 Oscar Carranza-Castañeda
Oscar Carranza-Castañeda
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci. , 21 May 2019
Sec. Paleontology
Volume 7 - 2019 | https://doi.org/10.3389/feart.2019.00089
This article is part of the Research Topic Examining Evolutionary Trends in Equus and its Close Relatives from Five Continents View all 14 articles
In the advances of the research of the sedimentary basins from central Mexico, it is significant the discovery of the first records of equids of Clarendonian and early Hemphillian age that are mentioned. The specimens were collected in southwest deposits of the San Miguel de Allende basin, the ages are verified by radiometric dates. The largest collection of Dinohippus mexicanus skulls jaws and isolated teeth recovered, have shown the wide geographical distribution in faunas of central Mexico. The study of these records from localities of Coecillos, Rinconada, Arroyo Emilio, and Rancho el Ocote faunas of the San Miguel Allende, Guanajuato state, Tecolotlán, Jalisco state and Juchipila basin Zacatecas state, have shown the variability and transitional features they have in the structures of the skull and upper and lower molars. The skull from Tecolotlán and Rancho El Ocote, of the late Hemphillian age present a well-defined dorsal fossa, in contrast the oldest skull from early-late Hemphillian of Juchipila, the fossa is just a depression in the Maxillary and nasal bone, however, the skull from Coecillos has structures that differentiate it from others. The most relevant records are the lower and upper molars collected at the top of the biostratigraphic sequence of Rancho El Ocote, in the sites named GTO 2 site D and F. These specimens present features that differentiate them from Dinohippus and suggest a primitive Equus. Upper teeth from the Blancan age of Tecolotlán basin, have a high crown but a pronounced curvature like the Dinohippus mexicanus collected in the late Hemphillian strata of this basin.
The research on paleontology of the sedimentary basins started at the Instituto de Geología has continued for more than 20 years at the Centro de Geociencias, Campus Juriquilla; both institutions belong to the Universidad Nacional Autónoma de México. The main objective of this project is to study the biostratigraphy of the Late Neogene sedimentary basins in the central region of Mexico.
These basins are in the Transmexican Volcanic Belt and north to it between the parallels 19° and 23°. The results obtained have demonstrated the importance of these sedimentary basins to complement the biostratigraphy of Mexico and North America.
The abundance and diversity of mammals that have been collected, with reliable stratigraphic and geographical information, have shown two faunal associations of different ages, Hemphillian and Blancan NALMA (North American Land Mammals Ages). These records allow us to understand important geological and biological events like the dispersal of mammals from the Temperate North America faunas to the central region of Mexico, the dispersal of the antilocaprids Cosoryx and Plioceros described in the Juchipila fauna in the early-late Hemphillian age, and the equids Calippus martini and the antilocaprid Merycodus, described from the la Nopalera locality in ages of 6.77 ± 0.21 My; these discoveries imply a reconsideration of the concepts of extinction, stratigraphic range and geographical distribution (Carranza-Castañeda et al., 2013; Robles-Rivera, 2015).
Another important result of the work in these basins refers to the most important biological event among the Americas, the Great American Biotic Interchange to (GABI). The recent discoveries of Neotropical immigrants in the Tecolotlán basin in deposits of 4.89 My confirm that there was more than one invasion of Neotropical faunas toward North America, prior to closing of the Panamanian land bridge (McDonald and Carranza-Castañeda, 2017). Additional contributions to these results refer to the exchange and displacement of mammals between the faunas of temperate North America toward the faunas of the central Mexico in both directions. Recent descriptions of neotropical mammals suggest that the western part of Mexico was an important migration route (Sussman et al., 2016; McDonald and Carranza-Castañeda, 2017; Carranza-Castañeda, 2018).
The above-mentioned information is based on an extensive collection of fossil mammals that has been integrated with the results of research in the sedimentary basins that have been studied, however, very little has been mentioned of the advances in the knowledge of the equids that have allowed to complement the biostratigraphic information of the strata that contain neotropical immigrants and mammals from the Great Plains of North America.
The objectives of this work are: to mention the first records of equids collected in the Clarendonian and early Hemphillian deposits, located south of the San Miguel de Allende basin, state of Guanajuato; to comment and describe the geographical distribution of Dinohippus mexicanus in the faunas of the central region of Mexico, defining the great variability that occurs in the facial region and the occlusal surface of upper and lower molars; to describe partially the stratigraphy of the most important faunas where diagnostic material of D. mexicanus was collected. In addition, the associated fauna in the stratigraphic levels where they were collected with the criterion that the age is determined by radiometric dates will be mentioned. The age of the records mentioned are determined to early-late Hemphillian, late Hemphillian (Hh3), and latest Hemphillian (Hh4). the analysis on interbedded volcanic ash and the associate fauna have determined the age of the fossils (Carranza-Castañeda et al., 2013; Figure 1).
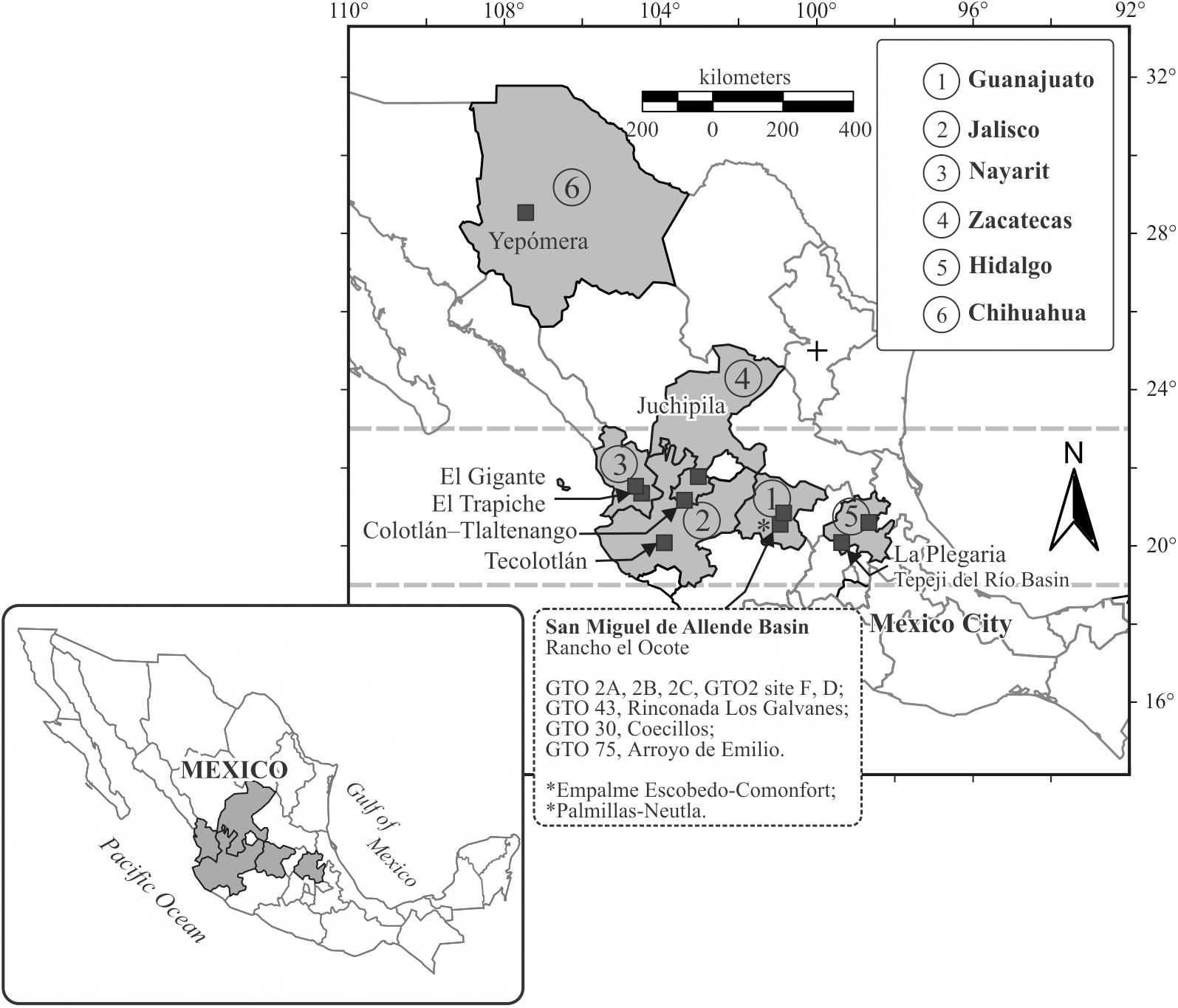
Figure 1. On the map are the locations explained in the paper.
All the fossil materials mentioned in this paper are housed in the fossil collection of the Instituto de Geología, Ciudad Universitaria and the Centro de Geociencias, UNAM in Juriquilla. Querétaro.
To avoid the repetition of names, of the associated fauna with records of D. mexicanus is mentioned in Supplementary Table 1.
The terminology used in the description is from Lance (1950), MacFadden (1984, 2016).
The abbreviations used in the text and tables are: Ap, Anteroposterior, DPOF, Dorsal Preorbital fossa; Colo, Colotlán; HC, High crown; IGM, Instituto de Geología; Ii incisive; Jalp, Jalpan; Juch, Juchipila; JalTeco, JT Jalisco Tecolotlán; LACM, Natural History Los Angeles County Museum; L, Left; MPGJ, Museo de Paleontología Geociencias Juriquilla; Mm, molar; MSTHT, Mesostyle Crown Height; NMMNH, New Mexico Museum Natural History; Pp, Premolar; R, Right; Tab, Tabasco; Tr, transverse; Zac, Zacatecas; UNAM, Universidad Nacional Autónoma de México.
The continuity of the investigations in southern San Miguel de Allende basin, have demonstrated new records of equids that correspond to Clarendonian and early Hemphillian ages, being the first time that in Mexico there are mentioned equids of unequivocal Clarendonian age, whose age is not inferred, but discovered a few meters below the andesites that have been dated.
In the city of Comonfort, 25 km south of San Miguel de Allende, two new localities have been discovered, the locality with the older fauna is Locality GTO 92 Empalme Escobedo, located 5 km south of the town of the same name. The fossils were collected in a quarry of materials for road construction. The thickness of the sedimentary sequence is greater than 50 m and it is composed of different strata of igneous rocks and gravels at the base, covered by lacustrine sediments and layers of clay with sands. The fossils were collected in these layers. This sequence has a thickness of 5 m, is composed of clay and clays with gravel only 6 m above are the strata of andesites that cover the entire sequence.
The age of the sedimentary sequence is determined by the radiometric dates that were made in the andesites located 6 m above, by the method of 40Ar/39Ar, which produced the result of 10.6 ± 0.3 My (Cerca-Martínez et al., 2000). These results confirm the Clarendonian age for this fauna. This is the only one known fauna of this age in central Mexico with well-determined ages.
The fossil material are two teeth, MPGJ 3031 assigned to Protohippus et al. sp., the other, MPGJ 3032, is referred to Hipparion (Robles-Rivera, 2015).
The molar MPGJ 3031, M1, is hypsodont and curved, the protocone is small rounded with no heel, and is joined to the protoselene, by the isthmus that is constricted. The prefossette has multiple plications and a well-developed prefossette loop; the postfossette with the anterior border very simple and large plicaballin, with elongated hypocone.
MPGJ 3032 M1 fragment of upper molar, by the wear corresponds to an old adult, the main character for its assignment is the isolated and rounded protocone and simple fossettes without plications.
The second locality, GTO 91 Palmillas, is near the village of the same name. The area is very small; however, the fossils were collected in the deposits of the La Nopalera Formation (Ledezma, 1960). They consist of layers of clay with different concentrations of sand. These strata are covered by a layer of volcanic ash that was analyzed by the U/Pb method, produced a result of 6.77 ± 0.21 My, which refers them to the early Hemphillian.
The associated fauna includes remains of the antilocaprid Merycodus sp. and an incisor referred to Megatylopus (Robles-Rivera, 2015).
Upper molars: MPGJ 1713 P2, MPGJ 1706 P34, MPGJ 1707 M2, MPGJ 1705 M3, MPGJ 712 M3, MPGJ 1708 M1, MPGJ 1709 M12, MPGJ 1711 M1, and MPGJ 1710 M3. Lower molar MPGJ 1824 m2. The main characters that justify the assignment to C. martini, refer to the protocone, arranged slightly diagonal, joined to the protoselene, and the shape change from oval to elongated; the curvature of the molars is moderate to pronounced, the fossettes with more plications in early stages of wear, hypocone open lobular to lenticular in old individuals.
The comparable C. hondurensis that has been described in faunas of central Mexico, differs considerably in size and the transverse and anteroposterior axes that are smaller compared with C. martini. The age for these deposits dated by U/Pb method is 6.77 ± 0.21 My, early Hemphillian (Robles-Rivera, 2015).
In all the Hemphillian faunas described in the sedimentary basins of central Mexico, there is a large diversity of mammals among which the equids are widely represented. However, C. hondurensis and Astrohippus ansae are present only in the early-late Hemphillian faunas. Neohipparion eurystyle and D. mexicanus are present in all sedimentary sequences, from the early-late and late Hemphillian, although in the deposits of the latest Hemphillian (Blanco Layer of Rancho El Ocote), only D. mexicanus survives until the unconformity of Blancan age (Carranza-Castañeda, 1989, 1992; Carranza-Castañeda et al., 2013).
The lake sediments fill the basin and a few sites have fluvial deposits. The study began north of the city of Colotlán to its limit to the south in the village of Tepechitlán where some metapodial fragments were collected and that is at the end of the basin.
The most important is the locality Zac 9 Sanidad Animal (Carranza-Castañeda, 2006), where isolated teeth of C. hondurensis were recovered, besides D. interpolatus, however, by comparison with new material collected in Juchipila basin, they have been reassigned to D. mexicanus. The age was determined to the early-late Hemphillian by the presence of Neohipparion and Calippus (Carranza-Castañeda, 2006).
It is the most relevant area in the state of Zacatecas by the stratigraphy and the diversity and number of fossils collected with an extent of 165 and 15 km in width in the NNE direction. The first works on geology were made by students from the University of New Orleans, Lahiere (1982) and López (1991), who informally named the fluvial lacustrine deposits as Juchipila Formation. However, in their reports on the geology they only mention a molar of a peccary that they assigned to Platygonus rex. At the beginning of our investigations several important sites were discovered with different kinds of mammal fossils, as well as traces of birds, camelids, proboscideans, and equids in the lacustrine sediments. The relationships of the main localities were established (Aranda-Gómez et al., 2008).
The most important localities are in the central part of the basin, between the Tabasco city in the north and Moyahua in the south. The greatest extent is found in the middle part at the localities: El Mixtón and La Perdida, Cofradía and the Pitahaya area; and the Juchipila area with the localities El Resbalón, La Copa y El Epazote. In the northern part, the Tabasco area, the localities are La Escalera, Patos y Madera and the Wang locality. The associated fauna that has been described in previous works has been referred to the early-late Hemphillian age, was collected and supported by the radiometric date obtained in the analysis of volcanic ashes by the method of U/Pb which gave results of 6.95 ± 0.27 My in the lower and 5.59 ± 011 in the upper ash, ages that bracket the fauna in the early-late Hemphillian age (NALMA) (Aranda-Gómez et al., 2008; Carranza-Castañeda et al., 2013; Carranza-Castañeda, 2018).
The diversity of equids presented in these localities is remarkable. C. hondurensis, maxillaries, jaws and isolated upper and lower teeth, are widely distributed in all the localities of the basin; this is the largest representation of this equid in early-late Hemphillian faunas in the central region of Mexico, and so far, never is associated with Nannippus (Carranza-Castañeda and Espinosa-Arrubarena, 1996; Carranza-Castañeda et al., 2013).
The genus N. eurystyle in the Mixtón locality was collected in the same stratigraphic level; both parts of the maxillary of the same individual (catalog MPGJ 1870) and several isolated teeth have been collected.
The only jaw of A. ansae known in Mexican faunas was collected in the locality Zac Juch 48, La Copa. It consists of a complete left jaw with p2-m2. The main difference with A. stockii is the measurements of the Ap and Tr axis that are bigger and the paralophid is stouter, the protoconid and hypoconid are more rounded. The most important find was collected in the Mixtón sequence. It is a partial skull, MPGJ 1682, with the craniofacial region almost complete in both sides but better preserved in the right side where the dorsal fossa is more evident; the dentition includes part of the P2, and complete P3-M3, molars on the right side. The specimen has been referred to D. mexicanus; in addition, isolated upper molars collected in Zac Juch 47 El Resbalón, have a pronounced curvature, although are very short high crown (Figures 2A,B).

Figure 2. MPGJ 1982 Dinohippus mexicanus, Zac Juch 30 Mixtón. (A) Right side of the skull, the nasal notch is on M2, the maxillary fossa is shallow, it is not pocketed posteriorly, the edges are poorly defined and the infraorbital foramen is included in the anterior part. (B) MPGJ 1683, ventral view of P3-M3, we observed the protocone with small anterior heel and P4-M1 the lingual edge is concave.
At the locality Zac Tab 34, Wang, located 45 km north of the Mixtón, a mandible referred to D. mexicanus was collected in a sequence of clays with different concentrations of fine sand. The jaw includes the complete right side with well-preserved p2-m3 and a whole symphysis including the incisors (Figure 3).

Figure 3. MPGJ 1683 of Zac Tab 34 Wang Location, ventral view of the jaw, symphysis and incisors are complete, the incisors have wide infundibulum, the linguaflexid is wide, the metacónid and metastilid are lobular, the lingual edge of the protoconid and hypoconid is rounded.
Zac Juch 30 Mixtón. MPGJ 1982, partial cranium with right region facial well preserved and P3-M3 and fragment of P2. Zac Juch 47 El Resbalón MPGJ 4067 P4-M1, MPGJ 4095 M1, and MPGJ 4067 P4. Lower jaw, Zac Juch 34 Wang MPGJ 1683 belongs to young individual, with p2-m3 and i1-3, both sides and complete symphysis.
The specimen MPGJ 1682 partial skull collected in the locality Zac Juch 30 El Mixtón.
The jaw MPGJ 1683 from the locality Zac Tab 34 Wang, State of Zacatecas, México; early-late Hemphillian age. Collectors, Xiaoming Wang, Jorge Aranda-Gómez and the author.
The skull (MPGJ 1682) on the left side is slightly squashed, the facial region on the right side is complete without deformation. The occipital region is also incomplete, although some of the structures are observed. The nasal notch is above the posterior part of P2, the row of P3-M3 has a length of 126 mm. The infraorbital foramen is located 68 mm above the alveolus of P4. The malar crest is remarkable and strong, it ends between P4 and the anterior part of M1. The malar fossa is absent. The nasomaxillary fossa (DPOF) is shallow, is 58 mm distant from the orbit, is a slight depression not pocketed posteriorly and the anterior end is above the M2. The fossa has poorly defined and rounded borders, the wider part is 38 mm and the infraorbital foramen is included. The distance from the orbit to the DPOF is 57 mm. The height from the alveolus of M2 to the orbit is 114 mm.
The premolar P2 is preserved. It has a prominent parastyle and mesostyle, the enamel is moderately thick and the fossettes are almost without plication by wear.
On the P3 (Ap 26.90 mm and Tr 27.18 mm), the prefossette is very worn and the postfossette is without plications. The hypocone is broad and the hypoconal groove is of moderate depth. They persist in the whole dentition but only reduce their size. The preprotoconal groove is constricted, the protocone is broad and moderately long, the anterior part forms a rounded heel, and the lingual border is flat and parallel to the lingual part of the alveoli. P4 (Ap 24.45 mm and Tr 25 mm), presents the parastyle and mesostyle that are of almost equal widths and flat. The protocone has a remarkable and blunt heel, the posterior part is long and rounded and the lingual border is concave in a “wooden shoe shape.”
M1 (Ap 22.95 mm and Tr 25.44 mm), The fossettes are very worn, the protocone has the “wooden-shoe shape,” posteriorly it is elongated and wide, the anterior with a notable heel, and the lingual border is concave. M2 (Ap 23.7 and Tr 23.06), it has a trapezoidal shape because the parastyle side is longer. The protocone is broad and elongated, with the posterior part triangular, and the anterior part forms an evident heel of rounded shape. On M3 (Ap 24.94 mm and Tr 20.46 mm) the most notable difference of the molar is reduction of the hypocone and hypoconal groove. The protocone in the anterior part forms a prominent heel, posteriorly is elongated and the lingual border is horizontal (Figures 2A,B).
Lower jaw. The specimen MPGJ 1683 preserves the right side, is broken on the horizontal edge, and only has part of the ascendant border of the coronoid process. The symphysis is complete with R and L i1-3, all have a wide infundibulum filled with cement. The canine is very small, which suggests a female. The pre-canine diastema is 8.13 mm, and the post-canine diastema is 74.45 mm. The maximum width between the L I3 and R 13 is 57.16 mm. The dentition p2-m3 is 147.5 length, the Ap of p3-p4 is 54.7 mm and Tr is 18.8; the Ap m1-m2 46.8 and Tr 16.3. The protoconid and hypoconid are rounded in molars and premolars. The metaconid and metastylid in the premolars are rounded almost with similar size and separated by a wide linguaflexid. In the molars, the metaconid is rounded, but the metastylid is smaller and slightly sharp posteriorly. The ectoflexid in the molars is deeper to almost touch the enamel-edge of the linguaflexid; in premolars, it does not penetrate the isthmus. The assigned age is determined associate fauna and analysis of the upper ash the result 5.9 My placed the jaw to early late Hemphillian.
According to the stratigraphic age the skull of Juchipila, early late Hemphillian, a protocone like D. interpolatus or D. leidyanus could be expected, without heel and inclined toward the lingual margin, however, the shape and disposition of the protocone, is like Dinohippus mexicanus of late Hemphillian (Figure 3).
The first reference to D. mexicanus is associated with the description of the equids from Rancho El Ocote fauna, however, it has had a complicated taxonomic history (Arellano, 1951; Mooser, 1958, 1960, 1965, 1973a,b; Carranza-Castañeda and Ferrusquía-Villafranca, 1978; Dalquest and Mooser, 1980; Carranza-Castañeda, 1992).
In the new investigations in Rancho El Ocote and other Hemphillian localities, the material of A. stockii, N. eurystyle, and D. mexicanus collected in different stratigraphic layers was compared with the Yepomera collection housed in the Museum of Los Angeles (LACM), arriving at the conclusion they are the same species, for which N. floresi and N. arellanoi are considered as junior synonym of N. eurystyle (Carranza-Castañeda and Ferrusquia-Villafranca, 1979; MacFadden, 1984), and D. ocotensis (Dalquest and Mooser, 1980; Dalquest, 1988), is a junior synonym of D. mexicanus from the faunas of central Mexico. All the records known in central Mexico are assigned to the same species as Yepómera (Carranza-Castañeda and Ferrusquia-Villafranca, 1979; MacFadden, 1984; Carranza-Castañeda, 1989, 1992, 2006).
The first mention of horses from Rancho El Ocote was made by Arellano (1951). Later several papers (Mooser, 1958, 1968, 1963) describe different species, summarized in the last of these works like D. ocotensis (Dalquest and Mooser, 1980); they suggest that the locality represents only one age, late Hemphillian, and that was younger than the Yepomera fauna, however, they have never justified this comment. The Rancho el Ocote Local Fauna has been considered as the reference of the late Hemphillian faunas in central Mexico, by the diversity and abundance of the Hemphillian fossil indexes.
The results of new field work in this locality and studies of taphonomy have demonstrated two stratigraphic levels occur, each one with its own lithology and faunal content that corresponds to the late (Hh3) and latest (Hh4) Hemphillian; covered in erosional unconformity by deposits that contain a fauna of early Blancan age (Carranza-Castañeda, 1989, 2006; Carranza-Castañeda et al., 2013).
The lower layer is named the Rhino layer (GTO2B), because its contains: Teleoceras, Machairodus, Borophagus secundus, Agriotherium, Pseudaelurus, Alforjas and the equids N. eurystyle and Nannippus aztecus, which are the last records in all the central México Hemphillian faunas, A. stockii is recorder in the Rhino layer and in contact with the Blanco layer. These are the best representation of late Hemphillian fauna in central Mexico and have been referred to Hh3. D. mexicanus is well represented by skull, jaws, isolated upper, and lower molars, incisors and postcranial elements; the age of the fauna represents the late Hemphillian age. The result of the analysis of the ash by the methods of fission track in zircons gave the result of 4.8 ± 0.2 My that confirms the assigned age (Kowallis et al., 1999; Carranza-Castañeda et al., 2013).
The Rhino level is covered by the Blanco layer (GTO2 A), the lithology is a fine sand deposit and volcanic ash lenses, which contain a special distribution of fauna. The A. stockii material is recorder only at the base represented by a few teeth. D. mexicanus is found throughout the White layer, associated with Stegomastodon, Megatylopus, and Hexobelomeryx and some rodents and the oldest record of the South American immigrant Paramylodon garbani (Montellano-Ballesteros and Carranza-Castañeda, 1986; Carranza-Castañeda et al., 2013).
Above the Blanco layer, there are two-three meters of early Blancan sediments where N. peninsulatus, Equus teeth and a few osteoderms of the South American immigrant Plaina have been recovered. The age was obtained by the analysis of volcanic detritus by 40Ar/39Ar 4.7 ± 0.02 My; this result bracket the Hemphillian-Blancan boundary in Mexico in 4.74 ± 0.14 and 4.89 ± 0.16 (Kowallis et al., 1999; Flynn et al., 2005).
In the upper part of the Blanco level, there are small deposits of clay containing upper and lower molars that have the axes Ap and Tr larger than those collected in the Rhino level. The fossils from these sites have been cataloged as GTO 2 site D and GTO 2 site F (Figure 4).
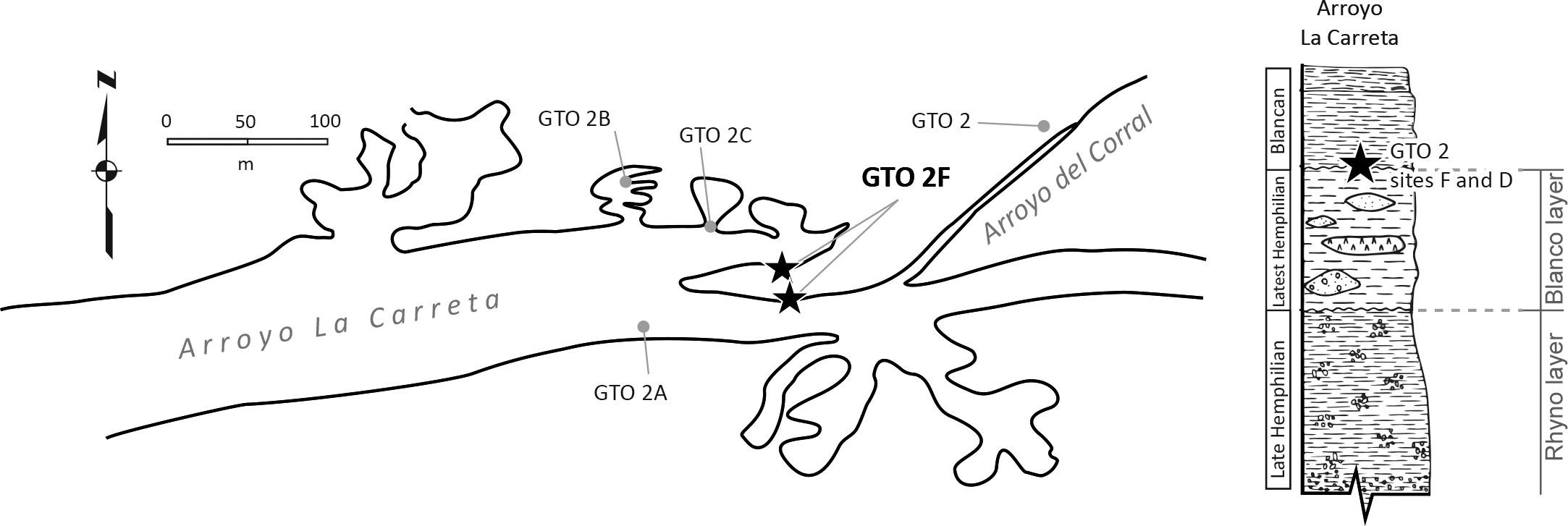
Figure 4. Map of Arroyo La Carreta, where the fossil localities of Rancho El Ocote are located. Note that the GTO 2 site D and F are on top of Blanco layer, the stratigraphic column show the sites in the Blanco layer (Latest Hemphillian).
Skull IGM 7596, skull with R I3, P2-M3, L I2-I3, P2-M3.
Lower Jaw. MPGJ 3739 P2-M3 GTO 2A, MPGJ 3872 L p2-p4 GTO 2b, Rhino layer, MPGJ 3946 R p2-m3, symphysis Ri 1-3 L 2-3, MPGJ 5125 GTO 2 A, Blanco layer, with p2-m1, canine and l 2-3 incisive the R I 2-3 broken, MPGJ 3018 R i1-3 Li 1-3.
Upper Molars, GT0 2A Blanco Layer: MPGJ 5131 M3, MPGJ 5151 M1, MPGJ 5132 P4, MPGJ 5153 P34. GTO 2B Rhino layer; MPGJ 510 M3, MPGJ 650 M1, MPGJ 2056 M1, MPGJ 2075 P4, MPGJ 2915 M1, MPGJ 3793 P4, MPGJ 3822 GTO M2, MPGJ 3841 P4, MPGJ 5128 P4, M3, MPGJ 5131M3, MPGJ 5135 M1, MPGJ 11500.
UPPER MOLARS GTO 2 site D, MPGJ 2825 p4, MPGJ 5111 P34, MPGJ 5113 P3, MPGJ 5126 P4, MPGJ 3947 M1.
UPPER MOLARS GTO 2 site F: MPGJ 4040 M1, MPGJ 5146 P4IGCU 5180 M2, IGCU 11503 P4, IGCU 11504 M1.
LOWER MOLARS GTO 2A; site D: MPGJ 5109 m3; MPGJ 2825 p4.
LOWER MOLARS GTO 2 SITE F, MPGJ 495 p3, MPGJ 2285 p34, MPGJ 2285 p43, MPGJ 3548 p3; IGCU 3858 p3; MPGJ 5116 p4, MPGJ 5117 p34, MPGJ 5118 p4, MPGJ 5119 m2, MPGJ 5120 p34, MPGJ 5129 p4, MPGJ 5130 p2, MPGJ 5134 p4, MPGJ 5144 p3, MPGJ 5145 p4; MPGJ 5147 p4, MPGJ 5149 m2, MPGJ 5150 m2.
The skull has been described and discussed extensively by MacFadden and Carranza-Castañeda (2002). In this part only comparisons will be made with other skulls collected in the faunas mentioned in this paper.
Lower Jaw referred. The jaw MPGJ 3739 from GTO 2 A, collected from the upper part of Rhino layer, conserves the left side, the horizontal border is complete. The maximum depth is at the beginning of the angle of the mandible below the m3 with 114 mm. The Ap length of p2-m3 is 166 mm. To determine the relationship between jaw depth and the crown height at p4 and m2, an X-Ray of the jaw was made. It is remarkable that there is no relationship between jaw depth (m2) and the height of the crown, which justifies the larger size of the molars of the GTO 2 D and F sites.
The jaw MPGJ 3946 collected in GTO 2 B from the Rhino layer, preserves the symphysis, the canine is partially broken, the incisors form an almost curved row, R i1-3, and L i2-3. The i2, is wider. The infundibulum limited by enamel and filled with cement. The post-canine diastema is 89.74 mm, the pre-canine diastema is ∼7 mm. The length R i2- L i2 is 43.70 mm; the length Ap of p2-m3 is 156 mm, the premolars and molars are straight and hypsodont, p2 67 mm long, p4 is 76 mm high, m2 47.4 mm. The metaconid and metastylid are rounded almost the same size. All the teeth with a remarkable cover of cement, protoconid and hypoconid roughly flat.
MPGJ 5125 is a fragment of left jaw collected in the GTO 2 A White layer, retains a robust symphysis with strong canines that are broken and the incisors with small infundibula without enamel. The jaw has its maximum depth of 101.3 mm, from the linguaflexid of p4 to the horizontal edge. The metaconid is lobular, and metastylid slightly pointed toward the lingual margin, V-shaped linguaflexid in molars, the ectoflexid is deep into the isthmus, the protoconid and hypoconid with well-defined flat edge in the labial border.
MPGJ 3872, is a fragment of left jaw with p2-p4. The most significant features are the metastylid pointed and the slightly rounded protoconid and hypoconid.
The jaw MPGJ 3946 from GTO 2B is very deep, the maximum depth is in p4 and m1. To determine the length of these molars in situ, a complete X-Ray was made, in which the length (HC) of the teeth was measured, and compared with the length of the mandible MPGJ 3739 from GTO 2 A. The size is like isolated molars collected in the same localities. The measurement of the molars included in the alveolus of the mandible is smaller compared with the specimens of the GTO 2 sites D and F (Figure 5).
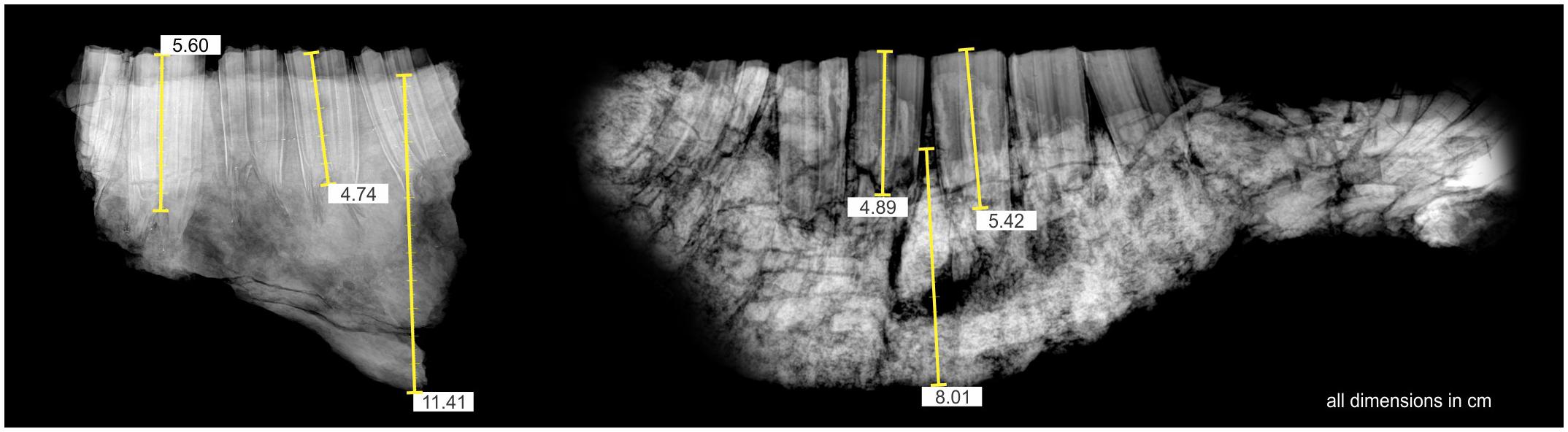
Figure 5. MPGJ 3739 Jaw fragment from GTO 2A collected in the Blanco layer. The jaw is very deep with full dentition. The X-Ray was taken to know the HC of p4-m1 and to determine its real size in the alveolus, to compare with the height of the crown of the molars of the F site of GTO2.
In the same stratigraphic sequence, at the Rhino and Blanco layers, numerous isolated molars have been collected. The analysis show similarities with the description mentions in lower molars that have been described in the jaws. However, at the top of the stratigraphic sequence, in clay deposits named GTO 2 site D and GTO 2 site F, lower and upper molars have been collected. The preservation is deficient, especially in length of the molars, however, the differences in size of the anteroposterior and transverse axis on the occlusal surface is remarkable.
The metaconid and metastylid are rounded and almost of the same size, the entoconid is round, the linguaflexid is open in V shape and wider in molars. Specimen MPGJ 5134 presents a pli caballinid, the protoconid and hypoconid large and flat in the lingual edge. The upper molars also differ from those collected from the Rhino layer: the crown is higher and straight, and the protocone developed. The Ap and Tr measurements of the p3-m2 of the mentioned jaws from GTO 2A and B have been plotted versus the molars of GTO 2 site D-F. It is remarkable the separation they have with the specimens collected in the sites GTO 2 D-F.
The MPGJ 5145 p4 from GTO 2 site F, was compared with MPGJ 5136 p4 of Equus simplicidens from GTO 6 Arrastracaballos locality early Blancan. The measurements are not very different, GTO 2 F Ap 29.87 Tr 17.46, the same measurements in MPGJ 5136 p4 Ap 32.53 Tr 20.23, which suggests that GTO 2 F specimens are a transitional group between the late Hemphillian Dinohippus and early Blancan Equus (Figures 6A,B).
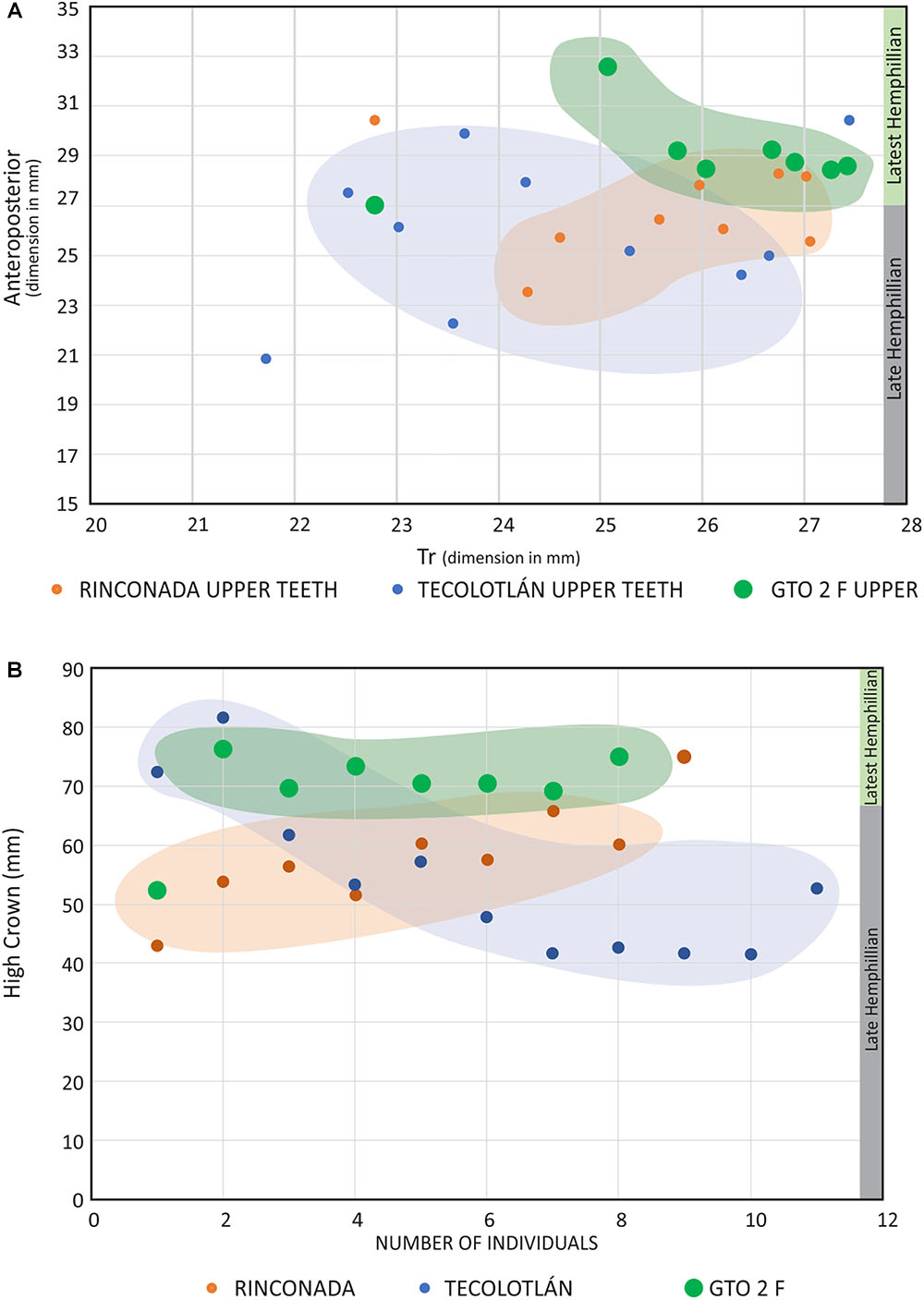
Figure 6. (A) All measurements of the Ap and Tr of M1 in upper molars including those of GTO 2 site F, have been plotted. (B) The measurements of the HC of P4-M1 were also graphed. The separation of GTO 2 F in the two graphs is remarkable, which suggests a different population.
The locality is situated 12 km southeast of Rancho El Ocote. The basal layer GTO 30, is a clay layer covered by ashy material, dated 5.32 ± 0.34 My. Above there is a sequence of clays of yellow color that contain Dinohippus material, besides abundant A. stockii and a few N. aztecus teeth. The upper part of the sedimentary sequence, located GTO 75, is covered by layers of compacted clay with abundant concretions. In these deposits, a fragment of skull and several jaws and isolated molars was collected isolated of D. mexicanus. It is the same stratigraphic sequence where the skunk Conepatus sanmiguelensis was recovered, which participated in previous GABI events. These sediments are covered by late Blancan-Irvingtonian deposits where Glyptotherium texanum was collected (Wang and Carranza-Castañeda, 2008; Gillette et al., 2016).
At the top of these deposits, the ash layer that covers the sequence was dated by the U/Pb method; the result is 4.54 ± 1.5 My. This means that the records of Dinohippus are bracketed with the lower age dated by 5.32 ± 0.34 My, results that gave the importance to these records.
GTO 75 Arroyo de Emilio MPGJ 103, skull fragment with palate and pat of the facial area. MPGJ 131 R P4-M3 and L P4-M1 from GTO 75 Same individual. MPGJ 857 R jaw p2-m3, L i1-3, Ri, 2-3. MPGJ 729 M1, MPGJ 1559 P4, MPGJ 5439. P4.
Lower teeth. GTO 30: MPGJ 310 L p2-m3, IGCU 5201 p2-m1 -m3, c, L i1-3, R i2-3. Isolated lower teeth, GTO 30 5306 p4, IGCU 5308 m2, IGCU 5593 m1, GTO 30. Upper teeth: IGCU 5437 P4, IGCU 5593 M1MPGJ 5438 P3, MPGJ 5578 M1, IGCU 5580 P4.
In deposits of the Coecillos area. GTO 30 Coecillos and GTO 75, Arroyo de Emilio localities, late Hemphillian age.
The skull MPGJ 103 from GTO 75 Arroyo de Emilio of the Coecillos area, consists of a fragment that preserves the complete palate and molars of both sides, the maximum length P3-M3 is 121 mm. The maxillary in the right side is the best preserved part including the wall of the orbit; however, there is no evidences of DPOF. The malar crest is well developed and ends in the anterior part of M1; there is no evidence of a malar fossa. The palate is well preserved in the ventral part, the posterior nasal orifices, the palatine bones, vomer, the anterior palatine holes and the palatal grooves. The maximum length Ap of the choana is 108 mm, the maximum transverse on the back of M3 is 49 mm. The maximum width of the palate between the lingual edge of M3 is 72 mm, maximum width between lingual edge of the protocone of P4 is 62 mm.
This skull MPGJ 103, is an old individual that does not present evidence of DPOF. According to the stratigraphic position almost at the top of the Coecillos sequence, the deposits where it was collected are only one meter below the ash that was analyzed and gave the result of 4.5 ± 1.5 My. It is possible to argue that corresponds to a primitive Equus with the support of the character of the teeth, however, the mandible MPGJ 857 that was collected at the same stratigraphic level presents characters in the dentition that are within the variability of D. mexicanus, although the protoconid and hypoconid are flat in the premolars and a little less in molars (Figures 7A,B).

Figure 7. (A) MPGJ 103, locality GTO 75 Arroyo de Emilio, we observe the facial region on the right side, part of the orbit, the nasal and maxillary bone without evidence of DPOF. The molar crest is strong and ends in P4. (B) MPGJ 857 from the Coecillos area, right jaw with p2-m3, c and R and L i1-3. The symphysis is complete, it is observed that the incisors do not have a defined infundibulum, the hypoconid is slightly flatted in premolars and the protoconid is more rounded.
Located almost in middle of the San Miguel Allende basin, limited by the Rancho Viejo hills in the east and the Tequisquiapan in the west. The fossiliferous deposits are 7 m thick.
The basal layer is unconsolidated sandy clay, covered by a lacustrine deposit about 1–2 m thick. This layer represents the bottom of the ancient lake, around 25 km long. The lithology is a sandy clay layer cemented by calcite and silica with a lens of volcanic ash and paleochannels of fine sand and gravel. All the fossils have been collected in this layer. A sandy clay layer covers the lake, and the top of the sequence is overlain by the unconformity of Hemphillian-Blancan age with a thickness of 1–2 m.
The equids collected in these sediments, N. eurystyle, A. stockii, and D. mexicanus, are associated with the best collection of carnivore that have been collected in the Hemphillian localities.
The faunal association is a typical of the late Hemphillian fauna Hh3. The result of radiometric analysis of the volcanic ash by the method of fission track of zircons is 4.4 ± 0.3 confirming this age (Carranza-Castañeda, 1992; Kowallis et al., 1999; Carranza-Castañeda et al., 2013).
GTO 56 MPGJ 856 of skull fragment with two Maxillaries with P2-M3, corresponds to an old adult individual by the wear of the molars. Preorbital dorsal fossa (DPOF), is in the nasal and maxillary bones.
The locality GTO 56, late Hemphillian age, is located about 700 m south of the Rinconada Locality.
Locality GTO 43 Rinconada upper teeth: MPGJ 5137 M1, MPGJ 5138 P4, MPGJ 5139 P4, MPGJ 5140 P4, MPGJ 5141 P4, MPGJ 5142 M1, IGCU 8180 m2. IGCU 8879 P3, IGCU 9590 P4, MPGJ 5143 P4-M1, MPGJ 3764 flat Incisor row i1-3 R and L.
MPGJ 856, two maxillaries with P2-M3, and fragment of nasal-maxillary bones with a dorsal fossa (DPOF), not pocketed posteriorly, like a shallow wide groove, the edges are smooth and poorly defined, with a maximum length of 60 mm.
The right maxillary is the most complete. The crest of the malar bone ends 25 mm above the anterior part of M1. The dentition is complete with P1-M3, (P2-M3, 159 mm, P1 is not included). The teeth are simple without folding in the fossettes, the hypocone and the groove are not present, the protocone is round and small in the premolars and slightly longer in M2-M3. On M1 it is worn away (Figure 8).

Figure 8. (A) Left and right maxillaries, notice the teeth without hypocone. (B) Nasal bone with shallow DPOF, and smooth borders. The three bones belongs to the same individual.
Lower Dentition. GTO 43 Rinconada locality, MPGJ 5132 jaw fragment Rp3-m2, MPGJ 5133 Rp2-m2 jaw fragment, 5136 p2-m3, MPGJ 10071 p4-m3. Isolated molars, GTO 43 Rinconada locality IGCU 8180 m2.
The jaws of this locality present some characters that differentiate them from the jaws collected in Rancho El Ocote. Among the important differences is the depth of the mandibular ramus. The specimen MPGJ 5136 of GTO 43, the maximum depth between p4-m1 is 72.43 mm, and on MPGJ 5132 with only p2-m2, the depth between p4-m1 is 75.56. In contrast, the mandible MPGJ 3749, of GTO 2A, the depth is 11.41 cm, and in MPGJ 3946 it is ∼101.48 cm. Besides the linguaflexid in the jaws, MPGJ 5132, IGCU 5133 and MPGJ 5136 is wide and shallow. The metaconid is lobular and the metastylid is rounded with on acute and small lingual border. The entoconid is lobular with an open hypoconulid. In IGCU 6963, p3 and p4, have a small pli caballinid. The ectoflexid is deeper in m1 and m2. The protoconid and hypoconid are flat in premolars and relatively rounded in molars. On MPGJ 5136, MPGJ 5132, and MPGJ 5133, lower jaws, the protoconid and hypoconid are variable from flat to slightly rounded, and the linguaflexid is shallow and open (Figures 9A–C).
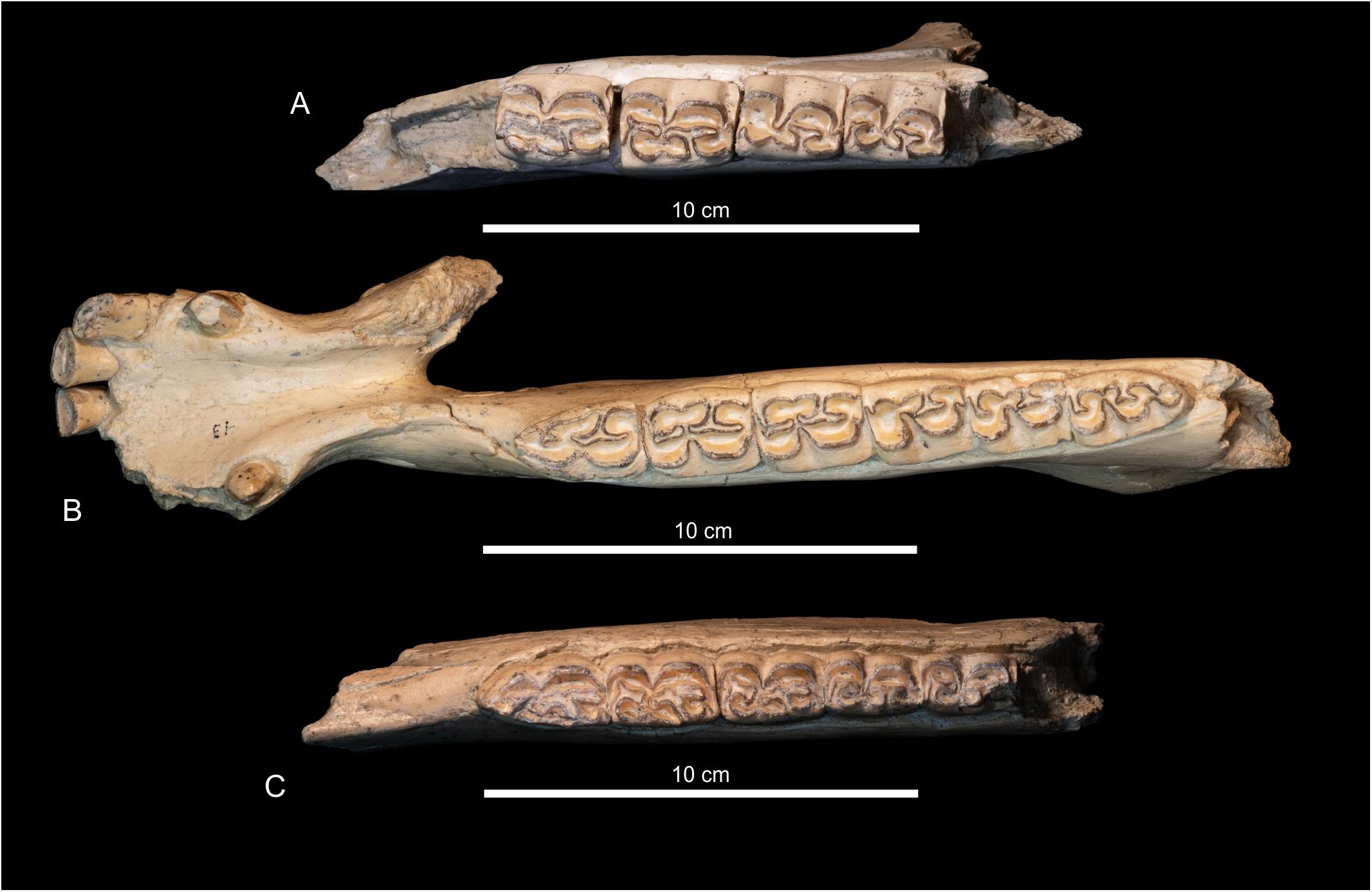
Figure 9. Jaws from GTO 43 Rinconada locality. (A) MPGJ 5132 p3-m2. (B) 5132 p2-m3. (C) MPGJ 5133 p2-m2. The linguaflexid is wide and shallow, the protoconid and hypoconid show inconsistency in the form since flatted to slightly rounded, the symphysis is not complete rounded.
The upper and lower molars were widely discussed and compared between the localities of San Miguel de Allende and the material of the Yepomera fauna that is housed in LACM, reaching the conclusion that they correspond to the same species. However, in this work it has been observed that in the lower molars, the linguaflexid, protoconid and hypoconid are different and that separates them from the specimens collected in the Rhino level of Rancho el Ocote and from the specimens at the base of the sequence of Coecillos (Carranza-Castañeda, 1992).
This basin is 90 km from the city of Guadalajara and is filled with fluviolacustrine deposits contain abundant North American Mammalian faunas, which represent the late Hemphillian age. The stratigraphic sequence has a thickness of 60 m and several sites have been mentioned where the late Hemphillian material has been collected. However, the stratigraphic sequence has been identified in two areas of main importance: Santa María sequence at the base and San José deposits that cover them. Resting in erosional unconformity are found the San Buenaventura deposits, thought to be of Blancan age (Carranza-Castañeda, 2016; Kowallis et al., 2017; McDonald and Carranza-Castañeda, 2017).
The mammalian fauna between the two areas, Santa María and San José, is almost the same. D. mexicanus and N. eurystyle remains are common in all the localities. The major difference is related to the presence of N. aztecus and A. stockii that so far never have been collected in the San José sequence. In these deposits are the last records of Machairodus, B. secundus, Teleoceras, Neohipparion, and Nannippus, that are well represented in the late Hemphillian fauna in central Mexico; among which has been described one of the first records of South American immigrants Zacatzontli tecolotlanensis that document the initial events of the GABI (McDonald and Carranza-Castañeda, 2017).
The age of the fauna has been determined by radiometric dating of the ashes that are interbedded with the fossiliferous strata. The ash found in the upper part of the San José area, was dated by the method of 40Ar/39Ar, and gave an age of 4.95 ± 0.02 My and (Kowallis et al., 2017). In recent works, a new date was made by the Laboratorio de Estudios Isotopicos, Centro de Geociencias, UNAM (LA-ICPMS), from a different site, by the method of U/Pb using zircons, which produced a date of 4.85 ± 0.1 My for the ash (McDonald and Carranza-Castañeda, 2017).
In erosional discordance are the deposits of the San Buenaventura sequence which has a thickness of ∼30 m of gravels and paleochannels of sand and clays. The contact with the deposits of San Jose is not exposed. The age of the sequence of San Buenaventura has been referred to the Blancan (Kowallis et al., 2017).
This sequence is important in the transition Dinohippus-Equus because some isolated teeth collected in the locality of Jal Teco 7 Las Gravas present more progressive characters by their grade of hypsodont and curvature that resembles Equus, and have been referred to the Blancan age.
MPGJ 1967 JT 35 late Hemphillian age skull fragment of young individual, with region facial and brain case of the right side well preserved. The M1-M2 first stages of wear, P4-M3 in the alveolus without wear.
Late Hemphillian. Isolated teeth: Jal Teco 1, MPGJ M1 Fragment. JalTeco 48: MPGJ 4044 M1, MPGJ 4045 M1, MPGJ 4046 P4.
Jal Teco 37, MPGJ 5025 M2, MPGJ 5029 p4, JalTeco 26 MPGJ 3541 M1; JalTeco 12 MPGJ 5001 M3, MPGJ 5002 P4. MPGJ 5003 p4, MPGJ 5004 m3. JalTeco 33 MPGJ 5027 p4, MPGJ 5028 m2, MPGJ 5026 M1. JalTeco 35, MPGJ 5102 M1. JalTeco 47 MPGJ 5072 m1-2. MPGJ 5069 m2.
Isolated Teeth. Jal Teco 7 Las Gravas locality Blancan age: Upper Teeth: MPGJ 3227 M1, MPGJ 3698 M3, MPGJ 3699 M12, MPGJ 3987 M1, MPGJ 4021 M1, MPGJ 5057 M3, MPGJ 5022 M1, MPGJ 5058 P4, MPGJ 5059 M3.
Lower Teeth, MPGJ 3246 p4, MPGJ 3226 p34, MPGJ 3985 m3, MPGJ 5052 m2, MPGJ 5049 m1, MPGJ 5050 m3, MPGJ 5051 m1, MPGJ 5052 m1, MPGJ 5053 m2, MPGJ 5054 p4, MPGJ 5136 p4.
MPGJ 1967 skull fragment, JalTeco 35 Locality, late Hemphillian age collector Mr. Harley Garbani and the author.
Specimen MPGJ 1967 is a skull fragment of a young individual that conserves only the left facial part with the frontal bones and part of the parietals. In the left facial part, in the anterior of the nasal and maxillary bones a well-defined deep dorsal fossa with the delimited edges, pocketed posteriorly with delimited edges; the fossa is wider above the P4. The orbit is complete, the zygomatic apophysis of the temporal bone, the temporal bone and the zygomatic apophysis of the malar are complete. The malar crest is prominent and ends between P4 and M1.
The dentition is reduced to M1 and M2 because P4 and M3 are erupting from the alveolus. The fossettes are simple, almost without plications, the prefossette of M1 presents a small loop, the postfossette has with small plications in the anterior edge and small pli hypostyle. The hypocone is wide with an open and deep furrow. A small and thin plicaballin is present in M1, and the protocone is elongated, without a heel (Figure 10).

Figure 10. MPGJ 1967 from JalTeco 35, left side, the facial fossa is pocketed with well-defined borders and strong malar crest.
Isolated teeth, the molar MPGJ 3987 M1 has a high crown, but the curvature is pronounced, the protocone larger the lingual border is concave with “wooden-shoe shape,” and present a small pointed heel. MPGJ 5022 M1, that correspond to mature individual, the curvature is similar but the protocone is small rounded and without heel. MPGJ 3698 M3 although is an M3 the protocone is larger and the tooth is curved like MPGJ M3 5001 late Hemphillian from locality JT 12 Hilda’s late Hemphillian locality of San Jose sequence in Tecolotlán basin.
The specimen MPGJ 5058 P3 from JalTeco 7 Ap 30.32 Tr 26.67 HC 54.25 mm in comparison with MPGJ 5141 P4 GTO 43 Ap 28.08 Tr 26.16, and HC 65.43 mm, they are similar in the structures and the size is less than 10%, however the protocone has the same shape, concave in the lingual border, developed heel, pli caballin in both and fossettes with same plications.
It is important to point out that the JalTeco 7 molars present the overlap of progressive characters, of a primitive Equus, with high crown hipsodoncia, protocone with heel and evident curvature, characters of the Blancan molars. In comparison to other teeth from the sequence of Santa María and San Jose, which have more “primitive” characters of Dinohippus. The overlap of characters in molars of typical Dinohippus to transitional molars of a primitive Equus, had already been noticed by MacFadden (personal communication, MacFadden and Carranza-Castañeda, 2002; Figure 11).
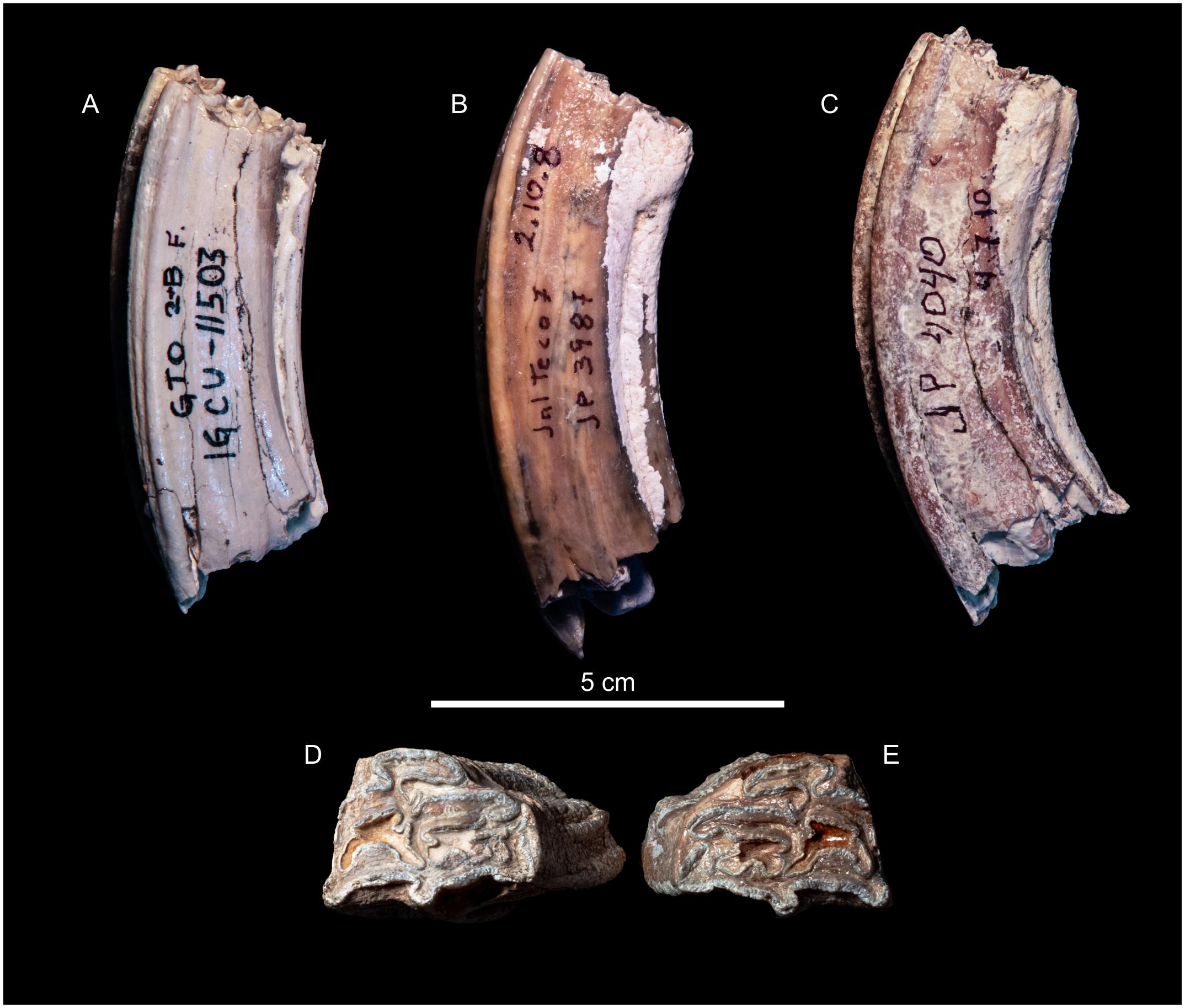
Figure 11. (A) IGCU 11503 P4, from GTO 2 site F Latest Hemphillian. (B) MPGJ 3987 M1 JalTeco 7 Blancan age. (C) MPGJ 4040 M1 GTO 2 site F. Latest Hemphillian. Comparison in the height of the crown and the curvature of the molars, in (A) the height of the crown is more straight than in (B), and in (C) the curvature of the crown is greater than in (A) and (B), and the height of the crown is also greater than in (A) and (B). However, molar (B) (Blancan age from Tecolotlán), shows intermediate characters. (D) (MPGJ 3698 M3 JalTeco 7 Blancan age) and (E) (MPGJ 5001 M3 JalTeco 12 late Hemphillian age) is remarkable the similarity of occlusal surface structures in molars of different ages from the same area.
Lower Teeth. The specimen that have been collected are scarce and damaged, correspond to adult individuals only one is a young premolar. In general, the structures of the occlusal surface have small affinities in comparison with lower molars from the GTO 2 site F, from the Blanco layer in Rancho el Ocote. Hemphillian localities. The linguaflexid is shallow and width in specimens MPGJ 3246 p4, 5050 m2, 5049 p4, 5053 p43, and 5059 p3, all these teeth the most significant difference is the protoconid and hypoconid flat, metaconid lobular and metastylid is smaller and pointed lingually. Deep ectoflexid in molars, short not passing the isthmus in premolars, the paralophid is high until the middle of the metaconid, and some teeth presents a small pli caballinid.
La Plegaria Locality. In the southwestern region of the state of Hidalgo, southwest of the city Tepeji Del Río, several localities with fossil remains were found. The most important is La Plegaria.
The mammal fossils are at the top of the lake sediments that have a thickness greater than 100 m and are evidence of the huge lakes that existed in central Mexico during the late Miocene. The fluvial sediments are at the top of the sequence. This is an area of little extension that is covered by vegetation, however, the fossils are found in clay, sand channels and gravel. D. mexicanus is an important part of these fauna (Padilla Gutiérrez, 2004).
El Valle del Gigante. At the top of the Sierra Madre Occidental, is the rancheria of El Valle del Gigante, which was initially explored and investigated by Dr. Theodore Downs and Mr. Harley J. Garbani from LACM, in Valle de El Gigante. Years later, the author continued the research in this area and discovered a new locality Nay 1 El Gigante. The continuity of the research in this area, led to the discovery of other localities. In the town of Huajimic Nay 2 El Huichol, there is material of upper and lower molars of D. mexicanus, and molars that were referred to as Rhynchotherium (?). The research in the north part of Nay 1, a new location Nay 3 El Trapiche, was discovered in the rancheria of the same name. Preliminary results are a collection of upper molars of D. mexicanus and fragments of the upper molar of Teleoceras, besides fragments of proboscideans. The age of this fauna has been referred to late Hemphillian, by analysis of the volcanic ash collected in the town of El Trapiche was analyzed by the method of U/Pb and gave an age of 5.50 ± 0.3 My.
All the skulls collected in the early-late and late Hemphillian faunas in central Mexico present a wide range of variability in the DPOF. The skull from Juchipila considered the oldest record of D. mexicanus in central Mexico, was compared with a mold 41478-1 of D. leidyanus housed in the collection of the University of Texas at Austin. The main differences in D. leidyanus cast, the dorsal (DPOF) and malar fossa are well developed. In addition, the protocones are slightly inclined toward the lingual edge.
In the Mexican specimen, the protocone is larger and horizontal to the maxillary lingual border. This is an important characteristic that eliminates the similarity between them (Matthew and Stirton, 1930; Azzaroli, 1988). The comparison with illustrations of D. interpolatus (Matthew and Stirton, 1930), also demonstrate important differences because the DPOF is always well developed and bounded at its edges, which does not occur in the skull of Juchipila which has smooth edges and is very shallow. Also in D. interpolatus the malar fossa is constant and absent in the skull of Juchipila and all the Mexican skulls. Regarding the curvature of the molars, it is difficult to determine this characteristic in the Juchipila skull because the molars are included in the maxilla. However, the isolated teeth of MPGJ 4067 Zac Juch 47 M12 from El Resbalón, are short and curved, the protocone is rounded, small and slightly oriented to the lingual edge. That reminds us of D. interpolatus, but this does not always occur in the molars of Juchipila. Most of the teeth from Juchipila have the anterior border developing a small heel, and the lingual border is flat and horizontal.
The skull from Juchipila was compared with the description of the skull of D. mexicanus collected in the Rhino level of Rancho El Ocote locality GTO 2B, (MacFadden and Carranza-Castañeda, 2002). Some similarities are evident in both specimens.
The nasal notch is in the same position, posterior to P2; the infraorbital foramen is 68 mm above P4. The malar fossa is absent, and the malar crest is strong and ends between P4-M1. The biggest difference is the DPOF. In the Guanajuato specimen it is pocketed posteriorly, deeper and the edges are well defined and is 80-mm anterior to the orbit. The specimen from Juchipila has only a faint depression 58 mm in front the orbit, and is shallow, with poorly defined borders. However, when compared the facial region of the Tecolotlán skull, the fossa is like the Rancho El Ocote specimen which is well delimited posteriorly, deep, and the edges are well defined.
The skulls differ in the size of the bar separating the orbit of the DPOF margin. The Juchipila skull is ∼58 mm, the specimen from Tecolotlán it is 61 mm, and the Rancho el Ocote is ∼80 mm. The skull from the GTO 56 Rinconada area has differences that distinguish it from the other skulls, the DPOF, is a slight depression with soft edges. In addition, the absence of the hypocone and hypoconal grooves possibly represent a primitive form of Equus (Figure 12).

Figure 12. Comparison of the ventral view of the Dinohippus skulls. (A) MPGJ 1982 from ZAC Juch 30. (B) IGM 7596 Rancho el Ocote. (C) MPGJ. 856 GTO 56 Rinconada área. The differences in the structures of the molars are evident, specially observe the variability in the protocone and absence of hypocone in (C).
The skull MPGJ 103 from GTO 75 Arroyo de Emilio, not present evidence of DPOF, as the structures and size suggest a transitional form between Dinohippus and a primitive Equus. However, the upper volcanic ash 4.54 ± 15 My and the associated fauna including Dinohippus mexicanus, N. eurystyle, and N. aztecus with the jaw MPGJ 857 collected in the lower layer, dated 5.32 ± 0.14 My; these ages undoubtedly encloses all the fauna in late Hemphillian age. Also, the C. sanmiguelensis specimen was collected in the same stratigraphic level (Wang and Carranza-Castañeda, 2008).
The comparison between the rows of teeth P2/p2 and M3/m3, shows a similarity in size. When comparing the P2-M3 row in the IGM 7596 from Rancho El Ocote fauna is 157.9 mm, the P2 with a prominent anterostyle. The same measurement in the Juchipila specimen is 150∼, though the anterostyle of P2 is broken. In the skull MPGJ 856 from Rinconada area, the length P2-M3 is 150 mm. The similarity in size of the three specimens is remarkable. The most important differences are the pronounced anterostyle of Rancho el Ocote, and the presence of P1 in the Rinconada maxillary that is not present in the other specimens.
Regarding the protocone, a great variability is also observed in the shape and size of the upper molars. The most noticeable difference is observed when comparing the protocones of the Rancho el Ocote skull where the anterior border is round or very small, the heel is almost non-existent, like the Rinconada specimen. The specimen from Tecolotlán MPGJ 1967 is a young individual, and some differences are observed. The anterior border of the protocone of M1 is rounded, in M2 which is incompletely erupted, the anterior edge is elongated and sharp forming a small heel. The posterior part is elongated and pointed. These variations can be explained because the skull belongs to a young individual. On the contrary, the dentition from Juchipila skull, referred to the early late Hemphillian age, the protocones should be more related to the shape of D. interpolatus and D. leidyanus, with its axis slightly inclined to the lingual border, and the anterior edge without heel. However, in the skull of Juchipila, the shape of the protocones is elongated with a horizontal and concave lingual border and with a prominent and unexpected heel. Nevertheless, the specimen MPGJ 4067 P4-M1 R and L of the locality Zac Juch 47 El Resbalón of the same age, the protocones are small, lobular without vestiges of a heel and are different from those molars of the Mixtón teeth. These differences show the wide variability that exists in the protocone in individuals of the same age and stratigraphic level.
In comparison the Rancho El Ocote skull of late Hemphillian age, the protocones of the maxilla do not present a heel although the shape is larger. However, strong differences with the specimen MPGJ 131 at locality GTO 75 Arroyo de Emilio, collected in the upper part of the stratigraphic sequence of the Coecillos area were present. These teeth have elongated protocones with concave lingual border. The shape is like a wooden shoe and is remarkable in the R and L M1, but the anterior side is oval without a heel.
The curvature is another important characteristic. The specimen MPGJ 5112 of GTO 2, site F, is larger, in comparison to MPGJ 131 from late Hemphillian of Arroyo Emilio. The contrast between them in curvature and size is outstanding. The specimens MPGJ 5112 M1 site F, is high crowned, the axis Ap 30.84 and Tr 29.12 mm, has been compared with IGCU 5652 M1 referred to Equus, an old individual collected in GTO 39 Tlaloc, early Blancan. The measurements Ap 27.56 mm, TR 30.12, show similarity between them.
The Blancan specimen MPGJ 3987 M1 referred to E. simplicidens that correspond to a young individual, collected in the locality JalTeco 7 Las Gravas M1 referred to Equus, share some features with IGCU 11503 of Rancho El Ocote: the curvature and height crown are similar, both has elongated protocone with an evident triangular heel and concave lingual margin.
The specimens MPGJ 5126 P4 from GTO 2B-F, with minimum wear, the molar is straight and long like IGCU 11503 M1, when compared with MPGJ 4067 P4-M1 R and L from the locality Zac Juch 47, that have lobular and small protocone without vestiges of a heel. It is evident that there is great diversity of shape and size in the protocone of D. mexicanus in the faunas of central Mexico, even in young and old specimens of the same locality and stratigraphic level.
Lower Teeth. The jaws, maxillary and isolated teeth, show wide range of variability, but there are no differences from that already mentioned in previous works (Lance, 1950; Quinn, 1955; MacFadden, 1984, 2006, 2016; Carranza-Castañeda, 1992). However, several differences are observed. The metaconid and metastylid are lobular in molars and in premolars, the metastylid is smaller and often the lingual margin is pointed. The entoconid is lobular, the hypoconulid short although it is not constant or very reduced in the jaw MPGJ 857 Arroyo de Emilio and is absent in MPGJ 5136 of Rinconada. The linguaflexid in the MPGJ 5136 from Rinconada is open and shallow, however, the jaw IGCU 6963 that corresponds to a young individual, the linguaflexid is also deep. The specimen MPGJ 857 Arroyo Emilio has a V shape and is more closed, like the mandibula MPGJ 3946 of GTO 2 B level Rhino. The ectoflexid is deep in molars, the protoconid and hypoconid in the jaws GTO 2A MPGJ 5125 and MPGJ 5132 from GTO 43 Rinconada and jaw 310 from GTO 30 Coecillos are horizontal and flat. However, in the specimen MPGJ 5136 and 5132 from Rinconada, the protoconid and hypoconid are horizontal and flat in the premolars. Some of the jaws that are described retain part of the symphysis and the incisors. The differences between them are not significant in relation to their size.
Some of the incisors have an infundibulum limited by enamel and filled with cement, however, others only present a small lake without enamel. The disposition of the incisors is arranged in a circular or flat row, possibly this difference is related to grazing or a mixed feeding (Figure 13 and Supplementary Table 2).

Figure 13. The structures of the symphysis are compared, (A) MPGJ 5129 from GTO2B, (B) MPGJ 5126 from GTO 43, (C) MPGJ 857 from GTO 75, and (D) MPGJ 1683 from Zac Tab 34. Observe the infundibulum of the incisors that do not present enamel. The row of the incisors is rounded in (C), in (A), and (D) the row is more flatted.
The outstanding result of the research over the material in Rancho El Ocote, without a doubt, is the specimens that have been collected in the GTO 2 sites F and D. These differ in their measurements of the anteroposterior (Ap) and transverse (Tr) axis which are clearly distinguished from those collected in the stratigraphic sequence of the Rhino and White levels. All the teeth from GTO 2 Site F and D, the hypoconid and protoconid have their labial edges more flattened and straight. The ectoflexid is narrow but very deep in the molars, almost touching the inner edge of the linguaflexid and less deep in the premolars. The linguaflexid is wide and shallow. Another difference is the metastylid that tends to be small and more pointed at its lingual edge. In addition, Pli caballinid is frequent but very small.
All the measurements of the teeth in the jaws described in this paper were plotted on a graph. It is remarkable that all the material from GTO 2 site D-F are separated from the typical Hemphillian teeth, and suggest a transitional group between Dinohippus and primitive Equus, as seen in the Figures 6, 14.
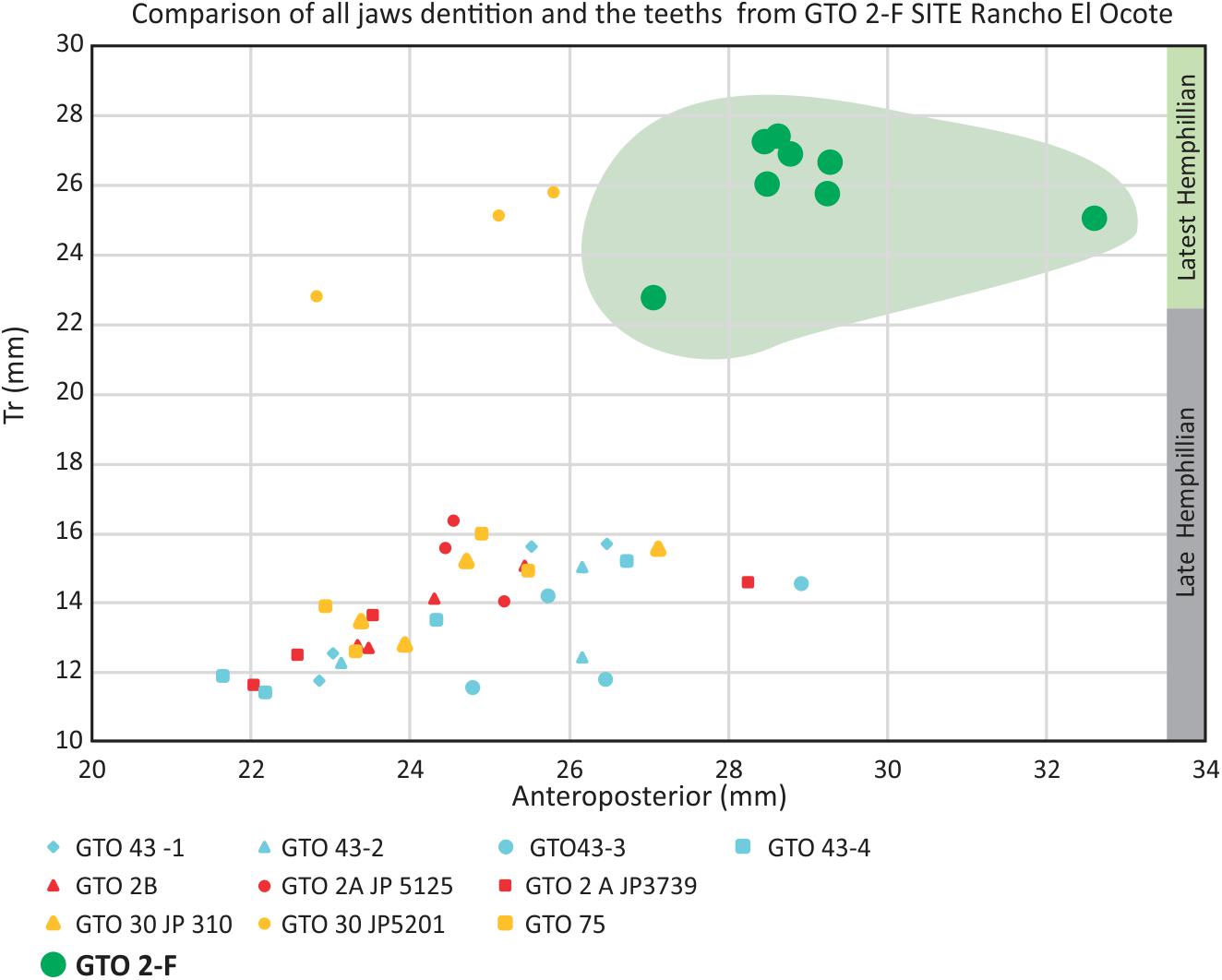
Figure 14. All the teeth p3-m2 of the jaws are plotted in the graph, including those of GTO2 site F. Is remarkable the separation all the teeth from site F. The result suggests that the molars of GTO 2 F and D correspond to a group of individuals different from those found in the late Hemphillian localities.
(1) Dinohippus mexicanus, has a wide geographical distribution in the central region of Mexico between the parallels 19° and 22°, only the most important localities due to the number of specimens, presence of skull, jaws and isolated upper and lower teeth by the variability they present in their structures and the stratigraphic position have been mentioned in this work. The main localities: early-late Hemphillian of Juchipila Zacatecas, late Hemphillian of Arroyo de Emilio Rinconada, Coecillos and Rhino layer of Rancho El Ocote; and latest Hemphillian in the Blanco layer, at Rancho El Ocote, basin of San Miguel de Allende, Guanajuato, late Hemphillian-early Blancan of Tecolotlán basin, Jalisco. The associate fauna always with stratigraphic indexes of late Hemphillian age and confirmed by radiometric ages.
(2) In D. mexicanus the morphology of the fossa dorsal (DPOF), was considered an important character in the diagnosis and systematics of the Pliohippus group of the Hemphillian age. However, the new records of skull collected in the faunas of early-late Hemphillian and late Hemphillian of the central region of Mexico, have shown that this structure has a wide variation in shape and size.
(3) The greatest differences observed in the skulls are more evident in the MPGJ 103 specimen of Arroyo de Emilio older individual, which has the largest facial region without evidence of DPOF, and the hypocone is not clear because the natural wear. These differences are comparable with the skull MPGJ 856 old individual from Rinconada, where the DPOF is reduced to a narrow and shallow groove and the hypocone is absent. It is possible to argue that these variants are determined by the ontogenetic age and natural wear. However, it is remarkable that these skulls were collected in deposits of 4.5 ± 15 My, (MPGJ 103) and in 4.4 ± 0.3 My (MPGJ 856), and in both the affinity with Equus is noticeable. However, the associated fauna of these two compared specimens, undoubtedly contains stratigraphic indices of the late Hemphillian age (Supplementary Table 1).
(4) The sample of skulls referred to Dinohippus mentioned in this work is the largest that has been described the differences observed in the DPOF induce to reconsider its value in the diagnosis of Dinohippus. The malar fossa is never present or insinuated in the facial region in the sample of skulls.
(5) The height of the crown in upper and lower molars is greater than 82.34 mm (MPGJ 4040, GTO 2B), 76.65 (MPGJ 5143, GTO 43) and in lower molars is 82.73 mm (MPGJ 5116 GTO 2 site F), these values are higher than the Yepomera samples and those mentioned for the fauna of Bone Valley, Florida (MacFadden, 2016). Always the cement covers the crown.
(6) The protocone has a wide variation of shape and size, even in molars collected at the same stratigraphic level vary from rounded to elongated although always the lingual edge is horizontal to the jaw. The anterior edge very often forms a heel that is not constant in all molars inclusive from the same locality, such as the molars of the skull of Juchipila and the molars MPGJ 4607 (ZJ 47).
(7) The MPGJ 856 skull from the Rinconada area, with shallow DPOF, very simple fossettes and absence of hypocone are features that make it distinctive to another skull. Also, the jaws: MPGJ 5136 P2-M3, MPGJ 5132 P3-M2, and MPGJ 6963 P2-M3 of the locality GTO 43 Rinconada, with the wide and shallow linguaflexid, protoconid and hypoconid are more flattened and straight; are features distinguish them from the mandibles of El Ocote and Coecillos. These differences suggest that the skull MPGJ 856 and the mandibles of GTO 43 Rinconada have significant affinity with Equus.
(8) The upper and lower molars collected in the Rhino and Blanco layers, have differences in size compared to those collected in the GTO 2 site D and F, on their AP and Tr axes and the height of the crown, indicating a “Group” with transitional characters between Dinohippus and a primitive Equus.
(9) The Graphs 6 and 14 have been plotted the measurements Ap, Tr and crown height of the upper and lower molars that have been discussed in this work, including the upper and lower molars of the sites GTO 2 site D and F. The result of this analysis suggests that D. mexicanus makes up a distinctive group closer (transitional) to Equus.
(10) In this work it has been shown the variability of the D. mexicanus collected in the Hemphillian localities from Zacatecas, Guanajuato and Jalisco localities: the curvature of the upper molars, the height of the crown in upper and lower molars besides the complexity of the DPOF and the variability of characters of the teeth. All these variations are evidences of the transition from D. mexicanus to Equus, in the faunas of the central region of Mexico in the boundaries Hemphillian-Blancan of Rancho El Ocote and Tecolotlán faunas.
Also, this has been observed too in the fauna of Yepomera (late Hemphillian) and La Concha (early Blancan) in the state of Chihuahua, where some lower molars cannot be reliably referred to Dinohippus (Lindsay, 1984).
(11) The specimens referred to D. mexicanus collected in the boundaries of late Hemphillian-early Blancan age, are the last records of D. mexicanus in Mexican faunas.
OC wrote the manuscript, conceived, interpreted, and collected the data for the graphics, and it is the main collector of the fossil material.
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The research of the sedimentary basins in the central region of Mexico began at the Instituto de Geología and have continued with the support of the Centro de Geociencias (Universidad Nacional Autónoma de México). I thank DGAPA, Project PAPIIT IN 102817, Dr. Xiaoming Wang Curator of LACM for the facilities provided to study the Yepomera collection, Dr. Jesús Alvarado, Director of the Collection Nacional de Paleontología, housed in the Instituto de Geología, for his assistance, Dr. Spencer Lucas of the MNHNM and Dr. Federico Angolin, by the review of this manuscript and their comments, Eduardo Jimenez for his comments in the review of this work, Ing. Jesus Silva-Corona for the photographs, graphics that illustrate this work, and edition, Biologist Hilda Troncoso-A. who made a large part of the preparation, assistance during the preparation of this manuscript, and her collaboration during the field work, MVZ Gerardo Carranza-Troncoso for the X-Ray radiograph, Dr. Carlos Ortega-Obregón of the Laboratorio de Estudios Isotópicos for the determination of radiometric ages of the area of Coecillos, Ing. Cesar Alfredo Contreras-Zamora for his collaboration in the solution of the problems of computation. Dr. Mariano Cerca, Centro de Geociencias, for his comments about the Clarendonian age, and the Earth Watch volunteer for the contributions in the Mexican Megafauna and Fossils of the Sierra Madre Projects.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2019.00089/full#supplementary-material
Aranda-Gómez, J. J., Carranza-Castañeda, O., Cervantes Corona, J., Wang, X., Tseng, J. J., Troncoso Altamirano, H., et al. (2008). Los sedimentos continentales (Formación Juchipila), en la región entre apozol y tabasco zacatecas. Unión Geofisíca Mexicana 28:190.
Arellano, A. R. V. (1951). Research on the continental neogene of Mexico. Am. J. Sci. 249, 604–616. doi: 10.2475/ajs.249.8.604
Azzaroli, A. (1988). On the equid genera dinohippus quinn 1955 and pliohippus marsh 1874. Boll. Soc. Paleontol. Ital. 27, 61–72.
Carranza-Castañeda, O. (1989). Rinocerontes de la fauna del Rancho El Ocote, Mioceno tardío (Henfiliano tardío) de Guanajuato, México. Univ. Nal. Autón. Méx. Inst. Geol. Rev. 8, 88–99.
Carranza-Castañeda, O. (1992). Una nueva localidad del henfiliano tardío de la mesa central. Univ. Nal. Autón. Mex. Rev. Inst. Geol. 10, 151–159.
Carranza-Castañeda, O. (2006). “Late Tertiary fossil localities in central Mexico between 19° 23°N, en Carranza-Castañeda,” in Advances in Late Tertiary Vertebrate Paleontology in Mexico and the Great American Biotic: Interchange. National Autonomous University of Mexico, Vol. 4, ed. E. H. Lindsay (Alexandria, VA: Instituto de Geología y Centro de Geociencias), 45–60.
Carranza-Castañeda, O. (2016). Roedores caviomorfos (Rodentia Hydrochoeridae) del blancano temprano-tardío – irvingtoniano de los estados de guanajuato, jalisco y sonora, México: relación con phugatherium dichroplax. Rev. Mex. de Cien. Geol. 33, 297–315.
Carranza-Castañeda, O. (2018). Early and late Hemphillian records of Gomphotherium hondurensis (Mammalia, Proboscidea) from central Mexico. New Mexico Mus. Nat. Hist. Sci. Bull. 79, 83–98.
Carranza-Castañeda, O., Aranda-Gómez, J., Wang, X., and Iriondo, A. (2013). The early late Hemphillian (Hh2) faunal assemblage from juchipila basin, state of zacatecas, México and its biostratigraphic correlation with other hemphillian faunas in central México. Los Angeles County Museum Cont. Sci. 521, 13–49.
Carranza-Castañeda, O., and Espinosa-Arrubarena, L. (1996). Late Tertiary equids from the State of Hidalgo, México. Univ. Nal. Autón. Méx. Inst. Geol. Rev. 11, 182–192.
Carranza-Castañeda, O., and Ferrusquía-Villafranca, I. (1978). Nuevas investigaciones sobre la fauna Rancho El Ocote Plioceno medio de Guanajuato, México; informe preliminar. Univ. Nac. Autón. de México Inst. Geol. Rev. 2, 163–166.
Carranza-Castañeda, O., and Ferrusquia-Villafranca, I. (1979). El Género Neohipparion (Mammalia-Perissodactyla) de la fauna local Rancho El Ocote (Plioceno medio) de Guanajuato, México. Univ. Nal. Autón. México Inst. Geol. Rev. 3, 29–38.
Cerca-Martínez, L. M., Aguirre-Díaz, G. and López-Martínez, M. (2000). The geologic evolution of the southern sierra de Guanajuato, Mexico: a documented example of the transition from the sierra madre occidental to the Mexican volcanic belt. Inter. Geol. Rev. 42, 131–151. doi: 10.1080/00206810009465073
Dalquest, W. W. (1988). Astrohippus and the origin of the Blancan and Pleistocene horses. Occas. Pap. Mus. Tex. Tech. Univ. 116, 1–23.
Dalquest, W. W., and Mooser, O. (1980). Late hemphillian mammals of the ocote local fauna, guanajuato, Mexico. Pearce Sellards Ser. 32, 1–25.
Flynn, J. J., Kowallis, B. J., Nuñez, C., Carranza-Castañeda, O., Miller, W. E., Swisher, C. C., et al. (2005). Geochronology of hemphillian-blancan aged strata, guanajuato, mexico, and implications for timing of the great american biotic interchange. J. Geol. 113, 287–307. doi: 10.1086/428806
Gillette, D. D., Carranza-Castañeda, O., White, S. R., Morgan, S. G., Trascher, C. L., McCord, R., et al. (2016). Ontogeny and sexual dimorphism of Glyptotherium texanum (Xenarthra- Cingulata) from the pliocene and pleistocene (Blancan-Irvingtonian NALMA) of Arizona, New Mexico and Mexico. J. Mammal. Evol. 22:3. doi: 10.1007/s10914-015-9309-6
Kowallis, B. J., Christiansen, E. H., Carranza-Castañeda, O., Miller, W. E., Ross, K. T., and Tingey, D. G. (2017). The geology of the tecolotlán graben, Jalisco, México. Geol. Soc. Am. Digital Map Chart 22:37. doi: 10.1130/2017.DMCH022.TXT
Kowallis, J. B., Swisher, C. C., Carranza-Castañeda, O., Miller, E. W., and Tingey, G. D. (1999). “Fission-track and single crystal 40 AR/39AR laser fusion ages from volcanic ash layers in fossil-bearing pliocene sediments in central México,” in Tectonic and Magmatic Evolution of México during the Cenozoic, Vol. 15, ed. Aguirre-Díaz (Santiago de Querétaro: Revista Mexicana De Ciencias Geologicas),157–160.
Lahiere, L. (1982). Petrology of Lacustrine Deposits, Juchipila Quadrangle, Zacatecas. Master thesis, University of New Orleans, New orleans.
Lance, J. F. (1950). Paleontología y estratigrafía del Plioceno de Yepómera, Estado de Chihuahua 1a parte: equidos, excepto Neohipparion. Univ. Nac. Autón. México Inst. Geol. 54, 1–81.
Ledezma, O. (1960). Bosquejo Geológico de la zona de Neutla, Guanajuato. Ph.D. thesis, Universidad Nacional Autónoma México, Mexico city.
Lindsay, E. H., Opdyke, N. D., and Johnson, N. M., (1984). Blancan-hemphillian land mammal ages and late cenozoic mammal dispersal events. Ann. Rev. Earth Planet. Sci. 12, 445–488.
López, J. C. (1991). Stratigraphy and Petrology of Miocene-Pliocene lacustrine Deposits, Juchipila Quadrangle, Zacatecas, México. Masters thesis, University or New Orleans, Lousiana.
MacFadden, B. J. (1984). Astrohippus and dinohippus from the yepómera local fauna late hemphillian, Mexico and implications for the phylogeny of one-toed horses, in origin and evolution of the cenozoic vertebrate fauna of Middle America, ed MacFadden. J. Vert. Paleont. 4, 273–283. doi: 10.1080/02724634.1984.10012009
MacFadden, B. J. (2006). “Early Pliocene (latest Hemphillian) horses from the Yepómera Local fauna, Chihuahua, Mexico,” in Advances in late Tertiary vertebrate paleontology in Mexico and the Great American Biotic Interchange, Vol. 4, eds O. Carranza-Castañeda and E. H. Lindsay (Alexandria, VA: Instituto de Geología y Centro de Geociencias), 33–43.
MacFadden, B. J. (2016). Late hemphillian monodactyl horses (Mammalia, Equidae) from the bone valley formation of central florida. J. Paleont. 60, 466–475. doi: 10.1017/S0022336000021995
MacFadden, B. J., and Carranza-Castañeda, O. (2002). Cranium of Dinohippus mexicanus (Mammalia, Equidae) from the early Pliocene (latest Hemphillian) of Central Mexico, and the origin of Equus. Bull. Fl. Mus. Nat. Hist. 43, 163–185.
Matthew, W. D., and Stirton, R. A. (1930). Equidae from the pliocene of texas: university of california Pub. Dept. Geol. Sci. Bull. 19, 509–556.
McDonald, G., and Carranza-Castañeda, O. (2017). Increased xenarthran diversity of the great american biotic interchange: a new genus and species of ground sloth (Mammalia, Xenarthra, Megalonychidae) from the hemphillian (late Miocene) of Jalisco, Mexico. J. Paleontol. 19, 1069–1082. doi: 10.1017/jpa.2017.45
Montellano-Ballesteros, M., and Carranza-Castañeda, O. (1986). Descripción de un milodóntido del Blancano temprano de la Mesa Central de México. Univ. Nac. Autón. Mexico Inst. Geol. Rev. 6, 193–203.
Mooser, O. (1958). Una cebra fósil de la mesa central de México. Anal. Inst. Biol. México 28, 359–363.
Mooser, O. (1960). Un équido fósil del género Neohipparion de la Mesa Central de México. Anal. Inst. Biol. México 30, 375–388.
Mooser, O. (1963). Neohipparion monias n. sp. équido fósil del Plioceno de la Mesa Central de México. Anal. Inst. Biol. México 34, 393–396.
Mooser, O. (1965). Una nueva especie de equido del genero Protohippuset al. del Plioceno de la Mesa Central de México. Anal. Inst. Biol. México 35, 157–158.
Mooser, O. (1968). Fossil equidae from the middle pliocene of the central plateau of Mexico. Southwest. Nat. 13, 1–12.
Mooser, O. (1973a). Neohipparion monias n. sp. équido fósil del Plioceno de la Mesa Central de México. Anal. Inst. Biol. México 34, 393–396.
Mooser, O. (1973b). Pliocene horses of the ocote local fauna, central plateau of México. Southw. Nat. 18, 257–268.
Padilla Gutiérrez, J. M. (2004). Bioestratigrafía del Area de Las Golondrinas, Terciario tardío de Tepeji del Río, Estado de Hidalgo, México. Master thesis, Universidad Nacional Autónoma de México, Ciudad de México.
Quinn, J. H. (1955). Miocene equidae of the texas gulf coastal plain. Univ. Tex. Bur. Econ. Geol. 5516, 1–102.
Robles-Rivera, C. (2015). Mastofaúnula del Clarendoniano tardío-Henfiliano temprano (Mioceno tardío), del área de Neutla, Municipio de Comonfort, estado de Guanajuato, México. Master thesis, Centro de Geociencias, Universidad Nacional Autónoma de México, Querétaro.
Sussman, R., Croxen, F. W., McDonald, H. G., and Shaw, C. A. (2016). Fossil porcupine (Mammalia-Rodentia, Erethizontidae) from El Golfo de Santa Clara, Sonora, Mexico, with a review of the taxonomy of the North American erethizontids. Los Angeles County Museum Cont. Sci. 524, 1–29.
Keywords: Dinohippus mexicanus, Equus, Hemphillian, central Mexico, Clarendonian
Citation: Carranza-Castañeda O (2019) Dinohippus mexicanus (Early-Late, Late, and Latest Hemphillian) and the Transition to Genus Equus, in Central Mexico Faunas. Front. Earth Sci. 7:89. doi: 10.3389/feart.2019.00089
Received: 01 February 2019; Accepted: 10 April 2019;
Published: 21 May 2019.
Edited by:
Leonardo Dos Santos Avilla, Universidade Federal do Rio de Janeiro, BrazilReviewed by:
Spencer G. Lucas, New Mexico Museum of Natural History and Science, United StatesCopyright © 2019 Carranza-Castañeda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Oscar Carranza-Castañeda, Y2FycmFuemFAZ2VvY2llbmNpYXMudW5hbS5teA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.