 Mariana Nabais
Mariana Nabais Anna Rufà
Anna Rufà Marina Igreja3,6
Marina Igreja3,6
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Archaeol., 24 July 2024
Sec. Zooarchaeology
Volume 3 - 2024 | https://doi.org/10.3389/fearc.2024.1411853
This article is part of the Research TopicNeanderthal Complex Behaviour Through the Lens of Faunal ResourcesView all 6 articles
The origin of complex behaviour amongst early humans is a subject of heated debate within the scientific community, and the study of small prey remains has become a significant aspect when examining such modern behaviour. Nonetheless, the consumption of small prey by human populations poses analytical difficulties due to the often negligible, or entirely absent, traces on bone surfaces. To address this difficulty, an experimental study focusing on terrestrial avifauna has been prepared, and here we present a preliminary phase of this research. The aim is to distinguish potential modifications on bird bone surfaces and fracture patterns that might facilitate the recognition of human manipulation of avian skeletal remains. Building upon the challenges encountered in the study of archaeological findings recovered from recent excavations in Iberian Middle Palaeolithic sites, the experimental protocol was formulated to encompass the processing of two uncooked and three roasted birds; and the lithic use-wear analysis of the flint flake used in the processing of raw birds. The results showcase distinct patterns of bone surface modifications and breakage between cooked and uncooked birds. Higher numbers of cut marks and manual disarticulation breaks are found on raw animals, whereas roasted animals show no cut marks, local-specific burns and higher bone loss. This pilot-study provides a baseline for future research to further explore the role of avifauna in Neanderthal subsistence and food processing, which may help highlight cultural choices.
The investigation of human dietary behaviours has long been a central theme in palaeoanthropological research, offering crucial insights into the adaptive strategies and subsistence practices of early human populations. Among the various components of prehistoric diets, the consumption of birds by Neanderthals emerges as an interesting and, yet, scarcely studied facet. This dietary choice, involving the deliberate use of avian resources, represents a vital area of inquiry, as it holds the potential to illuminate essential aspects of Neanderthal culture, adaptation, and ecological interactions.
Neanderthals inhabited a diverse range of environments in Eurasia for over 200,000 years. Their ability to thrive across a broad geographical expanse has prompted investigations into the means by which these humans secured their sustenance. While the consumption of large game has received considerable attention (e.g., Smith, 2015; Weyrich et al., 2017; Gaudzinski-Windheuser et al., 2023), the role of birds in Neanderthal diet has been overshadowed. However, recent archaeological discoveries and advancements in analytical techniques have allowed the opportunity to reassess the importance of avian use by Neanderthals. Birds offer a complementary dietary resource that may have played an essential role in Neanderthal adaptation and survival (e.g., Rufà and Laroulandie, 2021; Blasco et al., 2022; Nabais et al., 2023). Additionally, the use of bird feathers and other bird products carries different implications, encompassing technological, social, and symbolic dimensions (e.g., Finlayson et al., 2012; Morin and Laroulandie, 2012; Radovcic et al., 2015; Rodríguez-Hidalgo et al., 2019).
To assess the importance of birds in Neanderthal lifeways, and diets in particular, it is imperative to have a multidisciplinary approach. Therefore, this study promotes the application of archaeological experimentation to investigate and document taphonomic alterations on avian remains through detailed examination of bird bone surface modifications. This experimental endeavour is essential for identifying human-related bone surface alterations—indicative of bird use, cooking, butchering and consumption—so they can be distinguished from natural processes and carnivore-induced damage. By simulating early human cooking and butchering techniques, this pilot-study will provide a baseline for further experimentation and research in the identification and understanding of the human-signatures of avian exploitation. It offers the potential to shed light on the technological capabilities, cultural choices and cognitive faculties of early humans' avian resource use.
Bird specimens were recovered from the Wildlife Ecology, Rehabilitation and Surveillance Centre (CERVAS), which is part of the ICNF Serra da Estrela Natural Park, located in Gouveia (Portugal). The birds died under natural conditions in this rehabilitation centre, following veterinary checks and then frozen in order to preserve for scientific research after death. Information regarding the bird species and date of death used were collected by Dr. Ricardo Brandão from CERVAS.
The selection of birds tried to represent species commonly associated with Neanderthal diets in the Iberian Peninsula (e.g., Blasco et al., 2014, 2016; Martínez Valle et al., 2016; Nabais et al., 2023). However, given that the obtainability of wild animals depends on their natural death and availability, the species used were the nearest equivalents to those discovered in archaeological sites—i.e., taxonomically close relatives, similar in size to other bird species consumed by Neanderthals. Hence, the species used were Corvus corone (carrion crow; Birds 2 and 4), Columba palumbus (common wood-pigeon; Bird 5), and Streptopelia decaocto (Eurasian collared-dove; Birds 1 and 3), with a total of five individuals used in the study (Table 1; Supplementary material 1). Bird specimens were let to defrost 24 h prior the start of the experimentation so that carcasses could resemble their original raw state and appearance.
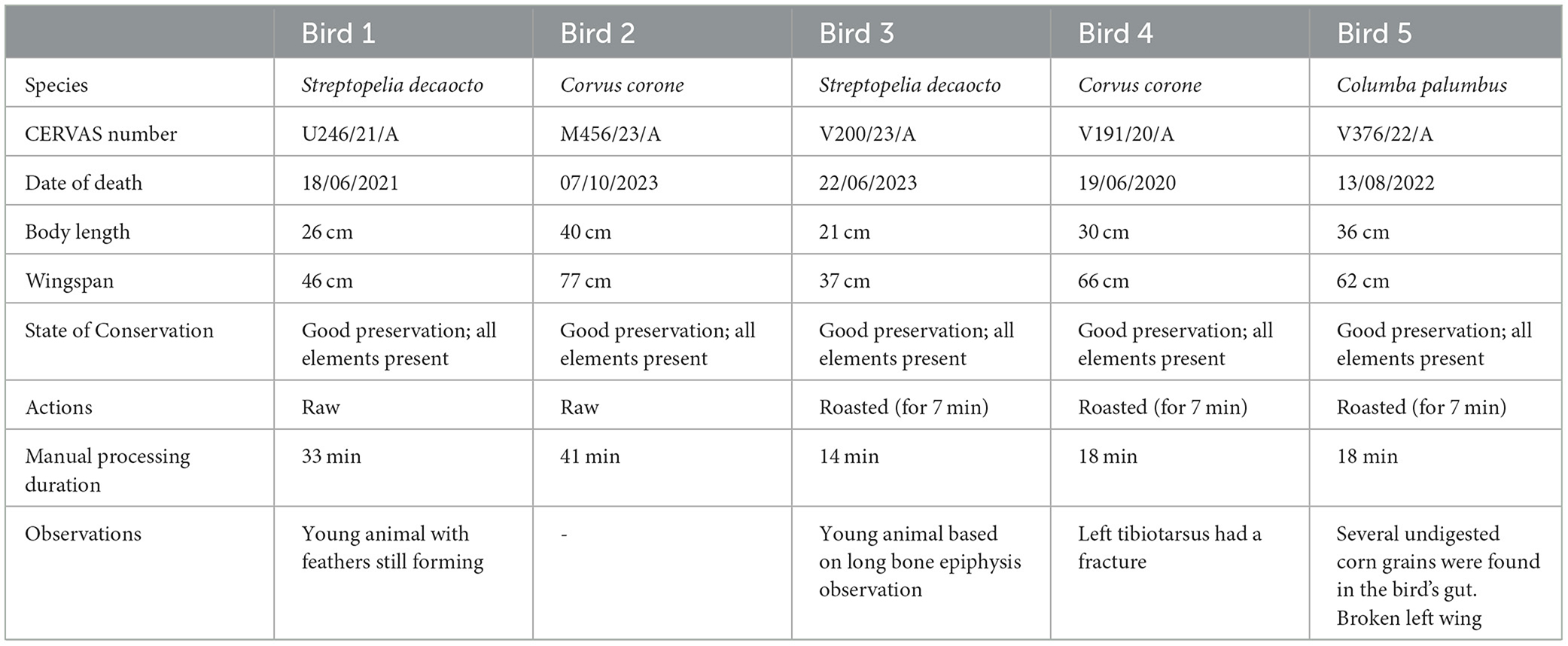
Table 1. Summary of bird specimens used in the experimental study detailing species name, corresponding rehabilitation centre (CERVAS) identifiers, date of death, physical measurements (body length and wingspan), state of conservation at the time of study, processing actions undertaken and duration of each experiment, and additional observations noted prior to and during the experimental procedure.
The birds were photographed and measured prior to processing for experimental purposes (Table 1; Supplementary material 1). The archaeological experimentation was conducted at ICArEHB research centre (Faro, Portugal), in a controlled research environment designed to replicate Neanderthal techniques for the treatment and culinary preparation of avian species. Consequently, birds were prepared, cooked and processed according to archaeological evidence and ethnographic data (e.g., Osgood, 1971; Nicolaysen, 1980; Laroulandie, 2001; Blasco and Fernández Peris, 2009). To acquire a better understanding regarding diverse potential Neanderthal consumption practices, two individuals were butchered uncooked (S. decaocto and C. corone, corresponding to Birds 1 and 2), whereas the other three were butchered after cooking (S. decaocto, C. corone, and C. palumbus; or Birds 3, 4, and 5). All avian specimens started by being manually defeathered and then, in the case of the raw individuals, immediately butchered. The other three specimens were cooked by roasting in direct contact with the coals whenever the temperature reached 500°C; they were cooked defeathered and complete, without being broken before heat exposure (Figure 1). Temperature was permanently measured using an Infrared Thermometer HS 960D. Complete birds were initially roasted with their bellies on the coals for 4 min, and then turned around and cooked for another 3 min. Cooking durations were determined empirically, based on the assessment of the birds' meat's doneness, which occurs quickly when directly exposed to the coals.
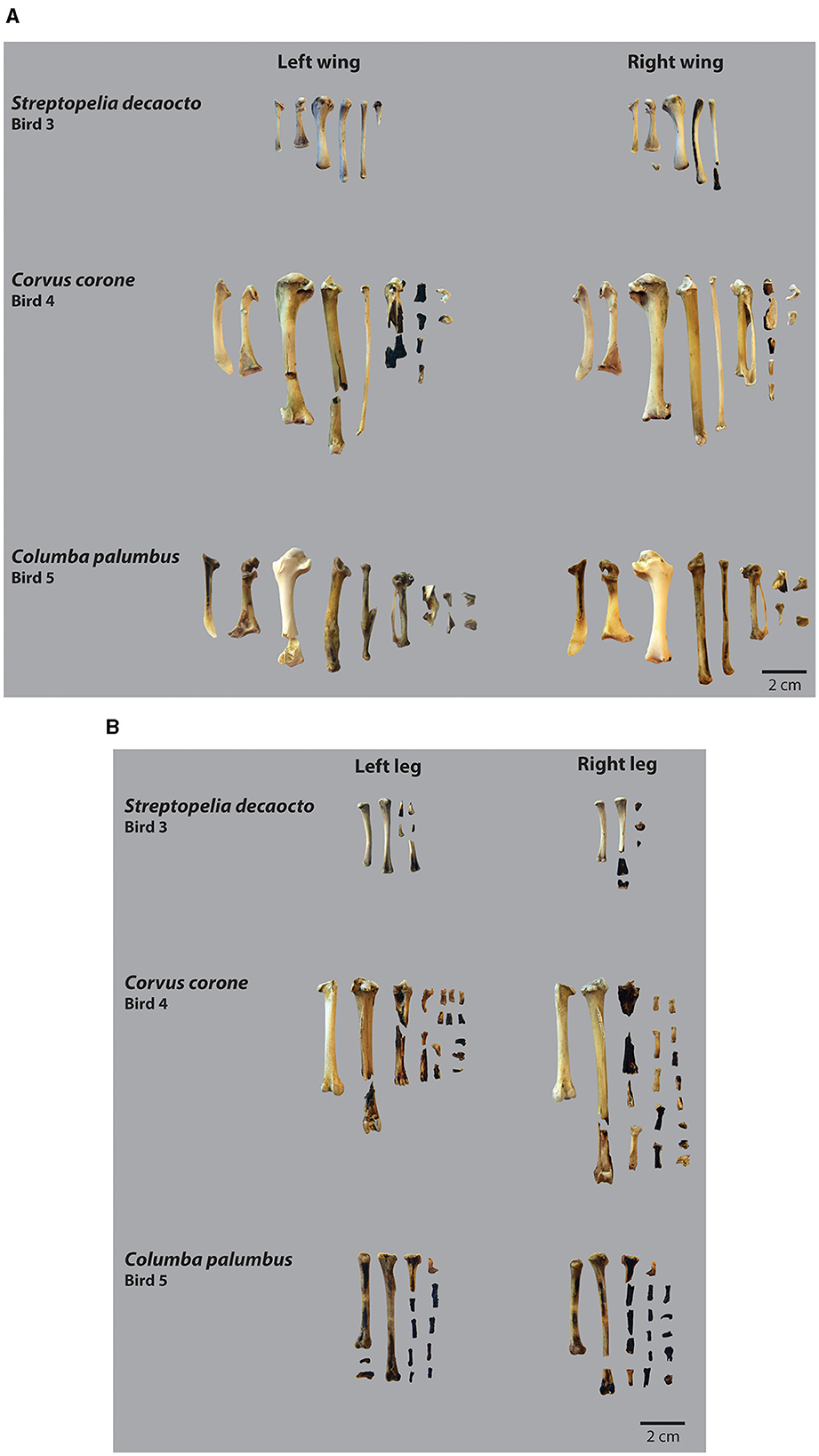
Figure 1. (A) Thermal alterations and impact of heat exposure on wing bones after experimental exposure to heat of three bird specimens, Streptopelia decaocto, Corvus corone, and Columba palumbus. (B) Thermal alterations and impact of heat exposure on leg bones after experimental exposure to heat of three bird specimens, Streptopelia decaocto, Corvus corone, and Columba palumbus.
The butchering techniques employed used manual handling, and movements such as flexion, twisting, pulling and overextension. Whenever needed, such actions were aided by an experimental flint flake (Supplementary material 1). The latter was produced by ICArEHB students during their practical sessions in lithic technology. Two of us (AR and MN) processed one raw bird specimen each (Birds 1 and 2, respectively). MN processed one roasted animal (Bird 5), and AR processed two (Birds 3 and 4). After processing, each bird's bones were placed in separate laundry bags to soak in Neutrase enzyme solution. This facilitates tissue removal within a stable 34–55°C range over 3–12 h (Simonsen et al., 2011). Post-enzyme action, bones were cleaned, brushed, and sun-dried to deactivate the enzymes.
The birds' remains underwent macroscopic and microscopic analysis (using a Hirox HR 5000E in the LARC-Archaeosciences Laboratory, in Lisbon) to document modifications on bone surfaces. This included recording all marks such as cuts, their distribution, location, and orientation, as well as patterns of breakage like fragment size, fracture outlines, angles, and edges. Given that most avian skeletal remains unearthed from Palaeolithic contexts predominantly consist of appendicular elements (e.g., Mourer-Chauviré, 1983; Ericson, 1987; Blasco et al., 2016; Romero et al., 2017; Rufà et al., 2018), this pilot-study focused on these anatomical components, specifically the bones associated with the wings and legs.
All experimental actions were recorded with a Nikon D5300 camera, an iPhone 11 and an Android Pixel 4a, and the video files are archived at the CORA repository (https://dataverse.csuc.cat/dataset.xhtml?persistentId=doi%3A10.34810%2Fdata1216) for future reference and potential replication of this experimentation.
Finally, it is important to acknowledge that the experimental setting may not fully replicate the exact environmental conditions experienced by Neanderthals, and our butchering experience may not mimic the exact movements used by these early humans. Therefore, such limitations should be considered when interpreting our results.
Use-wear analysis refers to the study of traces present on the artefact's surfaces that result from the wear produced by human use. It takes into account the fact that repeated actions performed with a stone tool leave micro- and macroscopic evidence of friction. It heavily relies on experimental reference collections that aim to replicate past potential uses and understand the formation process of identifiable diagnostic traces, to which the archaeological material can be compared.
Based on experimental replication, researchers showed that these modifications are known to be the result of the interaction between different surfaces (i.e., tool and worked material), which causes a gradual removal or deformations of the natural surface. The character of the discipline is based on a pattern recognition method when assessing the typology, location, and distribution of the different types of wear traces (Semenov, 1964; Tringham et al., 1974; Kamminga, 1979; Keeley, 1980; Plisson, 1985; Gonzales Urquijo and Ibanez Estevez, 1994; Andrefsky, 2005; McPherron et al., 2014; Claud et al., 2019a,b). Comparing the use-wear observed on replicated tools used in experimental actions to use-wear found on archaeological artefacts, wear patterns found on artefacts have been successfully classified. It is a key discipline and currently the most reliable method for obtaining direct evidence on tool use.
Successful interpretations of stone tool use depend on as many lines of evidence as possible and must rely on the observation of both micro- and macroscopic use-wear. The present study combines the analysis of macroscopic features (scarring, edge rounding, and fractures) seen with low magnification with microscopic use-wear (such as polishes and striae) observed with high magnification microscopy and relies on comparisons with extensive experiments that have been previously completed on a wide range of rocks, including flint, and are broadly recognised as reliable references (Keeley, 1980; Gonzales Urquijo and Ibanez Estevez, 1994; Claud et al., 2019a).
The experimentally made flint flake (Supplementary material 1) used for processing the uncooked birds was submitted to macro- and microscopic analysis of tools' edges and probable use zones in LARC-Archaeosciences Laboratory (Lisbon, Portugal).
Firstly, the flake was cleaned in order to remove any grease and residues that could adhere to it. Subsequently, the tool's edges and surfaces were examined under low and high magnifications by combining optical bright field reflected light and digital microscopy: the flake was first examined using an Olympus SZX12 and a Hirox HR 5000E, in order to macroscopically identify the used zones. Edges and surfaces were then analysed at high magnification with a Microscope Olympus BX60 for the observation and identification of the microscopic use-wear features indicative of specific materials worked and actions performed.
All birds underwent initial manual plucking of feathers from the head, neck, and body by gripping the feathers at the skin joint and pulling them sharply away. Subsequently, the processing of both raw and roasted animals involved a similar sequence of actions. Manual handling was used for all, but roasted animals disarticulated more readily, without the need of a cutting tool. The birds' heads were manually detached by twisting and pulling them free from the cervical vertebra. The chest skin was then slit longitudinally with the flint flake along the sternum's centre line. Flesh was cut away and internal organs were carefully detached and removed intact.
Wings were detached by twisting, and for raw birds, a flint flake was used to sever tendons, which is particularly evident on the cuts found on the scapula, coracoid and humerus (Figure 2, Table 2). Regardless of whether the bird was raw or roasted, the scapula and coracoid stayed attached to the axial skeleton, separating from the wing bones. The humerus was disarticulated from the radius and ulna through flexion in a direction opposing the joint's natural movement. In some cases, this action resulted in minor fractures at the distal end of the humerus (Figure 2A–F) and the proximal end of the ulna (Figure 2A–I, L). Furthermore, the distal humerus occasionally exhibited a peculiar type of perforation (Figure 2B–D). These observations are consistent with the wrenching, squashing and notching previously documented by Laroulandie et al. (2008). A similar flexion motion was used to separate the radius and ulna from the carpometacarpus, resulting in wrenching observed on the proximal or distal joints of the bones (Figure 2B–E, G).
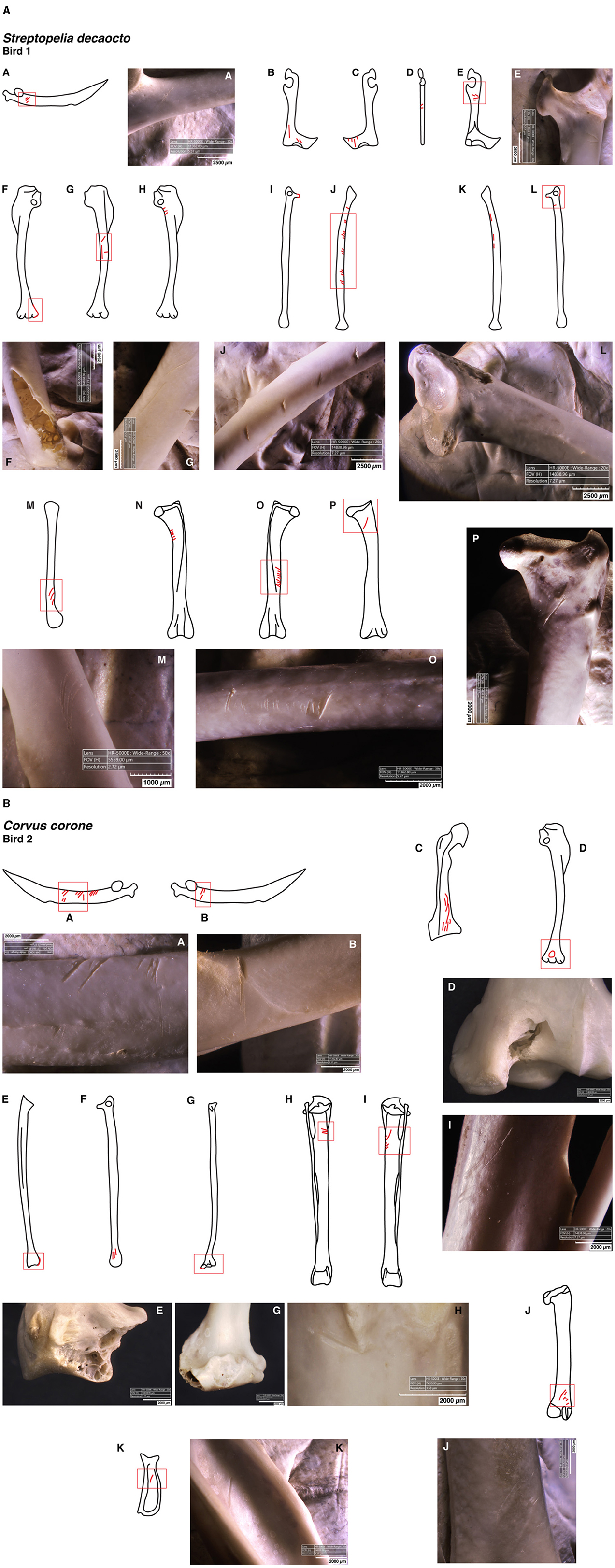
Figure 2. (A) Bone surface modifications on the appendicular skeleton of Streptopelia decaocto after being experimentally processed uncooked. Illustrations are not drawn to scale and should be interpreted as conceptual sketches; they are not suitable for species identification. A - Right scapula (lateral side): cut marks. B - Left coracoid (dorsal side): cut marks. C - Right coracoid (dorsal side): cut marks. D - Right coracoid (medial side): cut marks. E - Right coracoid (ventral side): cut marks. F - Left humerus (posterior side): notching. G - Left humerus (anterior side): cut marks. H - Right humerus (posterior side): cut marks. I - Left ulna (anterior side): wrenching mark. J - Left ulna (medial side): cut marks. K - Right ulna (medial side): cut marks. L - Right ulna (anterior side): wrenching and cut marks. M - Left radius (medial side): cut marks. N - Left femur (cranial side): cut marks. O - Right femur (cranial side): cut marks. P - Right femur (caudal side): cut marks. (B) Bone surface modifications on the appendicular skeleton of Corvus corone after being experimentally processed uncooked. Illustrations are not drawn to scale and should be interpreted as conceptual sketches; they are not suitable for species identification. A - Left scapula (lateral side): cut marks. B - Right scapula (lateral side): cut marks. C - Right coracoid (dorsal side): cut marks. D - Right humerus (posterior side): squashing mark. E - Right ulna (lateral side): wrenching mark. F - Right ulna (anterior side): cut marks. G - Right radius (lateral side): wrenching mark. H - Left tibiotarsus (caudal side): cut marks. I - Right tibiotarsus (caudal side): cut marks. J - Right femur (caudal side): cut marks. K - Right major digit phalanx 1: cut mark.
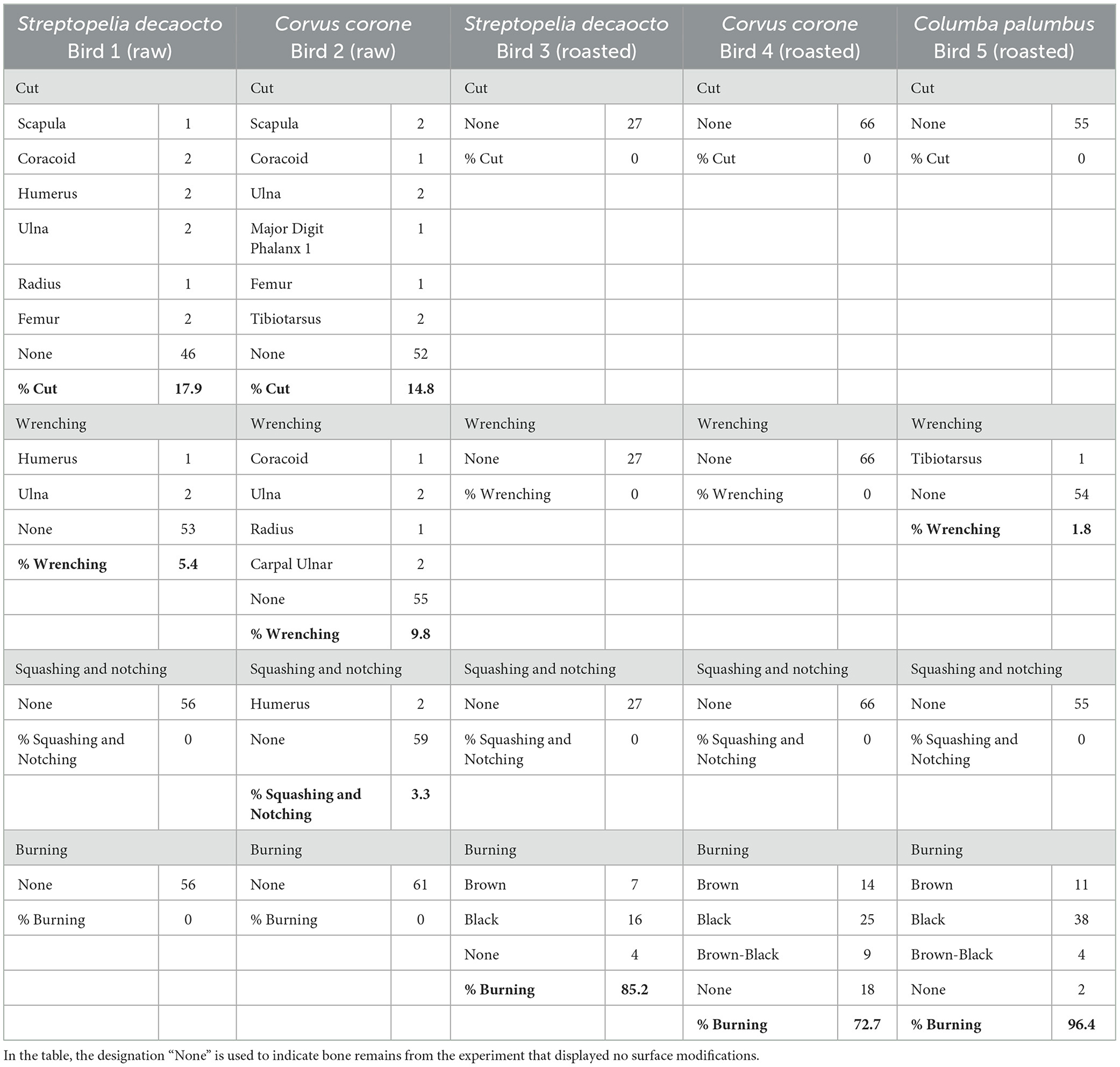
Table 2. Bone surface modifications of the five bird specimens following the experimental processing.
The legs were removed in a manner akin to the wings, using twisting motions and applying flexion and overextension to disarticulate the femur from the pelvis. On uncooked animals, tendons were cut using a flint flake, leaving cut marks on the proximal and mid-shaft of the femur (Figure 2A-N, P). Disarticulation from the tibiotarsus was achieved by flexing the femur against the joint's natural direction and through overextension, as well as with the aid of the flint flake resulting on marks on the proximal shaft of the tibiotarsus (Figure 2B–H, I). A similar technique was employed to separate the tibiotarsus from the tarsometatarsus without the necessity of using a flake. Flexion and overextension movements did not seem to cause any marks on the surface of leg bones, except for the wrenching identified on the roasted Columba palumbus' proximal tibiotarsus (Figure 1B, Table 2).
Through macroscopical observation, carrion crows were found to have minimal meat, in contrast to collared-doves, and wood pigeons in particular, which offered generous portions of breast filets and a significant amount of meat surrounding the femur. However, it is important to consider that the birds used were rescues and, as such, the meat yield from these animals might not accurately reflect that of birds in prime health. In roasted specimens, the meat fell away effortlessly, with no need for flint flakes. Conversely, processing raw specimens needed such tools for cutting the tendons, leaving cut marks on the long bones. Longitudinal cuts aimed at defleshing were made primarily on the humerus and femur, yet these did not imprint on bone surfaces. Therefore, only the transverse marks, indicative of tendon cutting or joint disarticulation, were observable.
A total of 265 bird bone remains, corresponding to complete and fragmented wing and leg elements, from five distinct individuals were collected following the experimental and cleaning procedures. Animals that underwent processing in their raw state retained the entirety of their skeletal representation, while those that were roasted exhibited a predisposition to bone fracture and loss (Table 2, Supplementary material 2). The roasted dove (Bird 3) presented a skeletal loss of 57.1%, corresponding to a total of 29 remains that disappeared during burning. Conversely, the roasted crow (Bird 4) had an increase in the number of remains, due to higher bone breakage (Figure 1, Supplementary material 2). Bones subjected to heat tend to fracture more readily even in the absence of burning marks. Observations indicate that the content of the inner cavity of the bone also combusts internally increasing its fragility. This phenomenon is particularly evident in the wing bones of the carrion crow and the wood pigeon (Figure 1A), where a dark longitudinal stain is commonly found inside the long bones.
Evidence of charring was found on nearly all the bird bones subjected to heat (Figure 1; Table 2). Exceptions included the two humeri of the Columba palumbus, the scapula, ulnar carpal, and femur of the Streptopelia decaocto, and eighteen skeletal parts of the Corvus corone, comprising the scapula, coracoid, humeri, ulnae, radii, both ulnar and radial carpals, the first phalanx of the minor digit, femora, and the second and third phalanges. Leg bones tend to be more frequently and intensively charred than wing bones. All thermal alterations are either brown or black burns. Long bones generally exhibit a brown discolouration from heat, with localised blackened areas, mainly on their epiphyses and along the shafts. While the majority of charred bones exhibit a single colouration from burning, instances of double-coloured charring have also been observed.
The experimentally replicated flake used in the processing of the two uncooked bird specimens, Streptopelia decaocto and Corvus corone, shows micro- and macroscopic recognisable use-wear typical of cutting up meat in the scope of butchering. Use-wear traces are tenuous, which is consistent to the working of a light butchery and over a short time. The set of use-wear features observed are similar to those mentioned by other authors on experimental pieces used in experimental butchery (Semenov, 1964; Keeley, 1980; Plisson, 1985; Gonzales Urquijo and Ibanez Estevez, 1994; Claud et al., 2019a; Costamagno et al., 2019).
Particularly noteworthy is the presence, on both sides, of the used edge of small isolated scars. These are “half-moon” shaped scars distributed along the edge and associated with a reticular shaped polish that can be observed at low and high magnifications. The edge removals result from the contact between the flake and the fibrous textures of avian muscle tissue that occasionally touched more resistant material, such as bone or cartilage (Figure 3).

Figure 3. Microscopic use-wear patterns on a flint flake following experimental processing. The image highlights the scars and polish distributed along the used edge of the experimental flint flake visible at low magnification using a digital microscope Hirox HR500E (a, b). Details of the same polish at high magnification (200x) using an Olympus BX60 optical microscope (c, d).
No striations nor any edge rounding is observed. Edge rounding is usually associated with skin processing rather than with meat cutting (Keeley, 1980; Gonzales Urquijo and Ibanez Estevez, 1994).
This pilot study reveals some initial patterns concerning the processing of bird remains that bear on archaeological interpretations of similar assemblages. We observed that when processing the carrion crow, it needed a longer processing time than the collared-dove, in particular when processed uncooked, which could be related to the larger size of this bird species. This aligns with the practical understanding of meat preparation, where cooked specimens provide easier meat access with, for instance, breast meat cutlets coming out effortlessly. However, larger birds, like the carrion crow, do not necessarily provide more substantial meat yields. The wood pigeon, noted for its ample meat quantity, especially around the breast filets and femur, supports the observations (1) that smaller birds can yield more edible meat than larger birds, and (2) that larger birds are not always sought after primarily for their meat content. This is consistent with findings from Gibraltar, where pigeons, and some corvids, were targeted for their meat (Blasco et al., 2014, 2016), while raptors were valued for their feathers (Finlayson et al., 2012). However, there are instances when raptors were also used for food (Gómez-Olivencia et al., 2018). Nonetheless, a growing body of Middle Palaeolithic research demonstrates that large birds were frequently sought for their feathers, and also for their phalanges (e.g., Peresani et al., 2011; Morin and Laroulandie, 2012; Radovcic et al., 2015; Rodríguez-Hidalgo et al., 2019), even if some use for food is also suggested (see references in Gómez-Olivencia et al., 2018). These practices have been further substantiated by various experimental studies (e.g., Pedergnana and Blasco, 2016; Romandini et al., 2016; Rufà et al., 2023).
In our experimentation, most bird specimens were processed by hand, without tools. The evidence of this manual processing is apparent in the form of twisting, crushing, and indenting marks, which were defined by Laroulandie et al. (2008) as wrenching, squashing and notching marks. These are evident on the distal humerus (Figure 2A–F, B–D), and on the ends of ulnas and radius (Figure 2A–I, L, B–E, G). Moreover, no tools were used to process the roasted specimens, other than cutting their chest line, indicating that cooking facilitated meat removal, thereby reducing/removing the need for their use. Additionally, the use of hammerstones was not required at any stage of our experimentation, which led to a total lack of percussion marks or breakage patterns associated with such actions (i.e., impact flakes or percussion notches).
Conversely, a lithic tool was necessary for processing raw birds. This is noted by the presence of cut marks on some uncooked appendicular skeleton bones. A flake was mainly used for cutting the tendons and to disarticulate bone joints. These actions are particularly visible by the transversal straight cuts that cluster on the scapulas' proximal and mid-shafts (Figure 2A–A, B–A, B), as well as on coracoids (Figure 2A–B–E, B–C). The same applies to the femurs and tibiotarsus from our uncooked sample, with predominance of oblique and transverse cuts in their shafts (Figure 2A–N–P, B–H–J). Although scapulas tend not to preserve well in archaeological contexts, the incisions found on coracoids, femurs and tibiotarsus agree with food provisioning actions, such as those found in Gorham's Cave (Blasco et al., 2016), for example.
The colouration of the burnt bones, exclusively brown and black, is consistent with the fact that these remains are associated with cooking activities (e.g., Nicholson, 1993). The absence of grey and white burns, which are characteristic of exposure to higher temperatures and longer burning times, supports the idea that the bird bones from the current experiment—which show brown and black burns—were subjected to controlled cooking rather than accidental or uncontrolled fire exposure. Notably, some bones, like the humerus, do not exhibit extensive burning marks, which is attributable to the thicker flesh surrounding them that may act as a barrier, insulating the bone from direct heat exposure. Conversely, bones with less surrounding meat, such as those in the limbs' extremities, display a greater propensity to burn, as evidenced by their charred appearance (as also noted by other studies, such as Cassoli and Tagliacozzo, 1997; Laroulandie, 2005; Blasco and Fernández Peris, 2009; Blasco et al., 2013, 2014).
The latter observation also highlights issues of skeletal loss. This is particularly evident in the roasted collared-dove (Bird 3), where the bird's extremities, especially the phalanges, became so fragile and brittle that they shattered into tiny, unrecoverable fragments. Similarly, higher bone breakage was noted in the roasted carrion crow (Bird 4), although this breakage did not lead to the complete disappearance of bones. Another observation made while handling the bones post-heat exposure was that some of the intact bones exhibited black staining on their inner surfaces. This occurred in instances where there appeared to be no direct contact between the bone's inner cavity and the heat source, since bones were not broken prior to heat exposure. Nonetheless, it seems that the inner structure of the bones, specifically the thin bar-like struts, had been burnt, rendering them more fragile and susceptible to breakage. These observations underscore the fact that bones from roasted birds tend to be underrepresented in archaeological assemblages. As burnt bird bones are prone to breakage and loss, roasting activities may therefore go undetected in archaeological sites.
The analysis of use-wear patterns on lithic artefacts provides invaluable insights into the technological behaviours and subsistence strategies of past human populations. Our use-wear results are consistent with the observations made by Pedergnana and Blasco (2016) during the defeathering of a Circaetus gallicus and a Gyps fulvus, since the use-wear observed on the flake that was used in our pilot experiment is under developed as well. This phenomenon may be attributed to the fact that we worked with less robust animals, which likely resulted in less damage to the flake used. Additionally, the flake shows few “half-moon” shaped scars distributed along the used edge, associated with polish.
All these observations not only have implications for the efficiency of food preparation but may also influence the archaeological visibility of butchery practices. Following the research line of this pilot study, future experimental approaches related to Neanderthal exploitation of birds are aimed to focus on expanding the variety of bird species examined in order to capture the range of avian life that early humans might have encountered and used. This would allow for a detailed comparison of processing times, meat yields, and bone preservation across species of different sizes and ecological roles. Nutritional analysis of the different bird species and preparation methods would also shed light on their contribution to Neanderthal diets. Additionally, amplifying the array of taphonomic studies, including human and other carnivores' bird bone chewing, as well as bird processing for the use of non-food elements (like feathers, tendons or some specific bones) could deepen our understanding of the role birds played within early human diets, but also beyond mere sustenance.
This pilot study has contributed to the growing body of experimental evidence providing insights into the replication of early human behaviour concerning the preparation and consumption of birds, focusing particularly on the implications of cooked vs. raw processing methods. Raw birds presented several cut marks, mainly associated with bone disarticulation and tendon cutting. The use of a flint flake for food processing was also detected by lithic use-wear observations. Conversely, cooked birds exhibited no cut marks and greater bone loss when compared to raw birds. This was due to heat exposure, which makes the disarticulation process and the removal of meat much easier and led to bone fragility and breakage. Such observations suggest that cooking methods significantly affect the preservation of skeletal remains in archaeological contexts, potentially influencing the archaeological visibility of certain cooking practices.
Our study contributes to the broader discourse on Neanderthal subsistence strategies by offering experimental data that can help interpret cut mark patterns and cooking-related bone alterations found in archaeological assemblages. By simulating Neanderthal cooking and butchering techniques, this research provides a baseline for distinguishing human-related modifications from those caused by natural processes or other predators. However, acknowledging the limitations of our experimental setup, including the small sample size and a single cooking technique, this study calls for further research incorporating a wider range of avian species and different cooking methods. Such expansions are essential to fully understand the range of Neanderthal dietary practices, their complex interactions with their environment, and their adaptive strategies and cultural development across different ecological changes.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because these were animals recovered dead from a rehabilitation centre as well described in the methods section of the paper.
MN: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AR: Conceptualization, Funding acquisition, Investigation, Methodology, Validation, Writing – review & editing. MI: Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Financial support has been provided by MN' 2-year postdoc contract for project “Neanderthal and Anatomically Modern Human interactions with small prey in Atlantic Iberia throughout the changing environments of the Pleistocene,” as part of the European Union's Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No. 101034349, and from the State Research Agency of the Spanish Ministry of Science and Innovation through the Program Maria de Maeztu Unit of Excellence (CEX2019-000945-M). Additional support has been given by UIDB/00698/2020 and UIDP/00698/2020, Fundação para a Ciência e Tecnologia (FCT), Portugal. AR is currently a beneficiary of a CEEC - 3rd Edition research contract promoted by the Portuguese FCT (reference: 2020.00877.CEECIND) and participates in the Spanish MICIIN projects PID2020-114462GB-I00, and the Generalitat de Catalunya projects CLT009/22/00044, CLT009/22/000045, and CLT009/22/00024. MN and AR also participate in the Spanish MICINN project PID2022-138590NB-C41.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2024.1411853/full#supplementary-material
Andrefsky, W. (2005). Lithics: Macroscopic Approaches to Analysis, Second edition Cambridge: Cambridge University Press. doi: 10.1017/CBO9780511810244
Blasco, R., Cochard, D., Colonese, A. C., Laroulandie, V., Meier, J., Morin, E., et al. (2022). “Chapter 8 – Small animal use by Neanderthals,” in Updating Neanderthals. Understanding Behavioural Complexity in the Late Middle Palaeolithic, eds. F. Romagnoli, F. Rivals, and S. Benazzi (New York: Elsevier Academic Press), 123–143. doi: 10.1016/B978-0-12-821428-2.00010-X
Blasco, R., and Fernández Peris, J. (2009). Middle Pleistocene bird consumption at Level XI of Bolomor Cave (Valencia, Spain). J. Archaeol. Sci. 36, 2213–2223. doi: 10.1016/j.jas.2009.06.006
Blasco, R., Finlayson, C., Rosell, J., Sánchez Marco, A., Finlayson, S., Finlayson, G., et al. (2014). The earliest pigeon fanciers. Sci. Rep. 4:5971. doi: 10.1038/srep05971
Blasco, R., Rosell, J., Fernández Peris, J., Arsuaga, J. L., Bermúdez, de Castro, J. M., and Carbonell, E. (2013). Environmental availability, behavioural diversity and diet: a zooarchaeological approach from the TD10-1 sublevel of Gran Dolina (Sierra de Atapuerca, Burgos, Spain) and Bolomor Cave (Valencia, Spain). Quat. Sci. Rev. 70, 124–144. doi: 10.1016/j.quascirev.2013.03.008
Blasco, R., Rosell, J., Rufà, A., Sánchez Marco, A., and Finlayson, C. (2016). Pigeons and choughs, a usual resource for the Neanderthals in Gibraltar. Quater. Int. 421, 62–77. doi: 10.1016/j.quaint.2015.10.040
Cassoli, P. F., and Tagliacozzo, A. (1997). Butchering and cooking of birds in the palaeolithic site of Grotta Romanelli (Italy). Int. J. Osteoarchaeol. 7, 303–320. doi: 10.1002/(SICI)1099-1212(199707/08)7:4<303::AID-OA380>3.0.CO;2-R
Claud, E., Thiébaut, C., Costamagno, S., Deschamps, M., Soulier, M. C., Brenet, M., et al. (2019b). The practices used by the Neanderthals in the aquisition and exploitation of plant and animal resources and the function of the sites studied: summary and discussion. Palethnologie 10:4179. doi: 10.4000/palethnologie.4179
Claud, E., Thiébaut, C., Coudenneau, A., Deschamps, M., Mourre, V., Brenet, M., et al. (2019a). Le référentiel des outils lithiques. Palethnologie 10:5142. doi: 10.4000/palethnologie.5142
Costamagno, S., Claud, E., Thiébaut, C., Chacón-Navarro, M., and Soulier, M. C. (2019). L'exploitation des ressources végétales et animales au Paléolithique: quels outils méthodologiques pour quelles questions? Palethnologie 10:3866. doi: 10.4000/palethnologie.3866
Ericson, G. P. (1987). Interpretations of archaeological bird remains: a taphponomic approach. J. Archaeol. Sci.14, 64–75. doi: 10.1016/S0305-4403(87)80006-7
Finlayson, C., Brown, K., Blasco, R., Roseli, J. N. J, Bortolotti, G., et al. (2012). Birds of a feather: neanderthal exploitation of raptors and corvids. PLoS ONE 7, 1–9. doi: 10.1371/annotation/5160ffc6-ec2d-49e6-a05b-25b41391c3d1
Gaudzinski-Windheuser, S., Kindler, L., Macdonald, K., and Roebroeks, W. (2023). Hunting and processing of straight-tusked elephants 125.000 years ago: implications for Neanderthal behavior. Sci. Adv. 9:eadd8186. doi: 10.1126/sciadv.add8186
Gómez-Olivencia, A., Sala, N., Núñez-Lahuerta, C., Sanchis, A., Arlegi, M., Rios-Garaizar, J., et al. (2018). First data of Neandertal bird and carnivore exploitation in the Cantabrian Region (Axlor; Barandiaran excavations; Dima, Biscay, Northern Iberian Peninsula). Sci. Rep. 8, 1–14. doi: 10.1038/s41598-018-28377-y
Gonzales Urquijo, J. E., and Ibanez Estevez, J. J. (1994). Metodologia de analisis functional de instrumentos tallados en silex. Bilbao: Universidad de Deusto, 301.
Kamminga, J. (1979). “The nature of use polish and abrasive smoothing on stone tools,” in Lithic Use-wear Analysis, ed. B. Hayden (New York: Academic Press), 143–158.
Keeley, L. H. (1980). Experimental Determination of Stone Tool Uses. A Microwear Analysis. Chicago: University of Chicago Press.
Laroulandie, V. (2001). ≪ Les traces liées à la boucherie, à la cuisson et à la consummation d'oiseaux: apport de l'expérimentation,” in Préhistoire et approche expérimentale. Collection préhistoire 5, eds. L. Bourguignon, I. Ortega, M.-C. Frère-Sautot (Montagnac : Monique Mergoual), 97–108.
Laroulandie, V. (2005). “Bird exploitation pattern: the case of Ptarningan Lagopus sp. in the Upper Magdalenian site of la Vache (Ardiège, France),” in Feathers, Grid and Symbolism Birds and Humans in the Ancient Old and New Worlds, eds. G. Grupe, J. Peters (Munich: Proceedings of the 5th. Meeting of the ICAZ), 165–178.
Laroulandie, V., Costamagno, S., Cochard, D., Mallye, J. B., Beauval, C., Castel, J. C., et al. (2008). When disarticulation causes bone damage. The case of the hyperextension of the elbow. Ann. Paléontol. 94, 287–302. doi: 10.1016/j.annpal.2008.09.002
Martínez Valle, R., Guillem Calatayud, P. M., and Villaverde Bonilla, V. (2016). Bird consumption in the final stage of Cova Negra (Xátiva, Valencia). Quater. Int. 421, 85–102. doi: 10.1016/j.quaint.2016.01.068
McPherron, S., Braun, D., DogandŽi,ć, T., Archer, W., and Desta, D. (2014). An experimental assessment of the influences on edge damage to lithic artifacts: a consideration of edge angle, substrate grain size, raw material properties, and exposed face. J. Archaeol. Sci. 49, 70–82. doi: 10.1016/j.jas.2014.04.003
Morin, E., and Laroulandie, V. (2012). Presumed symbolic use of diurnal raptors by Neanderthals. PLoS ONE 7:e32856. doi: 10.1371/journal.pone.0032856
Mourer-Chauviré, C. (1983). “Les oiseaux dans les habitats paléolithiques: gibier des hommes ou proies des rapaces?” in Animals and Archaeology: 2. Shell Middens, Fishes and Birds, eds. C. Grigson, J. Clutton-Brock (Oxford: BAR International Series 183), 111–124.
Nabais, M., Pimenta, C., and Zilhão, J. (2023). Human-bird interaction in last Interglacial Iberia: a combined approach using skeletal part analysis, bone surface modification, bird ethology and ethnography. J. Archaeol. Sci.49:104023. doi: 10.1016/j.jasrep.2023.104023
Nicholson, R. (1993). A morphological investigation of burnt animal bone and an evaluation of its utility in archaeology. J. Archaeol. Sci. 20, 411–428. doi: 10.1006/jasc.1993.1025
Nicolaysen, R. (1980). Arctic nutrition. Perspect. Biol. Med. 1980, 295–310. doi: 10.1353/pbm.1980.0061
Osgood, C. (1971). The Han Indians: A Compilation of Ethnographic and Historical Data on the Alaska-Yukon Boundary Area. Yale University Publications in Anthropology, Number 74. New Haven: Department of Anthropology Yale University.
Pedergnana, A., and Blasco, R. (2016). Characterising the exploitation of avian resources: An experimental combination of lithic use-wear, residue and taphonomic analyses. Quater. Int. 421, 255–269. doi: 10.1016/j.quaint.2015.07.025
Peresani, M., Fiore, I., Gala, M., Romandini, M., and Tagliacozzo, A. (2011). Late Neandertals and the intentional removal of feathers as evidenced from bird bone taphonomy at Fumane Cave 44 ky B.P., Italy. Proc. Nat. Acad. Sci. 108, 3888–3893. doi: 10.1073/pnas.1016212108
Plisson, H. (1985). Etude fonctionnelle d'outillages lithiques prehistoriques par l'analyse des micro-usures: recherche méthodologique et archéologique. Uni-versité Paris I, thèse doctorat, sciences humaines, 357.
Radovcic, D., Oros Srsen, A., Radovcic, J., and Frayer, D. W. (2015). Evidence for Neandertal Jewelry: Modified White-Tailed Eagle Claws at Krapina. PLoS ONE 10:e0119802. doi: 10.1371/journal.pone.0119802
Rodríguez-Hidalgo, A., Morales, J. I., Cebrià, A., Courtenay, L. A., Fernández-Marchena, J. L., García-Agudo, G., et al. (2019). The Châtelperronian Neanderthals of Cova Foradada (Calafell, Spain) used imperial eagle phalanges for symbolic purposes. Sci. Adv. 5:eaax1984. doi: 10.1126/sciadv.aax1984
Romandini, M., Fiore, I., Gala, M., Cestari, M., Guida, G., Tagliacozzo, A., et al. (2016). Neanderthal scraping and manual handling of raptor swing bonés: Evidence from Fumane Cave. Experimental activities and comparison. Quater. Int. 421, 154–172. doi: 10.1016/j.quaint.2015.12.078
Romero, A. J., Díez, J. C., and Brugal, J. P. (2017). Aves de caza. Estudio tafonómico y zooarqueológico de los restos avianos de los niveles musterienses de Pié Lombard (Alps-Maritimes, Francia). Munibe 68, 73–84. doi: 10.21630/maa.2017.68.14
Rufà, A., Blasco, R., Roger, T., Rué, M., and Daujeard, C. (2018). A rallying point for different predators: the avian record from a Late Pleistocene sequence of Grotte des Barasses II (Balazuc, Ardèche, France). Archaeol. Anthropol. Sci. 10, 1459–1476. doi: 10.1007/s12520-017-0469-6
Rufà, A., and Laroulandie, V. (2021). Unravelling the taphonomic stories of bird bones from the middle pleistocene layer VIII of Grotte Vaufrey, France. Quaternary 4:30. doi: 10.3390/quat4040030
Rufà, A., Martin, C., and Laroulandie, V. (2023). “Searching for raptor feathers and talons: Results of an experimental approach,” in Hiatus, lacunes et absences: identifier et interpreter les vides archéologiques, Actes du 29e Congrès préhistorique de France, 31 mai – 4 juin 2021, Toulouse, 25–40.
Semenov, S. A. (1964). Prehistoric Technology: An Experimental Study of the Oldest Tools and Artefacts from Traces of Manufacture and Wear. London: Cory, Adams and Mackay.
Simonsen, K. P., Rasmussen, A. R., Mathisen, P., Petersen, H., and Borup, F. (2011). A fast preparation of skeletal materials using enzyme maceration. J. Foren. Sci. 56, 480–484. doi: 10.1111/j.1556-4029.2010.01668.x
Smith, G. (2015). Neanderthal megafaunal exploitation in Western Europe and its dietary implications: a contextual reassessment of La Cotte de St Brelade (Jersey). J. Hum. Evol. 78, 181–201. doi: 10.1016/j.jhevol.2014.10.007
Tringham, R., Cooper, G., Odell, G., and Voytek, B. (1974). Experimentation in the formation of edge damage: a new approach to lithic analysis. J. Field Archeol. 1, 171–196. doi: 10.1179/jfa.1974.1.1-2.171
Keywords: experimental archaeology, taphonomy, use-wear, cooked vs. raw meat processing, avifauna
Citation: Nabais M, Rufà A and Igreja M (2024) Experimental replication of early human behaviour in bird preparation: a pilot-study focusing on bone surface modification and breakage patterns. Front. Environ. Archaeol. 3:1411853. doi: 10.3389/fearc.2024.1411853
Received: 03 April 2024; Accepted: 30 May 2024;
Published: 24 July 2024.
Edited by:
Camille Daujeard, UMR7194 Histoire naturelle de l'Homme préhistorique (HNHP), FranceReviewed by:
Aurélia Borvon, UMR 7041 Archeologies and Antiquity Sciences (ArScAn), FranceCopyright © 2024 Nabais, Rufà and Igreja. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariana Nabais, bWFyaWFuYW5hYmFpc0BnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.