 Flávio M. M. Mota
Flávio M. M. Mota Kyle D. Kittelberger
Kyle D. Kittelberger Cristian Flórez-Paí3
Cristian Flórez-Paí3 Çağan H. Şekercioğlu
Çağan H. Şekercioğlu- 1Applied Ecology and Conservation Lab, Programa de Pós-graduação em Ecologia e Conservação da Biodiversidade, Universidade Estadual de Santa Cruz, Ilhéus, Brazil
- 2School of Biological Sciences, University of Utah, Salt Lake City, UT, United States
- 3Conserve Birds Colombia Program, Fundación Ecológica Los Colibríes de Altaquer (FELCA) - KBA Río Ñambí, Barbacoas, Colombia
- 4Department of Molecular Biology and Genetics, Koç University, Istanbul, Türkiye
Introduction: Climate change poses a significant threat to bird communities, especially forest-dwelling and narrowly distributed species, which are expected to experience severe range contractions and higher extinction risks compared to widely distributed and open-area species. The Chocó region in southwestern Colombia, known for its rich bird endemism, is particularly vulnerable.
Methods: We analyzed potential distribution shifts for 27 endemic and near-endemic bird species in the Chocó region using eBird occurrence records and climate projections. We modeled species distributions under low and high greenhouse gas emission scenarios for 2050 and 2070, comparing these projected distributions to current forested and protected areas to evaluate future conservation needs.
Results: Our findings indicate that nearly all species are projected to lose climate-suitable areas under at least one future scenario, resulting in a regional decline in species richness. Changes in species richness are most pronounced near the Colombia-Ecuador border, suggesting a shift to higher elevations. Notably, the Scarlet-and-white Tanager (Chrysothlypis salmoni) is predicted to suffer the greatest losses in climate-suitable area, both within protected and forested regions.
Discussion: These results highlight the urgency of expanding the protected area network and conserving key forested regions to help species adapt to climate change. By providing projected distribution maps and potential range shifts, our study underscores the importance of modeling future distributions to support conservation strategies for at-risk species and the ecological services they provide in tropical montane regions.
1 Introduction
Climate change is expected to cause population-level modifications to bird communities, leading to contractions, expansions, or shifts in their distributions due to global warming (Şekercioğlu et al., 2008; Wormworth and Şekercioğlu, 2011; Mota et al., 2022). The response to climate change may vary among species, with those inhabiting tropical forests and having restricted distributions expected to experience more drastic reductions in their ranges (Şekercioğlu et al., 2012; de Moraes et al., 2020), as these species are unlikely to keep pace with rapid changes in precipitation and temperature patterns (Román-Palacios and Wiens, 2020). Conversely, generalist species, such as those found in degraded environments or across wide elevational or latitudinal gradients, may experience expansions in their ranges due to the capacity to exploit a broad variety of resources (Sales et al., 2020). As a result, the expected change in species richness in future climate scenarios poses a challenge for conservation planning (Vale et al., 2018), as currently protected areas could become unsuitable for species that play essential ecological roles (Şekercioğlu, 2006; Littlefield et al., 2019), thereby threatening natural systems.
Tropical forests, recognized for being the world’s most biodiverse regions (Raven et al., 2020), have lost most of their original cover to agricultural and pasturelands (Taubert et al., 2018), with the majority of the remaining forest still unprotected (Stan et al., 2024). Climate change may intensify this scenario, as many regions are anticipated to experience increased warmth and aridity (Sales et al., 2020), posing a dire risk to species already threatened by habitat degradation. Additionally, there is uncertainty about whether protected areas can still match new climate suitable areas for species in the future (Littlefield et al., 2019), especially in the understudied tropics (Harris et al., 2011). For example, endemic monkey species in the eastern Amazonian rainforest are projected to vanish from most of their current distribution in the future when considering both forest loss and climate (da Silva et al., 2022). In particular, the Chocó ecoregion in northwestern South America emerges as a crucial conservation priority, as it still retains a substantial amount of forest cover (Anaya et al., 2020). The Chocó Endemic Bird Area (EBA) is located across much of the Pacific slope of the Andes in western Colombia and northwestern Ecuador, and is also part of the larger Tumbes-Chocó-Magdalena biodiversity hotspot that extends from the Darién lowlands of eastern Panama to northwestern Peru (Devenish et al., 2009; Sánchez-Nivicela and Montenegro-Pazmiño, 2022). Recent discoveries of undescribed species and new distributional records of other species in the area emphasize the significance of this region as a hotspot (Guevara Andino and Fernandez-Fernandez, 2020; Amaya-Vallejo et al., 2021; Velandia Perilla et al., 2021). The Chocó is the ninth most biodiverse vegetation hotspot globally, with close to 3,000 endemic species and around 11,000 total plant species in an area covering around 0.2% of the planet’s surface (Pérez-Escobar et al., 2019); among birds, the Chocó has the highest number of endemic bird species of any EBA on the continent, with over 60 species recognized as endemic to the Chocó region (Devenish et al., 2009; Sánchez-Nivicela and Montenegro-Pazmiño, 2022; BirdLife International, 2024b). However, the Chocó has been negatively affected by major habitat loss through deforestation as well as other anthropogenic pressures such as hunting and poaching, making this region a critical conservation priority (Sánchez-Nivicela and Montenegro-Pazmiño, 2022; BirdLife International, 2024a). Furthermore, projections already indicate the risk of local extinctions of bird species in this region in the coming decades (Velásquez-Tibatá et al., 2013), underscoring the urgency of addressing this understudied issue in the region to support future conservation efforts.
Birds, known for their sensitivity to environmental changes, play a crucial role in revealing patterns essential for future biodiversity conservation (Triviño et al., 2018). These avian species contribute to ecosystem functioning by providing a range of ecological services (Şekercioğlu et al., 2016). They facilitate natural forest regeneration by moving seeds between fragments, promote genetic variability through pollination, regulate populations of other species like insects, and prevent disease outbreaks by scavenging on carcasses (Şekercioğlu, 2006; Mäntylä et al., 2011; Ogada et al., 2012; Martínez and García, 2017; Wessinger, 2021). However, recent projections indicate that many bird species may struggle to find suitable climate conditions in the future, leading to a depletion in richness and alterations in community composition (Miranda et al., 2019; Mota et al., 2022), which could compromise these essential ecological services. Additionally, the loss of frugivorous birds may severely limit plants’ capacity to track climate change due to reduced long-distance seed dispersal (Fricke et al., 2022). For Western Hemisphere landbirds, intermediate extinction estimates by Şekercioğlu et al. (2008), based on climate-induced changes in distributions, range from 1.3% (1.1°C warming) to 30.0% (6.4°C warming). Birds confined to the Chocó ecoregion could experience even more drastic reductions in their current distributions compared to species found elsewhere, as their restricted geographic ranges make them particularly vulnerable to losing suitable climate conditions (Velásquez-Tibatá et al., 2013). For example, the loss of climate-suitable areas could result in an approximately 75% reduction in the distribution of the Choco Vireo (Vireo masteri), a species exclusively found in the region, while the loss is expected to be smaller for the Wattled Guan (Aburria aburri; ca. 28%), a species whose distribution also extends into Peru (Velásquez-Tibatá et al., 2013). Furthermore, information on changes in the distribution of Chocó endemic birds is relatively scarce, limiting the implementation of appropriate conservation strategies to ensure their long-term persistence.
In this study, we used ecological niche models to predict the distribution of 27 endemic or near-endemic birds within the Chocó region, with a focus on species occurring in southwest Colombia, under different climate scenarios. We focus on the ranges of these species not only within this targeted Colombian region but also across the Chocó and the entirety of a species’ current distribution. Specifically, we aimed to assess distributional changes and conservation needs of these species, considering current forested and protected areas. We anticipated a reduction in species distributions, with variation across species that may be more sensitive and specialized, and a lower proportion of suitable areas within forested and protected areas in future scenarios. We predict that some Chocó endemics will likely disappear from southwestern Colombia, and we expect to observe heightened population fragmentation across species’ ranges.
2 Methods
2.1 Study area
We assess species distributions, both present and future, across two spatial scales. In our first, broad spatial scale, we examine distributions across the entirety of the Chocó region and, to some extent, part of the Tumbes-Chocó-Magdalena hotspot that occurs in western Colombia and Ecuador (this is due to the fact that some of the species noted as Chocó endemics (BirdLife International, 2024b) have eBird records that fall outside of the established Endemic Bird Area (EBA)). There are various interpretations in the literature of what specifically constitutes the Chocó region (i.e., Pérez-Escobar et al., 2019), so we have chosen to interpret the Chocó area specifically as that which encompasses the designated EBA (Devenish et al., 2009; Sánchez-Nivicela and Montenegro-Pazmiño, 2022). For our second spatial scale, we focused on the region centered around the Nariño department in southwestern Colombia, where there are several mid-elevation reserves in the Chocó that are focused on avian conservation and protection, including Nature Reserve La Nutria-Pimán, La Planada Nature Reserve, OECM (other effective area-based conservation measure) Bangsias BirdLodge, OECM San Antonio, and KBA (Key Biodiversity Area) Río Ñambí.
2.2 Species selection
When determining which species to include in this analysis, we selected 27 species (Table 1) that we have been actively capturing and therefore studying during bird banding operations in the Colombian Chocó, from 2021 to 2024, at the Río Ñambí and Bangsias reserves located near Altaquer in the Barbacoas Municipality. Our selected species therefore largely consisted of the more abundant and less threatened birds within the Chocó (at least via bird banding), though our list does include some range-restricted and threatened species as well (i.e., Hoary Puffleg [Haplophaedia lugens] and Cloud-forest Pygmy-Owl [Glaucidium nubicola], respectively). This approach allows us to use these largely widespread and banding-sampled species as best-case indicators for predicting potential changes that may also affect less frequently encountered and more cryptic species (i.e., Choco Vireo). Our list also includes species that feed on a variety of items, including fruits, invertebrates, vertebrates, and nectar, meaning these species play important roles in different ecological functions (Table 1).
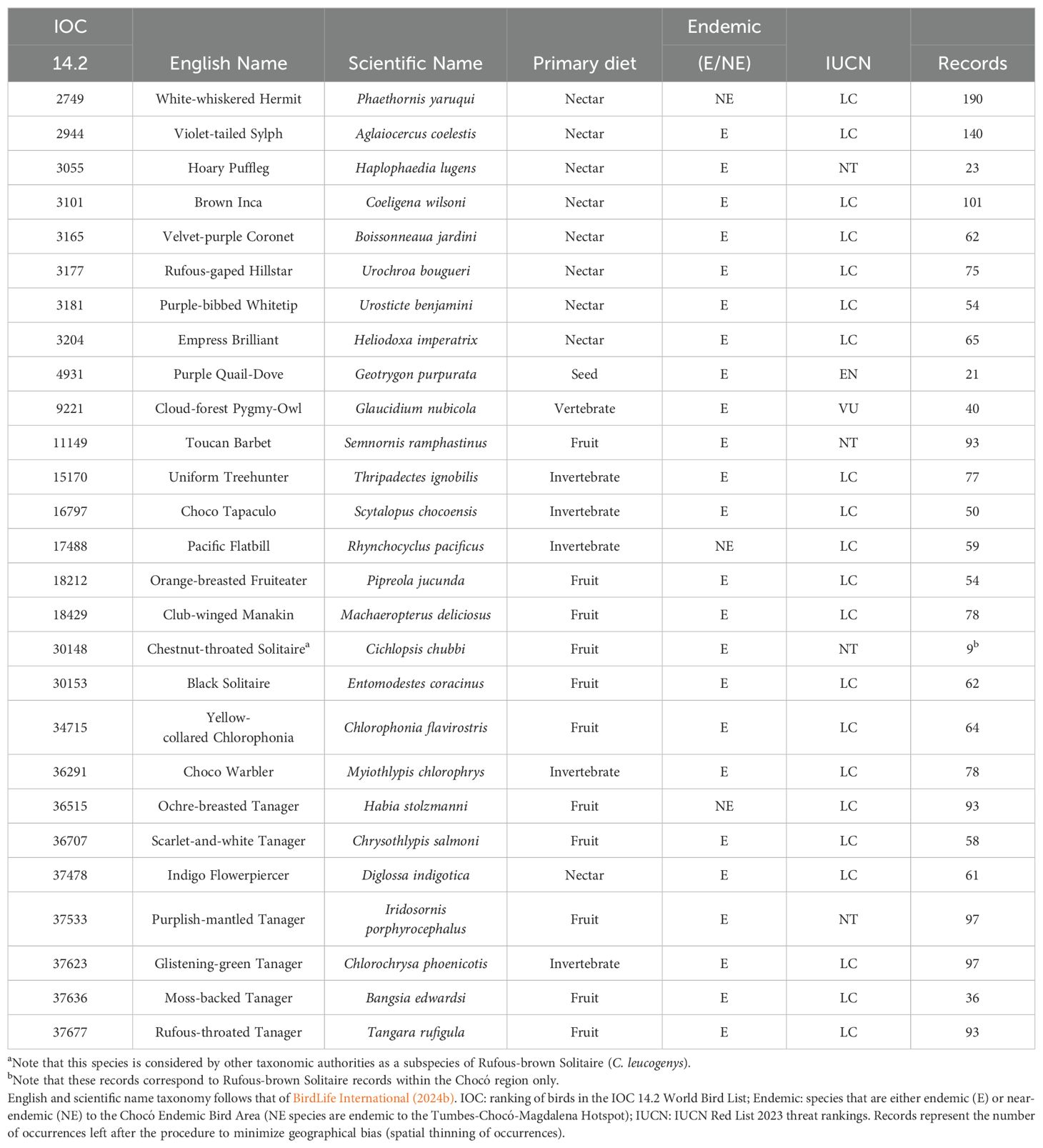
Table 1. Bird species included in the study.
All 27 species are either endemic or near-endemic to the Chocó Endemic Bird Area (EBA). Endemic species were identified using the BirdLife International (2024b) definition, which classifies a species as endemic if its range is entirely confined to the EBA. For species not currently listed as endemic by BirdLife International, we performed an additional review by cross-referencing the species’ projected ranges with the boundaries of the Chocó EBA to determine if they could be considered endemic based on geographical restriction. Near-endemic species, on the other hand, are those that are not exclusively confined to the Chocó EBA but have a significant portion of their range within this region. Specifically, the three near-endemic species included in this study are restricted to the greater Tumbes-Chocó-Magdalena biodiversity hotspot and have distributions that largely overlap with the Chocó EBA.
2.3 Avian data preparation
We downloaded the complete eBird world dataset (www.ebird.org) until the end of 2022 (ebd_prv_relDec-2022) and filtered the records for 27 Chocó bird species using the R package auk (Strimas-Mackey et al., 2023). We considered only complete checklists with all species reported and approved. Then, we eliminated inconsistencies such as duplicates, records at sea, and those matching centroids of municipalities using the R package CoordinateCleaner (Zizka et al., 2019). Finally, we filtered the records of each species to obtain the maximum number of occurrences that were at least 5 km apart, to avoid geographical bias, using the R package spThin (Aiello-Lammens et al., 2015). The final number of records for each species in the study are listed in Table 1.
While the eBird taxonomy follows that of the Clements checklist, we followed the avian taxonomic classifications of BirdLife International and the global bird checklist of the BirdLife Taxonomic Working Group (BirdLife International, 2024a) since it is used by the IUCN Red List for designating global threat assessments of species (IUCN, 2024). The only main discrepancy between these two taxonomies is for Chestnut-throated Solitaire (Cichlopsis chubbi), which is currently recognized, along with two other highly disjunct populations in South America that are treated by BirdLife as separate species, by Clements/eBird as a subspecies of Rufous-brown Solitaire (C. leucogenys). This taxonomic discrepancy was accounted for during the eBird data filtering process, since the chubbi population is the only one among the Rufous-brown complex that occurs in the Chocó region.
2.4 Climate data and ecological niche modeling
We obtained 19 bioclimatic variables (Table 2) for the baseline and future scenarios (2050 and 2070) with a spatial resolution of 2.5 arcminutes from the WorldClim platform (Fick and Hijmans, 2017). These variables are derived from precipitation and temperature measurements and largely used in ecological studies (Prieto-Torres et al., 2020; Oliveira-Silva et al., 2022). We considered three Global Climate Models (GCMs): IPSL-CM6A-LR, MIROC6, and MPI-ESM1-2-HR, and two Shared Socio-economic Pathways (SSPs): 245 and 585. The selection of these GCMs was based on their better performance with respect to frequency and persistence errors in the South America region (Cannon, 2020). The SSPs represent optimistic and pessimistic greenhouse gas emission scenarios, with the optimistic scenario corresponding to a global temperature increase of up to 3°C on average, and the pessimistic scenario corresponding to an increase of up to 5°C on average compared to the preindustrial period (O’Neill et al., 2014). For each species, we used the R package ENMwizard (Heming et al., 2019) to perform a Pearson correlation analysis considering all bioclimatic variables and selected those below the cutoff value of 0.7 to minimize multicollinearity (Dormann et al., 2013).
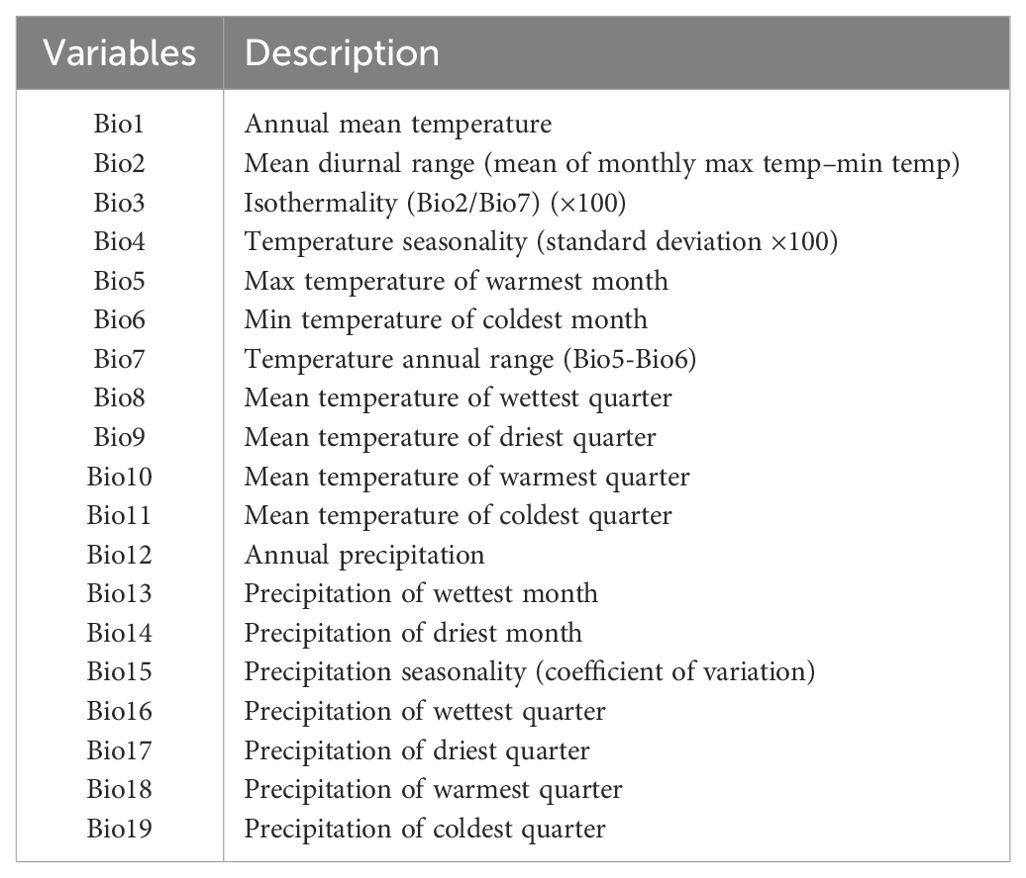
Table 2. Bioclimatic variables derived from temperature and precipitation data obtained from WorldClim – Global climate and weather data (www.worldclim.org).
We modeled the ecological niche of Chocó bird species using the MaxEnt algorithm (Phillips et al., 2006). This algorithm is widely used in the literature (Velásquez-Tibatá et al., 2013; Corrêa et al., 2019; Zhu et al., 2021), and its superior performance over other algorithms has already been demonstrated (Mota et al., 2022). We optimized two important parameters of MaxEnt: regularization multiplier and feature classes, thus generating 70 models per species. We employed the “block” and “jackknife” methods for cross-validation (Shcheglovitova and Anderson, 2013; Roberts et al., 2017). Subsequently, we ranked the best models based on the lowest omission rate (OR) and the highest area under the curve (AUC) (Boria et al., 2017). Next, we projected the models onto future climate scenarios (GCMs and SSPs) and generated consensus models by averaging the best models for each species and scenario (see details in Mota et al., 2022). We used the Maximum Training Sensitivity plus Specificity threshold to convert the final models into binary maps of 2.5 spatial arcminutes resolution representing “suitable” or “unsuitable” climate areas. This threshold is indicated for presence data only (Liu et al., 2013) and is more restrictive than other thresholds, which aligns with the projections for our species. Subsequently, we created a buffer around occurrence records for each species with a radius of 100 km and cropped the binary maps to reduce overprediction (Mendes et al., 2020). The buffer size was selected based on the distance between the western Andean slope and the coast, which we assumed to be accessible areas for the studied species. We used these cropped binary maps to calculate species richness by climate scenario using the R package divraster (Mota et al., 2023). The chosen variables by species and their percentages of contribution are listed in Supplementary Table S1.
2.5 Total, forested, and protected areas
We used the binary maps to calculate the total climate suitable area for each species and climate scenario. We acquired Land Cover Type 1 for the year 2022 from the MCD12Q1 product Version 6.1, available in the U.S. Geological Survey Earth Explorer platform (www.earthexplorer.usgs.gov). We extracted the forest cover class and converted it to a spatial resolution of 2.5 arcminutes. We overlaid the binary maps with the forested areas to calculate the climate suitable area within the current forested area.
Additionally, we acquired the maps of protected areas from the World Database on Protected Areas platform (www.protectedplanet.net) and converted them from a vector to a raster file with a spatial resolution of 2.5 arcminutes. We overlaid the binary maps with the protected areas to calculate the climate suitable area within the current protected areas.
We used the R package terra (Hijmans et al., 2023) to transform the data and to measure the area in square kilometers. All calculations and graphing were conducted in R (version 4.4.1; R Core Team, 2024) and QGIS (version 3.34; QGIS, 2024).
3 Results
Among the 27 Choco bird species of southwestern Colombia in our study, species fed mostly on fruits (n = 11), followed by nectar (n = 9), invertebrates (n = 5), seeds (n = 1), and vertebrates (n = 1). Their ecological niche models showed a good fit, with the Area Under the Curve (AUC) higher than 0.7 for all but one species (Pacific Flatbill, Rhynchocyclus pacificus), which showed an intermediate fit with an AUC of 0.68. The average AUC value was 0.78, while the Omission Rate (OR) value was 0.17, ranging from 0.06 to 0.36 (Supplementary Table S2).
Our projections indicate that the highest concentration of species in the baseline scenario is located in the southern Chocó region and on the west and east sides of the Andes Mountains in southwestern Colombia and northwestern Ecuador (Figure 1A). Nearly all species are forecast to lose suitable climate areas in at least one future scenario in these same areas, leading to an overall reduction in species richness (Figures 1B–E). A small increase in the number of species is projected in some currently less diverse regions, including highland areas near the Colombia-Ecuador border and the east side of the Andes Mountains in southern Ecuador (Figures 1B–E). These delta richness patterns are consistent across all future scenarios but become more evident in the 2070 pessimistic scenario, with up to 22 species losses and 10 species gains (Figures 1B–E).
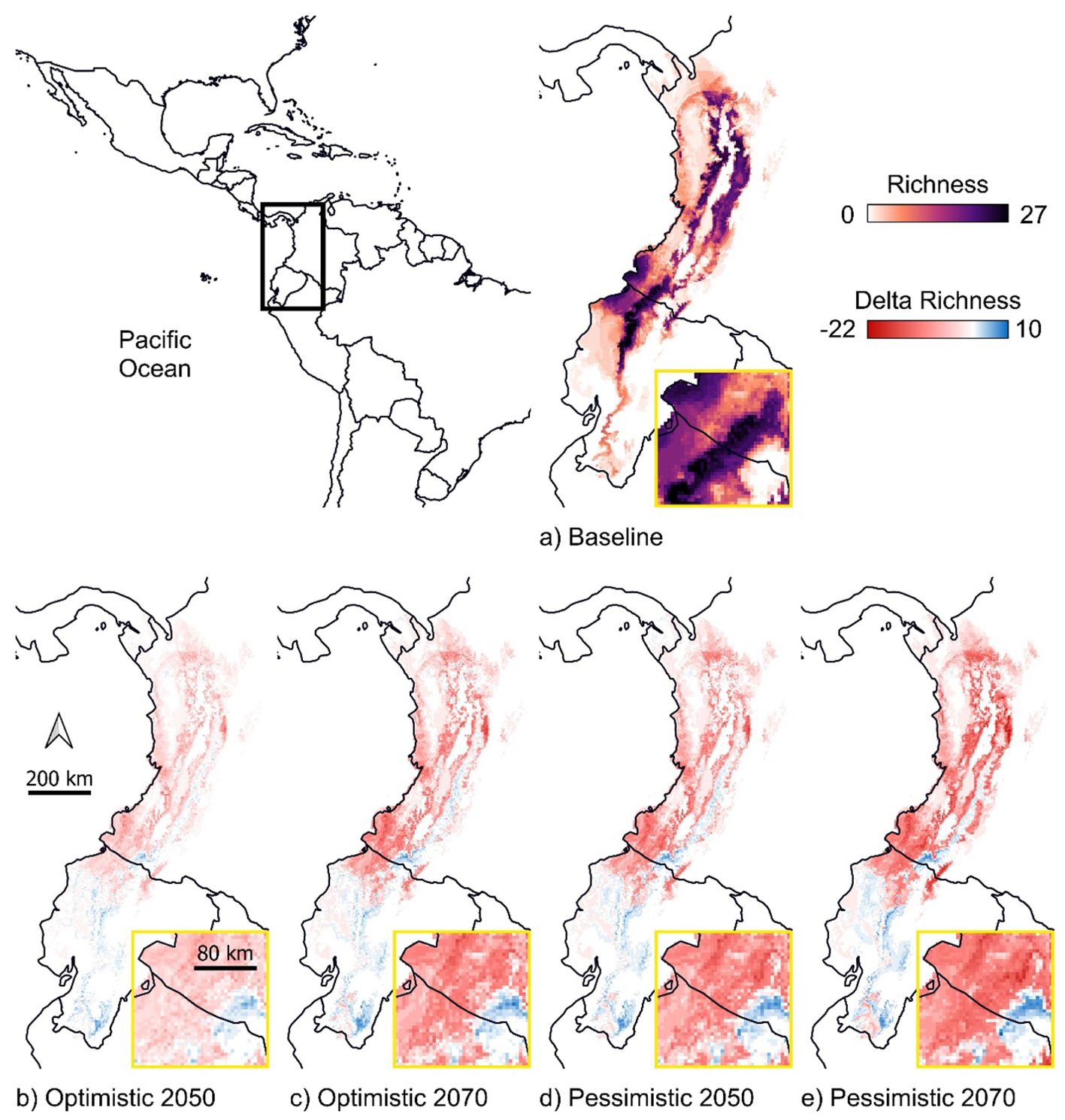
Figure 1. Species richness in the baseline scenario (A) and the difference between the richness in the baseline and future scenarios [delta richness; (B–E)] for 27 endemic or near-endemic bird species across the Chocó in Colombia and Ecuador, with a subset region depicted that focuses on southwestern Colombia. In the baseline richness map (A), darker colors represent areas with suitable climate conditions for a higher number of species. In the delta richness maps (B–E), red indicates areas where species are projected to contract their distributions, while blue indicates areas where species are projected to expand. The future scenarios represent optimistic (SSP245) and pessimistic (SSP585) greenhouse gas emission pathways for 2050 and 2070.
The average of the total climate suitable areas is 2.7–2.8 times larger than the suitable area within forested areas, 3.6–3.9 times larger than the suitable area within protected areas, and 7.3–7.7 times larger than the suitable area within both forested and protected areas for all climate scenarios evaluated. Additionally, the average suitable area decreases from the baseline to future scenarios for all classes. This reduction is higher from the baseline to the 2070 pessimistic scenario, whereas the reduction is lower but similar from the baseline to the optimistic scenarios and to the 2050 pessimistic scenario (Figure 2) (Table 3).
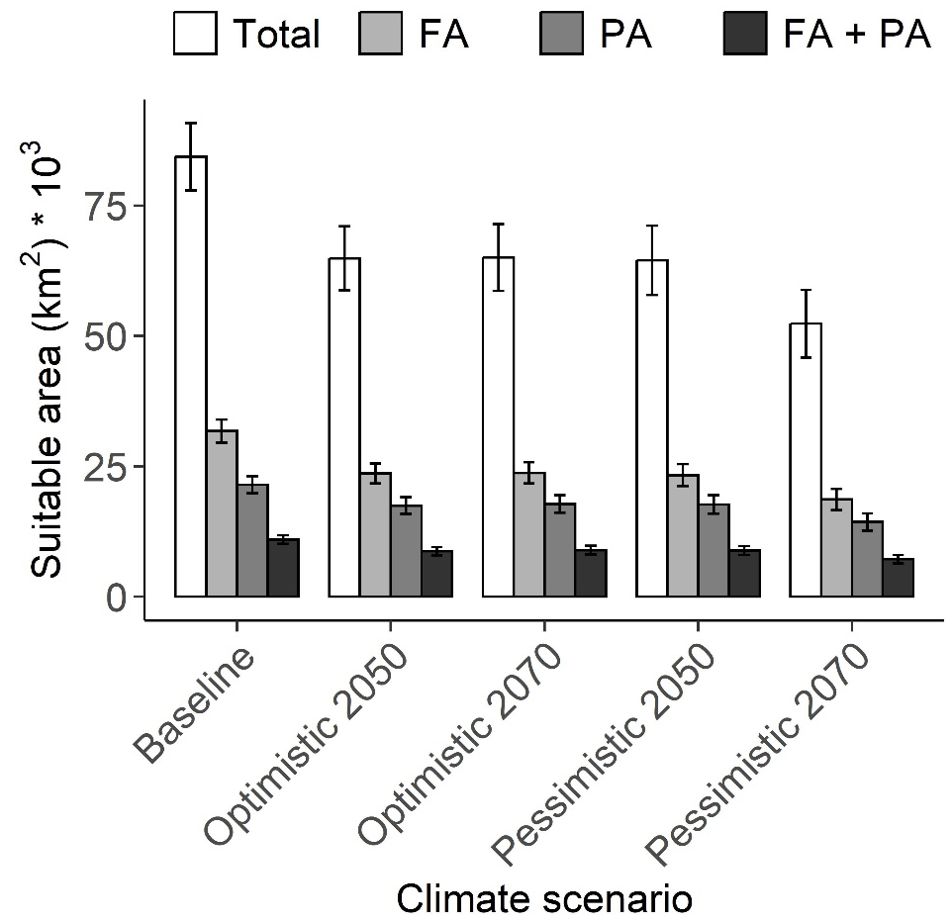
Figure 2. The average climate suitable area in squared kilometers for 27 endemic or near-endemic bird species across the Chocó in Colombia and Ecuador by climate scenario. Total: whole suitable area, FA: suitable area within forested areas, PA: suitable area within protected areas, and FA + PA: suitable area within forested and protected areas simultaneously. Vertical bars indicate the standard error of the mean. The future scenarios represent optimistic (SSP245) and pessimistic (SSP585) greenhouse gas emission pathways for 2050 and 2070.
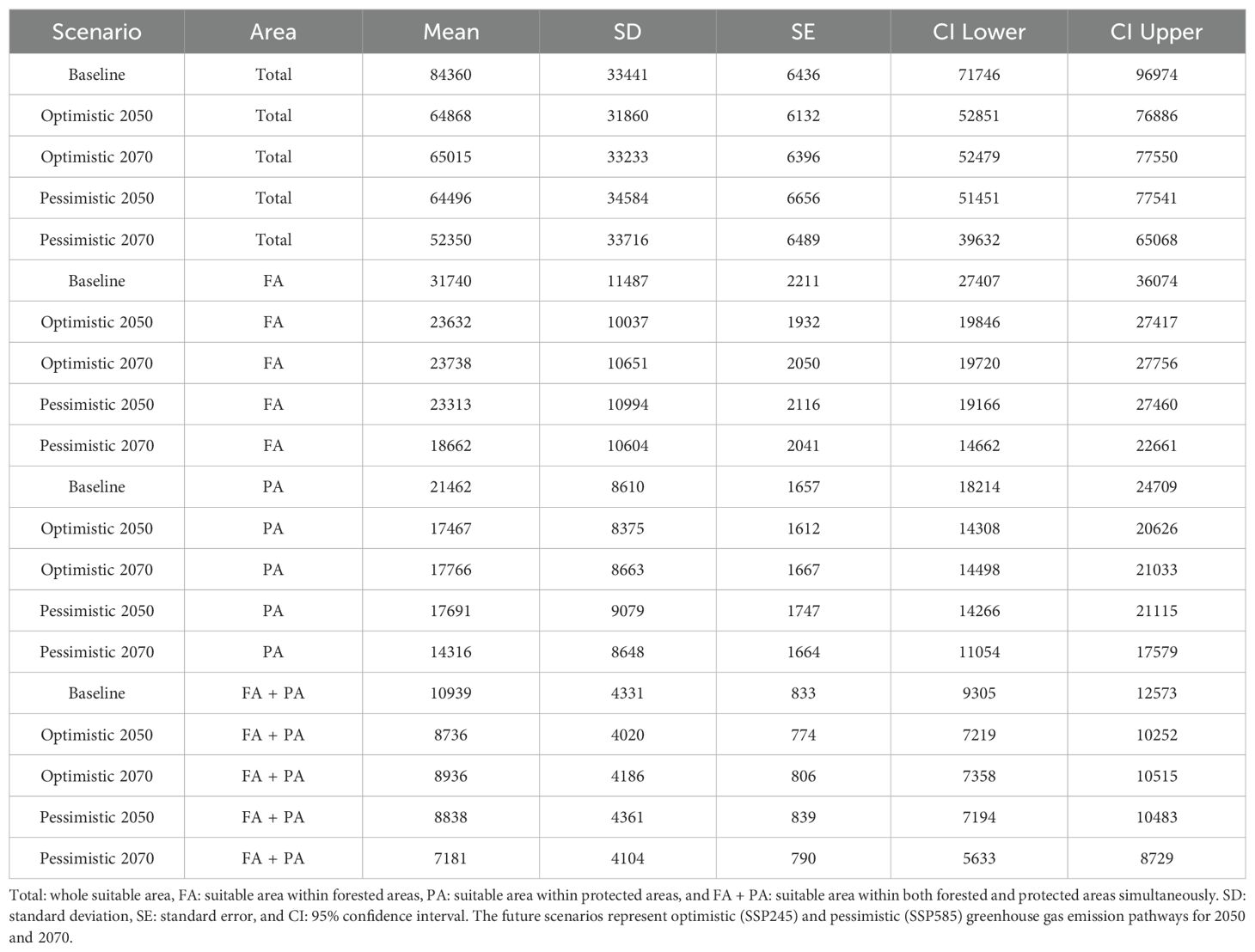
Table 3. The average climate suitable area in square kilometers for 27 endemic or near-endemic bird species across the Chocó in Colombia and Ecuador is presented by climate scenario.
Choco Warbler (Myiothlypis chlorophrys) is projected to lose the largest total climate-suitable area in the optimistic scenarios (2050: 53%; 2070: 55%) and the pessimistic 2050 scenario (60%). In the pessimistic 2070 scenario, however, greater losses are expected for Scarlet-and-white Tanager (Chrysothlypis salmoni) (84%). Both species are also projected to lose a significant portion of their suitable areas within forests. The Yellow-collared Chlorophonia (Chlorophonia flavirostris) is expected to lose the highest amount of suitable area within protected areas in the optimistic scenarios (2050: 41%; 2070: 42%), whereas greater losses in the pessimistic scenarios are anticipated for Scarlet-and-white Tanager (2050: 54%; 2070: 84%). When considering forested and protected areas together, the largest contractions in suitable area are projected for the Moss-backed Tanager (Bangsia edwardsi) in the optimistic scenarios (2050: 55%; 2070: 57%) and the pessimistic 2050 scenario (55%). In contrast, the Purple Quail-Dove (Geotrygon purpurata) is projected to be the least affected by climate change, proportionally, with a small gain in suitable area ranging from 4% (341 km²) within forested areas in the pessimistic 2050 scenario to 25% (534 km²) within forested and protected areas combined in the pessimistic 2070 scenario. The Ochre-breasted Tanager (Habia stolzmanni) is also expected to be less affected in terms of total suitable area under the pessimistic scenarios, with a modest gain of 3-5% (3218-6985 km²).
4 Discussion
Our results indicate that climate change is projected to induce significant changes in the species richness of birds that are endemic or near-endemic to the Chocó, especially in southwestern Colombia, with three-quarters of these bird species [in our study] at risk of extinction. Almost all species are expected to experience a reduction in their global distributions in at least one future climate scenario, resulting in a decline in the average suitable area. In addition, the suitable area within forested and protected areas is much smaller compared to the total suitable area, highlighting the urgent need to expand the protected area network in the region.
The change in total species distribution across all future climate scenarios ranged from -84% to +6%, with an average loss of 28%, resulting in a reduction of 81% in species richness (Figure 1, Supplementary Table S3). The potential loss of species was expected, given the already restricted distributions of Chocó birds, and our findings align with other studies that reported drastic losses for endemic birds (Hoffmann et al., 2020), as well as a positive relationship between range size and suitable area loss (Mota et al., 2022). Furthermore, the regions projected to experience high species losses and high species gains are predominantly at intermediate and high elevations, suggesting potential upslope shifts of birds in the future. This pattern mirrors trends observed in other parts of the world (Freeman and Class Freeman, 2014; Freeman et al., 2018, 2021; Neate-Clegg et al., 2020, 2021; Neate-Clegg and Tingley, 2023). These results emphasize the value of modelling future distributions of montane species to forecast potential elevational range shifts and climate-driven extirpations (Freeman et al., 2018; Nowak et al., 2019; Hoffmann et al., 2020). Additionally, the protected area Cotacachi Cayapas (WDPA, 2024), which represents the highest species richness in the baseline scenario, is located near a region projected to remain stable in the future (Figure 1). Expanding this protected area southwards could be an effective conservation strategy to address climate change impacts.
Supporting our prediction, the proportion of suitable area within forested and protected areas is considerably smaller compared to the total suitable area, regardless of the climate scenario, and it is also expected to decrease in the future (Figure 2; Supplementary Table S3). This suggests that, despite the availability of sites presenting favorable climate conditions, without vegetation, forest birds will not persist. This is supported by the higher vulnerability of forest species to climate change, as predicted in other studies (Miranda et al., 2019; Sales et al., 2020). Increasing forest cover in southwestern Colombia may help mitigate these impacts, not only because of the benefits of microclimatic conditions provided by natural vegetation but also to promote connections between potential refuge areas (Flórez-Paí, 2016; Frey et al., 2016; Guevara et al., 2016). Furthermore, the proportion of suitable areas within protected areas is even lower, representing only one-fourth of the total on average (Supplementary Table S3). This underrepresentation of species distribution within protected areas has been reported previously, highlighting the need to expand the coverage of these areas (Vale et al., 2018; Ivanova and Cook, 2020; Critchlow et al., 2022). Moreover, as a consequence of suitable area loss induced by climate change, Chocó birds are expected to show more fragmented distributions (Figure 3), a pattern already observed for other Colombian species (Velásquez-Tibatá et al., 2013). The expansion of the protected area network may prevent the isolation of bird populations and their risk of extinction by serving as corridors between suitable areas (Littlefield et al., 2017) that increase the connectivity of critical habitat at local and landscape scales (Şekercioğlu, 2009; Palacio et al., 2020).
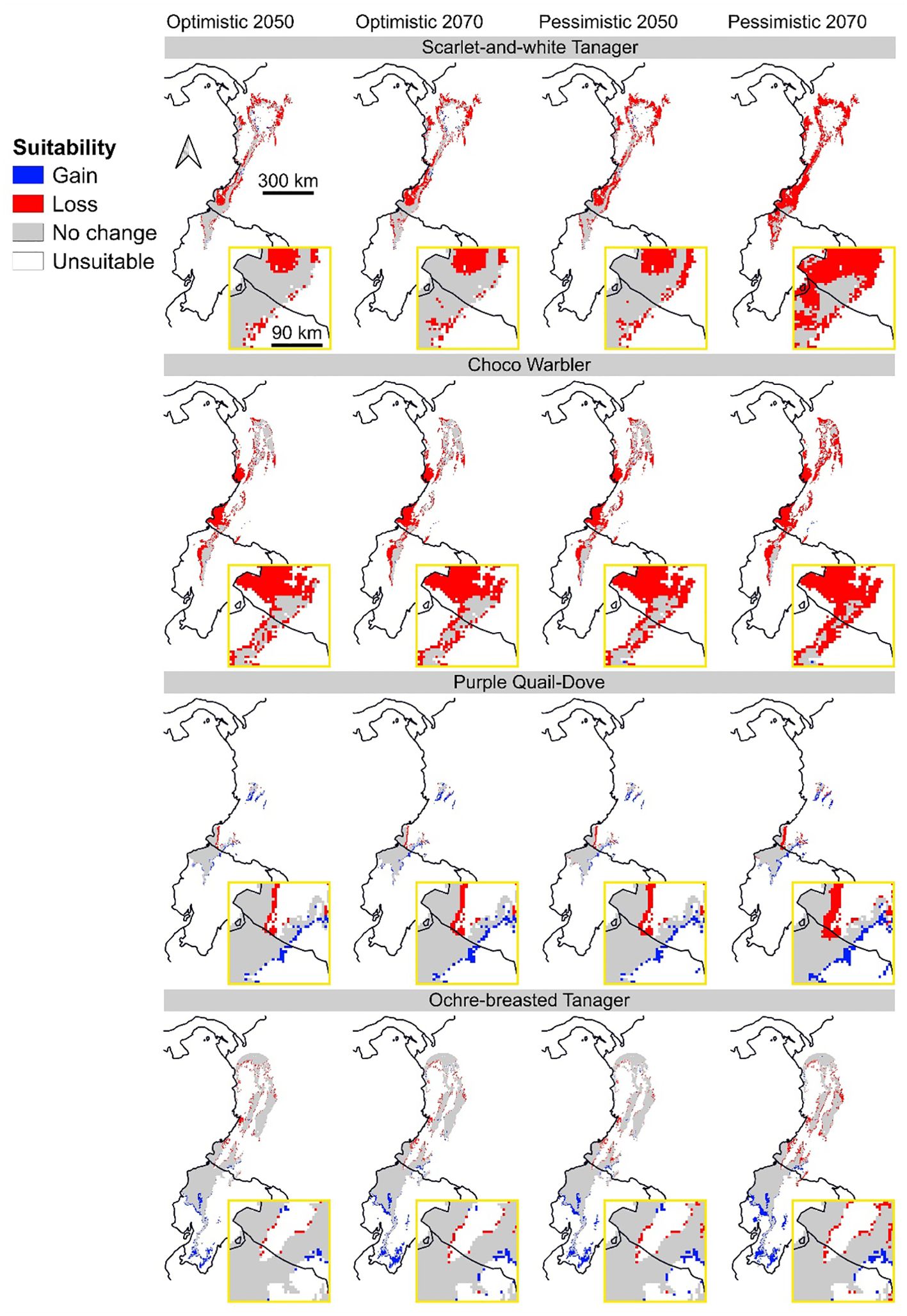
Figure 3. Changes in climate suitable areas for the Scarlet-and-white Tanager (Chrysothlypis salmoni), Choco Warbler (Myiothlypis chlorophrys), Purple Quail-Dove (Geotrygon purpurata), and Ochre-breasted Tanager (Habia stolzmanni) across the Chocó region in Colombia and Ecuador. Colors indicate changes in climate suitable area between baseline and future scenarios, with red indicating contraction, blue expansion, gray stability, and white no suitable climate conditions. The future scenarios represent optimistic (SSP245) and pessimistic (SSP585) greenhouse gas emission pathways for 2050 and 2070. The suitability change for all species is available in Appendix A.
As we expected, changes in birds’ suitable areas vary among species (Supplementary Table S3). The species expected to suffer larger contractions on average in their distributions feed primarily on invertebrates (Choco Warbler; -61%) and fruits (Yellow-collared Chlorophonia; -59%). Similarly, forest frugivore birds from the Atlantic and Amazon forest are also expected to lose a significant portion of their current distributions (Miranda et al., 2019; Mota et al., 2022). Moreover, species such as White-flanked Antwren (Myrmotherula axillaris) and Choco Poorwill (Nyctiphrynus rosenbergi; this species has been observed but not yet captured at the Río Ñambí and Bangsias reserves by the authors), are also expected to lose climate suitable area greater than 80% in the future corroborating the high risk of forest insectivore birds as well (Velásquez-Tibatá et al., 2013; Miranda et al., 2019). The loss of these species could affect the provision of essential ecological functions, particularly seed dispersal and insect control, by reducing the natural regeneration of forests and increasing leaf damage in plants (Şekercioğlu, 2006; Mäntylä et al., 2011; Wenny et al., 2016; Martínez and García, 2017). Furthermore, Purple Quail-Dove seems to be favored by future climatic conditions in the Chocó region, though this gain is still small in absolute terms, representing a 25% increase within forested and protected areas, totaling only 534 km² (Supplementary Table S3). In this case, protected areas such as Serranía de Los Paraguas and Cotacachi Cayapas are particularly important for the species’ conservation, especially since the Purple Quail-Dove is already considered endangered due to habitat loss (del Hoyo et al., 2020). Indeed, birds in the Colombian Andes have declined across a century due primarily to reductions in forest cover resulting from land-use change, deforestation, and habitat fragmentation (Palacio et al., 2020), indicating that suitable climatic conditions alone cannot ensure the species’ persistence.
5 Conclusion
Our study represents the first assessment of future climate change impacts on endemic and near-endemic Chocó birds in southwestern Colombia, offering valuable insights for conservation planning. While a higher number of records could increase the predictive power of our models, and taking into account other factors such as dispersal scenarios and species-specific traits could provide deeper insight into species’ responses to climate change, our findings nevertheless essentially contribute to the limited information on climate change impacts in this biodiverse region. Importantly, we provide projected distribution maps and potential elevational range shifts, for 27 Neotropical bird species under different climate scenarios (Figure 3, Appendix B) – these projections will be useful for conservation efforts and better understanding trends in resident bird populations. The potential loss of these species could jeopardize essential ecological services such as seed dispersal, pollination, and insect control, leading to cascading effects on other species affecting the entire community and reducing its resilience to disturbances. Moreover, eBird, one of the most well-known and widely used citizen and community science initiatives around the world (Kittelberger et al., 2023), serves as an important source of bird occurrence data globally and is an important tool for forecasting species distributions in the future (Delfino, 2023; Liu et al., 2023). To mitigate the potential losses of suitable habitats and facilitate species’ range shifts, reforestation and habitat restoration should be prioritized alongside the expansion of protected areas throughout the biome. These strategies are vital for enhancing habitat connectivity and creating corridors that allow species to move between fragmented areas, thereby maintaining biodiversity and the ecosystem services they provide. Such combined efforts will be crucial in sustaining bird populations and their ecological roles in the Chocó region under future climate scenarios.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
FM: Writing – original draft, Writing – review & editing, Conceptualization, Data curation, Formal analysis, Methodology, Visualization. KK: Writing – original draft, Writing – review & editing, Conceptualization, Visualization. CF-P: Writing – review & editing. ÇŞ: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was supported by funding from the Exploration Fund of The Explorers Club and a fellowship granted by the Brazilian Federal Foundation for the Support and Evaluation of Graduate Education (CAPES - 88881.846347/2023-01) to FM.
Acknowledgments
We thank H. Batubay Özkan and Barbara Watkins for their continued support of the Biodiversity and Conservation Ecology (BCE) Lab with the School of Biological Sciences at the University of Utah. We also are appreciative of the University of Utah’s Graduate Research Fellowship for providing support to KK for this research. Special thanks to the local communities for their conservation efforts for endemic birds since 1991 at OECM Bangsias BirdLodge, OECM San Antonio, and KBA Río Ñambí.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2024.1412440/full#supplementary-material
References
Aiello-Lammens M. E., Boria R. A., Radosavljevic A., Vilela B., Anderson R. P. (2015). spThin: an R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 38, 541–545. doi: 10.1111/ecog.01132
Amaya-Vallejo V., Bota-Sierra C., Novelo-Gutiérrez R., Sanchez-Herrera M. (2021). Two new species of Archaeopodagrion (Odonata, Philogeniidae) from the western foothills of the Tropical Andes, with biological observations and distributional records. Zookeys 1036, 21–38. doi: 10.3897/zookeys.1036.64230
Anaya J. A., Gutiérrez-Vélez V. H., Pacheco-Pascagaza A. M., Palomino-Ángel S., Han N., Balzter H. (2020). Drivers of forest loss in a megadiverse hotspot on the pacific coast of Colombia. Remote Sens (Basel) 12, 1235. doi: 10.3390/rs12081235
BirdLife International (2024a). BirdLife Data Zone. Available online at: http://datazone.birdlife.org (Accessed January 26, 2024).
BirdLife International (2024b). Endemic Bird Areas factsheet: Chocó. Available online at: http://datazone.birdlife.org/eba/factsheet/42 (Accessed January 26, 2024).
Boria R. A., Olson L. E., Goodman S. M., Anderson R. P. (2017). A single-algorithm ensemble approach to estimating suitability and uncertainty: cross-time projections for four Malagasy tenrecs. Divers. Distrib 23, 196–208. doi: 10.1111/ddi.12510
Cannon A. J. (2020). Reductions in daily continental-scale atmospheric circulation biases between generations of global climate models: CMIP5 to CMIP6. Environ. Res. Lett. 15, 064006. doi: 10.1088/1748-9326/ab7e4f
Corrêa L. L. C., Oliveira S. V., Silva D. E., Petry M. V. (2019). Occurrence and predictive distribution of Crypturellus noctivagus (Aves, tinamidae) in Brazil. Oecologia Australis 23, 548–561. doi: 10.4257/oeco.2019.2303.13
Critchlow R., Cunningham C. A., Crick H. Q. P., Macgregor N. A., Morecroft M. D., Pearce-Higgins J. W., et al. (2022). Multi-taxa spatial conservation planning reveals similar priorities between taxa and improved protected area representation with climate change. Biodivers Conserv. 31, 683–702. doi: 10.1007/s10531-022-02357-1
da Silva L. B., Oliveira G. L., Frederico R. G., Loyola R., Zacarias D., Ribeiro B. R., et al. (2022). How future climate change and deforestation can drastically affect the species of monkeys endemic to the eastern Amazon, and priorities for conservation. Biodivers Conserv. 31, 971–988. doi: 10.1007/s10531-022-02373-1
Delfino H. C. (2023). A fragile future for pink birds: habitat suitability models predict a high impact of climate change on the future distribution of flamingos. Emu - Austral Ornithology 123, 310–324. doi: 10.1080/01584197.2023.2257757
del Hoyo J., Collar N., Kirwan G. M., Garcia E., Sharpe C. (2020). “Purple quail-dove (Geotrygon purpurata),” in Birds of the world. Eds. del Hoyo J., Elliott A., Sargatal J., Christie D., de Juana E. (Ithaca, NY, USA: Cornell Lab of Ornithology). doi: 10.2173/bow.sapqud1.01
de Moraes K. F., Santos M. P. D., Gonçalves G. S. R., de Oliveira G. L., Gomes L. B., Lima M. G. M. (2020). Climate change and bird extinctions in the Amazon. PLoS One 15, e0236103. doi: 10.1371/journal.pone.0236103
Devenish C., Díaz Fernandéz D. F., Clay R. P., Davidson I., Yépez Zabala I. (2009). Important bird areas of the Americas-priority sites for biodiversity conservation, vol. 31. (Quito, Ecuador: BirdLife International).
Dormann C. F., Elith J., Bacher S., Buchmann C., Carl G., Carré G., et al. (2013). Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46. doi: 10.1111/j.1600-0587.2012.07348.x
Fick S. E., Hijmans R. J. (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatology 37, 4302–4315. doi: 10.1002/joc.5086
Flórez-Paí C. (2016). Lineamientos para la priorización de áreas de importancia para la conservación en el suroccidente de Colombia con base en grupos bióticos amenazados, aves y plantas (Bogotá: Pontificia Universidad Javeriana).
Freeman B. G., Class Freeman A. M. (2014). Rapid upslope shifts in New Guinean birds illustrate strong distributional responses of tropical montane species to global warming. Proc. Natl. Acad. Sci. 111, 4490–4494. doi: 10.1073/pnas.1318190111
Freeman B. G., Scholer M. N., Ruiz-Gutierrez V., Fitzpatrick J. W. (2018). Climate change causes upslope shifts and mountaintop extirpations in a tropical bird community. Proc. Natl. Acad. Sci. 115, 11982–11987. doi: 10.1073/pnas.1804224115
Freeman B. G., Song Y., Feeley K. J., Zhu K. (2021). Montane species track rising temperatures better in the tropics than in the temperate zone. Ecol. Lett. 24, 1697–1708. doi: 10.1111/ele.13762
Frey S. J. K., Hadley A. S., Johnson S. L., Schulze M., Jones J. A., Betts M. G. (2016). Spatial models reveal the microclimatic buffering capacity of old-growth forests. Sci. Adv. 2. doi: 10.1126/sciadv.1501392
Fricke E. C., Ordonez A., Rogers H. S., Svenning J.-C. (2022). The effects of defaunation on plants’ capacity to track climate change. Sci. (1979) 375, 210–214. doi: 10.1126/science.abk3510
Guevara O., Abud M., Trujillo A. F., Suárez C. F., Cuadros L., López C., et al. (2016). Plan Territorial de Adaptación al Cambio Climático del departamento de Nariño (Cali, Colombia: Cali).
Guevara Andino J. E., Fernandez-Fernandez D. (2020). A new rare and endemic species of Sloanea (Elaeocarpaceae) from the Chocó region of Ecuador. PhytoKeys 160, 131–139. doi: 10.3897/phytokeys.160.54993
Harris J. B. C., Şekercioğlu Ç.H., Sodhi N. S., Fordham D. A., Paton D. C., Brook B. W. (2011). The tropical frontier in avian climate impact research. Ibis 153, 877–882. doi: 10.1111/j.1474-919X.2011.01166.x
Heming N. M., Dambros C., Gutiérrez E. E. (2019). ENMwizard: Advanced techniques for Ecological Niche Modeling made easy. Available online at: https://github.com/HemingNM/ENMwizard (Accessed October 15, 2024).
Hijmans R. J., Bivand R., Pebesma E., Sumner M. D. (2023). terra: Spatial Data Analysis. Available online at: https://CRAN.R-project.org/package=terra (Accessed October 18, 2024).
Hoffmann D., de Vasconcelos M. F., Wilson Fernandes G. (2020). The fate of endemic birds of eastern Brazilian mountaintops in the face of climate change. Perspect. Ecol. Conserv. 18, 257–266. doi: 10.1016/j.pecon.2020.10.005
IUCN (2024). The IUCN Red List of Threatened Species. Version 2023-1. Available online at: https://www.iucnredlist.org (Accessed January 26, 2024).
Ivanova I. M., Cook C. N. (2020). The role of privately protected areas in achieving biodiversity representation within a national protected area network. Conserv. Sci. Pract. 2, e307. doi: 10.1111/csp2.307
Kittelberger K. D., Tanner C. J., Orton N. D., Şekercioğlu Ç.H. (2023). The value of community science data to analyze long-term avian trends in understudied regions: The state of birds in Türkiye. Avian Res. 14, 100140. doi: 10.1016/j.avrs.2023.100140
Littlefield C. E., Krosby M., Michalak J. L., Lawler J. J. (2019). Connectivity for species on the move: supporting climate-driven range shifts. Front. Ecol. Environ. 17, 270–278. doi: 10.1002/fee.2043
Littlefield C. E., McRae B. H., Michalak J. L., Lawler J. J., Carroll C. (2017). Connecting today’s climates to future climate analogs to facilitate movement of species under climate change. Conserv. Biol. 31, 1397–1408. doi: 10.1111/cobi.12938
Liu Z., Sandoval L., Sherman L. B., Wilson A. M. (2023). Vulnerability of elevation-restricted endemic birds of the Cordillera de Talamanca (Costa Rica and Panama) to climate change. Neotrop Biodivers 9, 115–127. doi: 10.1080/23766808.2023.2261196
Liu C., White M., Newell G. (2013). Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr 40, 778–789. doi: 10.1111/jbi.12058
Mäntylä E., Klemola T., Laaksonen T. (2011). Birds help plants: a meta-analysis of top-down trophic cascades caused by avian predators. Oecologia 165, 143–151. doi: 10.1007/s00442-010-1774-2
Martínez D., García D. (2017). Role of avian seed dispersers in tree recruitment in woodland pastures. Ecosystems 20, 616–629. doi: 10.1007/s10021-016-0043-6
Mendes P., Velazco S. J. E., de Andrade A. F. A., De Marco P. (2020). Dealing with overprediction in species distribution models: How adding distance constraints can improve model accuracy. Ecol. Modell 431, 109180. doi: 10.1016/j.ecolmodel.2020.109180
Miranda L. S., Imperatriz-Fonseca V. L., Giannini T. C. (2019). Climate change impact on ecosystem functions provided by birds in southeastern Amazonia. PLoS One 14, e0215229. doi: 10.1371/journal.pone.0215229
Mota F. M. M., Alves-Ferreira G., Talora D. C., Heming N. M. (2023). divraster: an R package to calculate taxonomic, functional and phylogenetic diversity from rasters. Ecography 2023, e06905. doi: 10.1111/ecog.06905
Mota F. M. M., Heming N. M., Morante-Filho J. C., Talora D. C. (2022). Climate change is expected to restructure forest frugivorous bird communities in a biodiversity hot-point within the Atlantic Forest. Divers. Distrib 28, 2886–2897. doi: 10.1111/ddi.13602
Neate-Clegg M. H. C., Jones S. E. I., Tobias J. A., Newmark W. D., Şekercioğlu Ç.H. (2021). Ecological correlates of elevational range shifts in tropical birds. Front. Ecol. Evol. 9. doi: 10.3389/fevo.2021.621749
Neate-Clegg M. H. C., O’Brien T. G., Mulindahabi F., Şekercioğlu Ç.H. (2020). A disconnect between upslope shifts and climate change in an Afrotropical bird community. Conserv. Sci. Pract. 2, e291. doi: 10.1111/csp2.291
Neate-Clegg M. H. C., Tingley M. W. (2023). Building a mechanistic understanding of climate-driven elevational shifts in birds. PLoS Climate 2, e0000174. doi: 10.1371/journal.pclm.0000174
Nowak L., Kissling W. D., Bender I. M. A., Dehling D. M., Töpfer T., Böhning-Gaese K., et al. (2019). Projecting consequences of global warming for the functional diversity of fleshy-fruited plants and frugivorous birds along a tropical elevational gradient. Divers. Distrib 25, 1362–1374. doi: 10.1111/ddi.12946
O’Neill B. C., Kriegler E., Riahi K., Ebi K. L., Hallegatte S., Carter T. R., et al. (2014). A new scenario framework for climate change research: the concept of shared socioeconomic pathways. Clim Change 122, 387–400. doi: 10.1007/s10584-013-0905-2
Ogada D. L., Torchin M. E., Kinnaird M. F., Ezenwa V. O. (2012). Effects of vulture declines on facultative scavengers and potential implications for mammalian disease transmission. Conserv. Biol. 26, 453–460. doi: 10.1111/j.1523-1739.2012.01827.x
Oliveira-Silva A. E., Piratelli A. J., Zurell D., da Silva F. R. (2022). Vegetation cover restricts habitat suitability predictions of endemic Brazilian Atlantic Forest birds. Perspect. Ecol. Conserv. 20, 1–8. doi: 10.1016/j.pecon.2021.09.002
Palacio R. D., Kattan G. H., Pimm S. L. (2020). Bird extirpations and community dynamics in an Andean cloud forest over 100 years of land-use change. Conserv. Biol. 34, 677–687. doi: 10.1111/cobi.13423
Pérez-Escobar O. A., Lucas E., Jaramillo C., Monro A., Morris S. K., Bogarín D., et al. (2019). The origin and diversification of the hyperdiverse flora in the chocó Biogeographic region. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01328
Phillips S. J., Anderson R. P., Schapire R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecol. Modell 190, 231–259. doi: 10.1016/j.ecolmodel.2005.03.026
Prieto-Torres D. A., Lira-Noriega A., Navarro-Sigüenza A. G. (2020). Climate change promotes species loss and uneven modification of richness patterns in the avifauna associated to Neotropical seasonally dry forests. Perspect. Ecol. Conserv. 18, 19–30. doi: 10.1016/j.pecon.2020.01.002
QGIS (2024). QGIS Geographic Information System. QGIS Association. Available online at: https://qgis.org/en/site/forusers/index.html (Accessed October 22, 2024).
Raven P. H., Gereau R. E., Phillipson P. B., Chatelain C., Jenkins C. N., Ulloa Ulloa C. (2020). The distribution of biodiversity richness in the tropics. Sci. Adv. 6. doi: 10.1126/sciadv.abc6228
R Core Team (2024). R: The R Project for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.r-project.org/.
Roberts D. R., Bahn V., Ciuti S., Boyce M. S., Elith J., Guillera-Arroita G., et al. (2017). Cross-validation strategies for data with temporal, spatial, hierarchical, or phylogenetic structure. Ecography 40, 913–929. doi: 10.1111/ecog.02881
Román-Palacios C., Wiens J. J. (2020). Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. 117, 4211–4217. doi: 10.1073/pnas.1913007117
Şekercioğlu Ç.H. (2006). Increasing awareness of avian ecological function. Trends Ecol. Evol. 21, 464–471. doi: 10.1016/j.tree.2006.05.007
Şekercioğlu Ç.H. (2009). Tropical ecology: riparian corridors connect fragmented forest bird populations. Curr. Biol. 19, R210–R213. doi: 10.1016/j.cub.2009.01.006
Şekercioğlu Ç.H., Primack R. B., Wormworth J. (2012). The effects of climate change on tropical birds. Biol. Conserv. 148, 1–18. doi: 10.1016/j.biocon.2011.10.019
Şekercioğlu Ç.H., Schneider S. H., Fay J. P., Loarie S. R. (2008). Climate change, elevational range shifts, and bird extinctions. Conserv. Biol. 22, 140–150. doi: 10.1111/j.1523-1739.2007.00852.x
Şekercioğlu Ç.H., Wenny D. G., Whelan C. J. (2016). Why Birds Matter. (Chicago, IL, USA: University of Chicago Press). doi: 10.7208/chicago/9780226382777.001.0001
Sales L. P., Galetti M., Pires M. M. (2020). Climate and land-use change will lead to a faunal “savannization” on tropical rainforests. Glob Chang Biol. 26, 7036–7044. doi: 10.1111/gcb.15374
Sánchez-Nivicela M., Montenegro-Pazmiño E. (2022). Conservation Investment Strategy for Resident and Migratory Birds of the Chocó-Andean Region in Northwestern Ecuador (Quito: irdLife International; Aves y Conservación, Fundación Jocotoco). Available at: https://www.birdlife.org/papers-reports/conservation-investment-strategy-for-resident-and-migratory-birds-of-the-choco-andean-region-in-northwest-Ecuador/.
Shcheglovitova M., Anderson R. P. (2013). Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Modell 269, 9–17. doi: 10.1016/j.ecolmodel.2013.08.011
Stan K. D., Sanchez-Azofeifa A., Hamann H. F. (2024). Widespread degradation and limited protection of forests in global tropical dry ecosystems. Biol. Conserv. 289, 110425. doi: 10.1016/j.biocon.2023.110425
Strimas-Mackey M., Miller E., Hochachka W., Ornithology C. L. (2023). auk: eBird Data Extraction and Processing in R. Available online at: https://cran.r-project.org/package=auk (Accessed October 15, 2024).
Taubert F., Fischer R., Groeneveld J., Lehmann S., Müller M. S., Rödig E., et al. (2018). Global patterns of tropical forest fragmentation. Nature 554, 519–522. doi: 10.1038/nature25508
Triviño M., Kujala H., Araújo M. B., Cabeza M. (2018). Planning for the future: identifying conservation priority areas for Iberian birds under climate change. Landsc Ecol. 33, 659–673. doi: 10.1007/s10980-018-0626-z
Vale M. M., Souza T. V., Alves M. A. S., Crouzeilles R. (2018). Planning protected areas network that are relevant today and under future climate change is possible: the case of Atlantic Forest endemic birds. PeerJ 6, e4689. doi: 10.7717/peerj.4689
Velandia Perilla J. H., Guerrero-Chacon A. L., Ruiz-Espinoza A. (2021). New record of the brown sac-wing bat, balantiopteryx infusca (Chiroptera, emballonuridae) in Colombia. Mammalogy Notes 7, 194. doi: 10.47603/mano.v7n1.194
Velásquez-Tibatá J., Salaman P., Graham C. H. (2013). Effects of climate change on species distribution, community structure, and conservation of birds in protected areas in Colombia. Reg. Environ. Change 13, 235–248. doi: 10.1007/s10113-012-0329-y
WDPA (2024). The World Database on Protected Areas. Available online at: https://www.protectedplanet.net/ (Accessed December 18, 2023).
Wenny D. G., Şekercioğlu Ç.H., Whelan C. J. (2016). “Seed dispersal by fruit-eating,” in Why Birds Matter (Chicago: University of Chicago Press), 107–146.
Wessinger C. A. (2021). From pollen dispersal to plant diversification: genetic consequences of pollination mode. New Phytol. 229, 3125–3132. doi: 10.1111/nph.17073
Wormworth J., Şekercioğlu Ç.H. (2011). Winged Sentinels. 1st Edn (New York, NY, USA: Cambridge University Press). doi: 10.1017/CBO9781139150026
Zhu G., Papeş M., Giam X., Cho S.-H., Armsworth P. R. (2021). Are protected areas well-sited to support species in the future in a major climate refuge and corridor in the United States? Biol. Conserv. 255, 108982. doi: 10.1016/j.biocon.2021.108982
Keywords: global warming, population decline, forest cover, neotropics, tropical ornithology, KBA Río Ñambí, conservation biology
Citation: Mota FMM, Kittelberger KD, Flórez-Paí C and Şekercioğlu ÇH (2024) Climate-driven distributional shifts in Chocó endemic birds of southwest Colombia. Front. Conserv. Sci. 5:1412440. doi: 10.3389/fcosc.2024.1412440
Received: 04 April 2024; Accepted: 31 October 2024;
Published: 27 November 2024.
Edited by:
Ricardo Lourenço-de-Moraes, Federal University of Paraíba, BrazilReviewed by:
Felipe S. Campos, Ecological and Forestry Applications Research Center (CREAF), SpainStella De La Torre, Universidad San Francisco de Quito, Ecuador
Copyright © 2024 Mota, Kittelberger, Flórez-Paí and Şekercioğlu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Flávio M. M. Mota, ZmxhdmlvbW9jQGdtYWlsLmNvbQ==
†Present address: Flávio M. M. Mota, Evolutionary Ecology and Biodiversity Lab, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil