 Camila Alday
Camila Alday Maria Angélica García2
Maria Angélica García2 Carmela Alarcón
Carmela Alarcón
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Environ. Archaeol. , 07 November 2023
Sec. Archeobotany
Volume 2 - 2023 | https://doi.org/10.3389/fearc.2023.1251137
Bast fiber artifacts from the Pacific coast of South America are among the earliest evidence of fabric manipulation and the foundation of the millennial Andean textile tradition. Recent examinations of plant macro remains and plant fiber artifacts from La Yerba II (7570–6674 cal BP) and La Yerba III (6485–5893 cal BP) sites provide insight into how plant fabric technologies and pre-loom textile traditions unfolded over the Middle Preceramic (or Middle Archaic; 8000–5000 BP) on the south coast of Peru. Using light microscopy and scanning electron microscopy (SEM), we conduct comparative analyses of modern and archaeological plants to assess the taxonomy and morphological distinctions between them to discriminate species. We identify materials from wild-gathered plants in the Cyperaceae, Typhaceae, and Apocynaceae families and show how they were used to make fishing nets, looped bags, mats, and other artifacts. Evidence from tools and cut marks also enabled us to reconstruct parts of the chaîne opératoire of the fiber production. We discuss the significance of plants from wetland and estuarine habitats in the interrelated dimensions of fabric manufacture and food consumption among these coastal hunter-gatherers.
Plants have been used for making fabrics for thousands of years (Nadel et al., 1994; Hardy, 2007; Hurcombe, 2007; Kvavadze et al., 2009; Hardy et al., 2020), and many species have been gathered and eventually cultivated for this purpose (Barber, 1991; Gleba and Harris, 2019; Rast-Eicher et al., 2021). Direct and indirect evidence of bast fiber artifacts suggests that the development of these sophisticated technologies was critical to different dimensions of past lifestyles. This was the case for Middle Preceramic hunter-gatherers of the south coast of Peru.
The arid Pacific coast of South America has exceptional preservation environments for organic remains and, therefore, offers unique opportunities to study changes in plant textiles. Macro remains of plant and bast fibers, residues of fiber production, and material culture have been recovered from very early coastal archaeological contexts here (Sandweiss, 2003; Dillehay, 2017: Beresford-Jones et al., 2018; Martens and Cameron, 2019; Alday, 2022; Santos and Standen, 2022). The use of wild-gathered plants from the Cyperaceae, Typhaceae, and Apocynaceae families for fibers along the Pacific coast seems to date back to the earliest human occupation during the Terminal Pleistocene (e.g., Jolie et al., 2011; Connolly et al., 2016; Gilligan, 2018; Adovasio and Dillehay, 2020) until today. The production of these plant fibers allowed for the manufacturing of a range of technologies key to the marine-focused economy of the Preceramic (or Archaic) Period, including bags and containers, nets, twined mats, fishing lines, and string for hafting. Most importantly, the wild-gathered plant technologies that underpinned early economies before 6000 BP evolved into later cotton cultivation and the production, spinning and weaving techniques, and technological forms that this new raw material required. The use of plant fibers defined an enduring tradition that developed during the Early Preceramic Period, if not before (Alday, 2023). For more than seven millennia, coastal hunter-gatherers used wild plants gathered from estuaries and wetlands, riparian oases, and, perhaps, lomas habitats to produce twined mats, nets, looped bags, skirts, ropes, and cordage (Engel, 1963; Standen, 2003; Martens and Cameron, 2019; Alday, 2022; Santos and Standen, 2022; Montt et al., 2023). Fiber industries likely constituted the necessary socio-economic and ecological contexts for the eventual shift from wild fiber plants to cotton fibers (Gossypium barbadense) as a supply for producing large nets and complex fabrics.
In turn, cotton fibers intensified marine subsistence by enabling the production and maintenance of larger and more effective seine fishing nets (Moseley, 1975; Quilter and Stocker, 1983; Moseley and Feldman, 1988; Beresford-Jones et al., 2018). Scholars have long noted the significance of cotton in the fiber technologies used to harvest the coast's rich marine resources in the emergence of complex societies in Peru during the Late Preceramic (Moseley and Feldman, 1988). Indeed, Moseley (1975) “Maritime Foundation of Andean Civilization” (MFAC) hypothesis proposed that Andean complex societies arose from the exploitation of marine resources, in which cotton nets played a crucial role in the maritime technology that facilitated the exploitation of small-schooling fish, such as anchovies (Moseley, 1992; Sandweiss, 2003). After ~5000 BP, when the intensified use of cotton became widespread along the coast of Peru, different skills compared to those used in bast fiber industries became necessary, presumably leading to the formation of more specialized labor and a call for innovations in the chaîne opératoire of fishing net, clothing, and fabric production, such as the shift from splicing long bast fibers by hand (cf. Gleba and Harris, 2019) to spinning short cotton fibers using spindle whorls (Bird et al., 1985; Splitstoser et al., 2016; Dillehay, 2017).
Bast fiber is defined, technically, as the use of “parts of the plants underneath the phloem surrounding the stem itself” (Gleba and Harris, 2019, p. 2329). The fibers need to be extracted and processed before being spun and worked into an artifact, thus calling for a specialized chaîne opératoire. While these have been well understood for important Old World bast fiber plants, such as flax, hemp, and jute, the tools and processes of procurement and the stages of production of plant fibers have been far less studied along the Pacific coast of South America (see Martens and Cameron, 2019; Santos and Standen, 2022, for northern Chile; see Alday, 2022, for southern Peru). Despite their critical significance to the exploitation of rich marine resources by the hunter-gatherer societies of the Preceramic and the excellent condition for the preservation of organic residues in our area of study, little attention has been paid to plant and bast fibers as critical raw materials.
In this article, we present findings from the examination and identification of plant remains, plant fiber artifacts, and techniques of production from two Middle Preceramic archaeological sites: La Yerba II and La Yerba III on the Río Ica estuary on the south coast of Peru (Figure 1). Using light microscopy and scanning electron microscopy (SEM), we conduct comparative analyses of modern and archaeological plants to assess the taxonomy and morphological distinctions between them so as to attempt to discriminate species. Following this, we attempt to characterize nets, fishing lines, and fabrics using digital microscopy for structural textile analysis and SEM for fiber determination. Finally, we use these data to emphasize the potential of plant fiber and bast fiber studies to begin to map out a landscape of Preceramic fiber production and use these to understand the seasonality of desert coast ecologies and, thereby, their hunter-gatherer ways of life. In this regard, ethnographic and ethnohistorical accounts of similar environments and utilization of bast fiber provide useful comparative data and some insight into the dynamic and multiple-level interactions between communities and local ecosystems.
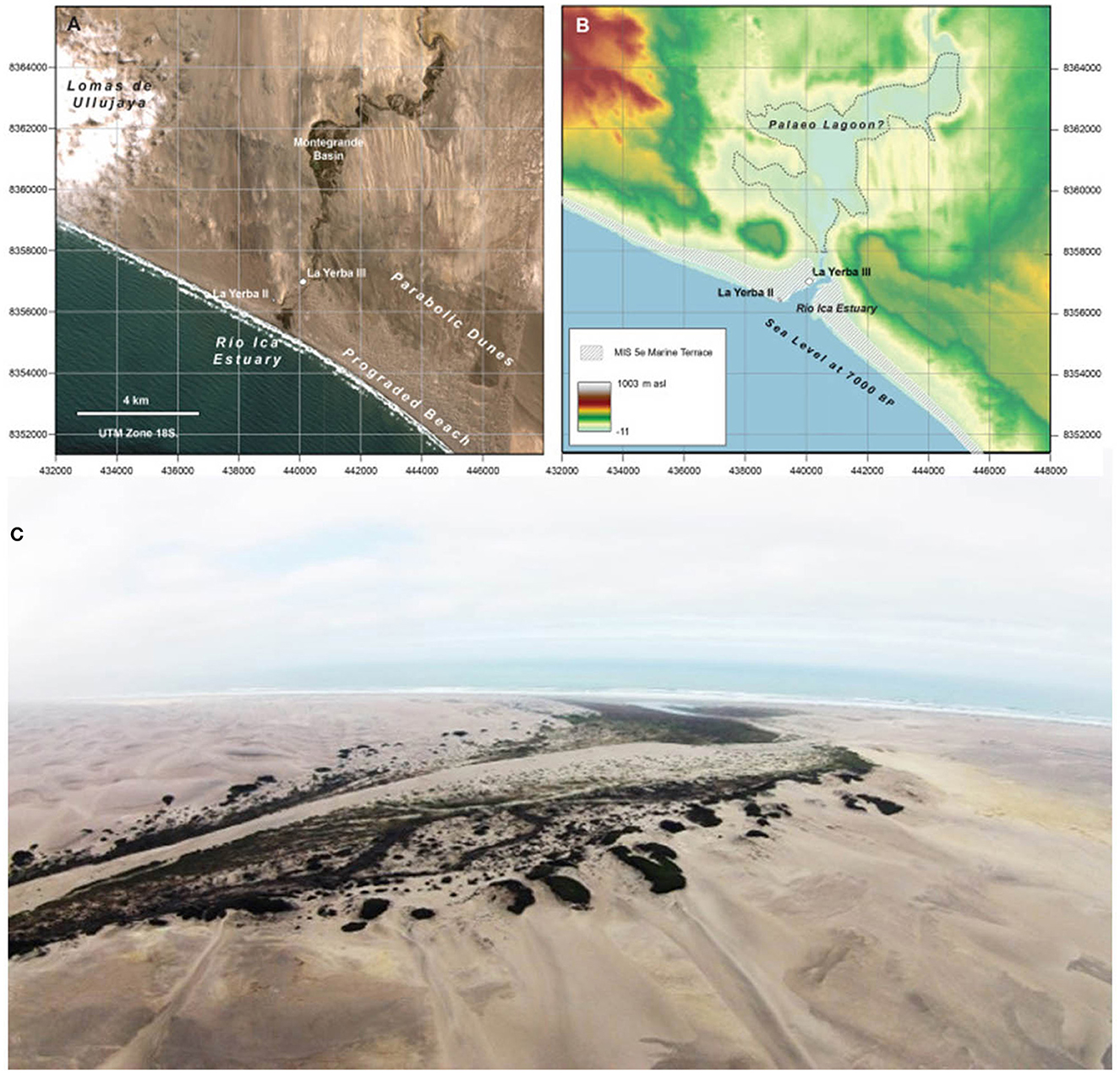
Figure 1. (A, B) Paleo-lagoon and location of La Yerba II and La Yerba III (Source: Beresford-Jones et al., 2022, Figure 12); (C) wetland and estuary habitats, La Yerba II and La Yerba III.
The desert regions of the coast of Peru have a unique and rich history of fabric technology and textiles that is intertwined with ecological change and adaptation. In the context of the south coast, the sites of La Yerba II and La Yerba III on the Río Ica estuary host exceptional evidence of plant fiber material culture not only for their excellent preservation but also for their contextual information in well-defined chronostratigraphies and midden and material culture analyses. Middle Preceramic archaeological sites are relatively few compared to those of later south coast periods, limiting the opportunities to study fiber production. The records of these two sites, however, enable the study of how these plant fiber technologies unfolded over some 1,500 years of occupation.
The La Yerba II shell midden mound is the outcome of an intense rhythm of repeated occupations by logistically mobile marine hunter-gatherers over some five centuries (7100–6600 cal BP), exploiting mainly marine resources such molluscs (especially Mesodesma donacium surf clams), fish, crustaceans, marine mammals, and kelp. While resources hunted and gathered from beach and rocky littoral habitats dominate the La Yerba II remains, its middens also contain terrestrial resources, such as remains of guanaco and deer, land snails (Bostyrx sp.), and many plants, including, in particular, bottle gourd (Lagenaria sp.) and charred and desiccated Cyperaceae esculentus rhizomes, which were frequent in certain contexts (Beresford-Jones et al., 2018, 2022). Fragments of yarn, looped bags, balls of thread and fiber, twined mats, and unspliced fibers have been recovered from dense midden and occupation surfaces. No true knotted net fragments were recovered from La Yerba II contexts and, indeed, its fish remains are comprised of 60% [by Minimum number of individuals (MNI)] various species of presumably line- or harpoon-caught drum (family Sciaenidae) and 25% mullet (Mugil cephalus), likely estuarine-trapped during summer spawning, with almost no small-schooling species (Beresford-Jones et al., 2018). These contexts sometimes also contained modified Choromytilus shells used as scraper tools (see Figure 9).
Shortly after, between 6392 and 6033 cal BP, the very different site of La Yerba III was occupied alongside the estuary ~1 km upstream, composed of semi-subterranean dwellings set amidst dense middens. Its population was an order of magnitude greater than La Yerba II, this village contains evidence suggesting permanent or semi-permanent settlement, including storage pits, structured burials beneath the floors, more substantial architecture, heavy grinding stones, and whalebone anvils. The site contains substantial amounts of obsidian lithic materials sourced to Quispisisa, 160 km away in the high Andes, at 4100 m asl (Beresford-Jones et al., 2023). At La Yerba III, multiple knotted net fragments were recovered, and fish remains included significant proportions (40% by MNI) of small-schooling, presumably net-caught species such as anchovies (Beresford-Jones et al., 2018). Bone isotope analyses show that the diets of La Yerba III inhabitants were, like those of their predecessors at La Yerba II, dominated by marine resources evident in its middens, but those middens also contain copious gathered and hunted terrestrial resources, including plants from wetlands and lomas fog oases, and crucially, early evidence of cultivated food plants, including lima beans (Phaseolus lunatus directly dated to 6275–5999 cal BP), jack beans (Canavalia ensiformis), guava (Psidium guajava), and animals such as guinea pigs (Cavia porcellus) and dogs (Canis familiaris; Beresford-Jones et al., 2021).
Both La Yerba II and III are located at the intersection of very different, highly productive habitats. Estuaries and wetlands, as the nexus of marine and https://www.sciencedirect.com/topics/earth-and-planetary-sciences/freshwater-ecosystem freshwater ecosystems, enjoy high levels of nutrient influx from each (Day et al., 2012, Figure 1). Present-day estuaries and wetlands formed after the stabilization of eustatic sea levels at ~5800 BP, but there is geomorphological evidence that subsequent, ongoing tectonic uplift in this area has greatly diminished the original extent of wetland habitats from those extant when those sites were occupied six millennia ago (Beresford-Jones et al., 2022, Figure 12). Alongside beach habitats critical for almost inexhaustible high-protein Mesodesma surf clam resources, this extended estuary and palaeo-lagoon environment enabled increased sedentism and increasing populations by providing other plentiful easily gathered resources and aquatic plants for critical fiber resources.
Desiccated and well-preserved plant macros and fiber artifacts (Figure 2) were extracted from the excavated contexts by dry sieving and flotation to produce 2-mm and 0.5-mm fractions and stored at the Museo Regional de Ica Peru (Supplementary material 2). Reference materials used for botanic identification of plant fiber and bast fibers were obtained in fieldwork during 2018–2019, along with plant materials from the Economic Botany Collection at the National Museum of Natural History in Chile.

Figure 2. (A) La Yerba II Trench 1 SU 1017 twined mat two raw materials (vertical element cf. Scirpus and horizontal element: cf. Asclepias), (B) Typha scf raw material; (C) La Yerba III, Trench 1 SU 9506 closed twined mat (cf. Asclepias); (D) La Yerba III Trench 3 SU 7003 looped structure bag (cf. Scirpus); (E) La Yerba III Trench 1 SU 9505 fragment of yarn for hafting; (F) La Yerba III Trench 2 SU 9756 net (cf. Asclepias); (G) La Yerba II Trench 2 looped structure (cf. Scirpus); (H) La Yerba II Trench 1 SU 1010 spliced yarns.
The particular challenge in the plant identification of bast fibers is that many diagnostic features, such as nodes and cross-markings, are shared across most bast fibers, making it difficult to discriminate taxonomy at least to plant species (Bergfjord and Holst, 2010; Patterson et al., 2017). Supplementary material 1 notes consequent uncertainties in our identifications. Moreover, besides variations in fiber diameter, lumen, or plant epidermis characteristics, many features may be lost from bast fibers since they have been modified or altered through preparation processes such as decortication and retting. We adapted Gleba and Harris (2019) methods for the materials and samples in this study, allowing us to suggest new categories of analysis for the structure of fibers and plants (Figure 3).
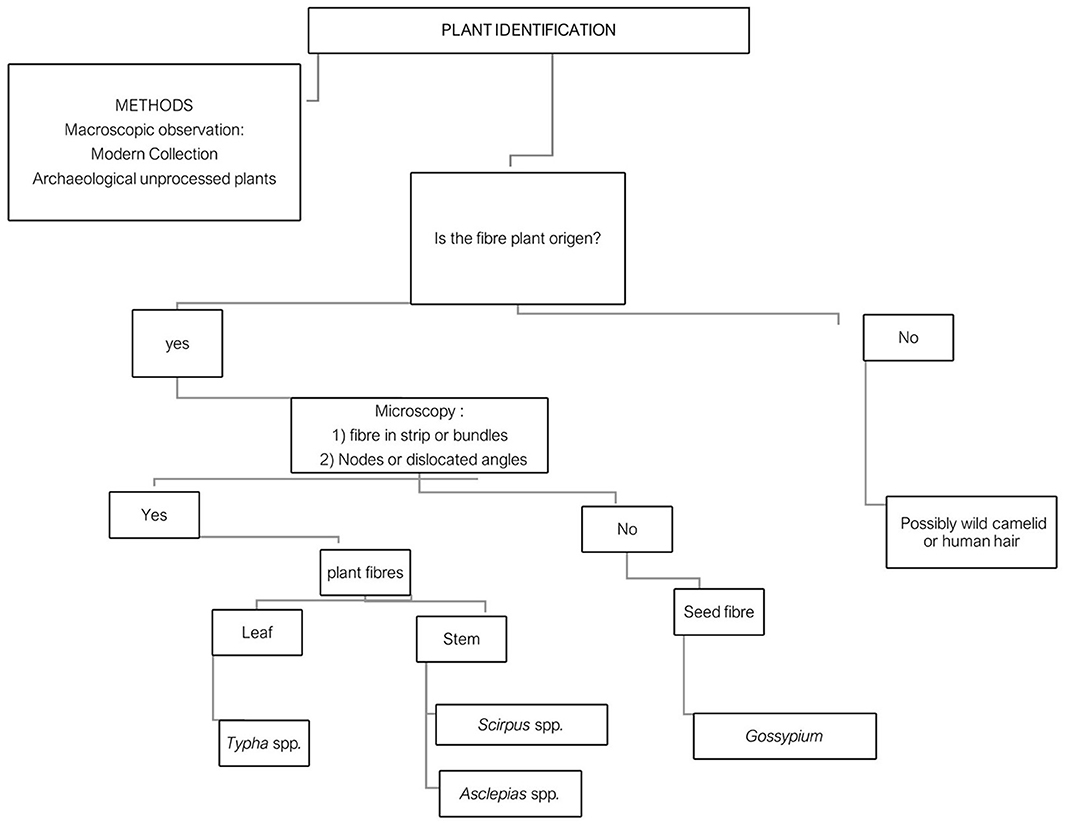
Figure 3. A flowchart that explains the process of identification of bast fiber remains from archaeological materials (based on Gleba and Harris, 2019, p. 2340, Figure 13).
The terminology used in our work as to plant fiber technologies refers to the use of different parts of the plants that have been worked into fibers for textiles, fabrics, basketry, and cordages (Dodge, 1897; Emery, 1966; Adovasio, 1977; Florian et al., 1991). In our analyses, we draw the distinction between bast fiber from cf. Asclepias and plant fibers from cf. Typha and cf. Scirpus.
We conducted microscopic observation of archaeological and modern reference materials, and we registered the presence and/or absence of particular stomata cells and nodes (dislocations) and cross-marking to attempt to identify plants at least at the family level (Sadrmanesh and Chen, 2019). Samples were mounted and viewed through both transmission light and reflected via a light microscope (LM) at magnifications ranging from 10 × to 50 × . Samples were also analyzed using Hitachi TM3000 TableTop SEM at the McDonald Institute for Archaeological Research, University of Cambridge. The following instrumental settings were used: analytical condition mode at 15.00 kV accelerating voltage, compositional imaging and working distance of 5–10 mm.
Observations revealed varying characteristics of the plant fibers of different species (see Figure 6). Cross-markings were distinctly visible on bast fibers of Asclepias sp. when observed through an LM. By contrast, nodes were prominently discernible on Scirpus spp. when examined via SEM, whereas both cross-markings and nodes on Typha spp. were relatively faint. Whether such variations in nodes and cross-marking visibilities arise from different processing techniques, usage-induced degradation of plant structures or aging will require further investigation.
The origin of nodes or dislocations which are found typically in close proximity to the bast fiber surface is uncertain and still debated (Bergfjord et al., 2010; Suomela et al., 2018). Sometimes referred to in the literature as kink bands, some argue that the quality of these nodes, influenced by fiber processing, offers an avenue to explore the correlation between different techniques of fiber processing and plant characteristics (Shah and Bourmad, personal communication 2023). Kink bands, for instance, observed in flax from Egyptian textiles (Melelli et al., 2021) are not inherent features of the fiber but rather a result of processing. These seem to arise from mechanical stresses during fiber extraction and processing, possibly related to retting. Such stresses may also arise from the rigorous decortication, scutching, and thread-making processes reported for flax bast fibers (Melelli et al., 2021). Some nodes in our samples have a resemblance to kink bands seen in flax (Shah and Bourmad, personal communication 2023), although their systematic analysis to identify specific plant processing stages is still pending. We hypothesize, however, that, like flax bast fibers, features of South American bast fiber node morphology, such as those noted in Asclepias, might be linked to plant-processing steps like extraction, decortication, and subtle retting.
These features are rarely sufficient to distinguish fiber plants at higher taxonomic levels on their own (Lukesova and Holst, 2021), so we combined these initial analyses with the identification of reproductive organs such as inflorescence, seeds, and other macro remains to enhance botanical identification of, for instance, Asclepias spp., Scirpus spp., and Typha sp. (Supplementary material 2, Figures 4, 5). Additionally, the analysis of almost-complete pieces of Typha spp. and Scirpus spp. woven into some artifacts with minimal processing (Figure 7) assisted the identification.

Figure 4. (A) La Yerba III, Trench 1 SU 9506 cf. Typha); (B) cf. Typha details of the longitudinal section of the vascular bundles and fiber cables (magnification 100× DinoLite); (C) T. dominguensis reference collections (local wetland, recollected in 2018); (D) Cyperaceae stems; (E) Scirpus spp. inflorescence on fragment of mat (cf. Typha) La Yerba III Trench 1 SU 9506 (magnification 100× DinoLite); (F) S. americanus reference collection (local wetland, recollected in 2018).

Figure 5. (A) Asclepias reference collection; (B) cf. Asclepias Apocynaceae bast fibers La Yerba II (magnification 50× DinoLite); (C, D) cross-markings and nodes on cf. Asclepias bast fibers (La Yerba II Trench 1 SU 1010).
Nets, fishing lines, and fabrics were characterized using digital microscopy for structural textile analysis and SEM for fiber determination. We first observed how plant fiber and bast fiber bundles were worked into threads, for example, dislocations or nodes, which are exclusive to bast fibers remain adjacent to each other when plants have been processed manually. Then, the quality of the nodes as a result of fiber processing allowed us to interrogate the relationship between processing techniques (e.g., decortication or retting) and raw materials. Furthermore, here, we also focused on finding evidence relative to thread-making techniques, degrees of fiber processing, and any other new information that could help us reconstruct stages of the technological process.
In the following, we set out the detailed identification of plant fiber artifacts recovered from La Yerba II and III contexts. Table 1 summarizes those findings alongside the identification of other plant macros from each context (see Supplementary material 1, 2).
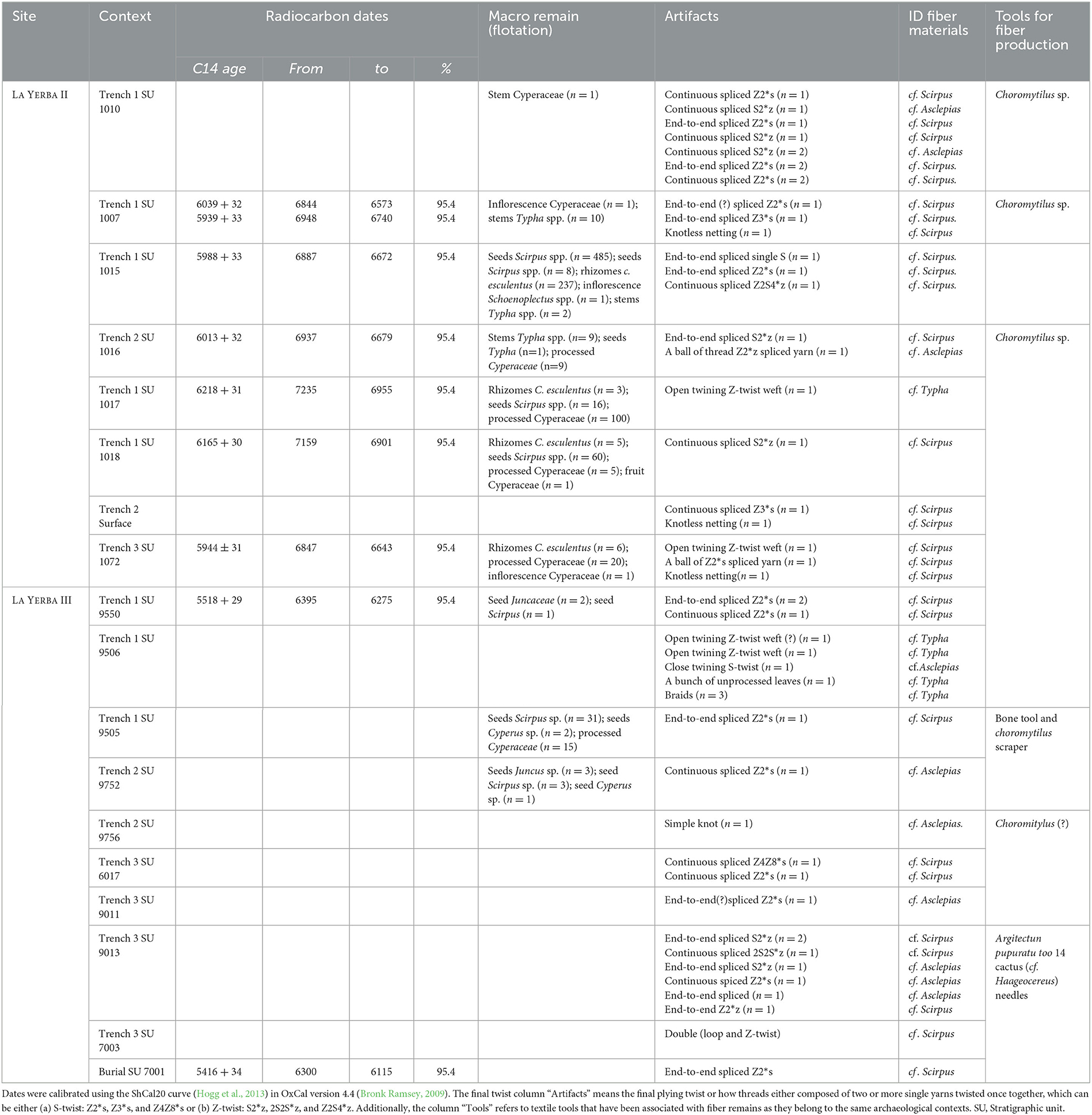
Table 1. Macro and plant fiber materials analyzed for this study and their distribution.
Plant macro remains of cf. Scirpus and cf. Typha aquatic plants from estuaries and wetlands; plant fiber artifacts, including some showing evidence of processing, such as balls of cf. Scirpus fiber, cf. Asclepias stalks (Figures 5, 6) and processed fibers of cf. Scirpus and cf. Typha (Figures 2, 8); and diverse tools for fabric manipulation (Figure 7) offer clear evidence for the significance of plant fiber processing at both La Yerba II and La Yerba III. Scirpus spp. inflorescences, charred rhizomes of Cyperaceae esculentus, and leaves and capsules of Typha spp. also offer further evidence of plant processing and consumption as food at both sites. The presence of Scirpus inflorescences and Typha capsules suggests these plants were likely harvested in summer. Few cf. Typha artifacts (n = 5) were identified in the contexts of both sites. Yet cf. Typha plant capsules (reproductive organs; see Figure 7) were recovered from contexts at La Yerba II (Trench 2 SU 1016), which might suggest the use of Typha pollen as a food source as noted among North American peoples (Morton, 1975; Saunders, 1978 in Linskens and Jorde, 1997), at Chaco Canyon (Arenas and Scarpa, 2003), and elsewhere worldwide (Zhang et al., 2020).

Figure 6. (A, B) Knot from a bast fiber fishing net (cf. Asclepias) La Yerba III, Trench 2 UE 9756; (B) detail of the bast fiber; (C) parenchyma reference collection for Typha (T. Dominguesis); (D) trachery cf. Scirpus; (E) stomata in parenchyma tissues cf. Scirpus.

Figure 7. Wetlands plants from La Yerba II and La Yerba III: (A) capsules of cf. Typha; (B) Cyperaceae stem; (C) La Yerba II Trench 1 SU 1015 charred C. esculentus (rhizome); (D, E) grinding stones for food processing (La Yerba II Trench 1 SU 1001; La Yerba III Trench 1 SU 9542); (F) La Yerba II Trench 3 SU 1050 Cyperaceae inflorescence; (G) La Yerba II Trench 3 1076 Schoenoplectus (likely S. americanus).
A diverse pattern of Cyperaceae plant parts, including seeds, inflorescences, stems, and rhizomes is documented in the archaeological record of La Yerba II and La Yerba III (over 800 specimens). This suggests that the Cyperaceae plants were transported to these sites, for their processing for the purposes of food and fiber technology. C. esculentus rhizomes, seeds of Scirpus spp., and inflorescences of Schoenoplectus spp., collectively, represent diverse activities taking place at La Yerba II and La Yerba III. Those activities likely involved peeling and cleaning rhizomes, preparing the stems and extracting bast fibers, and preparing starchy rhizomes for eating. Cyperaceae and Typhaceae plants were, we suggest, harvested by families for food consumption and fiber production simultaneously during the peak summer growing season.
Despite being identified in the fibers of nets, cordages, and twined fabric (n = 10), other plant parts of Asclepias spp. were not identified, aside from one stalk evidencing cut marks (Figure 5). This might suggest that the extraction and processing of cf. Asclepias bast fibers took place off-site. Finally, diverse tools offer evidence of specialized fiber production and manipulation, as we discuss shortly.
The use of various taxa of estuarine and wetland plants reflects the sophisticated ecological understanding of the ancient inhabitants of La Yerba II and III and highlights the critical significance of such ecologies in fostering diverse economic and social dynamics observed in many regions worldwide (Ramsey and Rosen, 2016; Ramsey et al., 2017; Ramsey, 2023).
The Cyperaceae family provided a number of resources that were critical for the marine hunter-gatherers at La Yerba. Plants from this family have prolific underground vegetative growth that consists of a network of rhizomes, tubers, and basal bulbs offering various reliable wetland resources with year-round availability. The family's taxonomy suffers from numerous revisions and synonymy. Species within this family commonly known as sedges have fibrous stems making them suitable for stripping into cordage and ropes used in numerous artifacts (see Table 1) and have long, flexible leaves suitable for splicing.
Some species of the Cyperaceae family, such as C. esculentus, also produce rhizomes that offer a starchy food source. Charred and desiccated C. esculentus rhizomes were recovered in some quantities from some La Yerba II contexts alongside their inflorescences (Table 1, Figure 7, Supplementary material 2). Similarly, Eleocharis flavescens and cf. Schoenoplectus have been reported at other Preceramic archaeological sites along this Pacific coast (Weir and Dering, 1986; Dillehay and Rossen, 2002; Benfer, 2008; Pearsall, 2009). C. esculentus is a perennial sedge that is 30–80 cm tall with triangular stems with fibrous roots. The plant can regenerate completely if even a single tuber is left in place. Tubers typically survive up to ~3–4 years. A single plant can produce thousands of tubers annually, while the carrying capacity is estimated at 1,030 million tubers/ha without crop interference. In tropical regions, where there is less seasonal variation, C. esculentus may have continuous growth throughout the year, whereas in temperate regions with distinct seasons, it may have a more pronounced growth period during the warmer months (Thullen and Keeley, 1979; Johnson and Mullinix, 1999; De Castro et al., 2015). Surface flow in dryland rivers like the Río Ica is highly seasonal and erratic, although its estuary has subsurface supplies all year round. Cyperaceae family plants can, therefore, be expected to persist all year round but thrive particularly during the austral summer when the river flows and soil moisture levels are high. Cf . Scirpus (Schoneoplectus) is typically a perennial or rhizomatous geophyte, which in temperate regions is often deciduous, undergoing dormancy during winter months, although roots and rhizomes remain alive underground. In a cool tropical maritime desert environment like the Río Ica estuary, its active growth season will follow the arrival of summer river flow when lush, grass-like foliage and inflorescences or flower spikes emerge. Small, clustered flowers can sometimes be used to identify Cyperaceae plants at the species level, but they are often clustered together. Flowers are followed by the production of seeds, which can contribute to the plant's spread and propagation.
Cf . Typha (T. domingensis) is a helophyte and grows primarily in the subtropical biome, where it provides medicine, fuel, and food (Arenas and Scarpa, 2003). Where water and nutrient supplies are sufficient, they can rapidly spread and colonize wetland areas. With warmer summer temperatures and enhanced river flow, Typha's growth will be promoted in the Río Ica estuary, and it will flower. As the dry season approaches, Typha plants may start to decline or enter a dormant phase but will rely on groundwater and nutrients and reserves stored in underground rhizomes to survive. Plants of the Typhaceae family are also distributed along the coast of Peru and Chile (Towle, 1961; Leon and Young, 1996; Beresford-Jones, 2011; Whaley et al., 2019) and some members also provide edible rhizomes and roots (Towle, 1961). The ethnobotany of Gran Chaco's Indigenous people (Arenas and Scarpa, 2003, p. 184–185, Figure 2) describes how “women go to the wetlands to cut the flowering stalks of cf. Typha and prepare the pollen at the edges of the wetlands, which is later consumed as a kind of yellow powder like sulfur” (Arenas and Scarpa, 2003, p. 184; Figure 7A). Typhaceae, likely T. dominguensis remains, have been reported in other Preceramic contexts along the coast of Peru (Weir and Dering, 1986; Dillehay and Rossen, 2002; Benfer, 2008; Pearsall, 2009). Typha spp. fibers are typically soft and have good tensile strength (Liu et al., 2017), making them ideal for weaving and twining structures into fabric artifacts.
Notably, different parts of Scirpus and Typha spp. plants were abundant in seasonally available starchy carbohydrates and fibers. Their harvesting may have entailed, beyond their seasonal exploitation, incipient wetland management to promote a continuous supply of food from edible components of tubers, seeds, and young shoots. Most importantly, however, they offered a consistent supply of fibers for crafting twined mats, looped bags cordage, and other essential artifacts. This all unfolded within the broader economic and ecological contexts of the Middle Preceramic, a time of significant shifts in consumption patterns and resource intensification dynamics (Beresford-Jones et al., 2021). These included an increased reliance on pelagic or pelagic-consuming marine resources, accompanied by the growing importance of cultivated food crops, not least high-protein beans. Throughout wetland resources, Scirpus and Typha complemented and enabled the harvesting technologies of Preceramic marine-based diets. Their exploitation doubtless shaped and influenced socio-economic structure. The multi-functional significance of both Scirpus and Typha plants may have promoted more efficient harvesting practices, assisted by their overlapping seasons for both food and fibers. This might have encompassed strategies such as seasonal, rotational harvesting practices alongside other mechanisms for regulating resource access and utilization, thereby enabling the hunter-gatherers of La Yerba II and III to manage these critical wetland resources adeptly and sustainably.
Bast fiber yarns at La Yerba are likely to be of Asclepias spp. (milkweed), or a closely related plant of the dogbane (Apocynaceae) family (Endress et al., 2014). This family includes a number of climbing vines of the tribe Asclepiadeae native to Peru, such as Asclepias curassavica and Funastrum clausum, recorded today in the Ica Valley (Whaley et al., 2019).
As hinted at by vernacular names such as “white twine vine,” “silkweed,” and, in Peru, “algodóncillo” (little cotton) and “flor de seda” (silk flower), plants of the Apocynaceae family have long been an important resource for fiber production throughout the Americas (Whiting, 1943). This plant yields copious seed floss – also known as vegetable silk or milkweed floss that can be harvested in late autumn, although at La Yerba people used the plant to produce fibers from the stems and not from the seeds.
Although Asclepias spp. seed pods have been reported at a number of Preceramic sites on the central (Patterson and Moseley, 1968; Cohen, 1978) and northern coasts of Peru, including at Huaca Prieta (Bird et al., 1985; Splitstoser et al., 2016), information on the use of its bast fibers is limited. Bird et al. (1985, p. 14) notes that “(Asclepias fabrics have) not made entirely of these fibers suggest(s) that either the supply was limited or that it was not very satisfactory.” We suggest, however, that the under-representation of Asclepias bast fibers in the archaeological record may be a problem of identification. Since these fibers are processed differently compared to those of cf. Scirpus and cf. Typha, their quality and color are also very different. Spliced fine Asclepias bast fibers look white and fluffy to the naked eye, so their initial classification is rather problematic. Even under low-definition microscopy, noting any residues of plant tissues and assigning them to a plant family without reference material is challenging. It is only when fibers are examined under a high-definition microscopy camera that we can make accurate observations of nodes (dislocation) and cross-marking in the fibers, these being key features for the classification of Apocynaceae bast fibers (Figure 5). These features of Asclepias bast fiber make spliced yarns from them all too easily mistaken for cotton (G. barbadense), so we suspect that some yarns and fibers previously identified as cotton without careful observation under a high-definition microscopy camera and proper contrasting with distinctive G. barbadense fibers (the seed fibers of which have, for instance, no nodes and dislocation features) may well be made of Asclepias.
Here, we emphasize riparian and estuarine habitats not only for the critical fiber plants they provided for fabric technologies but also for starchy foods for the Middle Preceramic coastal hunter-gatherers at La Yerba II and III. Before they were utterly changed by agriculture and the removal of much of their riparian woodlands, these offered very different floral and faunal landscapes from those that prevail today (Beresford-Jones, 2011). We suggest then that the spatial and ecological relationships that Preceramic people established with these habitats through the changing seasons helped define their patterns of occupation here.
Estuaries and coastal wetlands provided significant biomass of the low-ranked resources, such as the edible rhizomes of Cyperaceae esculentus and other plants of cf. Scirpus and cf. Typha (cf. Santoro et al., 2017). These were harvested, processed, and consumed probably by grinding them into flour or pounding them to extract the starch. Many mortars (batanes) have been recovered from La Yerba contexts (Figure 7). Such starch-rich food likely played an important role in coastal hunter-gatherers' diet and subsistence economy, not least likely to dilute the nitrogen load of very high-protein marine diets (cf. Speth and Spielmann, 1983; Dillehay, 2017; Beresford-Jones et al., 2018, 2021). For plant fiber technologies, evidently, coastal wetlands were ecological points that provided critical raw materials for the development of fiber production, specifically the bast fibers of cf. Scirpus and cf. Typha. These plants were harvested and processed to extract the strong fibers which were then spliced to create threads of varying thickness, as explained in the next section. Additionally, estuaries are composed of fine sediments that offer habitats for fish spawning, such as lisa (Mugil cephalus), which can be easily trapped as river levels fall, alongside habitats vital for nesting, breeding, and spawning of various coastal fauna and stopping points for migratory shorebirds. These ecological phenomena generally occur in summer when estuarine levels are high. Estuarine rhizomes provided an important dietary supplement alongside other food sources, including fish, shellfish, game, and foraged plants, together enabling sustainable human subsistence alongside marine, lomas fog oasis, riverine oasis, and estuary environments.
The utilization of riverine and riparian resources, however, must have required careful management and adaptation to the arid environment to ensure the sustainability of water and ecosystem resources in this challenging desert landscape. Along the river floodplains or other areas with groundwater sources, riparian vegetation is densely concentrated, providing habitats for hunted game, leguminous Prosopis woodlands with edible fruit, and a host of valuable resources, such as bottle gourd (Lagenaria sp.) for fishing floats or diverse containers; flexible willow (Salix humboltiana) wood for house construction and wood for fuel, and Apocynaceae vines for bast fibers. More widely, riparian vegetation performs important ecological functions, including serving as terrestrial and aquatic habitats, ensuring stream-bank stability, and providing natural corridors for larger migratory wildlife (Beresford-Jones, 2011).
Somewhat further afield from estuarine and riverine oases, Middle Preceramic patterns of marine hunter-gatherer occupation at La Yerba were also shaped by their interdependence with ephemeral winter-blooming lomas (Eichler and Londoño, 2013; Beresford-Jones et al., 2018). Lomas habitats provided seasonally critical terrestrial resources, including starchy rhizomes and land snails (Bostryx sp., 25% of total mollusc assemblage by MNI at La Yerba II), alongside hunted guanaco (Lama guanicoe), deer (Odocoileus virginianus), and vizcachas (Lagidium viscacia), all noted in the midden remains of both La Yerba II and III. Engel (1957, p. 275) suggests that Tillandsia sp. plants from fog oases were sources of fibers, although without providing much detail or evidence for the idea. Presumably, he meant fibers stripped from their leaf rosettes, but we have no evidence of distinctive Tillandsia trichome cells in plant fibers from La Yerba II or III. However, lomas habitats, nonetheless, required another set of ecological knowledge, where the temporality of subsistence strategies merged with seasonal exploitation of coastal resources through alternating cycles of austral winter and summer.
The knowledge of seasonal cycles and growth patterns of plants, which, in turn, enabled specialized knowledge of the availability of edible rhizomes and other plant foods, undoubtedly helped organize subsistence strategies on this Pacific coast. As Cyperaceae and Typhaceae families have both food and fiber applications, we suppose that “traditional knowledge” and practices related to their identification, harvest, processing, and use must have turned on peak abundance during summer: seasonal productivity defining, in turn, seasonal harvesting. Yet in times of need, edible Cyperaceae rhizomes could have provided a critical and readily available food resource that could be harvested throughout the year, even when other food sources may have been scarce. Meanwhile, seasonal rounds in fog oases habitats in winter when vegetation was at peak abundance called for other sets of “traditional knowledge” and different strategies, as discussed by Beresford-Jones et al. (2018). Winter–when estuaries, wetlands, and riverine are less productive–could have also been the ideal time to work fibers that had been harvested and processed during the spring and summer.
Ecological relationships defined by the seasonal growth of Scirpus and Typha create a framework for the temporalities of bast fiber technologies. If we imagine bast fiber production as an activity initiated when aquatic environments were managed through actions such as trimming, propagating, and collecting these plants through the austral summer, then, while ecological conditions are crucial to sustaining bast fiber production, this is also governed by the organization of people of La Yerba II and La Yerba III, with its roots in the social structures of the Middle Preceramic. These social structures are, therefore, intertwined with the economic and ecological contexts of marine and aquatic subsistence strategies. In this scenario, bast fiber technologies are not more than merely an aspect of Middle Preceramic technology but are integrated through various dimensions of technology, craft, and food preparation to shape the society itself.
There is little evidence of a highly specialized society at the La Yerba sites (e.g., Beresford-Jones et al., 2018), which in turn complements our idea of bast fiber production as incipient craft specialization as part of an organized system of co-dependent tasks, in which different activities were integrated and shared within the same society. While some contend that the specialized nature of sustaining reliable subsistence from the sea demands tools, skills, and knowledge that are incompatible with part-time fishing (Moseley, 1992, p. 22; Reitz et al., 2008, p. 129), modern ethnographies often emphasize the seasonal and part-time nature of fishing (Quilter, 1992; Tietze et al., 2011), which is evident also among contemporary onshore fishermen at the mouth of the Río Ica. Indeed, the essential compatibility of fishing and farming is, we suggest, central to the MFAC hypothesis (Beresford-Jones et al., 2018).
We suggest this integration or co-dependency of activities defined the seasonal organization of social groupings at La Yerba II and III. This means, for instance, that periods of reduced activity before and after the peak fishing season would have been used by the sedentary population of La Yerba III for the planting and eventually harvesting of crops, such as beans and gourds, and targeting other seafood less bound by specific seasons, including diverse gastropods, crustaceans, echinoderms, and seaweeds. Currently, extended families along the lower courses of the Río Ica similarly engage in such seasonal pursuits interspersed with a sophisticated schedule of various inshore and beach-based fishing activities. Estuarine and wetland plants show the most significant growth during spring and summer when river surface flows rise, likely defining the timing of plant gathering from these ecotones. Other aquatic resources require different timings. Strong river flows in summer can bring so much debris that net fishing at the river estuary can become impossible. Toward the end of summer, however, freshwater crayfish and estuary-spawning lisa become easy to catch using nets or even baskets as they become trapped by falling river levels (Beresford-Jones et al., 2018, p. 406).
The plant fiber chaîne opératoire begins with the collection of plants from which to extract bast fibers from the local estuary, wetlands, and river valley environments, which are doubtless entangled with other economic and subsistence practices. Interactions between Preceramic people and wetland plants in the context of their gathering likely intertwined different purposes: collecting plants for food, technology, and medicine. As Typhaceae, Cyperaceae, and Apocynaceae family plants grow in patches of vegetation circumscribed to sources of water across the desert coast, fiber artisans congregated around these to collect the bast fibers. As mentioned previously, coastal wetland, estuary, and riparian oasis flow regimes are characterized by surface flow during summer precipitation in the highlands, with periods of reduced water inputs during the rest of the year. Therefore, it seems likely that summertime was the most appropriate time to gather bast fibers, likely coinciding with other activities, such as gathering starchy rhizomes food of Cyperaceae esculentus and other plants of Typha and Scirpus (cf. Towle, 1961; Cohen, 1978; Reinhard et al., 2011).
Ethnographic and historical data provide insights into plant fiber technologies, including strategies, plant gathering, and management of aquatic plants. Tierra del Fuego hunter-gatherers collected Poaceae and Cyperaceae family plants that grow in low-lying water-rich habitats, like shallow pools, streams, and lake margins to produce baskets and for food (Belmar et al., 2007; Berihuete-Arizon, 2013). This seemed to have also been a seasonal exploitation that mostly took place in spring.
S. californicus (syn. Scirpus) is cultivated in certain regions of Ecuador and Peru to produce totora products. In the wild, S. californicus reproduces either vegetatively or from seed. However, in cultivation, the process begins right after harvesting, where the rhizomes from the harvested plants are permanently planted without allowing them to dry. To ensure optimal growth, the plants need to be submerged throughout the year. The green aerial stems of totora (S. californicus) are harvested between July and October when the inflorescence has matured and the plant has reached its maximum development (Heiser, 1979, in Macia and Balslevl, 2000, p. 86).
Another example can be found at the Medanos la Joyada, a midden site located in northern Peru. Based on the analysis of macro remains of marine fauna and archaeobotanical evidence, the inhabitants engaged in a diverse economy that included cultivating crops such as totora (S. californicus) alongside maritime resources. The sunken fields for totora cultivation at Medanos La Joyada were occupied from the Pre-Columbian to the colonial era, as documented by Kautz and Keatinge (1977). Rondón et al. (2003), Paredes and Hopkins (2018), and Prieto (2018) have documented that in the Huanchaco communities of northern Peru, Poaceae and Cyperaceae family plants, including S. californicus (syn. Scirpus), have been traditionally collected for the construction of boats and other crafts. The communities not only gathered and utilized these materials but also planted S. americanus rhizomes in sunken gardens near the beach to propagate and manage this valuable resource (Paredes et al., 2020).
Ethnohistoric documents from the seventeeth century, reviewed by Rostworoski (1981, in Banack et al., 2004), indicate the cultivation of totora (syn. Scirpus) in various parts of Peru. These documents, including those from the General Archives, mention locations such as Huaman, a pueblo near Trujillo, and the pueblo of Quilcay in the Lurin Valley. They suggest that indigenous communities employed a strategy of plant propagation and management by cultivating these aquatic plants that are critical for crafting seacrafts.
It is conceivable, we suggest, that incipient cultivators of the Middle Preceramic also planted Cyperaceae seeds and rhizomes in this way, thereby beginning this millennial tradition of manipulating estuary and wetland environments.
The processing of plant bast fibers involves various stages, including retting, splitting, peeling, grinding, and decortication, documented in various plant species, including bamboo (Xhauflair et al., 2023), nettles, hemp, and flax (Harris, 2014; Harris et al., 2017), for diverse purposes of cordage, baskets, and clothing (Hurcombe, 2007). Retting is the process of decomposition of non-fibrous materials surrounding bast fibers, typically through the methods of dew (or field) retting or water retting. In the former, plants are left on the ground for the requisite time for micro-organisms (e.g., fungi) to produce enzymes enabling a partial dissociation of stem tissue (Tahir et al., 2011; Liu et al., 2017). In water retting, plants are soaked in water and are colonized by anaerobic bacteria to accomplish the same dissociation. The specific retting process employed is not easily discerned from the morphology of archaeological fibers and depends very much on preservation and environmental context (Andresen and Karg, 2011; Gleba and Harris, 2019). Moreover, it is easily confused with the degradation of fibers (Lukesova and Holst, 2021). To identify retting, evidence of decomposition, such as signs of microbial degradation, including the presence of bacteria or fungi, lignin breakdown, or other alterations, in cell structures, needs to be identified (Djemiel et al., 2020).
We discerned distinct levels of processing in the bast fibers of La Yerba II and III, including varying degrees of epidermis detachment and different levels of decortication (see Supplementary material 1) that reflect diverse forms of bast fiber extraction, influencing fiber texture and appearance.
Scirpus and Typha fibers seem to require less extensive processing for their particular end uses, principally, decertification, probably without retting at all, which is evident from the identification of their strips or ribbons glued together. The absence of, or sometimes subtle retting of these bast fibers likely reflects a technological choice to retain plant pectins and gums, thus facilitating the fiber-making process (Gleba and Harris, 2019). The presence of epidermal tissue adhering to fibers reported by Alday (2023) from Preceramic sites of northern Chile also suggests this. Presumably, Scirpus stems were ripped off and separated, while Typha (cf. T. dominguesis) stems and leaves were processed by ripping them into strips, peeling them, and scrapping them. As these are aquatic plants, no true bark needs to be scrapped, rather, only gentle scrapping on the plant surfaces is needed to obtain different ranges of fiber width (Figures 8, 9).
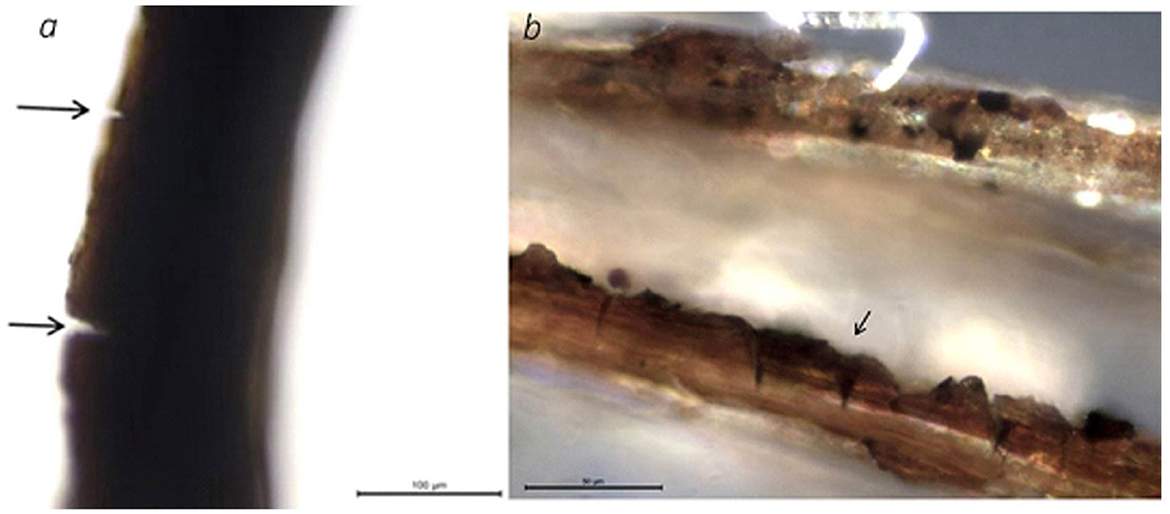
Figure 8. Cut marks cf. Scirpus: (a) La Yerba II Trench 1 SU 1015 and (b) La Yerba III Trench 1 SU 9550.

Figure 9. Specialized tools for fiber production: (A) La Yerba III Trench 1 SU 9522 net-making bone needles; (B) La Yerba III Trench 1 SU 9523 wooden needle; (C) La Yerba III Trench 3 SU 9013 14 cactus (cf. Haageocereu) needles; (D) La Yerba II Trench 1 SU 1005 modified Choromytilus sp. Shell; (E) La Yerba Trench 1 SU 9505 net-making bone needles; (F) La Yerba II, Trench 1 SU 1005 Choromytilus sp; (G) La Yerba II Trench 3 SU 1072 balls of raw material (cf. Scirpus); (H) La Yerba II Trench 1 UE 1016 ball of threads (cf. Scirpus).
We observe different processing to separate bast fibers from the stem in Asclepias fibers than in Scirpus or Typha spp. Under microscopic examination, these bast fibers appear significantly paler and finer. We presume that herbaceous perennial vines of the Apocynaceae family (cf. Asclepias) underwent partial retting before decortication and that vines were then crushed to extract the pulp and then cleaned. Stems would be crushed gently and bast fiber cracked off (see Figure 8). After retting, fine fibers were separated, presumably by hackling or combing, to be processed and spliced (Hurcombe, 2007), as reported for other bast fibers in Europe (Beugnier and Crombé, 2007; Harris, 2014; Loudon et al., 2023) and indeed other species of Apocynaceae in the Americas (Hulle et al., 2015).
These observations highlight distinct and deliberate technological choices. For instance, the absence of evidence of retting in Scirpus and Typha bast fibers suggests that coastal hunter-gatherers employed freshly harvested plants, which are easier to work into fibers, strengthening interpretations that these plants were collected during the summer and promptly utilized, eliminating the need for storage and avoiding surplus materials.
Although direct evidence from residue or micro-wear analyses is lacking, we suggest that lithics and grinding stones evident in both sites (Figure 9) were used in part for the preparation of plant material into fibers. Grinding stones are, for instance, used to soften maguey (Agavaceae family) fibers in northern Peru (Lucrezia Milillo, personal communication 2023). Finally, while other potential fiber-processing techniques like braking, scutching, and heckling are described elsewhere (Hurcombe, 2014; Harris et al., 2017), we propose that these methods would be more applicable when fibers are spun rather than spliced, as our samples have been, as we discuss next.
Once plants were cleaned by hand and epidermal material had been separated, fibers were used to produce spliced threads. Splicing is a technological process (sometimes called “thigh-spinning”), whereby fibers are twisted together without the use of a spindle necessary for spinning shorter yarns (Gleba and Harris, 2019). Just as in other ancient fabric technologies, such as those in Egypt, the Mediterranean, and across Europe (Barber, 1991; Gleba and Harris, 2019), splicing from bast fiber was the first technology of thread production in the Americas. Our study indicates that the threads are all spliced regardless of the raw material. Thicker spliced threads were used in mats and skirts, whilst finer spliced threads were used in bags and nets. The technical gesture is described as “[t]o twist the yarn, thigh-spinners use friction between a hand and a leg to roll fibers into a yarn” (Tiedemann and Jakes, 2006, p. 295).
Splicing as a core technique to make threads is also correlated with how plants are processed since the length of leaves and stems are directly linked to the thread-making process (see Gleba and Harris, 2019). In splicing, artisans used long strips of fibers and twisted them tightly together, which was a strenuous and highly time-consuming activity (Tiedemann and Jakes, 2006). As a manual technique, splicing does not require spinning equipment and so generates variation in the appearance of threads (Gleba and Harris, 2019).
When threads are made using splicing technology, two methods can be used: (1) “continuous splicing, in which individual fiber strips are added continuously into a single twisting thread. While the thread is twisted, new fiber strips are laid adjacent to fiber strips already caught into the thread, thereby becoming incorporated into the single thread. As part of the same procedure, two singles may be twisted together into a plied yarn which will be plied in the opposite direction to the singles” (Gleba and Harris, 2019, p. 2330), and (2) end-to-end splicing, in which the end of one fiber strip (or fiber ribbon) is joined to the end of a second fiber strip, which in turn is joined to a third and so on, creating a continuous thread. As the joins are inherently weak, typically two such spliced threads are twisted together. The twisting is a two-stage process of splicing the singles end-to-end, then plying them” (Gleba and Harris, 2019, p. 2330). As Table 1 summarizes, spliced threads have different diameters and angles of twist. The diameter of the threads varies, with all threads displaying a diameter of <1 mm. The final plying shows a strong preference for S-direction (n = 22) while Z-direction was shown in 13 samples.
End-to-end splicing and continuous splicing were represented almost equally among our samples (see Supplementary material 1). Thread making was a time-consuming component of fiber manufacture and, therefore, an important dimension of the economic and social activities of the hunter-gatherers of La Yerba II and La Yerba III. Most of the yarns in our samples, for instance, in fragments of nets and looped bags, are quite fine and seem to have been continuously spliced because the ends of their bundle of bast fibers are difficult to discern. By contrast, end-to-end spliced threads are more common on fragments of mats. Finally, spliced and plied yarns were used to produce close-weft twined mats, open-weft twined mats, double-looped bags, and nets (Alday, 2023, p. 65).
The earliest specialized tools associated with textile production are spindles used to spin fibers into yarn and reported at various Late Preceramic sites after 5000 BP (Engel, 1963; Moseley, 1975; Bird et al., 1985; Doyon-Bernard, 1990; Quilter et al., 1991; Dillehay et al., 2017). Such equipment is necessary for the use of slippery protein animal fibers, such as camelid fiber, and relatively short plant fibers such as cotton (Beresford-Jones et al., 2018). Spinning equipment was not, however, part of the technological repertoire of the Middle Preceramic coastal hunter-gatherers at La Yerba II and III and has not, to our knowledge, been reported at any other site of this period. Bast fiber technologies of the Middle Preceramic therefore required pre-spindle whorl techniques such as splicing using hands and thighs.
Three modified Choromytilus sp. mussel shells modified and used as scraper tools at the La Yerba II site, two modified mussel shells, and one modified Argitecten purpuratus shell at La Yerba III might represent another dimension of plant fiber production because, alongside denticulated lithics, they might have been used to extract bast fibers and carry out decortication (Figure 9). Similarly, sub-circular needles made from cactus spines of tubular bird bones would likely have been used to fabricate more delicate fabric structures (Gabriel et al., 2005). Many net-making needles have also been recovered in La Yerba III contexts (Figure 9), used by passing through a pre-arranged looping structure to create a continuous interconnected pattern (Figure 9). Such needles were used to make and maintain the fishing nets also evidenced at La Yerba III. Twined mats are fabricated through twining, which involves twisting or wrapping fibers around each other to form a stable structure (Alday, 2022; Figure 9). Thus, we suggest that needle artifacts were used for both one-element structures, looping and knotting, while two-element structures such as twining were done using finger manipulation and other artifacts that we have not found in our contexts.
The existence of specialized tools for net manufacture at the permanently settled village of La Yerba III stands in marked contrast to the contexts of the earlier La Yerba II shell midden: a contrast also marked in the very different assemblages of fish remains at each site (Beresford-Jones et al., 2018). The use of twinned mats was, however, crucial to the construction of shelters and working surfaces at both sites. Craft specialization as a social activity likely involved all members of a group, either directly or indirectly. Coastal hunter-gatherers were participants in the technological process either as producers or as consumers so the participation of both individuals and social groupings became an essential element of the craft production (Figure 10).
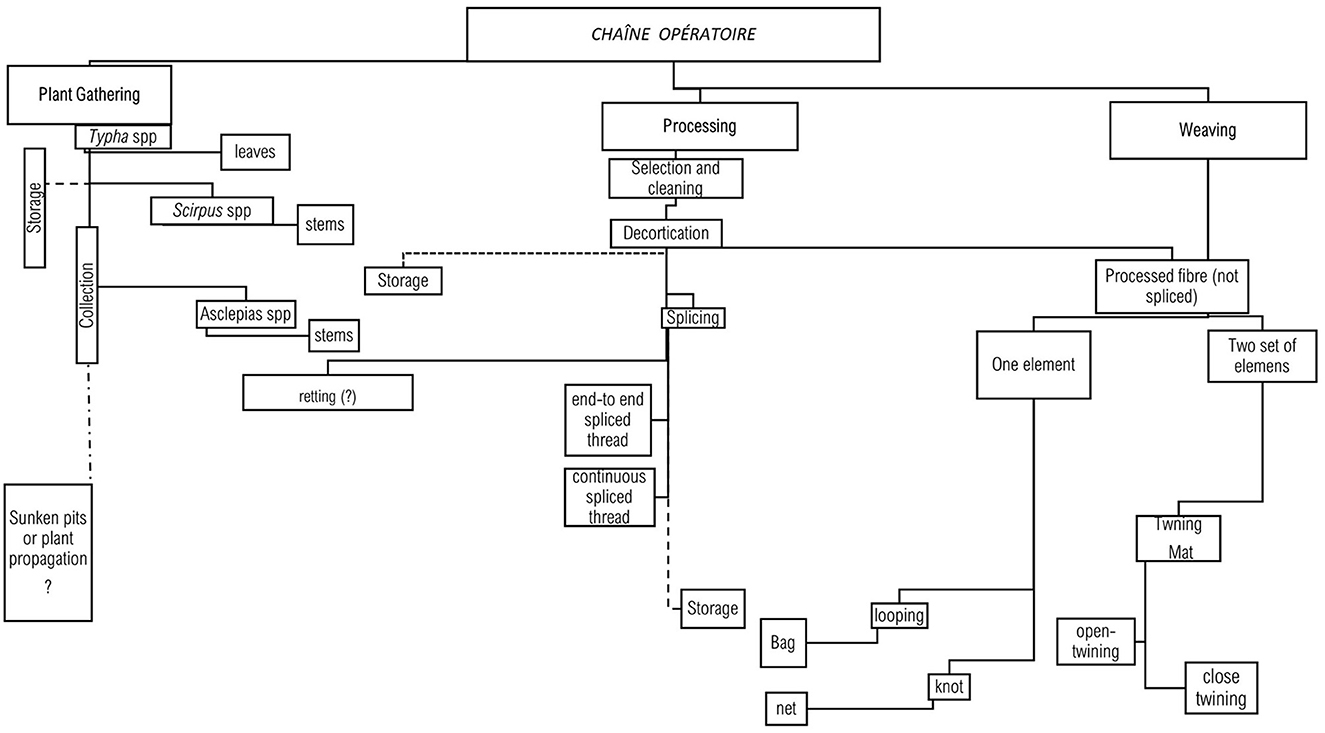
Figure 10. Reconstruction of the chaîne opératoire.
We invoke the concept of “craft specialization” to reflect how bast fiber technologies–encompassing an array of techniques, tools, and gestures–comprise a comprehensive technological system that fosters a sense of community by bringing together skilled bodies, specialized knowledge, and plant materials into an intricate chaîne opératoire. In both instances of La Yerba II and La Yerba III, this technological framework yielded social and technological advantages. It is likely that individuals within these groups organized various activities: tending aquatic environments, propagating resources, and collecting edible and fibrous raw materials in conjunction with other coastal activities in seasonal rounds like fishing, diverse seafood collection, hunting, and other terrestrial foraging. While we infer that social groupings within these societies might have organized themselves according to age, skills, and social identities, there might be other, even less visible, social categories that also defined roles in these ancient societies.
It has long been assumed that women played a significant–albeit not exclusive–role in these social dynamics in activities such as monitoring, picking, weeding, watering, watching, trimming, tending, and replanting plants (e.g., Reiter, 1975; Binford, 1982; Jochim, 1988; Hastorf, 1998). As Hastorf (1998, p. 77) notes, women were likely consistently and intimately engaged in the task of plant tending, nurturing, and the earliest forms of plant management, emerging from their primary engagement with the foraging of wild plant resources. While Hastorf primarily discusses plant domestication and cultivation for food purposes, she also highlights the broader importance of plants in early human societies, including their roles in technology, medicine, and so on. Indeed, gender almost certainly emerges as a crucial social category in the context of early bast fiber production. This significance is not necessarily restricted to the physical fabrication of artifacts but, rather, to the wider potential roles women may have played in nurturing wetlands, estuaries, and riverbanks as productive zones for both food and raw materials, within the context of Middle Preceramic lifestyles along the southern coast of Peru.
While roles often stereotypically ascribed to women, such as plant gathering, weaving, and plant tending, were once perceived as secondary contributions to the past economy and, therefore, less defining of social life, such notions have long been challenged (see, e.g., Mason and Hather, 2002). We suggest that on the Pacific coast of South America, bast fiber production, if anything, occupied a central position within the social, economic, and ecological relationships of the long Preceramic Period. Conceivably, most craft production toward the creation of bast fiber artifacts was carried out by women, with children and older adults perhaps also contributing in various capacities. We refrain, however, from proposing a strict binary division of labor between men and women (e.g., Jochim, 1988). Nonetheless, our perspective serves to underscore the need to shed further light on women's often less visible roles in the Preceramic Period without embracing axioms calling for new theoretical perspectives beyond the binary women/men division of society.
Middle Preceramic marine hunter-gatherers at both La Yerba II and III foraged local plants such as sedges (Cyperaceae), cattails (Typhaceae), and milkweeds (Apocynaceae) for manufacturing early fabrics on the Pacific coast. The procurement of these plants for fiber production represents a dimension of coastal hunter-gatherer cultural and ecological knowledge, not least in identifying plant species, their growth patterns, and seasonal availability, as well as recognizing their different physical characteristics for their specific purposes.
Estuaries and wetlands provided a relatively stable and predictable food supply throughout the year including starchy rhizomes of C. esculentus and Typha capsules and, therefore, likely contributed to settlement patterns in close proximity to such habitats, illustrating the adaptation of these hunter-gatherers to the specific opportunities and challenges presented by their local ecosystems. Moreover, reconstruction of that environment suggests a much more extensive palaeo-lagoon and estuarine marsh area during the mid-Holocene at the time when La Yerba II and III were occupied (Beresford-Jones et al., 2022, and cf. raised coastal embayment at Otuma 120 km to the north; Craig, 1968, p. 100).
Among plant remains from the La Yerba II and III contexts, those of the Cyperaceae family have a particular significance evidenced by their frequent occurrence in the archaeological contexts of both La Yerba II and III in the form of Scirpus spp. seeds and inflorescences, desiccated and charred C. esculentus rhizomes as food source, and cf. Scirpus stems and processed fibers as valuable raw materials for looped bags, a huge variety of yarns, and some knotting and twined structures. Similarly, rhizomes of cf. Typha, although not as abundant as Cyperaceae, were perhaps consumed, and its fibers certainly were used in twined mats. Apocynaceae bast fibers were, by comparison, used to fabricate fewer, generally finer artifacts for specific purposes such as fishing nets and lines. These would have required a more demanding manufacturing process including retting compared with straightforward decortication of Scirpus spp. and Typha spp.
These processes of fiber production relied on coastal hunter-gatherers' understanding of optimal times and methods for harvesting and processing of these plants. After gathering fibers, these were separated through decortication and retting to remove non-fibrous tissues and ease the extraction of plant fibers using scrapers. Once the plants were processed into fine and flexible fibers, they were spliced into yarn and interlaced into looped and/or knotted nets using both sub-circular and flat net-making needles at La Yerba III.
The archaeobotanical remains from La Yerba II and La Yerba III thereby illustrate an extended Middle Preceramic knowledge of plants for both technology and food: a knowledge that extended beyond individual plants to surely embrace entire ecosystems. This evidence contributes to a wider understanding of the significance of wetland and estuarine habitats to Mesolithic hunter-gatherers. Moreover, ancient riparian environments before the advent of agriculture provided different, wooded landscapes that served as important corridors for the movement of larger wildlife between winter lomas and summer highland hinterlands in seasonal patterns of migration that likely shaped hunter-gatherer movements too.
Finally, we emphasize how marine subsistence strategies along this Pacific coast entailed different adaptation strategies to ensure a sustainable supply of food and resources, which may have included incipient cultivation of Cyperaceae rhizomes or seeds, for critical food and fiber supplies. Bast fiber production as a craft specialization, created collective technological and ecological knowledge. This interplay not only shaped the relationship between coastal hunter-gatherers and their coastal and terrestrial environments but also facilitated an ongoing coastal lifestyle, performed in various tasks, fostering cooperative labor and defining social structures. We may still lack a comprehensive picture of their impacts at the family or kinship level, but it is nonetheless evident that these fiber artifacts were critical to the Middle Preceramic technological repertoire and catalyzed technological and ecological transformations, especially as the subsequent Late Preceramic Period introduced new ways of harvesting the sea.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
CAld: paper design and writing, plant fiber analyses, and interpretations. CAla and MG: analyses of archaeobotanical data. DB-J: paper writing, director of archaeological fieldwork, interpretations, and project funding. All authors contributed to the article and approved the submitted version.
This research was supported by the Leverhulme Trust (grant number RPG-117) and Doctoral Research: ANID-Becas Chile/Cambridge Trust, University Fieldwork Fund (2018–2019), Wolfson College Travel Fund (2018), and Simón Bolívar Fund, University of Cambridge (2018).
We wish to thank the two reviewers of this article; Catherine Kneale (University of Cambridge) for their help with microscopy; the members of the One River Project particularly George Chauca, Oliver Huaman, Lauren Cadwallader, and Sandy Pullen; the Director of the Museo Regional de Ica Susana Arce for access to samples, Alberto Benavides G. and the people of Samaca; and the Ministerio de Cultura del Perú for granting permission for fieldwork and the export of samples for dating.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2023.1251137/full#supplementary-material
Adovasio, J. (1977). Basketry Technology. A Guide to Identification and Analysis. Los Angeles, CA: Left Coast Press.
Adovasio, J., and Dillehay, T. (2020). Perishable Technology and the successful peopling of South America. PaleoAmerica 6, 21022. doi: 10.1080/20555563.2019.1686849
Alday, C. (2022). “Chapter 5: early fibre production in the west coast of south america: the case of plant-fibre technology of the south coast of Peru,” in Exploring Ancient Textiles: Pushing the Boundaries of Established Methodologies: 40, eds A. Dickey, M. Gleba, S. Hitchens, and G. Longhitano (Oxford: Oxbows), 55–68.
Alday, C. (2023). Fibre Production Among the Coastal Hunter-gatherers of South America's West Coast: New Narratives from Plant Fibre Technologies. doi: 10.17863/CAM.94903
Andresen, S. T., and Karg, S. (2011). Retting pits for textile fibre plants at Danish prehistoric sites dated between 800 BC and AD 1050. Veg. History Archaeobot. 20, 517–526. doi: 10.1007/s00334-011-0324-0
Arenas, P., and Scarpa, G. F. (2003). The consumption of Typha domingensis pers. (Typhaceae). Pollen among the ethnic groups of the Gran Chaco, South America. Econ. Bot. 57, 181–188. doi: 10.1663/0013-0001(2003)057[0181:TCOTDP]2.0.CO;2
Banack, S. A., Rondón, X. J., and Diaz-Huamanchumo, W. (2004). Indigenous cultivation and conservation of totora (Schoenoplectus californicus, Cyperaceae). in Peru. Econ. Bot. 58, 11–20. doi: 10.1663/0013-0001(2004)058[0011:ICACOT]2.0.CO;2
Barber, E. J. W. (1991). Prehistoric Textiles: The Development of Cloth in the Neolithic and Bronze Ages With Special Reference to the Aegean. Princeton, NJ: Princeton University Press.
Belmar, C., Mendez, C., and Reyes, O. (2007). Hunter-gatherer plant resources used during the Holocene in central western Patagonia (Aisen, Chile, South America). Veg. History Archaeobot. 26, 607–625. doi: 10.1007/s00334-017-0632-0
Benfer, R. A. (2008). “Early villages,” in Encyclopedia of Archaeology, ed D. M. Pearsall (New York, NY: Academic Press), 368–80.
Beresford-Jones, D. G. (2011). The Lost Woodlands of Ancient Nasca. Oxford: Oxford University Press.
Beresford-Jones, D. G., Friesem, D. E., Sturt, F., Pullen, A., Chauca, G. E., Moat, J., et al. (2022). Insights into changing coastlines, environments and marine hunter-gatherer lifestyles on the Pacific Coast of South America from the La Yerba II shell midden, Río Ica Estuary, Peru. Quaternary Sci. Rev. 285, 107509. doi: 10.1016/j.quascirev.2022.107509
Beresford-Jones, D. G., Mader, C., Lane, K. J., Cadwallader, L., Gräfingholtf, B., Chauca, G. E., et al. (2023). Beyond Inca roads: archaeological mobilities from the high Andes to the Pacific in Southern Peru. Antiquity 97, 194–212. doi: 10.15184/aqy.2022.168
Beresford-Jones, D. G., Pomeroy, E., Alday, C., Benfer, R. A., Quilter, J., O'Connell, T., et al. (2021). Diet and Lifestyle in the First Villages of the Middle Preceramic: Insights from stable isotope and osteological analyses of human remains from Paloma, Chilca I, La Yerba III and Morro I. Latin Am. Antiq. 32, 741–759. doi: 10.1017/laq.2021.24
Beresford-Jones, D. G., Pullen, A. G., Chauca Iparraguirre, G., Cadwallader, L., García, M., Salvatierra, I., et al. (2018). Refining the maritime foundations of andean civilization: how plant fiber technology drove social complexity during the preceramic period. J. Archaeol. Method Theor. 25, 393–425. doi: 10.1007/s10816-017-9341-3
Bergfjord, C., and Holst, B. (2010). A procedure for identifying bast fibres using microscopy: flax, nettle/ramie, hemp and jute. Ultramicroscopy 110, 1192–1197. doi: 10.1016/j.ultramic.2010.04.014
Bergfjord, C., Karg, S., Rast-Eicher, A., Nosch, M. L., Mannering, U., Allaby, R. G., et al. (2010). Comment on 30,000-year-old wild flax fibres. Science 328, 1634. doi: 10.1126/science.1186345
Berihuete-Arizon, M. (2013). First archaeobotanical approach to plant use among Selknam hunter-gatherer (Tierra del Fuego, Argentina). Archaeol. Anthropol. Sci. 5, 255–266. doi: 10.1007/s12520-013-0137-4
Beugnier, V., and Crombé, P. (2007). Plant Processing from a Prehistoric and Ethnographic Perspective. BAR International Series 1718. London: John and Erica Hedges Ltd.
Binford, L. (1982). The archaeology of place. J. Anthropol. Archaeol. 1, 5–31. doi: 10.1016/0278-4165(82)90006-X
Bird, J., Hyslop, J., and Skinner, M. (1985). The preceramic excavations at the Huaca Prieta Chicama Valley, Peru. Anthropol. Pap. Am. Museum Nat. History 62, 1–294.
Bronk Ramsey, C. (2009). Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360. doi: 10.1017/S0033822200033865
Cohen, M. (1978). Archaeological plant remains from the central coast of Peru. Ñawpa Pacha J. Andean Archaeol. 16, 23–50. doi: 10.1179/naw.1978.16.1.003
Connolly, T. J., Barker, P., F owler, C. S., Hattori, E. M., Jenkins, D. L., and Cannon, W. J. (2016). Getting beyond the point: textiles of the terminal pleistocene/early holocene in the north western great basin. Am. Antiq. 81, 490–514. doi: 10.7183/0002-7316.81.3.490
Craig, A. K. (1968). Marine Desert Ecology of Southern Peru, Vol. 1. Arlington, VA: Office of Naval Research, Geography Branch.
Day, J., Hunter, R., Keim, R. F., DeLaune, R., Shaffer, G., Evers, E., et al. (2012). Ecological response of forested wetlands with and without large-scale Mississippi River input: implications for management. Ecol. Eng. 46, 57–67. doi: 10.1016/j.ecoleng.2012.04.037
De Castro, D., Gargiulo, O., Del Guacchio, R., Caputo, E. P., and De Luca, P. (2015). A molecular survey concerning the origin of Cyperus esculentus (Cyperaceae, Poales): two sides of the same coin (weed vs. crop). Annal. Bot. 115, 733–745. doi: 10.1093/aob/mcv001
Dillehay, T. (2017). Where the Land Meets the Sea: Fourteen Millennia of Human History at Huaca Prieta, Peru. Austin, TX: University of Texas Press.
Dillehay, T. D., Goodbred, S., Pino, M., Vásquez Sánchez, V. F., Tham, T. R., Adovasio, J., et al. (2017). Simple technologies and diverse food strategies of the late pleistocene and early holocene at Huaca Prieta, coastal Peru. Sci. Adv. 3, e1602778. doi: 10.1126/sciadv.1602778
Dillehay, T. D., and Rossen, J. (2002). Plant Food and its Implications for the Peopling of the New World: A View From South America. The First Americans: The Pleistocene Colonization of the New World. San Francisco, CA: California Academy of Sciences, 237–253.
Djemiel, C., Goulas, E., Badalato, N., Chabbert, B., Hawkins, S., and Grec, S. (2020). Targeted metagenomics of retting in flax: the beginning of the quest to harness the secret powers of the microbiota. Front. Genet. 11, 581664. doi: 10.3389/fgene.2020.581664
Dodge, C. R. (1897). A Descriptive Catalogue of Useful Fiber Plants of the World, Vol. 9. New York, NY: US Government Printing Office.
Doyon-Bernard, S. J. (1990). From twining to triple cloth: experimentation and innovation in ancient Peruvian weaving (ca. 5000-400 BC). Am. Antiq. 55, 68–87. doi: 10.2307/281493
Eichler, T., and Londoño, A. (2013). ENSO impacts on lomas formation in south coastal Peru: Implications for the pliocene?. Adv. Meteorol. 2, 1–7. doi: 10.1155/2013/175947
Endress, M. E., Liede-Schumann, S., and Meve, U. (2014). An updated classification for Apocynaeae. Phytotaxa. 159, 175–194. doi: 10.11646/phytotaxa.159.3.2
Engel, F. (1957). Early sites on the Peruvian coast. Southwestern J. Anthropol. 11, 13546. doi: 10.1086/soutjanth.13.1.3629157
Engel, F. (1963). A preceramic settlement on the central coast of Peru: Asia, unit−1. Trans. Am. Philos. Soc. New Serie 53, 1–139. doi: 10.2307/1005956
Florian, M., Dale, P., and Kronkright Norton, R. (1991). The conservation of Artifacts Made from Plant Materials. Los Angeles, CA: Getty Publications.
Gabriel, O., Lange, K., Dahm, E., and Wendt, T. (2005). Fish Catching Methods of the World, 4th Edn. Oxford: Blackwell Publishing.
Gilligan, I. (2018). “Agriculture and textiles in the Americas,” in Climate, Clothing, and Agriculture in Prehistory: Linking Evidence, Causes, and Effects, ed Gilligan I (Cambridge: Cambridge University Press), 161–173.
Gleba, M., and Harris, S. (2019). The first plan bast fibre technology: identifying splicing in archaeological textiles. Archaeol. Anthropol. Sci. 11, 2329–2346. doi: 10.1007/s12520-018-0677-8
Hardy, B., Moncel, M., Kerfant, C., Lebon, M., Bellot-Gurlet, L., and Mélard, N. (2020). Direct evidence of Neanderthal fibre technology and its cognitive and behavioral implications. Sci. Rep. 10, 4889. doi: 10.1038/s41598-020-61839-w
Hardy, K. (2007). “Where would we be without string? Ethnographic and prehistoric evidence for the use, manufacture, and role of string in the upper paleolithic and mesolithic of northern Europe,” in Plant Processing From a Prehistoric and Ethnographic Perspective BAR International Series, eds Beugnier, V and Combe, P. (Ghent: Ghent University), 9–22.
Harris, S. (2014). “Flax fibres: innovation and change in the early neolithic a technological and material perspective. Textile society of America,” in Biennal Symposium Proceedings: New Directions: Examining the Past, Creating the Future. Los Angeles, CA, 10–14.
Harris, S., Haigh, S., Handley, A., and Sampson, W. (2017). Material choices for fibre in the Neolithic: an approach through the measurement of mechanical properties. Archaeometry 59, 574–591. doi: 10.1111/arcm.12267
Hastorf, C. (1998). The cultural life of early domestic plant use. Antiquity 72, 773–782. doi: 10.1017/S0003598X00087366
Heiser, C. (1979). The totora (Scirpus Califonicus). in Ecuadro and Peru. Economic Botany. 32, 222–236. doi: 10.1007/BF02864698
Hogg, A., Hua, Q., Blackwell, P., Niu, M., Buck, C., Guilderson, T., et al. (2013). SHCal13 Southern Hemisphere calibration, 0-50,000 years cal BP. Radiocarbon 55, 1889–1903. doi: 10.2458/azu_js_rc.55.16783
Hulle, A., Kadole, P., and Katkar, P. (2015). Agave Americana leaf fibers. Fibers 3, 64–75. doi: 10.3390/fib3010064
Hurcombe, L. (2007). “Plant processing for cordage and textiles using serrated flint edges: new chaine operatoires suggested by combining ethnographic, archaeological and experimental evidence for bast fibres processing,” in Plant Processing From a Prehistoric and Ethnographic Perspectives, BAR International Series, eds V. Beugnier and P. Crombe (Ghent: Ghent University), 1718.
Hurcombe, L. (2014). Perishable Material Culture in Prehistory: Investigating the Missing Majority. London: Routledge.
Jochim, M. (1988). Optimal foraging and the division of labor. Am. Anthropol. 90, 130–136. doi: 10.1525/aa.1988.90.1.02a00100
Johnson, W. C., and Mullinix, B. G. (1999). Cyperus esculentus interference in Cucumis sativus.Weed Sci. 47, 327–331. doi: 10.1017/S0043174500091852
Jolie, E. A., Lynch, T. F., Geib, P. R., and Adovasio, J. M. (2011). Cordage, textiles, and the late Pleistocene peopling of the Andes. Curr. Anthropol. 52, 285–296. doi: 10.1086/659336
Kautz, R. R., and Keatinge, R. W. (1977). Determining site function: a north Peruvian coastal example. Am. Antiq. 42, 86–97. doi: 10.2307/279464
Kvavadze, E., Bar-Yosef, O., Belfer-Cohen, A., Boaretto, E., Jakeli, N., Matskevich, Z., et al. (2009). 30,000-year-old wild flax fibers. Science. 325,1359–1359. doi: 10.1126/science.1175404
Leon, B., and Young, K. R. (1996). Aquatic plants of Peru: diversity, distribution and conservation. Biodiv. Conserv. 5, 1169–1190. doi: 10.1007/BF00051570
Linskens, H., and Jorde, W. (1997). Pollen as food and medicine: a review. Econ. Bot. 78–86. doi: 10.1007/BF02910407
Liu, J., Zhang, Z., Yu, Z., Liang, Y., Li, X., Ren, L., et al. (2017). The structure and flexural properties of Typha leaves. Appl. Bionics Biomech. 2017, 870. doi: 10.1155/2017/1249870
Loudon, N. N., Wollstonecroft, M., and Fuller, D. Q. (2023). Plants to textiles: local bast fiber textiles at pre-pottery neolithic Çatalhöyük. J. Archaeol. Sci. Rep. 49, 103940. doi: 10.1016/j.jasrep.2023.103940
Lukesova, H., and Holst, B. (2021). Is cross-section shape a distinct feature in plant fibre identification? Archaeometry 63, 216–226. doi: 10.1111/arcm.12604
Macia, M., and Balslevl, H. (2000). (Schoenoplectus califonicus, Cyperacea). in Ecuador. Econ. Bot. 54, 82–89. doi: 10.1007/BF02866602
Martens, T., and Cameron, J. (2019). Early coastal fiber technology from the Caleta Vitor archaeological complex in northern Chile. Latin Am. Antiq. 30, 287–299. doi: 10.1017/laq.2018.78
Mason, S. L. R., and Hather, J. G. (2002). Hunter-Gatherer Archaeobotany. Perspectives From the Northern Temperate Zone. London: UCL Press.
Melelli, A., Shah, D. U., Hapsari, G., Cortopassi, R., Durand, S., Arnould, O., et al. (2021). Lessons on textile history and fibre durability from a 4,000-year-old Egyptian flax yarn. Nat. Plants 7, 1200–1206. doi: 10.1038/s41477-021-00998-8
Montt, I., Valenzuela, D., Cases, B., Santoro, C. M., Capriles, J. M., Standen, V. G., et al. (2023). Chinchorro fibre management in the atacama desert and its significance for understanding Andean textilization processes. J. Anthropol. Archaeol. 71, 101530. doi: 10.1016/j.jaa.2023.101530
Morton, J. (1975). Cattails (Typha spp.)-weed problem or potential crop? Econ. Bot. 29, 7–29. doi: 10.1007/BF02861252
Moseley, M. (1975). The Maritime Foundations of Andean Civilisation. Menlo Park: Cummings Publishing Company.
Moseley, M. (1992). Maritime foundations and multilinear evolution: retrospect and prospect. Andean Past. 3, 1–5.
Moseley, M. E., and Feldman, R. A. (1988). “Fishing, farming and the foundations of Andean civilization,” in The Archaeology of Prehistoric Coastlines, eds G.Bailey and J. Parkington (Cambridge: Cambridge University Press), 125–134.
Nadel, D., Danin, A., Werker, E., Schick, T., Kislev, M. E., Stewart, K., et al. (1994). 19, 000.-year-old twisted fibers from Ohalo II. Curr. Anthropol. 35, 451–458. doi: 10.1086/204303
Paredes, R., and Hopkins, A. L. (2018). Dynamism in traditional ecological knowledge persistence and change in the use of totora (Schoenoplectus californicus). for subsistence in Huanchaco, Peru. Ethnobiol. Lett. 9, 169–179. doi: 10.14237/ebl.9.2.2018.1176
Paredes, R., Hopkins, A. L., and Villanueva, F. (2020). Ethnobotany in the north coast of Peru: use of plants in the fishing community of huanchaco for subsistence. Econ. Bot. 74, 32–45. doi: 10.1007/s12231-020-09486-0
Patterson, R., Lowe, B. J., and Smith, C. (2017). Light microscopy: an old technique casts new light on maori textile plantas. Archaeometry 59, 965–979. doi: 10.1111/arcm.12281
Patterson, T. C., and Moseley, M. E. (1968). Late preceramic and early ceramic cultures of the central coast of Peru. Ñawpa Pacha J. Andean Archaeol. 6, 115–133. doi: 10.1179/naw.1968.6.1.006
Pearsall, D. M. (2009). Investigating the transition to agriculture. Current Anthropol. 50, 609–613. doi: 10.1086/605406
Prieto, G. (2018). The social dynamics and economic interactions of the households at gramalote, a small-scale residential settlement during the second millennium BC on the NorthCoast of Peru. Latin Am. Antiq. 29, 532–551. doi: 10.1017/laq.2018.10
Quilter, J. (1992). To fish in the afternoon: Beyond subsistence economies in the study of early Andean civilization. Andean Past 3, 1–10.
Quilter, J., Ojeda, B., Pearsall, D. M., Sandweiss, D. H., Jones, J. G., Wing, E. S., et al. (1991). Subsistence economy of El Paraíso, an Early Peruvian site. Science 251, 277–283. doi: 10.1126/science.251.4991.277
Quilter, J., and Stocker, T. (1983). Subsistence economies and the origins of Andean complex societies. Am. Anthropol. 85, 545–562. doi: 10.1525/aa.1983.85.3.02a00030
Ramsey, M. (2023). Ecological-cultural inheritance in the wetlands: the non-linear transition to plant food production in the southern Levant. Veg. History Archaeobot. 24, 1–18. doi: 10.1007/s00334-023-00908-2
Ramsey, M., and Rosen, A. (2016). Wedded to wetlands: exploring Late Pleistocene plant-use in the eastern levant. Q. Int. 396, 5–19. doi: 10.1016/j.quaint.2015.10.109
Ramsey, M., Rosen, A., and Nadel, D. (2017). Centered on the wetlands: integrating new phytolith evidence of plant-use from the 23, 000.-year-old site of Ohalo II, Israel. Am. Antiq. 82, 702–722. doi: 10.1017/aaq.2017.37
Rast-Eicher, A., Karg, S., and Jørgensen, L. (2021). The use of local fibres for textiles at Neolithic Çatalhöyük. Antiquity 95, 1129–1144. doi: 10.15184/aqy.2021.89
Reinhard, K., Le Roy Toren, S., and Arriaza, B. (2011). Where have all the plant foods gone? The search for refined dietary reconstruction from chinchorro mummies yearbook of mummy studies. Nat. Res. 1, 139–151.
Reitz, E., Andrus, C., and Sandweiss, D. (2008). Ancient fisheries and marine ecology of coastal Peru. Hum. Impact. Anc. Mar. Ecosyst. Glob. Persp. 29, 125–146. doi: 10.1525/9780520934290-008
Rondón, X. J., Banack, S. A., and Diaz-Huamanchumo, W. (2003). Ethnobotanical investigation of caballitos (Schoenoplectus californicus: Cyperaceae). in Huanchaco, Peru. Econ. Bot. 57, 35–47. doi: 10.1663/0013-0001(2003)057[0035:EIOCSC]2.0.CO;2
Rostworoski, W. (1981). Recursos Naturals Renova- bles y Pesca, Siglos XVI y XVII. Lima: Instituto de Estudios Peruanos.
Sadrmanesh, V., and Chen, Y. (2019). Bast fibres: structure, processing properties and applications. Int. Mat. Rev. 64, 381–406. doi: 10.1080/09506608.2018.1501171
Sandweiss, D. (2003). Terminal Pleistocene through Mid-Holocene archaeological sites as paleoclimatic archives for the Peruvian coast. Palaeogeogr. Palaeoclimatol. Palaeoecol. 194, 23–40. doi: 10.1016/S0031-0182(03)00270-0
Santoro, C. M., Capriles, J. M., Gayo, E. M., De Porras, D., Maldonado, M. E., Standen, A., et al. (2017). Continuities and discontinuities in the socio-environmental systems of the Atacama Desert during the last 13, 000. years. J. Anthropol. Archaeol. 46,28–39. doi: 10.1016/j.jaa.2016.08.006
Santos, M., and Standen, V. (2022). El temprano arte de la tradición de tejer esteras en fibra vegetal en la sociedad Chinchorro (10.000-3500 aP): Extremo norte de Chile. Latin Am. Antiq. 33, 355–375. doi: 10.1017/laq.2021.59
Speth, J. D., and Spielmann, K. A. (1983). Energy sources, protein metabolism, and hunter-gatherer subsistence strategies. J. Anthropol. Archaeol. 2, 1–31. doi: 10.1016/0278-4165(83)90006-5
Splitstoser, J. C., Dillehay, T. D., Wouters, J., and Claro, A. (2016). Early pre-Hispanic use of indigo blue in Peru. Sci. Adv. 2, e1501623. doi: 10.1126/sciadv.1501623
Standen, V. G. (2003). Bienes funerarios del cementerio Chinchorro Morro 1: descripción, análisis e interpretación. Chungará 35, 175–207. doi: 10.4067/S0717-73562003000200002
Suomela, J., Vajanto, K., and Räisänen, R. (2018). Seeking nettle textiles–utilizing a combination of microscopic methods for fibre identification. Stu. Conserv. 63, 412–422. doi: 10.1080/00393630.2017.1410956
Tahir, P. M., Ahmed, A. B., SaifulAzry, S. O., and Ahmed, Z. (2011). Retting process of some bast plant fibres and its effect on fibre quality: a review. BioResources 6, 5260–5281. doi: 10.15376/biores.6.4.5260-5281
Thullen, R. J., and Keeley, P. E. (1979). Seed production and germination in Cyperus esculentus and C. rotundus. Weed Science. 27, 502–505. doi: 10.1017/S0043174500044489
Tiedemann, E., and Jakes, K. (2006). An exploration of prehistoric spinning technology: spinning efficiency and technology transition. Archaeometry 48, 293–307. doi: 10.1111/j.1475-4754.2006.00257.x
Tietze, U., Lee, R., Siar, S., Moth-Poulsen, T., and Båge, H. E., (eds.). (2011). Fishing With Beach Seines. Fisheries and Aquaculture Technical Paper No. 562. Rome: FAO.
Weir, G., and Dering, J. (1986). “The lomas of paloma: human-environment relations in a central peruvian fog oasis: archaeobotany and palynology,” in Monographs in Archaeology No. 27: Andean Archaeology, eds R. Matos M., S. A. Turpin, and H. H. Eling (Los Angeles, CA: Institute of Archaeology, University of California), 18–44.
Whaley, O., Orellana, A., and Pecho-Quispe, O. (2019). An annotated checklist to vascular flora of the ica region, Peru—with notes on endemic species, habitat, climate and agrobiodiversity. Phytotaza. 389, 1–125. doi: 10.11646/phytotaxa.389.1.1
Whiting, G. (1943). A Summary of the Literature on Milkweeds (Asclepias spp.)., and Their Utilisation (No. 2). Bibliographical Bulletin. Washington, DC: United States Department of Agriculture
Xhauflair, H., Jago-On, S., Vitales, T., Manipon, D., Amano, N., Callado, J., et al. (2023). The invisible plant technology of Prehistoric Southeast Asia: indirect evidence for basket and rope making at Tabon Cave, Philippines, 39–33,000 years ago. PLoS ONE 18, e0281415. doi: 10.1371/journal.pone.0281415
Keywords: plant fibers, hunter-gatherers, Pacific coast, South America, paleoethnobotany, Cyperaceae, estuaries, wetlands
Citation: Alday C, García MA, Alarcón C and Beresford-Jones D (2023) The use and context of fiber plants during the middle preceramic: evidence from La Yerba II and III, south coast of Peru. Front. Environ. Archaeol. 2:1251137. doi: 10.3389/fearc.2023.1251137
Received: 30 June 2023; Accepted: 02 October 2023;
Published: 07 November 2023.
Edited by:
Alison Crowther, The University of Queensland, AustraliaReviewed by:
Justin Bradfield, University of Johannesburg, South AfricaCopyright © 2023 Alday, García, Alarcón and Beresford-Jones. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Camila Alday, Y2NhMjhAY2FtLmFjLnVr
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.