerophytic and mesophytic plants, and Cheirolepidiaceae prefer arid environments. Cycadopites acme is an indicator of the CPE, together with Aulisporites astigmousus acme, in the northern calcareous Alps (Mueller et al., 2016). However, there are no acmes of Cycadopites or A. astigmousus in the boreal region—for example, Central Spitsbergen (Mueller et al., 2015). Therefore, it is difficult to provide a key palynological indicator for the CPE. Meanwhile, the carbon isotopic stratigraphy of the Ladinian and Carnian deposits in the western and eastern Tethys shows at least four negative excursion episodes (Dal Corso et al., 2012; Dal Corso et al., 2015; Dal Corso et al., 2018; Mueller et al., 2016; Sun et al., 2016; Miller et al., 2017). Recent carbon isotopic chemostratigraphy in the Ordos Basin, together with cyclostratigraphy and isotopic dating, has revealed several negative excursions in the YC8 and YC7 members; however, whether these results are related to Ladinian or Carnian events remains controversial (Zhang K. et al., 2021; Jin et al., 2021).
The latest palynological research in the Carnian amber-bearing section of the Dolomite Alps in Italy sheds light on the primary criteria for identifying the CPE. The coexistence of Majonicaceae (Lueckisporites), Voltziaceae (Triadispora), Araucariaceae (Araucariacites), and Pinaceae (Abietineaepollenites) pollen may provide a mark of the CPE (Roghi et al., 2022). In this study, Lueckisporites in the Yunmeng profile were only found in sample STYM-04 and Araucariacites only in sample STYM-12; moreover, they did not coexist. This might indicate that sample STYM-04 was not the last observation of Lueckisporites. Meanwhile, the possibility of pioneering Araucariacites cannot be avoided (Tokunaga et al., 1977). If we consider Abietineaepollenites for calibration, the lowest record of the genus was in the Erqiao Formation in Guizhou Province, China, which is younger than the Carnian Neogondolella polygnathiformis zone (Yang et al., 1995; Shang, 2011); however, the Araucariacites were neither recorded there nor in the younger Late Triassic horizons. As the uppermost Falang Formation is aged early Carnian due to the N. polygnathiformis zone (Yang et al., 1995), the overlying Erqiao and Banan formations are most probably late Julian–Tuvalian in age (Tong et al., 2019). Therefore, the coexistence of Dictyophyllidites, Lueckisporites, and Abietineaepollenites might be regarded as an indicator of the CPE. Furthermore, Classopollis in the Erqiao Formation may be another reference for the event. Therefore, it is reasonable to regard the existence of Classopollis and Dictyophyllidites as indications of the CPE. Thus, the CPE on the Yunmeng profile in the Ordos Basin can be roughly located in the CG-1 sub-zone (Figure 8). The humid pulse can be recognized by the humidity index of the lowland plants, which is directly related to the fine organic-rich deposit of deep and semi-deep lacustrine environments. At least three humid pulses can be observed on the Yunmeng profile, including two in the early Carnian CG-1 sub-zone (marked by green dashed lines in Figure 8).
6 Conclusion
Detailed palynological investigation of the Yunmeng profile in the Ordos Basin, China, was performed to determine the age and reconstruct its vegetation and climate. The result revealed: 1) two palynological assemblages for the YC8-1 and YC7-3 sub-members, L. watangensis–T. combinatus and L. communis–D. granulus, respectively; 2) correlation of the GSSP of the Carnian and the Late Triassic floras in South China with marine conodont fossils allowing the assignment of the two assemblages to the latest Ladinian and early Carnian ages, respectively; 3) the coexistence of Cyathidites minor (Couper, 1953), Dictyophyllidites harrisii (Couper, 1958), Apiculatisporis bulliensis (Helby ex De Jersey 1971), Aratrisporites xiangxiensis (Li and Shang, 1980), Piceaepollenites omoriciformis (Bolkh.) (Xu and Zhang, 1980), Podocarpidites ornatus (Pocock, 1962), Discisporites granulus (Zhang, 1984), and Classopollis (Pflug, 1953) is proposed as an indicator of the Carnian age in the North China palynofloral realm; 4) vegetation changes are obvious, especially in the boundary between the Ladinian and Carnian, in which 70% of ferns and >30% of gymnosperm species were lost; the vegetation changes suggest a strong seasonal arid climate, as indicated by the emergence of Cheirolepidiaceae and Pinuspollenites; 5) the climate during latest Ladinian and early Carnian was “hot house,” with evergreen shrubs, bushes, and arbor trees flourishing in the tropical Ordos Basin, which was isolated by the Qinling–Dabie orogenic belt mountains from the Tethys to the south; 6) the warm seasonal arid climate might have been interrupted by a monsoonal pluvial pulse, and three strong pulses are signaled in the latest Ladinian and early Carnian based on the humidity index of lowland plants.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding author.
Author contributions
YS and XL proposed the study, collected the samples, identified most part of the palynomorphs, and wrote the original draft; YZ edited the figures; XS proofread the manuscript; SZ identified some of the sporopollen. All authors have read and agreed to the final version of the manuscript.
Funding
This work is financially supported by the National Natural Science Foundation of China (Grant nos. 41972012 and 31700183) and the State Key Laboratory of Shale Oil and Gas Enrichment Mechanisms and Effective Development (No. GSYKY-B09-33).
Acknowledgments
The authors express their sincere thanks to Prof. Jinnan Tong (China University of Geosciences, Wuhan) for providing constructive discussion on the Triassic deposits in China and to Prof. Quanyou Liu (Institute of Energy, Peking University), Peng Li and Rui Zhang (Petroleum Exploration and Production Research Institute, SINOPEC), and Mingda Zhang (Jilin University, Changchun) for observing organic cumulations and collecting samples in the field. They also thank the two reviewers for their careful and thorough review of the manuscript; their constructive suggestions have considerably strengthened the main topic of the manuscript. Finally, they express their special thanks to Prof. Xing Xu (Associate Editor-in-Chief) for graciously handling this report.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2022.1008707/full#supplementary-material
SUPPLEMENTARY APPENDIX S1 | Stratigraphy of the Yunmeng profile showing isotopic dating and productive palynological samples. Results of the quantitative sporo-pollen analysis (number).
SUPPLEMENTARY APPENDIX S2 | Description of the species in the Yunmeng Profile.
References
Arjang, B. (1975). The Rhaetian-Jurassic flora of Iran and Afghanistan. 1. The microflora of the Rhaetian-Jurassic deposits of the kerman basin (central Iran). Palaeontogr. Abt. B 152 (4), 85–148.
Balme, B. E. (1995). Fossil in situ spores and pollen grains: An annotated catalogue. Rev. Palaeobot. Palynology 87, 81–323. doi:10.1016/0034-6667(95)93235-X
Balme, B. E. (1963). Plant microfossils from the Lower Triassic of western Australia. Palaeontology 6 (1), 12–40.
Benton, M. J., and Newell, A. J. (2014). Impacts of global warming on Permo-Triassic terrestrial ecosystems. Gondwana Res. 25, 1308–1337. doi:10.1016/j.gr.2012.12.010
Bharadwaj, D. C., and Singh, H. P. (1964). An Upper Triassic miospore assemblage from the coals of Lunz, Austria. Palaeobotanist 12 (1), 28–44. doi:10.54991/jop.1963.641
Bolkhovitina, N. A. (1956). Atlas of spores and pollen from Jurassic and Lower Cretaceous deposits of the vilyui depression. USSR Transactions of the Institute of Geology Academy of Sciences, 1–188. (in Russian).
Boucot, A. J., Chen, X., Scotese, C. R., and Fan, X. X. (2009). Phanerozoic paleoclimate reconstruction of the world. Beijing: Science Press.
Brandner, R. (1984). Meeresspiegelschwankungen und Tektonik in der Trias der NW–Tethys. Jahrb. Geol. Bundesanst. 126 (4), 435–475.
Bureau of Geology and Mineral Resources of Shaanxi Province, (1998). Stratigraphy (lithostratic) of Shaanxi Province. Wuhan: China University of Geosciences Press.
Césari, S. N., and Colombi, C. (2016). Palynology of the Late Triassic ischigualasto formation, Argentina: Paleoecological and paleogeographic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 449, 365–384. doi:10.1016/j.palaeo.2016.02.023
Chitaley, S. D., and McGregor, D. C. (1988). Bisporangiostrobus harrisii gen. et sp. nov. an eligulate lycopsid cone with Duosporites megaspores and Geminospora microspores from the Upper Devonian of Pennsylvania. U.S.A. Palaeontogr. Abt. B 210 (4–6), 127–149.
Clarke, R. F. A. (1965). Keuper miospores from worcetershire, England. Palaeontology 8 (2), 294–321.
Colombi, C., Martínez, R. N., Césari, S., Alcober, O., and Montanez, I. (2021). A high-precision U-Pb zircon age constraints the timing of the faunistic and palynofloristic events of the Carnian Ischigualasto Formation, San Juan, Argentina. J. S. Am. Earth Sci. 111 (6), 103433. doi:10.1016/j.jsames.2021.103433
Cookson, I. C. (1947). Plant microfossils from the lignites of kerguelen archipelago. New Zealand Antarctic Res Expedition, 127–142. Reports Series A2.
Couper, R. A. (1958). British Mesozoic microspores and pollen grains. A systematic and stratigraphic study. Palaeontogr. Abt. B -Stuttgart- 103 (4–6), 75–179.
Couper, R. A. (1953). Upper Mesozoic and Cainozoic spores and pollen grains from New Zealand. Paleontol. Bull. N. Z. Geol. Surv. 22, 1–77.
Cuneo, N. R., Taylor, E. L., Taylor, T. N., and Krings, M. (2003). In situ fossil forest from the upper Fremouw Formation (Triassic) of Antarctica: Paleoenvironmental setting and paleoclimate analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 197, 239–261. doi:10.1016/S0031-0182(03)00468-1
Dal Corso, J., Gianolla, P., Newton, R. J., Franceschi, M., Roghi, G., Caggiati, M., et al. (2015). Carbon isotope records reveal synchronicity between carbon cycle perturbation and the “Carnian Pluvial Event” in the Tethys realm (Late Triassic). Glob. Planet. Change 127, 79–90. doi:10.1016/j.gloplacha.2015.01.013
Dal Corso, J., Gianolla, P., Rigo, M., Franceschi, M., Roghi, G., Mietto, P., et al. (2018). Multiple negative carbon-isotope excursions during the carnian pluvial episode (late triassic). Earth-Science Rev. 185, 732–750. doi:10.1016/j.earscirev.2018.07.004
Dal Corso, J., Mietto, P., Newton, R. J., Pancost, R. D., Preto, N., Roghi, G., et al. (2012). Discovery of a major negative δ13C spike in the Carnian (Late Triassic) linked to the eruption of Wrangellia flood basalts. Geology 40 (1), 79–82. doi:10.1130/G32473.1
De Jersey, N. J. (1972). Triassic miospores from the esk beds. Publ. Geol. Surv. Queensl., 357, Palaeontol. Pap. 32, 1–40.
De Jersey, N. J. (1971). Triassic miospores from the tivoli formation and kholo sub-group. Publ. Geol. Surv. Queensl. 353, 1–40.
De Jersey, N. J. (1964). Triassic spores and pollen grams from the Bundamba Group. Publ. Geol. Surv. Queensl. 321, 1–21.
De Jersey, N. J. (1962). Triassic spores and pollen grams from the Ipswich Coalfield. Publ. Geol. Surv. Queensl. 307, 1–18.
Deng, S. H., Lu, Y. Z., Luo, Z., Fan, R., Li, X., Zhao, Y., et al. (2018). Subdivision and age of the Yanchang Formation and the Middle/Upper Triassic boundary in Ordos Basin, North China. Sci. China Earth Sci. 61, 1419–1439. doi:10.1007/s11430-017-9215-3
Domeier, M., and Torsvic, T. H. (2014). Plate tectonics in the Late Paleozoic. Geosci. Front. 5, 303–350. doi:10.1016/j.gsf.2014.01.002
Donofrio, D. A. (1991). “Radiolaria and porifera spiculae from the Upper Triassic of aghdarband (ne–Iran),” in The Triassic of aghdarband, AqDarband ne–Iran, and its pre-Triassic frame. Editor A. W. Ruttner (Abhandlungen der Geologischen Bundesanstalt), 38, 205–223.
Du, B. A. (1985). Middle triassic palynological assemblage in the Wangjiashn basin of Jingyuan, Gansu and its stratigraphical significance. Acta Bot. Sin. 27 (5), 538–544. (in Chinese with English abstract).
Dubiel, R. F., Parrish, J. T., Parrish, J. M., and Good, S. C. (1991). The pangaean megamonsoon: Evidence from the Upper Triassic chinle formation, Colorado plateau. Soc. Sediment. Geol. 6 (4), 347–370. doi:10.2307/3514963
ECSDC (Editorial Committee of Stratigraphic Dictionary of China) (2000). Stratigraphic dictionary of China:Triassic system. Beijing: Geological Publishing House.
Fedo, C. M., Sircombe, K. N., and Rainbird, R. H. (2003). Detrital zircon analysis of the sedimentary record. Rev. Mineralogy Geochem. 53, 277–303. doi:10.2113/0530277
Fisher, M. J., and Bujak, J. (1975). Upper Triassic palynofloras from arctic Canada. Proc. Annu. Meet. Am. Assoc. Stratigr. Palynologists 11 (6), 87–94. doi:10.2307/3687224
Gair, H. S., Norris, G., and Ricker, J. (1965). Early Mesozoic microfloras from Antarctica. N. Z. J. Geol. Geophys. 8 (2), 231–235. doi:10.1080/00288306.1965.10428109
Geiger, M. E., and Hopping, C. A. (1968). Triassic stratigraphy of the southern North sea basin. Philosophical Trans. R. Soc. B Biol. Sci. 254 (790), 1–36. doi:10.1098/rstb.1968.0012
Gradstein, F. M. (1971). The age of Beds in the Lettenkohle facies in South-East France (A palynological approach). Pollen Spores 13 (1), 169–178.
Grauvogel-Stamm, L., and Lugardon, B. (2004). The spores of the Triassic lycopsid Pleuromeia sternbergii (münster) corda: Morphology, ultrastructure, phylogenetic implications, and chronostratigraphic inferences. Int. J. Plant Sci. 165 (4), 631–650. doi:10.1086/386562
Halle, T. G. (1908). Bidrag till kannedomen om skandinaviens Amphipoda gammaridea. K. Sven. vetenskapsakademiens Handl. 43i (1), 1–104. doi:10.5962/bhl.part.28297
Harris, T. M. (1937). The fossil flora of scoresby sound east Greenland: Part 5. Stratigraphic relations of the plant beds. Meddelel Ser. Om. Grønl. 112, 82–86.
Hart, G. F. (1964). A review of the classification and distribution of the Permian miospore: Disacciti Striatiti. Congrès Int. de Stratigr. de géologie du Carbonifère 5, 1171–1199.
Hornung, T., and Brandner, R. (2005). Biochronostratigraphy of the Reingraben turnover (Hallstatt facies belt), local black shale events controlled by regional tectonics, climatic change and plate tectonics. Facies 51, 460–479. doi:10.1007/s10347-005-0061-x
Horowitz, A. (1973). Triassic miospores from southern Israel. Rev. Palaeobot. Palynology 16 (3), 175–207. doi:10.1016/0034-6667(73)90045-6
IGCAGS (Institute of Geology, Chinese Academy of Geological Sciences) (1980). Mesozoic stratigraphy and palaeontology of Ordos Basin. Beijing: Geological Publishing House Press.
Jansonius, J. (1962). Palynology of Permian and Triassic sediments, peace river area, western Canada. Palaeontogr. Abt. B Band. 110, 35–98.
Ji, L. M., and Meng, F. W. (2006). Palynology of Yanchang Formation of Middle and Late Triassic in eastern Gansu Province and its paleoclimatic significance. J. China Univ. Geosciences 17 (3), 209–220. doi:10.1016/S1002-0705(06)60030-7
Ji, L. M., and Zhu, Y. H. (2013). Sporo-pollen assemblages and paleoclimate of the Yanchang Formation in the Xifeng area, southwestern Ordos Basin, Gansu Province, NW China. Acta Micropalaeontologica Sin. 30 (4), 367–378.
Ji, L. X., Liu, F., Luo, W., and Ouyang, S. (2017). Spores-pollen from Early Triassic deposits in Qinghai Province and their stratigraphic significance. J. Stratigr. 41 (4), 450–466. (in Chinese with English abstract). doi:10.19839/j.cnki.dcxzz.2017.04.012
Ji, L. X., Liu, F., Luo, W., and Ouyang, S. (2015). Triassic sporopollen and acritarchs from northern Qilian Mountain (Qinghai) and their stratigraphical significance. J. Stratigr. 39 (4), 367–379. (in Chinese with English abstract). doi:10.19839/j.cnki.dcxzz.2015.04.003
Jin, X., Baranyi, V., Caggiati, M., Franceschi, M., Wall, C. J., Liu, G. L., et al. (2021). Middle Triassic lake deepening in the Ordos Basin of North China linked with global sea-level rise. Glob. Planet. Change 207 (2), 103670. doi:10.1016/j.gloplacha.2021.103670
Jin, X., McRoberts, C. A., Shi, Z. Q., Mietto, P., Rigo, M., Roghi, G., et al. (2019). The aftermath of the CPE and the Carnian–Norian transition in northwestern sichuan basin, south China. J. Geol. Soc. 176 (1), 179–196. doi:10.1144/jgs2018-104
Kent, D. V., and Olsen, P. E. (2000). Magnetic polarity stratigraphy and paleolatitude of the Triassic–Jurassic blomidon Formation in the fundy basin (Canada): Implications for early Mesozoic tropical climate gradients. Earth Planet. Sci. Lett. 179 (2), 311–324. doi:10.1016/S0012-821X(00)00117-5
Kimyai, A. (1977). Further information on the palynological stratigraphy of the Mesozoic Coaly sediments from Kerman. IranTehran: Iranian Petroleum Institute, Proceeding of the 2nd Geological Symposium of Iran, 191–217. (in Persian).
Klaus, W. V. (1960). Sporen der karnischen stufe der ostalpinen Trias. Jahrb. Geol. Bundesanst. 5, 107–183.
Krassilov, V. A., Afonin, S. A., and Naugolnykh, S. V. (1999). Permotheca with in situ pollen grains from the Lower Permian of the urals. Palaeobotanist 49 (1), 19–25. doi:10.54991/jop.1999.1287
Kustatscher, E., and Van Konijnenburg-van Cittert, J. H. A. (2010). Seed ferns and cycadophytes from the Triassic flora of thale (Germany). Neues Jahrb. für Geol. Paläontologie - Abh. 258 (2), 195–217. doi:10.1127/0077-7749/2010/0097
Kutzbach, J. E., and Gallimore, R. G. (1989). Pangaean climates: Megamonsoons of the megacontinent. J. Geophys. Res. 93 (3), 3341–3357. doi:10.1029/JD094iD03p03341
Lele, K. M. (1964). Studies in the talchir flora of India: 2. Resolution of the spore genus nuskoisporites pot. & KL. Palaeobot. 12 (2), 147–168. doi:10.54991/jop.1963.651
Leschik, G. (1955). Die Keuperflora yon Neuwelt bei Basel. II. Die Iso-und Mikrosporen. Schweiz. Paläontologische Abh. 71, 1–70.
Li, L. Q., Wang, Y. D., Kürschner, W. M., Ruhl, M., and Vajda, Vivi. (2020). Palaeovegetation and palaeoclimate changes across the Triassic–Jurassic transition in the Sichuan Basin, China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 556 (15), 109891. doi:10.1016/j.palaeo.2020.109891
Li, W. B., and Shang, Y. K. (1980). Sporo-pollen assemblages from the Mesozoic coal series of Western hubei. Acta Palaeontol. Sin. 19 (3), 201–219. (in Chinese with English abstract). doi:10.19800/j.cnki.aps.1980.03.004
Li, Y., Yao, J. X., Wang, S. E., and Pang, Q. Q. (2016). Middle-Late Triassic terrestrial strata and establishment of stages in the Ordos Basin. Acta Geosci. Sin. 37 (3), 267–276. doi:10.3975/cagsb.2016.03.02
Liu, Q. Y., Li, P., Jin, Z. J., Sun, Y. W., Hu, G., Zhu, D. Y., et al. (2022). Organic-rich formation and hydrocarbon enrichment of lacustrine shale strata: A case study of chang 7 member. Sci. China Earth Sci. 65, 118–138. doi:10.1007/s11430-021-9819-y
Liu, Z. S., Li, L. Q., and Wang, Y. D. (2015). Late triassic spore-pollen assemblage from the Xujiahe Formation in hechuan of chongqing, China. Acta Palaeontol. Sin. 54 (3), 279–304. (in Chinese with English abstract). doi:10.19800/j.cnki.aps.2015.03.001
Liu, Z. S., Shang, Y. K., and Li, W. B. (1981). Triassic and Jurassic spore-pollen assemblages from some localities of Shaanxi and Gansu, north-west China. Bull. Nanjing Inst. Geol. Palaeontol. Chin. Acad. Sci. 3, 131–210. (in Chinese).
Lu, J., Zhang, P. X., Dal Corso, J., Yang, M. F., Wignall, P. B., Greene, S. E., et al. (2021). Volcanically driven lacustrine ecosystem changes during the carnian pluvial episode (Late Triassic). Proc. Natl. Acad. Sci. U. S. A. 118 (40), e2109895118. doi:10.1073/pnas.2109895118
Ma, X. H., Xing, L. S., Yang, Z. Y., Xu, S. J., and Zhang, J. X. (1993). Paleomagnetic study since late Paleozoic in the Ordos Basin. Acta Geophys. Sin. 36 (1), 68–79. (in Chinese with English abstract).
Mädler, K. (1964). Die geologische Verbreitungvon Sporen und Pollen in der deutschen Trias. Beih. Zum Geol. Jahrb. 65, 1–145.
Malyavkina, V. S. (1949). Determination key of spores and pollen, Jurassic-Cretaceous. Tr. Vsesoyuznogo Neft. Naucho-Issledovatel Skogo Geologo-Razvedochnogo Instituta 33, 1–137.
Mamay, S. H. (1950). Some American Carboniferous fern fructifications. Ann. Mo. Botanical Gard. 37 (3), 409–476. doi:10.2307/2394515
Mander, L., Kurschner, W. M., and Mcelwain, J. C. (2010). An explanation for conflicting records of Triassic-Jurassic plant diversity. Proc. Natl. Acad. Sci. 107 (35), 15351–15356. doi:10.1073/pnas.1004207107
Miao, S. J., Yu, J. X., Qu, L. F., Zhang, W. P., Zhang, Q. H., and Zhang, D. H., (1984). Mesozoic spores and pollen. In Tianjin Institute of Geology and mineral Resources. Palaeontological Atlas of North China, 3. Micropaleontoloy Geological Publishing House Press, 440–638 (in Chinese with English abstract).
Mietto, P., Manfrin, S., Preto, N., Rigo, M., Roghi, G., Furin, S., et al. (2012). The global boundary stratotype section and point (GSSP) of the carnian stage (Late Triassic) at prati di Stuores/stuores wiesen section (southern Alps, NE Italy). Episodes 35 (3), 414–430. doi:10.18814/epiiugs/2012/v35i3/003
Miller, C. S., Peterse, F., da Silva, A. C., Baranyi, V., Reichart, G. J., and Kürschner, W. M. (2017). Astronomical age constraints and extinction mechanisms of the late Triassic Carnian crisis. Sci. Rep. 7 (1), 2557. doi:10.1038/s41598-017-02817-7
Mueller, S., Hounslow, M. W., and Kurschner, W. M. (2015). Integrated stratigraphy and palaeoclimate history of the carnian pluvial event in the boreal realm: New data from the Upper Triassic kapp toscana group in central spitsbergen (Norway). J. Geol. Soc. 173 (1), 186–202. doi:10.1144/jgs2015-028
Mueller, S., Krystyn, L., and Kurschner, W. M. (2016). Climate variability during the carnian pluvial phase - a quantitative palynological study of the Carnian sedimentary succession at lunz am see, northern calcareous Alps, Austria. Palaeogeogr. Palaeoclimatol. Palaeoecol. 441, 198–211. doi:10.1016/j.palaeo.2015.06.008
Mutti, M., and Weissert, H. (1995). Triassic monsoonal climate and its signature in Ladinian-Carnian carbonate platforms (Southern Alps, Italy). J. Sediment. Res. 65 (3), 357–367.
Nilsson, T. (1958). Uber das Vorkommen eines mesozoischen Sapropelgesteins in Schonen. Sweden: Lunds Universitets Arsskrift Press.
Ouyang, S., Ji, L. X., and Luo, W. (2011). Early Triassic spore-pollen assemblages from the Hongshuichuan Group of central-Western Qinghai. Acta Palaeontol. Sin. 50 (2), 187–218. (in Chinese with English abstract). doi:10.19800/j.cnki.aps.2011.02.005
Ouyang, S., and Li, Z. P. (1980). “Microflora from the Kayitou Formation of Fuyuan district, E. Yunnan and its bearing on stratigraphy and palaeobotany,” in Nanjing Institute of Geology and palaeontology, academia sinica (Beijing: Stratigraphy and Palaeontologyof Upper Permian Coal-Bearing Formation in Western Guizhou and Eastern YunnanScience Press), 123–194. (in Chinese).
Ouyang, S. (1982). Upper Permian and Lower triassic palynomorphs from eastern yunnan, China. Can. J. Earth Sci. 19 (1), 68–80. doi:10.1139/e82-006
Pan, Z. X. (1934). Oil shale geology in northern Shaanxi. Central Geological Survey, Geological Report.
Pang, J. G., Li, W. H., and Chen, Q. H. (2010). Characteristics and deposition of marker beds in the Yanchang Formation of the northern Shaanxi. J. Stratigr. 34 (2), 173–178. (in Chinese with English abstract). doi:10.19839/j.cnki.dcxzz.2010.02.011
Parrish, J. T. (1993). Climate of the supercontinent Pangea. J. Geol. 101 (2), 215–233. doi:10.1086/648217
Paterson, N. W., Mangerud, G., and Mørk, A. (2016). Late triassic (early carnian) palynology of shallow stratigraphical core 7830/5-U-1, offshore kong karls land, Norwegian arctic. Palynology 41 (1), 230–254. doi:10.1080/01916122.2016.1163295
PCOC (PetroChina Changqing Oilfield Company) (1992). Petroleum geology of China, Vol. 12. Beijing: Petroleum Industry Press. (in Chinese).
Pflug, H. D. (1953). Origin and development of the angiosperm pollen in Earth history. Palaeontogr. Abt. B 95, 61–171.
Planderova, E. (1972). A contribution to palynological research of lunz beds in west carpathian region. Geol. Pr. Spravy 58, 57–77.
Playford, G., and Dettmann, M. E. (1965). Rhaeto-liassic microfossils from the leigh creek coal measures, south Australia. Senckenberg; Lethaea 46 (2/3), 127–181.
Playford, G. (1965). Plant microfossils from triassic sediments near poatina, tasmania. J. Geol. Soc. Aust. 12 (2), 173–210. doi:10.1080/00167616508728592
Pocock, S. A. J. (1962). Microfloral analysis and age determination of strata at the Jurassic-Cretaceous boundary in the Western Canada Plains. Palaeontogr. Abt. B 111 (1–3), 1–95.
Pocock, S. A. J. (1970). Palynology of the jurassic sediments of Western Canada. Part I and II. Palaeontogr. Abt. B 130, 12–136.
Potonié, R., and Klaus, W. (1954). Einige Sporengattungen des alpinen Sazgebirges. Geol. Jahrb. 68, 517–544.
Preto, N., and Hinnov, L. A. (2003). Unraveling the origin of carbonate platform cyclothems in the Upper Triassic durrenstein formation (dolomites, Italy). J. Sediment. Res. 73 (5), 774–789. doi:10.1306/030503730774
Preto, N., Kustatscher, E., and Wignall, P. B. (2010). Triassic climates-state of the art and perspectives. Palaeogeogr. Palaeoclimatol. Palaeoecol. 290 (1–4), 1–10. doi:10.1016/j.palaeo.2010.03.015
Qu, L. F. (1984). “Palaeontological atlas of North China, 3. Micropaleontology,” in Tianjin Institute of Geology and mineral Resources ((Beijing: Ceological Publishing House Press), 538–573. (in Chinese with English abstract).
Qu, L. F. (1982). The palynological assemblage from the Liujiagou formation of Jiaocheng, Shanxi. Bull. Geol. Inst. Chin. Acad. Geol. Soc. 4, 83–93. (in Chinese with English abstract).
Qu, L. F. (1980). “Triassic spores and pollen,” in Mesozoic stratigraphy and palaeontology of the shanxi-cansu-ningxia basin (1)Institute of ceology, Chinese academy of geological sciences (Beijing: Geological Publishing House Press), 115–143. (in Chinese).
Qu, L. F., and Wang, Z. (1990). “Triassic palynological assemblages in North xinjiang,” in Permian to tertiary strata and palynological assemblages in the North of xinjiang (Xinjiang Petroleum Administration (Beijing: Institute of Geology, Chinese Academy of Geological Sciences, Research Institute of Petroleum Exploration and DevelopmentChina Environment Science Press), 37–56. (in Chinese with English abstract).
Qu, L. F., and Wang, Z. (1986). “Triassic sporopollen,” in Permian and Triassic strata and fossil assemblages in the Dalongkou area of Jimsar (Xinjiang, (Beijing: Institute of Geology, Chinese Academy of Geological Sciences & Institute of Geology, Xinjiang Bureau of Geology and Mineral Resources, Geological Publishing House Press), 111–173.
Retallack, G. J. (1997). Earliest Triassic origin of Isoetes and quillwort evolutionary radiation. J. Paleontology 71, 500–521. doi:10.1017/S0022336000039524
Rigo, M., Preto, N., Roghi, G., Tateo, F., and Mietto, P. (2007). A rise in the carbonate compensation depth of Western Tethys in the Carnian (Late Triassic): Deep-water evidence for the carnian pluvial event. Palaeogeogr. Palaeoclimatol. Palaeoecol. 246 (2–4), 188–205. doi:10.1016/j.palaeo.2006.09.013
Roghi, G., Gianolla, P., Kustatscher, E., Schmidt, A. R., and Seyfullah, L. (2022). An exceptionally preserved terrestrial record of LIP effects on plants in the Carnian (Upper Triassic) amber-bearing section of the dolomites, Italy. Front. Earth Sci. 10, 900586. doi:10.3389/feart.2022.900586
Roghi, G., Gianolla, P., Minarelli, L., Pilati, C., and Preto, N. (2010). Palynological correlation of carnian humid pulses throughout western Tethys. Palaeogeogr. Palaeoclimatol. Palaeoecol. 290, 89–106. doi:10.1016/j.palaeo.2009.11.006
Rouse, C. E. (1959). Plant microfossils from kootenay coal-measures strata of British columbia. Micropalaeontology 5 (3), 303–324. doi:10.2307/1484422
Ruttner, A. W., Brandner, B., and Kirchner, E. (1991). “Geology of the aghdarband area (kopet dagh, ne–Iran),” in The triassic of aghdarband (AqDarband), ne–Iran, and its pre–triassic frame. Editor A. W. Ruttner (Abhandlungen der Geologischen Bundesanstalt), 38, 7–81.
Sakulina, G. V. (1973). “Middle and Late Triassic miospores from southeastern Kazakhstan,” in Mikrofossilii drevneishikh otlozhenii. Proceedings of the 3rd international palynological conference. Editors T. F. Vozzhennikova, and T. B. Timofeev (Moscow: Nauka), 33–38.
Scheuring, B. W. (1970). Palynologische und palynostratigraphische untersuchungen des Keupers im bolchentunnel (Solothurner Jura). Schweiz. Palaontologische Abh. 88, 1–119.
Scheuring, B. W. (1966). Preliminary report on pollen analitical investigations of the upper meridekalke on mount san giorgio (sudtessin). Eclogae Geol. Helvetiae 59 (2), 964–965.
Schlager, W., and Schollnberger, W. (1974). Das prinzip stratigraphischer wenden in der Schichtfolge der Nordlichen kalkalpen. Mitteilungen der Geol. Gesellschaft Wien 66/67, 165–193.
Schulz, E. (1964). Spores and pollen of the middle buntsandstein of the German basin. Monatsberichte Dtsch. Akad. Wisssenschaften, Berl. 6 (8), 597–606.
Schulz, E. (1967). Study of fossil spores from Rhaetian-Liassic Layers in the central part of the German Basin. Palaeontogr. Abt. B. Palaobotanik 2 (3), 543–644.
Schuurman, W. M. (1977). Aspects of Late Triassic palynology. 2. Palynology of the “Gres et Schiste a Avicula contorta” and “Argiles de Levallois” (Rhaetian) of northeastern France and southern Luxemburg. Rev. Palaeobot. Palynology 23 (3), 159–253. doi:10.1016/0034-6667(77)90007-0
Sedova, M. A. (1956). The definition of four genera of disaccate Striatiti. Material Palaeontol. 12, 246–249.
Semenova, E. V. (1987). Miospores from the Triassic deposits in the South-Western regions of the Ukraine. (The fore-dobrogean trough). Geol. Zhurnal 47 (5), 66–70.
Seyfullah, L., Kustatscher, E., and Taylor, W. A. (2013). The first discovery of in situ Verrucosisporites applanatus spores from the Middle Triassic flora from Bromsgrove (Worcestershire, UK). Rev. Palaeobot. Palynology 197, 15–25. doi:10.1016/j.revpalbo.2013.04.004
Shang, Y. K. (2011). Late Triassic palynology of Yunnan and Guizhou. China. Beijing: Geological Publishing House Press. (in Chinese).
Shi, X., Lang, J. B., Shu, W. C., Li, H., and Yu, J. X. (2021). Fossil woods from the olenekian (late Early Triassic) Shaofanggou Formation in the Junggar basin, northern Xinjiang, north-west China. Geol. J. 56 (12), 6223–6230. doi:10.1002/gj.4193
Shi, Z. Q., Ou, L. H., Luo, F. Z., Li, Y., and Qian, L. J. (2009). Black shale event during the Late Triassic Carnian age: Implications from sedimentary and palaeontological records in Longmen Mountains region. J. Palaeogeogr. 11 (4), 375–383. (in Chinese with English abstract).
Shugaevskaya, O. V. (1969). Spores Duplexisporites in upper Mesozoic deposits of the river gerbikan (uda depression). Fossil Fauna Flora ofthe Far East, 1, 153–160.
Simms, M. J., and Ruffell, A. H. (1990). Climatic and biotic change in the Late Triassic. J. Geol. Soc. 147 (1), 321–327. doi:10.1144/gsjgs.147.2.0321
Singh, C. (1964). Microflora of the Lower Cretaceous mannville group, east-central alberta. Res. Counc. Bull. Alta. 15, 1–239.
Smith, A. H. V., and Butterworth, M. A. (1967). Miospores in coal seams of the carboniferous of great britain. Special Pap. Palaeontol. 1, 1–324.
Smith, D. G. (1974). Late Triassic pollen and spores from the kapp toscana formation, hopen, svalbard: A preliminary account. Rev. Palaeobot. Palynology 17, 175–178. doi:10.1016/0034-6667(74)90098-0
Song, Z. C., Shang, Y. K., Liu, Z. S., Huang, P., Wang, X. F., Qian, L. J., et al. (2000). “The Mesozoic spores and pollen,” in Fossil spores and pollen of China. Z. C. Song, Y. K. Shanget al. (Beijing: Science Press), 2. (in Chinese).
Strullu-Derrien, C., McLoughlin, S., Philippe, M., Mork, A., and Strullu, D. G. (2012). Arthropod interactions with bennettitalean roots in a triassic permineralized peat from hopen, svalbard archipelago (arctic). Palaeogeogr. Palaeoclimatol. Palaeoecol. 348–349, 45–58. doi:10.1016/j.palaeo.2012.06.006
Sun, G., Meng, F. S., Qian, L. J., and Ouyang, S. (1995). “Triassic floras,” in Floras of geological age in China. Editor X. X. Li (Guangzhou: Guangdong Science and Technology Publishing House Press), 229–259. (in Chinese with translation into English by the authors).
Sun, Y. D., Wignall, P. B., Joachimski, M. M., Bond, D. P. G., Grasby, S. E., Lai, X. L., et al. (2016). Climate warming, euxinia and carbon isotope perturbations during the Carnian (Triassic) crisis in South China. Earth Planet. Sci. Lett. 444, 88–100. doi:10.1016/j.epsl.2016.03.037
Sun, Y. W., Li, X., Liu, Q. Y., Zhang, M. D., Li, P., Zhang, R., et al. (2020). In search of the inland carnian pluvial event: Middle–Upper Triassic transition profile and U-Pb isotopic dating in the Yanchang Formation in Ordos Basin, China. Geol. J. 55 (7), 4905–4919. doi:10.1002/gj.3691
Taylor, T. N., Taylor, E. L., and Krings, M. (2009). Paleobotany: The Biology and evolution of fossil plants. 2nd ed. San Diego: Academic Press.
Tokunaga, S., Oshima, H., and Ito, Y. (1977). Recent status and problems of pollen stratigraphy. J. Geogr. (Chigaku Zasshi) 86 (2), 73–79. doi:10.5026/jgeography.86.2_73
Tong, J. N., Chu, D. L., Liang, L., Shu, W. C., Song, H. J., Song, T., et al. (2019). Triassic integrative stratigraphy and timescale of China. Sci. China Earth Sci. 62, 189–222. (in Chinese). doi:10.1007/s11430-018-9278-0
Tuzhikova, V. I. (1975). “New data on spore-pollen complexes of the Ladinian deposts of the urals,” in New miospores, foraminifera, ostracods and conodonts of the paleozoic and mesozoic of the urals. Collected articles on the problems of stratigraphy. Trudy uralskii nauchnyi tsentralnykh, instituta geologii i geokhimii. Editors N. P. Malachova, and B. I. Chuvashev (Akademiya Nauk SSR), 119, 3–13.
Tuzhikova, V. I. (1979). “On the age and extent of the surakaisk suite and bukobaisk series of the southern cis-urals,” in Triassic stratigraphy of the urals and cis-urals. Symposium on the problems of stratigraphy. Trudy uralskii nauchnyi tsentralnykh, instituta geologii i geokhimii. Editors V. I. Tuzhikova, and G. N. Papulov (Sverdlovsk: Akademiya Nauk SSR), 147, 26–51.
Tuzhikova, V. I.(1980). “On the dating of the deposits of the turinsk phytostratigraphy horizon, turinsk and bitkuyevskaya suites of the suhoyaksk region chelyabinsk basin,” in New data on the triassic stratigraphy of the paleo-urals. Akademiya nauk SSSR, uralskii nauchnyi tsentralnykh, trudy instituta geologii i geokhimii. Editors V. I. Tuzikova, and G. N. Papulov
Varyukhina, L. M. (1961). Spores and pollen of triassic deposites of the southern pechora basin. Dokl. Akad. Nauk. SSSR 138 (3), 631–634.
Vasilevskaya, N. D. (1972). “The Late Triassic flora of svalbard,” in Mesozoic deposits in svalbard. Editors V. N. Sokolov, and N. D. Vasilevskaya (NIIGA: Leningrad Press), 27–63.
Vinogradova, K. V. (1974). Miospore assemblages of Triassic sediments of southern mangyshlak. Dokl. Akad. Nauk. SSSR 215 (1), 15–18.
Visscher, H., and Brugman, W. A. (1981). Ranges of selected palynomorphs in the alpine Triassic of Europe. Rev. Palaeobot. Palynology 34 (1), 115–128. doi:10.1016/0034-6667(81)90069-5
Visscher, H., and van der Zwan, C. J. (1981). Palynology of the circum-mediterranean triassic: Phytogeographical and palaeoclimatological implications. Geol. Rundsch. 70, 625–634. doi:10.1007/BF01822140
Wang, Y. D., Jiang, D. X., and Xie, X. P. (2003). Late Triassic palynoflora and its environmental significance of Tuweihe Shaanxi. Acta Sedimentalogica Sin. 21 (3), 434–440. (in Chinese with English abstract). doi:10.14027/j.cnki.cjxb.2003.03.011
Wang, Y. Y., Zhang, B., Shi, Z. Q., and Yi, H. S. (2012). Oxygen and carbon isotopic records of the Late Triassic Carnian pluvial event deposits in northwestern Sichuan Province. J. Palaeogeogr. 14 (3), 375–382. (in Chinese with English abstract). doi:10.7605/gdlxb.2012.03.011
Wang, Z. Q. (1991). Advances on the permo-triassic lycopods in North China. I. An Isoetes from the mid-Triassic in northern Shaanxi Province. Palaeontogr. Abt. B 222 (1–3), 1–30.
Warrington, G. (1979). A derived late permian palynomorph assemblage from the Keuper marl (late triassic), of west somerset. Proc. Ussher Soc. 4 (3), 299–302.
Warrington, G. (1970). “Palynology of the trias of the langford lodge borehole,” in Geology of belfast and the lagan valley. Editors P. I. Manning, J. A. Robbie, H. E. Wilson, and E. Hull (Geological Survey of Northern Ireland), 44–51.
Wilson, K. M., Pollard, D., Hay, W. W., Thompson, S. L., and Wold, C. N. (1994). General circulation model simulations of Triassic climates: Preliminary Results, 288. Special Paper of the Geological Society of America, 91–116. doi:10.1130/SPE288-p91
Xie, X. Y., and Heller, P. L. (2013). U-Pb detrital zircon geochronology and its implications: The early late triassic Yanchang Formation, south Ordos Basin, China. J. Asian Earth Sci. 64, 86–98. doi:10.1016/j.jseaes.2012.11.045
Xu, Y. L., and Zhang, W. P. (1980). “Jurassic spores and pollen,” in Institute of Geology, Chinese academy of geological SciencesMesozoic stratigraphy and palaeontology of the shaanxi-gansu-ningxia basin (Beijing: Geological Publishing House Press), 1, 144–186. (in Chinese).
Yang, R. C., Jin, Z. J., van Loon, A. J., Han, Z. Z., and Fan, A. P. (2017). Climatic and tectonic controls of lacustrine hyperpycnite origination in the Late Triassic Ordos Basin, central China: Implications for unconventional petroleum development. AAPG Bull. 101, 95–117. doi:10.1306/06101615095
Yang, S. R., Liu, J., and Zhang, M. F. (1995). Conodonts from the "Falang Formation" of southwestern Guizhou and their age. J. Stratigr. 19 (3), 161–198. (in Chinese with English abstract). doi:10.19839/j.nki.dcxzz.1995.03.001
Zhang, J. G., Lenz, O. K., Wang, P. J., and Hornung, J. (2021a). The eco-plant model and its implication on Mesozoic dispersed sporomorphs for bryophytes, pteridophytes, and gymnosperms. Rev. Palaeobot. Palynology 293, 104503. doi:10.1016/j.revpalbo.2021.104503
Zhang, K., Liu, R., Liu, Z. J., and Li, L. (2021b). Geochemical characteristics and geological significance of humid climate events in the Middle-Late Triassic (Ladinian-Carnian) of the Ordos Basin, central China. Mar. Petroleum Geol. 131 (5), 105179. doi:10.1016/j.marpetgeo.2021.105179
Zhang, L. J. (1984). “Late Triassic spores and pollen from central Sichuan,” in Palaeontologia sinica (Beijing: Science Press). (in Chinese with English abstract).
Zhang, R., Jin, Z. J., Liu, Q. Y., Li, P., Huang, Z. K., Shi, J. Y., et al. (2019). Astronomical constraints on deposition of the Middle Triassic chang 7 lacustrine shales in the Ordos Basin, central China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 528, 87–98. doi:10.1016/j.palaeo.2019.04.030
Zhao, X. D., Zheng, D. R., Xie, G. W., Jenkyns, H. C., Guan, C. G., Fang, Y. N., et al. (2020). Recovery of lacustrine ecosystems after the end-Permian mass extinction. Geology 48 (6), 609–613. doi:10.1130/G47502.1
Zhu, Z. C., Kuang, H. W., Liu, Y. Q., Benton, M. J., Newell, A. J., Xu, H., et al. (2020). Intensifying aeolian activity following the end-Permian mass extinction: Evidence from the Late Permian–Early Triassic terrestrial sedimentary record of the Ordos Basin, North China. Sedimentology 67, 2691–2720. doi:10.1111/sed.12716
Keywords: Carnian pluvial event, arid climate, megamonsoon, Triassic, Ordos inland basin, palynology, Ladinian–Carnian boundary, Yanchang Formation
Citation: Li X, Zhang Y, Sun Y, Shi X and Zhang S (2023) Vegetation changes and climate shift during the latest Ladinian to the early Carnian: Palynological evidence from the Yanchang Formation, Ordos Basin, China. Front. Earth Sci. 10:1008707. doi: 10.3389/feart.2022.1008707
Received: 01 August 2022; Accepted: 21 November 2022;
Published: 01 February 2023.
Edited by:
Xing Xu, Yunnan University, ChinaReviewed by:
Li Tian, China University of Geosciences Wuhan, ChinaDaoliang Chu, China University of Geosciences Wuhan, China
Copyright © 2023 Li, Zhang, Sun, Shi and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuewu Sun, c3VueXVld3VAamx1LmVkdS5jbg==