
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Environ. Archaeol. , 14 August 2023
Sec. Zooarchaeology
Volume 2 - 2023 | https://doi.org/10.3389/fearc.2023.1208908
This article is part of the Research Topic Editor's Showcase: Zooarchaeology View all 4 articles
 Edouard Masson-MacLean1*
Edouard Masson-MacLean1* Sesilia Niehaus1
Sesilia Niehaus1 Elizabeth Fraser1,2
Elizabeth Fraser1,2 Veronica Lee3Shalen Prado4Orsolya Czére1Jovita Fawcett1James O'Driscoll1
Veronica Lee3Shalen Prado4Orsolya Czére1Jovita Fawcett1James O'Driscoll1 Linus Girdland-Flink1Gordon Noble1
Linus Girdland-Flink1Gordon Noble1 Kate Britton1
Kate Britton1Our knowledge of the use of livestock in early medieval Scotland is fragmentary and relies on a handful of well-studied faunal assemblages, with few from Pictland (north-east Scotland), an important and enigmatic group and latterly kingdom in Northern Britain that existed between the 3rd and 9th centuries AD. The assemblages that have been recovered and studied mainly occur at the limits of this territory, beyond the heartland of the Picts in the northern and eastern mainland. Recent archaeological excavations at three high-status sites in eastern Scotland have unexpectedly yielded well-preserved faunal remains providing a unique and long-awaited opportunity to explore further human-animal relationships and the use of animals in Pictish society. This paper presents new data from the initial study of these assemblages. It discusses the implications in terms of animal economy in Pictland, the potential of these sites to yield larger faunal assemblages and the directions of future research. Results show that cattle were a pivotal element of the economy, playing a multi-faceted role (beef and secondary products), pigs ranked second in frequency which likely reflects the high status of these sites and sheep appear as a marginal resource and were primarily raised for consumption. Results also suggest that these sites may have operated within an integrated network rather than functioning solely as self-sufficient entities.
Domestic animals played a central role in the early medieval societies of north-west Europe. As pillars of the economy, they provided food, wealth, raw materials, and labor (McCormick, 2014; O'Connor, 2014), and also played key symbolic roles in human lifeworlds (Crabtree, 1995; Bond, 1996). The remains of livestock are therefore a valuable material to study human-animal relationships during the 1st Millennium AD and are of far greater significance than just proxies for diet (O'Connor, 2011). While there is a growing body of research on early medieval animal use in Europe (Castillo, 2014), northern Britain and Pictland (Figure 1), in particular, appear as zooarchaeological blackholes when compared to contemporary neighbors like Ireland and England (O'Connor, 2011, 2014; McCormick, 2014; McCormick and Murray, 2017). In Pictland, the main underlying reason is the limited number of sites that have been identified and investigated (Foster, 2014, p. 105–106; Noble et al., 2019a; Noble, 2020), combined with poor bone preservation due to acidic soils. Faunal assemblages large enough to generate valuable interpretative data have been limited to a handful of sites located on coastal or insular sandy soils, mostly in the northwestern-most edges of Pictland (Dockrill et al., 2010) or lie in other adjacent kingdoms such as Dál Riata (Lane and Campbell, 2000; Murray et al., 2004) or be Lothians/Northumbria (Perry, 2000; Crone et al., 2016). Furthermore, unlike Ireland and England, there is a lack of historical sources for Pictland that could provide valuable complementary information on animal use (Foster, 2014, p. 62; Noble and Evans, 2022, 1–22). Amplified by the scarce zooarchaeological evidence, our understanding of the role of domestic animals in Pictish society and the economy more broadly remains poorly characterized.
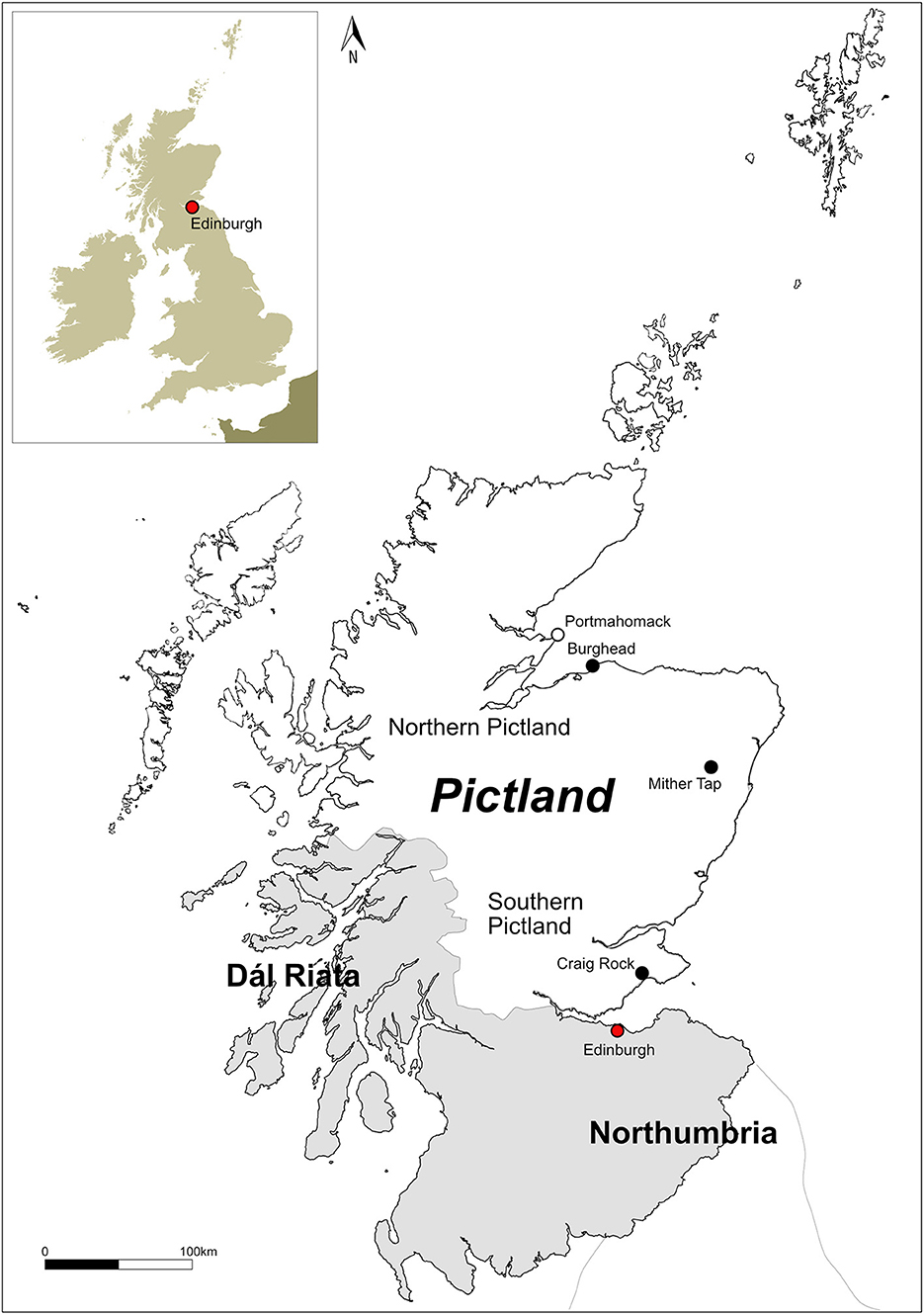
Figure 1. Map of Pictland and modern Scotland showing sites (black) and placenames (white) mentioned in the text.
First mentioned in late Roman sources as a collective name for people living north of the Roman frontier, the Picts went on to dominate a large part of present-day Scotland until the late first millennium AD (Noble and Evans, 2022). The emergence of the Pictish over-kingdom, the precursor of the kingdom of Scotland, was part of broader changes in northern Europe that laid the foundations for the modern states of Europe. The major legacies of the Picts include iconic symbol stones (e.g., RCAHMS, 2007), but other than this, the archaeological and historical record for this region c. 300–900 AD has been traditionally diffuse and difficult—famously dubbed the “Problem of the Picts” (Wainwright, 1955). Our understanding of the socio-political organization of Pictland is poor, but there are securely documented Pictish kings from the 6th century onwards and there was an overkingship of Pictland from at least the 7th century AD, centered initially around the Moray Firth region (Woolf, 2006), with the center of power shifting southwards in the Viking Age (Noble and Evans, 2022, p. 290–293). With regards to the archaeology, recent excavations, have begun to reveal a whole host of new sites and evidence providing important insights into the functioning of Pictish society and the international connections that rulers in Pictland were able to draw upon (Carver et al., 2016; Noble et al., 2019a,c, 2020).
Since 2010, the University of Aberdeen has investigated a range of power-centers including previously unknown examples (Noble et al., 2013, 2019b,c). Since late 2017, the Leverhulme/Historic Environment Scotland-funded Comparative Kingship Project has targeted over 40 sites for excavation, including hillforts, promontory forts, ecclesiastic enclosures, cemeteries, and the location of in-situ carved stones, providing a wealth of new knowledge (Noble and Evans, 2022). At most sites, faunal remains were either absent or limited to a few bone and teeth fragments, but in some rare cases, well-preserved animal bone were recovered. As most excavations consisted of small-scale evaluations, these new faunal assemblages remain small in comparison to contemporary English and Irish sites. Nevertheless, they provide a rare opportunity to add to the body of evidence regarding animal use in Pictish society and address some key research questions. What economic practices were livestock involved in at these sites and is there inter-site variability? Is the animal economy at Pictish elite sites a self-sufficient operation or is it part of network of producer and consumer sites? These new assemblages also highlight the potential of some of these sites to yield larger faunal assemblages of greater interpretative value. This paper presents the first archaeozoological data from the secular elite sites of Burghead, Mither Tap, and Craig Rock (Figure 1) and discusses animal exploitation in eastern Pictland, the archaeozoological potential of these sites, and highlights future research directions.
The sites presented here (Burghead, Craig Rock and Mither Tap) comprise fortified high-status hill- or promontory forts (Noble et al., 2019c, p. 58), monument types that are often associated with elite occupation and interpreted as centers of power and authority. The size, location, and material culture from these sites correspond well with this narrative, with excavations producing evidence for monumental buildings, high-status metal-working and other elite indicators.
Burghead is a promontory fort located on the Moray Firth coast of Scotland (Figure 1). It is one of the largest (5.5 ha) known Pictish sites (Foster, 2014, p. 43) and is defined by massive timber laced ramparts protecting both the seaward and landward sides. Despite a large portion of the site being destroyed by 19th century construction of the modern village and harbor (Noble and O'Driscoll, 2019, p. 28), the site contains a rich archaeological record and is best known for more than 30 bull carvings (Young, 1890, 1891, 1893; Ralston, 2004; Oram, 2007; RCAHMS, 2007). The fort was occupied between the 6th and 10th centuries AD and appears to have been abandoned in the Viking Age. There is a long history of excavations at Burghead extending back to the 19th century, including the University of Aberdeen (2015 to present).
The hillfort of Craig Rock, Fife, was occupied between the 7th and 9th Century AD. Overlooking the Firth of Forth, the site lies within an area of dense Pictish activity and a few kilometers from the barrow cemetery at Lundin Links (Greig et al., 2002; Lock and Ralston, 2017). The site has a complex system of enclosing elements (2.16 ha) surrounding a flat oval summit. Prior to the University of Aberdeen's investigations, Craig Rock had not been investigated, and had only been recognized as a “possible hillfort” through aerial photography. Small-scale excavations from 2019 confirmed Craig Rock as an early medieval site with excellent preservation of deposits.
Mither Tap is complex hillfort with multiple enclosing elements, located at the summit of Bennachie (518 m AOD), Aberdeenshire. Dating to the 5th−9th Centuries AD it comprises a large outer rampart and smaller inner rampart which surround an imposing granite tor. Antiquarian work revealed the limited areas where habitation was possible were densely occupied. However, it wasn't until the early 2000s that rescue excavations by the Forestry Commission confirmed the site dated to the early medieval period (Atkinson, 2007; Noble and O'Driscoll, 2019, p. 45). A single season of excavation at Mither Tap in 2019 confirmed the presence of numerous internal structures and occupation deposits and provided more precise dating evidence for the construction and use of the hillfort.
For all three sites, animal bone was hand-collected combined with on-site dry-sieving using a 10 mm mesh and bulk samples were taken from selected contexts. Dogs do not appear to have been a major taphonomic factor in the formation of the faunal assemblages with gnawing observed on 4.4% of the bones from Burghead (NISP = 76), 2.4% from Craig Rock (NISP = 8) and none from Mither Tap. Bone weathering was extremely rare suggesting the rapid burial of bone after discard and only 2 specimens at Burghead and 10 at Craig Rock (3% NISP) showed evidence of Stage 1 cracking (Behrensmeyer, 1978). Burnt bone, charred, and calcined, was also infrequent with only 3.3% at Craig Rock (NISP = 11), under 1% at Burghead (NISP = 8), and Mither Tap (NISP = 1) at Burghead, the more extensive excavations from 2018, 2019, and 2021 produced the largest assemblage, with over 1,600 specimens identified beyond class level with the sandy soils contributing to the excellent bone preservation at the site. The evaluation trenches at Craig Rock produced an assemblage of 189 identifiable specimens from an occupation layer and midden deposits abutting the rampart wall on the northern side of the summit, where soil conditions appear conducive to bone preservation. The Mither Tap assemblage was also small, with 161 bone fragments identified. Except for some well-preserved faunal remains from midden deposits at the foot of the southern rampart, bone survival was generally very poor. It is possible that the large amount of bone originally deposited near the rampart altered the soil chemistry rendering burial conditions favorable to bone preservation.
The analysis of the faunal material was undertaken using standard zooarchaeological methods and techniques (Reitz and Wing, 2008; Driver, 2011). Taxonomic identifications were made by comparison to the zoological reference collection at the University of Aberdeen, complemented with the use of identification guides (Schmid, 1972; Cannon, 1987; Cohen and Serjeantson, 1996; Watt et al., 1997). To estimate the economic significance of each taxa, relative abundance was calculated using Number of Identifiable Specimens (NISP) and Minimum Number of Individuals (MNI). Age at death estimates were based on long bone fusion using Silver (1969) and tooth eruption and wear for cattle, pigs and ovicaprids (Payne, 1973; Grant, 1982; Greenfield and Arnold, 2008; Jones and Sadler, 2012; Lemoine et al., 2014). Given the limited number of mandibles with complete or partial tooth rows, isolated lower deciduous fourth premolars (dp4) and permanent third molars (M3) were also included in the analysis (Gillis et al., 2022, p. 9).
The main domesticates dominated all three assemblages, representing approximately 90% of identified remains (NISP; Table 1). Cattle (Bos taurus) was the most common taxon, representing at least half of all specimens. Cattle were slightly more frequent at Burghead (56.27%) than at Mither Tap (50.63%) or Craig Rock (49.21%). Pigs (Sus scrofa) ranked second, representing ~30% NISP on average, though they appear to be less frequent at Burghead (14.26%) where pigs and sheep (Ovis aries) and/or goat (Capra hircus) are almost equally represented, though pigs are more frequent based on MNI. Ovicaprids ranked third with a mean NISP of 14.9%.
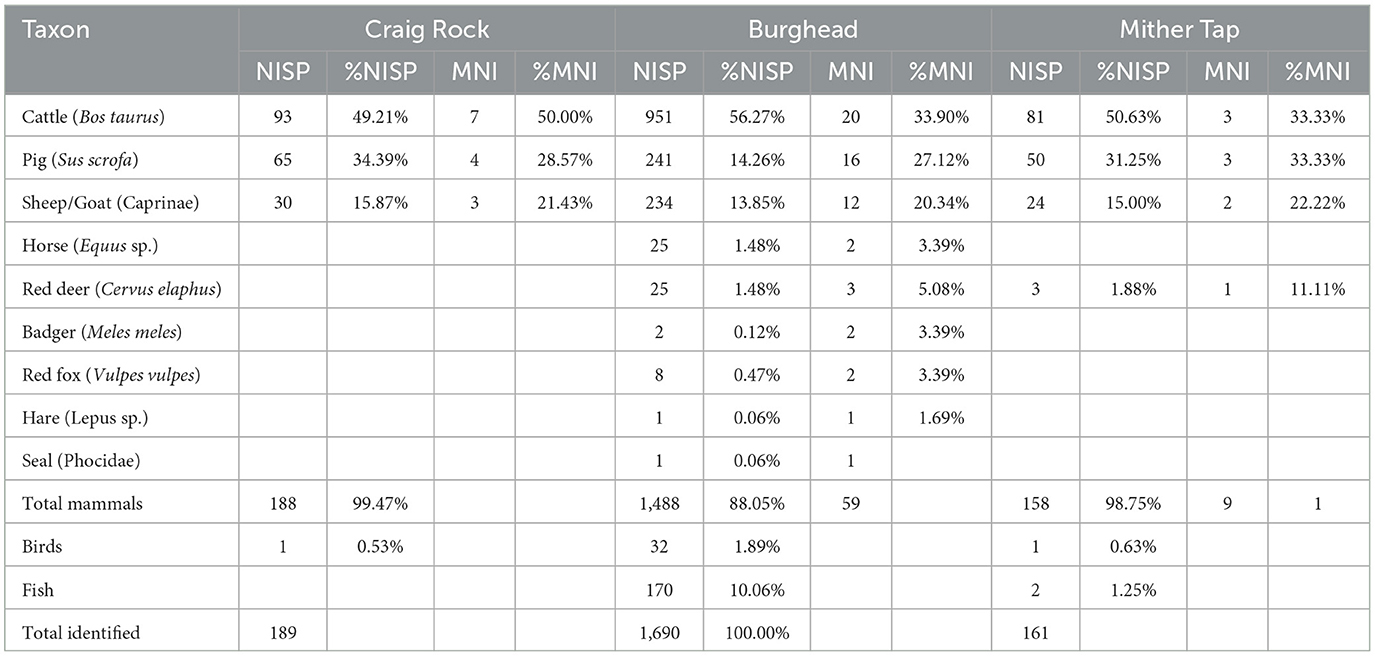
Table 1. Taxonomic identifications from Burghead, Mither Tap and Craig Rock.
All parts of the carcass were present in the assemblages. At Burghead, meat-bearing bones were well represented and cranial elements and metapodials were present but not overrepresented (Supplementary Figure 1). At both Mither Tap and Craig Rock, meatier limb bones (femur, humerus) were better represented.
Except for Burghead, the age at death dataset was of limited interpretative value due to the small size of the samples. There is an apparent deficit in juveniles and immature individuals in all assemblages based on tooth and long bone data, though occasional fetal/neonatal remains were identified at Burghead (5.7% cattle NISP). Subadult cattle (1.5–3 years) appear to be more frequent at Mither Tap and Craig Rock than at Burghead where adults and mature adults seem to predominate the assemblage (Figure 2). Tooth data for Craig Rock was limited to just five isolated maxillary M3, which had wear indicative of individuals under 3.5 years. Unfortunately, no cattle teeth suitable for wear analysis were recovered from Mither Tap. At Burghead, cattle appear to have been primarily culled as adults (over 3.5 yrs) with older adults more frequent. Out of 13 lower M3s, eight had dentine exposure on the distal cusp indicative of adult animals over 5 years of age (Mulville et al., 2005, p. 176).
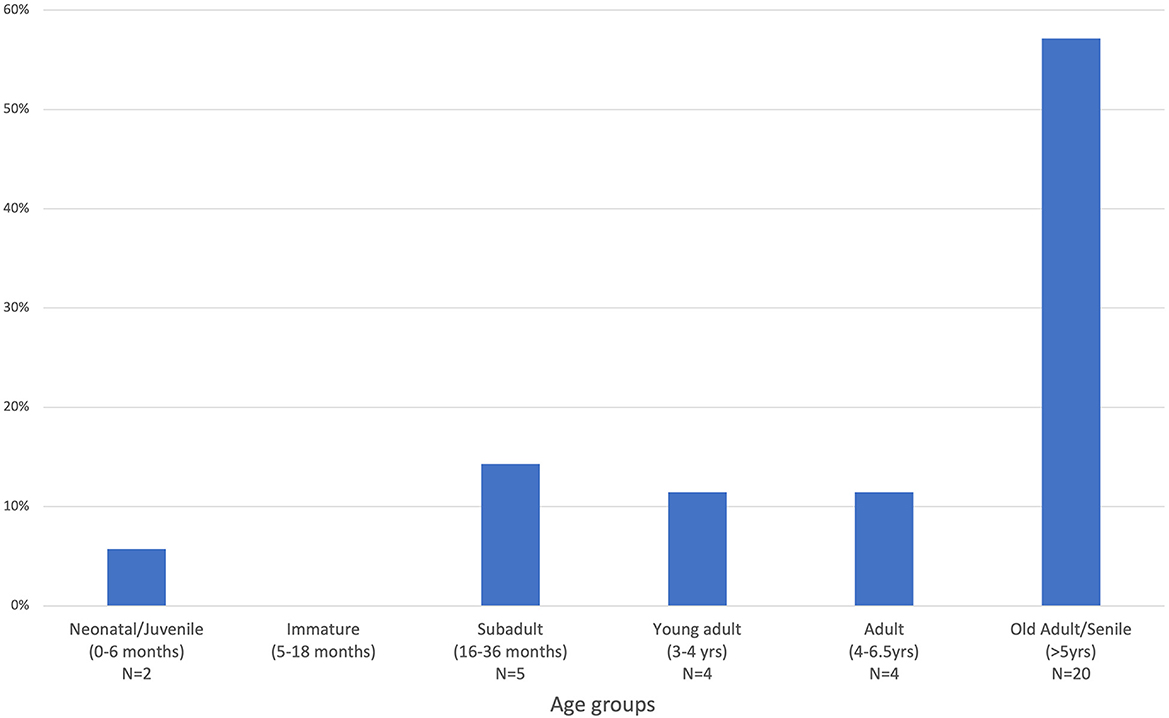
Figure 2. Cattle age at death profile for Burghead based on mandibles and isolated teeth (N = 35). Age groups were defined following Halstead (1985, p. 219), O'Connor (2003, p. 160), and Jones and Sadler (2012).
Long bone fusion data seems to largely mirror tooth data. Unfused long bones were more frequent at Mither Tap (17.3%) and Craig Rock (41.5%) than at Burghead (12.13%) where there is a higher cattle survival rate based on long bones (Figure 3, Supplementary Table 1). This indicates a greater proportion of animals slaughtered before reaching skeletal maturity at Craig Rock, possibly Mither Tap too, than at Burghead. This pattern is only tentative for Mither Tap due to the small number of long bone fusion data points but is more discernible at Craig Rock (Figure 3). Unfused early fusing bones were also rare, suggesting most cattle were slaughtered after 2 years of age at all three sites. Though adults and older adults are predominant at Burghead, there is evidence based on long bone fusion that some cattle were slaughtered as subadults around 2–3 years of age (Supplementary Figure 2) a trend also visible in the tooth data.
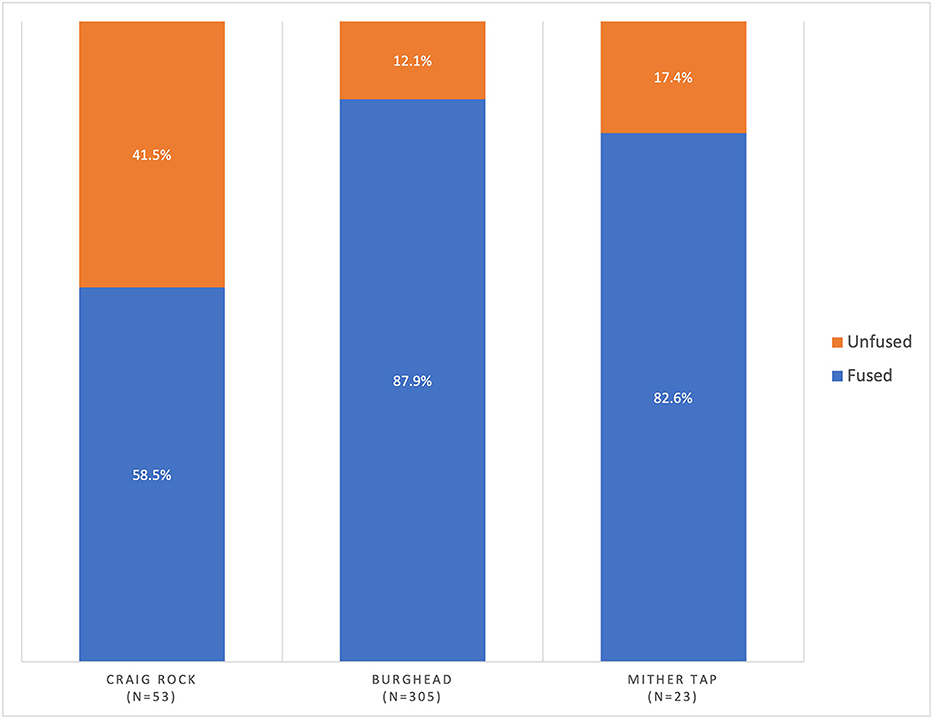
Figure 3. Proportion of unfused cattle bones at Burghead, Mither Tap and Craig Rock (N represents number of fusion points).
A small number of cattle specimens from Burghead were suitable for sex determination and appear to indicate a predominance of cows. These include three pelvises all identified as female based on the height of the medial border of the acetabulum (Greenfield, 2006) and all six measurable distal metacarpals width <55 mm and regarded as female (after McCormick, 1992).
Pigs ranked second at all sites in terms of NISP and MNI. All body parts appear present in the assemblages, although the most frequent elements were mandibles and long bones. The age at death dataset currently available is small and renders the identification and interpretation of pig culling patterns at the sites difficult. To increase sample size, mandibles and isolated M3 were combined and assigned to broad age groups (Supplementary Table 2), Group I (0–12 months), Group II (12–52 months), Group III (52–96 months), Group IV (over 96 months) following Lemoine et al. (2014). All groups were present, though pigs from Group IV were rare. Group II predominated (42.9%) indicating most individuals were slaughtered between 12 and 52 months. Group I and Group III were equally represented (25%−28%). Data for Group I show piglets were slaughtered at all sites and where more accurate mandibular data was available, the majority were slaughtered after 6 to 8 months (with the exception of a single neonate from Craig Rock).
Caprines appear less well-represented than cattle and pigs. The small dataset available seems to suggest that most body parts are present. Sample sizes for age at death determination is currently small for ovicaprids at Craig Rock, Burghead and Mither Tap. At Mither Tap and Craig Rock the data available is reduced to a single mandible and a few long bone fusion points per site making any interpretations speculative. At both sites the remains come from sheep/goat slaughtered between 2 and 4 years. At Burghead, based on 15 mandibles, there appears to be a lack of immatures (N = 1) and seniles (N = 1) with subadults/young adults (2–4 years; N = 5) and older adults (N = 5) potentially predominant and the presence of some juveniles (N = 3) was also observed.
The only other domestic mammal identified was horse (Equus sp.) represented by a handful of specimens from Burghead (1.48%). Dogs (Canis familiaris) and cats (Felis catus) were not identified in the studied assemblages, though a dog mandible has recently been identified in the 2022 Burghead material (E. Masson-MacLean unpublished data). Gnawing marks were also observed on bones confirming the presence of dogs at Burghead and Craig Rock. Wild mammals, birds and fish were absent or a rare occurrence the majority of which were identified from Burghead, which also produced a sizeable fish assemblage of mainly gadids and salmonids (V. Lee and S. Niehaus unpublished data). Taxonomic richness, the number of species represented (Gifford-Gonzalez, 2018, p. 488), was therefore very low at all sites, though slightly higher at Burghead which could be the effect of sample size.
The predominance of cattle remains at the sites adds to the body of evidence regarding the importance of cattle in the Pictish economy (Dockrill et al., 2010; Foster, 2014, p. 201–208; Carver et al., 2016) and echoes the ubiquitous pattern identified throughout early medieval Britain and Ireland (McCormick, 2008; Holmes, 2014; O'Connor, 2014). Based on the available data, cattle were raised for secondary products, and beef production. The presence of all skeletal components indicates that livestock are likely to have been brought to sites “on the hoof.” Contrary to Mither Tap and Craig Rock, multiple age groups of cattle were represented at Burghead, which may indicate husbandry at or near the site. At present, the absence of cattle fetal and/or neonatal remains (under 1 month) at Craig Rock and Mither Tap may indicate a lack of breeding close to these sites. On the other hand, the presence of this age group at Burghead, albeit in low numbers, suggests some calving took place within the fort or nearby and most likely represented accidental deaths. The lack of calves in the assemblages is an indication that intensive dairying and calf skin exploitation were not carried out at these sites (Halstead and Isaakidou, 2017, p. 119; Gidney, 2018, 4). This contrasts with the monastic site of Portmahomack where a greater proportion of juveniles was related to vellum production (Carver et al., 2016). The presence of subadults and young adults in all assemblages attest that a proportion of cattle were slaughtered for meat production, likely representing young males that were surplus to requirements (McCormick, 2014, p. 122). At Craig Rock and Mither Tap this age group appears to be better represented, though the picture is incomplete due to the absence of age data based on teeth. Nevertheless, it cannot be ruled out that Craig Rock and Mither Tap are consumer sites and the provisioning of beef to these hillforts was an integral part of the local economy. Of particular interest was the apparent difference in the proportion of 2–4-year-old cattle with Burghead. Though there is a “peak” in subadults and young adults, older adults predominated, indicating that cattle had a primary function other than meat production such as dairying, labor and breeding as part of a mixed strategy (Holmes et al., 2021, 5). There appears to be a lack or absence of older adult cattle at the hillforts, though the dearth of suitable teeth for aging hinders the reconstruction of detailed age profiles.
In the absence of reliable sex data, it is not yet possible to ascertain the role of older cattle at Burghead. However, the few specimens for which sex could be determined suggest females may be predominant and be indicative of the presence of dairy cows. Individuals over 8 years are likely to represent the overhaul of dairy stock (Balasse et al., 2000, p. 44; Kamjan et al., 2021, 5). The current consensus is that dairying was an important component of a mixed economy of pastoral and arable farming in Iron Age and early medieval Scotland (Mulville et al., 2005; Foster, 2014, p. 167). The importance of draft animals cannot be underestimated, however, considering the role of cereal production in generating surplus and wealth (McCormick, 2008, p. 219). Other by-products of cattle would also have been valuable, such as hides, bones and horns as raw materials and there is growing evidence of craft activities taking place at Burghead.
Pigs were the second most important domesticate on all three sites, though appear to be better represented at Craig Rock and Mither Tap. Pigs are often associated with high-status sites and are considered an animal of choice for feasting (Smith, 2000, p. 720; Madgwick et al., 2019) and the apparent predominance of age Group II (1–4 years) indicates pigs were generally slaughtered once they reached optimal size. The presence of older individuals at all sites could represent breeding stock and pig rearing but also animals given as rents or payments (McCormick, 2014, p. 123). Ovicaprids ranked third and appear to be of minor economic importance, contrasting with the higher frequency of sheep found in insular or “marginal” environments (Dockrill et al., 2010; McCormick, 2014, p. 124) or in England (Holmes, 2014, 2017; O'Connor, 2014). Sheep/goat appear to have been primarily slaughtered for meat at 2–4 years old. The presence of various age groups at Burghead, including lambs and mature adults, implies the presence and management of a sheep flock nearby for meat and possibly the small-scale production of wool and milk as well as for reproduction.
Compared to Pictland, Ireland and England have numerous and sizeable early medieval faunal assemblages, providing opportunities to identify producer/consumer and consumer sites (McCormick et al., 2011, p. 99–105; Holmes, 2014). There is a shift in England between the 7th and 9th Century AD from a self-sufficient economy to an emerging market-based economy (Holmes, 2014, p. 125). Increasing political, economic and social complexity led to the development of producer and consumer sites involving the provisioning of high-status secular and ecclesiastical centers and proto-urban settlements (Crabtree, 1996, p. 73; Holmes, 2014, p. 123–125). That level of resolution for early medieval Scotland is well beyond the capabilities of the current available data. Hoverer, as new datasets emerge, a more comprehensive and nuanced understanding of the Pictish animal economy will be gained. The possibility that smaller hillforts such as Mither Tap and Craig Rock may have restricted age groups, at least for cattle, could indicate high-status consumer sites (Crabtree, 1990, p. 162), perhaps supplemented by local pig rearing. A large fort such as Burghead would certainly have had the capacity to keep some livestock, and some local production cannot be excluded considering the greater variety of age groups present at the site, though this may be in part due to a larger sample. The high-status nature of this major power center would certainly make it an ideal candidate for a consumer or a producer-consumer site. Certainly, the extent of local cattle rearing around Burghead is nebulous and cattle were possibly used as currency and received as tribute payment to elites, as in early medieval Ireland (Kelly, 2016, p. 44) where a dairy cow, pregnant or with a calf, commanded a higher value (McCormick, 2014, p. 121). Burghead would have been in a position to receive tax payments in the form of cattle from its hinterland or the wider region it had influence on. Determining the geographical provenience of livestock, through strontium isotope analysis for example (Britton, 2020), will be key to better understanding the geographical “catchment” of power centers such as Burghead.
While the current study confirms that cattle were the predominant domestic animal in Pictland, future excavations at these three sites (or at others) are needed to yield the more expansive datasets to support the initial findings and synthesis outlined here. Bigger samples are necessary (Crabtree, 2018) to explore, for example, potential differences between larger lowland and coastal sites and hillforts in terms of the economic context they operated in. Although the findings here are a welcome addition to the thin body of evidence, husbandry practices and herd management are also still poorly understood. For example, while there is currently no evidence of specialized milk production based on age data from any of the sites, this does not mean that dairying did not occur. In addition to increasing the size of assemblages through further excavations, isotopic approaches, such as oxygen isotope analysis, combined with intra-tooth sampling, could be used to explore cattle birth seasonality and dairying. Such approaches have identified multiple birth seasons at first millennium AD Pool in Orkney, implying the economic importance of ongoing provision of fresh milk (Towers et al., 2017). It is plausible, perhaps, that in Pictland, as in Iron Age north-west Europe, cattle were part of a mixed farming subsistence strategy where small-scale dairying was undertaken for local production ensuring year-round availability of milk by increasing the birthing season (Groot et al., 2021, p. 14). Age and isotopic data can be integrated with sex ratios, inferred from morphological traits and biometric study or ancient DNA analysis (Svensson et al., 2008; Ruscillo, 2014) to further explore the prevalence and intensity of dairying. An ongoing pilot study has successfully extracted aDNA from Burghead and Mither Tap, showing good preservation of DNA in the faunal assemblages (Girdland-Flink, unpublished data), which confirms this could be a fruitful line of enquiry.
Stable isotope analyses and other specialized approaches may also be pivotal in better understanding other aspects of animal husbandry in Pictland, such as grazing and foddering strategies. Carbon and nitrogen isotope data from faunal bone collagen from the monastic site of Portmahomack, for example, have highlighted that salt-marsh grazing may have been practiced in the management of sheep (Curtis-Summers et al., 2014, 2020). Unpublished stable isotope data from Burghead sheep suggest that this may have also been the case at the site (Czére et al., 2021), although these coastal sites are not representative of how sheep may have been raised in more inland or central areas. Further isotopic studies are required to not only explore salt-marsh grazing but also other strategies such as leaf-foddering or even sea-weed grazing of ungulates and the provisioning of other domesticates such as dogs. Preliminary analysis of pigs, for example, suggest a spread in carbon isotopic data which could be indicative of a range of diets, including some animals feeding in wooded environments (Fawcett, Czére and Britton, unpublished data). This could suggest a diversity of managements strategies (e.g., “backyard”, panage), though the occasional exploitation of wild boar cannot be excluded. The analysis of microbotanical residues, such as phytoliths and starch grains, extracted from external dental surfaces can also be used to explore the plants an animal consumes before death (Middleton and Rovner, 1994; Weber and Price, 2016). A pilot study has successfully extracted phytoliths (grass, arboreal, and dicot) and starch grains (barley and oat) from cattle, pig and caprines molars from Burghead, Mither Tap, and Craig Rock (Prado, unpublished data) demonstrating the significant potential of microbotanical residues for understanding livestock feeding regimes in Pictland. Beyond biomolecular analysis, the recovery and study of larger faunal assemblages must be an ongoing priority. If more comprehensively excavated, Burghead could produce the largest faunal assemblage for the early medieval period in Scotland. While the preservation of animal bone was poorer at Mither Tap and Craig Rock, there appears to have been significant accumulation of bone at the foot of ramparts creating an alkaline environment conducive to bone preservation. The discard of large quantities of bone at the foot of ramparts also raises important social aspects for the construction of these deposits. In Ireland, for example, the symbolic value of settlement enclosures made them a location of choice for discarding food waste, particularly from feasts (O'Sullivan and Nicholl, 2011). It may be that the deposition of faunal materials at these high-status sites in north-east Scotland was also a conscious decision, reflecting their role as power centers, as opposed to a reflection of expediency in waste management. A social zooarchaeology of Pictland, incorporating biomolecular analysis, historical sources and animal depictions on Pictish symbol stones, may help to disentangle both the economic and more esoteric aspects of Pictish human-animal relationships.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
EM-M designed the study (with GN), undertook the analysis, and wrote the initial draft of the manuscript, with input from KB and GN. EF, VL, and SN contributed to the analysis and provided critical reviews of the manuscript. GN, JO'D, EM-M, and SN led the excavations of the sites presented here and provided critical review of and contributions to subsequent versions of the manuscript. KB, JF, and OC were undertaking isotope analysis of the material presented here and provided critical review of and contributions to subsequent versions of the manuscript. SP was undertaking ongoing analysis of microbotanical remains and provided critical review of and contributions to subsequent versions of the manuscript. L-GF was undertaking ongoing analysis of aDNA and contributed to this manuscript. All authors provided critical review of and contributions to subsequent versions of the manuscript.
This research was part of the Northern Picts Project (2009–2015), The Comparative Kingship Project (since 2017) funded by the Leverhulme Trust as part of a Research Leadership Award under Grant RL-2016-069, and Historic Environment Scotland Citadel project—code is RG15531-10. KB was supported by the Leverhulme Trust (PLP-2019–284).
We wish to thank the two reviewers for their constructive comments that have improved the original version of this paper. We thank the University of Aberdeen students and volunteer excavators who participated in the excavations and the onsite collection of samples as well as helping clean and sort the faunal material. We also wish to thank Zena Timmons and Jerry Herman (National Museum of Scotland) for access to NMS collections and for their time. Thanks to funding from Don and Elizabeth Cruickshank that allowed many of the excavations referenced here to take place and enabled publication of results.
EF was employed by AOC Archaeology Group.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) KB declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2023.1208908/full#supplementary-material
Supplementary Table 1. Cattle long bone fusion data at Burghead.
Supplementary Table 2. Distribution of pig mandibles and isolated M3s by age groups following Lemoine et al. (2014).
Supplementary Figure 1. Cattle element representation at Burghead (%MAU).
Supplementary Figure 2. Cattle survival rates based on long bone fusion at Burghead.
Atkinson, D. (2007). Mither Tap Fort, Bennachie, Aberdeenshire (Oyne Parish), watching brief, radiocarbon dating. Discovery and Excavation in Scotland, 8.
Balasse, M., Tresset, A., Bocherens, H., Mariotti, A., and Vigne, J.-D. (2000). “Un abattage “post-lactation” sur des bovins domestiques néolithiques. Étude isotopique des restes osseux du site de Bercy (Paris, France),” in Bassano B., Giacobini G. and Peracino V. (eds), La gestion démographique des animaux à travers le temps. Actes du VIème colloque international de l'association “L'Homme et l'Animal. Société de Recherche Inter-disciplinaire”, Turin, 16-18 septembre 1998. Anthropozoologica 31, 39–48.
Behrensmeyer, A. K. (1978). Taphonomic and ecologic information from bone weathering. Paleobiology 4, 150–162. doi: 10.1017/S0094837300005820
Bond, J. M. (1996). Burnt offerings: animal bone in Anglo-Saxon cremations. World Archaeol. 28, 76–88. doi: 10.1080/00438243.1996.9980332
Britton, K. (2020). “Isotope analysis for mobility and climate studies,” in Archaeological Science: An Introduction, eds K. Britton, and M. P. Richards (Cambridge: Cambridge University Press), 99–124. doi: 10.1017/9781139013826.005
Cannon, D. Y. (1987). Marine Fish Osteology: A Manual for Archaeologists. Burnaby, BC: Simon Fraser University.
Carver, M., Garner-Lahire, J., and Spall, C. (2016). Portmahomack on Tarbat Ness: Changing Ideologies in North-east Scotland, Sixth to sixteenth Century AD. Edinburgh: Society of Antiquaries of Scotland. doi: 10.1515/9780748699971
Castillo, J. A. Q. (2014). Agrarian archaeology in early medieval Europe. Quat. Int. 346, 1–6. doi: 10.1016/j.quaint.2014.08.007
Cohen, A., and Serjeantson, D. (1996). A Manual for the Identification of Bird Bones from Archaeological Sites. London: Archetype Publications.
Crabtree, P. J. (1990). Zooarchaeology and complex societies: some uses of faunal analysis for the study of trade, social status, and ethnicity. Archaeol. Method Theory 2, 155–205.
Crabtree, P. J. (1995). “The symbolic role of animals in anglo-saxon england: evidence from burials and cremations,” in The Symbolic Role of Animals in Archaeology, eds. K. Ryan, and P. J. Crabtree (Philadelphia, PA: University of Pennsylvania Press), 21–26. doi: 10.9783/9781949057003-003
Crabtree, P. J. (1996). Production and consumption in an early complex society: animal use in Middle Saxon East Anglia. World Archaeol. 28, 58–75. doi: 10.1080/00438243.1996.9980331
Crabtree, P. J. (2018). “The value in studying large faunal collections using traditional zooarchaeological methods: a case study from Anglo-Saxon England,” in Zooarchaeology in Practice: Case Studies in Methodology and Interpretation in Archaeofaunal Analysis, eds C. M. Giovas, and M. J. LeFebvre (Berlin: Springer), 173–188. doi: 10.1007/978-3-319-64763-0_9
Crone, A., Hindmarch, E., and Woolf, A. (2016). Living and Dying at Auldhame, East Lothian: The Excavation of an Anglian Monastic Settlement and Medieval Parish Church. Edinburgh: Society of Antiquaries of Scotland.
Curtis-Summers, S., Janet, M., and Carver, M. (2014). Stable isotope evidence for dietary contrast between pictish and medieval populations at Portmahomack, Scotland. Mediev. Archaeol. 58, 21–43. doi: 10.1179/0076609714Z.00000000030
Curtis-Summers, S., Pearson, J. A., and Lamb, A. L. (2020). From picts to parish: stable isotope evidence of dietary change at medieval Portmahomack, Scotland. J. Archaeol. Sci. Rep. 31, 102303. doi: 10.1016/j.jasrep.2020.102303
Czére, O., Müldner, G., Hall, M., Will, B., Mitchell, J., Noble, G., et al. (2021). Multi-isotope analysis of the human skeletal remains from Blair Atholl, Perth and Kinross, Scotland. Tayside Fife Archaeol. J. 27, 31–44.
Dockrill, S. J., Bond, J. M., Turner, V. E., Brown, L. D., Bashford, D. J., Cussans, J. E., et al. (2010). Excavations at Old Scatness, Shetland. Volume 1: The Pictish Village and Viking Settlement. Lerwick: Shetland Heritage Publications.
Driver, J. C. (2011). Identification, classification and zooarchaeology. Ethnobiol. Lett. 2, 19–39. doi: 10.14237/ebl.2.2011.32
Gidney, L. (2018). “The animal in late Medieval Britain,” in The Oxford Handbook of Later Medieval Archaeology in Britain, eds C. Gerrard, and A. Gutiérrez (Oxford: Oxford University Press), 102–115. doi: 10.1093/oxfordhb/9780198744719.013.65
Gifford-Gonzalez, D. (2018). An Introduction to Zooarchaeology. Berlin: Springer International Publishing. doi: 10.1007/978-3-319-65682-3
Gillis, R. E., Gaastra, J. S., Linden, M. V., and Vigne, J.-D. (2022). A species specific investigation into sheep and goat husbandry during the early European Neolithic. Environ. Archaeol. 27, 8–19. doi: 10.1080/14614103.2019.1615214
Grant, A. (1982). “The use of tooth wear as a guide to the age of domestic animals,” in Ageing and Sexing Animal Bones from Archaeological Sites BAR British series 109, eds B. Wilson, C. Grigson, and S. Payne (Oxford: Archaeopress), 91–108.
Greenfield, H. J. (2006). “Sexing fragmentary ungulate acetabulae,” in Recent Advances in Ageingand Sexing Animal Bones. Proceedings of the 9th Conference of the International Council of Archaeozoology, Durham, August 2002, ed D. Ruscillo (Oxford: Oxbow), 68–86.
Greenfield, H. J., and Arnold, E. R. (2008). Absolute age and tooth eruption and wear sequences in sheep and goat: determining age-at-death in zooarchaeology using a modern control sample. J. Archaeol. Sci. 35, 836–849. doi: 10.1016/j.jas.2007.06.003
Greig, C., Greig, M., Ashmore, P., Campbell-Wilson, M., Lorimer, D., Smart, I. H. M., et al. (2002). Excavation of a cairn cemetery at Lundin Links, Fife, in 1965-6. Proc. Soc. Antiq. Scotl. 130, 585–636. doi: 10.9750/PSAS.130.585.636
Groot, M., Albarella, U., Eger, J., and Evans, J. (2021). Cattle management in an Iron Age/Roman settlement in the Netherlands: archaeozoological and stable isotope analysis. PLoS ONE 16, e0258234. doi: 10.1371/journal.pone.0258234
Halstead, P. (1985). “A study of mandibular teeth from Romano-British contexts at Maxey,” in Archaeology and Environment in the Lower Welland Valley East Anglian Archaeology, eds F. Pryor, and C. French (Norwich: East Anglian Archaeology), 219–224.
Halstead, P., and Isaakidou, V. (2017). “Calf mortality and milking: was Tony Legge right after all?,” in Economic Zooarchaeology: Studies in Hunting, Herding and Early Agriculture, eds P. Rowley-Conwy, D. Serjeantson, and P. Halstead (Oxford: Oxbow Books), 187–195.
Holmes, M. (2014). Animals in Saxon and Scandinavian England: Backbones of Economy and Society. Leiden: Sidestone Press.
Holmes, M. (2017). Southern England: A Review of Animal Remains from Saxon, Medieval and Post-Medieval Archaeological Sites. Portsmouth: Historic England.
Holmes, M., Hamerow, H., and Thomas, R. (2021). Close companions? A zooarchaeological study of the human–cattle relationship in medieval England. Animals 11, 1174. doi: 10.3390/ani11041174
Jones, G. G., and Sadler, P. (2012). Age at death in cattle: methods, older cattle and known-age reference material. Environ. Archaeol. 17, 11–28. doi: 10.1179/1461410312Z.0000000002
Kamjan, S., de Groene, D., van den Hurk, Y., Zidarov, P., Elenski, N., Patterson, W. P., et al. (2021). The emergence and evolution of Neolithic cattle farming in southeastern Europe: New zooarchaeological and stable isotope data from DŽuljunica-Smărdeš, in northeastern Bulgaria (ca. 6200–5500 cal. BCE). J. Archaeol. Sci. Rep. 36, 102789. doi: 10.1016/j.jasrep.2021.102789
Kelly, F. (2016). “Cattle in ancient Ireland: early Irish legal aspects,” in Cattle in Ancient and Modern Ireland, eds M. O'Connell, F. Kelley, and J. H. McAdam (Newcastle upon Tyne: Cambridge Scholars Publishing), 44–50.
Lemoine, X., Zeder, M. A., Bishop, K. J., and Rufolo, S. J. (2014). A new system for computing dentition-based age profiles in Sus scrofa. J. Archaeol. Sci. 47, 179–193. doi: 10.1016/j.jas.2014.04.002
Lock, G., and Ralston, I. (2017). Mapping hillforts in Britain and Ireland. Br. Archaeol. 157, 46–51.
Madgwick, R., Grimes, V., Lamb, A. L., Nederbragt, A. J., Evans, J. A., and McCormick, F. (2019). Feasting and mobility in iron age Ireland: multi-isotope analysis reveals the vast catchment of Navan Fort, Ulster. Sci. Rep. 9, 19792. doi: 10.1038/s41598-019-55671-0
McCormick, F. (1992). Early faunal evidence for dairying. Oxford J. Archaeol. 11, 201–210. doi: 10.1111/j.1468-0092.1992.tb00264.x
McCormick, F. (2008). The decline of the cow: agricultural and settlement change in early medieval Ireland. Peritia 20, 209–224. doi: 10.1484/J.Peri.3.632
McCormick, F. (2014). Agriculture, settlement and society in early Medieval Ireland. Quat. Int. 346, 119–130. doi: 10.1016/j.quaint.2013.10.040
McCormick, F., Kerr, T., McClatchie, M., and O’Sullivan, A. (2011). “The archaeology of livestock and cereal production in early Medieval Ireland, AD 400– 1100.” in Early Medieval Archaeology Project (EMAP 2) Report 5:1. Irish National Strategic Archaeological Research (INSTAR) programme 2011.
McCormick, F., and Murray, E. (2017). “The zooarchaeology of Medieval Ireland,” in The Oxford Handbook of Zooarchaeology, eds U. Albarella, M. Rizzetto, H. Russ, K. Vickers, and S. Viner-Daniels (Oxford: Oxford University Press), 195–213. doi: 10.1093/oxfordhb/9780199686476.013.15
Middleton, W. D., and Rovner, I. (1994). Extraction of opal phytoliths from herbivore dental calculus. J. Archaeol. Sci. 21, 469–473. doi: 10.1006/jasc.1994.1046
Mulville, J., Bond, J. M., and Oliver, C. (2005). “The white stuff: milking in the outer Scottish Isles,” in The Zooarchaeology of Fats, Oils, Milk and Dairying. Proceedings of the 9th Conference of the International Council of Archaeozoology, eds J. Mulville, and A. K. Outram (Oxford: Oxbow Books), 167–182.
Murray, E., McCormick, F., and Plunkett, G. (2004). The food economies of Atlantic island monasteries: the documentary and archaeo-environmental evidence. Environ. Archaeol. 9, 179–188. doi: 10.1179/env.2004.9.2.179
Noble, G. (2020). The problem with the Picts. Searching for a lost people in northern Scotland. Curr. Archaeol. 364, 28–35.
Noble, G., Cruickshanks, G., Dunbar, L., Evans, N., Hall, D., Hamilton, D., et al. (2019a). Kinneddar: a major ecclesiastical centre of the picts. Proc. Soc. Antiq. Scotl. 148, 113–145. doi: 10.9750/PSAS.148.1271
Noble, G., Evans, N., and Campbell, E. (2019b). The King in the North: The Pictish Realms of Fortriu and Ce: Collected Essays Written as Part of the University of Aberdeen's Northern Picts Project. Edinburgh: Birlinn Ltd.
Noble, G., Evans, N., Hamilton, D., MacIver, C., Masson-MacLean, E., and O'Driscoll, J. (2020). Dunnicaer, Aberdeenshire, Scotland: a roman iron age promontory fort beyond the frontier. Archaeol. J. 177, 256–338. doi: 10.1080/00665983.2020.1724050
Noble, G., Gondek, M., Campbell, E., and Cook, M. (2013). Between prehistory and history: the archaeological detection of social change among the picts. Antiquity 87, 1136–1150. doi: 10.1017/S0003598X00049917
Noble, G., Gondek, M., Campbell, E., Evans, N., Hamilton, D., and Taylor, S. (2019c). A powerful place of pictland: interdisciplinary perspectives on a power centre of the 4th to 6th centuries AD. Mediev. Archaeol. 63, 56–94. doi: 10.1080/00766097.2019.1588529
Noble, G., and O'Driscoll, J. (2019). “Fortified settlement in early medieval Northern Britain and Ireland,” in Hillforts: Britain, Ireland and the Nearer Continent. Papers from the Atlas of Hillforts of Britain and Ireland Conference, June 2017, eds. G. Lock, and I. Ralston (Oxford: Archaeopress), 97–116. doi: 10.2307/j.ctvnb7r0b.13
O'Connor, T. (2011). “Animal husbandry,” in The Oxford Handbook of Anglo-Saxon Archaeology, eds D. A. Hinton, S. Crawford, and H. Hamerow (Oxford: Oxford University Press), 361–376.
O'Connor, T. (2014). Livestock and animal husbandry in early medieval England. Quat. Int. 346, 109–118. doi: 10.1016/j.quaint.2013.09.019
O'Connor, T. P. (2003). The Analysis of Urban Animal Bone Assemblages: A Handbook for Archaeologists. London: The Council for British archaeology.
Oram, R. (2007). “Capital tales or Burghead bull,” in A Grey Eye Looks Back: A Festschrift in Honour of Colm Ó Baoill, eds S. Arbuthnot, and K. Hollo (Perthshire: Clann Tuirc), 241–262.
O'Sullivan, A., and Nicholl, T. (2011). Early medieval settlement enclosures in Ireland: dwellings, daily life and social identity. Proc. R Ir. Acad. C Archaeol. Celt. Stud. Hist. Linguist. Lit. 111C, 59–90. doi: 10.1353/ria.2011.a810654
Payne, S. (1973). Kill-off patterns in sheep and goats: the mandibles from Asvan Kale. Anatol. Stud. 23, 281–303. doi: 10.2307/3642547
Perry, D. R. (2000). Castle Park, Dunbar: Two Thousand Years on a Fortified Headland. Edinburgh: Society Antiquaries Scotland.
Ralston, I. (2004). The Hill-forts of Pictland since the Problem of the Picts. Fortrose: Groam House Museum.
RCAHMS (2007). In the Shadow of Bennachie: A Field Archaeology of Donside, Aberdeenshire. Edinburgh: Royal Commission on the Ancient and Historical Monuments of Scotland and Society of Antiquaries of Scotland.
Reitz, E. J., and Wing, E. S. (2008). Zooarchaeology. Cambridge: Cambridge University Press. doi: 10.1017/CBO9780511841354
Ruscillo, D. (2014). “Zooarchaeology: methods of collecting age and sex data,” in Encyclopedia of Global Archaeology, ed. C. Smith (Berlin: Springer), 8000–8010. doi: 10.1007/978-1-4419-0465-2_2163
Schmid, E. (1972). Atlas of Animal Bones: for Prehistorians, Archaeologists and Quaternary Geologists. Amsterdam: Elsevier.
Silver, I. A. (1969). “The ageing of domestic animals,” in Science in Archaeology: A Survey of Progress and Research, eds. D. R. Brothwell, E. S. Higgs, and G. Clark (New York, NY: Basic Books, Inc., Publishers), 283–302.
Smith, C. (2000). A grumphie in the sty: an archaeological of pigs in Scotland, from their earliest domestication to the agricultural revolution. Proc. Soc. Antiq. Scotl. 130, 705–724. doi: 10.9750/PSAS.130.705.724
Svensson, E. M., Götherström, A., and Vretemark, M. (2008). A DNA test for sex identification in cattle confirms osteometric results. J. Archaeol. Sci. 35, 942–946. doi: 10.1016/j.jas.2007.06.021
Towers, J., Mainland, I., Montgomery, J., and Bond, J. (2017). Calving seasonality at Pool, Orkney during the first millennium AD: an investigation using intra-tooth isotope ratio analysis of cattle molar enamel. Environ. Archaeol. 22, 40–55. doi: 10.1080/14614103.2015.1116214
Watt, J., Pierce, G. J., and Boyle, P. R. (1997). Guide to the Identification of North Sea Fish Using Prermaxillae and Vertebrae. Copenhagen: International Council for the Exploration of the Sea.
Weber, S., and Price, M. D. (2016). What the pig ate: a microbotanical study of pig dental calculus from 10th−3rd millennium BC northern Mesopotamia. J. Archaeol. Sci. Rep. 6, 819–827. doi: 10.1016/j.jasrep.2015.11.016
Woolf, A. (2006). Dén Nechtain, fortriu and the geography of the picts. Scott. Hist. Rev. 85, 182–201. doi: 10.3366/shr.2007.0029
Young, H. W. (1890). The ancient bath at Burghead, with remarks on its origin, as shewn by existing baths of the same shape and design. Proc. Soc. Antiq. Scotl. 24, 147–156. doi: 10.9750/PSAS.024.147.156
Young, H. W. (1891). Notes on the ramparts of Burghead, as revealed by recent excavations. Proc. Soc. Antiq. Scotl. 25, 435–447. doi: 10.9750/PSAS.025.435.447
Keywords: animal remains, agriculture, pastoralism, subsistence, diet, economy
Citation: Masson-MacLean E, Niehaus S, Fraser E, Lee V, Prado S, Czére O, Fawcett J, O'Driscoll J, Girdland-Flink L, Noble G and Britton K (2023) New zooarchaeological evidence from Pictish sites in Scotland: implications for early medieval economies and animal-human relationships. Front. Environ. Archaeol. 2:1208908. doi: 10.3389/fearc.2023.1208908
Received: 19 April 2023; Accepted: 24 July 2023;
Published: 14 August 2023.
Edited by:
Nimrod Marom, University of Haifa, IsraelReviewed by:
Peter Rowley-Conwy, Durham University, United KingdomCopyright © 2023 Masson-MacLean, Niehaus, Fraser, Lee, Prado, Czére, Fawcett, O'Driscoll, Girdland-Flink, Noble and Britton. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Edouard Masson-MacLean, ZWRvdWFyZC5tYXNzb24tbWFjbGVhbkBhYmRuLmFjLnVr
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.