Abstract
Background:
In the cardiovascular (CV) system, overactivation of the angiotensin converting enzyme (ACE) may trigger deleterious responses derived from angiotensin (Ang)-II, which can be attenuated by stimulation of ACE2 and subsequent Ang-(1-7) metabolite. However, ACE2 exhibits a high degree of genetic polymorphism that may affect its structure and stability, interfering with these cardioprotective actions. The aim of this study was to analyse the relationship of ACE2 polymorphisms with cardiovascular risk factors in children.
Methodology:
Five ACE2-single nucleotide polymorphisms (SNP), rs4646188, rs2158083, rs233575, rs879922, and rs2074192, previously related to CV risk factors, were analyzed in a representative sample of 12–16-year-old children and tested for their potential association with anthropometric parameters, insulin levels and the lipid profile.
Results:
Girls (N = 461) exhibited lower rates of overweight, obesity, blood pressure, and glycemia than boys (N = 412), though increased plasma lipids. The triglycerides (TG)/HDL-C ratio was, however, lower in females. Interestingly, only in girls, the occurrence of overweight/obesity was associated with the SNPs rs879922 [OR 1.67 (1.02–2.75)], rs233575 [OR 1.98 (1.21- 3.22)] and rs2158083 [OR 1.67 (1.04–2.68)]. Also, TG levels were linked to the rs879922, rs233575, and rs2158083 SNPs, and the TG/HDL-C ratio was associated with rs879922 and rs233575. Levels of TC and LDL-C were associated with rs2074192 and rs2158083. Furthermore, the established cut-off level for TG ≥ 90 mg/dL was related to rs879922 [OR 1.78 (1.06–2.96)], rs2158083 [OR 1.75 (1.08–2.82)], and rs233575 [OR 1.62 (1.00–2.61)]. The cut-off level for TC ≥ 170 mg/dL was associated with rs2074192 OR 1.54 (1.04–2.28) and rs2158083 [OR 1.53 (1.04–2.25)]. Additionally, the haplotype (C-G-C) derived from rs879922-rs2158083-rs233575 was related to higher prevalence of overweight/obesity and TG elevation.
Conclusion:
The expression and activity of ACE2 may be essential for CV homeostasis. Interestingly, the ACE2-SNPs rs879922, rs233575, rs2158083 and rs2074192, and the haplotype (C-G-C) of the three former could induce vulnerability to obesity and hyperlipidemia in women. Thus, these SNPs might be used as predictive biomarkers for CV diseases and as molecular targets for CV therapy.
Introduction
The Angiotensin converting enzyme-2 (ACE2) is a significant regulator of the Renin-Angiotensin-Aldosterone system (RAAS), which plays key roles in the control of cardiovascular (CV) system (1). In the canonical pathway, the RAAS precursor, angiotensinogen, is degraded to the octapeptide angiotensin-II (Ang-II) by consecutive digestions of renin and angiotensin convertase enzyme (ACE). Ang-II could be then converted by aminopeptidase-A to Ang-III, which conserves similar proprieties mostly mediated by two distinct G protein-coupled receptors named AT1R and AT2R (2). Overactivation of the ACE/Ang-II/AT1R pathway has been associated with several CV pathologies, including hypertension, heart failure, vascular inflammation and remodeling, coagulation, and atherosclerosis (3). However, activation of AT2R can trigger anti-inflammation and vasodilation. In this regard, the stimulation of the non-canonical axis of the RAAS may compensate ACE/Ang-II/AT1R actions. The mono-carboxypeptidase ACE2 can convert Ang-II into angiotensin 1–7 [Ang (1-7)], which binds to the Mas receptor (MasR) (1). The ACE2/Ang (1-7)/MasR axis can reduce blood pressure and CV hypertrophy and fibrosis, stimulating vasoactive prostaglandins and lessening redox imbalance (4). Ang (1-7) also protects ACE-degradation of bradykinins and ameliorates inflammatory responses, and vascular permeability (1). Moreover, ACE2 also degrades Ang l into Ang (1–9), decreasing Ang-II levels. Given the opposite effects of Ang (1-7) and Ang-II in the CV homeostasis, it might be imperative to maintain a minimum ACE2 level or activity.
Interestingly, the ACE2 gene is highly conserved in mammals and expressed in testes, renal and CV system, and especially in gastrointestinal tissues (5). It encodes a type I membrane-bounding glycoprotein composed by 805 amino acids divided in three functional domains including a C-terminal transmembrane anchoring region, a N-terminal signal peptide motif, and an HEXXH zinc-binding metalloprotease region (Figure 1). ACE2 maps in chromosome Xp22 and contain 17 introns and 18 exons, where a high degree of genetic polymorphism can be found (6). There are at least 510 valid variants in ACE2 (mostly intronic regions) (7), and some single nucleotide polymorphisms (SNP) may affect ACE2 structure and stability, interfering with its cardioprotective functions. In adults with different gender, ethnic and presence of pre-existing CV disease, the rs4646188, rs2158083, rs233575, rs879922, and rs2074192 SNPs (Figure 1) have been related to hypertension, ventricular hypertrophy, or type-2 diabetes mellitus (T2DM), and increased plasma total cholesterol (TC), triglycerides (TG), low-density lipoprotein-cholesterol (LDL-C), or reduced high-density lipoprotein-cholesterol (HDL-C) and Ang (1-7) (8–11). In children or adolescent cohorts, where existence of confusing factors (i.e., epigenetic alterations) is lower, boys but not girls showed association of rs2158083, rs233575 and rs2074192 with hypertension (12, 13). Nowadays, regulation of the ACE2 expression and activity has become essential for Coronavirus disease (COVID)-19 subjects, particularly for those with cardiovascular complications (6, 14). Thus, further studies analyzing ACE2-SNPs and related haplotypes in young populations, could suggest predictive markers for CV diseases and molecular targets for CV therapy.
Figure 1
In our study, we examine five ACE2 SNPs in relationship with the presence of overweight and obesity in a representative sample of 12-to 16-year-old children, and we assess the potential association of the polymorphisms with the lipid profile and insulin levels.
Methodology
Subjects
The present study comprised 873 adolescents (11–17 years old) recruited in the “Four Provinces study” during 2004–08 in whom information on biochemical variables and DNA samples were available (15). This cross-sectional study was designed to analyze CV risk factors in general population of adolescents from Spain (461 girls and 412 boys). Children suffering from any endocrine, metabolic, hepatic, or renal disorder were excluded from the study. Specialized physicians and nurses oversaw the anthropometric characterization and blood extractions. The investigation fulfills the principles contained in the Declaration of Helsinki and subsequent reviews, as well as the prevailing Spanish legislation on clinical research in human subjects. Parents were required to sign a written consent form allowing their children to participate. The study protocol was approved by the Ethics Committee of Clinical Investigation of the Fundación Jiménez Díaz (Code number: PIC016-2019 FJD).
Variables of the Study
Anthropometric and Blood Pressure Measurements
The weight and height were taken in barefoot children, wearing light clothing. Both weight and height were approached to the nearest 0.1 unit (kg or cm, respectively) and were used to estimate the body mass index (BMI), as kilograms divided by height in squared meters (kg/m2). The age- and sex-specific cut-off points of BMI for overweight and obesity were established according to Cole et al. (16). In addition, the blood pressure (systolic and diastolic) was quantified in subjects as average of two measurements in the right arm by using a mercury sphygmomanometer and after 15 min resting in seated position.
Plasma Parameters
Blood samples were obtained from subjects in the morning after a 12-h fasting period by using EDTA-collection tubes (Vacutainer, BD). Plasma was separated by centrifuging blood at 1,500 g at 4°C for 20 min. The upper fraction was stored at −80°C until use. Total cholesterol and triglycerides were enzymatically quantified in a RA-1000 Autoanalyzer (Technicon, Luton, UK). HDL-C concentrations were measured after precipitation of apo B-containing lipoproteins with phosphotungstic acid and Mg++ (Boehringer Mannheim, Baden-Wurttemberg, Germany). LDL-C levels were calculated according to Friedewald's formula. Also, the glucose concentration was evaluated by the glucose oxidase method, and insulin levels were achieved by using a RIA commercial kit (BI-Insulin IRMA, Bio-Rad, France). To estimate insulin resistance, the homeostatic model assessment of insulin resistance (HOMA-IR) index was calculated following formula: fasting insulin (μU/mL) x fasting glucose (mmol/liter)/22.5.
Single Nucleotide Polymorphism (SNPs) Determinations
Genomic DNA was also obtained from blood samples. After plasma separation, the interphasic fraction (leukocytes) was used to isolate total DNA by adaptation of a classic procedure, which involves salting out of the cellular proteins by dehydration and precipitation with a saturated NaCl solution (17). The quantity and quality of DNA was assessed by UV-spectrophotometry using the Nanodrop spectrophotometer ND-1000. Five SNPs located in intronic sequences of the ACE2 gene (A/G [rs4646188], G/C [rs879922], A/G [rs233575], C/T [rs2074192] and T/C [rs2158083]) were genotyped by using predesigned TaqMan® SNP Genotyping Assays from Applied Biosystems (C_32336232_10, C_8877953_10, C_2260459_10, C_16163821_10 and C_16141434_10, respectively). A StepOnePlus™ Real-Time PCR System (Applied Biosystems) was used for allelic discrimination. PCR was performed with a mixture of 10 ng of genomic DNA, TaqMan® SNP Genotyping Assay (20X), and TaqPath™ ProAmp™ Master Mix (Applied Biosystems). Samples were cycled under the recommended conditions: 95 °C for 10 min, 95 °C for 15 s and 60 °C for 1 min, repeated over 40 cycles. The data presented in the study are deposited in the dbSNP repository, submission batch ID 1063361 (dbSNP Build ID: B156). The genotyping data were upload in a public repository (accession number).
Haplotype Analysis
The five SNPs (rs4646188, rs879922, rs233575, rs2074192, and rs2158083) were selected to construct haplotypes. Linear generalized models were used to compare outcomes levels between haplotypes. The comparisons were summarized by coefficients and odds ratios for continuous and categorical outcomes respectively, and with 95% confidence intervals and p-values. These analyses were performed using the haplo.glm function from the haplo.stats R package. Haplotypes with frequencies less than 5% were included in the category “other”.
Statistical Analysis
Qualitative variables were included as absolute and relative frequencies. Associations between qualitative variables were studied by the chi-square test and odds ratio. On the other hand, normality of quantitative variables was analyzed by the Kolmogorov–Smirnov test. On one hand, variables with normal distribution (age and HDL-C) were compared using a Student's t-test and ANOVA one way, followed by Tukey (assuming equal variances) or Games-Howell (equal variances not assumed) post-hoc test. Also, they were summarized as mean values and 95% confidence interval (CI). Some variables with non-normal distribution were normalized by log base-10 transformation [systolic (SBP) and diastolic (DBP) blood pressures, TC, TG, LDL-C, TG/HDL] and were expressed as geometric mean values and 95% confidence interval. Other variables with non-normal distribution (glucose, insulin, and HOMA-IR) were compared using the Mann–Whitney U test and Kruskal-Wallis test followed by Dunn's (post-hoc). These variables were summarized by median and interquartile range. The associations of haplotypes with anthropometric, blood pressure and plasma parameters were analyzed by regression models. For continuous variables, we used linear models, whereas dichotomous coded variables were examined by logistic models. The former were summarized by coefficients (coef.) while the later were summarized by odds ratio (OR). Also, a 95% CI and p-values were reported. Statistical analyses were performed using the statistical language R version 4.0.5 (R Foundation for Statistical Computing, Vienna, Austria).
Results
Characterization of the Population
We analyzed a population of 873 adolescents aged 13.9 years with a mean BMI of 21.5 kg/m2. In the study, we found that 71.8 percent of children were normo-weight, whereas 28.2 percent presented an excessive weight (Table 1). The prevalence of obesity in girls and boys was 4.3 and 7.6 %, respectively. The mean BMI Z-score was 0.00 (−0.067–0.067) for the global population, and −0.018 (−0.10–0.07) and 0.021 (−0.08–0.12), for girls and boys, respectively (p = 0.569). Also, according to the latest clinical guides of the American College of Cardiology/American Heart Association/American Diabetes Association for pediatric population (18, 19), they were, on average, in the normotensive range and showed unaltered levels of the glycemic and lipid profiles (Table 1). However, further analysis pointed out significant differences between females and males. Females (52.8%) exhibited lower rates of overweight and obesity, SBP and glycemia, but higher levels of TC, LDL-C, and HDL-C than boys (Table 1). The ratio of TG and HDL-C, as a predictor of metabolic syndrome and CV diseases (20), was significantly lower in female individuals.
Table 1
| Variables | Global (N = 873) | Female (N = 461) | Male (N = 412) | P-value |
|---|---|---|---|---|
| Age (years) | 13.9 (13.8–13.9) | 13.9 (13.7–14.0) | 13.9 (13.7–14.0) | 0.873 |
| BMI (kg/m2) | 21.5 (21.3–21.8) | 21.5 (21.1–21.8) | 21.6 (21.2–21.9) | 0.720 |
| NW % (n) | 71.8 (625) | 75.7 (349) | 67.3 (276) | 0.006 |
| OW + Obesity % (n) | 28.2 (246) | 24.3 (112) | 32.7 (134) | 0.006 |
| SBP (mmHg) | 113.9 (113.0–114.7) | 111.5 (110.3–112.6) | 116.6 (115.3–117.8) | <0.001 |
| DBP (mmHg) | 64.7 (64.0–65.4) | 64.2 (63.2–65.2) | 65.4 (64.4–66.3) | 0.099 |
| Glucose mg/dL | 90.5 (10.6) | 89.0 (10.2) | 92.3 (10.4) | <0.001 |
| Insulin (μU/mL) | 7.44 (4.4) | 7.54 (4.28) | 7.32 (4.67) | 0.249 |
| HOMA–IR | 1.65 (1.06) | 1.64 (0.98) | 1.66 (1.14) | 0.867 |
| TC (mg/dL) | 163.9 (162.0–165.8) | 168.1 (165.6–170.7) | 159.2 (156.6–161.9) | <0.001 |
| TG (mg/dL) | 72.3 (70.5–74.0) | 71.7 (69.4–74.0) | 73.0 (70.2–75.8) | 0.477 |
| LDL–C (mg/dL) | 94.9 (93.2–96.6) | 97.3 (94.9–99.7) | 92.3 (89.9–94.7) | 0.004 |
| HDL–C (mg/dL) | 52.5 (51.5–53.5) | 54.6 (53.2–55.9) | 50.2 (48.8–51.7) | <0.001 |
| TG/HDL–C ratio | 1.43 (1.38–1.48) | 1.36 (1.30–1.43) | 1.51 (1.43–1.60) | 0.004 |
Anthropometric and biochemical characterization of a Spanish population of adolescents.
Data are expressed as %, median and interquartile range (IR), mean or geometric mean (CI 95%). Variables with normal distribution (Age, BMI, SBP, DBP, TC, TG, LDL-C, HDL-C and TG/HDL-C ratio) were compared using t-Test, whereas variables with non-normal distribution (Glucose, Insulin, and HOMA-IR) were compared using the Mann–Whitney U test. Also, qualitative variables (NW and OW + Obesity) were studied by the Pearson's chi-square test. In bold, statistically significant data (p < 0.05).
NW and OW, as normal weight and overweight, respectively. HOMA-IR, as Homeostatic Model Assessment of Insulin Resistance. TC, TG, LDL-C and HDL-c, for total cholesterol, triglycerides, low-density lipoprotein-cholesterol, and high-density lipoprotein-cholesterol, respectively.
Genotypic and Allelic Frequencies of ACE2 SNPs
Five single nucleotide polymorphisms (SNPs) of the ACE2 gene previously related to CV injuries (9, 10, 21) were analyzed in our study. The genotype frequencies of rs4646188, rs879922, rs233575, rs2074192, and rs2158083 were found in a Hardy's-Weinberg equilibrium. The prevalence of the minor alleles of the studied SNPs in girls and boys, respectively, were: (G) in rs4646188, 10.1% and 8.4%; (C) in rs879922, 42.4% and 39.3%; (G) in rs233575, 39.2% and 37.3%; (T) in rs2074192, 40.10% and 41.5%, and (C) in rs2158083, 38.45% and 36.3% (Table 2).
Table 2
| ACE2SNPs | Female | Male | |||
|---|---|---|---|---|---|
| Genotypes | n | Allele (%) | n | Allele (%) | |
| frequencies % (n) | |||||
| rs4646188 A/G | AA: 80.7 (359) | 445 | A: 89.9 | 391 | A: 91.6 |
| AG: 18.4 (82) | G: 10.1 | G: 8.4 | |||
| GG: 0.9 (4) | |||||
| rs879922 G/C | GG: 33.2 (144) | 434 | G: 57.6 | 374 | G: 60.7 |
| GC: 48.8 (212) | C: 42.4 | C: 39.3 | |||
| CC: 18.0 (78) | |||||
| rs233575 A/G | AA: 36.8 (164) | 446 | A: 60.8 | 389 | A: 62.7 |
| AG: 48.0 (214) | G: 39.2 | G:37.3 | |||
| GG: 15.2 (68) | |||||
| rs2074192 C/T | CC: 37.0 (164) | 443 | C: 59.9 | 390 | C: 58.5 |
| CT: 45.8 (203) | T: 40.10 | T: 41.5 | |||
| TT: 17.2 (76) | |||||
| rs2158083 T/C | TT: 38.4 (171) | 445 | T: 61.55 | 391 | T: 63.7 |
| TC: 46.3 (206) | C: 38.45 | C: 36.3 | |||
| CC: 15.3 (68) | |||||
Genotype and allele frequencies of ACE2-SNPs.
Five SNPs (rs4646188, rs879922, rs233575, rs2074192, and rs2158083) of the ACE2 gene, related to cardiovascular risk factors, were described in the population of adolescents as absolute and relative frequencies.
Association Between ACE2 SNPs and Overweight and Obesity
We tested whether these five ACE2 SNPs (rs4646188, rs879922, rs233575, rs2074192, and rs2158083) might associate with the anthropomorphic variables of the population. Remarkably, none of the SNPs were significantly related to the SBP or DBP, either in girls or boys (not shown). However, in females, the occurrence of overweight or obesity was found to be significantly associated with three of the studied SNPs (Table 3). In particular, the minor allele C of the rs879922 variant was present in 26.9% of overweight or obese females [OR 1.67 (95% CI: 1.02–2.75), p = 0.042]. Similarly, the G allele of rs233575 and the C allele of rs2158083 were present in 28.0% [OR 1.98 (1.21- 3.22), p = 0.006] and 27.0% [OR 1.67 (1.04–2.68), p = 0.032] of overweight or obese girls, respectively (Table 3). In contrast, the overweight or obesity condition was not related to any SNPs in the male population (Supplementary Table 1). Thus, the presence of rs879922, rs233575, and rs2158083 were associated with overweight and obesity only in adolescent females.
Table 3
| ACE2SNPs | NW % (n) | OW+ | Pearson | OR (95%IC) |
|---|---|---|---|---|
| Obesity % (n) | Chi-Square | |||
| rs4646188 A/G | ||||
| AA | 76.6 (275) | 23.4 (84) | 0.841 | - |
| AG + GG | 75.6 (65) | 24.4.0 (21) | ||
| rs879922 C/G | ||||
| GG | 81.9 (118) | 18.1 (26) | 0.042 | 1.67 (1.02–2.75) |
| GC + CC | 73.1 (212) | 26.9 (78) | ||
| rs233575 A/G | ||||
| AA | 83.5 (137) | 16.5 (27) | 0.006 | 1.98 (1.21–3.22) |
| AG + GG | 72 (203) | 28.0 (79) | ||
| rs2074192 C/T | ||||
| CC | 72.6 (119) | 27.4 (45) | 0.131 | - |
| CT + TT | 78.9 (220) | 21.1 (59) | ||
| rs2158083 T/C | ||||
| TT | 81.9 (140) | 18.1 (31) | 0.032 | 1.67 (1.04–2.68) |
| TC + CC | 73.0 (200) | 27.0 (74) |
Association of the ACE2 rs4646188, rs879922, rs233575, rs2074192, and rs2158083 genotypes with overweight/obesity in female adolescents.
The presence of rs4646188, rs879922, rs233575, rs2074192, and rs2158083 SNPs was analyzed in normo-weight (NW), and in overweight (OW)+obese girls. Data were described as absolute and relative frequencies (%) and associations between variables were analyzed by the Pearson's chi-square test, followed by logistic models. In bold, significant associations between variables (p < 0.05).
Association Between ACE2 SNPs and the Glucose and Lipid Profiles
Further potential relationships between the SNPs and biochemical variables were also assessed. Glycemic parameters such as glycemia, plasma insulin and the HOMA-IR were not significantly associated with the presence of any SNPs in any sex (not shown). Lipid parameters did not show any relationship with any SNPs in male adolescents. However, again in females, four out of five SNPs were related to the lipid profile. The highest levels of TG were significantly associated with the presence of heterozygous genotypes of rs879922 (p = 0.020), rs233575 (p = 0.017) and rs2158083 (p = 0.036) (Table 4). The TG/HDL-C ratio also linked with rs879922 and rs233575. In addition, the established cut-off levels of TG (over 90 mg/dL; 75th percentile) for adolescents (10–19 years-old) (18) were related to rs879922 [OR 1.78 (95% CI: 1.06-2.96)], rs2158083 [OR 1.75 (1.08–2.82)], and rs233575 [OR 1.61 (1.00–2.62)] (Figure 2A). Furthermore, the highest levels of TC and LDL-C were related to the heterozygous genotypes of rs2074192 (p = 0.003 and p = 0.03, respectively) and rs2158083 (p = 0.008 and p = 0.019, respectively) (Table 4). Similarly, the cut-off levels of TC (over 170 mg/dL; 75th percentile), but not that of LDL-C levels, for adolescents (10–19 years-old) (18) were associated with rs2074192 [OR 1.54 (95% CI: 1.04–2.28) and rs2158083 [OR 1.53 (1.04–2.25)] (Figure 2B). The HDL-C levels were, however, independent of these ACE2 SNPs. Therefore, the presence of ACE2 SNPs rs879922, rs233575 and rs2158083 may be useful to predict elevated levels of TG and TC in girls.
Table 4
| ACE2SNPs (n) | TC (mg/dL) | TG (mg/dL) | LDL–C (mg/dL) | HDL–C (mg/dL) | TG/HDL–C ratio |
|---|---|---|---|---|---|
| rs879922 C/G | |||||
| GG (143) | 166.4 (162.4–170.4) | 66.9 (63.2–70.7) | 95.4 (91.6–99.2) | 55.72 (53.5–57.9) | 1.2 (1.2–1.3) |
| GC (209) | 171.5 (167.3–175.7) | 74.0 (70.7–77.6)* | 100.0 (96.3–103.9) | 54.6 (52.6–56.6) | 1.4 (1.3–1.5)* |
| CC (78) | 163.2 (157.4–169.1) | 71.4 (66.7–76.4) | 93.0 (87.8–98.5) | 53.96 (50.3–57.6) | 1.4 (1.3–1.6) |
| rs233575 A/G | |||||
| AA (163) | 166.3 (162.6–170.0) | 67.6 (64.1–71.3) | 95.4 (91.7–98.8) | 55.6 (53.5–57.7) | 1.3 (1.2–1.3) |
| AG (211) | 171.0 (166.3–175.2) | 74.7 (71.3–78.2)* | 100.4 (96.5–104.2) | 54.0 (52.0–55.9) | 1.4 (1.4–1.5)* |
| GG (68) | 163.8 (157.4–170.5) | 71.8 (66.7–77.2) | 93.5 (87.8–99.6) | 53.9 (50.0–57.9) | 1.4 (1.2–1.6) |
| rs2074192 C/T | |||||
| CC (164) | 164.4 (159.9–168.9) | 72.7 (69.2–76.2) | 94.4 (90.4–98.6) | 53.5 (51.2–55.8) | 1.4 (1.3–1.5) |
| CT (200) | 173.4 (169.6–177.3)* | 72.2 (68.8–75.7) | 101.1 (97.6–104.8)* | 55.8 (53.7–57.8) | 1.3 (1.3–1.4) |
| TT (75) | 164.4 (159.4–169.6)# | 66.9 (61.6–72.6) | 95.3 (90.7–100.2) | 54.0 (51.5–56.6) | 1.3 (1.1–1.4) |
| rs2158083 T/C | |||||
| TT (170) | 165.2 (161.4–169.0) | 67.9 (64.7–71.4) | 95.2 (91.9–98.7) | 54.7 (52.7–56.6) | 1.3 (1.2–1.4) |
| TC (203) | 172.6 (168.6–176.8)* | 74.3 (70.8–78.0)* | 101.0 (97.3–104.9) | 54.7 (52.6–56.8) | 1.4 (1.3–1.5) |
| CC (68) | 163.0 (156.7–169.6)# | 72.0 (67.1–77.5) | 92.6 (86.7–98.7)# | 53.8 (50.2–57.4) | 1.4 (1.2–1.6) |
Lipid levels by ACE2 rs4646188, rs879922, rs233575, rs2074192, and rs2158083 genotypes in girls.
The occurrence of rs4646188, rs879922, rs233575, rs2074192, and rs2158083 SNPs was studied in females classified according with the plasma levels of total cholesterol (TC), triglycerides (TG), low-density lipoprotein (LDL-C), high-density lipoprotein (HDL-C) and the TG/HDL-C ratio. Results are expressed as mean or geometric mean (CI 95%).
Variables were compared by using an ANOVA one way test, followed by Tukey (assuming equal variances) or Games-Howell (non-equal variances) post-hoc test. In bold, significant associations between variables (p < 0.05).
p < 0.05 heterozygous vs. major allele.
p < 0.05 heterozygous vs. minor allele.
Figure 2
Association Between ACE2-haplotypes and Overweight/Obesity and TG Levels
Since rs879922, rs233575, rs2074192, and rs2158083 were related to overweight/obesity and lipid alterations in females, we examined whether combinations of these SNPs might also associate with higher risk of both pathologies. In particular, the haplotype composed by the minor alleles of rs879922, rs233575, and rs2158083 (C-G-C) was overrepresented in girls (34%) and linked to significantly higher BMI [coef. 0.01 (0.001, 0.019; 95% CI), p = 0.038], in comparison with the haplotype composed by the major alleles (G-A-T) (Table 5). The C-G-C haplotype was also associated with the presence of overweight/obesity [OR 1.41 (1.01, 1.97; 95% CI), p = 0.044], and elevated plasma TG [coef. 0.023 (0.002, 0.044; 95% CI), p = 0.031] and TG/HDL ratio [(coef. 0.031 (0.001, 0.061; 95% CI), p = 0.045]. Interestingly, after adjusting by BMI, the C-G-C haplotype maintained its association with higher TG levels [coef. 0.023 (0.002, 0.044 95% CI), p = 0.034] (Table 5). On the other hand, the haplotype composed by the minor alleles of rs22074192 and rs2158083 (C-C) was overrepresented in 37% subjects but did not significantly associate with either BMI or lipid levels (not shown). Thus, the C-G-C haplotype of rs879922, rs233575, and rs2158083 SNPs could be considered a risk marker for obesity and dyslipemia in females.
Table 5
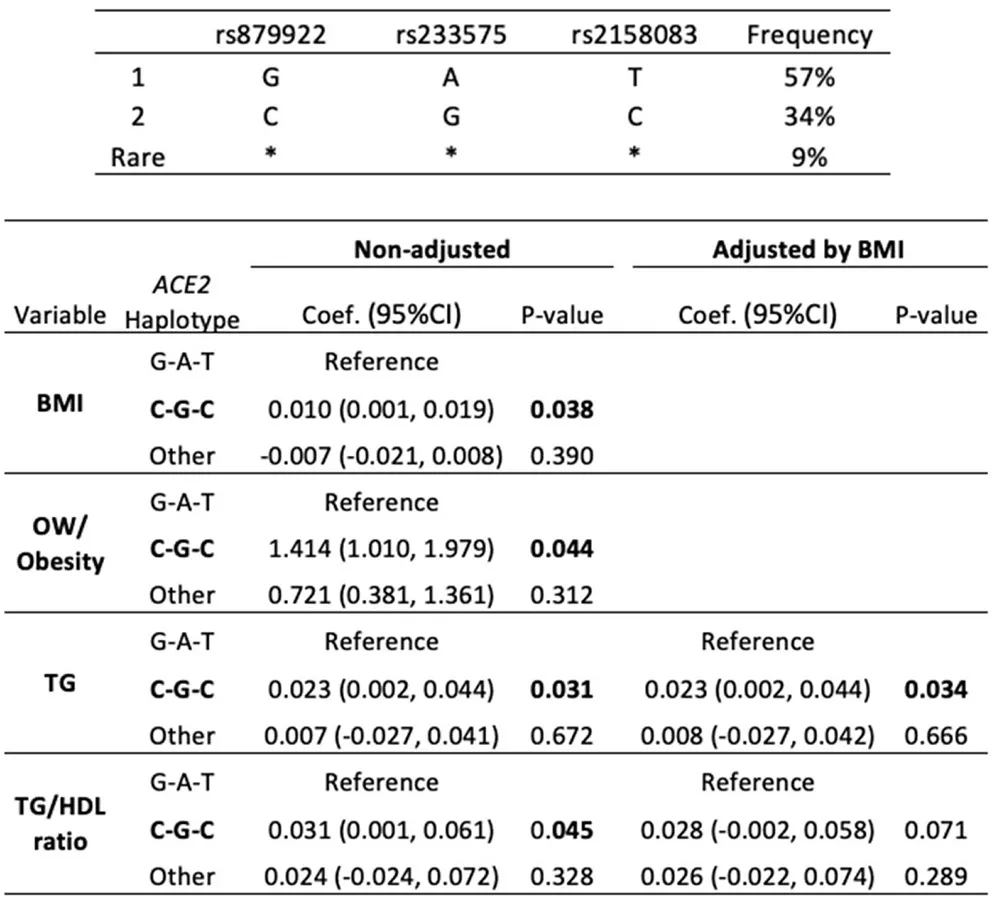 |
ACE2-haplotypes and the risk of obesity and elevated TG levels.
The three ACE2-SNPs (rs879922, rs233575, and rs2158083) related to OW/obesity and elevated plasma lipids in females were analyzed as haplotype. The frequency of minor C-G-C and major G-A-T alleles (top) and their associations with BMI, OW/obesity, TG, and the TG/HDL ratio (bottom) were included. BMI, TG y TG/HDL are shown as coefficient of linear regression. OW/Obesity is shown as Odds ratio of logistic regression.
In bold, statistically significant data (p < 0.05).
Discussion
In this work we have analyzed five SNPs in the ACE2 gene in a general population of apparently healthy adolescents from Spain. The prevalence of these polymorphisms in females and males was comparable to previous findings (12, 22, 23), but the occurrence of some of them was associated with CV risk factors only in girls.
At these ages, females showed lower rates of overweight and obesity, SBP and glycemia than males. Although girls showed higher TC and LDL-C, they enhanced HDL-C, as compensation. They also exhibited lower TG/HDL ratio, which is a predictor of metabolic syndrome and CV diseases (i.e., coronary artery disease or atherosclerosis), and an estimator of LDL-C particle size (20). Thus, theoretically, young girls may be cardio-protected, as previously described (24, 25). Spite of the differences in hormonal regulation between sexes (26), a potential explanation of this cardio-protection could be based on the expression of ACE2 and subsequent effects on the RAAS activity. Since ACE2 is encoded in chromosome X, and specifically in a region that escape from X-inactivation, ACE2 may be upregulated in girls. However, this escape gene also shows heterogeneous sex bias that may lead to lower expression (and tissue-specific) and its activity can be regulated by hormone release (27). Nevertheless, the ACE2/Ang (1-7)/MasR activation could preserve pancreas, skeletal muscle, adipose tissue or the CV system from oxidation, inflammation, and fibrosis (28). Moreover, the elevation of ACE2 involve a reduction of Ang-II, which enhances the NADH/NADPH oxidase activity and reduces the nitric oxide-dependent vasodilatation at systemic level (29). Also, Ang-II stimulates cholesterol biosynthesis, overexpresses lipid receptors (i.e., LRP1, LOX-1), facilitates the uptake and oxidation of LDL-C, and induces pro-inflammatory matrix proteins and matrix-degrading enzymes (i.e., MCP-1, IL-18, PAI-1) (30, 31). In addition, ACE2 may defend the CV system by direct actions. ACE2 increases mitochondrial ATP production and reduces pro-oxidant factors such as NOX4. It also ameliorates lipid deposition and endoplasmic reticule stress via GRP78/eIF2α/XBP-1/ATF4/CHOP expression (32).
Importantly, a reduction of ACE2 levels and subsequent Ang (1-7)-MasR signaling can promote CV failures (33). ACE2 knockout mice showed increased lipid accumulation, ER stress and mitochondrial dysfunction in skeletal muscle, and overexpression of ACE2 reduced these responses and improved glucose and lipid metabolism through the IKKβ/NFkB/IRS-1 pathway (32, 34). Similarly, MasR knockout exhibited dyslipemia and insulin resistance, along with decreased adiponectin secretion and glucose uptake (35). Furthermore, during hypertension or diabetes, a deficiency of ACE2 and increased ACE/ACE2 ratio have been noted (36), and cardioprotective treatments with ACE inhibitors and Ang-II receptor blockers exert their effects partly by increasing ACE2 levels (37). In this regard, we propose that in those girls who carried specific intergenic ACE2 SNPs, the integrity, activity and/or stability of the ACE2 RNA-messenger and resultant protein could have been affected (38, 39), and Ang-II signaling may overcome those of ACE2/Ang (1-7)/MasR. In fact, the existence of rs879922, rs233575, and rs2158083 SNPs of ACE2 was associated with overweight/obesity and elevated plasma TG in Spanish girls. The rs879922 and rs233575 polymorphisms were also related to upper TG/HDL-C ratio, and the rs2074192 and rs2158083 with higher TC levels. Reinforcing our findings, we observed that the C-G-C haplotype of rs879922, rs233575, and rs2158083 was associated with excessive weight and elevated TG levels. Previous data on ACE2 haplotypes linked higher risk of cardiac hypertrophy with minor alleles of rs2074192 and rs2106809 in females (10) or with those of rs2106809 and rs6632677 in males (40).
Little is known about rs2158083 and rs233575. The former is encoded in ACE2's intron 4 and was previously associated with higher blood pressure (12). Similarly, rs233575 (intron 16) was related to higher blood pressure in adolescents and its interaction with obesity was present only in females (13). It was also linked to increased LDL-C (≥1.8 mmol/L) (8) and T2DM (11) and associated with higher LVMI (41). The rs233575 SNP may alter binding of regulatory factors such as RNA-binding proteins involved in splicing (e.g., ARID5A), which is stimulated under inflammation (42). Moreover, rs879922 (intron 11) can be considered a significant quantitative trait locus for essential hypertension in female Chinese, where it also associated with increased TC (≥5.2 mmol/L) and LDL-C (≥1.8 mmol/L) (8). rs879922 was linked with T2DM (11) and cardiac hypertrophy (41), and importantly, the relative expression of ACE2 was significantly lower in subjects with the CC genotype of rs879922 compared with those carrying the GG/CG genotype (43). Finally, rs2074192 (intron 16) has been associated with CV risk (i.e., LVH, carotid arteriosclerosis stenosis, microalbuminuria), retinopathy in T2DM individuals, hypertension, and cardiac hypertrophy (10–12). rs2074192 was also linked to increased TC (≥5.2 mmol/L) in hypertensive population (8), and neonates with rs2074192 were more likely to be born as small for gestational age babies, which contribute to metabolic syndrome and CV diseases in later life (44). Interestingly, rs2074192 was associated with reduced circulating Ang (1-7) levels (22), and in COVID-19 patients, rs2074192 correlated to more severe outcomes (i.e., bilateral pneumonia, dyspnea, high fever) (13, 45).
This study lays the groundwork for subsequent trials in patients and experimental animals. The ACE2-SNPs, rs879922, rs233575, rs2158083, and rs2074192, could help to classify girls at risk of obesity and dyslipidemia in adulthood. The presence of these SNPs should be confirmed in populations of adult women with obesity, dyslipidemia and/or cardiovascular diseases. Also, this work suggests that variability of the ACE2 gene could predispose patients to certain metabolic or cardiovascular conditions that are key in the evolution of other diseases (i.e., COVID-19). Furthermore, these ACE2-mutations might be generated in rodent models with/without obesity and cardiovascular failures in order to characterize atherogenic responses under ACE2 modulation.
Limitations of the Study
At these ages, girls and boys may produce different sexual hormones and factors, which can influence on the lipid profile. Thus, their puberal stage may add important information to explain differences between sex. Also, the precise biological mechanism and other possible factors underlying the association of the ACE2 gene with overweight/obesity or plasma lipids elevation remain to be clarified. Finally, functional investigations of the association between C-G-C haplotype with overweight/obesity or TG elevation could be required.
Conclusion
The ACE2 gene may become a double-sword factor for CV protection, at least in women. Those female adolescents who carry the wild-type ACE2 gene may be somehow protected from increased adiposity and hyperlipidemia. However, Spanish girls carrying the rare alleles of the ACE2-SNPs rs879922, rs233575, rs2158083, or rs2074192 may be vulnerable to future obesity and CV injuries. The haplotype rs879922-rs233575-rs2158083 may be considered a valid biomarker for both pathologies. Therefore, these ACE2-SNPs might be addressed for therapeutic and prognostic purposes against CV diseases. In this sense, new studies might evaluate their role as predictors of worse evolution in COVID-19 subjects, where CV homeostasis may be seriously damaged.
Funding
This research was funded by SPACE2-CV-COVID-CM grant from REACT-EU, Comunidad de Madrid and European Regional Development Fund, and by the Fondo de Investigación Sanitaria-IS. Carlos III (ref.: PI18/01016 and PI20/00923), Biobank (ref.: FEDERRD09/0076/00101), and Ciberdem (ref.: CB07/08/2007). JL-C was granted by FPI contract from Universidad Autónoma, Madrid.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Statements
Data availability statement
The data presented in the study are deposited in the dbSNP repository, submission batch ID 1063361 (dbSNP Build ID: B156).
Ethics statement
The study protocol was approved by the Ethics Committee of Clinical Investigation of the Fundación Jiménez Díaz (Code number: PIC016-2019 FJD). Written informed consent to participate in this study was provided by the participants' legal guardian/next of kin.
Author contributions
JL-C, CV-V, and IP-N quantified the gene expression of ACE2-SNPs and analyze its associations with clinical variables. IM-F performed the statistic studies. LS-G, OL, and CG designed and wrote the manuscript. All authors participate in the discussion of the work.
Acknowledgments
The article is dedicated to the late Prof. Manuel de Oya as the warmest homage to his memory.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcvm.2022.888830/full#supplementary-material
- ACE
angiotensin convertase enzyme
- ACE2
Angiotensin converting enzyme 2
- Ang (1-7)
angiotensin 1–7
- Ang (1-9)
angiotensin 1–9
- Ang I
angiotensin-I
- Ang-II
angiotensin-II
- ARID5A
AT-Rich Interaction Domain 5A
- AT1R
angiotensin II receptor type 1
- AT2R
angiotensin II receptor type 2
- ATP
Adenosine triphosphate
- BMI
body mass index
- COVID
Coronavirus disease
- CV
cardiovascular
- DBP
diastolic blood preassure
- GRP78/eIF2α/XBP-1/ATF4/CHOP, Glucose-Regulated Protein
78-KD/Eukaryotic Initiation Factor 2/X-box binding protein 1/C/EBP Homologous Protein
- HDL-C
high-density lipoprotein-cholesterol
- HOMA-IR
homeostatic model assessment of insulin resistance
- IKKβ/NFkB/IRS-1
IκB Kinase-beta/nuclear factor-kappa B/insulin receptor substrate 1
- IL-18
interleukin-18
- LDL-C
low-density lipoprotein-cholesterol
- LOX-1
lectin-like OxLDL receptor 1
- LRP1
LDL Receptor Related Protein 1
- LVMI
left ventricular mass index
- MasR
Mas receptor
- MCP-1
Monocyte chemoattractant protein-1
- NAD(P)H
nicotinamide adenine dinucleotide (phosphate) hydrogen
- NOX4
NADPH oxidase 4
- NW
normal weight
- OR
odds ratio
- OW
overweight
- PAI-1
Plasminogen activator inhibitor-1
- RAAS
Renin-Angiotensin-Aldosterone system
- SBP
systolic blood pressure
- SNP
single nucleotide polymorphism
- T2DM
type-2 diabetes mellitus
- TC
total cholesterol
- TG
triglycerides.
Abbreviations
References
1.
SantosRASSampaioWOAlzamoraACMotta-SantosDAleninaNBaderMet al. The ACE2/Angiotensin-(1-7)/MAS axis of the renin-angiotensin system: focus on angiotensin-(1-7). Physiol Rev. (2018) 98:505–53. 10.1152/physrev.00023.2016
2.
JugduttBI. Expanding saga of the renin-angiotensin system. Circulation American Heart Association. (2015) 131:1380–3. 10.1161/CIRCULATIONAHA.115.016328
3.
SriramKInselPA. A hypothesis for pathobiology and treatment of COVID-19: The centrality of ACE1/ACE2 imbalance. Br J Pharmacol. (2020) 177:4825–44. 10.1111/bph.15082
4.
PovlsenALGrimmDWehlandMInfangerMKrügerM. The Vasoactive Mas Receptor in Essential Hypertension. J Clin Med. (2020) 9:267. 10.3390/jcm9010267
5.
HikmetFMearLEdvinssonAMickePUhlenMLindskogC. The protein expression profile of ACE2 in human tissues. Mol Syst Biol. (2020) 16:e9610. 10.15252/msb.20209610
6.
LimaRSRochaLPCMoreiraPR. Genetic and epigenetic control of ACE2 expression and its possible role in COVID-19. Cell Biochem Funct. (2021) 39:713–26. 10.1002/cbf.3648
7.
CurtisD. Variants in ACE2 and TMPRSS2 Genes Are Not Major Determinants of COVID-19 Severity in UK Biobank Subjects. Hum Hered. (2020) 85:66–8. 10.1159/000515200
8.
PanYWangTLiYGuanTLaiYShenYet al. Association of ACE2 polymorphisms with susceptibility to essential hypertension and dyslipidemia in Xinjiang, China. Lipids Health Dis. (2018) 17:241. 10.1186/s12944-018-0890-6
9.
FanXWangYSunKZhangWYangXWangSet al. Polymorphisms of ACE2 gene are associated with essential hypertension and antihypertensive effects of Captopril in women. Clin Pharmacol Ther. (2007) 82:187–96. 10.1038/sj.clpt.6100214
10.
FanZWuGYueMYeJChenYXuBet al. Hypertension and hypertensive left ventricular hypertrophy are associated with ACE2 genetic polymorphism. Life Sci. (2019) 225:39–45. 10.1016/j.lfs.2019.03.059
11.
LiuCLiYGuanTLaiYShenYZeyaweidingAet al. ACE2 polymorphisms associated with cardiovascular risk in Uygurs with type 2 diabetes mellitus. Cardiovasc Diabetol. (2018) 17:127. 10.1186/s12933-018-0771-3
12.
MalardLKakinamiLO'LoughlinJRoy-GagnonM-HLabbeAPiloteLet al. The association between the angiotensin-converting enzyme-2 gene and blood pressure in a cohort study of adolescents. BMC Med Genet. (2013) 14:117. 10.1186/1471-2350-14-117
13.
HametPPausovaZAttaouaRHishmihCHalouiMShinJet al. SARS-CoV-2 receptor ACE2 gene is associated with hypertension and severity of COVID 19: interaction with sex, obesity, and smoking. Am J Hypertens. (2021) 34:367–76. 10.1093/ajh/hpaa223
14.
BaderMTurnerAJAleninaN. ACE2, a multifunctional protein - from cardiovascular regulation to COVID-19. Clin Sci Lond Engl 1979. (2020) 134:3229–32. 10.1042/CS20201493
15.
de OyaISchoppenSLasunciónMALopez-SimonLRiestraPde OyaMet al. Sex hormone-binding globulin levels and metabolic syndrome and its features in adolescents. Pediatr Diabetes. (2010) 11:188–94. 10.1111/j.1399-5448.2009.00559.x
16.
ColeTJBellizziMCFlegalKMDietzWH. Establishing a standard definition for child overweight and obesity worldwide: international survey. BMJ. (2000) 320:1240–3. 10.1136/bmj.320.7244.1240
17.
MillerSADykesDDPoleskyHF, A. simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. (1988) 16:1215. 10.1093/nar/16.3.1215
18.
GrundySMStoneNJBaileyALBeamCBirtcherKKBlumenthalRSet al. 2018 AHA/ACC/AACVPR/AAPA/ABC/ACPM/ADA/AGS/APhA/ASPC/NLA/PCNA guideline on the management of blood cholesterol: a report of the American College of Cardiology/American Heart Association task force on clinical practice guidelines. Circulation. (2019) 139:e1082–143. 10.1161/CIR.0000000000000624
19.
American Diabetes Association. Children and adolescents: standards of medical care in diabetes-2020. Diabetes Care. (2020) 43:S163–82. 10.2337/dc20-S013
20.
CorderoAAndrésEOrdoñezBLeónMLaclaustraMGrimaAet al. Usefulness of triglycerides-to-high-density lipoprotein cholesterol ratio for predicting the first coronary event in men. Am J Cardiol. (2009) 104:1393–7. 10.1016/j.amjcard.2009.07.008
21.
ItoyamaSKeichoNHijikataMQuyTPhiNCLongHTet al. Identification of an alternative 5'-untranslated exon and new polymorphisms of angiotensin-converting enzyme 2 gene: lack of association with SARS in the Vietnamese population. Am J Med Genet A. (2005) 136:52–7. 10.1002/ajmg.a.30779
22.
ChenYYZhangPZhouXMLiuDZhongJCZhangCJet al. Relationship between genetic variants of ACE2 gene and circulating levels of ACE2 and its metabolites. J Clin Pharm Ther. (2018) 43:189–95. 10.1111/jcpt.12625
23.
PatelSKWaiBOrdMMacIsaacRJGrantSVelkoskaEet al. Association of ACE2 genetic variants with blood pressure, left ventricular mass, and cardiac function in Caucasians with type 2 diabetes. Am J Hypertens. (2012) 25:216–22. 10.1038/ajh.2011.188
24.
MercuroGDeiddaMPirasADessalviCCMaffeiSRosanoGMC. Gender determinants of cardiovascular risk factors and diseases. J Cardiovasc Med Hagerstown Md. (2010) 11:207–20. 10.2459/JCM.0b013e32833178ed
25.
BarstadLHJúlíussonPBJohnsonLKHertelJKLekhalSHjelmesæthJ. Gender-related differences in cardiometabolic risk factors and lifestyle behaviors in treatment-seeking adolescents with severe obesity. BMC Pediatr. (2018) 18:61. 10.1186/s12887-018-1057-3
26.
KittnarO. Selected sex related differences in pathophysiology of cardiovascular system. Physiol Res. (2020) 69:21–31. 10.33549/physiolres.934068
27.
TukiainenTVillaniA-CYenARivasMAMarshallJLSatijaRet al. Landscape of X chromosome inactivation across human tissues. Nature. (2017) 550:244–8. 10.1038/nature24265
28.
CaoXSongL-NYangJ-K. ACE2 and energy metabolism: the connection between COVID-19 and chronic metabolic disorders. Clin Sci Lond Engl 1979. (2021) 135:535–54. 10.1042/CS20200752
29.
St PaulACorbettCBOkuneRAutieriMV. Angiotensin II, hypercholesterolemia, and vascular smooth muscle cells: a perfect trio for vascular pathology. Int J Mol Sci. (2020) 21:E4525. 10.3390/ijms21124525
30.
SendraJLlorente-CortésVCostalesPHuesca-GómezCBadimonL. Angiotensin II upregulates LDL receptor-related protein (LRP1) expression in the vascular wall: a new pro-atherogenic mechanism of hypertension. Cardiovasc Res. (2008) 78:581–9. 10.1093/cvr/cvn043
31.
ZhangFHuYXuQYeS. Different effects of angiotensin II and angiotensin-(1-7) on vascular smooth muscle cell proliferation and migration. PLoS One. (2010) 5:e12323. 10.1371/journal.pone.0012323
32.
CaoXLuX-MTuoXLiuJ-YZhangY-CSongL-Net al. Angiotensin-converting enzyme 2 regulates endoplasmic reticulum stress and mitochondrial function to preserve skeletal muscle lipid metabolism. Lipids Health Dis. (2019) 18:207. 10.1186/s12944-019-1145-x
33.
RabeloLATodirasMNunes-SouzaVQadriFSzijártóIAGollaschMet al. Genetic deletion of ACE2 induces vascular dysfunction in c57bl/6 mice: role of nitric oxide imbalance and oxidative stress. PLoS ONE. (2016) 11:e0150255. 10.1371/journal.pone.0150255
34.
XuanXGaoFMaXHuangCWangYDengHet al. Activation of ACE2/angiotensin (1-7) attenuates pancreatic β cell dedifferentiation in a high-fat-diet mouse model. Metabolism. (2018) 81:83–96. 10.1016/j.metabol.2017.12.003
35.
SantosSHSFernandesLRMarioEGFerreiraAVMPôrtoLCJAlvarez-LeiteJIet al. Mas deficiency in FVB/N mice produces marked changes in lipid and glycemic metabolism. Diabetes. (2008) 57:340–7. 10.2337/db07-0953
36.
SrivastavaPBadhwarSChandranDSJaryalAKJyotsnaVPDeepakKK. Imbalance between Angiotensin II - Angiotensin (1-7) system is associated with vascular endothelial dysfunction and inflammation in type 2 diabetes with newly diagnosed hypertension. Diabetes Metab Syndr. (2019) 13:2061–8. 10.1016/j.dsx.2019.04.042
37.
KrisztaGKrisztaZVáncsaSHegyiPJFrimLErossBet al. Effects of angiotensin-converting enzyme inhibitors and angiotensin receptor blockers on angiotensin-converting enzyme 2 levels: a comprehensive analysis based on animal studies. Front Pharmacol. (2021) 12:254. 10.3389/fphar.2021.619524
38.
WysockiJYeMSolerMJGurleySBXiaoHDBernsteinKEet al. ACE and ACE2 activity in diabetic mice. Diabetes. (2006) 55:2132–9. 10.2337/db06-0033
39.
ZainaSPérez-LuqueELLundG. Genetics Talks to Epigenetics? The interplay between sequence variants and chromatin structure. Curr Genomics. (2010) 11:359–67. 10.2174/138920210791616662
40.
WangSFuCZouYWangHShiYXuXet al. Polymorphisms of angiotensin-converting enzyme 2 gene associated with magnitude of left ventricular hypertrophy in male patients with hypertrophic cardiomyopathy. Chin Med J (Engl). (2008) 121:27–31. 10.1097/00029330-200801010-00006
41.
LiebWGrafJGötzAKönigIRMayerBFischerMet al. Association of angiotensin-converting enzyme 2 (ACE2) gene polymorphisms with parameters of left ventricular hypertrophy in men. Results of the MONICA Augsburg echocardiographic substudy. J Mol Med Berl Ger. (2006) 84:88–96. 10.1007/s00109-005-0718-5
42.
NyatiKKZamanMM-USharmaPKishimotoT. Arid5a, an RNA-binding protein in immune regulation: RNA stability, inflammation, and autoimmunity. Trends Immunol. (2020) 41:255–68. 10.1016/j.it.2020.01.004
43.
ZhangQCongMWangNLiXZhangHZhangKet al. Association of angiotensin-converting enzyme 2 gene polymorphism and enzymatic activity with essential hypertension in different gender. Medicine (Baltimore). (2018) 97:e12917. 10.1097/MD.0000000000012917
44.
HeJLuY-PLiJLiT-YChenXLiangX-Jet al. Fetal but not maternal angiotensin converting enzyme (ACE)-2 gene Rs2074192 polymorphism is associated with increased risk of being a small for gestational age (SGA) newborn. Kidney Blood Press Res. (2018) 43:1596–606. 10.1159/000494449
45.
CafieroCRosapepeFPalmirottaRReAOttaianoMPBenincasaGet al. Angiotensin system polymorphisms' in SARS-CoV-2 positive patients: assessment between symptomatic and asymptomatic patients: a pilot study. Pharmacogenomics Pers Med. (2021) 14:621–9. 10.2147/PGPM.S303666
Summary
Keywords
ACE2, SNP, haplotype, cardiovascular, obesity, hyperlipidemia
Citation
Lumpuy-Castillo J, Vales-Villamarín C, Mahíllo-Fernández I, Pérez-Nadador I, Soriano-Guillén L, Lorenzo O and Garcés C (2022) Association of ACE2 Polymorphisms and Derived Haplotypes With Obesity and Hyperlipidemia in Female Spanish Adolescents. Front. Cardiovasc. Med. 9:888830. doi: 10.3389/fcvm.2022.888830
Received
04 March 2022
Accepted
28 March 2022
Published
02 May 2022
Volume
9 - 2022
Edited by
George W. Booz, University of Mississippi Medical Center School of Dentistry, United States
Reviewed by
Eric Lazartigues, Louisiana State University, United States; Tlili Barhoumi, King Abdullah International Medical Research Center (KAIMRC), Saudi Arabia
Updates
Copyright
© 2022 Lumpuy-Castillo, Vales-Villamarín, Mahíllo-Fernández, Pérez-Nadador, Soriano-Guillén, Lorenzo and Garcés.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Oscar Lorenzo olorenzo@fjd.es
†These authors have contributed equally to this work
This article was submitted to Cardiovascular Genetics and Systems Medicine, a section of the journal Frontiers in Cardiovascular Medicine
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.