 Bruce S. Rubidge
Bruce S. Rubidge Michael O. Day
Michael O. Day Julien Benoit
Julien Benoit.svg){kind=link}

95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci. , 01 June 2021
Sec. Paleontology
Volume 9 - 2021 | https://doi.org/10.3389/feart.2021.668143
This article is part of the Research Topic Origin and Early Evolution of Amniotes View all 16 articles
Lanthanostegus is an unusual dicynodont known from only two partial skulls from a single locality near Jansenville in the Eastern Cape Province of South Africa. Although these specimens can be constrained to near the base of the late middle Permian (Guadalupian) Abrahamskraal Formation, their precise age is uncertain as a result of diachroneity of the base of the Formation and the absence, in the Jansenville area, of index taxa to correlate this horizon with the biostratigraphy established in the Western Cape Province. Here, we describe a third skull that we identify as Lanthanostegus, which we recently discovered from a locality north of Laingsburg, on the western side of the main Karoo Basin. This skull reveals morphological details of the palate, occiput, and lower jaw that are not preserved in the described specimens of Lanthanostegus mohoii and will advance understanding of this poorly known dicynodont. This discovery provides the first direct correlation between the lower Abrahamskraal Formation at Jansenville on the eastern side of the basin and the southwestern part of the basin, and suggests that Lanthanostegus occurs in the lowest Tapinocephalus Assemblage Zone (AZ), or possibly to a new assemblage transitional between the Eodicynodon and Tapinocephalus AZs. This supports earlier work proposing that the Eodicynodon AZ is present only on the western side of the Karoo Basin and that the transition from a marine to continental depositional environment occurred later toward the East.
Systematic fossil collecting from the rocks of the middle Permian Abrahamskraal Formation of the lower Beaufort Group of South Africa over four decades has revealed more than ten new basal therapsid genera, with most of them being dicynodonts (Day and Rubidge, 2020; Rubidge and Day, 2020). These fossils, together with other middle Permian taxa, have enabled subdivision of the Abrahamskraal Formation into three dicynodont-based biozones defined by the ranges of (from the base upwards) Eodicynodon, Eosimops, and Diictodon (Day and Rubidge, 2020; Rubidge and Day, 2020).
The Eodicynodon Assemblage Zone (AZ) is present only in the southwestern part of the Karoo Basin in the basal Combrinkskraal and Grootfontein members of the Abrahamskraal Formation (Rubidge and Day, 2020). Because of the paucity of fossils in the lower part of the Abrahamskraal Formation, determining its lateral and vertical boundaries is challenging, especially in the area to the south and east of Aberdeen, where this is compounded by a lack of good rock outcrop. Nevertheless, targeted collecting efforts to the east and the north of the known extent of the Eodicynodon AZ have resulted in the discovery of several new anomodont species that are different from the species described from the Eodicynodon AZ. These include Anomocephalus africanus, Colobodectes cluveri, and Prosictodon dubei. The most bizarre species, which has anteriorly oriented orbits, is Lanthanostegus mohoii, and is known from only two fragmentary specimens from the farm Mandalay, north of Klipplaat in Jansenville district of the Eastern Cape Province (Modesto et al., 2002; Modesto and Rubidge, 2003).
The only two known specimens of Lanthanostegus, which are from the same stratigraphic horizon, were found together 52 m above the base of the Abrahamskraal Formation. When the specimens were described, they were considered to come from the Tapinocephalus AZ (Modesto et al., 2002), but because of the small number of identifiable fossils and the lack of diagnostic taxa to compare with the better-sampled western part of the basin, this has remained tentative.
During fieldwork in 2018 in the Moordenaarskaroo, north of Laingsburg on the western side of the basin, our team discovered several vertebrate fossils from a horizon in the lower Leeuvlei Member of the Abrahamskraal Formation (Figure 1). This horizon lies above the units known to be associated with the Eodicynodon AZ (Combrinksrkaal and Grootfontein members) but below the units most associated with the Tapinocephalus AZ (Koornplaats, Swaerskraal, Moordenaars, and Karelskraal members). One of the specimens discovered by JB displays diagnostic characters, allowing it to be referred to Lanthanostegus mohoii. In this article, we describe this new specimen and discuss its stratigraphic significance.
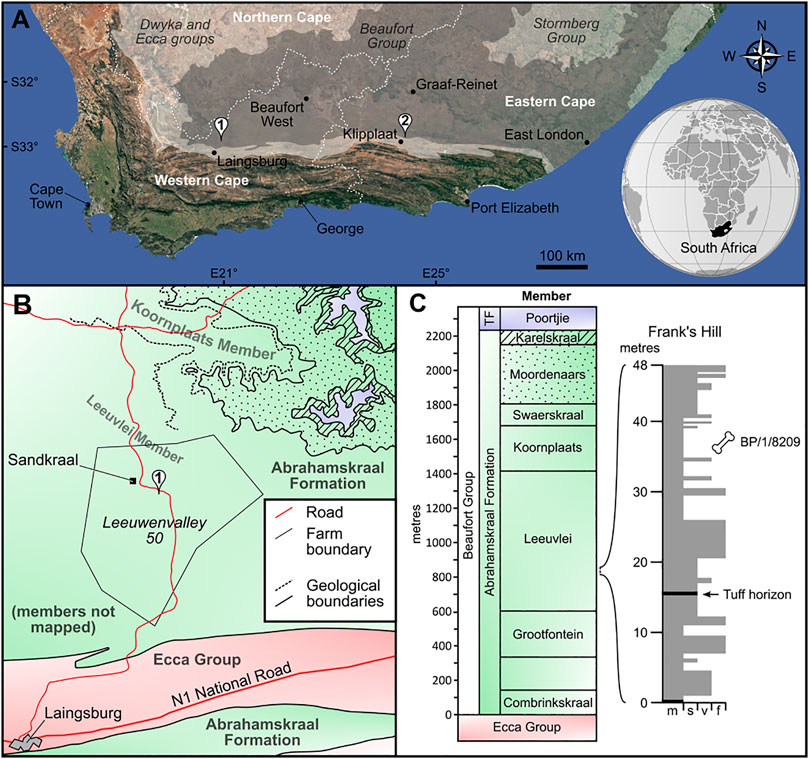
FIGURE 1. Locality and stratigraphic information for Lanthanostegus. (A) Map showing the localities of BP/1/8209 (1) and of the type specimens of Lanthanostegus (2) within the Karoo Basin of South Africa. (B) Geological map of the southern Moordenaarskaroo showing the locality of BP/1/8209 at “Frank’s Hill” on Sandkraal (1). The only members of the Abrahamskraal Formation to have been mapped throughout the area are the Karelskraal and Moordenaars members; lower members of this formation have been mapped in a limited area north from Sandkraal (dashed lines) by Loock et al. (1994). (C) Stratigraphic divisions and thicknesses of the Abrahamskraal Formation in the Moordenaarskaroo (from Day and Rubidge, 2014) and stratigraphic section from “Frank’s Hill” showing the position of BP/1/8209 and a tuffaceous horizon. TF, Teekloof Formation; f, fine sandstone; m, mudrocks; s, siltstone; v, very fine sandstone. Images modified from Google Earth and Wikimedia Commons (https://commons.wikimedia.org/wiki/File:South_Africa_(orthographic_projection).svg).
The specimen (skull BP/1/8209) was mechanically prepared using compressed air-driven engravers fitted with a tungsten carbide tip. As the enclosing matrix is extremely hard, tests were undertaken to determine whether the fossil could be prepared using acetic or formic acid, but they proved to be ineffective. Although differentiation between the bone and matrix is difficult in the specimen, the skull has been fully prepared.
It is extremely weathered in the dorsal view, many of the external features of the bones have been removed through erosion, and it is difficult to determine sutures. Over most of the specimen, sutures between bones are not easily distinguishable; however the midline suture which has been traced over much of the skull roof and part of the palate helps to orientate the various sections of the skull relative to the midline. This is valuable considering that the specimen is distorted with the ventral part being displaced toward the left relative to the skull roof. In the ventral view, the palate is well exposed but is so heavily encrusted with pyrite toward the anterior end that anatomical details are masked. The posterior end of the lower jaw is preserved in articulation, but, like the rest of the skull, the surface of the bone is heavily weathered.
Therapsida Broom, 1905
Dicynodontia Owen, 1859
Lanthanostegus mohoiiModesto et al., 2002
“NMQR 3396, the holotype, is a partial skull comprising the right circumorbital region, the posterior skull table, occiput, and adjacent regions of the braincase. NMQR 3398 is a partial skull comprising the circumorbital regions, anterior skull table, posterior palate with epipterygoid, and partial braincase” (Modesto et al., 2002 p 1756). Both specimens are from Mandalay farm, Jansenville district (close to the village of Klipplaat), Eastern Cape Province, South Africa (32°57′30″S; 24°24′12″E), lowermost Abrahamskraal Formation of the Beaufort Group.
BP/1/8209 is a distorted and crushed skull. The tip of the snout was inadvertently cut off during excavation. From Sandkraal, Leeuwen Valley 50, Laingsburg district, Western Cape Province.
A dicynodont anomodont that is distinguished from other anomodonts by a laterally facing excavation on the dorsal surface of the postfrontal, by extensive, dorsally exposed contacts between the parietals and the supraoccipital, anterior orientation of the orbits, and by extensive ossification of the lateral wall of the braincase that is formed mainly by the prootic. Single postcanine tooth positioned laterally on the maxilla, alongside the longitudinal ridge of the maxilla.
The preserved part of the skull (BP/1/8209) is dorsoventrally compressed, deformed to the right side, and is 20 cm long, which is much as the same size as the two type specimens of Lanthanostegus (Modesto et al., 2002; Modesto and Rubidge, 2003). It is more complete than the previously described specimens and was collected as a single fragment extending from the occiput posteriorly to the snout in front of the canine teeth anteriorly, but the occiput is not preserved. It resembles the previously described Lanthanostegus specimens in the extensive ossification of the lateral wall of the braincase formed mainly by the prootic and anteriorly oriented orbits (Modesto et al., 2002). As the new specimen is the most complete, it provides new anatomical information, particularly of the palate, and its description is important to enhance the understanding of the cranial morphology of this rare and enigmatic dicynodont genus.
Although the extreme anterior end of the skull is not preserved, a part of the maxilla in the anteorbital region is present, and, as in other dicynodonts, it formed the lateral surface of the face (Figures 2–4). The posterior sutural contact of the maxilla with the lacrimal and prefrontal is not evident. The skull roof is very weathered, and the tip of the snout was inadvertently destroyed by an angle grinder in the process of field excavation, but a cross section of a large canine tooth is present in the right maxilla, as well as the worn base of a single postcanine, which is present on the palatal exposure of the maxilla.
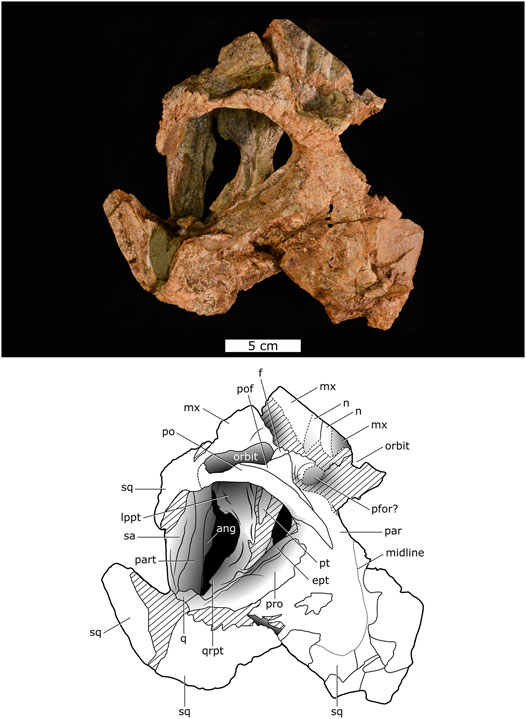
FIGURE 2. Dorsal view of Lanthanostegus specimen BP/1/8209.
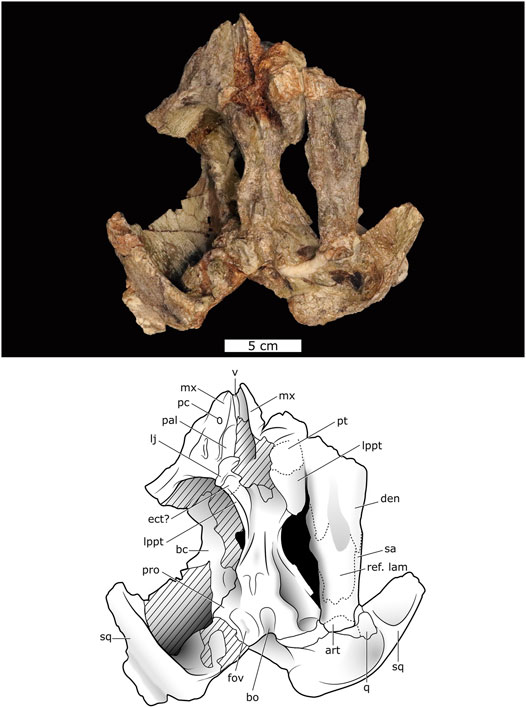
FIGURE 3. Ventral view of Lanthanostegus specimen BP/1/8209.
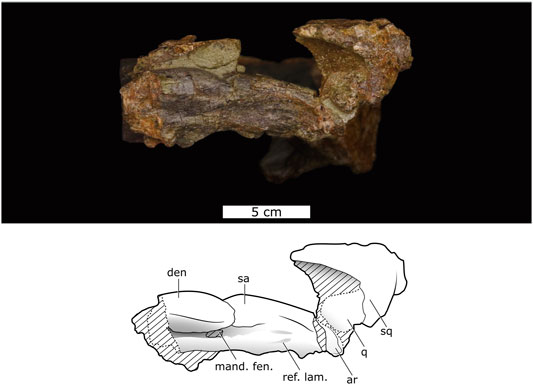
FIGURE 4. Lateral view of Lanthanostegus specimen BP/1/8209 showing the lower jaw.
A feature of the skull is the pronounced and characteristic anterior orientation of the orbits as in Lanthanostegus (Modesto et al., 2002), which is not the result of deformation. The supraorbital area of the skull roof is not well preserved. A prominent depression is present on the weathered skull roof posterodorsal of the orbits (Figure 2), and we suspect that the pineal foramen is present, although not visible, at the posterior end of this depression as in the paratype of Lanthanostegus mohoii (NMQR 3398). We interpret the preparietal as forming the posterior part of the floor of this depression and the paired frontal as forming most of the floor of this depression, and forming the dorsal margin of the orbit anterior to the frontal is the impression of a thin paired elongate bone which we interpret as the nasal, which contacts the maxilla laterally in a longitudinal trough-like depression. The posterior sutural contact of the frontal with the postfrontal is not clear, but the latter triangular bone, which tapers posteriorly between the frontal and postorbital, is positioned at the posterodorsal end of the orbit.
The postorbital is heavily weathered but is intact and forms most of the postorbital bar. As in the paratype of Lanthanostegus mohoii (NMQR 3398), it tapers posterodorsally between the parietal and the temporal opening. It appears that the postorbital overlies the parietal, and the dorsal contact between the two bones is visible as a posterodorsally oriented straight suture, but this contact is not well defined in BP/1/8209.
The dorsal surface of the parietal is extremely weathered and has been squashed toward the right. Judging by the midline suture, it has wide exposure on the skull roof, but no surface details are preserved. The squamosal, as in all dicynodonts, is a large bone that forms the posterior and ventral margins of the temporal fenestra, and it has a broad occipital exposure. It is poorly preserved on the dorsal surface, but the posterior left side is intact, displaying the typical dicynodont squamosal morphology. A prominent external auditory meatus is evident on the occipital surface and has dorsoventrally oriented grooves on its posterior surface, but the rest of the occiput is poorly preserved. The double articulatory condyles of the quadrate are visible in the posterior view below the auditory meatus, but the dorsal process of the quadrate is not evident.
Most of the constituent elements of the braincase are either indistinguishably fused or their contacts are obscured by damage. As in the two type specimens, the prootic forms a thick posteromedial wall that medially closes the posterior half of the temporal fenestra. The anterior border of the prootic is subvertical and notched at mid-height as in the types. No pila antotica is preserved.
The palate (Figure 3) is relatively complete, and its anterior end manifests morphological detail, which is not evident in the holo- and paratypes. The high and vaulted nature of the palate is reminiscent of the situation in Endothiodon (Cox and Angielczyk, 2015). In the ventral view, the right maxilla (the only side which is well preserved) forms the lateral border of the choana, has a relatively smooth surface texture, and has a longitudinal trough in which a single postcanine tooth is present halfway down the lateral side of the trough. Lateral to this tooth, a sharp longitudinal ridge separates the palatal exposure of the maxilla from the facial portion, with the latter having a rugose surface texture. The anterior contact with the premaxilla is not visible, but more posteriorly, a line extending obliquely, lateromedially from the internal naris, is interpreted as the suture between the maxilla and palatine, with the latter bone having a ventrally rounded and more rugose surface.
The palatine forms the posteromedial border of the internal naris and, as is evident from the left side of the skull where the anterior end of the palatine is not preserved, overlies the maxilla ventrally. The posteromedial end of the palatine is encrusted with goethite pseudomorphs of pyrite, which obscures morphological details.
The vomer, which is broad anteriorly and thin posteriorly, is similar to that described for some Eodicynodon oosthuizeni specimens (Rubidge, 1990a). In the ventral view, the two sides of the vomer slope dorsomedially with a straight sutural contact along the midline as is the situation described for the posterior end of the vomer in the paratype of Lanthanostegus (Modesto et al., 2002). The morphology of the posterior end of the vomer cannot be determined as it is heavily coated with goethite encrustation. As the interpterygoid vacuity cannot be determined with certainty it is not possible to ascertain whether the vomers form its anterior margin.
A crescentic line posteroventral to the rugose facial surface of the maxilla is suggestive of the suture between the maxilla and the ectopterygoid, which borders medially with the palatine and anteriorly with the maxilla. Posteriorly, an unclear vertical lineation on the side of the lateral process is considered the suture between the ectopterygoid and pterygoid. The anterior sutural contact between the pterygoid and palatine in the palatal view is not visible and is obscured by a bone fragment that may have been part of the right dentary, and more medially is obscured by goethite.
In this specimen, an elongated cylindrical cast of matrix extending down the midline is possibly a mold of the interpterygoid vacuity, as it is positioned in the appropriate position between the lateral processes but is much smaller than would be expected. As this area is covered by goethite encrustation, it is not possible to determine finer details. The lateral process of the pterygoid is a very prominent feature as is the case in Eodicynodon and the known specimens of Lanthanostegus (Modesto et al., 2002). The left pterygoid is particularly wide in the ventral view, but this may be distortion resulting from the lower jaw being appressed against this part of the skull. A longitudinal ridge down the posterior edge of the lateral process extends posteriorly on the ventrolateral side of the pterygoid and down the quadrate ramus, as in the paratype of Lanthanostegus mohoii (Modesto et al., 2002). Another longitudinal ridge extends posteriorly down the midline of the pterygoid as far as two small elongate posterior pterygoid ridges on either side of the anterior tip of the parabasisphenoid as in the paratype of Lanthanostegus (Modesto et al., 2002). The parabasisphenoid, although present, is not well preserved, and its sutural contacts are not evident. As is typical in dicynodonts, an elongated U-shaped trough extends down the midportion of the parabasisphenoid onto the basioccipital. The parabasisphenoid and basioccipital are flanked on either side by the fenestra ovalis.
The pterygoid girder is a robust bone that is well exposed in the dorsal view. Anteriorly, it is broad and fan-shaped to form a prominent lateral process as in the described specimens of Lanthanostegus, which is particularly well exposed on the left side in the palatal view. In the dorsal view, a longitudinal midline trough extends between the left and right pterygoids. Posteriorly, the ptergygoid narrows, and then expands laterally as the quadrate ramus. A tall but thin epipterygoid column extends anterodorsally from the baseplate and swells dorsally, where it meets the prootic. This structure is much thinner than that of the paratype, where it is uncharacteristically robust for a dicynodont. In the holotype, only the dorsal portion of the epipterygoid column is preserved and is of the same thickness as that of BP/1/8209.
The large base plate of the epipterygoid, which tapers anteriorly and posteriorly, has extensive sutural contact with the dorsal side of the pterygoid girder. Contra to what is figured for the paratype (NMQR 3398) by Modesto et al. (2002), where the anterior extension of the baseplate is short, that of BP/1/8209 is long. Our observations of NMQR 3398 indicate that the anterior process is broken, and there is a triangular impression on the dorsolateral side of the pterygoid girder to indicate the original position of the anterior process in the specimen, showing that the anterior process in this specimen was also long.
As in the Lanthanostegus specimen NMQR 3398, the only other specimen to have a quadrate preserved, the quadrate is poorly preserved in BP/1/8209, and only the condyles of the left quadrate are visible in the ventral view in articulation with the posterior end of the lower jaw. The dorsal part of the quadrate is poorly preserved and extends dorsally from the condyles.
Only the posterior portion of the left lower jaw is preserved and is in articulation with the quadrate, but this is the only lower jaw known of Lanthanostegus. The bone surface of the jaw is not well preserved, but it is possible to determine the configuration of most of the various elements. In the lateral view, the dentary forms the largest component of the mandible, but the anterior end of the dentary is not preserved, and because the lower jaw is occluded to the skull, it is not possible to see details of the transversely expanded dorsal surface of the dentary. It is also not possible to determine whether teeth are present on the dorsal surface of the dentary. Posteriorly, the dentary has a low coronoid process (Figure 4).
The dentary, which is an extremely robust and broad bone in cross section, forms most of the dorsal and anterior margins of the large mandibular fenestra, but appears to be excluded from the ventral margin by the angular (Figure 4). Posterodorsally, the dentary meets the surangular, but the sutural contact is not clear. The pre-articular extends over the medial surface of the dentary. Because only the posterior end of the lower jaw is preserved, the splenial is not present.
The surangular forms the dorsal part of the postdentary region of the mandible. Its anterior contact with the dentary is not clear, but in the lateral view there is a break in the topography of the dorsal side of the lower jaw, which appears to coincide with the contact. The surangular forms the posterodorsal and posterior margin of the mandibular fenestra on both the lateral and medial sides of the jaw. Posteriorly, the surangular forms the broadly rounded dorsal edge of the postdentary region and much of its medial surface. The lateral surface of the surangular appears to be mostly overlapped and hidden by the angular, but this is unsure as the suture between the angular and surangular is not evident. The surangular is fused to the articular, and a distinct suture between the elements is not visible. Medially, the broad pre-articular overlaps the surangular and angular on the medial surface of the mandible.
The angular forms most of the ventral portion of the postdentary region. Anteriorly, the angular is overlapped laterally by the dentary and medially by the pre-articular and splenial. Laterally parts of the element are visible along the ventral and posterior margins of the mandibular fenestra, and on the medial surface below the pre-articular. Posteriorly, the angular overlaps the surangular and forms most of the lateral surface of the mandible between the mandibular fenestra and the articular. As a result of postdepositional lateral compression, the reflected lamina has become appressed to the lateral side of the angular and also protrudes below the ventral margin of the angular.
As is typical in dicynodonts, the pre-articular is a strap-like bone that extends along most of the medial surface of the mandible. Anteriorly, it contacts the dentary and presumably the splenial as in other dicynodonts, but this contact is not preserved, whereas posteriorly, it overlaps parts of the surangular and angular, and is indistinguishably fused to the articular.
The articular is positioned at the posterior end of the lower jaw, but most of it is not visible because the jaw is preserved in articulation with the quadrate. An articular recess appears to be typically present in front of the main articular surfaces, near the junction of the surangular and the articular, and a robust retroarticular process is present on the posteroventral end of the articular.
Lanthanostegus mohoii was previously known from only two fragmentary specimens from the farm Mandalay north of Klipplaat in Jansenville district in the Eastern Cape Province (Modesto et al., 2002; Modesto and Rubidge, 2003). The genus is distinguished from other dicynodonts by the presence of three autapomorphies; a conspicuous laterally facing excavation on the dorsal surface of the postfrontal, dorsal expansions of the supraoccipital that contact the parietals, and extensive ossification of the lateral wall of the braincase (Modesto et al., 2002). Of these autapomorphies, only the extensive ossification of the lateral wall of the braincase is present in BP/1/8209, and the other two are not preserved.
A conspicuous feature of Lanthanostegus is the anterior orientation of the orbits (Modesto et al., 2002), a character present in all three specimens. The most obvious feature of the parietal of the holotype of Lanthanostegus is the thickened rim of the pineal foramen, forming a boss that projects above the dorsal surface of the parietal (Modesto et al., 2002). The pineal foramen cannot be determined in either the paratype and BP/1/8209 as this area of the skull is damaged, and as the foramen cannot be determined, the pineal boss is also not evident. However, it is possible that the pineal foramen could be present in the indentation on the skull roof above the orbits.
The palate is not preserved in the Lanthanostegus mohoii holotype (NMQR 3396) and only partially preserved in the paratype NMQR 3398. The palate of the new specimen (BP/1/8209) is more complete than the other specimens and preserves more of the anterior part than the paratype. The anterior portion of the vomers is preserved and is sutured down the midline as was suggested by Modesto et al. (2002) for the posterior end of the vomers, which is preserved on the Lanthanostegus paratype (NMQR 3398) and as in Eodicynodon oosthuizeni (Barry, 1974; Barry, 1975; Cluver and King, 1983, Rubidge 1990a, Rubidge 1990b). Because of goethite encrustation in the new specimen, it is not possible to determine whether a lateral palatal foramen is present between the palatine and ectopterygoid as in the paratype of Lanthanostegus mohoii (Modesto et al., 2002 p1759). Also, the pterygoid is a conspicuous bone of the palate, and the most obvious part of this bone is the large lateral process (Modesto et al., 2002), which is slightly less prominent than that of Eodicynodon (Rubidge, 1990a). In the new specimen, the anterior contact of the pterygoid with the vomer is obscured by goethite pseudomorph encrustation, and, as in the paratype, the pterygoids meet posteroventrally to form a low posteromedial sagittal ridge (Modesto et al., 2002).
One of the three autapomorphies recognized for Lanthanostegus, namely, the extensive ossified braincase formed mainly by the prootic, which has not yet been recognized in any other dicynodont, is present in BP/1/8209. Furthermore, all dicynodont genera from the Abrahamskraal Formation (Eodicynodon and Tapinocephalus AZ) are small, with the exception of Lanthanostegus. In determining the taxonomic affinity of BP/1/8209, these facts, in addition to the forward-pointing orbits, depression on the frontal above the orbits, lateral position of the single postcanine, robust and large lateral process of pterygoid, the dorsoventrally tall quadrate ramus of the pterygoid, sutured rather than fused vomers, and the ventral meeting of the pterygoids to form a low sagittal ridge posteromedially, suggest that BP/1/8209 belongs to Lanthanostegus. The only difference (from the preserved portions of the skulls) between the newly discovered specimen and paratype of Lanthanostegus mohoii is that the epipterygoid of the paratype is broad, whereas that of the new specimen is narrow. We do not consider this single character difference significant enough to erect a new species of Lanthanostegus and think it is safer to assign BP/I/8209 to the species L. mohoii.
Over the past decade, understanding of the taxonomy, diversity, phylogeny, and stratigraphic ranges of dicynodonts from the Middle Permian Eodicynodon and Tapinocephalus AZ has been greatly clarified (Modesto et al., 2002; Modesto et al., 2003; Angielczyk et al., 2005; Angielczyk and Rubidge, 2009; Angielczyk and Rubidge, 2010; Angielczyk and Rubidge, 2013; Rubidge and Angielczyk, 2009; Smith et al., 2012; Day, 2013). Eodicynodon, the only dicynodont genus from the Eodicynodon AZ (Rubidge and Day, 2020), includes the species E. oosthuizeni (e.g., Barry, 1974; Barry, 1975; Rubidge, 1984, Rubidge, 1990a; Rubidge et al., 1994), and the more enigmatic E. oelofseni (Rubidge, 1990b), which may represent a distinct genus (e.g., Modesto et al., 2003; Kammerer et al., 2011; Castanhinha et al., 2013). Day and Rubidge (2020), provided an up-to-date compilation of dicynodonts from the Tapinocephalus AZ and recognized nine genera: Brachyprosopus, Colobodectes, Diictodon, Emydops, Eosimops, Lanthanostegus, Pristerodon, Prosictodon, and Robertia. Lanthanostegus, which is much larger than any of the other dicynodonts (medium sized as opposed to small), has mostly been considered to be part of the Tapinocephalus zone fauna (Modesto et al., 2002; Modesto and Rubidge, 2003), although Day (2013) suggested that the strata in which it originated could correlate with the Eodicynodon Assemblage Zone.
Eodicynodon (Rubidge, 1990a; Rubidge, 1990b), Brachyprosopus (Angielczyk et al., 2016), Emydops (Cluver and King, 1983; Keyser, 1993; King and Rubidge, 1993; Fröbisch and Reisz, 2008), Eosimops (Angielczyk and Rubidge, 2013), Pristerodon (Fourie, 1993; Keyser, 1993; King and Rubidge, 1993), and Lanthanostegus (Modesto et al., 2002; Modesto and Rubidge, 2003) are all distinctive middle Permian basal dicynodont genera having a broad intertemporal region and “postcanine” teeth. Lanthanostegus can be differentiated from Eodicynodon by the presence of a shelf lateral to the maxillary “postcanine” teeth, smaller ventral flanges on the anterior pterygoid rami, raised edges surrounding the interpterygoid vacuity, larger ossified braincase, anteriorly oriented orbits, and its larger size. Lanthanostegus retains a midline vomerine suture, which is an ancestral character state otherwise known only amongst the dicynodonts Eodicynodon, Colobodectes, and Brachyprosopus (Barry, 1974; Modesto et al., 2002; Modesto et al., 2003; Angielczyk and Rubidge, 2016). In addition, Lanthanostegus possesses a strong pineal boss and lacks a contact between the squamosal and the maxilla (Modesto et al., 2002), and the parts of the skull where these latter characters should be present are very poorly preserved in the newly discovered specimen.
Pristerodon, Emydops, Eosimops, Robertia, and Brachyprosopus can be differentiated from Lanthanostegus by having more medial placement of the maxillary “postcanine” teeth, less prominent ventral flange on the anterior pterygoid ramus, more laterally directed orbits, less plate-like postorbital bar, wider exposure of the parietals on the skull roof (Angielczyk et al., 2016), and a less anteroposteriorly extensive braincase lateral walls (Modesto et al., 2002).
The lower Beaufort Group in the southern part of the Moordenaarskaroo has been affected by folding resulting from the Cape orogenesis, and an anticline structure runs approximately east–west through Sandkraal, close to the homestead, which in places is associated with localized faulting (Loock et al., 1994). The anticline brings to the surface strata of the Leeuvlei Member (Loock et al., 1994), which represents the mid-to-lower part of the Abrahamskraal Formation. This locality is one of the few areas where rocks this low in the Abrahamskraal Formation are exposed at a relatively low angle of dip. The horizons from which BP/1/8209 and the other fossils collected from this locality were found are approximately 220–240 m above the base of the Leeuvlei Member (Figure 1C), and around 800 m above the base of the Abrahamskraal Formation (Loock et al., 1994).
The fossil locality itself is situated on the southern limb of the anticline in the lower-mid Leeuvlei Member. Because so few fossils have been found from this member, or from strata of the middle part of the Abrahamskraal Formation generally, the biostratigraphic provenance of BP/1/8209 is not well constrained; however, it occurs in the stratigraphic interval in which the boundary between the Eodicynodon and Tapinocephalus AZs is expected to be. Other fossils discovered by us from this locality include the weathered partial skull of an undescribed basal anomodont, several dinocephalians, including the skull of a small tapinocephalid showing similarities to some derived tapinocephalids, and some indeterminate temnospondyl bones. We found no fossils of Eodicynodon, which would be expected if the horizon was within the Eodicynodon AZ given the preponderance of this taxon there. Although the assemblage is unusual in lacking any pylaecephalid dicynodonts or Pristerodon, and further collecting is required to clearly demonstrate its correlation, we tentatively assign this horizon to the lower Tapinocephalus AZ on the basis of the tapinocephalid (BP/1/8125).
The type specimens of Lanthanostegus were recovered from a horizon only 52 m above the base of Abrahamskraal Formation (previously the Koonap Formation), at Mandalay near Klipplaat in the Eastern Cape Province (Modesto et al., 2002). These authors attributed them to the Tapinocephalus AZ on the basis of the absence of Eodicynodon and on tapinocephalid dinocephalian fossils from the lower Beaufort on several other farms in the Jansenville area, which were determined to be more advanced than Tapinocaninus (Modesto et al., 2001). A younger age for the base of the Abrahamskraal Formation in the Eastern Cape also came from the area north of Grahamstown, along the Great Fish River, where the dinocephalian Moschognathus whaitsii (Neumann, 2020; Benoit et al., 2021) and the reptile Eunotosaurus africanus had been found in the lower Abrahamskraal Formation (Gow and de Klerk, 1997; Modesto et al., 2001). However, the Abrahamskraal Formation by the Great Fish River is thinner than that in the vicinity of Jansenville, where it is more comparable in thickness to the southwest of the basin (Day and Rubidge, 2014). The other fossils recovered from the lowest Abrahamskraal Formation around Jansenville are either unique occurrences (Lanthanostegus mohoii and Pachydectes elsi; Modesto et al., 2002; Rubidge et al., 2006), or known from both the Eodicynodon and Tapinocephalus AZs (a scylacosaurid therocephalian; Modesto et al., 2001), but Day and Rubidge (2020) accepted a tentative assignment to the lowest Tapinocephalus AZ.
The discovery of Lanthanostegus in the Moordenaarskaroo provides a direct link between the lower Abrahamskraal Formation in the east and west of the Karoo Basin. To our knowledge, the Sandkraal locality is the stratigraphically lowest collection attributed to the Tapinocephalus AZ in the southwest of the basin. Because the Eodicynodon AZ is present at this longitude and is known to be up to 1,100 m thick in the vicinity of Prince Albert Road, 50 km to the east (Jinnah and Rubidge, 2007; Rubidge and Angielczyk, 2009), we infer that it falls within the lowest part of the AZ (see Day and Rubidge, 2020). The presence of Lanthanostegus supports correlation with the base of Abrahamskraal Formation at Jansenville, and thus that the Eodicynodon AZ is not present so far east in the basin as was previously suggested by Modesto et al. (2001).
Given that so few fossils have been found in the lowest Tapinocephalus AZ, near to where it is expected to contact the underlying Eodicynodon AZ, the presence of two unique taxa is remarkable. Additional undescribed anomodont material from the Sandkraal locality (BP/1/8131) suggests the presence of a third taxon restricted to this horizon. If this is the case, then these taxa may be indicative of a distinct subassemblage of the Tapinocephalus AZ, or possibly a distinct AZ in its own right.
Of the three described specimens of Lanthanostegus, BP/1/8209 is the most complete and provides new information on the palate, skull roof, and lower jaw. The discovery of Lanthanostegus in the Leeuvlei Member of the Beaufort Group on the western side of the basin supports earlier work proposing that the Eodicynodon AZ is not present to the east of the Willowmore basinal high and that the transition to nonmarine deposition occurred later there. The new specimen provides the first direct correlation between the lower Abrahamskraal Formation at Jansenville and the southwest of the basin, suggesting Lanthanostegus belongs to the lowest Tapinocephalus AZ or to an assemblage transitional between the Eodicynodon and Tapinocephalus AZs.
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
All authors contributed to the manuscript. BR and JB were mainly responsible for the description, and MD for the stratigraphy. JB discovered the specimen.
Palaeontological Scientific Trust (PAST) funded fieldwork and radiometric dating of lower Beaufort rocks for this project. The DSI/NRF Center of Excellence for Palaeosciences assisted to fund fieldwork. African Origins Platform of the NRF funded preparation of fossils.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Victor Rademacher for doing the anatomical illustrations of Lanthanostegus, and acknowledge the financial support of the Palaeontological Scientific Trust (PAST) and its Scatterlings of Africa program; the National Research Foundation (NRF); the DST/NRF Center of Excellence in Palaeosciences (CoE-Pal); and the University of the Witwatersrand.
ang, angular; art, articular; BP, Evolutionary Studies (formerly Bernard Price) Institute, University of the Witwatersrand, Johannesburg; bc, braincase; den, dentary; ept, epipterygoid; f, frontal; fov, fenestra ovalis; j, jugal; lct, ectopterygoid; lj, lower jaw; lppt, lateral process of pterygoid; mand. fen, mandibular fenestra; mx, maxilla; n, nasal; pbs, parabasisphenoid; NMQR, National Museum, Bloemfontein; pal, palatine; par, parietal; part, pre-articular; pfor, pineal foramen; po, postorbital; pof, postfrontal; pro, prootic; pt, pterygoid; q, quadrate; qrpt, quadrate ramus of pterygoid; ref. lam, reflected lamina of angular; sa, surangular; sq, squamosal; v, vomer.
Angielczyk, K. D., Fröbisch, J., and Smith, R. M. H. (2005). On the Stratigraphic Range of the Dicynodont Taxon Emydops (Therapsida, Anomodontia) in the Karoo Basin, South Africa. Palaeontol. africana 41, 23–33.
Angielczyk, K. D., and Rubidge, B. S. (2010). A New Pylaecephalid Dicynodont (Therapsida, Anomodontia) from theTapinocephalusAssemblage Zone, Karoo Basin, Middle Permian of South Africa. J. Vertebr. Paleontol. 30, 1396–1409. doi:10.1080/02724634.2010.501447
Angielczyk, K. D., Rubidge, B. S., Day, M. O., and Lin, F. (2016). A Reevaluation ofBrachyprosopus broomiandChelydontops Altidentalis, Dicynodonts (Therapsida, Anomodontia) from the Middle PermianTapinocephalusAssemblage Zone of the Karoo Basin, South Africa. J. Vertebr. Paleontol. 36, e1078342. doi:10.1080/02724634.2016.1078342
Angielczyk, K. D., and Rubidge, B. S. (2013). Skeletal Morphology, Phylogenetic Relationships and Stratigraphic Range of Eosimops Newtoni Broom, 1921, a Pylaecephalid Dicynodont (Therapsida, Anomodontia) from the Middle Permian of South Africa. J. Syst. Palaeontology 11, 191–231. doi:10.1080/14772019.2011.623723
Angielczyk, K. D., and Rubidge, B. S. (2009). The Permian Dicynodont Colobodectes Cluveri (Therapsida, Anomodontia), with Notes on its Ontogeny and Stratigraphic Range in the Karoo Basin, South Africa. J. Vertebr. Paleontol. 29, 1162–1173. doi:10.1671/039.029.0431
Barry, T. H. (1974). A New Dicynodont Ancestor from the Upper Ecca (Lower Middle Permian) of South Africa. Ann. South Afr. Mus. 64, 117–136.
Barry, T. H. (1975). “Affinities and Systematic Position of the South African Lower Middle Permian Dicynodonts (Therapsida: Dicynodontidae),” in Gondwana Geology: Papers Presented at the Third Gondwana Symposium. Editor K. S. W Campbell (Canberra: Australian National University Press), 475–479.
Benoit, J., Kruger, A., Jirah, S., Fernandez, V., and Rubidge, B. (2021). Palaeoneurology and Palaeobiology of the Dinocephalian Anteosaurus Magnificus. App 66 (1), 2021: 29-39. doi:10.4202/app.00800.2020
Castanhinha, R., Araújo, R., Júnior, L. C., Angielczyk, K. D., Martins, G. G., Martins, R. M. S., et al. (2013). Bringing Dicynodonts Back to Life: Paleobiology and Anatomy of a New Emydopoid Genus from the Upper Permian of Mozambique. PLoS One 8 (12), e80974. doi:10.1371/journal.pone.0080974
Cluver, M. A., and King, G. M. (1983). A Reassessment of the Relationships of Permian Dicynodontia (Reptilia, Therapsida) and a New Classification of Dicynodonts. Ann. South Afr. Mus. 91, 191–273.
Cox, C. B., and Angielczyk, K. D. (2015). A New Endothiodont Dicynodont (Therapsida, Anomodontia) from the Permian Ruhuhu Formation (Songea Group) of Tanzania and its Feeding System. J. Vertebr. Paleontol. 35, e935388. doi:10.1080/02724634.2014.935388
Day, M. O. (2013). “Middle Permian continental Biodiveristy Changes as Reflected in the Beaufort Group of South Africa: A Bio- and Lithostratigraphic Review of the Eodicynodon, Tapinocephalus and Pristerognathus Assemblage Zones,”. Unpublished Ph.D. Thesis (Johannesburg: University of the Witwatersrand), 387.
Day, M. O., and Rubidge, B. S. (2014). A Brief Lithostratigraphic Review of the Abrahamskraal and Koonap Formations of the Beaufort Group, South Africa: towards a basin-wide Stratigraphic Scheme for the Middle Permian Karoo. J. Afr. Earth Sci. 100, 227–242. doi:10.1016/j.jafrearsci.2014.07.001
Day, M. O., and Rubidge, B. S. (2020). Biostratigraphy of the Tapinocephalus Assemblage Zone (Beaufort Group, Karoo Supergroup), South Africa. South. Afr. J. Geology 123, 149–164. doi:10.25131/sajg.123.0012
Fourie, H. (1993). A Detailed Description of the Internal Structure of the Skull of Emydops (Therapsida: Dicynodontia). Palaeontol. africana 30, 103–111.
Fröbisch, J., and Reisz, R. R. (2008). A New Species of Emydops (Synapsida, Anomodontia) and a Discussion of Dental Variability and Pathology in Dicynodonts. J. Vertebr. Paleontol. 28, 770–787. doi:10.1671/0272-4634(2008)28[770:ansoes]2.0.co;2
Gow, C. E., and de Klerk, B. (1997). First Record of Eunotosaurus (Amniota: Parareptilia) from the Eastern Cape. Palaeontol. africana 34, 27–31.
Jinnah, Z. A., and Rubidge, B. S. (2007). A Double-Tusked Dicynodont and its Biostratigraphic Significance. South Afr. J. Sci. 103, 51–53.
Kammerer, C. F., Angielczyk, K. D., and Fröbisch, J. (2011). A Comprehensive Taxonomic Revision of Dicynodon (Therapsida, Anomodontia) and its Implications for Dicynodont Phylogeny, Biogeography, and Biostratigraphy. J. Vertebr. Paleontol. 31, 1–158. doi:10.1080/02724634.2011.627074
Keyser, A. W. (1993). A Re-evaluation of the Smaller Endothiodontidae. Geol. Surv. South Africa Memoir 82, 1–53.
King, G. M., and Rubidge, B. S. (1993). A Taxonomic Revision of Small Dicynodonts with Postcanine Teeth. Zoolog. J. Linn. Soc. 107, 131–154. doi:10.1111/j.1096-3642.1993.tb00218.x
Loock, J. C., Brynard, H. J., Heard, R. G., Kitching, J. W., and Rubidge, B. S. (1994). The Stratigraphy of the Lower Beaufort Group in an Area north of Laingsburg, South Africa. J. Afr. Earth Sci. 18, 185–195. doi:10.1016/0899-5362(94)90003-5
Modesto, S. P., Rubidge, B. S., De Klerk, W. J., and Welman, J. (2001). A Dinocephalian Therapsid Fauna on the Ecca-Beaufort Contact in Eastern Cape Province, South Africa. South Afr. J. Sci. 97, 161–163.
Modesto, S. P., Rubidge, B. S., and Welman, J. (2003). Erratum: A Replacement Name for Lanthanocephalus Modesto, Rubidge, and Welman 2002, Not Lanthanocephalus Williams and Starmer 2000. Can. J. Earth Sci. 40 (3), 467. doi:10.1139/e03-900
Modesto, S. P., Rubidge, B. S., Welman, J., and Welman, J. (2002). A New Dicynodont Therapsid from the Lowermost Beaufort Group, Upper Permian of South Africa. Can. J. Earth Sci. 39, 1755–1765. doi:10.1139/e02-091
Modesto, S., Rubidge, B., Visser, I., and Welman, J. (2003). A New Basal Dicynodont from the Upper Permian of South Africa. Palaeontology 46, 211–223. doi:10.1111/1475-4983.00295
Neumann, S. (2020). “Taxonomic Revision of the Short-Snouted Tapinocephalid Dinocephalia (Amniota – Therapsida)- the Key to Understanding Middle Permian Tetrapod Biodiversity,”. Unpublished PhD Thesis (Johannesburg: University of the Witwatersrand), 410pp.
Owen, R. (1859). On the Orders of Fossil and Recent Reptilia and Their Distribution in Time. Rep. Br. Assoc. Adv. Sci. for 1859, 153–166.
Rubidge, B., and Angielczyk, K. D. (2009). Stratigraphic Ranges of Tapinocephalus Assemblage Zone Dicynodonts: Implications for Middle Permian continental Biostratigraphy. Palaeontol. africana 44, 134–135.
Rubidge, B. S., and Day, M. O. (2020). Biostratigraphy of the Eodicynodon Assemblage Zone (Beaufort Group, Karoo Supergroup), South Africa. South. Afr. J. Geology 123, 141–148. doi:10.25131/sajg.123.0010
Rubidge, B. S., King, G. M., and Hancox, P. J. (1994). The Postcranial Skeleton of the Earliest Dicynodont Synapsid Eodicynodon from the Upper Permian of South Africa. Palaeontology 37, 397–408.
Rubidge, B. S. (1990a). Redescription of the Cranial Morphology of Eodicynodon Oosthuizeni (Therapsida: Dicynodontia). Navorsinge van die Nasionale Mus. Bloemfontein 7, 1–25.
Rubidge, B. S., Sidor, C. A., and Modesto, S. P. (2006). A New Burnetiamorph (Therapsida: Biarmosuchia) from the Middle Permian of South Africa. J. Paleontol. 80 (4), 740–749. doi:10.1666/0022-3360(2006)80[740:anbtbf]2.0.co;2
Rubidge, B. S. (1984). The Cranial Morphology and Palaeoenvironment of Eodicynodon Barry (Therapsida: Dicynodontia). Navorsinge van die Nasionale Mus. Bloemfontein 4 (14), 325–402.
Rubidge, B. S. (1990b). The Cranial Morphology of a New Species of the Genus Eodicynodon (Therapsida, Dicynodontia). Navorsinge van die Nasionale Mus. Bloemfontein 7, 29–41.
Smith, R., Rubidge, B., and van der Walt, M. (2012). “Therapsid Biodiversity Patterns and Paleoenvironments of the Karoo Basin, South Africa,” in Forerunners of Mammals: Radiation, Histology, Biology. Editor A. Chinsamy-Turan (Bloomington, Indiana: Indiana University Press), 30–62. Submitted 8 February, 2021;.
Keywords: Guadalupian, Tapinocephalus, Beaufort Group, Permian, Abrahamskraal Formation
Citation: Rubidge BS, Day MO and Benoit J (2021) New Specimen of the Enigmatic Dicynodont Lanthanostegus mohoii (Therapsida, Anomodontia) from the Southwestern Karoo Basin of South Africa, and its Implications for Middle Permian Biostratigraphy. Front. Earth Sci. 9:668143. doi: 10.3389/feart.2021.668143
Received: 15 February 2021; Accepted: 10 May 2021;
Published: 01 June 2021.
Edited by:
Jörg Fröbisch, Museum of Natural History Berlin (MfN), GermanyReviewed by:
Michael David Gottfried, Michigan State University, United StatesCopyright © 2021 Rubidge, Day and Benoit. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bruce S. Rubidge, YnJ1Y2UucnViaWRnZUB3aXRzLmFjLnph
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.