 Jianping Zhang1,2*
Jianping Zhang1,2* Ying Lv1
Ying Lv1 Lupeng Yu3*Miao Tang4Ming Huang4Konglan Shao1
Lupeng Yu3*Miao Tang4Ming Huang4Konglan Shao1 Xiujia Huan1Changhui Wen1
Xiujia Huan1Changhui Wen1 Yajie Dong1Ming Jiang4Kunyu He4Xue Yan4Maolin Ye5,6Naiqin Wu1
Yajie Dong1Ming Jiang4Kunyu He4Xue Yan4Maolin Ye5,6Naiqin Wu1 Houyuan Lu1,2,7
Houyuan Lu1,2,7- 1Key Laboratory of Cenozoic Geology and Environment, Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing, China
- 2Innovation Academy of Earth Science, Chinese Academy of Sciences, Beijing, China
- 3Luminescence Research Laboratory, Shandong Provincial Key Laboratory of Water and Soil Conservation and Environmental Protection, School of Resource and Environmental Sciences, Linyi University, Linyi, China
- 4Institute of Cultural Relics and Archaeology of Chengdu, Chengdu, China
- 5School of History and Culture, Lanzhou University, Lanzhou, China
- 6Institute of Archaeology, Chinese Academy of Social Sciences, Beijing, China
- 7College of Earth and Planetary Sciences, University of Chinese Academy of Sciences, Beijing, China
Identifying when agricultural expansion has occurred and how it altered the landscape is critical for understanding human social survival strategies as well as current ecological diversity. In the present study, phytolith records of three profiles from the Baodun site area were dated to the period from 7,500 to 2,500 aBP by optically stimulated luminescence and 14C dating, providing the first evidence that the Asian cultivated rice (Oryza sativa) progenitor was distributed in the Chengdu Plain as early as 7,500 aBP. The percentage of rice bulliform with ≥9 scales and the concentration of rice phytoliths sharply increased by approximately 4,200 aBP, suggesting that rice cultivation occupied a dominant position in survival strategy no later than approximately 4,200 aBP, which might be driven by climate deterioration in eastern China. The results further showed that the proportion of Bambusoideae phytoliths increased synchronously with the increase in the proportion of rice phytoliths, suggesting that the vegetation structure near the site was changed intentionally as a consequence of increasing rice agricultural activity since 4,200 aBP. The present study contributes to a deeper understanding of the distribution of wild rice and rice farming throughout the Baodun culture in the Chengdu Plain, and it also provides a glimpse of how humans intentionally changed the vegetation landscape on a local scale.
Introduction
Agricultural expansion is one of the major drivers of human culture development (Gowdy and Krall, 2014). Reconstructing the time of the spread of domesticated crops can help us understand the climatic and other environmental factors that governed the range of expansion of these crop species (Gutaker et al., 2020), as well as the relationship between farming activities and the consequent landscape changes (Ellis, 2011; Woodbridge et al., 2014; Zheng et al., 2021).
Rice (Oryza sativa L.) is a major staple crop for half of the global population (Wang et al., 2018). It is a highly diversified crop cultivated in a wide ecological range, spanning tropical and temperate zones of Asia, from permanent to seasonal wetlands (Fuller et al., 2010; Fuller et al., 2011). However, the distribution of wild rice (Oryza rufipogon and/or O. nivara) in China remains unclear. The reports of possible wild rice distribution can only be traced back to the Tang and Song Dynasties (You, 1987), while evidence of an earlier distribution can only be obtained from agroclimatic studies (Fuller et al., 2010).
While the origins of rice have been the focus of intensive study, less attention has been paid to its spread after domestication, especially from its cradle Middle-Lower Yangtze to southwest China, which contains a variety of ecosystems and diverse landscapes of the Yunnan-Guizhou Plateau and the low lying Chengdu Plain (d’Alpoim Guedes et al., 2013). There is a further lack of description of the process of landscape modification by rice farming among earlier small-scale societies, as evidence of these processes is sparse (Pavlik et al., 2021).
The Chengdu Plain has a suitable climate for rice cultivation which attracted earlier rice farming populations that widely occupied the area (Sun, 2009; Huang et al., 2017). It is believed that the introduction of rice farming into the Chengdu Plain occurred relatively late, in the time of the Baodun culture (4,600–3,700 cal aBP) from the Middle-Lower Yangtze (Jiang et al., 2002; Jiang et al., 2011; Qin, 2012; d’Alpoim Guedes and Wan, 2015), which was supported by increasing archaeological (Chen et al., 2015) and DNA evidence (Castillo et al., 2016). The development of rice farming in the Chengdu Plain has influenced the regional landscape since the late Neolithic and played a significant role in the formation of prehistoric cultures in Southwestern China, such as Baodun and Sanxingdui cultures (Chen et al., 2015). Although recent studies have outlined the time that rice arrived the plain (d’Alpoim Guedes et al., 2013), however, because of the lack of comparable chronological records from natural deposits and culture layers, the development of rice agriculture and its impact on vegetation are still unclear. In particular, whether wild rice spread into the Chengdu Plain during the Holocene has not been clarified to date.
Phytoliths can be used not only to identify individual species but also to reconstruct the process of crop domestication in order to reveal the intensity of agricultural activities (Piperno, 2006). Specifically, the proportion of bulliform phytoliths with ≥9 fish scale patterns largely differ between wild rice habitats and cultivated rice paddies (Huan et al., 2018; Huan et al., 2020). Bulliform phytoliths can be used to assess the degree and rate of rice domestication to some extent (Huan et al., 2015; Huan et al., 2018). Reconstruction of the quantitative relationship between crop domestication and natural vegetation changes in the same profiles of the same site can provide new evidence for human impact on the ecological environment.
Here, we reported the results of optically stimulated luminescence (OSL) and radiocarbon dating of phytoliths derived from archaeological deposits of the Baodun site approximately 4,500–2,500 cal aBP and a successive natural deposit profile that extends to 7500 aBP in the Chengdu Plain. Based on phytolith assemblage identification, this study revealed the earliest record of wild rice in the Chengdu Plain to date and established a complete rice domestication sequence combined with natural plant changes, which provides new evidence for understanding how humans intentionally influenced their environment.
Materials and Methods
Regional Setting
The Chengdu Plain, which covers an area of approximately 8,400 km2, is the largest fluvial plain in Southwestern China (Liang et al., 2014; Huang et al., 2019a) (Figure 1A). It is a composite alluvial plain formed by the Minjiang River, Tuojiang River, and its tributaries in the Northwest Sichuan Plateau. The plain extends from northeast to southwest, with an altitude of approximately 400–750 m, and belongs to the subtropical monsoon climate (Liu et al., 2004). The average annual temperature is 15–16 °C, and the average annual precipitation is 1,000–1,300 mm (Luo et al., 2008). At present, this area is an important rice farming area in Southwestern China.
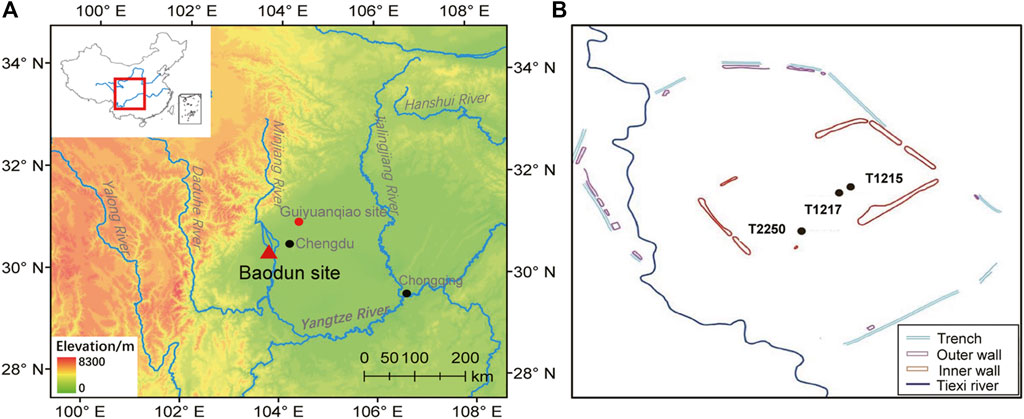
FIGURE 1. (A) Location of the study site, and (B) sampling positions of the three profiles in the Baodun site. The black circles in (A) indicate modern cities, and the red circle indicates the archaeological site in this study.
The chronological order of Neolithic cultures in the Chengdu Plain is: 1) the Baodun culture (4,600–3,700 aBP), which is equivalent to the Longshan period of the Central Plains (Huang et al., 2019b); 2) the Sanxingdui culture (3,700–3,000 aBP), an important archaeological culture after the Baodun culture and is equivalent to the fourth phase of the Erlitou culture in the Central Plains and to the second phase of Yinxu, represented by the Sanxingdui site in Guanghan (Liu et al., 2004) (Figure 1); and 3) the Shierqiao culture (3,200–2,600 aBP), from the early Western Zhou Dynasty to the early Spring and Autumn period, represented by the Shierqiao and Jinsha sites (Li et al., 1987; Zhu et al., 2004).
The Baodun culture (4,600–3,700 aBP), which is named by the site of Baodun, is currently the earliest archaeological culture found in the Chengdu Plain in Southwestern China. The Baodun site (E103°45′, N30°26′, 472–474 m) is located in Baodun village, Xinping Town, which is ∼5 km northwest of Xinjin County, Chengdu City, Sichuan Province (Figure 1). The site is currently the biggest prehistoric site that characterises Baodun culture.
Sample Collection
In the present study, we analysed 105 phytolith samples from three profiles. Among the profiles, two were from excavations T1217 and T2250, and the third was from natural deposits beneath the oldest culture layers of T1215. All three profiles had comparable stratigraphic sequences.
Deposits from the first profile (T1217, 130 cm in thickness) could be divided into 12 layers from bottom to top according to the structure of the stratigraphy, soil colour, and archaeological remains: 1) layers 12–11 (190–170 cm), grey-green clay with brown soil blocks and Fe-Mn nodules, transition layers from natural deposits to the early Baodun culture; 2) layers 10–9 (170–123 cm), brown clay with Fe-Mn nodules, middle to late Baodun culture; 3) layers 8–5 (123–70 cm), grey-brown clay, Han Dynasty; 4) layers 4–3 (70–41 cm), brown clay, Tang-Song Dynasty; 5) layer 2 (41–33 cm), brown clay, Ming-Qing Dynasty; 6) layer 1 (33–0 cm), modern cultivated layer. Four samples were taken at 5 cm intervals in the 12th layer, 30 samples were taken at 2 cm intervals in the 11th–8th layer, 16 samples were taken at 5 cm intervals in the 7th–2nd layer, and three samples were taken at 10 cm intervals in the 1st layer. Two charcoals for dating were collected at 140–142 cm and 136–138 cm (Figure 2).

FIGURE 2. Stratigraphic sequences of the three sampling profiles (T2250, T1217, and T1215).
T1215 (75 cm in thickness) is a natural deposit profile just beneath the oldest culture layer of the Baodun site. It could be divided into four layers from bottom to top: 1) layers 4 (235–230 cm), black-dark grey clay; 2) layers 3 (230–205 cm), grey clay; 3) layers 2 (205–170 cm), grey clay grey clay with few Fe-Mn nodules; 4) layers 1 (170–160 cm), grey clay with Fe-Mn nodules, which are transition layers from natural deposit to early Baodun culture. Fifteen samples were taken at 5 cm intervals in the 4th–1st layer, and three OSL samples were collected for dating at 228, 204, and 168 cm (Figure 2).
Profile T2250 (220 cm in thickness) could be divided into seven layers from bottom to top: 1) layers 7 (200–180 cm), grey-green clay, natural deposit; 2) layers 6 (180–120 cm), grey-green clay with brown soil blocks and Fe-Mn nodules, Baodun culture; 3) layers 5–3 (120–64 cm), grey-green clay to brown clay with few Fe-Mn nodules, Han Dynasty; 4) layers 2 (64–30 cm), grey-green clay, Tang-Song Dynasty; 5) layer 1 (30–0 cm), modern cultivated layer. Five samples were taken at 5 cm intervals in the 7th layer, 30 samples were taken at 2 cm intervals in the 6th–5th layer, 18 samples were taken at 5 cm intervals in the 4th–2nd layer, and three samples were taken at 10 cm intervals in the 1st layer. Four OSL samples were collected for dating at 200, 173, 116, and 62 cm. We analysed a total of 37 samples from layers 7 to 5 in T2250 (Figure 2).
Phytolith Extraction and Identification
Phytoliths were extracted from soil samples according to established methods (Zhao and Piperno, 2000; Piperno, 2006) with minor modifications. Initially, approximately 2 g of each sample was weighed and treated with 30% H2O2 and 15% HCl to remove organic matter and carbonates. The samples were then subjected to heavy liquid flotation using ZnBr2 (density = 2.35 g/cm3) to separate the phytoliths, which were subsequently mounted on a slide using Canada Balsam. After air drying, the phytoliths on the slide were counted and identified using a Leica microscope at ×400 magnification. More than 400 phytolith particles were identified in each sample and recorded according to published references and criteria (Wang and Lu, 1993; Lu et al., 2002; Lu et al., 2006). Phytolith abundance was expressed as percentages of all phytoliths counted. In particular, for samples with rice phytoliths, the slides were scanned until 50 rice bulliforms with clear and countable scales were observed (except in 16 out of 105 samples that contained minor phytoliths) in order to calculate the proportion of rice bulliform phytoliths with ≥9 scales (Huan et al., 2015; Huan et al., 2020). The abundance of bulliform with ≥9 scales was expressed as percentages of only bulliform phytoliths counted.
Methods of Chronology Assessment
A total of nine dating samples were collected from the three profiles, of which two were used for 14C dating by the Center for Applied Isotope Studies, University of Georgia, and the rest were tested by OSL from the Luminescence Research Laboratory, Linyi University, China. The dates that were obtained by the 14C method were calibrated by OxCal 4.4 using the IntCal 20 atmospheric curve (Reimer et al., 2020) and are presented in Table 1 (Lv et al., 2021).
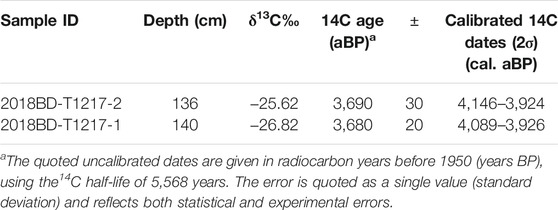
TABLE 1. 14C dating results from the Baodun site (Lv et al., 2021).
For OSL dating, quartz was purified by the following process: after the removal of carbonates and organic materials, the samples were subjected to wet sieving (63–90 μm), density liquid (SPT, 2.62–2.70 g/cm3), HF etching (40 min), HCl rinse (40 min), and dry sieving (63 μm) (An et al., 2020). Small aliquots (2 mm diameter) were used to effectively demonstrate the scatter in equivalent dose (De). The Des were measured with a Risø TL/OSL DA-20 reader, and the Single Aliquot Regenerative dose (SAR) (Murray and Wintle, 2000; 2003) protocol and the standardised growth curve (SGC) method (Roberts and Duller, 2004) were applied. The detailed protocol and parameters followed were referred from An et al. (2020). For each sample, five aliquots were measured with the SAR protocol, and the five growth curves of each aliquot of a sample were averaged to build the SGC for each sample. Then normalised natural signals of additional 14–16 aliquots were matched to their SGCs to calculate SGC Des. Finally, the De of each sample was calculated based on the SAR Des and SGC Des (n = 19–21) using the central age model (Galbraith et al., 1999). The decay curves (dominated by the fast component), growth curves, and SGCs of samples are displayed in Figures 3A,B. De distribution plots (Abanico plots) (Dietze and Kreutzer, 2020) of all the samples (Figures 3C–I) suggested that the OSL signals were well bleached before the burial. The content of elements was measured using ICP-MS (U and Th) and ICP-OES (K). Considering the high groundwater level, water content was estimated based on the actual measured and saturated values (Table 1). The dose rates and ages are calculated by the Dose Rate and Age Calculator (Durcan et al., 2015) and are shown in Table 2.
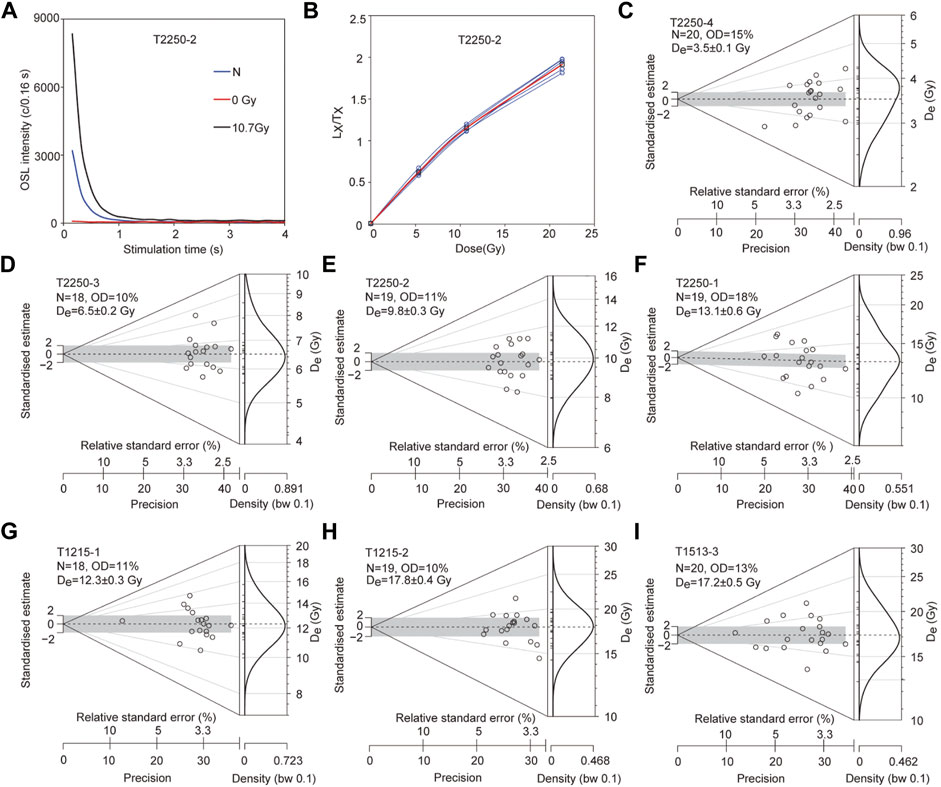
FIGURE 3. (A) Decay curves, (B) growth curves (red for SGC, and (C–I) De distributions. SGC: Standardised growth curve; De: The scatter in equivalent dose.
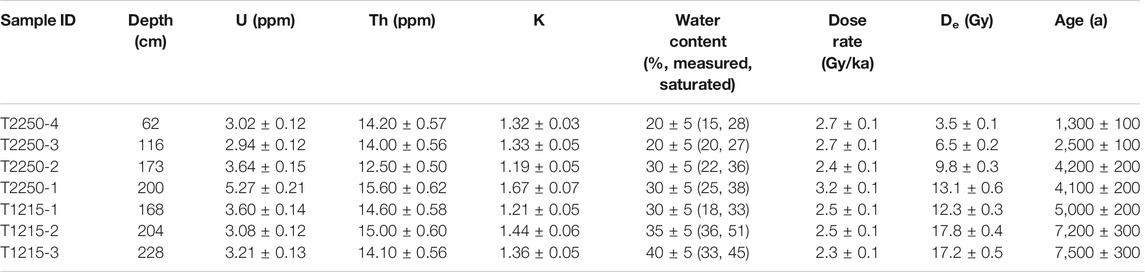
TABLE 2. Optically stimulated luminescence (OSL) dating results of the Baodun Site.
Results
All nine dating samples from the sites successfully yielded credible dates (Tables 1, 2). Generally, all these direct dates fit well with the cultural affiliations of their contexts. Dates of T1215 were found to range from 7,500 to 5,000 BP, which is completely beyond the cultural period of the site. The oldest age of the lowest layer of T1215 (7500 BP) is consistent with that of the palaeowood remains dated at 7,400 cal BP (Huang et al., 2019b). Two dating results of T1217 (4,000 cal aBP) are corresponding with the late Baodun period. Dates of T2250 fell into the range of 4,200–2,500 aBP, covering the periods of middle to late phases of the Baodun culture.
All 105 samples from the Baodun sites yielded abundant phytoliths (Figure 4). In total, 26 phytolith morphotypes were identified, of which four could be confirmed from crops, including double-peaked, bulliform, and parallel-bilobate phytolith types of rice, and epidermal long cell phytoliths from the upper lemma and palea of millets (broomcorn and foxtail millet) (Figure 4). Other typical morphotypes that could be identified to the subfamily level and lower levels were the long saddle phytoliths of Bambusoideae, rondel and trapeziform sinuate phytoliths of Pooideae, bulliform phytoliths of Arundiaceae, and bilobate phytoliths of Panicoideae, as well as woody phytoliths and Cyperaceae phytoliths.
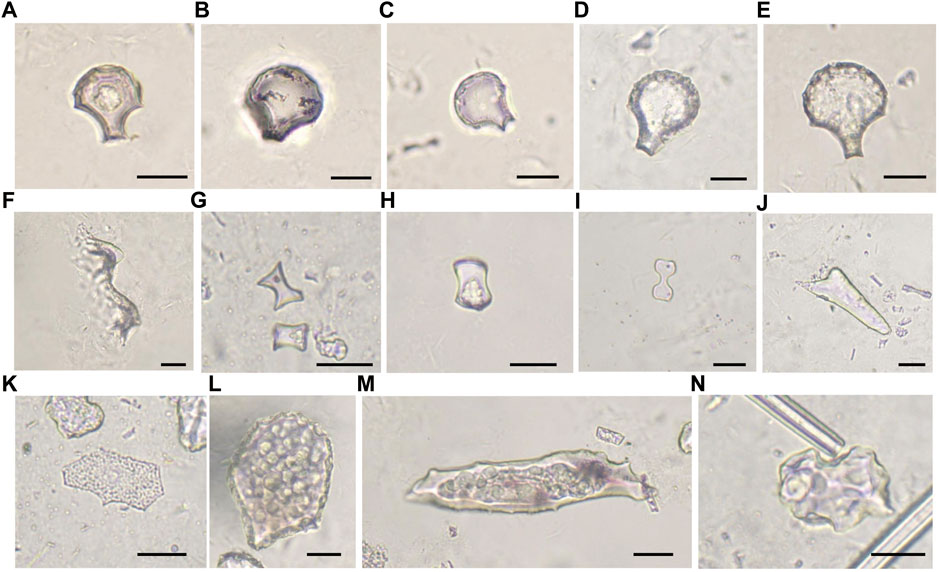
FIGURE 4. Main phytolith morphotypes at the Baodun site. (A–E) rice bulliform phytoliths from rice leaves with increased scale numbers, (F) double-peaked cells from rice husk, (G–H) long saddle phytoliths, (I) bilobate phytoliths, (J) acicular hair cell phytoliths, (K) achene phytoliths belonging to Cyperaceae, (L) scutiform-bulliform phytoliths belonging to reeds, and (M–N) woody phytoliths.
The most significant feature of crop phytolith assemblages from the three profiles was the presence of abundant rice phytoliths (rice bulliform and double-peaked types), ranging from approximately 0.2–5.5%, in almost every sample 7,500 aBP (Figure 5). To reconstruct the farming development process, all 105 samples were further analysed to count the number of scales on the edge of the rice bulliform phytoliths. For each sample, almost 50 bulliform phytoliths with clear and countable scales were carefully observed. The average proportion of bulliform phytoliths with ≥9 scales remained at a low level of 20.2 ± 5.1% between 7,500 and 5,000 aBP in T1215. However, their proportion increased over 25% during 5,000–4,000 aBP in T1217, including the early-middle Baodun period (Figure 5), and sharply increased to 48.6 ± 8.5% after 4200 aBP, as evident in T2250 (Figure 6).
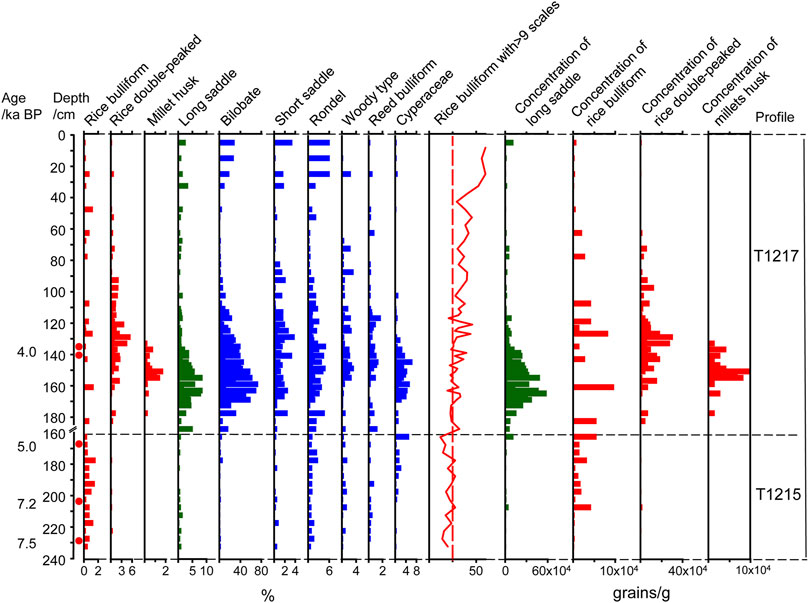
FIGURE 5. Phytolith assemblages of T1215 and T1217 profiles.
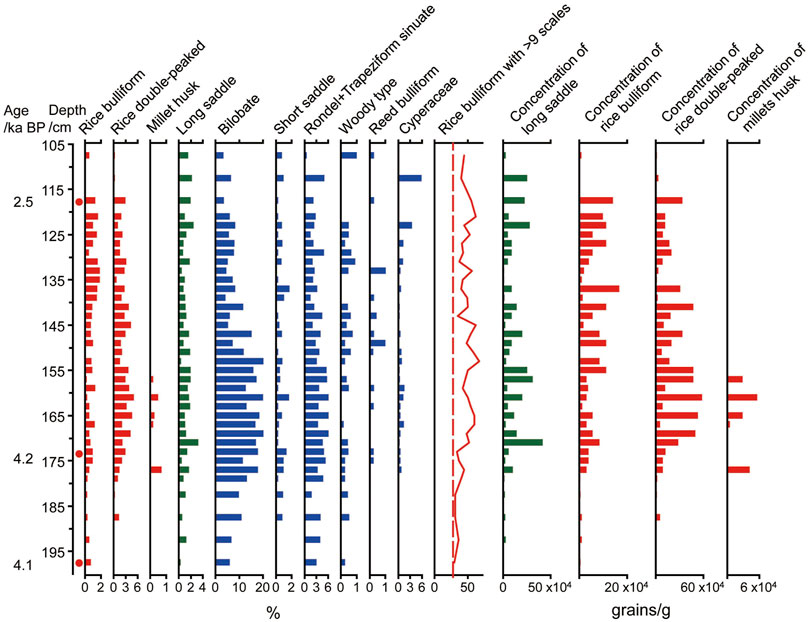
FIGURE 6. Phytolith assemblages of T2250 profile.
Similarly, the percentage and concentration of rice phytoliths and millet husks increased synchronously in both T1217 and T2250 (Figures 5, 6). Especially, the concentrations of rice bulliform, double-peaked and millet husk in T1217 simultaneously reached the peak of 9.6 × 104, 30 × 104, and 9.6 × 104 grains/g at approximately 4,000 aBP, respectively. The same trend was also evident in T2250, as the concentrations of rice double-peaked and millet husk reached peaks of 57.9 × 104 and 5.5 × 104 grains/g at approximately 4,200 aBP.
It is also worth noting that the assemblage of phytoliths varied greatly from natural to cultural layers (i.e., in T1215 and T1217). It is characterised by a synchronous increase in the abundance of rice phytoliths and long saddle phytoliths belonging to Bambusoideae, bilobate and cross phytoliths belonging to Panicoideae, and rondel and trapeziform sinuate phytoliths belonging to Poaceae, Cyperaceae, and woody phytoliths. In particular, the concentration of rice phytoliths and long saddles also changed synchronously in the profiles of T1217 and T2250, which reached the highest value soon after 4,200 cal aBP, after which it decreased again, revealing a strong relationship between agricultural activities and natural changes in vegetation near the site. The concentration of long saddles sharply increased from 5.8 to 57.9 grains/g and peaked synchronously with the concentration of rice phytoliths at approximately 4,000 aBP in T1217. Similarly, in T2250, the concentration of long saddles obviously increased after 4,200 aBP, from 2.3 to 41.3 grains/g, along with the increase in rice phytolith concentration.
Discussion
Studies on Neolithic agriculture in the Chengdu Plain have focused primarily on documenting the cultivation and spread of crops and were limited to the Baodun period (d’Alpoim Guedes et al., 2013; d’Alpoim Guedes and Wan, 2015). Since there are a limited amount of palaeovegetation records before and after the Baodun period, the introduction and development of rice farming in the Plain, as well as the vegetation evolution history influenced by rice farming are still a subject of discussion (Jiang, 2009; Jiang et al., 2009; Qin, 2012). The phytolith records analysed in the present study dated from 7,500 to at least 2,500 aBP provide chronological records of rice farming development and landscape alteration near the study site.
Wild Rice Distribution
The proportion of bulliform phytoliths with ≥9 fish scale patterns was largely different in wild rice soils (17.5 ± 8.3%) and domesticated rice soils (57.6 ± 8.7%), which can be used as an indicator to assess the degree and rate of rice domestication to some extent (Huan et al., 2015). According to our records, the proportion of rice bulliform phytoliths with ≥9 fish scales was maintained at approximately 15–25% throughout the T1215 profile, which was the first substantial evidence of the presence of wild rice in the Chengdu Plain as early as 7,500 BP. Our results, combined with the presence of Cyperaceae and reed phytoliths from the T1215 profile and other palaeoenvironmental evidence (Huang et al., 2019b), indicated that the environment near the Baodun site was permanent to seasonal wetlands in association with lake and marsh biomes, which contained a spectrum of ecological niches suitable for the rice wild progenitor complex (Oryza rufipogon and O. nivara).
The Development of Rice Farming
The observed sharp increase in the percentage of rice phytoliths with ≥9 fish scales beyond the distribution range of wild rice data (more than 50%), with simultaneous increases in the percentage of both rice husk and leaves after 4,200 cal BP in T 2250 and T1217 (Figures 5, 6), suggested an increase in the area of domesticated rice habitat and a decrease in the area of wild rice habitat. This, combined with the synchronous increase in the percentage and concentration of rice phytoliths, indicated that rice farming with a minor millet cultivation survival strategy occupied a dominant position no later than 4,200 aBP at the Baodun site.
We suggested that this might be a consequence of social development, and it may more likely be attributed to the rapid expansion of rice cultivation as suggested in a previous study (Gutaker et al., 2020). Gutaker et al. (2020) speculated that the climatic change (aridification) might have resulted in the southward dispersal of rice (agricultural communities). This study used absolute dating and confirmed the abrupt increases in rice abundance and domesticated traits since approximately 4,200 aBP in sedimentary archives in southwestern China, offering archaeological, biological, and chronological evidence for this suspension.
Since 4,200 aBP, Neolithic cultures and rice agriculture (Wang, 2004; An et al., 2005; Guedes et al., 2015; Wang et al., 2016) declined in East Asia/eastern China, and debates remained on what drove the decline, e.g., climate deterioration (An et al., 2005; Marcott et al., 2013), flood and wars (Wang 2004; Shen et al., 2015), and from where the people migrated (Gutaker et al., 2020). The rapid development of rice cultivation in Chengdu Plain in 4,200 aBP may shed light on this question. The aridification since approximately 4,200 aBP could not sustain the former agricultural systems and cause the southward shift of the agricultural belts, which further drove the southward migration of people, resulting in the southward dispersal of rice agricultural communities (Silva et al., 2015; Castillo et al., 2018; Gutaker et al., 2020). At this time, the climate in the Chengdu Plain was relatively mild (Luo et al., 2007; Luo et al., 2008) with ample water sources, presenting an ideal environment for wetland rice production (d’Alpoim Guedes et al., 2013). However, owing to the lack of regional high-resolution evidence of environmental change and population migration, further research is needed.
Although rice has appeared as early as approximately 4,600 BP at the site of study (d’Alpoim Guedes et al., 2013), the present study indicated low proportions of rice bulliform phytoliths with ≥9 scales between 5,000 and 4,200 aBP. We propose that the presence of wild rice had caused inevitable hybridisation with the domesticated rice (Wang et al., 2017) and the wild rice might have degenerated the traits of domesticated rice. Moreover, the area of rice cultivation during the primary stage of the Baodun culture was small, which may also be a reason for the reduced proportion of rice bulliform phytoliths with ≥9 scales. A previous archaeobotanical study also indicated that domesticated rice accounted for only 50% of the total rice remains during the Baodun period (d’Alpoim Guedes et al., 2013). However, rice remains were not distinguished by culture phases in that study. Therefore, the proportion of domesticated rice cultivated during the early stage of the Baodun culture was not exactly determined. Another evidence of a single broken rice spikelet was found in the Guiyuanqiao site in the Chengdu Plain (d’Alpoim Guedes and Wan, 2015). However, it was also difficult to determine whether it was wild or domesticated.
Landscape Changes as a Consequence of Neolithic Rice Cultivation
Our data further indicated that human-induced vegetation changes in our study region can be traced back to as early as 4,200 aBP. The vegetation transformations that occurred at this time involved a simplification of the Poaceae vegetation structure, particularly the increase in Bambusoideae (long saddle phytoliths), Panicoideae (bilobate and cross phytoliths), and Pooideae (rondel, trapeziform sinuate phytoliths), based on a comparison with phytolith records from T1215, T1217 and T2250 (Figures 5, 6). It is suggested that because of the development of rice farming, ancient people altered the surrounding vegetation, for example the irrigation expanded the habitats suitable for hydrophilous plants, such as reeds and some Cyperaceae species. Specifically, agriculture might benefit the expansion of bamboo. On the one hand, the expansion of rice paddy caused deforestation, which provided space for the expansion of bamboo. On the other hand, the utilisation of bamboo as agricultural tools and building materials promoted the development of rice production. Our records, combined with previous charcoal analyses, indicated that bamboo was widely utilised at the Baodun site (Yan et al., 2016).
The combined increase in the amount of evidence of rice and Bambusoideae further suggests that the farmhouse forest (Linpan in Chinese) landscape with the bamboo forest as one of the main vegetation types (Dajiang et al., 2011) may have appeared no later than 4,200 aBP. It also contributes to understanding the evolution of the Linpan community, which is a traditional rural settlement unit that has close relations among rice farming, daily life, and landscape and has outstanding agricultural heritage conservation values in the Chengdu Plain (Fang and Li, 2011; Yuan et al., 2020).
It should be pointed out that information relating to past plant diversity revealed by phytolith data is limited and incomplete, mainly because different plant taxa have different phytolith productivities (Piperno, 2006). In particular, woody taxa yield less phytoliths that can be identified to the genus level (Wang and Lu, 1993). Although the inherent biases of phytolith analyses cannot be eliminated, phytolith records still represent one of the best data sources to describe the surrounding landscape, especially the herbaceous vegetation landscape, in a wide ecological range, from dry to humid regions (Dickau et al., 2013; Zhang et al., 2013; Liu et al., 2021). The phytolith records described in the present study provide a glimpse of the development of rice farming throughout the Baodun culture in the Chengdu Plain and a description of Poaceae floral compositional changes since 7,500 aBP.
Conclusion
In the present study, phytolith remains collected from natural deposits and culture layers dated from 7,500 to 2,500 aBP at the Baodun site, Southwestern China, were analysed. The evidence indicated that wild rice was already distributed in the area as early as 7,500 aBP. The comparison of phytolith records from natural and archaeological layers indicated that the survival strategy of rice cultivation for the Baodun people occupied a dominant position no later than approximately 4,200 aBP. We suggest that this may be due to the expansion of rice cultivation to southwestern China caused by late Holocene climate deterioration and the subsequent decline of Neolithic culture in eastern China. The intensified farming activities further induced Poaceae vegetation alternation, particularly the expansion of the bamboo forest in areas near the Baodun site. These findings have implications in understanding the distribution of wild rice during the Holocene, as well as in understanding the development of rice farming and human-induced vegetation and landscape changes in past small-scale societies of Southwestern China. Thus, this study contributed to the understanding of the formation and continuance of the ancient Sichuan civilisation.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Author Contributions
JZ conceived and designed the study. JZ and LY compiled archaeological dates. JZ, YL, and LY analysed the data. JZ wrote the manuscript. LY, MT, MH, KS, XH, KH, XY, CW, MY, NW, and HL revised the manuscript.
Funding
This study was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB26000000), National Natural Science Foundation of China (41830322), National Key R&D Program of China (2020YFC1521605), and Youth Innovation Promotion Association of the Chinese Academy of Sciences.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer (XZ) declared a past co-authorship with one of the authors (JZ) to the handling Editor.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank Editage (www.Editage.cn) for English language editing.
References
An, C.-B., Tang, L., Barton, L., and Chen, F.-H. (2005). Climate Change and Cultural Response Around 4000 Cal Yr B.P. In the Western Part of Chinese Loess Plateau. Quat. Res. 63 (3), 347–352. doi:10.1016/j.yqres.2005.02.004
An, P., Yu, L., Wang, Y., Miao, X., Wang, C., Lai, Z., et al. (2020). Holocene Incisions and Flood Activities of the Keriya River, NW Margin of the Tibetan Plateau. J. Asian Earth Sci. 191, 104224. doi:10.1016/j.jseaes.2019.104224
Castillo, C. C., Fuller, D. Q., Piper, P. J., Bellwood, P., and Oxenham, M. (2018). Hunter-gatherer Specialization in the Late Neolithic of Southern Vietnam - the Case of Rach Nui. Quat. Int. 489, 63–79. doi:10.1016/j.quaint.2016.11.034
Castillo, C. C., Tanaka, K., Sato, Y.-I., Ishikawa, R., Bellina, B., Higham, C., et al. (2016). Archaeogenetic Study of Prehistoric rice Remains from Thailand and India: Evidence of Early Japonica in South and Southeast Asia. Archaeol Anthropol. Sci. 8 (3), 523–543. doi:10.1007/s12520-015-0236-5
Chen, T., Jiang, Z., He, K., Yang, Y., Guedes, J. d. A., Jiang, H., et al. (2015). Phytolith Analysis from the Baodun Archaeological Site, Xinjin, Sichuan. Acta Anthropol. Sin. 34 (2), 225–233. 10.16359/j.cnki.cn11-1963/q.2015.0023 .
Dajiang, S., Qibing, C., Tingxing, H., Guangli, L., Dayuan, S., and Qigao, L. (2011). Community Types and the Biodiversity of Farmhouse Forest in Western Sichuan Province. J. Sichuan Agr. Univ. 29 (1), 22–28. 10.3969/j.issn.1000-2650.2011.01.005 .
d’Alpoim Guedes, J., Jiang, M., He, K., Wu, X., and Jiang, Z. (2013). Site of Baodun Yields Earliest Evidence for the Spread of rice and Foxtail Millet Agriculture to South-West China. Antiquity 87 (337), 758–771. doi:10.1017/s0003598x00049449
d’Alpoim Guedes, J., Jin, G., and Bocinsky, R. K. (2015). The Impact of Climate on the Spread of Rice to North-Eastern China: A New Look at the Data from Shandong Province. PLOS ONE 10 (6), e0130430. doi:10.1371/journal.pone.0130430
d’Alpoim Guedes, J., and Wan, J. (2015). Flotation Analysis of Guiyuanqiao Site in Shifang City, Sichuan Province. Sichuan Cult. Relics (5), 81–87+94.
Dickau, R., Whitney, B. S., Iriarte, J., Mayle, F. E., Soto, J. D., Metcalfe, P., et al. (2013). Differentiation of Neotropical Ecosystems by Modern Soil Phytolith Assemblages and its Implications for Palaeoenvironmental and Archaeological Reconstructions. Rev. Palaeobotany Palynology 193, 15–37. doi:10.1016/j.revpalbo.2013.01.004
Dietze, M., and Kreutzer, S. (2020). “plot_AbanicoPlot(): Function to Create an Abanico Plot. Function Version 0.1.11,” in Luminescence: Comprehensive Luminescence Dating Data Analysis. R Package Version 0.9.10. Editors S. Kreutzer, C. Burow, M. Dietze, M. C. Fuchs, C. Schmidt, M. Fischeret al.
Durcan, J. A., King, G. E., and Duller, G. A. T. (2015). DRAC: Dose Rate and Age Calculator for Trapped Charge Dating. Quat. Geochronol. 28, 54–61. doi:10.1016/j.quageo.2015.03.012
Ellis, E. C. (20111938). Anthropogenic Transformation of the Terrestrial Biosphere. Phil. Trans. R. Soc. A. 369, 1010–1035. doi:10.1098/rsta.2010.0331
Fang, Z., and Li, X. (2011). On Causes for Formation of Linpan Culture in West Sichuan Plain. (5), 45–49. doi:10.3969/j.issn.1004-342X.2011.05.013
Fuller, D. Q., van Etten, J., Manning, K., Castillo, C., Kingwell-Banham, E., Weisskopf, A., et al. (2011). The Contribution of rice Agriculture and Livestock Pastoralism to Prehistoric Methane Levels. The Holocene 21 (5), 743–759. doi:10.1177/0959683611398052
Fuller, D., Sato, Y.-I., Castillo, C., Qin, L., Weisskopf, A., Kingwell-Banham, E., et al. (2010). Consilience of Genetics and Archaeobotany in the Entangled History of rice. Archaeol. Anthrop. Sci. 2 (SI), 115–131. doi:10.1007/s12520-010-0035-y
Galbraith, R. F., Roberts, R. G., Laslett, G. M., Yoshida, H., and Olley, J. M. (1999). Optical Dating of Single and Multiple Grains of Quartz from Jinmium Rock Shelter, Northern Australia: Part I, Experimental Design and Statistical Models. Archaeometry 41, 339–364. doi:10.1111/j.1475-4754.1999.tb00987.x
Gowdy, J., and Krall, L. (2014). Agriculture as a Major Evolutionary Transition to Human Ultrasociality. J. Bioecon 16 (2), 179–202. doi:10.1007/s10818-013-9156-6
Gutaker, R. M., Groen, S. C., Bellis, E. S., Choi, J. Y., Pires, I. S., Bocinsky, R. K., et al. (2020). Genomic History and Ecology of the Geographic Spread of rice. Nat. Plants 6 (5), 492–502. doi:10.1038/s41477-020-0659-6
Huan, X., Lu, H., Wang, C., Tang, X., Zuo, X., Ge, Y., et al. (2015). Bulliform Phytolith Research in Wild and Domesticated Rice Paddy Soil in South China. PLOS ONE 10 (10), e0141255. doi:10.1371/journal.pone.0141255
Huan, X., Lu, H., Wang, C., and Zhang, J. (2020). Progress of rice Bulliform Phytolith Research on Wild–Domesticated Characteristics. Acta Palaeontol. Sin. 59 (4), 467–478. 10.1371/journal.pone.0141255 .
Huan, X., Lu, H., Zhang, J., and Wang, C. (2018). Phytolith Assemblage Analysis for the Identification of rice Paddy. Sci. Rep. 8, 10932. doi:10.1038/s41598-018-29172-5
Huang, M., Ma, C., and Zhu, C. (2017). Progress of the Mid-late Holocene Environmental Archaeology in Chengdu plain. J. Palaeogeog. 19 (6), 1087–1098. doi:10.7605/gdlxb.2017.06.084
Huang, M., Zhu, C., Ma, C., He, K., and Xu, J. (2019a). Paleoenvironmental Context of the Evolution of the Baodun Culture at Chengdu Plain, Sichuan Province, China. The Holocene 29 (11), 1731–1742. doi:10.1177/0959683619862031
Jiang, M., Dai, Y., Zhang, Q., and He, K. (2009). Brief Report on Analysis of Flotation Results of Archaeological Excavation in 2009 Year of Baodun Site in Xinjin. Archaeol. Discov. Chengdu (0), 72–86+548–549.
Jiang, M., d’Alpoim Guedes, J., and He, K. (2011). “Flotation Analysis of the Archaeological Excavation of Baodun Site in Xinjin in 2009,” in Archaeological Discoveries of Chengdu in 2009Institute of Cultural Relics and Archaeology of Chengdu (Beijing: Science Press).
Jiang, Z. (2009). Agricultural Transformation and Settlement Changes in Chengdu Plain during the Pre-qin Period. Forum Chinese Cult, China. S2. 187–189.
Jiang, Z., Wang, Y., and Zhang, Q. (2002). A Preliminary Research on the Pre-qin Culture in the Chengdu Plain. J. Archaeol. (1), 1–22.
Li, Z., Weng, S., Zhang, X., Jiang, z., Liu, Z., and Zhou, K. (1987). A Brief Report on the First Phase of Excavation of the 12th Qiao Shang Dynasty Architectural Site in Chengdu. Cult. Relics (12), 1–23+37+99–101.
Liang, B., Zhu, B., and Wang, Q. (2014). Quaternary Geology and Environment in the Chengdu Plain. Beijing: Science Press.
Liu, Y., Chen, J., Chen, Y., Yang, X., Liu, S., Chen, P., et al. (2004). Trial Excavation of Sanxing Village Ruins in Qingbai District, Chengdu. Archaeol. Discov., 255–282.
Liu, Y., Liu, H., Jie, D., Gao, G., Meng, M., and Zhang, G. (2021). Phytolith Morphotypes of Woody Plants and Their Preservation in Soil in the Warm Temperate Humid Zones of China. Quat. Int. 599-600, 158–169. doi:10.1016/j.quaint.2021.03.017
Lu, H.-Y., Wu, N.-Q., Yang, X.-D., Jiang, H., Liu, K.-b., and Liu, T.-S. (2006). Phytoliths as Quantitative Indicators for the Reconstruction of Past Environmental Conditions in China I: Phytolith-Based Transfer Functions. Quat. Sci. Rev. 25, 945–959. doi:10.1016/j.quascirev.2005.07.014
Lu, H., Jia, J., Wang, W., Wang, Y., and Liu, K.-B. (2002). On the Meaning of Phytolith and its Classification in Gramineae. Acta Micropalaeontol. Sin. (4), 389–396. doi:10.3969/j.issn.1000-0674.2002.04.006
Luo, L., Zhu, L., Xiang, F., Yang, W., Kan, A., Luo, H., et al. (2008). Spore-pollen Assemblage and Environmental Changes of the Chengdu Plain during the Late Holocene. Acta Palaeontol. Sin. 47 (2), 195–202. doi:10.3969/j.issn.0001-6616.2008.02.006
Luo, L., Zhu, L., Yang, W., Zhang, Q., Wang, C., Kan, A., et al. (2007). Magnetic Susceptibility Characteristics and Paleoclimate Explanation of Chengdu Plain since 4000a. J. Chengdu Univ. Technol. (Sci. Technol. Edit.) 34 (3), 327–330. doi:10.3969/j.issn.1671-9727.2007.03.017
Lv, Y., Zhang, J., Tang, M., Yang, Z., Huang, M., Jiang, M., et al. (2021). Phytolith Analysis Reveals the Agricultural Development and Environmental Background of the Pre-qin Period in Chengdu Plain-A Case Study of Baodun and Sanxingcun Sites. Quat. Sci. 41 (5), 1475–1488. 10.11928/j.issn.1001-7410.2021.05.21 .
Marcott, S. A., Shakun, J. D., Clark, P. U., and Mix, A. C. (2013). A Reconstruction of Regional and Global Temperature for the Past 11,300 Years. Science 339 (6124), 1198–1201. doi:10.1126/science.1228026
Murray, A. S., and Wintle, A. G. (2003). The Single Aliquot Regenerative Dose Protocol: Potential for Improvements in Reliability. Radiat. Meas. 37 (4-5), 377–381. doi:10.1016/s1350-4487(03)00053-2
Murray, A. S., and Wintle, A. G. (2000). Luminescence Dating of Quartz Using an Improved Single-Aliquot Regenerative-Dose Protocol. Radiat. Measurements 32 (1), 57–73. doi:10.1016/s1350-4487(99)00253-x
Pavlik, B. M., Louderback, L. A., Vernon, K. B., Yaworsky, P. M., Wilson, C., Clifford, A., et al. (2021). Plant Species Richness at Archaeological Sites Suggests Ecological Legacy of Indigenous Subsistence on the Colorado Plateau. Proc. Natl. Acad. Sci. USA 118 (21), e2025047118. doi:10.1073/pnas.2025047118
Piperno, D. R. (2006). Phytolith: A Comprehensive Guide for Archaeologists and Paleoecologists. New York: AltaMira Press.
Qin, L. (2012). “Archaeobotany Research and Expectationon on Chinese Argiculture Origin,” in Archaeological ResearchSchool of Archeaology and Museology Peking University & the Center of the Study of Chinese Archaeology in Peking University (Beijing: Cultural Relics Press), 260–315.
Reimer, P. J., Austin, W. E. N., Bard, E., Bayliss, A., Blackwell, P. G., Bronk Ramsey, C., et al. (2020). The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0-55 Cal kBP). Radiocarbon 62 (4), 725–757. doi:10.1017/rdc.2020.41
Roberts, H. M., and Duller, G. A. T. (2004). Standardised Growth Curves for Optical Dating of Sediment Using Multiple-Grain Aliquots. Radiat. Measurements 38 (2), 241–252. doi:10.1016/j.radmeas.2003.10.001
Shen, H., Yu, L., Zhang, H., Zhao, M., and Lai, Z. (2015). OSL and Radiocarbon Dating of Flood Deposits and its Paleoclimatic and Archaeological Implications in the Yihe River Basin, East China. Quat. Geochronol. 30, 398–404. doi:10.1016/j.quageo.2015.03.005
Silva, F., Stevens, C. J., Weisskopf, A., Castillo, C., Qin, L., Bevan, A., et al. (2015). Modelling the Geographical Origin of Rice Cultivation in Asia Using the Rice Archaeological Database. PLOS ONE 10 (9), e0137024. doi:10.1371/journal.pone.0137024
Sun, H. (2009). Evolution of Prehistoric Grain Species in Sichuan basin: Information Mainly Derived from Archaeological Cultural Interactions. Forum Chin. Cult (2), 147–154.
Wang, H., Vieira, F. G., Crawford, J. E., Chu, C., and Nielsen, R. (2017). Asian Wild rice Is a Hybrid Swarm with Extensive Gene Flow and Feralization from Domesticated rice. Genome Res. 27 (6), 1029–1038. doi:10.1101/gr.204800.116
Wang, J., Sun, L., Chen, L., Xu, L., Wang, Y., and Wang, X. (2016). The Abrupt Climate Change Near 4,400 Yr BP on the Cultural Transition in Yuchisi, China and its Global Linkage. Sci. Rep. 6, 27723. doi:10.1038/srep27723
Wang, W., Mauleon, R., Hu, Z., Chebotarov, D., Tai, S., Wu, Z., et al. (2018). Genomic Variation in 3,010 Diverse Accessions of Asian Cultivated rice. Nature 557 (7703), 43–49. doi:10.1038/s41586-018-0063-9
Wang, W. (2004). The Dusscussion on the Causes of Large-Scale Cultural Changes in China Around 2000 BC. Archaeology (1), 67–77.
Woodbridge, J., Fyfe, R. M., Roberts, N., Downey, S., Edinborough, K., and Shennan, S. (2014). The Impact of the Neolithic Agricultural Transition in Britain: a Comparison of Pollen-Based Land-Cover and Archaeological 14C Date-Inferred Population Change. J. Archaeological Sci. 51, 216–224. doi:10.1016/j.jas.2012.10.025
Yan, X., Wang, S., Jiang, M., and He, K. (2016). Preliminary Study of Charcoal Remains Unearthed from Baodun Site in 2013-2014. South. Ethno. Archaeo. 2, 311–328.
Yuan, L., Qingwen, M., Liyun, Y., Fanxu, M., and Wanquan, Y. (2020). Historical Changes of Plant Community Structure and Biodiversity of Linpan: A Case Study in Pi County of Chengdu. Chin. Agr. Sci. Bul. 36 (7), 44–49. 10.11924/j.issn.1000-6850.casb18110111 .
Zhang, J., Lu, H., Wu, N., Qin, X., and Wang, L. (2013). Palaeoenvironment and Agriculture of Ancient Loulan and Milan on the Silk Road. The Holocene 23 (2), 208–217. doi:10.1177/0959683612455541
Zhao, Z., and Piperno, D. R. (2000). Late Pleistocene/Holocene Environments in the Middle Yangtze River Valley, China and rice (Oryza Sativa L.) Domestication: The Phytolith Evidence. Geoarchaeology 15 (2), 203–222. doi:10.1002/(sici)1520-6548(200002)15:2<203:aid-gea5>3.0.co;2-j
Zheng, Z., Ma, T., Roberts, P., Li, Z., Yue, Y., Peng, H., et al. (2021). Anthropogenic Impacts on Late Holocene Land-Cover Change and Floristic Biodiversity Loss in Tropical southeastern Asia. Proc. Natl. Acad. Sci. USA 118 (40), e2022210118. doi:10.1073/pnas.2022210118
Keywords: phytolith, agriculture, Oryza sativa, domestication, bambusoideae, vegetation, luminescence dating, chengdu plain
Citation: Zhang J, Lv Y, Yu L, Tang M, Huang M, Shao K, Huan X, Wen C, Dong Y, Jiang M, He K, Yan X, Ye M, Wu N and Lu H (2021) Neolithic Rice Cultivation and Consequent Landscape Changes at the Baodun Site, Southwestern China. Front. Earth Sci. 9:807626. doi: 10.3389/feart.2021.807626
Received: 02 November 2021; Accepted: 16 November 2021;
Published: 30 November 2021.
Edited by:
Ying Guan, Institute of Vertebrate Paleontology and Paleoanthropology (CAS), ChinaReviewed by:
Wuhong Luo, University of Science and Technology of China, ChinaXinxin Zuo, Fujian Normal University, China
Copyright © 2021 Zhang, Lv, Yu, Tang, Huang, Shao, Huan, Wen, Dong, Jiang, He, Yan, Ye, Wu and Lu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jianping Zhang, anB6aGFuZ0BtYWlsLmlnZ2Nhcy5hYy5jbg==; Lupeng Yu, eXVsdXBlbmczMTlAMTI2LmNvbQ==