 Armaan Patel
Armaan Patel David W. Mulder
David W. Mulder Dieter Söll
Dieter Söll Natalie Krahn
Natalie Krahn- 1Department of Molecular Biophysics and Biochemistry, Yale University, New Haven, CT, United States
- 2National Renewable Energy Laboratory, Biosciences Center, Golden, CO, United States
- 3Department of Chemistry, Yale University, New Haven, CT, United States
Hydrogen is a clean, renewable energy source, that when combined with oxygen, produces heat and electricity with only water vapor as a biproduct. Furthermore, it has the highest energy content by weight of all known fuels. As a result, various strategies have engineered methods to produce hydrogen efficiently and in quantities that are of interest to the economy. To approach the notion of producing hydrogen from a biological perspective, we take our attention to hydrogenases which are naturally produced in microbes. These organisms have the machinery to produce hydrogen, which when cleverly engineered, could be useful in cell factories resulting in large production of hydrogen. Not all hydrogenases are efficient at hydrogen production, and those that are, tend to be oxygen sensitive. Therefore, we provide a new perspective on introducing selenocysteine, a highly reactive proteinogenic amino acid, as a strategy towards engineering hydrogenases with enhanced hydrogen production, or increased oxygen tolerance.
1 Introduction
C1-utilizing microbes, microorganisms which rely on one carbon molecule for survival, have been of interest to produce biofuels for industrial use (Du et al., 2011). Advances in metabolic engineering have led to the design of biosynthetic pathways as a means to efficiently use cellular machinery (Bar-Even et al., 2010). One application of these engineering strategies is to utilize the activity of [NiFe]- and [FeFe]-hydrogenases in C1 microbes. Hydrogenases are enzymes that catalyze the reversible oxidation of hydrogen and are used for hydrogen production, a renewable source of energy. To compete with existing chemical methods for hydrogen production, hydrogenases require a significant hydrogen production rate (Khanna and Lindblad, 2015). Furthermore, the highest hydrogen producing hydrogenases are also the most oxygen sensitive, reducing their efficiency within these microbial factories. Detailed studies on the factors driving hydrogen production and oxygen sensitivity have facilitated engineering strategies to overcome this (Wittkamp et al., 2018). More specifically, an investigation into the role of selenocysteine (Sec) in these key processes for the development of novel hydrogenases increases the applicability for industrial purposes (Marques et al., 2017; Evans et al., 2021).
Sec, a homolog of cysteine (Cys), is found in redox-associated enzymes across all domains of life (Li et al., 2014). With a single sulfur to selenium replacement compared to Cys, Sec retains similar chemistry but with enhanced chemical properties (i.e., increased nucleophilicity, decreased side-chain pKa, and increased oxidation which is often reversible) (Chung and Krahn, 2022). The distinct characteristics of this amino acid and its similarity to Cys suggests it is potential to affect the active site electronic properties, catalytic rate, or oxygen sensitivity of hydrogenases (Hondal et al., 2013; Marques et al., 2017; Evans et al., 2021).
Occurring naturally in bacteria, Sec is incorporated in proteins at a UGA codon that immediately precedes a hairpin loop (known as the Sec insertion sequence [SECIS] element) in the translated region of the mRNA. Biosynthesis of Sec occurs on tRNASec, where it is first aminoacylated with serine (Ser) by seryl-tRNA synthetase (SerRS) and then converted to Sec by selenocysteine synthase (SelA) (Fu et al., 2018). SelA uses selenophosphate as a substrate for this conversion, provided by selenophosphate synthetase (SelD) (Stock and Rother, 2009). The resulting Sec-tRNASec is recognized by a specialized elongation factor (SelB) for peptide elongation in the ribosome. SelB resembles the canonical elongation factor EF-Tu, but with a C-terminal extension for interaction with the SECIS element (Stock and Rother, 2009). This complex and highly regulated path for insertion of Sec (Figures 1A, B) has been an obstacle for recombinant selenoprotein production (Fu et al., 2018).
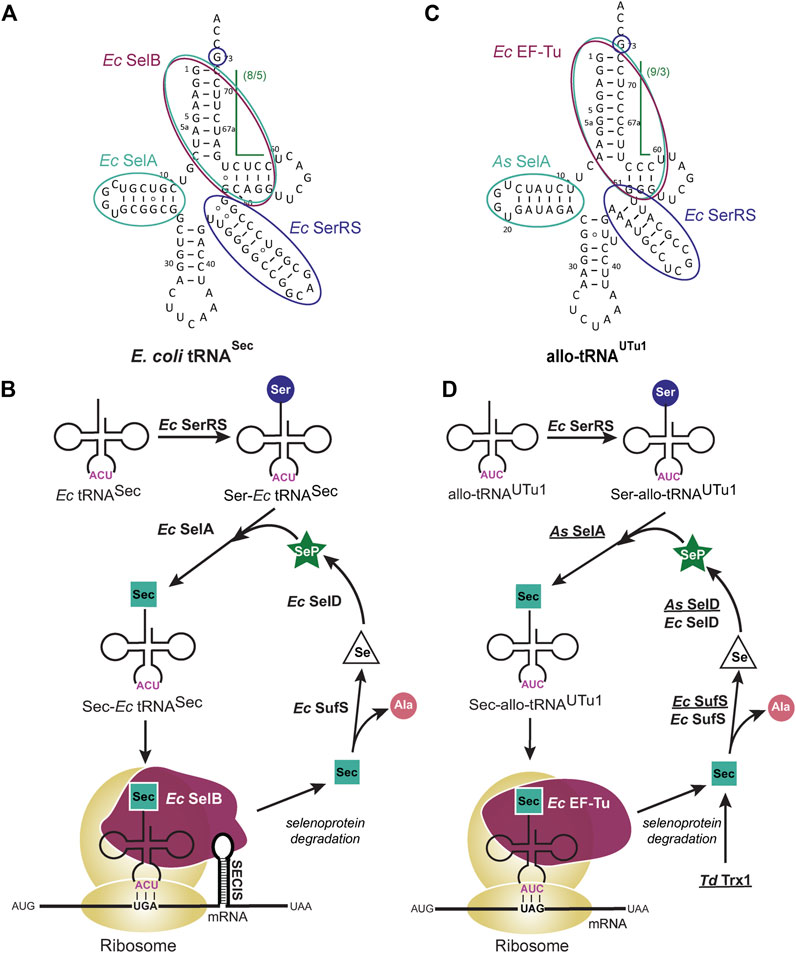
FIGURE 1. Selenocysteine (Sec) tRNA structure facilitates translation and insertion of Sec. (A) Escherichia coli (Ec) tRNASec secondary structure highlights the features which facilitate its natural biosynthesis and insertion into the growing polypeptide chain. The long variable arm and G73 discriminator base (dark blue) are recognized by Ec seryl-tRNA synthetase (SerRS) for initial aminoacylation with Ser. Conversion of Ser to Sec occurs through Ec selenocysteine synthase (SelA) which recognizes the D-arm and (8/5) acceptor domain (teal). This 13 bp acceptor domain discriminates tRNASec from canonical tRNAs and enables recognition by a specialized elongation factor, Ec SelB (maroon), for insertion into the ribosome. (B) Ec SelB also recognizes an mRNA hairpin [Sec insertion sequence (SECIS) element] that is immediately downstream of the UGA codon for insertion of Sec. Degradation of used selenoproteins releases Sec which is a substrate for Ec Sec lyase (SufS), converting the amino acid to alanine (Ala) and releasing selenium (Se). Se is then converted to selenophosphate (SeP) by selenophosphate synthetase (SelD) where it re-enters the biosynthesis path. (C) allo-tRNAUTu1 secondary structure highlights the features which facilitate a simpler biosynthesis and insertion path for Sec in bacteria. The long variable arm and G73 discriminator base (dark blue) are recognized by Ec SerRS while the D-arm and (9/3) acceptor domain (teal) are recognized by Aeromonas salmonicida (As) SelA. Elongation of allo-tRNAUTu1 and insertion of Sec into the polypeptide chain occurs with Ec EF-Tu (maroon) which does not require the restrictive SECIS element. (D) The pSecUAG-Evol plasmid harbors additional enzymes that are recombinantly expressed (underlined) to promote Sec insertion in this simplified path. Selenoprotein degradation, including that of the additional Treponema denticola (Td) Trx1 releases Sec which is converted to Ala by both recombinant and endogenous SufS. To increase conversion of Se to SeP, both As SelD and Ec SelD are present.
In this perspective, we discuss the details of an emerging technology for site-specific Sec insertion in Escherichia coli and how it can be adapted to cell factories. We focus on applying these cell factories for hydrogen production, highlighting recent evidence on the novel properties imparted by Sec on hydrogenases.
2 Engineering tRNAs for SECIS-independent selenocysteine insertion
The complicated biosynthesis of Sec is facilitated by the tRNASec structure, its main distinguishing factor being a 13 bp acceptor domain (acceptor stem and T-stem combined). This promotes recognition by SelB and reduces EF-Tu binding which favors the canonical 7/5 tRNA structure (12 bp acceptor domain) (Krahn et al., 2020). SelB also requires recognition of the SECIS element to facilitate insertion of Sec. This interaction controls the position of Sec within a protein, but it restricts the ability to overexpress these proteins or make novel selenoproteins (Cheng and Arnér, 2017). Therefore, to remove these restrictions, it was hypothesized that a tRNA could be engineered to biosynthesize Sec but be inserted in the ribosome by EF-Tu (Aldag et al., 2013; Miller et al., 2015; Thyer et al., 2015). Taking advantage of prior knowledge on the tRNASec elements required to interact with each protein in this pathway (Krahn et al., 2020), one can strategically engineer a tRNA for SECIS-independent Sec insertion.
The discovery of allo-tRNAs, a group of tRNAs with novel secondary structure (Mukai et al., 2017), provided a scaffold for engineering a tRNA to insert Sec in an EF-Tu dependent manner. Allo-tRNAs are found to have 12 bp acceptor domains in a 9/3 or 8/4 configuration, unlike the 7/5 configuration of canonical tRNAs (Figure 1C). This alternate arrangement of the acceptor stem and T-stem does not appear to limit its binding to EF-Tu (Mukai et al., 2017), nor does it affect the distance between the acceptor stem and anticodon (Prabhakar et al., 2022). The anticodon is not a recognition element for any of the involved enzymes in Sec biosynthesis (Krahn et al., 2020), therefore it can be manipulated for readthrough of Sec at a UAG codon in addition to its natural suppression at a UGA codon (Mukai et al., 2017; Mukai et al., 2018). Moreover, some allo-tRNAs have features that resemble tRNASec, namely the G73 discriminator base, long variable arm, and unique D-arm. The first two features are key identity elements required for SerRS recognition, while the latter is recognized by SelA (Figure 1C) (Krahn et al., 2020). For these reasons, allo-tRNAs became the backbone for tRNA engineering in a rewired Sec translation path (Figure 1D) (Mukai et al., 2018).
Since it was confirmed that allo-tRNAs could be efficiently recognized by EF-Tu (Mukai et al., 2017), it was imperative to ensure that only Sec would get inserted into the protein. Unlike SelB which preferentially recognizes Sec-tRNA over Ser-tRNA (Leibundgut et al., 2005), EF-Tu only requires the presence of an amino acid on the tRNA (Schrader et al., 2011). Therefore, a closer investigation of the SelA and tRNASec interaction was required to promote Ser to Sec conversion. In addition to the D-arm, E. coli SelA also recognizes the 13 bp acceptor domain of tRNASec (Krahn et al., 2020). To promote interaction with allo-tRNAs, Aeromonas salmonicida (As), a close relative of E. coli, was found to have a SelA enzyme able to recognize 12 bp acceptor domain tRNAs. This major change accommodated Sec conversion on allo-tRNAs and was further enhanced by engineering the D-arm of allo-tRNAs to resemble that of As tRNASec (Figure 1C) (Mukai et al., 2018).
The unique 9/3 structure of allo-tRNAs facilitate Sec biosynthesis and recognition by EF-Tu but does not provide any evidence as to how well it is accepted by the ribosome. Structural analysis and single-molecule translation studies revealed that the 9/3 acceptor domain does not interfere with translation. Instead, it was the rigidity of the variable arm affecting translocation of the allo-tRNA from the A- to the P-site of the ribosome. A single point mutation to disrupt the tertiary interaction in the central loop of the tRNA increased the flexibility of the variable arm to promote Sec insertion (Prabhakar et al., 2022).
The strategy of engineering allo-tRNAs for EF-Tu dependent translation has resulted in improved Sec incorporation compared to tRNAs engineered as a hybrid of tRNASec and tRNASer (Aldag et al., 2013; Miller et al., 2015; Thyer et al., 2015). The structure of allo-tRNAs has decoupled Sec translation from SelB and likewise the SECIS-element to facilitate Sec insertion into any position in the protein (Mukai et al., 2018). The versatility to site-specifically insert Sec and the enhanced chemical properties imposed by this amino acid makes it attractive for use in C1 microbial factories.
2.1 Adapting Sec translation in bacterial cell systems
The expression of allo-tRNAs in bacterial cell systems is mediated through a plasmid, specifically the pSecUAG-Evol plasmid series (Mukai et al., 2018; Chung et al., 2021). pSecUAG-Evol contains an allo-tRNA gene under an araC promoter plus other protein genes which have been found to enhance Sec insertion: i) As SelA to facilitate Ser to Sec conversion on allo-tRNAs and ii) As SelD, iii) Treponema denticola (Td) Trx1 and iv) mutant E. coli Sec lyase (SufS_C364A) to increase available selenium. With this plasmid, only two endogenous components from the host are required: i) SerRS for initial Ser aminoacylation and ii) EF-Tu to facilitate elongation in the ribosome. Endogenously expressed E. coli SelD and SufS are not essential to the path but can assist the recombinantly expressed proteins (As SelD and Ec SufS_C364A) to promote Sec insertion (Figure 1D).
The effectiveness of using allo-tRNAs for Sec insertion in E. coli suggests that they can also be adapted in bacterial cell factories. These factories can be composed of organisms from the genus Clostridium (Liew et al., 2016). For example, metabolically engineered Clostridium autoethanogenum is used to increase production of ethanol from carbon fixation (Liew et al., 2017). C. autoethanogenum, like E. coli, contains machinery for Sec incorporation, as seen by the presence of selenium containing formate dehydrogenase (Abubackar et al., 2015). Therefore, some of the proteins used for endogenous Sec insertion can also assist in recombinant expression. Furthermore, the >70% homology of both EF-Tu and SerRS in C. autoethanogenum compared to E. coli suggest that allo-tRNAs can potentially be serylated and recognized for elongation in these organisms.
The comparable machineries indicates that the pSecUAG-Evol plasmid system used in E. coli could be adapted for use in C. autoethanogenum. Optimization to determine the requirement or replacement of As SelD, Td Trx1 and Ec SufS_C364A to promote Sec insertion may be needed. These three proteins were added to increase the amount of available selenium in the cells without inducing cellular toxicity from additional selenium supplementation (Mukai et al., 2018). Codon usage in E. coli and Clostridium species also differs. Therefore, for optimal protein expression from the pSecUAG-Evol plasmid, codon optimization of these mRNA sequences would be preferable. As has been observed with E. coli, the expression levels of these additional proteins can be burdensome on the organism, therefore is should not be assumed that maximal expression levels are optimal (Mukai et al., 2018).
Another important consideration when adapting this system to bacterial cell factories, is the concentration and form of selenium donor that is being used. Some organisms tolerate different concentrations of sodium selenite (the selenium donor used in E. coli) before it inhibits their growth. However, other sources of selenium, such as selenomethionine can also be used and may be preferable for that organism (Mony and Larras-Regard, 2000).
3 Hydrogenases as a target for selenocysteine incorporation
Hydrogenases, found across diverse microorganisms, are a class of enzymes that carry out essential functions in hydrogen metabolism (Vignais and Billoud, 2007; Calusinska et al., 2010; Peters et al., 2015; Greening et al., 2016). Two major classes of hydrogenases named according to the metal centers in their active site, [NiFe]- and [FeFe]-hydrogenases, catalyze the reversible oxidation of H2 gas from electrons and protons (2H+ + 2e− ⇆ H2) (Figure 2) (Lubitz et al., 2014). A subset of [NiFe]-hydrogenases, the [NiFeSe]-hydrogenase, contains Sec in place of a Cys ligand in the active site (Garcin et al., 1999; Marques et al., 2010; Volbeda et al., 2013). A third class of hydrogenases, the Fe-hydrogenases, contain a mono-nuclear Fe active site and activates H2 in the presence of the substrate methylenetetrahydromethanopterin (Shima et al., 2008). Due to the ability of the [NiFe]- and [FeFe]-hydrogenases to achieve high reaction rates under ambient conditions, these enzymes and respective active-site structures serve as design models for bio-mimetic and bio-inspired catalysts (Simmons et al., 2014; Schilter et al., 2016; Dutta et al., 2018; Kleinhaus et al., 2021).
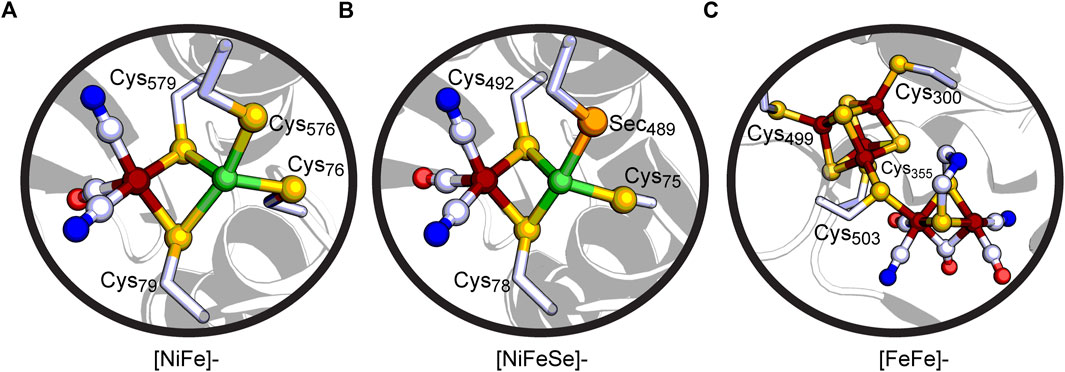
FIGURE 2. Cut-away views of (A) the active site of the [NiFe]-hydrogenase Hyd-1 from Escherichia coli (PDB: 5A4M), (B) the active site of the [NiFeSe]-hydrogenase from Desulfovibrio vulgaris Hildenborough (PDB: 5JSK), and (C) the H-cluster active site of the [FeFe]-hydrogenase CpI from Clostridium pasteurianum (PDB: 3C8Y). Coloring scheme: C, white; O, red; N, blue; S, yellow-orange; Se, orange; Ni, green; Fe, rust.
While the catalytic activation of H2 is reversible, the [NiFe]- and [FeFe]-hydrogenases often display a catalytic bias which is reflected by dis-proportionate reactions rates for the H2 oxidation and evolution directions (Mulder et al., 2021). This underscores the enzymes function in H2 metabolism, making them ideal targets for renewable H2 production and storage technologies through coupling to other reductive or oxidative reactions (Schuchmann et al., 2018). By example, electrons generated by the H2 oxidation reaction from [FeFe]-hydrogenase can be coupled to reductive processes such as CO2 reduction to formate (Schuchmann and Müller, 2013). Conversely, the H2 evolution reaction is often coupled to oxidation of mobile electron-carriers such as ferredoxin or NADPH in anaerobic fermentation. Green algae and cyanobacteria type hydrogenases play an important role in photosynthetic H2 production, through coupling electrons generated from the water splitting reaction to H2 evolution (Akhlaghi and Najafpour-Darzi, 2020; Kosourov et al., 2021).
3.1 [NiFe]- and [NiFeSe]-hydrogenases
[NiFe]-hydrogenases contain an active site with the Ni and Fe metals bridged together by Cys residues (Figure 2A) (Volbeda et al., 1995; Shafaat et al., 2013; Lubitz et al., 2014; Peters et al., 2015; Ogata et al., 2016). The Ni in the active site is coordinated by four Cys ligands: two bridging Cys thiolate ligands connect the Ni to the Fe and two terminal Cys thiolate ligands are located at the Ni. Furthermore, the Fe is coordinated by two CN− molecules and one CO molecule. The [NiFeSe]-hydrogenases share a similar active site (Figure 2B), whereas one of the Cys ligands is Sec. Nearby non-coordinating conserved residues by the protein scaffold play important roles in tuning the active site for reversible H2 oxidation reaction through hydrogen-bonding interactions (Brooke et al., 2017). Conserved Asp and Arg residues found distal to the bridging Cys ligands are necessary for efficient H2 oxidation (Evans et al., 2016; Vansuch et al., 2020), whereas a conserved Glu residue adjacent to the terminal Cys acts as the universal proton gate to the active site (Dementin et al., 2004; Evans et al., 2018). Outside of the active site environment, there exist FeS clusters which allow for long-range transfer of electrons. The O2-tolerant [NiFe]-hydrogenases (Group 1D) contain a unique proximal [4Fe-3S]6-Cys ligated cluster in relation to the active site that can reduce O2 to water, preventing a loss of catalytic activity (Fritsch et al., 2011; Goris et al., 2011; Shomura et al., 2011). On the other hand, O2-sensitive [NiFe]-hydrogenases do not have all 6 Cys residues present in this FeS cluster, which has been suggested to cause O2-sensitivity. [NiFeSe]-hydrogenases (Group 1A) are reported to have higher activity for producing H2 while [NiFe]-hydrogenases are typically biased for H2 oxidation (Parkin et al., 2008).
The [NiFe]-hydrogenases lose much of their catalytic activity in the presence of O2. In some [NiFe]-hydrogenases they regain activity once the O2 dissipates, but in others they require proton reduction before they become active again (Shafaat et al., 2013; Ogata et al., 2016). Those enzymes capable of oxidizing H2 in the presence of oxygen (albeit at a lower level) are deemed O2 tolerant. The catalytic mechanism of [NiFe]-hydrogenases has been well-studied for use in biotechnological applications, however the requirement to work in an aerobic environment reduces the feasibility of this endeavor.
As we begin to understand the role of Sec in [NiFeSe]-hydrogenases, there may be a solution unraveling to the O2 sensitivity roadblock. Initial studies looking at a natural [NiFeSe]-hydrogenase, investigated the effects of Sec to Cys substitution on its activity and O2 tolerance. The presence of a Cys instead of a Sec at the active site reduced Ni incorporation, H2 production activity, and O2 tolerance of the enzyme (Marques et al., 2017). In an opposite study, effects of a Cys to Sec substitution in a natural [NiFe]-hydrogenase were investigated (Evans et al., 2021). Regardless of the position of the Sec substitution in the active site, the H2 oxidation activity was reduced, and no H2 production activity was observed. On the other hand, substituting a Sec residue in the same position as what is observed in natural [NiFeSe]-hydrogenases, significantly increases the O2 tolerance (Evans et al., 2021).
These combined results suggest that the presence of Sec in the active site is not the only factor affecting a shift to H2 production activity. Rather the surrounding environment of the catalytic active site also influences the capability of [NiFe]-hydrogenases to oxidize or produce H2. However, the evidence is convincing that the presence of selenium increases O2 tolerance. This is the opposite to what one may expect given that selenium is readily oxidized. While this is true, it appears that the reversibility and ease in reduction of selenium oxides compared to sulfur oxides plays a role in this effect (Maroney and Hondal, 2018). One suggestion is that the selenium atom acts as a decoy, being preferentially oxidized, and therefore avoiding oxidation and inactivation of the [NiFe]-active site. Further investigation into the details of this chemical behavior still need to be uncovered. Conveying these two important details, [NiFeSe]-hydrogenases can be a solution towards the oxidation inactivation problem, and a promising target for growth in C1 microbes.
3.2 [FeFe]-hydrogenases
The second major group of hydrogenases, [FeFe]-hydrogenases, contain a complex metallocofactor active site termed the H-cluster and are known to be very active in H2 production (Lubitz et al., 2014; Peters et al., 2015). The H-cluster is comprised of a Cys ligated [4Fe-4S] cluster ([4Fe-4S]H) and binuclear iron site ([2Fe]H) (Figure 2C) (Peters et al., 1998; Nicolet et al., 1999). The two centers are bridged and electronically linked via a Cys thiolate ligand which is the only covalent attachment point for [2Fe]H to the protein scaffold (Popescu and Münck, 1999). This differs from the structure of the [NiFe]-hydrogenase active site, which contains multiple bridging Cys ligands connecting the active site to the protein scaffold. Similar to the [NiFe]-hydrogenase active site, biologically unique CO and CN− ligands coordinate the Fe atoms of [2Fe]H, in addition to a bridging azadithiolate ligand (Silakov et al., 2007; Berggren et al., 2013; Esselborn et al., 2013). The latter plays an important role in H2 activation. The bridgehead amine can function as a catalytic base (Fan and Hall, 2001) to the H+ binding site at the distal Fe atom of [2Fe]H, providing a key interaction point with a conserved proton-transfer pathway that terminates with a Cys within H-bonding distance of the ligand (Ginovska-Pangovska et al., 2014; Duan et al., 2018; Kisgeropoulos et al., 2022). Numerous site-directed mutagenesis studies have demonstrated the importance of the surrounding protein scaffold of the H-cluster in tuning its electronic properties and catalytic activity (Winkler et al., 2013; Stripp et al., 2022). Similar to [NiFe]-hydrogenases, long range potential effects on catalysis are possible through the presence of additional FeS clusters that function in electron-transfer (Greco et al., 2011; Rodriguez-Maciá et al., 2017; Caserta et al., 2018). These are present in different subgroups of [FeFe]-hydrogenases by means of ferredoxin-like binding domains present in a modular-like fashion (Mulder et al., 2011).
Attempts have been made to introduce exogenous hydrogenases into cyanobacteria that can either pair with the endogenous, bidirectional Hox [NiFe]-hydrogenase or operate in engineered strains devoid of Hox and other endogenous uptake [NiFe]-hydrogenases (Ducat et al., 2011; Gartner et al., 2012; Khanna and Lindblad, 2015; Avilan et al., 2018; Kosourov et al., 2021). Success was found by expressing an [FeFe]-hydrogenase from the energy demanding nitrogen-fixing Clostridium acetobutylicum in non-nitrogen-fixing Synechococcus elongatus sp. 7942 (Ducat et al., 2011). Without the need to use energy from the sun for nitrogen fixation, the sunlight to H2 conversion was increased. A significant challenge towards these types of approaches is the extreme O2 sensitivity of [FeFe]-hydrogenases. While certain [FeFe]-hydrogenases such as one from Clostridium beijerinckii (CbA5H) are emerging with unique O2 sensitivity properties (Morra et al., 2016; Corrigan et al., 2020; Winkler et al., 2021; Morra, 2022), the majority of characterized [FeFe]-hydrogenases are irreversibly inactivated by O2. As a result, biohydrogen production must be a delicate process to ensure that [FeFe]-hydrogenases are not inhibited by O2 produced from light-dependent reactions. This is typically approached by regular aerobic growth of cyanobacteria to generate internal stores of reductants before transferring to an anaerobic atmosphere (with inactivation of photosystem III), to facilitate hydrogenase activity (Ducat et al., 2011).
These efforts show the potential for increasing H2 production levels of [FeFe]-hydrogenases, though deeper understanding of the H2 metabolism in cyanobacteria is necessary (Khanna and Lindblad, 2015; Kosourov et al., 2021). A recurring problem is that cyanobacteria are unable to proceed with both light-dependent reactions and [FeFe]-hydrogenase activity at the same time. Taking into consideration the remarkable O2 tolerance gained by Sec substitution in the active site of a [NiFe]-hydrogenase, there is a prospective application for Sec insertion in the active site of [FeFe]-hydrogenases. Related studies have looked at selenium substitution at the sulfide positions of the H-cluster metal site (Noth et al., 2016; Kertess et al., 2017). This is made possible through semi-artificial reconstitution procedures which achieve an active [FeFe]-hydrogenase by addition of a synthetic [2Fe]H cluster {[Fe2(SCH2NHCH2S)(CO)4(CN)2]2−} into an apo-form of the enzyme only containing [4Fe-4S]H (Berggren et al., 2013; Esselborn et al., 2013). For one selenium derivative H-cluster containing selenium at the thiolate positions of the azadithiolate ligand (SeCH2NHCH2Se), a slight increase in catalytic bias toward H2 evolution was observed, however this was accompanied by a decrease in O2 stability compared to the native enzyme (Kertess et al., 2017). Another selenium derivative H-cluster containing selenium at the [4Fe-4S]H sulfide positions ([4Fe-4Se]H) displayed similar catalytic rates for H2 evolution compared to the native enzyme (Noth et al., 2016).
4 Outlook
We have highlighted an emerging recombinant technology for site-specific Sec insertion into proteins using novel allo-tRNAs in a specially designed plasmid. Furthermore, we have shown its application in hydrogenases and emphasize the transferability of this system into microbial cell factories. The unique chemical properties of Sec makes its incorporation beneficial to engineering hydrogenases with increased or tuned catalytic activity and, perhaps more difficult, O2 tolerance.
Evidence thus far has investigated the incorporation of Sec in the active site, however other conserved structures such as FeS clusters that function in electron-transfer may also play critical roles in regulating tolerance to oxidative damage. One such example for certain types of [NiFe]-hydrogenases which confer extreme O2 tolerance, is the presence of a unique [4Fe-3S] cluster proximal to the active site that contains 6 conserved Cys residues (Dementin et al., 2004; Evans et al., 2018). O2 tolerance is lost upon two Cys to Gly mutations and growth of the organisms in high O2 concentrations was affected (Goris et al., 2011). Although the tendency has been to substitute Sec for Cys in the active site, this suggests that other FeS cluster electron-transfer centers which are widely present among diverse [NiFe]- and [FeFe]-hydrogenases may also be suitable targets.
While we stress the substitution of Sec in hydrogenases and use in H2 production, the unique chemical properties of Sec can be harnessed in any protein when engineering microbial cell factories. Specifically, enhancing catalytic properties to enzymes which use Cys in their active site is on the top of the list.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
AP, DM, and NK wrote the manuscript. NK conceptualized the idea. NK and DS edited the manuscript.
Funding
This work was supported by grants from the National Institute of General Medical Sciences (R35GM122560-05S1 to DS) and the Department of Energy Office of Basic Energy Sciences (DE-FG0298ER2031 to DS). For research on hydrogenases, DM was supported by funds from the U.S. Department of Energy Office of Basic Energy Sciences, Division of Chemical Sciences, Geosciences, and Biosciences, Photosynthetic Systems Program. This work was authored in part by the Alliance for Sustainable Energy, LLC, the manager and operator of the National Renewable Energy Laboratory for the U.S. Department of Energy (DOE) under Contract No. DE-AC36-08GO28308. The U.S. Government retains and the publisher, by accepting the article for publication, acknowledges that the U.S. Government retains a non-exclusive, paid-up, irrevocable, worldwide license to publish or reproduce the published form of this work, or allow others to do so, for U.S. Government purposes.
Acknowledgments
We would like to thank Fraser Armstrong and Rhiannon Evans for critical discussions.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abubackar, H. N., Veiga, M. C., and Kennes, C. (2015). Carbon monoxide fermentation to ethanol by Clostridium autoethanogenum in a bioreactor with no accumulation of acetic acid. Bioresour. Technol. 186, 122–127. doi:10.1016/j.biortech.2015.02.113
Akhlaghi, N., and Najafpour-Darzi, G. (2020). A comprehensive review on biological hydrogen production. Int. J. Hydrogen Energy 45 (43), 22492–22512. doi:10.1016/j.ijhydene.2020.06.182
Aldag, C., Bröcker, M. J., Hohn, M. J., Prat, L., Hammond, G., Plummer, A., et al. (2013). Rewiring translation for elongation factor Tu-dependent selenocysteine incorporation. Angew. Chem. Int. Ed. Engl. 52 (5), 1481–1485. doi:10.1002/ange.201207567
Avilan, L., Roumezi, B., Risoul, V., Bernard, C. S., Kpebe, A., Belhadjhassine, M., et al. (2018). Phototrophic hydrogen production from a clostridial [FeFe] hydrogenase expressed in the heterocysts of the cyanobacterium Nostoc PCC 7120. Appl. Microbiol. Biotechnol. 102 (13), 5775–5783. doi:10.1007/s00253-018-8989-2
Bar-Even, A., Noor, E., Lewis, N. E., and Milo, R. (2010). Design and analysis of synthetic carbon fixation pathways. Proc. Natl. Acad. Sci. U. S. A. 107 (19), 8889–8894. doi:10.1073/pnas.0907176107
Berggren, G., Adamska, A., Lambertz, C., Simmons, T. R., Esselborn, J., Atta, M., et al. (2013). Biomimetic assembly and activation of [FeFe]-hydrogenases. Nature 499 (7456), 66–69. doi:10.1038/nature12239
Brooke, E. J., Evans, R. M., Islam, S. T., Roberts, G. M., Wehlin, S. A., Carr, S. B., et al. (2017). Importance of the active site "canopy" residues in an O2-tolerant [NiFe]-hydrogenase. Biochemistry 56 (1), 132–142. doi:10.1021/acs.biochem.6b00868
Calusinska, M., Happe, T., Joris, B., and Wilmotte, A. (2010). The surprising diversity of clostridial hydrogenases: A comparative genomic perspective. Microbiol. Read. 156 (6), 1575–1588. doi:10.1099/mic.0.032771-0
Caserta, G., Papini, C., Adamska-Venkatesh, A., Pecqueur, L., Sommer, C., Reijerse, E., et al. (2018). Engineering an [FeFe]-hydrogenase: Do accessory clusters influence O2 resistance and catalytic bias? J. Am. Chem. Soc. 140 (16), 5516–5526. doi:10.1021/jacs.8b01689
Cheng, Q., and Arnér, E. S. (2017). Selenocysteine insertion at a predefined UAG codon in a release factor 1 (RF1)-depleted Escherichia coli host strain bypasses species barriers in recombinant selenoprotein translation. J. Biol. Chem. 292 (13), 5476–5487. doi:10.1074/jbc.m117.776310
Chung, C. Z., and Krahn, N. (2022). The selenocysteine toolbox: A guide to studying the 21st amino acid. Arch. Biochem. Biophys. 730, 109421. doi:10.1016/j.abb.2022.109421
Chung, C. Z., Miller, C., Söll, D., and Krahn, N. (2021). Introducing selenocysteine into recombinant proteins in Escherichia coli. Curr. Protoc. 1 (2), e54. doi:10.1002/cpz1.54
Corrigan, P. S., Tirsch, J. L., and Silakov, A. (2020). Investigation of the unusual ability of the [FeFe] hydrogenase from Clostridium beijerinckii to access an O2-protected state. J. Am. Chem. Soc. 142 (28), 12409–12419. doi:10.1021/jacs.0c04964
Dementin, S., Burlat, B., De Lacey, A. L., Pardo, A., Adryanczyk-Perrier, G., Guigliarelli, B., et al. (2004). A glutamate is the essential proton transfer gate during the catalytic cycle of the [NiFe] hydrogenase. J. Biol. Chem. 279 (11), 10508–10513. doi:10.1074/jbc.m312716200
Du, J., Shao, Z., and Zhao, H. (2011). Engineering microbial factories for synthesis of value-added products. J. Ind. Microbiol. Biotechnol. 38 (8), 873–890. doi:10.1007/s10295-011-0970-3
Duan, J., Senger, M., Esselborn, J., Engelbrecht, V., Wittkamp, F., Apfel, U. P., et al. (2018). Crystallographic and spectroscopic assignment of the proton transfer pathway in [FeFe]-hydrogenases. Nat. Commun. 9 (1), 4726. doi:10.1038/s41467-018-07140-x
Ducat, D. C., Sachdeva, G., and Silver, P. A. (2011). Rewiring hydrogenase-dependent redox circuits in cyanobacteria. Proc. Natl. Acad. Sci. U. S. A. 108 (10), 3941–3946. doi:10.1073/pnas.1016026108
Dutta, A., Appel, A. M., and Shaw, W. J. (2018). Designing electrochemically reversible H2 oxidation and production catalysts. Nat. Rev. Chem. 2 (9), 244–252. doi:10.1038/s41570-018-0032-8
Esselborn, J., Lambertz, C., Adamska-Venkates, A., Simmons, T., Berggren, G., Noth, J., et al. (2013). Spontaneous activation of [FeFe]-hydrogenases by an inorganic [2Fe] active site mimic. Nat. Chem. Biol. 9 (10), 607–609. doi:10.1038/nchembio.1311
Evans, R. M., Ash, P. A., Beaton, S. E., Brooke, E. J., Vincent, K. A., Carr, S. B., et al. (2018). Mechanistic exploitation of a self-repairing, blocked proton transfer pathway in an O2-tolerant [NiFe]-hydrogenase. J. Am. Chem. Soc. 140 (32), 10208–10220. doi:10.1021/jacs.8b04798
Evans, R. M., Brooke, E. J., Wehlin, S. A., Nomerotskaia, E., Sargent, F., Carr, S. B., et al. (2016). Mechanism of hydrogen activation by [NiFe] hydrogenases. Nat. Chem. Biol. 12 (1), 46–50. doi:10.1038/nchembio.1976
Evans, R. M., Krahn, N., Murphy, B. J., Lee, H., Armstrong, F. A., and Söll, D. (2021). Selective cysteine-to-selenocysteine changes in a [NiFe]-hydrogenase confirm a special position for catalysis and oxygen tolerance. Proc. Natl. Acad. Sci. U. S. A. 118 (13), e2100921118. doi:10.1073/pnas.2100921118
Fan, H. J., and Hall, M. B. (2001). A capable bridging ligand for Fe-only hydrogenase: Density functional calculations of a low-energy route for heterolytic cleavage and formation of dihydrogen. J. Am. Chem. Soc. 123 (16), 3828–3829. doi:10.1021/ja004120i
Fritsch, J., Scheerer, P., Frielingsdorf, S., Kroschinsky, S., Friedrich, B., Lenz, O., et al. (2011). The crystal structure of an oxygen-tolerant hydrogenase uncovers a novel iron-sulphur centre. Nature 479 (7372), 249–252. doi:10.1038/nature10505
Fu, X., Söll, D., and Sevostyanova, A. (2018). Challenges of site-specific selenocysteine incorporation into proteins by Escherichia coli. RNA Biol. 15 (4-5), 461–470. doi:10.1080/15476286.2018.1440876
Garcin, E., Vernede, X., Hatchikian, E. C., Volbeda, A., Frey, M., and Fontecilla-Camps, J. C. (1999). The crystal structure of a reduced [NiFeSe] hydrogenase provides an image of the activated catalytic center. Structure 7 (5), 557–566. doi:10.1016/s0969-2126(99)80072-0
Gartner, K., Lechno-Yossef, S., Cornish, A. J., Wolk, C. P., and Hegg, E. L. (2012). Expression of Shewanella oneidensis MR-1 [FeFe]-hydrogenase genes in Anabaena sp. strain PCC 7120. Appl. Environ. Microbiol. 78 (24), 8579–8586. doi:10.1128/aem.01959-12
Ginovska-Pangovska, B., Ho, M. H., Linehan, J. C., Cheng, Y., Dupuis, M., Raugei, S., et al. (2014). Molecular dynamics study of the proposed proton transport pathways in [FeFe]-hydrogenase. Biochim. Biophys. Acta 1837 (1), 131–138. doi:10.1016/j.bbabio.2013.08.004
Goris, T., Wait, A. F., Saggu, M., Fritsch, J., Heidary, N., Stein, M., et al. (2011). A unique iron-sulfur cluster is crucial for oxygen tolerance of a [NiFe]-hydrogenase. Nat. Chem. Biol. 7 (5), 310–318. doi:10.1038/nchembio.555
Greco, C., Bruschi, M., Fantucci, P., Ryde, U., and De Gioia, L. (2011). Mechanistic and physiological implications of the interplay among iron-sulfur clusters in [FeFe]-hydrogenases. A QM/MM perspective. J. Am. Chem. Soc. 133 (46), 18742–18749. doi:10.1021/ja205542k
Greening, C., Biswas, A., Carere, C. R., Jackson, C. J., Taylor, M. C., Stott, M. B., et al. (2016). Genomic and metagenomic surveys of hydrogenase distribution indicate H2 is a widely utilised energy source for microbial growth and survival. ISME J. 10 (3), 761–777. doi:10.1038/ismej.2015.153
Hondal, R. J., Marino, S. M., and Gladyshev, V. N. (2013). Selenocysteine in thiol/disulfide-like exchange reactions. Antioxid. Redox Signal 18 (13), 1675–1689. doi:10.1089/ars.2012.5013
Kertess, L., Wittkamp, F., Sommer, C., Esselborn, J., Rüdiger, O., Reijerse, E. J., et al. (2017). Chalcogenide substitution in the [2Fe] cluster of [FeFe]-hydrogenases conserves high enzymatic activity. Dalton Trans. 46 (48), 16947–16958. doi:10.1039/c7dt03785f
Khanna, N., and Lindblad, P. (2015). Cyanobacterial hydrogenases and hydrogen metabolism revisited: Recent progress and future prospects. Int. J. Mol. Sci. 16 (5), 10537–10561. doi:10.3390/ijms160510537
Kisgeropoulos, E. C., Bharadwaj, V. S., Mulder, D. W., and King, P. W. (2022). The contribution of proton-donor pKa on reactivity profiles of [FeFe]-hydrogenases. Front. Microbiol. 13, 903951. doi:10.3389/fmicb.2022.903951
Kleinhaus, J. T., Wittkamp, F., Yadav, S., Siegmund, D., and Apfel, U. P. (2021). [FeFe]-Hydrogenases: Maturation and reactivity of enzymatic systems and overview of biomimetic models. Chem. Soc. Rev. 50 (3), 1668–1784. doi:10.1039/d0cs01089h
Kosourov, S., Böhm, M., Senger, M., Berggren, G., Stensjö, K., Mamedov, F., et al. (2021). Photosynthetic hydrogen production: Novel protocols, promising engineering approaches and application of semi-synthetic hydrogenases. Physiol. Plant 173 (2), 555–567. doi:10.1111/ppl.13428
Krahn, N., Fischer, J. T., and Söll, D. (2020). Naturally occurring tRNAs with non-canonical structures. Front. Microbiol. 11, 596914. doi:10.3389/fmicb.2020.596914
Leibundgut, M., Frick, C., Thanbichler, M., Böck, A., and Ban, N. (2005). Selenocysteine tRNA-specific elongation factor SelB is a structural chimaera of elongation and initiation factors. EMBO J. 24 (1), 11–22. doi:10.1038/sj.emboj.7600505
Li, F., Lutz, P. B., Pepelyayeva, Y., Arnér, E. S., Bayse, C. A., and Rozovsky, S. (2014). Redox active motifs in selenoproteins. Proc. Natl. Acad. Sci. U. S. A. 111 (19), 6976–6981. doi:10.1073/pnas.1319022111
Liew, F., Martin, M. E., Tappel, R. C., Heijstra, B. D., Mihalcea, C., and Köpke, M. (2016). Gas fermentation-a flexible platform for commercial scale production of low-carbon-fuels and chemicals from waste and renewable feedstocks. Front. Microbiol. 7, 694. doi:10.3389/fmicb.2016.00694
Liew, F., Henstra, A. M., Kӧpke, M., Winzer, K., Simpson, S. D., and Minton, N. P. (2017). Metabolic engineering of Clostridium autoethanogenum for selective alcohol production. Metab. Eng. 40, 104–114. doi:10.1016/j.ymben.2017.01.007
Lubitz, W., Ogata, H., Rüdiger, O., and Reijerse, E. (2014). Hydrogenases. Chem. Rev. 114 (8), 4081–4148. doi:10.1021/cr4005814
Maroney, M. J., and Hondal, R. J. (2018). Selenium versus sulfur: Reversibility of chemical reactions and resistance to permanent oxidation in proteins and nucleic acids. Free Radic. Biol. Med. 127, 228–237. doi:10.1016/j.freeradbiomed.2018.03.035
Marques, M. C., Coelho, R., De Lacey, A. L., Pereira, I. A., and Matias, P. M. (2010). The three-dimensional structure of [NiFeSe] hydrogenase from Desulfovibrio vulgaris Hildenborough: A hydrogenase without a bridging ligand in the active site in its oxidised, "as-isolated" state. J. Mol. Biol. 396 (4), 893–907. doi:10.1016/j.jmb.2009.12.013
Marques, M. C., Tapia, C., Gutiérrez-Sanz, O., Ramos, A. R., Keller, K. L., Wall, J. D., et al. (2017). The direct role of selenocysteine in [NiFeSe] hydrogenase maturation and catalysis. Nat. Chem. Biol. 13 (5), 544–550. doi:10.1038/nchembio.2335
Miller, C., Bröcker, M. J., Prat, L., Ip, K., Chirathivat, N., Feiock, A., et al. (2015). A synthetic tRNA for EF-Tu mediated selenocysteine incorporation in vivo and in vitro. FEBS Lett. 589 (17), 2194–2199. doi:10.1016/j.febslet.2015.06.039
Mony, M.-C., and Larras-Regard, E. (2000). Renal bioavailability of selenium after supplementation with different forms of selenium: Ion probe and mass spectrometry study. J. Trace Elem. Exp. Med. 13 (4), 367–380. doi:10.1002/1520-670x(2000)13:4<367::aid-jtra5>3.0.co;2-3
Morra, S., Arizzi, M., Valetti, F., and Gilardi, G. (2016). Oxygen stability in the new [FeFe]-hydrogenase from Clostridium beijerinckii SM10 (CbA5H). Biochemistry 55 (42), 5897–5900. doi:10.1021/acs.biochem.6b00780
Morra, S. (2022). Fantastic [FeFe]-hydrogenases and where to find them. Front. Microbiol. 13, 853626. doi:10.3389/fmicb.2022.853626
Mukai, T., Vargas-Rodriguez, O., Englert, M., Tripp, H. J., Ivanova, N. N., and Rubin, E. M. (2017). Transfer RNAs with novel cloverleaf structures. Nucleic Acids Res. 45 (5), 2776–2785. doi:10.1093/nar/gkw898
Mukai, T., Sevostyanova, A., Suzuki, T., Fu, X., and Söll, D. (2018). A facile method for producing selenocysteine-containing proteins. Angew. Chem. Int. Ed. Engl. 57 (24), 7215–7219. doi:10.1002/anie.201713215
Mulder, D. W., Peters, J. W., and Raugei, S. (2021). Catalytic bias in oxidation-reduction catalysis. Chem. Commun. (Camb) 57 (6), 713–720. doi:10.1039/d0cc07062a
Mulder, D. W., Shepard, E. M., Meuser, J. E., Joshi, N., King, P. W., Posewitz, M. C., et al. (2011). Insights into [FeFe]-hydrogenase structure, mechanism, and maturation. Structure 19 (8), 1038–1052. doi:10.1016/j.str.2011.06.008
Nicolet, Y., Piras, C., Legrand, P., Hatchikian, C. E., and Fontecilla-Camps, J. C. (1999). Desulfovibrio desulfuricans iron hydrogenase: The structure shows unusual coordination to an active site Fe binuclear center. Structure 7 (1), 13–23. doi:10.1016/s0969-2126(99)80005-7
Noth, J., Esselborn, J., Güldenhaupt, J., Brünje, A., Sawyer, A., Apfel, U. P., et al. (2016). [FeFe]-hydrogenase with chalcogenide substitutions at the H-Cluster maintains full H2 evolution activity. Angew. Chem. Int. Ed. Engl. 55 (29), 8536–8540. doi:10.1002/ange.201511896
Ogata, H., Lubitz, W., and Higuchi, Y. (2016). Structure and function of [NiFe] hydrogenases. J. Biochem. 160 (5), 251–258. doi:10.1093/jb/mvw048
Parkin, A., Goldet, G., Cavazza, C., Fontecilla-Camps, J. C., and Armstrong, F. A. (2008). The difference a Se makes? Oxygen-tolerant hydrogen production by the [NiFeSe]-hydrogenase from Desulfomicrobium baculatum. J. Am. Chem. Soc. 130 (40), 13410–13416. doi:10.1021/ja803657d
Peters, J. W., Lanzilotta, W. N., Lemon, B. J., and Seefeldt, L. C. (1998). X-ray crystal structure of the Fe-only hydrogenase (CpI) from Clostridium pasteurianum to 1.8 Angstrom resolution. Science 282 (5395), 1853–1858. doi:10.1126/science.282.5395.1853
Peters, J. W., Schut, G. J., Boyd, E. S., Mulder, D. W., Shepard, E. M., Broderick, J. B., et al. (2015). [FeFe]- and [NiFe]-hydrogenase diversity, mechanism, and maturation. Biochim. Biophys. Acta 1853 (6), 1350–1369. doi:10.1016/j.bbamcr.2014.11.021
Popescu, C. V., and Münck, E. (1999). Electronic structure of the H cluster in [Fe]-hydrogenases. J. Am. Chem. Soc. 121 (34), 7877–7884. doi:10.1021/ja991243y
Prabhakar, A., Krahn, N., Zhang, J., Vargas-Rodriguez, O., Krupkin, M., and Fu, Z. (2022). Uncovering translation roadblocks during the development of a synthetic tRNA. Nucleic Acids Res. 50 (18), 10201–10211. doi:10.1093/nar/gkac576
Rodriguez-Maciá, P., Pawlak, K., Rüdiger, O., Reijerse, E. J., Lubitz, W., and Birrell, J. A. (2017). Intercluster redox coupling influences protonation at the H-cluster in [FeFe] hydrogenases. J. Am. Chem. Soc. 139 (42), 15122–15134. doi:10.1021/jacs.7b08193
Schilter, D., Camara, J. M., Huynh, M. T., Hammes-Schiffer, S., and Rauchfuss, T. B. (2016). Hydrogenase enzymes and their synthetic models: The role of metal hydrides. Chem. Rev. 116 (15), 8693–8749. doi:10.1021/acs.chemrev.6b00180
Schrader, J. M., Sakes, M. E., and Uhlenbeck, O. C. (2011). “The specific interaction between aminoacyl-tRNAs and elongation factor Tu,” in Ribosomes. Editor M. V. Rodnina (Springer, Vienna: Wien and New York Spring-Verlag), 189–198. doi:10.1007/978-3-7091-0215-2_15
Schuchmann, K., Chowdhury, N. P., and Müller, V. (2018). Complex multimeric [FeFe] hydrogenases: Biochemistry, physiology and new opportunities for the hydrogen economy. Front. Microbiol. 9, 2911. doi:10.3389/fmicb.2018.02911
Schuchmann, K., and Müller, V. (2013). Direct and reversible hydrogenation of CO2 to formate by a bacterial carbon dioxide reductase. Science 342 (6164), 1382–1385. doi:10.1126/science.1244758
Shafaat, H. S., Rüdiger, O., Ogata, H., and Lubitz, W. (2013). [NiFe] hydrogenases: A common active site for hydrogen metabolism under diverse conditions. Biochim. Biophys. Acta 1827 (8-9), 986–1002. doi:10.1016/j.bbabio.2013.01.015
Shima, S., Pilak, O., Vogt, S., Schick, M., Stagni, M. S., Meyer-Klaucke, W., et al. (2008). The crystal structure of [Fe]-hydrogenase reveals the geometry of the active site. Science 321 (5888), 572–575. doi:10.1126/science.1158978
Shomura, Y., Yoon, K. S., Nishihara, H., and Higuchi, Y. (2011). Structural basis for a [4Fe-3S] cluster in the oxygen-tolerant membrane-bound [NiFe]-hydrogenase. Nature 479 (7372), 253–256. doi:10.1038/nature10504
Silakov, A., Reijerse, E. J., Albracht, S. P., Hatchikian, E. C., and Lubitz, W. (2007). The electronic structure of the H-cluster in the [FeFe]-hydrogenase from Desulfovibrio desulfuricans: A Q-band 57Fe-endor and HYSCORE study. J. Am. Chem. Soc. 129 (37), 11447–11458. doi:10.1021/ja072592s
Simmons, T. R., Berggren, G., Bacchi, M., Fontecave, M., and Artero, V. (2014). Mimicking hydrogenases: From biomimetics to artificial enzymes. Coord. Chem. Rev. 270-271, 127–150. doi:10.1016/j.ccr.2013.12.018
Stock, T., and Rother, M. (2009). Selenoproteins in archaea and gram-positive bacteria. Biochim. Biophys. Acta 1790 (11), 1520–1532. doi:10.1016/j.bbagen.2009.03.022
Stripp, S. T., Duffus, B. R., Fourmond, V., Léger, C., Leimkühler, S., Hirota, S., et al. (2022). Second and outer coordination sphere effects in nitrogenase, hydrogenase, formate dehydrogenase, and CO dehydrogenase. Chem. Rev. 122 (14), 11900–11973. doi:10.1021/acs.chemrev.1c00914
Thyer, R., Robotham, S. A., Brodbelt, J. S., and Ellington, A. D. (2015). Evolving tRNASec for efficient canonical incorporation of selenocysteine. J. Am. Chem. Soc. 137 (1), 46–49. doi:10.1021/ja510695g
Vansuch, G. E., Wu, C. H., Haja, D. K., Blair, S. A., Chica, B., Johnson, M. K., et al. (2020). Metal-ligand cooperativity in the soluble hydrogenase-1 from Pyrococcus furiosus. Chem. Sci. 11 (32), 8572–8581. doi:10.1039/d0sc00628a
Vignais, P. M., and Billoud, B. (2007). Occurrence, classification, and biological function of hydrogenases: An overview. Chem. Rev. 107 (10), 4206–4272. doi:10.1021/cr050196r
Volbeda, A., Amara, P., Iannello, M., De Lacey, A. L., Cavazza, C., and Fontecilla-Camps, J. C. (2013). Structural foundations for the O2 resistance of Desulfomicrobium baculatum [NiFeSe]-hydrogenase. Chem. Commun. (Camb) 49 (63), 7061–7063. doi:10.1039/c3cc43619e
Volbeda, A., Charon, M. H., Piras, C., Hatchikian, E. C., Frey, M., and Fontecilla-Camps, J. C. (1995). Crystal structure of the nickel-iron hydrogenase from Desulfovibrio gigas. Nature 373 (6515), 580–587. doi:10.1038/373580a0
Winkler, M., Duan, J., Rutz, A., Felbek, C., Scholtysek, L., Lampret, O., et al. (2021). A safety cap protects hydrogenase from oxygen attack. Nat. Commun. 12 (1), 756. doi:10.1038/s41467-020-20861-2
Winkler, M., Esselborn, J., and Happe, T. (2013). Molecular basis of [FeFe]-hydrogenase function: An insight into the complex interplay between protein and catalytic cofactor. Biochim. Biophys. Acta 1827 (8-9), 974–985. doi:10.1016/j.bbabio.2013.03.004
Keywords: Selenocysteine, cell factories, biofuel, allo-tRNA, Selenoprotein, C1 microbes, hydrogenase, hydrogen production
Citation: Patel A, Mulder DW, Söll D and Krahn N (2022) Harnessing selenocysteine to enhance microbial cell factories for hydrogen production. Front. Catal. 2:1089176. doi: 10.3389/fctls.2022.1089176
Received: 07 November 2022; Accepted: 13 December 2022;
Published: 22 December 2022.
Edited by:
Sanjay Kumar Singh Patel, Konkuk University, South KoreaReviewed by:
Zachary Tonzetich, University of Texas at San Antonio, United StatesIlka U. Heinemann, Western University, Canada
Copyright © 2022 Patel, Mulder, Söll and Krahn. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Natalie Krahn, bmF0YWxpZS5rcmFobkB5YWxlLmVkdQ==