 Alena McGuire
Alena McGuire Chase A. LaDue
Chase A. LaDue
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Conserv. Sci., 25 March 2024
Sec. Animal Conservation
Volume 5 - 2024 | https://doi.org/10.3389/fcosc.2024.1362313
This article is part of the Research TopicUnderstanding Rest-Activity Rhythms: Uncovering Factors Shaping Behavioural Rhythms in Diverse Ecological Contexts.View all 5 articles
Introduction: Monitoring overnight behavior is important in assessing the overall wellbeing of ex-situ elephant populations, with recumbent rest and stereotypy as key indicators of welfare. However, there have been few studies that address the overnight behavior of singly housed male elephants with a history of stereotypy. We conducted an opportunistic case study of the overnight behavior (i.e., rest and stereotypy) of a singly housed geriatric male Asian elephant at the Oklahoma City Zoo to identify his overnight behavioral rhythms in response to physiological (musth) and environmental changes (habitat access, automatic timed feeders) and guide management strategies.
Methods: Infrared cameras were utilized to conduct continuous focal-animal sampling of the elephant’s behavior between 20:00 and 08:00 in indoor and outdoor habitats. Sampling occurred from January 2023 to October 2023, with a total of 179 overnight observations.
Results: The elephant exhibited recumbent rest for 175.66 ± 6.80 (mean ± SE) minutes per night and was engaged in stereotypy for 175.88 ± 9.68 minutes per night. While there was no significant relationship between stereotypy and the sexual state of musth, musth was associated with reduced durations of recumbent rest. Access to both indoor and outdoor habitats was significantly related to decreased stereotypy compared to indoor-only treatments. Recumbent rest occurred most frequently between 02:00 and 05:00, and stereotypic behavior was most common between 06:00 and 08:00. Contrary to our expectations, automatic feeders did not significantly reduce stereotypy.
Discussion: The results of this study provide insight into the behavioral patterns of a geriatric male Asian elephant, demonstrating the importance of implementing an animal-centered approach to enhance animal wellbeing. As the zoo-housed Asian elephant population grows and continues to age, this case study emphasizes the importance of developing comprehensive welfare strategies for the elephants in our care.
Asian elephants (Elephas maximus) are endangered, and ex-situ populations (such as those housed by zoos, wildlife parks, camps, and similar facilities) are important for the long-term sustainability of the species (Riddle and Stremme, 2011). At the same time, zoos and other facilities have an obligation to deliver a high standard of care to elephants in these populations, including providing opportunities to thrive that ensure proper management and welfare (Association of Zoos and Aquariums, 2023). There is a great amount of interest in the wellbeing of zoo-housed elephants, and various studies have identified environmental and social factors that influence the welfare of elephants in zoos. For example, we now understand that positive indicators of welfare are associated with complex environments, diverse feeding opportunities, and softer flooring substrates (Meehan et al., 2016). Nonetheless, animal-centered approaches to welfare that consider the experiences of individual animals remain central to modern elephant management practices. Recently there has been scientific interest in quantifying the diurnal and nocturnal experiences of elephants and other species in zoos to provide “24/7” husbandry (Brando and Buchanan-Smith, 2018). To accomplish this, animal caretakers and researchers utilize a variety of tools to monitor the wellbeing of individual zoo elephants.
Monitoring animal behavior is an important welfare tool for the management of elephants and other zoo-housed species (Watters et al., 2009, 2019). Circadian rhythms of zoo elephants have been regularly described in the literature; elephants spend approximately 40% of the day foraging (Tresz and Wright, 2006; Lukacs et al., 2016) and typically sleep 25% of the day (Williams et al., 2015; Schiffmann et al., 2018, 2023). Most rest appears to happen at night, although smaller bouts of rest may occur during the day, especially among younger elephants. Rest has been commonly cited as a key indicator of elephant welfare (Walsh, 2017; Schiffmann et al., 2018; Kalirathinam et al., 2019). When resting, elephants may stand upright or be in a recumbent position, but they may only enter rapid eye movement (REM) sleep when recumbent, as they do not require active tension to support themselves against gravity (Schiffmann et al., 2018). REM sleep is important for an animal’s physiological health (Blumberg et al., 2020), and therefore inadequate levels of sleep may reflect a compromised welfare state. Furthermore, stereotypic behavior—any repetitive behavior with no obvious goal or function (Mason, 1991)—has also been commonly used as an important welfare indicator in elephants and other zoo-housed species (Mason and Rushen, 2006). Stereotypic behavior is relatively common in zoo elephants and most typically consists of swaying, head bobbing, and/or pacing (Rees, 2009), with previous studies finding that approximately 60% of elephants in zoos demonstrate some form of stereotypy (Mason and Veasey, 2010). Additionally, stereotypic behavior may be most common following periods of eating in zoo-housed elephants (Greco et al., 2016a), indicating a possible anticipatory function (Krebs et al., 2022). Studies that focus on sleep patterns and stereotypic behavior as two components of behavioral welfare may help us to further enhance elephant wellbeing in zoos.
Despite recent interest in understanding rest-activity rhythms in zoo-housed elephants, few long-term studies have been conducted that focus on overnight stereotypy. Additionally, studies on elephant sleep often focus on groups of elephants (Wilson et al., 2006; Evison et al., 2020; Finch et al., 2021; Thevarajah et al., 2021) rather than on singly housed individuals. Older male elephants are routinely housed separately from other elephants to replicate social structures observed in free-ranging populations (Keerthipriya et al., 2021; LaDue et al., 2022d). Furthermore, during the sexual state of “musth”—a period unique to male elephants that is characterized by elevated testosterone and concomitant behavioral changes—males may be housed separately from females as a precaution to minimize aggression toward conspecifics (LaDue et al., 2022a). Similarly, geriatric elephants may be housed singly for various lengths or frequencies of time to reduce the risk of injury or fatigue resulting from prolonged exposure to younger, more energetic conspecifics. Without prolonged social stimulation, these individually housed elephants may require specialized husbandry to ensure their behavioral repertoire is still conducive to a good welfare standard. In this regard, it can be particularly important to monitor the overnight behavior of these elephants during periods when caretakers are not at the zoo.
The aim of this case study was to evaluate factors that may be associated with the overnight behavior of a geriatric male elephant at the Oklahoma City Zoo and Botanical Garden (“OKC Zoo”, USA) who was housed individually and had a history of stereotypic behavior that presented as swaying. OKC Zoo houses a multi-generational herd of Asian elephants. For 14 hours each day (spanning from 17:00 in the evening to 07:00 the next morning), animal caretakers are off premises, leaving animals to exert a greater amount of choice in their nightly activities without direct intervention from caretakers. We implemented a behavioral monitoring protocol to quantify the impacts of several changes in the focal animal’s environment and physiological state, including prolonged feeding strategies, diverse habitat access, and musth. While caretakers at the Zoo utilize a broad ethogram to quantify behavioral rhythms that can be expected to be associated with these changes, in this case study we focus on recumbency and stereotypy as key indicators of this elephant’s overall welfare. Specifically, we hypothesized that prolonging food availability (through automatic overnight feeders) and increasing environmental complexity (by providing simultaneous access to multiple habitat spaces) would result in lower levels of stereotypic behavior overnight. Additionally, we hypothesized that periods of musth would be associated with higher levels of overnight stereotypy and shorter durations of rest (i.e., recumbency) overnight.
The study was conducted at the OKC Zoo from January 2023 through October 2023. At the time of the study, the Zoo housed three male elephants (including the focal elephant in this case study) and five female elephants, ranging from 1 years to 54 years in January 2023. The base of each elephant’s diet consisted of hay and leafy forage, with sparse fruits and vegetables provided intermittently for enrichment and animal training. Water for drinking was provided ad libitum. The elephant habitat complex at OKC Zoo consists of multiple outdoor habitats with grass and dirt substrate, and a large indoor barn with interconnected stalls (two of which had sand substrate, the others were concrete) (Figure 1).
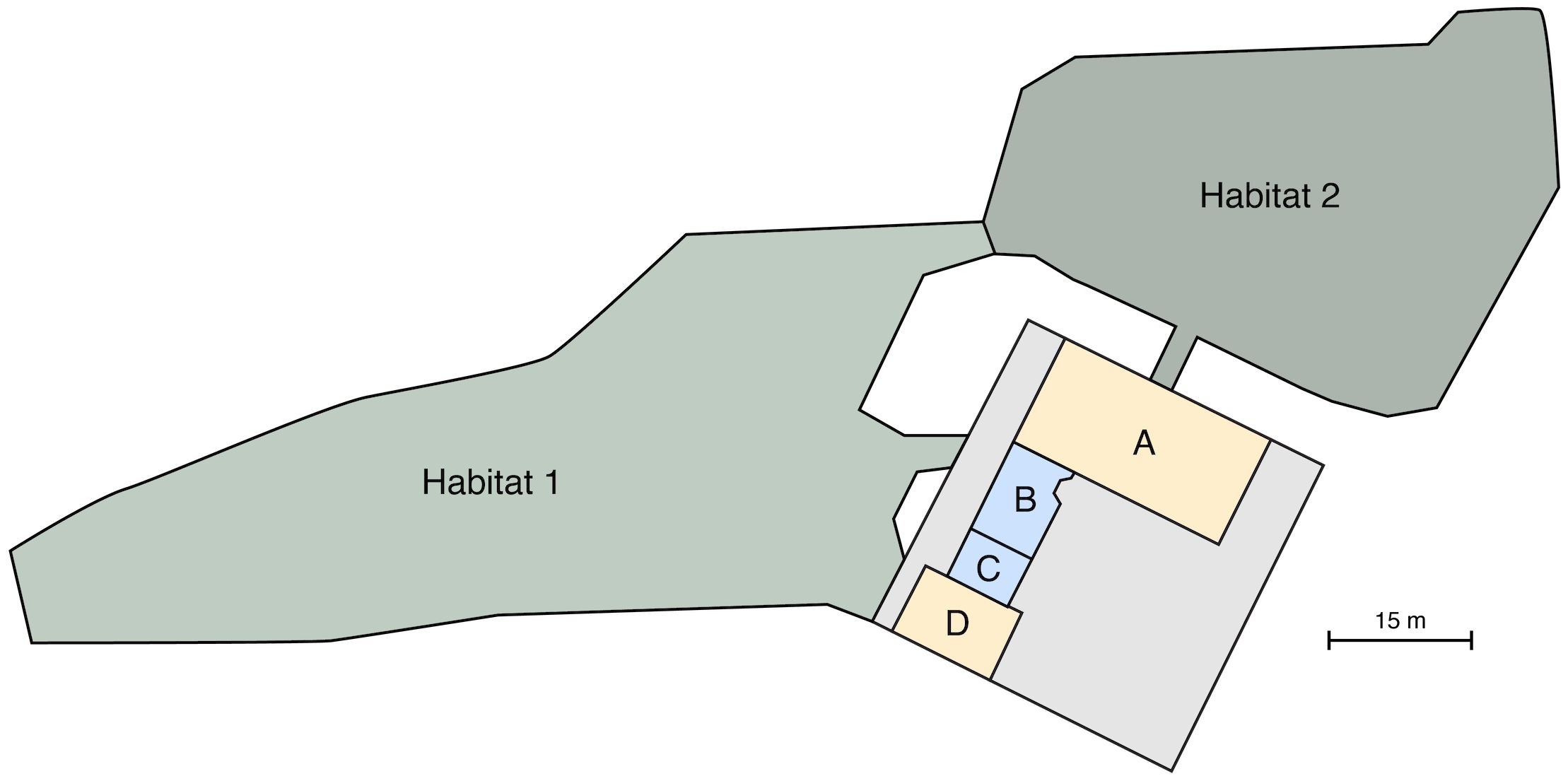
Figure 1 Overview of spaces available to focal elephant during study from 02 Jan 2023 to 30 Oct 2023, drawn to scale. Habitats 1 and 2 are outdoors, and indoor spaces are indicated with letters. Yellow indoor spaces (A, D) had a sand substrate, with automatic feeders installed on 11 Feb 2023. Blue indoor spaces had concrete floors, with no automatic feeders. For overnight observations, access was defined as outdoors (either Habitat 1 or Habitat 2), indoors (some combination of spaces A, B, C, and/or D), or both (some combination of indoor spaces, and either Habitat 1 or Habitat 2). Gray rectangle represents the entire indoor elephant complex; other indoor and outdoor spaces inaccessible to focal animal during this study are not shown.
The subject of this case study is the oldest elephant at the OKC Zoo, a geriatric, wild-born male that was 54 years old. Due to previous incompatibility with other elephants, he was housed individually (except during times of breeding), although he had regular tactile, olfactory, visual, and auditory access to other elephants at the Zoo across a barrier. This elephant has exhibited a long history of stereotypic behavior. Before the study began, he sustained age-related degenerative joint disease. Concomitantly, this elephant has presented with intermittent and recurring toenail abscesses, presumably due to abnormal distribution of weight during stereotypy. Furthermore, in 2017 he was diagnosed with a punctate cataract and corneal mineralization in one eye, presumably impairing his visual capacity.
This elephant’s housing alternated between two of the outdoor habitats (both of which contained a sand pile for resting, a pool, and various hanging and ground-level feeding stations for hay) and/or indoor stalls with sand substrate and concrete flooring. Depending on outdoor temperatures and weather overnight, the elephant in this study was housed outdoors (when overnight temperatures were above 7°C and lower than 37°C), indoors (during extreme weather or when overnight temperatures were lower than 2°C), or with simultaneous access to both indoor and outdoor spaces (when overnight temperatures were between 2°C and 7°C or above 37°C).
During the study period, automatic feeders (Stable Grazer® Generation II Automatic Hay Feeder, HayDay LLC, Hayden, Idaho, USA) were installed on 11 February 2023—approximately one month into the study—to dispense hay at unpredictable intervals in indoor stalls A and D between 04:00 and 06:45 (Table 1). Automatic feeders in Habitats 1 and 2 already had been present for several years before the study began, and they were present for the entire duration of the study as well. During overnight housing in Habitats 1 and 2, these feeders were originally programmed to dispense food between 18:00 and 00:00, shortly after caretakers left grounds each evening. Automatic feeders in Habitat 2 were then reprogrammed to dispense hay in early morning hours (between 04:00 and 06:45) on 03 August 2023. Additionally, the elephant displayed signs of musth (temporal gland secretions and/or urine dribbling) between March and July 2023. While we made no husbandry modifications due to musth, male Asian elephants may be expected to change activity budgets (e.g., decreased foraging, increasing walking and stereotypy) during this period (LaDue et al., 2022c).
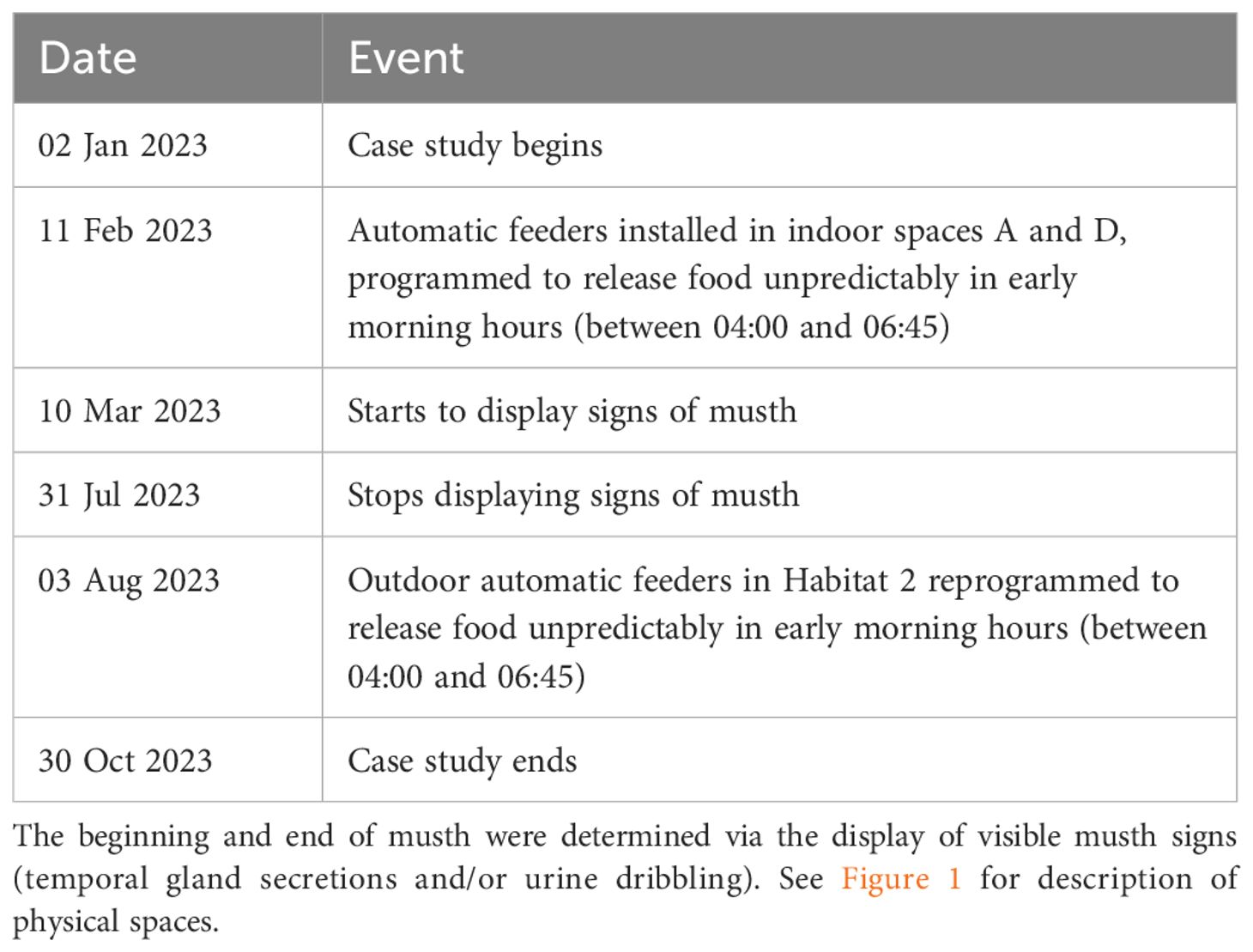
Table 1 Timeline of relevant events in the case study, including habitat modifications and occurrence of annual musth period.
We conducted continuous focal-animal sampling of the focal elephant’s behavior between 20:00 and 08:00 using infrared cameras present in indoor and outdoor habitats (Altmann, 1974), reviewing the camera footage within two days of each recording session. We aimed to collect data on at least four randomly selected nights each week. Over approximately ten months, we conducted 179 overnight observations (mean ± SD observations per month = 17.9 ± 4.0, min = 11, max = 24). During each observation, we noted the beginning and end of each bout of recumbency (i.e., “recumbent rest”) and stereotypy. Recumbent rest was defined as any instance when the body was touching the ground with at least two soles off the ground, encompassing both sternal and lateral recumbency. Bouts of stereotypic behavior were initiated with the start of repetitive swaying and ended when swaying stopped for a minimum of 30 seconds; swaying bouts were identified when the elephant was stationary in space on all four legs, visibly shifting weight from one side of the body to the other for at least two repetitions. No other forms of stereotypy were noted in the focal elephant before or during the study.
All analyses were carried out using R version 4.1.0 (R Core Team, 2021), with the following packages: AICc-modavg (Mazerolle, 2019), lme4 (Bates et al., 2015), and tidyverse (Wickham et al., 2019). For relevant analyses, statistical significance was set at α = 0.05.
For each overnight observation, we calculated the total time engaged in recumbent rest and stereotypy (separately), to the nearest minute. To determine putative effects of space access (indoors only, outdoors only, or both indoors and outdoors) and musth status on duration of recumbent rest or stereotypy, we implemented a linear mixed model approach using a ranked model selection approach (Johnson and Omland, 2004; Zuur and Ieno, 2016). For each behavior, we created a list of candidate models that included additive permutations of fixed effects (space access and musth status), with observation month as a random intercept in each model to account for temporal autocorrelation. Candidate models were ranked via Akaike Information Criterion (AIC) values with maximum likelihood estimation. We used Kruskal-Wallis rank sum tests for post-hoc analyses to identify behavioral differences between states (space access and/or musth status).
Each night of observations, we summed the number of minutes the elephant was in recumbent rest or engaging in stereotypy (separately) each hour from 20:00 to 08:00. Then for each night, we calculated the hourly proportion of recumbency or stereotypy by dividing each hour’s sum by the total number of recumbency/stereotypy for the night. To test for differences in the proportion of each night engaged in recumbent rest or stereotypy, we constructed linear mixed models with hour as a fixed effect and month as a random intercept. Significant differences between hours for each behavior were identified via Kruskal-Wallis rank sum tests with Bonferroni corrections applied to resulting P-values due to a high number of pairwise comparisons.
Additionally, to evaluate the impact of automatic feeders on mitigating high rates of stereotypy, we analyzed a subset of the data for the proportion of time spent swaying before and after the feeders were installed/programmed to dispense food. The automatic feeders for indoor spaces were installed on 11 February 2023. We identified nights during which the elephant had access to at least one of these feeders one month before and three months after the installation, only including nights in this period in which the elephant was in a space where feeders would be/were installed (n = 33). On each of these nights, we calculated the proportion of time engaged in stereotypy between 04:00 and 08:00, around the time when the feeders were programmed to dispense food (between 04:00 and 06:45). Automatic feeders in Habitat 2 were reprogrammed from late evening hours (18:00 to 00:00) to dispense food during early morning hours starting on 03 August 2023. Similarly, we identified nights when the elephant had access to these outdoors feeders one month before and approximately three months after this date and calculated time spent engaged in stereotypy between 04:00 and 08:00 (n = 31). For each of these scenarios (indoor feeders and outdoor feeders), we utilized Kruskal-Wallis rank sum tests to identify differences in stereotypy between each of the four months (one month prior to and three months following the automatic feeders) due to uneven sampling between months.
Over the study period, the elephant exhibited recumbent rest for 175.66 ± 6.80 (mean ± SE) minutes per night and was engaged in stereotypy for 175.88 ± 9.68 minutes per night. There were no changes in the overnight duration of either recumbent rest or stereotypy due to time of year (Supplementary Figure 1). However, there was a significant influence of musth status on recumbent rest (Supplementary Table 1A); our model selection process revealed that variation in recumbency was best explained by musth status but not habitat access, with shorter nightly durations of recumbency observed during musth compared to non-musth (Supplementary Table 2A; Figure 2A; χ21 = 4.186, P = 0.041). Conversely, variation in observed stereotypy was best explained by habitat access and not musth status (Supplementary Table 1B). In other words, there was a significant difference in the duration of stereotypic swaying between the three levels of habitat access (Supplementary Table 2B; χ22 = 7.505, P = 0.023). Post-hoc analyses revealed significantly more stereotypy in days with only indoor access compared to those where both indoor and outdoor access was permitted; there was no significant difference in stereotypy duration between days with indoor-only or outdoor-only access (Figure 2B).
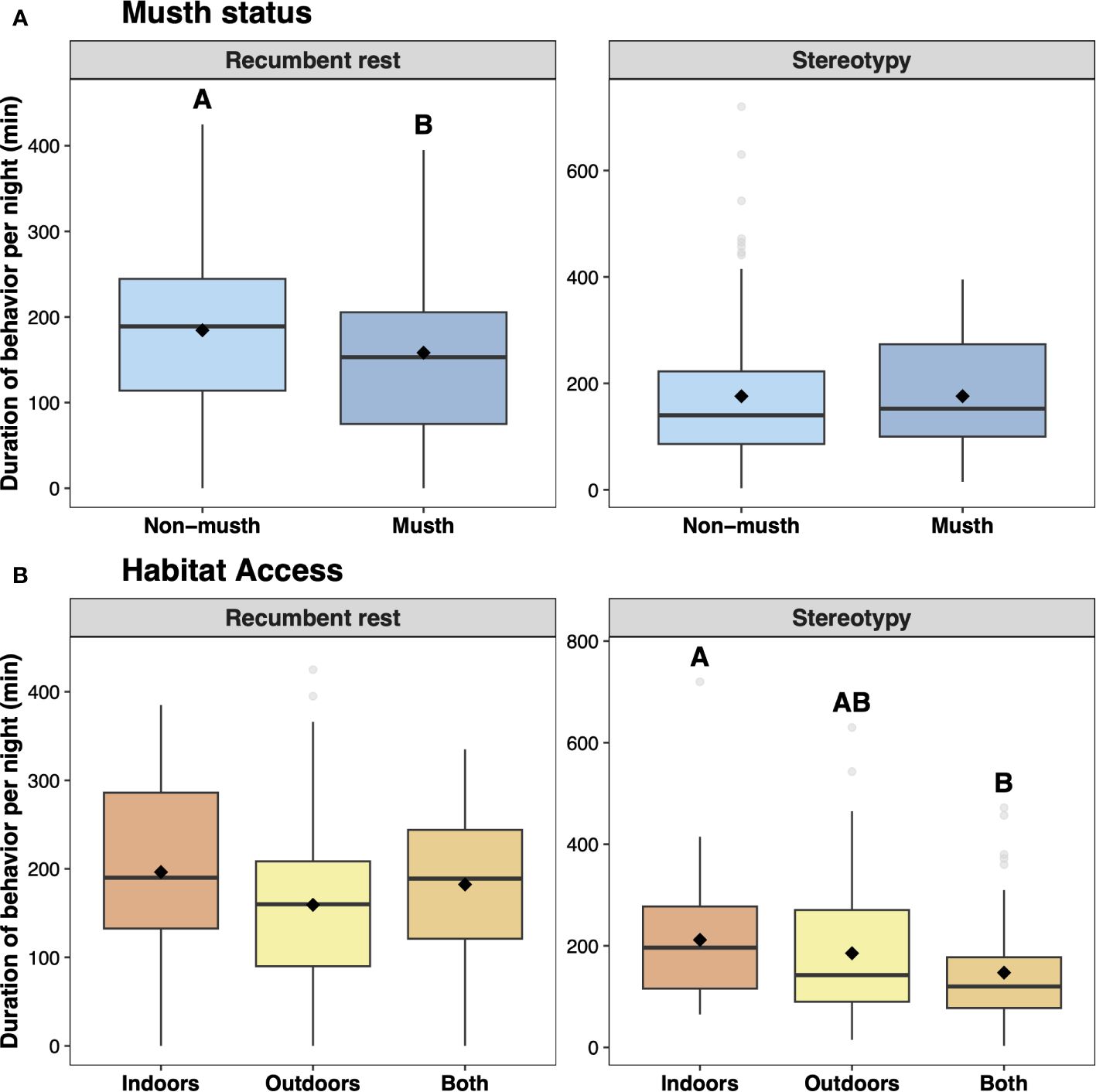
Figure 2 Boxplots of recumbent rest and stereotypy durations per night of a geriatric male Asian elephant based on (A) musth status and (B) habitat access. Boxes extend from the first to third quartiles, with median indicated by a thick line and mean by a diamond; fences extend to 1.5 times the interquartile range, and closed circles indicate values outside this range. Different letters above boxes indicate statistically significant differences between (A) non-musth and musth, and between (B) habitat access; absence of letters indicate no significant differences between these treatments.
There were clear behavioral rhythms across the night for both recumbent rest (χ211 = 531.13, P < 0.001) and stereotypy (χ211 = 557.80, P < 0.001) (Supplementary Table 3). The average time spent in recumbency increased with the night’s progression, with sleep most commonly occurring between 02:00 and 05:00 and decreasing until 08:00 (Figure 3A). Conversely, time spent in stereotypy was relatively minimal between 20:00 and 04:00, increasing dramatically between 06:00 and 08:00 (Figure 3B).
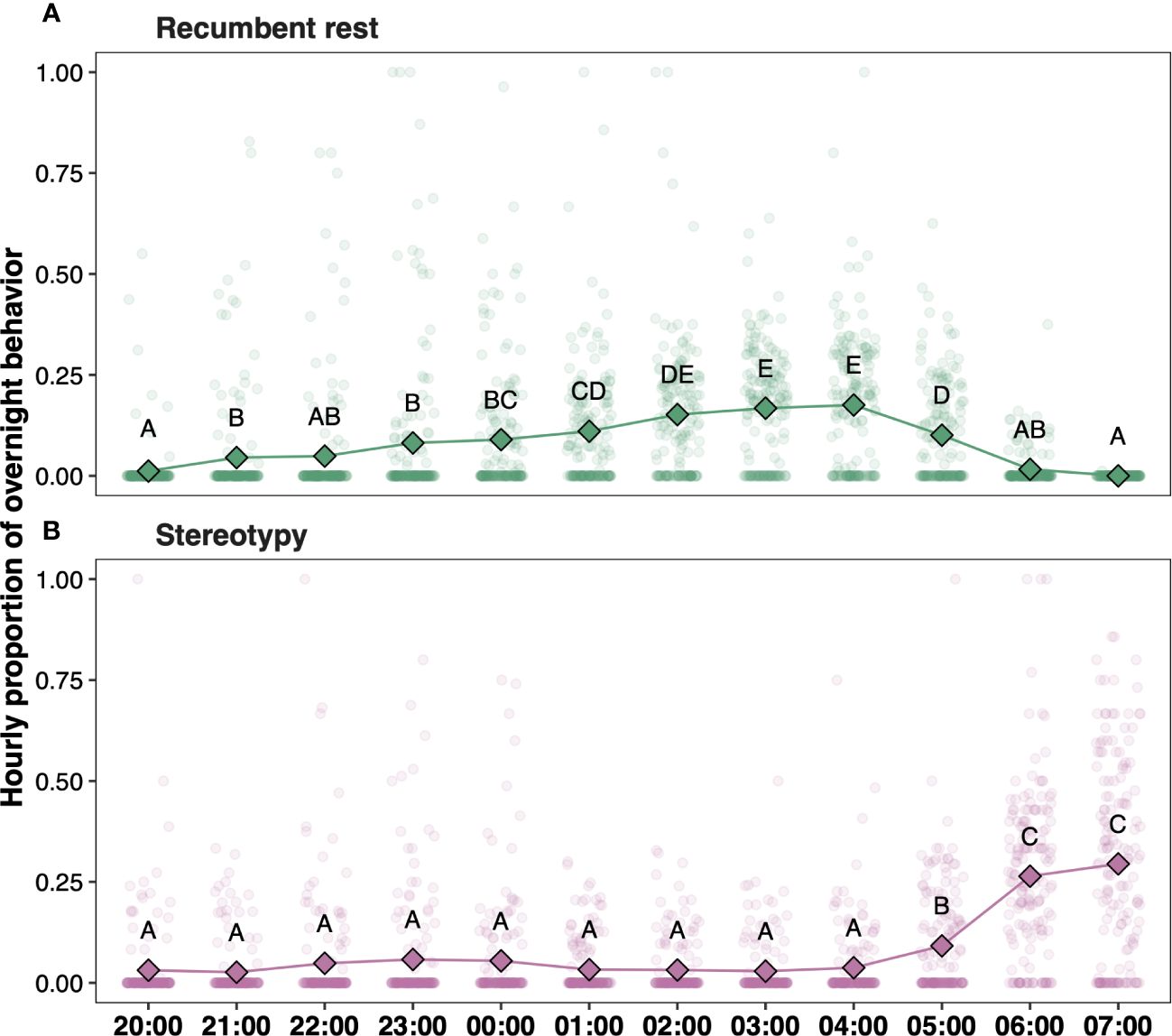
Figure 3 Hourly proportions per night engaged in (A) recumbent rest and (B) stereotypy. Diamonds represent average values for each hour across the study period, with points denoting individual observations (jittered for ease of interpretation). Letters indicate statistically significant hourly differences between (A) recumbent rest and (B) stereotypy.
Finally, while there was a decreasing trend in the amount of time engaged in stereotypy between 04:00 and 08:00 as a result of installing the automatic feeders in indoor spaces, there were no significant monthly differences between the four months we analyzed (χ23 = 5.79, P = 0.122) (Supplementary Figure 2A). Similarly, there was no apparent difference in stereotypy between 04:00 and 08:00 as a result of reprogramming the outdoor feeders (χ23 = 5.54, P = 0.136) (Supplementary Figure 2B).
This case study embodies the value of an animal-centered approach to behavioral monitoring to enhance zoo elephant wellbeing. Recumbent rest and stereotypy can be useful behavioral indicators with which to measure elephant welfare, and while we observed variation in the extent by which the focal elephant engaged in these behaviors overnight, we also identified factors associated with this variation. First, shorter durations of recumbent rest were associated with musth. Further, it appeared that habitat access (whether indoors, outdoors, or a combination of the two) influenced stereotypical behavior, with longer durations associated with indoor-only access and shorter durations with the choice of indoor and outdoor spaces. We also noted distinct behavioral rhythms associated with time of the night—recumbent rest was most common between 02:00 and 05:00, along with stereotypy increasing dramatically between 06:00 and 08:00. While we did not observe significant effects of automatic feeders on stereotypic behavior, there was a statistically nonsignificant trend of decreasing incidence of stereotypy with prolonged feeding opportunities in indoor spaces. While the results presented here represent a single elephant, this study provides an example of the value of behavioral monitoring in zoos using an animal-centered approach. With greater emphasis on the 24-hour experience of zoo animals, it also is important to monitor the behavior of animals overnight, when caretakers are not present. Furthermore, as zoo-housed elephant populations age and we produce more male elephants through ex-situ breeding efforts, our study will be representative of many of the elephants (i.e., geriatric, individually housed males) that may be a challenge for sustainable management strategies into the future.
Providing zoo-housed animals with choice over their environment has been suggested to have considerable impact on animal wellbeing (Powell and Vitale, 2016). Notably in this study, we found a decline in stereotypic behavior when the focal individual had access to both indoor and outdoor habitats even though stereotypy did not differ between indoor- and outdoor-only conditions, providing supportive evidence for the value of choice for elephants in zoos. Among zoo elephants, the ability to choose between indoor and outdoor experiences is associated with decreased stereotypic behavior (Greco et al., 2016a; Schiffmann et al., 2019). In implementing a data-driven approach to mitigate stereotypy in this elephant, we may manipulate environmental complexity (by providing access to multiple habitats) to enhance overall wellbeing (Scott and LaDue, 2019).
Anecdotally, we observed a preference in this focal elephant for indoor sand stalls as he often chooses to go inside to sleep, rather than resting on the outdoor sand piles or sloped ground. Additionally, we found that the focal elephant exclusively chose to sleep on substrate rather than concrete when housed indoors, corroborating the results of other studies on rest in zoo-housed elephants (Walsh, 2017). As elephants primarily sleep at night, it is unsurprising that we most commonly observed recumbent rest in this elephant between 02:00-05:00. Due to his geriatric age (54 years), it is important that the focal elephant consistently rests in a recumbent position, as older elephants may be reluctant to lie down due to hesitancy to get back up (Schiffmann et al., 2018). We did not observe many instances of recumbency-free nights in this elephant, and his average rest time was approximately 179 minutes per night. Moving forward as this elephant ages, we plan to use the measure of recumbency duration as an important indicator for wellbeing to guide the care that we provide to this elephant.
Even though we did not find a significant impact of musth on stereotypy in this case study, we did describe a significant negative association of musth with recumbency. Musth poses distinct challenges to elephant management (LaDue et al., 2022a, LaDue et al., 2022b), and we now understand the importance of providing male elephants with complex, engaging environments. In understanding how behavioral rhythms change with musth (including overnight behavior), we may provide targeted interventions to bolster animal health and wellbeing during this period that is physiologically taxing for male elephants. As described in previous studies, the focal male appeared to have a decreased appetite and increased locomotion with musth, changes that presumably are associated with altered hormonal profiles and increased reproductive activity (Keerthipriya et al., 2020; LaDue et al., 2022c). Wild male elephants in musth often traverse long distances to pursue estrous females (Fernando et al., 2008), and evidently this behavioral adaptation persists in ex-situ environments (LaDue et al., 2022c). In the present case study, this change in behavioral rhythm was manifested through decreased recumbent rest, and so now we know that it may be important to provide this elephant with environmental conditions that promote rest, especially given his advanced age.
We originally hypothesized that automatic feeders would decrease stereotypic swaying in the focal elephant, as the randomized times would prolong feeding time and decrease anticipation. Indeed, other studies in zoo-housed elephants have suggested that unpredictable and diverse feeding schedules can be a valuable tool in reducing stereotypy (Wilson et al., 2004; Greco et al., 2016b; Sullivan and Fazio, 2018). However, our case study revealed no significant relationship between the automatic feeders and stereotypy. Anecdotal observations indicate that the presence of the feeders instead caused the individual to engage in swaying throughout the night and reduced recumbent rest; we observed shorter durations of recumbent rest interrupted by bouts of activity (including stereotypy) to check or wait for the presence of food at the automatic feeders. Given the schedule of this elephant’s stereotypic swaying (with a high prevalence in the hours just before caretakers arrive for the day), there is a strong possibility that this behavior is anticipatory in nature (Krebs et al., 2022). In prolonging the possibility of food—and also providing smaller amounts of food at each feeding opportunity as a result—we may have inadvertently removed an otherwise reliable cue of a desired consequence, thereby increasing anticipation across a longer period of the night (Podturkin et al., 2023).
With these results in mind, we are not advocating for an abandonment of diverse feeding strategies; while statistically nonsignificant, our data indicate that automatic feeders may have a positive impact in simpler indoor spaces that may lack the complexity of outdoor spaces. Instead, we suggest that feeding strategies (including the use and timing of automatic feeders) be carefully considered and targeted with behavioral motivations and outcomes in mind. For the elephant in this study, we are now exploring ways that may decrease stereotypy while also promoting species-appropriate behavioral diversity.
For the focal elephant in this study, this work has played a principal role in challenging the assumptions we had about the temporal qualities and motivations of rest and stereotypy. By using an adaptive and data-driven approach, we have implemented targeted management strategies to enhance animal wellbeing, always evaluating the impact of these changes. In particular, we have gained better insight into the potential motivations behind this elephant’s stereotypy, even as his overnight behavior may be largely outside the influence of caretaker presence. While it is unlikely that overnight stereotypy will be completely eliminated given its semi-permanent neurophysiological nature (Mason and Rushen, 2006), this case study has been an important factor in providing comprehensive animal care for the focal elephant.
In understanding the behavioral rhythms of the animals in our care, this study demonstrates the value of overnight monitoring to promote animal wellbeing through an animal-centered approach. A true “24/7” approach to animal wellbeing requires that our management strategies grow and change with the animals in our care, and this is certainly relevant to ex-situ elephant populations. As we develop targeted interventions to enhance the lives of elephants in zoos (and aging male elephants in particular), further work is needed to investigate the relationship between musth and its impact on sleep and stereotypic behavior. Furthermore, as we better understand the causes and consequences of stereotypic behavior, future studies should evaluate factors that influence the success of automatic timed feeders and the impact they may have on overnight behavior of elephants. Animal-centered approaches challenge animal managers to evaluate even widely held assumptions. With the development of innovative and effective tools with which we can understand the lives of the animals in our care, it remains important to assess the influence of our management on individual animals.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was approved by Oklahoma City Zoo and Botanical Garden. The study was conducted in accordance with the local legislation and institutional requirements (Association for the Study of Animal Behaviour and Animal Behavior Society, 2023).
AM: Conceptualization, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing. MK: Conceptualization, Investigation, Methodology, Project administration, Writing – review & editing. RE: Conceptualization, Methodology, Project administration, Resources, Supervision, Writing – review & editing. CL: Data curation, Formal analysis, Methodology, Project administration, Supervision, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
We would like to acknowledge the OKC Zoo elephant team for supporting this research, and Sarah Fairchild for her assistance with data collection. We also thank Rebecca Snyder for reviewing an earlier draft of this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2024.1362313/full#supplementary-material
Altmann J. (1974). Observational study of behavior: sampling methods. Behaviour 49, 227–267. doi: 10.1163/156853974X00534
Association for the Study of Animal Behaviour, Animal Behavior Society. (2023). Guidelines for the ethical treatment of nonhuman animals in behavioural research and teaching. Anim. Behav. 195, i–xi. doi: 10.1016/j.anbehav.2022.09.006
Association of Zoos and Aquariums (2023). The Accreditation Standards & Related Policies (Silver Spring, MD: AZA).
Bates D., Maechler M., Bolker B., Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Blumberg M. S., Lesku J. A., Libourel P.-A., Schmidt M. H., Rattenborg N. C. (2020). What is REM sleep? Curr. Biol. 30, R38–R49. doi: 10.1016/j.cub.2019.11.045
Brando S., Buchanan-Smith H. M. (2018). The 24/7 approach to promoting optimal welfare for captive wild animals. Behav. Processes 156, 83–95. doi: 10.1016/j.beproc.2017.09.010
Evison E., McKenzie A., Holmes L. (2020). Social and environmental impacts on sleep in captive Asian elephants (Elephas maximus). Zoo Biol. 39, 397–404. doi: 10.1002/zoo.21568
Fernando P., Wikramanayake E. D., Janaka H. K., Jayasinghe L. K. A., Gunawardena M., Kotagama S. W., et al. (2008). Ranging behavior of the Asian elephant in Sri Lanka. Mamm. Biol. 73, 2–13. doi: 10.1016/j.mambio.2007.07.007
Finch K., Sach F., Fitzpatrick M., Rowden L. J. (2021). Insights into activity of zoo housed Asian elephants (Elephas maximus) during periods of limited staff and visitor presence, a focus on resting behaviour. J. Zoological Botanical Gardens 2, 101–114. doi: 10.3390/jzbg2010008
Greco B. J., Meehan C. L., Hogan J. N., Leighty K. A., Mellen J., Mason G. J., et al. (2016a). The days and nights of zoo elephants: Using epidemiology to better understand stereotypic behavior of African elephants (Loxodonta africana) and Asian elephants (Elephas maximus) in North American Zoos. PloS One 11, e0144276. doi: 10.1371/journal.pone.0144276
Greco B. J., Meehan C. L., Miller L. J., Shepherdson D. J., Morfeld K. A., Andrews J., et al. (2016b). Elephant management in North American zoos: Environmental enrichment, feeding, exercise, and training. PloS One 11, e0152490. doi: 10.1371/journal.pone.0152490
Johnson J. B., Omland K. S. (2004). Model selection in ecology and evolution. Trends Ecol. Evol. 19, 101–108. doi: 10.1016/j.tree.2003.10.013
Kalirathinam U. K., Elangkovan S., Kawi J., Cabana F. (2019). Sleep monitoring of an Asian elephant Elephas maximus calf at Night Safari, Singapore: testing whether sleep time is a significant predictor of cortisol or the onset of positive elephant endotheliotropic herpesvirus viraemia. Int. Zoo Yearbook 53, 128–137. doi: 10.1111/izy.12239
Keerthipriya P., Nandini S., Gautam H., Revathe T., Vidya T. N. C. (2020). Musth and its effects on male–male and male–female associations in Asian elephants. J. Mammal. 101, 259–270. doi: 10.1093/jmammal/gyz190
Keerthipriya P., Nandini S., Vidya T. N. C. (2021). Effects of male age and female presence on male associations in a large, polygynous mammal in southern India: the Asian elephant. Front. Ecol. Evol. 9, 616666. doi: 10.3389/fevo.2021.616666
Krebs B. L., Chudeau K. R., Eschmann C. L., Tu C. W., Pacheco E., Watters J. V. (2022). Space, time, and context drive anticipatory behavior: considerations for understanding the behavior of animals in human care. Front. Vet. Sci. 9, 972217. doi: 10.3389/fvets.2022.972217
LaDue C. A., Hunt K. E., Samaraweera M. G. S. M., Vandercone R. P. G., Kiso W. K., Freeman E. W. (2022a). Physical and behavioral indicators are associated with hormonal changes during the sexual state of musth in zoo-housed and free-ranging Asian elephants (Elephas maximus). Theriogenol. Wild 1, 100011. doi: 10.1016/j.therwi.2022.100011
LaDue C. A., Schulte B. A., Kiso W. K., Freeman E. W. (2022b). Musth and sexual selection in elephants: a review of signaling properties and potential fitness consequences. Behaviour 159, 207–242. doi: 10.1163/1568539X-bja10120
LaDue C. A., Vandercone R. P. G., Kiso W. K., Freeman E. W. (2022c). Behavioral characterization of musth in Asian elephants (Elephas maximus): defining progressive stages of male sexual behavior in in-situ and ex-situ populations. Appl. Anim. Behav. Sci. 251, 105639. doi: 10.1016/j.applanim.2022.105639
LaDue C. A., Vandercone R. P. G., Kiso W. K., Freeman E. W. (2022d). Social behavior and group formation in male Asian elephants (Elephas maximus): the effects of age and musth in wild and zoo-housed animals. Animals 12, 1215. doi: 10.3390/ani12091215
Lukacs D. E., Poulin M., Besenthal H., Fad O. C., Miller S. P., Atkinson J. L., et al. (2016). Diurnal and nocturnal activity time budgets of Asian elephants (Elephas maximus) in a zoological park. Anim. Behav. Cogn. 3, 63–77. doi: 10.12966/abc.01.05.2016
Mason G., Rushen J. (2006). Stereotypic Animal Behaviour: Fundamentals and Applications to Welfare. 2nd ed (Oxfordshire, UK: CABI).
Mason G. J. (1991). Stereotypies: a critical review. Anim. Behav. 41, 1015–1037. doi: 10.1016/S0003-3472(05)80640-2
Mason G. J., Veasey J. S. (2010). What do population-level welfare indices suggest about the well-being of zoo elephants? Zoo Biol. 29, 256–273. doi: 10.1002/zoo.20303
Mazerolle M. J. (2019). AICcmodavg: model selection and multimodel inference based on (Q)AIC(c), R package version 2.2-2 ed.
Meehan C. L., Mench J. A., Carlstead K., Hogan J. N. (2016). Determining connections between the daily lives of zoo elephants and their welfare: An epidemiological approach. PloS One 11, e0158124. doi: 10.1371/journal.pone.0158124
Podturkin A. A., Krebs B. L., Watters J. V. (2023). Quantifying animals’ perception of environmental predictability using anticipatory behavior. Zoo Biol. doi: 10.1002/zoo.21811
Powell D. M., Vitale C. (2016). Behavioral changes in female Asian elephants when given access to an outdoor yard overnight. Zoo Biol. 35 (4), 298–303. doi: 10.1002/zoo.21289
R Core Team. (2021). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing).
Rees P. A. (2009). Activity budgets and the relationship between feeding and sterotypic behaviors in Asian elephants (Elephas maximus) in a zoo. Zoo Biol. 28, 79–97. doi: 10.1002/zoo.20200
Riddle H. S., Stremme C. (2011). Captive elephants - an overview. J. Threatened Taxa 3, 1826–1836. doi: 10.11609/JoTT.o2620.1826-36
Schiffmann C., Clauss M., Hoby S. (2019). Impact of a new exhibit on stereotypic behaviour in an elderly captive African elephant (Loxodonta africana). J. Zoo Aquarium Res. 7, 37–43. doi: 10.19227/jzar.v7i1.358
Schiffmann C., Hellriegel L., Clauss M., Stefan B., Knibbs K., Wenker C., et al. (2023). From left to right all through the night: characteristics of lying rest in zoo elephants. Zoo Biol. 42, 17–25. doi: 10.1002/zoo.21693
Schiffmann C., Hoby S., Wenker C., Hård T., Scholz R., Clauss M., et al. (2018). When elephants fall asleep: a literature review on elephant rest with case studies on elephant falling bouts, and practical solutions for zoo elephants. Zoo Biol. 37, 133–145. doi: 10.1002/zoo.21406
Scott N. L., LaDue C. A. (2019). The behavioral effects of exhibit size versus complexity in African elephants: a potential solution for smaller spaces. Zoo Biol. 38, 448–457. doi: 10.1002/zoo.21506
Sullivan K., Fazio J. (2018). The future of managing elephants with use of automatic feeder systems. J. Elephant Managers Assoc. 29, 98–101.
Thevarajah S. J., Readyhough T. S., Davis M., Moresco A., Joseph S., Schreier A. L. (2021). Nighttime behavior and the length of social relationships in male Asian elephants. J. Appl. Anim. Welfare Sci. 26, 447–462. doi: 10.1080/10888705.2021.1998777
Tresz H., Wright H. (2006). Let them be elephants! How Phoenix Zoo integrated three ‘problem’ animals. Int. Zoo News 348, 154.
Walsh B. (2017). Sleep in Asian elephants (Elephas maximus): long-term quantitative research at Dublin Zoo. J. Zoo Aquarium Res. 5, 82–85. doi: 10.19227/jzar.v5i2.174
Watters J. V., Krebs B. L., Pacheco E. (2019). “Measuring welfare through behavioral observation and adjusting it with dynamic environments,” in Scientific Foundations of Zoos and Aquariums: Their Role in Conservation and Research. Eds. Kaufman A. B., Bashaw M. J., Maple T. L. (Cambridge University Press, Cambridge, UK), 212–240.
Watters J. V., Margulis S. W., Atsalis S. (2009). Behavioral monitoring in zoos and aquariums: a tool for guiding husbandry and directing research. Zoo Biol. 28, 35–48. doi: 10.1002/zoo.20207
Wickham H., Averick M., Bryan J., Chang W., McGowan L. D. A., François R., et al. (2019). Welcome to the tidyverse. J. Open Source Softw. 4, 1686. doi: 10.21105/joss.01686
Williams E., Bremner-Harrison S., Harvey N., Evison E., Yon L. (2015). An investigation into resting behavior in Asian elephants in UK zoos. Zoo Biol. 34, 406–417. doi: 10.1002/zoo.21235
Wilson M. L., Bashaw M. J., Fountain K., Kieschnick S., Maple T. L. (2006). Nocturnal behavior in a group of female African elephants. Zoo Biol. 25, 173–186. doi: 10.1002/zoo.20084
Wilson M. L., Bloomsmith M. A., Maple T. L. (2004). Stereotypic swaying and serum cortisol concentrations in three captive African elephants (Loxodonta africana). Anim. Welfare 13, 39–43. doi: 10.1017/S0962728600026646
Keywords: Asian elephant, rest, recumbency, sleep, stereotypy, welfare, wellbeing, zoo
Citation: McGuire A, Kienlen M, Emory R and LaDue CA (2024) Overnight monitoring reveals the behavioral rhythms of a geriatric male elephant: an animal-centered case study of rest and stereotypy. Front. Conserv. Sci. 5:1362313. doi: 10.3389/fcosc.2024.1362313
Received: 28 December 2023; Accepted: 13 March 2024;
Published: 25 March 2024.
Edited by:
Anton Baotic, Austrian Academy of Sciences (OeAW), AustriaReviewed by:
Christian Schiffmann, Tbz Pariv, GermanyCopyright © 2024 McGuire, Kienlen, Emory and LaDue. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chase A. LaDue, Y2hhc2UubGFkdWVAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.