
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
GENERAL COMMENTARY article
Front. Conserv. Sci., 22 August 2024
Sec. Plant Conservation
Volume 5 - 2024 | https://doi.org/10.3389/fcosc.2024.1437922
This article is part of the Research TopicReconciling Nature Conservation and Sustainability of Tropical EcosystemsView all 5 articles
 Lindsey Gillson1,2*†
Lindsey Gillson1,2*† Estelle Razanatsoa2†
Estelle Razanatsoa2† Andriantsilavo Hery Isandratana Razafimanantsoa2†
Andriantsilavo Hery Isandratana Razafimanantsoa2† Malika Virah-Sawmy3†
Malika Virah-Sawmy3† Anneli Ekblom4†
Anneli Ekblom4†A Commentary on
Commentary: The role of palaeoecology in reconciling biodiversity conservation, livelihoods and carbon storage in Madagascar
By Joseph GS and Seymour CL (2024). Front. Conserv. Sci. 5:1389498. doi: 10.3389/fcosc.2024.1389498
We welcome Seymour and Joseph’s commentary, which acknowledges the complexity of Malagasy landscapes (Joseph and Seymour, 2024). The multiple landscape elements that are present throughout most of the island provide a wealth of biodiversity habitats and ecosystem services (Bond et al., 2023; Silander et al., 2024). The climatic gradient from wettest in the northeast to the dry south west, as well as the interacting effects of fire and topography, means that only the northeast contains true forest, in the sense of a closed or near-continuous canopy (Straka, 1996; Moat and Smith, 2007). The rest of the island comprises a mosaic of dry forest, open woodland, savanna, ericoid heathland, and grassland, with tree cover varying from 0% to 75%, and patches of closed forest, largely confined to ravines and riverine areas (Robinson et al., 2021; Silander et al., 2024). The distinction between these vegetation types is important since they house different (though sometimes overlapping) suites of biodiversity. Palaeoecological work has shown the existence of these open, grassy, and mosaic ecosystems, as well as the presence of fire, long before human arrival in Madagascar (Burney, 1987; Gasse and Van Campo, 2001; Bond et al., 2008; Virah-Sawmy et al., 2009b; Vorontsova et al., 2016; Hackel et al., 2018; Vorontsova et al., 2020; Razafimanantsoa, 2022).
While some open grasslands have been derived from anthropogenic clearing and burning, others are ancient and rich in endemic species (Bond et al., 2019; Vetter, 2020; García-Dory et al., 2021; Lehmann et al., 2021). Understanding the landscape history and how it evolved with changing land use and environment is critical in planning appropriate restoration and management. The assumption that open and mosaic landscapes are degraded forests is still a dominant narrative in Madagascar and has left them vulnerable to afforestation using exotic species such as pine and Eucalyptus (Carriere and Randriambanona, 2007; Charles and Dukes, 2008; Stickler et al., 2009; Bond, 2019; Kull et al., 2019; Randriambanona et al., 2019). Despite growing awareness of the complexity of landscape mosaics, deforestation rates in Madagascar are still frequently measured relative to the erroneous baseline of 100% forest cover (Huff and Orengo, 2020; Gillson et al., 2023), or in other cases based on an overestimation of original forest cover (McConnell and Kull, 2014; Amelot, 2017). In the twentieth century, the binary categorization of forest versus all other vegetation types as “non forest” led to inflated estimates of tree cover losses. The combination of inflated forest loss and underlying assumptions regarding degradation in turn impacted communities who were then blamed for landscape mismanagement (Virah-Sawmy and Ebeling, 2010). We give some examples of the effects of these misunderstandings in Gillson et al. (2023).
Joseph and Seymour (2020, 2021) (hereafter J&S) set out to investigate the longevity of open grasslands in the Highlands of Madagascar, as proposed by Bond et al. (2008) through an inventory of species living in this environment, and came to the conclusion that treeless open grasslands were formerly restricted in distribution and are mainly recent and anthropogenic. The reason we used their paper as an example of the problem of a continued binary way of thinking around forest and non-forest was because, in their analyses, they divided taxa into two categories following IUCN species assessment. For example:
1) In Joseph and Seymour, 2020, Figure 1, there is a comparison of the proportion of endemic Madagascan vertebrate groups, Australian diprotodonts, polyprotodonts, and endemic rodents, as adapted to predominantly forest, grassland, or both habitats. To construct this table, J&S “designat[ed] species as either ‘grassland’ (grassland-limited) or ‘not’ (forest or either habitat)”, i.e., for both countries, the figures for vertebrates add up to 100%, suggesting that vertebrates must be adapted to either forest or grassland or both habitats. It follows then that any species that is adapted to heathland, woodland, savanna, thicket, or any other habitat must have been assigned to the forest or “both” categories. No other habitats were considered or acknowledged, which was why we described the classification as binary.
2) Similarly, in Joseph and Seymour, 2021, Figure 2, only two categories are shown, forest-limited or grassland-limited. In the underlying data (Supplementary Table S3C), only these two categories are used and no other habitat types are considered, which is again why we described the classification as binary.
These strict, binary allocations to forest and/or grassland habitats are problematic, because, as acknowledged in Joseph and Seymour, 2024, there are multiple vegetation types present in Malagasy landscapes, most of which comprise a matrix of grassy and woody elements. True forests are confined to the north east (Straka, 1996; Moat and Smith, 2007), elsewhere found along riverine areas and ravines protected from fire. The dichotomy of open treeless grassland versus true forest when assigning affinities for animals and plants is ecologically inaccurate. A suite of species can exist across the continuum of grassy, open, and mosaic ecosystems, characterized by the presence of C4 grasses, with degrees of tree cover varying from 0% to 75% (Robinson et al., 2021; Silander et al., 2024). The distinction between true forests and the continuum of other habitats is nicely illustrated in Silander et al. (2024) (Figure 1), reproduced here. This figure demonstrates how the attribution of species to open treeless grasslands versus “forests” is bound to encompass only a very small subset of the suite of species that are adapted to open and mosaic habitats, artificially inflating the number of “forest” species, which in J&S’s analyses (Joseph and Seymour, 2020, 2021) covers the full suite of other habitats that by default must include, heathlands, savannas, open woodlands, and dry forests with a grassy understory. It is in this context that we cited J&S as perpetuating a false dichotomy in our paper (Gillson et al., 2023).
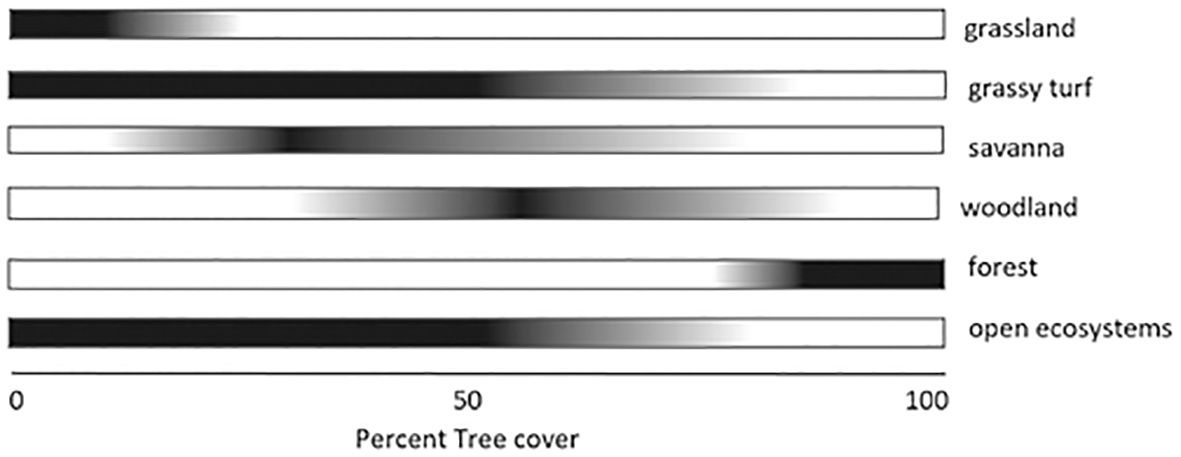
Figure 1. Schematic characterization of the continuous and fuzzy nature of defining open, grassy ecosystems versus closed forest ecosystems on a scale of continuous variation in tree cover. Reproduced from Silander et al., 2024, Creative Commons License.
The figures described above (Joseph and Seymour, 2020, 2021) are problematic as they conflate savanna, woodland heathland, and forest, which, as J&S point out in their current commentary (Joseph and Seymour, 2024), have very distinct ecologies. In contrast to the figures, the text of the Joseph and Seymour, 2020, 2021 papers and their current commentary (Joseph and Seymour 2024) acknowledges the presence of landscape mosaics. Habitats other than forest and grassland are shown elsewhere in their work. For example, five vegetation types are mentioned in J&S 2020, Figure 3A and in Figure 3B, and the woodland category is treated separately from forest (as it should be).
In a later paper (Joseph et al., 2022), J&S suggest that the lack of correlation between rainfall and tree cover provides evidence for the effects of anthropogenic fire. However, this decoupling of climate and tree cover is well known in the literature and is explained by the interacting effects of rainfall seasonality, fire, and herbivory which all limit tree cover to varying degrees and cause a non-linear relationship between tree cover and rainfall (e.g., see Bond et al., 2005; Sankaran et al., 2005; Staver et al., 2011; Higgins and Scheiter, 2012). Savannas are likely to occur in areas where rainfall is seasonal (as in the Highlands of Madagascar), as grass biomass accumulates in the wet season, dries out, and burns in the dry season, regardless of human sources of ignition and there being enough annual rainfall to support forest cover. Humans are capable of manipulating this relationship to encourage favoured habitat types (e.g., see Whitlock et al., 2010); however, attempts to suppress fire completely in seasonally rainy savannas have proved largely futile (e.g., see van Wilgen et al., 2004).
The main point in Gillson et al. (2023) is to stress the importance of understanding landscape history and the potential contribution of palaeoecology, especially in countering afforestation schemes that are planned for open and mosaic landscapes erroneously perceived as degraded (Gillson et al., 2023). We do not dispute that deforestation is occurring in some areas of Madagascar (Vieilledent et al., 2018). We also acknowledge the interacting effects of climate and fire from ca. 1,000 years ago that together increased the proportion of open grassy ecosystems compared with woody habitats. Our intention in advocating the use of palaeoecology is to help find out which areas were formerly forests, which were not, and what the landscape mosaic looked like prior to intensive human impact over the past 1,000 years. None of this runs contrary to the commentary on mosaic landscapes (Joseph and Seymour, 2024) but does highlight our justifiable concern with the figures in the papers we cited (Joseph and Seymour, 2020, 2021). We find some of the generalized statements (e.g., “several lines of evidence affirm the highland grassland is young” (Joseph and Seymour, 2020) that appear in their work problematic. The inference that almost all grasslands and savannas in the Central Highlands are of anthropogenic origin can potentially open the way for afforestation of the Highlands, as anthropogenically derived grasslands are perceived to be of little ecological value (e.g., see Joseph et al., 2022). The risk is that such habitats are assessed as fair targets for introduced fast-growing trees such as Eucalyptus and pine plantation instead of species that are native and historically beneficial for communities living around these areas.
Statements attesting to the restricted range and anthropogenic origin of grassy ecosystems do not take into account the range of ancient grasslands (decisively shown in palaeoecological records), grasslands with trees, woodlands, heathlands, and other open habitats (also present in prehuman paleoecological records). We respectfully suggest that if S&J want to avoid misunderstanding of their work, they need to acknowledge the range of vegetation types across the Malagasy highlands. In the case of the figures in Joseph and Seymour, 2020, 2021, we suggest dropping the strict criteria of treeless grassland and including the continuum of habitats mentioned in Silander (Silander et al., 2024). Woodland- and savanna-adapted species need to be removed from the “forest” category. A more representative suite of habitats, as outlined in Silander et al. (2024), that includes true forest as the outlier and the range of open and mosaic habitat types that includes grassland (with or without some trees), savanna, heathland, open woodland will be ecologically more realistic and more in harmony with their current commentary (Jospeh and Seymour, 2024).
Meanwhile, our main point stands; palaeoecological data are invaluable to understanding the history of Madagascar’s landscapes but remain relatively scarce (Gillson et al., 2023). In the drier areas, where these debates are most contentious, pollen preservation is often poor and chronologies discontinuous due to hiatuses in sediment deposition. Nevertheless, the records we have provide fascinating insights into the past landscapes of Madagascar (e.g., see Virah-Sawmy et al., 2009a, 2009b, 2016; Razanatsoa et al., 2021, 2022) and the changes taken place prior to human arrival and the interacting effects of climate and humans in shaping the landscapes we see today. Further palaeoecological work and careful interpretation based on knowledge of modern ecological affinities of pollen taxa (see Razafimanantsoa and Razanatsoa, 2024) will help to illuminate questions of how Madagascar’s mosaic landscapes looked and how forested and open landscapes have fluctuated over time in response to changing climate and disturbance by fire and herbivores and since the late Holocene, people. This information will be crucial in halting inappropriate afforestation schemes that utilize non-native species and in ensuring the conservation and restoration of open and mosaic habitats throughout the island.
Finally, the response to our paper and the ensuing academic exchange contribute to the research process, allowing us to refine our arguments and also clarify potential misunderstandings. Such exchanges raise questions regarding how to refine integrated methodologies that address the complexity of Madagascar’s landscapes, and provide a realistic underpinning for the management and restoration of landscapes based on site-specific and historic data. While there appears to be some consensus over complexity of Madagascar’s landscapes, narratives of degradation and economic imperatives continue to shape deforestation and reforestation research questions and methodologies (Gillson et al., 2024). New approaches are needed that are rooted in better exchanges between scientists, historians, and local communities (Gillson et al., 2024). Such exchange will ultimately lead to a better framework of understanding, which will contribute to the management of the island’s biodiversity, improving ecosystem services in those diverse landscapes that support the needs of its population.
LG: Conceptualization, Writing – original draft, Writing – review & editing. ER: Writing – review & editing. AR: Writing – review & editing. MV-S: Writing – review & editing. AE: Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Amelot X. (2017). “Dire la Nature à Madagascar: des mythes contre des cartes,” in Dire l'océan indien. Eds. Combeau Y., Gallat T., Rolland Y. (Université de la Réunion, Epica Editions, Saint-Denis, Réunion), 32–53.
Bond W. J. (2019). Open ecosystems: ecology and evolution beyond the forest edge (Oxford University Press). doi: 10.1093/oso/9780198812456.001.0001
Bond W. J., Silander J. A. Jr., Ranaivonasy J., Ratsirarson J. (2008). The Antiquity of Madagascar's grasslands and the rise of C4 biomes. J. Biogeography 35, 1743–1758. doi: 10.1111/j.1365-2699.2008.01923.x
Bond W. J., Silander J. A., Ratsirarson J. (2023). Madagascar's grassy biomes are ancient and there is much to learn about their ecology and evolution. J. Biogeography 50, 614–621. doi: 10.1111/jbi.14494
Bond W. J., Stevens N., Midgley G. F., Lehmann C. E. (2019). The trouble with trees: afforestation plans for Africa. Trends Ecol. Evol. 34, 963–965. doi: 10.1016/j.tree.2019.08.003
Bond W. J., Woodward F. I., Midgley G. F. (2005). The global distribution of ecosystems in a world without fire. New Phytol. 165, 525–538. doi: 10.1111/j.1469-8137.2004.01252.x
Burney D. A. (1987). “Pre-settlement vegetation changes at Lake Tritrivakely, Madagascar,” Palaeoecology of Africa and the surrounding islands (Rotterdam, Netherlands), 357–381.
Carriere S. M., Randriambanona H. (2007). Biodiversité introduite et autochtone: antagonisme ou complémentarité? Le cas l'eucalyptus à Madagascar. BOIS FORETS Des. TROPIQUES 292, 5–21. doi: 10.19182/bft2007.292.a20349
Charles H., Dukes J. S. (2008). “Impacts of invasive species on ecosystem services,” in Biological Invasions (Springer), 217–237.
García-Dory F., Houzer E., Scoones I. (2021). Livestock and climate justice: Challenging mainstream policy narratives. IDS Bulletin 53(4). doi: 10.19088/1968-2021.128
Gasse F., Van Campo E. (2001). Late Quaternary environmental changes from a pollen and diatom record in the southern tropics (Lake Tritrivakely, Madagascar). Palaeogeography,Palaeoclimatology,Palaeoecology 167, 287–308. doi: 10.1016/S0031-0182(00)00242-X
Gillson L., Hoffman M. T., Gell P. A., Ekblom A., Bond W. J. (2024). Trees, carbon, and the psychology of landscapes. Trends Ecol. Evol 39 (4). doi: 10.1016/j.tree.2023.11.008
Gillson L., Razanatsoa E., Razafimanantsoa A. H. I., Virah-Sawmy M., Ekblom A. (2023). The role of palaeoecology in reconciling biodiversity conservation, livelihoods and carbon storage in Madagascar. Front. Conserv. Sci. 4. doi: 10.3389/fcosc.2023.1286459
Hackel J., Vorontsova M. S., Nanjarisoa O. P., Hall R. C., Razanatsoa J., Malakasi P., et al. (2018). Grass diversification in Madagascar: in situ radiation of two large C3 shade clades and support for a Miocene to Pliocene origin of C4 grassy biomes. J. Biogeography 45, 750–761. doi: 10.1111/jbi.13147
Higgins S. I., Scheiter S. (2012). Atmospheric CO2 forces abrupt vegetation shifts locally, but not globally. Nat. advance Online publication. 408, 209–212. doi: 10.1038/nature11238
Huff A., Orengo Y. (2020). Resource warfare, pacification and the spectacle of ‘green’development: Logics of violence in engineering extraction in southern Madagascar. Political Geogr. 81, 102195. doi: 10.1016/j.polgeo.2020.102195
Joseph G. S., Rakotoarivelo A. R., Seymour C. L. (2022). Tipping points induced by palaeo-human impacts can explain presence of savannah in Malagasy and global systems where forest is expected. Proc. R. Soc. B 289, 20212771. doi: 10.1098/rspb.2021.2771
Joseph G. S., Seymour C. L. (2020). Madagascan highlands: originally woodland and forest containing endemic grasses, not grazing-adapted grassland. Proc. R. Soc. B 287, 20201956. doi: 10.1098/rspb.2020.1956
Joseph G. S., Seymour C. L. (2021). The unlikely A’ntiquity of Madagascar's grasslands’: Disproportionately forest-limited endemic fauna support anthropogenic transformation from woodland. J. Biogeography 48, 2111–2115. doi: 10.1111/jbi.14132
Joseph G. S., Seymour C. L. (2024). Commentary: The role of palaeoecology in reconciling biodiversity conservation, livelihoods and carbon storage in Madagascar. Front. Conserv. Sci. 5, 1389498. doi: 10.3389/fcosc.2024.1389498
Kull C. A., Harimanana S. L., Andrianoro A. R., Rajoelison L. G. (2019). Divergent perceptions of the ‘neo-Australian’forests of lowland eastern Madagascar: Invasions, transitions, and livelihoods. J. Environ. Manage. 229, 48–56. doi: 10.1016/j.jenvman.2018.06.004
Lehmann C. E., Solofondranohatra C. L., Vorontsova M. S. (2021). Beyond ancient versus anthropogenic for Madagascar's grassy ecosystems. A Reply to: Crowley. Proc. R. Soc. B 288, 20210388. doi: 10.1098/rspb.2021.0388
McConnell W. J., Kull C. A. (2014). “Deforestation in Madagascar: Debates over the island's forest cover and challenges of measuring forest change,” in Conservation and environmental management in Madagascar (Abingdon Oxon UK: Routledge), 67–104.
Moat J., Smith P. (2007). “Atlas of the vegetation of Madagascar,” in The Royal Botanic Gardens (Richmond, Surrey: Royal Botanic Garden-Kew).
Randriambanona H., Randriamalala J. R., Carrière S. M. (2019). Native forest regeneration and vegetation dynamics in non-native Pinus patula tree plantations in Madagascar. For. Ecol. Manage. 446, 20–28. doi: 10.1016/j.foreco.2019.05.019
Razafimanantsoa A. H. I. (2022). “Holocene vegetation history and environmental change in the forest-grassland mosaic of the Central Highlands of Madagascar,” in Doctoral. University of Cape Town (South Africa: Cape Town). doi: 10.1177%2F09596836211041731
Razafimanantsoa A. H. I., Razanatsoa E. (2024). Modern pollen rain reveals differences across forests, open and mosaic landscapes in Madagascar. Plants People Planet 6, 729–742. doi: 10.1002/ppp3.10487
Razanatsoa E., Gillson L., Virah-Sawmy M., Woodborne S. (2021). Pollen records of the 14th and 20th centuries AD from Lake Tsizavatsy in southwest Madagascar. Palaeoecol. Afr. 35, 309–315.
Razanatsoa E., Gillson L., Virah-Sawmy M., Woodborne S. (2022). Synergy between climate and human land-use maintained open vegetation in Southwest Madagascar over the last millennium. Holocene 31 (1-2), 57-69. doi: 10.1177/09596836211041731
Robinson J. R., Rowan J., Barr W. A., Sponheimer M. (2021). Intrataxonomic trends in herbivore enamel δ13C are decoupled from ecosystem woody cover. Nat. Ecol. Evol. 5, 995–1002. doi: 10.1038/s41559-021-01455-7
Sankaran M., Hanan N. P., Scholes J. R., Ratman J., Augustine D. J., Cade B. S., et al. (2005). Determinants of woody cover in African savannas. Nature 438, 846–849. doi: 10.1038/nature04070
Silander J. A. Jr., Bond W. J., Ratsirarson J. (2024). The grassy ecosystems of Madagascar in context: Ecology, evolution, and conservation. Plants People Planet 6, 94–115. doi: 10.1002/ppp3.10417
Staver A. C., Archibald S., Levin S. (2011). Tree cover in sub-Saharan Africa: Rainfall and fire constrain forest and savanna as alternative stable states. Ecology 92, 1063–1072. doi: 10.1890/10-1684.1
Stickler C. M., Nepstad D. C., Coe M. T., McGrath D. G., Rodrigues H. O., Walker W. S., et al. (2009). The potential ecological costs and cobenefits of REDD: a critical review and case study from the Amazon region. Global Change Biol. 15, 2803–2824. doi: 10.1111/j.1365-2486.2009.02109.x
Straka H. (1996). Histoire de la végétation de Madagascar oriental dans les derniers 100 millénaires. BiogeHographie Madagascar ORSTOM Paris 37.
van Wilgen B. W., Govener N., Biggs H. C., Ntsala D., Funda X. N. (2004). Response of savanna fire regimes to changing fire management policies in a large African national park. Conserv. Biol. 18, 1533–1540. doi: 10.1111/j.1523-1739.2004.00362.x
Vetter S. (2020). With power comes responsibility–A rangelands perspective on forest landscape restoration. Front. Sustain. Food Syst. 4, 225. doi: 10.3389/fsufs.2020.549483
Vieilledent G., Grinand C., Rakotomalala F. A., Ranaivosoa R., Rakotoarijaona J.-R., Allnutt T. F., et al. (2018). Combining global tree cover loss data with historical national forest cover maps to look at six decades of deforestation and forest fragmentation in Madagascar. Biol. Conserv. 222, 189–197. doi: 10.1016/j.biocon.2018.04.008
Virah-Sawmy M., Ebeling J. (2010). The difficult road toward real-world engagement: conservation science and mining in southern Madagascar. Conserv. Lett. 3, 288–289. doi: 10.1111/j.1755-263X.2010.00126.x
Virah-Sawmy M., Gillson L., Gardner C., Anderson A., Clark G., Haberle S. (2016). A landscape vulnerability framework for identifying integrated conservation and adaptation pathways to climate change: the case of Madagascar’s spiny forest. Landscape Ecol. 31, 637-654. doi: 10.1007/s10980-015-0269-2
Virah-Sawmy M., Gillson L., Willis K. J. (2009a). How does spatial heterogeneity influence resilience to climatic changes? Ecol. dynamics southeast Madagascar. Ecol. Monogr. 79, 557–574. doi: 10.1890/08-1210.1
Virah-Sawmy M., Willis K. J., Gillson L. (2009b). Threshold response of Madagascar's littoral forest to sea-level rise. Global Ecol. Biogeography 18, 98–110. doi: 10.1111/j.1466-8238.2008.00429.x
Vorontsova M. S., Besnard G., Forest F., Malakasi P., Moat J., Clayton W. D., et al. (2016). Madagascar's grasses and grasslands: anthropogenic or natural? Proc. R. Soc. B: Biol. Sci. 283, 20152262. doi: 10.1098/rspb.2015.2262
Vorontsova M. S., Lowry P. P., Andriambololonera S. R., Wilmé L., Rasolohery A., Govaerts R., et al. (2020). Inequality in plant diversity knowledge and unrecorded plant extinctions: An example from the grasses of Madagascar. Plants People Planet 3, 45–60. doi: 10.1002/ppp3.10123
Keywords: palaeoecology, fire, deforestation, afforestation, restoration, grasslands
Citation: Gillson L, Razanatsoa E, Razafimanantsoa AHI, Virah-Sawmy M and Ekblom A (2024) Response: Commentary: The role of palaeoecology in reconciling biodiversity conservation, livelihoods and carbon storage in Madagascar. Front. Conserv. Sci. 5:1437922. doi: 10.3389/fcosc.2024.1437922
Received: 24 May 2024; Accepted: 08 July 2024;
Published: 22 August 2024.
Edited by:
Maria M. Romeiras, University of Lisbon, PortugalReviewed by:
Juan Carlos López-Acosta, Universidad Veracruzana, MexicoCopyright © 2024 Gillson, Razanatsoa, Razafimanantsoa, Virah-Sawmy and Ekblom. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lindsey Gillson, bGluZHNleS5naWxsc29uQHlvcmsuYWMudWs=
†ORCID: Lindsey Gillson, orcid.org/0000-0001-9607-6760
Estelle Razanatsoa, orcid.org/0000-0002-7219-1411
Andriantsilavo Hery Isandratana Razafimanantsoa, orcid.org/0000-0002-9933-6991
Malika Virah-Sawmy, orcid.org/0000-0003-3646-5646
Anneli Ekblom, orcid.org/0000-0001-9248-5516
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.