 Eric A. VanderWerf
Eric A. VanderWerf Nick D. Holmes2
Nick D. Holmes2 Scott A. Morrison
Scott A. Morrison Lindsay C. Young
Lindsay C. Young- 1Pacific Rim Conservation, Honolulu, HI, United States
- 2The Nature Conservancy, Sacramento, CA, United States
Laysan (Phoebastria immutabilis) and Black-footed Albatrosses (P. nigripes) nest primarily on low-lying atolls in the Northwestern Hawaiian Islands that are threatened by inundation from sea level rise and increasing storm surge associated with climate change. Restoration or creation of breeding colonies on higher islands is among the highest priority conservation actions for these species. A previous structured decision-making analysis identified the California Channel Islands as a possible restoration site for Black-footed Albatross. The California Current is part of the natural foraging ranges of Laysan and Black-footed albatrosses. Archaeological evidence indicates both species were present in the California Channel Islands prehistorically, yet neither currently nests in the Channel Islands. We assessed the feasibility of creating albatross breeding colonies in the Channel Islands using social attraction and translocation, and the suitability of each island. We used a risk analysis framework developed for the U.S. National Park Service to evaluate the potential ecological risks of this action. Creating an albatross colony in the Channel Islands is feasible using available methods. Santa Barbara and San Nicolas islands would be most suitable for albatross. Social attraction is less expensive and might be effective for creating a Laysan Albatross colony because that species is already visiting some islands. Translocation would be necessary to create a Black-footed Albatross colony. The risks associated with attempting to establish albatross breeding colonies in the Channel Islands were deemed to be generally low, but the risk of no action is high to these albatrosses. This can be a useful assisted colonization case study that can inform decisions by land managers and agencies regarding conservation of North Pacific albatrosses and other species.
Introduction
More than 95% of the global populations of Laysan Albatross (Phoebastria immutabilis) and Black-footed Albatross (P. nigripes) nest on low-lying atolls in the Northwestern Hawaiian Islands that have a maximum elevation of just a few meters above sea level (U.S. Fish and Wildlife Service 2005 [USFWS], Arata et al., 2009). These islands and the biota they support are threatened by sea level rise and increasing storm surge associated with global climate change, as well as catastrophic events like tsunamis (USFWS, 2005, Baker et al., 2006; Reynolds et al., 2015; Reynolds et al., 2017). Protection of suitable breeding habitat and restoration or creation of breeding colonies on higher islands are among the highest priority conservation actions for increasing resiliency of these species (Flint et al., 2011; Young et al., 2012; VanderWerf et al., 2019).
Some species are responding naturally to climate change by shifting their range toward higher latitudes, higher elevations, and in other ways, but the capacity of many species to shift in range is limited by their physical ability, behavior, or geophysical barriers (Thomas et al., 2004; Freeman and Freeman, 2014; MacLean and Beissinger, 2017; Dunn and Møller, 2019). One conservation strategy to help facilitate range shifts is deliberately moving a species to a new location, which is referred to as managed relocation (Richardson et al., 2009; Karasov-Olson et al., 2021a), assisted colonization (Hoegh-Guldberg et al., 2008; Seddon, 2010), or assisted migration (McLachlan et al., 2007), all of which are sometimes termed conservation introductions (Seddon, 2010). We use the term assisted colonization because it is most relevant to albatrosses, which breed colonially, and because the action that is the subject of this paper would not be an introduction.
There has been much debate about whether the potential risks of this type of action outweigh the risks of possible extinction resulting from inaction (McLachlan et al., 2007; Ricciardi and Simberloff, 2009; Richardson et al., 2009; Maier and Simberloff, 2016). For example, managers may face difficult questions about the risk of species becoming invasive or other unforeseen consequences (Kostyack et al., 2011; Lawler and Olden, 2011; Wallingford et al., 2020). The potential benefits and risks of assisted colonization must be judged on a case-by-case basis, and there have been several efforts to provide methods for evaluating the value and risks associated with assisted colonization projects (McLachlan et al., 2007; Richardson et al., 2009). The most rigorous framework yet for assessing assisted colonization projects was provided by Karasov-Olson et al. (2021a); Karasov-Olson et al. (2021b), as part of a collaborative process funded by the U.S. National Park Service (NPS).
In 2011, the USFWS convened a structured decision-making workshop in Hawai’i to help guide management of the Black-footed Albatross to mitigate effects of climate change (Flint et al., 2011). The workshop identified several actions that could be undertaken to increase the resiliency of the species to climate change, including translocation to, and social attraction at, higher islands, and recommended this be done on three Mexican islands, two California islands, and three main Hawaiian Island sites. Flint et al. (2011) did not specify islands by name, but the focus of discussion regarding California islands centered on the Channel Islands of southern California (E. Flint, USFWS, pers. comm.). The Channel Islands are a logical place to evaluate potential albatross colony creation because of the importance of the California Current for these species’ foraging. The California Current is a productive cold-water current that flows southward along western North America, beginning off southern British Columbia, Canada and ending off the southern tip of the Baja Peninsula, Mexico. Laysan Albatrosses breeding on Guadalupe Island, roughly 350 km southwest of Ensenada, Mexico, forage primarily in the California Current and have higher reproductive output than their Hawaiian counterparts (Hernández-Montoya et al., 2014), highlighting the potential value of additional colonies in this region. The Farallon Islands, approximately 50 km offshore of San Francisco, California, are less suitable for albatrosses because they have little flat terrain and support high densities of breeding gulls, which are potential nest predators (see below). In the main Hawaiian Islands, efforts are underway to restore or create Laysan Albatross breeding colonies at James Campbell National Wildlife Refuge (JCNWR) on O’ahu, and Black-footed Albatross breeding colonies at JCNWR and Kaena Point, O’ahu (Young and VanderWerf, 2016; VanderWerf et al., 2019). In Mexico, translocation of Black-footed Albatrosses to Guadalupe Island began in 2021 (Pacific Rim Conservation and Grupo de Ecología y Conservación de Islas, unpublished data). No such actions have been evaluated or undertaken on any islands in California.
Methods for creation and restoration of seabird colonies are well established. There are two primary methods: social attraction and translocation. Social attraction involves attracting seabirds to a site with visual, auditory, and occasionally olfactory lures (Jones and Kress, 2012; VanderWerf et al., 2022; Spatz et al., 2023). Translocation involves physically moving birds from one location to another, usually when they are chicks, and caring for them until they fledge (Gummer, 2003; Deguchi et al., 2012; Jacobs et al., 2020; VanderWerf et al., 2022). These techniques have been used in at least 857 projects involving 138 seabird species at 550 sites around the world (Spatz et al., 2023). The effectiveness of social attraction and translocation for restoring or creating seabird breeding colonies depends on multiple factors, including the natural history of the species involved, the biotic and abiotic characteristics of the restoration site, and proximity to the nearest existing colony of the target species (Jones and Kress, 2012; Buxton et al., 2014; VanderWerf et al., 2019; VanderWerf et al., 2022). Social attraction alone is more likely to be effective in colonial species with weak natal philopatry and that require post-fledging parental care, and where existing colonies of the target species are close enough that birds are likely to fly near the site. Translocation is necessary more often in species with strong natal philopatry, including albatrosses, and in cases where there are no nearby colonies and thus a lower chance of visitation by prospecting birds (Jones and Kress, 2012; VanderWerf et al., 2022). Translocation is usually combined with social attraction (Spatz et al., 2023). One advantage of social attraction is that it often is less expensive and less labor intensive than translocation. Once attraction systems are deployed, they can operate independently with little labor required.
Here, we assess the potential role of the California Channel Islands in conservation of Laysan and Black-footed Albatrosses. Specifically, we examine: 1) the current and former status of albatrosses in the Channel Islands; 2) the suitability of each of the Channel Islands for albatrosses; 3) the feasibility of attempting to establish albatross breeding colonies in the Channel Islands by social attraction and translocation; 4) regulatory, logistical, and biological factors that could affect implementation of such a project, and 5) the potential benefits and risks of undertaking albatross conservation actions in the Channel Islands using the most current and rigorous scientific framework (Karasov-Olson et al., 2021a; Karasov-Olson et al., 2021b).
Methods
Study site
The Channel Islands consist of eight islands located within the continental shelf of North America, offshore of southern California (Figure 1; Table 1). The islands often are divided into two groups, the four Northern Channel Islands, which were connected as one large island called Santarosae until roughly 10,000 years ago when sea level was lower (Erlandson et al., 2011), and the four Southern Channel Islands. The Channel Islands are recognized as a biodiversity hotspot and support many endemic taxa (Schoenherr et al., 2003).
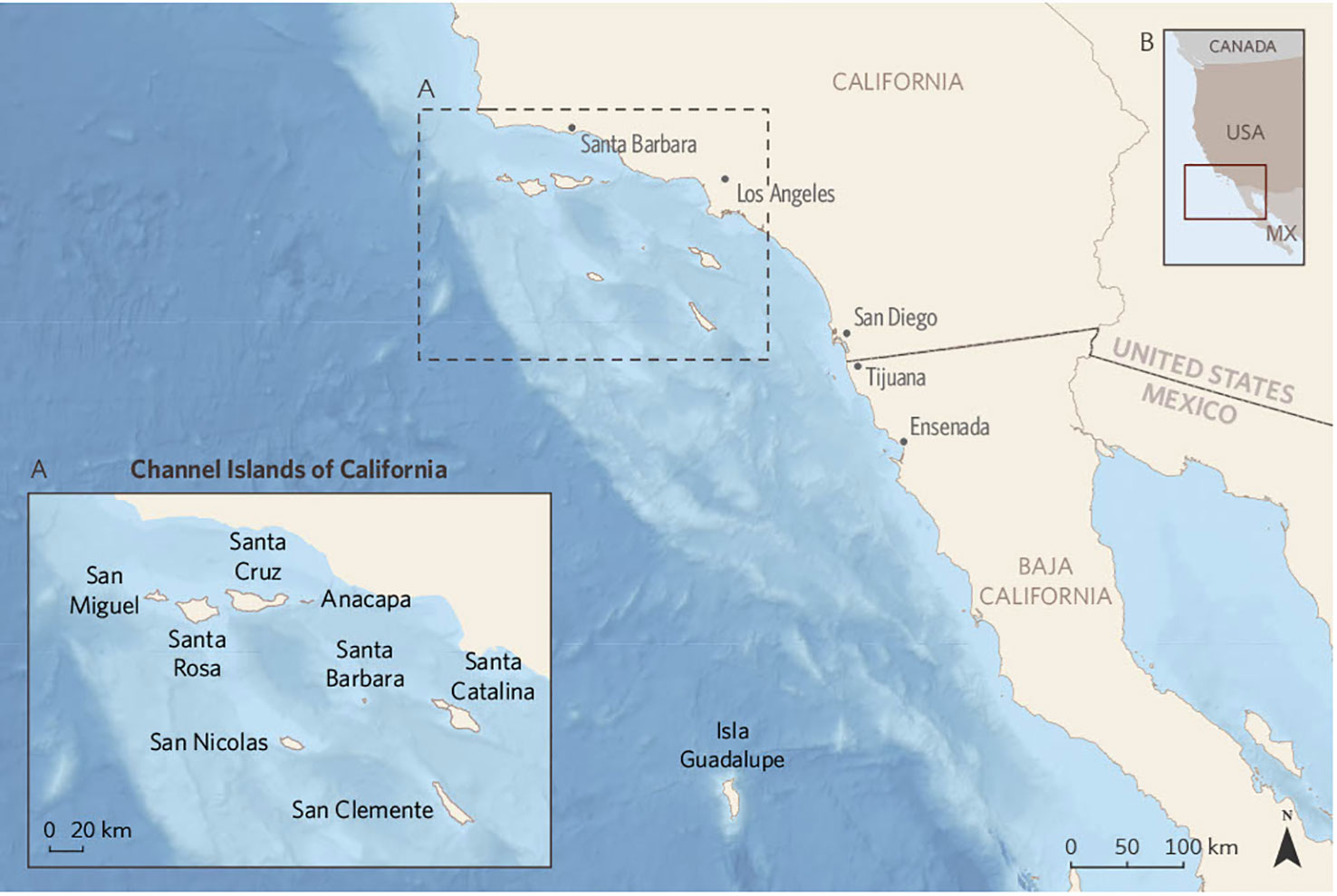
Figure 1 Map of the California Channel Islands. The continental shelf is indicated by the blue shading of the bathymetry. Inset B shows the location along the west coast of North America.
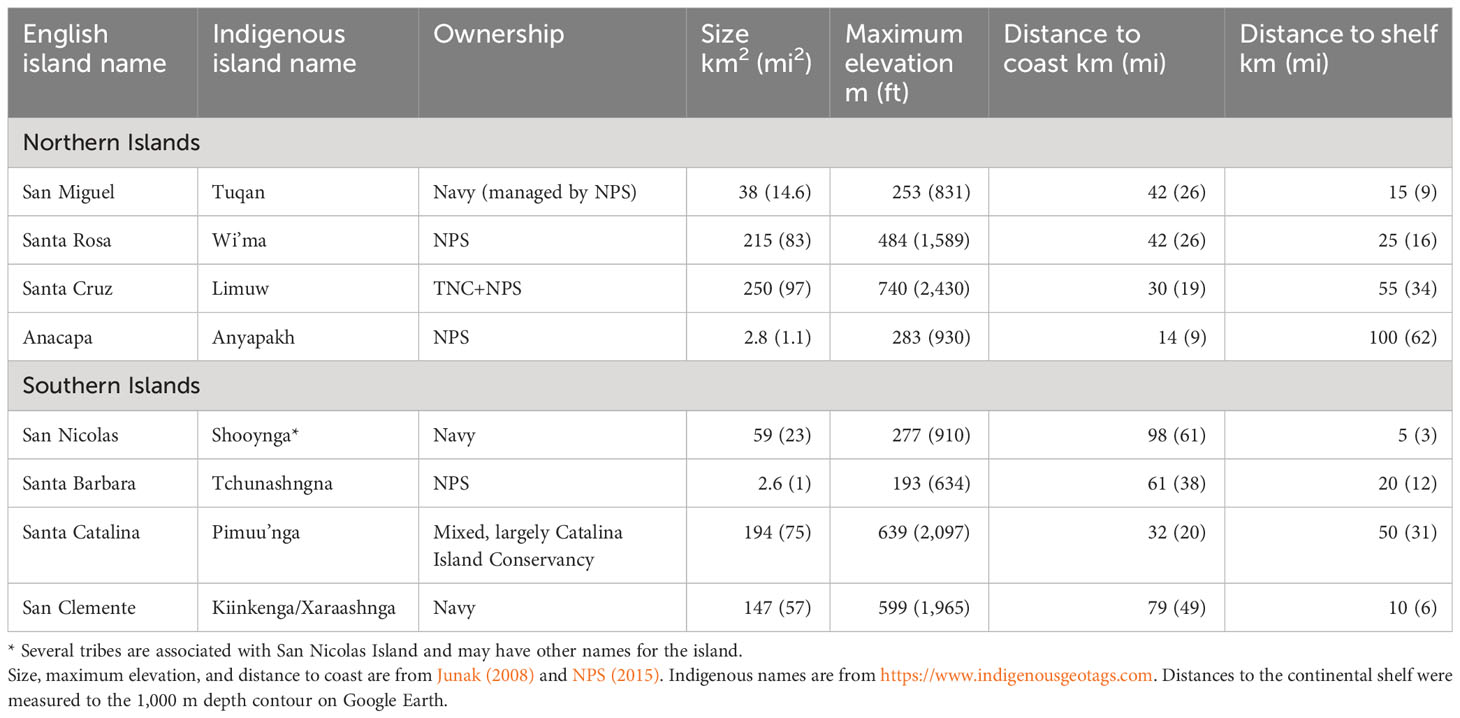
Table 1 Geographic summary of the California Channel Islands, listed from northwest to southeast.
The human history of the Channel Islands goes back at least 13,000 years, when native Americans are documented to have reached the islands (Glassow, 2010). Archaeological sites in the Channel Islands provide some of the earliest evidence of humans in North America (Erlandson et al., 2011). The early Channel Islands human inhabitants were proficient at traveling along the coast and between islands in large ocean-going canoes, and they inhabited all the Channel Islands and used them extensively (Glassow, 2010; Erlandson et al., 2011). The island ecosystems underwent many changes during this time, including the introduction of new species (e.g., foxes and mice), extinctions, and alteration of the nearshore marine invertebrate community (Rick et al., 2014).
Arrival of Europeans, including extraction of marine mammals and fishing, followed by an era of intensive ranching, had significant negative impacts on island ecosystems, including further introductions of invasive mammals (e.g., ungulates, feral cats, and rats), further extinctions and extirpations, and major habitat destruction. Over more recent decades, the islands have largely been transferred into some form of conservation management (Rick et al., 2014; NPS, 2015; McEachern et al., 2016). In 1976, all eight islands were designated a UNESCO Biosphere Reserve. The NPS owns and manages Santa Barbara, Anacapa, and Santa Rosa Islands, and the eastern 24% of Santa Cruz Island. The Nature Conservancy (TNC) owns and manages the remainder of Santa Cruz Island. The U.S. Navy owns San Miguel, San Nicolas, and San Clemente Islands, though San Miguel Island is managed by NPS. Santa Catalina Island is largely managed by the Catalina Island Conservancy. Channel Islands National Park extends 1 nautical mile offshore of NPS islands and Channel Islands National Marine Sanctuary extends 6 nautical miles from the islands’ shores. Significant ecological restoration has been accomplished on all the islands over the last several decades, including the eradication of most introduced ungulate populations (McEachern et al., 2016) and a subsequent recovery of many vegetation communities (e.g., Beltran et al., 2014).
Study species
Three species of albatrosses inhabit the North Pacific Ocean: Laysan, Black-footed, and Short-tailed Albatross (P. albatrus). These three albatross species are ecologically similar, but there are important differences in their population sizes, distributions, conservation status, and threats (Table 2). Although all three species were present in the Channel Islands waters prehistorically and potentially are suitable candidates for colony establishment in the Channel Islands, we focused on the Laysan Albatross and Black-footed Albatross because their breeding colonies in the Northwest Hawaiian Islands are at greater risk from climate change, and because of feasibility constraints of working with the Short-tailed Albatross. The Short-tailed Albatross currently has a much smaller population and a more limited distribution, but its population size and range are increasing because of a variety of conservation efforts in Japan, including translocation to a new island (Deguchi et al., 2012; Deguchi et al., 2017). All breeding colonies are in the Izu, Bonin, and Senkaku islands in the western Pacific. Translocation of the species outside that region would be very difficult for logistical and political reasons.
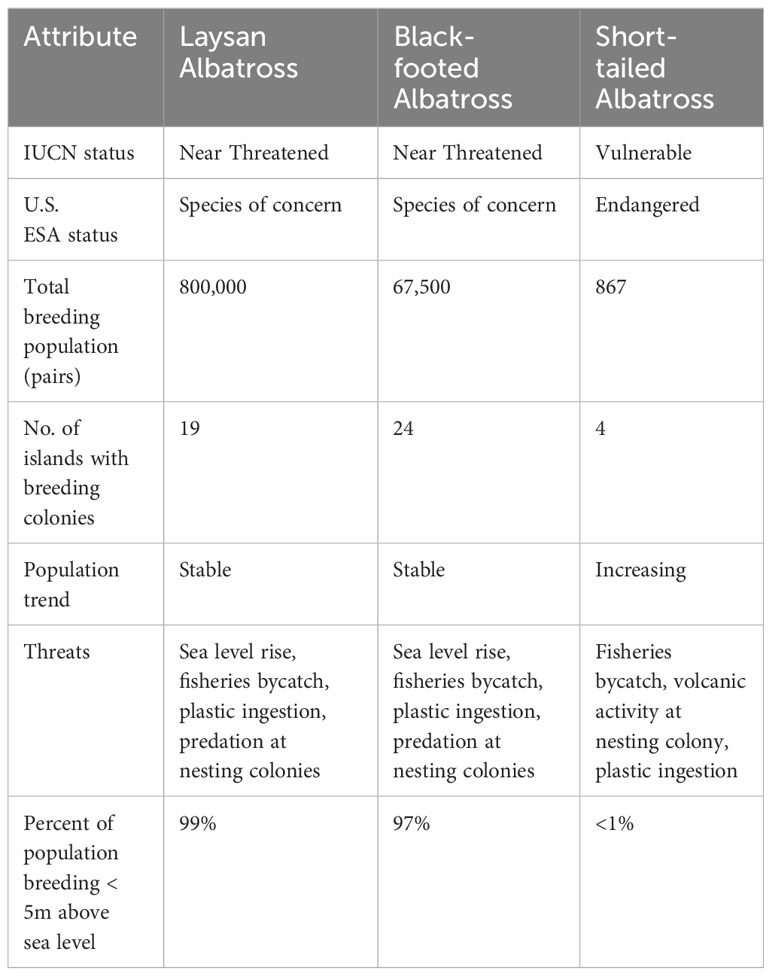
Table 2 Laysan, Black-footed, and Short-tailed Albatross population status, conservation status, and threats.
Laysan and Black-footed Albatrosses are relatively abundant, with breeding populations of 800,000 pairs and 67,500 pairs, respectively, and the population sizes of both species are thought to be stable currently (Table 2; Arata et al., 2009; BirdLife International, 2018). Both species are also widespread. The Laysan Albatross currently breeds on 19 islands, including 13 islands in Hawaii, four islands near Mexico (Guadalupe, Clarión, Alijos, and San Benedicto), and two islands in the western Pacific (Wake Island and Mukojima near Japan; Chiba et al., 2007; Hernández-Montoya et al., 2014; VanderWerf and Young, 2016; Henry et al., 2021). Over 99% of the global Laysan Albatross population nests in the Hawaiian Islands (VanderWerf and Young, 2016). The Black-footed Albatross currently breeds on 24 islands, including 11 islands in Hawaii and 13 islands in three island groups near Japan (Izu-Torishima, three of the Senkaku Islands, and nine of the Bonin Islands; Eda et al., 2008; VanderWerf and Young, 2011; VanderWerf and Young 2017). 97% of the population breeds in Hawaii, with about 1,500 pairs on the Japanese islands. The at-sea distributions of Laysan and Black-footed Albatrosses encompass pelagic areas across virtually the entire North Pacific Ocean, from the tropics to the Bering Sea and from Japan and Russia to the west coast of North America (Arata et al., 2009).
Although they currently are abundant, the populations of Laysan and Black-footed Albatrosses are expected to decline substantially over the next several decades due to inundation of the largest breeding colonies in the Northwestern Hawaiian Islands. Reynolds et al. (2015) modeled inundation and wave-driven flooding on Midway Atoll under various climate change scenarios and predicted that a 2.0-meter rise in sea level combined with wave-driven flooding events would result in loss of 61% and 60% of Laysan and Black-footed Albatross nests, respectively. Similarly, Baker et al. (2006) modeled habitat loss in the Northwestern Hawaiian Islands predicted to result from sea level rise by 2100, and found that expected inundation varied among islands, with 3% to 65% loss of habitat with a 48 cm rise in sea level, and 5% to 75% loss with an 88 cm rise in sea level, with some islands, such as French Frigate Shoals and Pearl and Hermes, expected to be completely lost. Some islands already have lost substantial area and some albatrosses already have been displaced (VanderWerf et al., 2019).
Island suitability
We assessed the suitability of each island using 16 criteria that encompassed biological, geographical, logistical, and regulatory and legal considerations that could influence the feasibility of establishing an albatross breeding colony. The criteria are listed and described below. More details about the criteria and their importance are provided by VanderWerf et al. (2022). We gave each island a plus (+) or minus (-) based on whether it was generally favorable regarding each factor, and then summed the number of pluses to obtain an overall score. This was intended to be a preliminary assessment, and the results of this evaluation alone will not determine which island, if any, might be selected. The criteria are intended to provide a simple list of factors that should be considered and the overall suitability of the islands based on each.
1. Permitting and environmental compliance. This project would require several permits and environmental review documents, and the requirements might depend on which island is chosen. Permits would be required from the USFWS Migratory Bird Program and the State of California’s Department of Fish and Wildlife (CDFW). If the island chosen supports plants or animals protected under the U.S. Endangered Species Act or the California Endangered Species Act, consultation with the USFWS and CDFW would be required. An Environmental Assessment (EA) or Environmental Impact Statement (EIS) might be required under the National Environmental Policy Act (NEPA), unless the action is covered under another document that already has completed environmental compliance. Review and approval of the California Coastal Commission also may be required, particularly if plans involve ground disturbance and construction within the coastal zone (e.g., installation of predator fencing). Consultation with Native American tribes also would be appropriate and advisable. On NPS islands, activities would need to be consistent with the Wilderness Act.
2. Compatibility with existing land uses and plans. This is a broad category intended to encompass compatibility with a variety of other activities and uses on the islands. Albatrosses are sometimes attracted to large, open areas like runways, of which there are several in the Channel Islands. In Hawai’i, Laysan Albatrosses are removed and hazed from airfields at the Pacific Missile Range Facility on Kaua’i (Anders et al., 2009) and at Marine Corps Base Hawai’i and U.S. Army Dillingham Airfield on O’ahu. Other potential compatibility issues include ability to continue important invasive plant removal and native plant restoration, and the compatibility of activities and associated infrastructure in wilderness areas. Compatibility with public use also should be considered on islands with public access. The presence of albatrosses could be an additional attraction to visitors, but proximity of visitors to albatrosses might need to be managed.
3. Compatibility with other native biota. Albatrosses are large birds, and though they generally are not aggressive toward other species, they potentially could have negative impacts on plants and other animals. Albatrosses nest on the surface and do not dig burrows, but they gather soil and plant material around them to form a nest cup. They require open ground for taking off and landing, and their repeated foot traffic could inhibit plant growth in a small area. On islands with large albatross colonies the density of nests can be high, up to 1 nest per square meter. If albatrosses increase in number, they could compete for space with other surface nesting birds such as Western Gulls (Larus occidentalis) and California Brown Pelicans (Pelecanus occidentalis californicus), and could affect endangered plants, such as the Santa Barbara Island live-forever (Dudleya traskiae).
4. Assisted colonization policy of landowner. Laysan and Black-footed Albatrosses commonly forage in waters off California, and in this sense they are native to the region, but there are no records of them nesting in the Channel Islands. There is thus some ambiguity as to whether attempting to establish a nesting colony of either species in the Channel Islands would be an assisted colonization, because the action being considered would not introduce an entirely new species, but instead facilitate use of the area for an additional life history stage or purpose (breeding instead of just foraging). Assisted colonization is consistent with USFWS policy, but NPS policy currently does not allow introduction of species not known to have inhabited an area previously, though this policy may change as awareness grows about the impacts of climate change and the importance of assisted colonization (Karasov-Olson et al., 2021a; Karasov-Olson et al., 2021b). Channel Islands National Park is facing similar questions in at least one other bird conservation management challenge, that of the Island Scrub-Jay (Aphelocoma insularis; Morrison, 2014).
5. Logistics. The ability to transport people, birds, and supplies safely to the island is crucial to successfully implementing this project. For translocation, birds or their eggs would need to be transported just once at the beginning of the breeding season, but regular access would be needed to support project staff and bring more supplies to the island. Transport could be by boat or aircraft; the most important determinants would be safety and reliability. If access might be affected by weather and some trips could be cancelled, this might be acceptable if there is sufficient infrastructure on the island to store extra supplies.
6. Island infrastructure. There must be sufficient infrastructure on the island to support the project, or the potential to build it. For translocation, this includes shelter, cooking, and bathroom facilities for 2-3 people to care for the chicks for five months, ability to keep bird food cold/frozen, and water for cooking and cleaning. It is essential to sterilize all equipment used to feed birds every day, and this requires some fresh water.
7. Readiness or time to implementation. This criterion represents the time and effort required to complete environmental review, obtain all necessary permits, and prepare the island for project implementation. This could include building or improving infrastructure, installing a predator exclusion fence, and conducting public outreach, the needs for which could vary among islands.
8. Capacity of landowner or manager. Establishing an albatross colony would take many years and require a long-term commitment, not only in the implementation phase but also in the long-term management of the colony. During and after project implementation it would be important to monitor albatrosses and manage any problems that might occur, such as presence of invasive species, impacts to other biological resources, and maintenance of a predator exclusion fence and social attraction equipment. This monitoring and management would require some staff time and resources from the landowner.
9. Island size/Suitable habitat area. A larger island would be preferable in general because it could support a larger albatross population, but habitat suitability for albatross is also affected by terrain and wind exposure. Laysan and Black-footed Albatrosses prefer flat ground for nesting, consistent winds to facilitate take off, and the presence of some soil to build a nest. They nest on slopes and rocky ledges in a few locations, but those habitat types are not preferred. On islands where it would be necessary to build a predator exclusion fence, the amount of suitable habitat would be the size of the fence, not the size of the entire island. Albatrosses do not require much space on land, just enough to build a nest out of reach of their neighbors, so many albatrosses can nest in a relatively small space. Midway Atoll is just 1,100 acres in size but supports over 600,000 pairs of breeding albatross (Arata et al., 2009).
10. Distance to continental shelf. Albatrosses forage primarily in deep water and areas of upwelling over the continental slope. Islands closer to the continental shelf would require a shorter commute for breeding adults and make it easier for fledglings to reach areas where they are more likely to find food. None of the distances involved are that long and albatrosses can cover them easily, but all else being equal, an island closer to the continental shelf would be preferable. For social attraction, proximity to the shelf is more important because albatrosses are more likely to naturally encounter such islands while foraging. Social attraction on islands far from the shelf is unlikely to succeed because albatrosses will rarely, if ever, visit such islands. The two islands visited by Laysan Albatrosses most often, San Nicolas and San Clemente (Collins et al., in press), are also the two islands closest to the continental shelf.
11. Distance to continental coast. Islands closer to the coast are more likely to be reached by predatory birds such as Bald Eagles (Haliaeetus leucocephalus) and Common Ravens (Corvus corax), and these species may prey on albatross eggs, chicks, and adults (see section below on predatory birds). On islands closer to the coast there also is greater potential for accidental introduction of ground predators as stowaways on private boats.
12. Presence of ground predators. Albatrosses and other Procellariform seabirds are naïve to ground predators and generally cannot coexist with them; this is the reason albatrosses nest primarily on predator-free islands (Spatz et al., 2014; Dias et al., 2019). The Island Fox (Urocyon littoralis) is considered endemic to the Channel Islands and currently occurs on six of the eight islands; it is absent from Anacapa and Santa Barbara. The Island Fox likely would be a threat to albatross eggs and chicks and possibly to breeding adults that did not flee from their nest. Non-native feral cats (Felis catus) occur on Santa Catalina Island and would be a threat to adult albatross. Island Spotted Skunks (Spilogale gracilis amphialus) occur on Santa Cruz and Santa Rosa islands, and non-native rats (Rattus spp.) occur on San Miguel, Santa Catalina, San Nicolas, and San Clemente islands, and could prey on eggs and chicks.If albatrosses were translocated to an island with Island Foxes, feral cats, or rats, it would be necessary to build a predator exclusion fence to protect albatrosses from predation. Such fences are generally effective if well sited and built correctly, but there could be occasional breaches of the fence caused by erosion or rockfall (Young et al., 2013; Young et al., 2018), and foxes might be able to dig under a fence depending on the substrate and their motivation.Native deer mice (Peromyscus maniculatus) in the Channel Islands are known to eat seabird eggs, but only eggs of smaller species that nest underground in burrows or rock crevices, such as Scripps’s Murrelet (Synthliboramphus scrippsi), and only when adult birds are absent from the nest and eggs are unattended (Murray et al., 1983; Millus et al., 2007). Predation by deer mice also has observed on Rhinoceros Auklet (Cerorhinca monocerata) eggs, which weigh 79 grams, on Triangle Island, Canada, primarily in years of low food abundance when the adult auklets spend more time foraging and leave eggs unattended (Blight et al., 1999). The risk of predation by deer mice on albatross eggs is negligible because albatross rarely leave eggs unattended and their eggs are much larger (average weight 278 grams in Laysan Albatross and 304 grams in Black-footed Albatross; Fisher, 1969). Invasive house mice (Mus musculus) are known to have injured and killed juvenile and adult albatrosses on Gough Island, Marion Island, and Midway Atoll (Jones and Ryan, 2010; Duhr et al., 2019), but in general mice are much less of a threat than larger predators, and deer mice have not been documented to prey on larger birds in the manner of house mice.Snakes are also a potential predator on albatross eggs and chicks, and several of the Channel Islands support one or more native snake species. On Clarion Island, Mexico, endemic Clarion Racers (Masticophis anthonyi) have been documented to kill Laysan Albatross chicks, even though they are too large for the snakes to swallow (Wanless et al., 2009, Daniel Portillo, GECI unpubl. data). In the Channel Islands, the Santa Cruz Gopher Snake (Pituophis catenifer pumilus) occurs on Santa Cruz and Santa Rosa, the Western Yellow-bellied Racer (Coluber constrictor) occurs on Santa Cruz, and the San Diego Nightsnake (Hypsiglena ochrorhyncha klauberi) occurs on Santa Cruz. The nightsnake is small and would not be a threat to albatrosses, but the gopher snake and the racer can be relatively large (up to 91 cm and 190 cm, respectively) and both are known to eat bird eggs and nestlings. The presence of these snakes makes Santa Cruz and Santa Rosa less suitable for albatross. Santa Catalina has several other native snake species.
13. Presence of predatory birds. Large predatory birds, particularly eagles, have been known to eat seabird eggs, chicks, and adults (Hipfner et al., 2012). In the Channel Islands, the bones of an adult Black-footed Albatross were found under an historical Bald Eagle nest on San Nicolas (Collins et al., in press). Following a successful conservation reintroduction program, Bald Eagles are now a common year-round resident in the Channel Islands, with 20 nesting pairs and over 60 individuals present in 2021 (NPS unpubl.). Newsome et al. (2010) found that seabirds were an important dietary component of Bald Eagles in the Channel Islands prehistorically, but their diet switched to non-native ungulates, primarily sheep, from 1850-1950 when seabirds declined and ranching was prevalent, and speculated that as eagle numbers recover, they could put increasing predation pressure on recovering seabird populations now that non-native ungulates have been removed. Newsome et al. (2015) found that recent Bald Eagle diets consisted of 40-45% seabirds in the Northern Channel Islands and 25-30% on Santa Catalina. In Hawai’i and other Pacific islands, adult Laysan, Black-footed, and Short-tailed albatrosses are known to have been preyed on by White-tailed Eagles (H. albicilla) and Steller’s Sea-Eagles (H. pelagicus; Balazs and Ralph, 1979; Zaun, 2009; Pyle and Pyle, 2017).Peregrine Falcons and a variety of owl species occur in the Channel Islands, but none of them are large enough to pose a predation risk to adult albatross. Peregrines and Barn Owls (Tyto alba) are present in areas of the Hawaiian Islands where Laysan Albatrosses nest, but have never been documented to prey on albatross eggs, chicks, or adults. Common Ravens (Corvus corax) on Isla Clarión off Mexico have learned to cooperatively prey on albatross eggs and chicks (Daniel Portillo, Grupo de Ecología y Conservación de Islas, unpubl. data). Ravens occur on most of the Channel Islands and could be a threat. On Guadalupe Island, Mexico, Western Gulls have preyed on a few albatross eggs, but only when the eggs are unattended, and have shown no interest in albatross chicks.
14. Parasite or pathogen transmission. Albatrosses moved or attracted to the Channel Islands could carry parasites or pathogens that could spread to other bird species on the island. In previous albatross translocations in Hawai’i and Mexico, chicks were treated with an external insecticide designed specifically for birds and an internal antibiotic to kill parasites, and this would greatly reduce the risk (VanderWerf et al., 2019). If eggs were moved, the risk would be lower because the eggshell is a barrier to parasites and most pathogens. This potential threat can be largely mitigated but should be considered.
15. Invasive Alien Species Introduction. The activities and transport of equipment associated with social attraction and translocation could result in the accidental introduction of invasive alien species. Precautions should be taken to avoid this, including thorough cleaning and inspection of all materials and dedicated clothing and equipment that would be used only on the specified island. The potential risk might be the same for all islands, but the consequences might be more severe on certain islands. There is an active biosecurity collaborative effort underway among managers of the Channel Islands that can advise on this subject.
16. Cultural Considerations. Albatrosses were important in the culture of the Native Americans that inhabited the Channel Islands, as evidenced by the abundance of albatross bones in archaeological sites. Albatross were used as food and a source of materials to make tools and decorations. Restoring albatrosses to the Channel Islands could help to restore a cultural connection with the species that has been lost. Hunting of albatross would not be appropriate, but viewing of albatross and cultural practices involving use of albatross feathers and bones might be possible through a permitting process, as it is done in Hawai’i. Public viewing of nesting albatrosses could enhance the recreational value of the islands.
Results
Current status of albatrosses in the Channel Islands
Collins et al. (in press) reported 10 recent observations of Laysan Albatrosses in the Channel Islands, including six on San Nicolas, two on San Clemente, and one each on San Miguel and Anacapa. These reports were judged by Collins (in press) to be sufficiently close (within 1 mile or reported as “close to” or “near”) to shore to suggest the bird was attracted to the island. There have been no recent reports of Black-footed Albatrosses on any of the Channel Islands.
Quantitative analysis of observations at sea during research cruises showed that Laysan and Black-footed Albatrosses were regular visitors to California waters (Leirness et al., 2021). Laysan Albatrosses were uncommon off California and occurred primarily in deeper water beyond the continental shelf and were somewhat more numerous in winter. Black-footed Albatrosses were fairly common in the California Current and occurred year-round but were more numerous in spring and summer and least numerous in winter. Black-footed Albatrosses occurred closer to the coast more often in spring and summer and occurred primarily farther offshore in fall and winter. While they occurred closer to the coast more often than Laysan Albatross, few occurred in the shallower nearshore waters within the continental shelf and around the Channel Islands (Leirness et al., 2021).
The number of Laysan Albatross sightings off southern California has increased since the 1980s for two reasons: 1) the growing population of Laysan Albatrosses on Guadalupe Island and other islands off the west coast of Mexico (Hernández-Montoya et al., 2014; Henry et al., 2021), and 2) greater observer effort; there has been an increase in the numbers of research cruises and recreational birding trips to offshore waters west and southwest of the Channel Islands (Leirness et al., 2021). Direct evidence that some of the Laysan Albatrosses seen in waters off California are from the breeding colony on Guadalupe was provided by Hernández Montoya et al. (2019), who tracked breeding Laysan Albatrosses from Guadalupe using GPS devices. These tracks showed that birds from Guadalupe commonly foraged off California, and that they occasionally ventured into shallower waters inside the continental shelf and farther west into the Central Pacific.
Former status of albatrosses in the Channel Islands
Extensive archaeological investigations in the Channel Island have revealed that all three North Pacific albatross species were present in the region and commonly were harvested by Native Americans (Porcasi, 1999; Glassow, 2010; Erlandson et al., 2011. P. Collins (unpublished) compiled all known records of albatrosses from archaeological sites in North America, which showed that albatross remains have been recovered from 44 sites on all eight of the Channel Islands, with the largest concentrations on San Miguel, San Nicolas, and San Clemente (see VanderWerf et al., 2022 for more details). The Short-tailed Albatross was the most abundant of the three species, comprising 97% of all identified specimens and 90% of all identified individuals. The high abundance of albatrosses compared to other bird species suggests they were especially targeted, or more easily captured using the methods available at that time, or both (Porcasi, 1999), and thus suffered disproportionate mortality.
Porcasi (1999) speculated that albatrosses may have nested in the Channel Islands, but no evidence of breeding, such as eggshells or juvenal or medullary bones, has been found in the Channel Islands (P. Collins, pers. comm. Sep 2021). It is possible that one or more of the species did breed in the Channel Islands, but the breeding sites have not yet been found, are now below sea level, or have been destroyed due to the widespread erosion that followed the vegetation loss during the ranching era. Other breeding seabirds are known to have been extirpated in the Channel Islands prehistorically, including a puffin species that was endemic to the Channel Islands (Guthrie et al., 2002). Gaps in fossil and archaeological records on the Channel Islands have been noted in other bird taxa, which are a reminder that absence of evidence is not evidence of absence (Collins et al., 2018; Morrison et al., 2018). Vokhshoori et al. (2019) used isotopic analysis of tissue samples from modern Short-tailed Albatrosses and ancient samples from archeological sites in Japan and the Channel Islands and found that ancient Short-tailed Albatrosses spent more time foraging in the California Current than modern individuals, and that ancient remains from the Channel Islands were isotopically distinct from those in Japan, indicating the species had a more complex population structure in the past.
Island suitability
Based on the 16 criteria described above, Santa Barbara and San Nicolas offer the best opportunities for establishing albatross breeding colonies in the Channel Islands. The suitability of each island is summarized below and in Table 3.
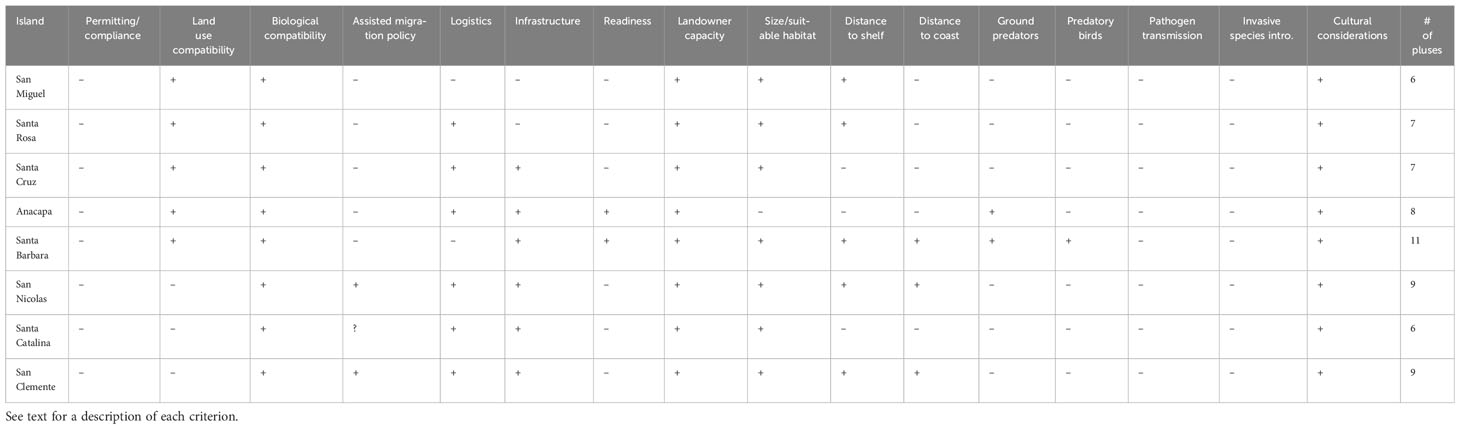
Table 3 Preliminary assessments of the eight California Channel Islands for albatross colony establishment using 16 selection criteria.
San Miguel is one of the less suitable islands for attempting to establish an albatross colony. It is relatively close to the coast, and there are ground predators, including Island Foxes and rats, and avian predators including Bald Eagles and Common Ravens. There is no regular boat service and there is no dock, but there are two airstrips so access by helicopter and fixed wing travel aircraft would be possible. There is little existing infrastructure apart from a NPS ranger station and a NOAA research station, which probably would not be available for albatross work. There is limited freshwater on the island, and it might be necessary to bring water which would be difficult and expensive.
Santa Rosa, like San Miguel, is one of the less suitable islands for an albatross colony because it is moderately close to the coast, and there are ground predators including Island Foxes and Island Spotted Skunks, and avian predators including Bald Eagles and Common Ravens. There is regular boat service to the island and a dock, and there is an airstrip so access by aircraft would be possible, so logistics would be easier than on San Miguel.
Santa Cruz has several desirable features but also several unfavorable aspects. It is moderately close to the continental shelf and distant from the coast, which is good for albatrosses. However, there are numerous ground predators, including Island Foxes, Island Spotted Skunks, gopher snakes and racers, and avian predators including Bald Eagles and Common Ravens. It is easily accessible by boat and has good existing infrastructure. Large portions of the island would be suitable for albatrosses but a predator fence would be needed, and albatross might be limited to the fenced area.
Anacapa has some desirable features, but overall it is one of the less suitable islands. It is the closest island to the coast and the most distant from the continental shelf. It is easily accessible by boat and has good existing infrastructure. There are no ground predators, but there is one pair of Bald Eagles on East Anacapa and a large colony of Western Gulls. Most of the island is steep and rocky and there is a limited amount of flat and gently sloping terrain that would be preferred for nesting by albatross.
Santa Barbara was the most suitable island for several biological and geographical reasons, but it also had some negative aspects. Santa Barbara is moderately close to the continental shelf and distant from the coast. There are no ground predators and currently no predatory birds large enough to be a threat to adult albatross, though there is a Western Gull breeding colony that could result in predation on unattended eggs. Although Santa Barbara is relatively small, no predator exclusion fence would be needed because no foxes are present and the entire island would be accessible to albatrosses; most of the island consists of suitable habitat for albatross, with gently sloping, open terrain. The island is sometimes accessible by boat, but the landing is currently challenging because the dock was damaged during a recent storm. NPS also regularly accesses the island by helicopter. Existing infrastructure at the NPS field station on Santa Barbara Island is good. There is no source of fresh water on the island, but there are large storage tanks and water is delivered by boat and pumped into the tanks. Public visitation is allowed and visitors are required to stay on existing trails, so public viewing of albatross would be possible. Habitat restoration efforts are underway, and there is some concern that actions of seabirds, potentially including albatrosses, could negatively affect native plants, particularly the endangered and endemic Santa Barbara Island live-forever (see Ecological Risk Assessment below).
San Nicolas is suitable for albatrosses in most respects, but there could be conflicts with the military uses of the island. It is the closest island to the continental shelf and the most distant from the coast, so it has the best geographic location of any of the Channel Islands for albatross. Laysan Albatrosses already have visited San Nicolas several times and may already prefer this island, and social attraction therefore has the best chance of succeeding there. It is easily accessible by airplane and has extensive existing infrastructure suitable for a translocation project. Much of the island consists of gently sloping terrain that would be suitable for albatrosses, but Island Foxes are present, so a predator fence would be needed, and albatross might be limited to the fenced area. Foxes would be a threat primarily to eggs and chicks, not adults. It might be possible to begin social attraction at one or more sites, and to build a fence around the site most visited by albatrosses once they begin to show signs of breeding. The flat terrain of the airfield might be attractive to albatrosses, where they could become a nuisance, but the airfield is located on the leeward side of the island which could reduce its attractiveness to albatrosses. On the other hand, the presence of albatrosses that have been intentionally and strategically established elsewhere on the island could reduce the chance that they visit the airfield.
Santa Catalina is not suitable because of the large human population, presence of several ground predators, particularly feral cats, rats and several snakes, and a variety of potentially conflicting land uses.
San Clemente is similar in most respects to San Nicolas but is less compatible in terms of existing land use because of more intensive military training over a larger portion of the island, and the presence of feral cats is a strong negative factor.
Colony establishment method
Social attraction would be relatively easy and inexpensive to implement on any of the Channel Islands, but the likelihood of establishing a colony through social attraction currently is relatively low on most islands because few albatrosses visit the shallow water inside the continental shelf. Decoys and sound playback systems are only effective if the birds come close enough to see and hear them; they cannot attract birds from far away. The chance of successful social attraction is highest for Laysan Albatross on San Nicolas, where they already have been observed on several occasions. Translocation is more likely to be needed to establish a breeding colony of Black-footed Albatrosses because they have never been observed on any of the Channel Islands, though they do tend to visit shallow water inside the continental shelf more often than Laysan Albatrosses.
The potential for successful social attraction is likely to increase over time. The number of Laysan Albatrosses in California waters is increasing because their population on Guadalupe Island is growing. The numbers of both species in California waters also are likely to increase as more individuals are displaced from colonies in Hawai’i that become inundated.
Ecological risk assessment
Based on an analysis using the framework provided by Karasov-Olson et al. (2021a), the risks associated with attempting to establish albatross breeding colonies in the Channel Islands were generally low. The only high or very high risk identified is to the target species (albatrosses) if no action is taken (Table 4; Supplementary Table 1). Four potential risks were rated as moderate, which were to the ecosystem, other native species, and land uses in the Channel Islands, each of which is described below.

Table 4 Visual summary of potential ecological risks of managed relocation of albatrosses to the Channel Islands.
There is some risk that translocated albatrosses could carry a parasite or pathogen to the Channel Islands that is not already present. This risk has been considered and mitigated to a large degree in previous albatross translocations in Hawai’i and Mexico by treating chicks with internal and external antibiotics that are safe, effective, and commonly used for veterinary purposes (VanderWerf et al., 2019). If these techniques were employed in the Channel Islands the risk would be considered low. Since albatrosses have already visited some islands and likely will continue to do so, this also is a naturally occurring risk.
Albatrosses could move from the island on which they were moved or attracted to different islands or locations where they are not wanted, and this can be regarded as a form of invasion beyond the intended recipient ecosystem. However, Laysan Albatrosses already have visited some of the Channel Islands, and it is not clear if social attraction or translocation would increase or decrease the chance of albatrosses visiting non-target islands or locations. The translocated individuals could visit other locations, but their presence at the restoration site could reduce the chance they visit and settle in other locations.
Albatrosses are sometimes attracted to airfields because of the flat terrain and wind conditions, and there is some risk that albatrosses could visit and attempt to nest on or near airfields in the Channel Islands, where they could pose a collision hazard with aircraft. On San Nicolas, the airfield is located on the leeward side of the island, which might make it less attractive to albatross. Albatrosses also could visit or attempt to nest near other infrastructure such as military training facilities and NPS maintenance and visitor facilities, which could require management of the albatrosses or of the activities occurring at the facilities.
Several iconic and culturally important species occur in the Channel Islands that have been the subject of much conservation work, including the Island Fox, Bald Eagle, and Santa Barbara Live-forever (Coonan et al., 2010; Newsome et al., 2010; Newsome et al., 2015). If albatrosses were attracted or translocated to an island that supported one or more of those species, it could create a potential management conflict. On islands with an Island Fox population, it would be necessary to build a predator exclusion fence to protect albatrosses from foxes. This would render a portion of the island inaccessible to foxes. If foxes were to dig under the fence or there was a fence break caused by erosion or rockfall, or if albatrosses settled outside the fence, it could create a conflict between the welfare of albatrosses and foxes. Similarly, if albatrosses were moved to an island that supported Bald Eagles, it is possible eagles would depredate them. On Santa Barbara, the actions of seabirds, including albatrosses, could damage plants and be counterproductive to restoration of the live-forever and other plant species.
Several other potential risks are low but warrant mentioning. There is increasing interest in offshore wind energy development in southern California, and wind turbines can have negatives effects on seabirds, including collisions, reduced habitat quality, and altered foraging behavior and displacement (Croll et al., 2022). Albatrosses are at less risk of collision with wind turbines than some seabirds because they typically fly low over the water, below the height of the turbine blades, and several methods are available to mitigate wind-energy impacts on seabirds (Croll et al., 2022). Moreover, Laysan and Black-footed Albatrosses are already present year-round in the offshore waters where wind energy projects may occur; establishment of a breeding colony in the Channel Islands would not substantially alter their abundance in this area.
Fishery bycatch and entanglement is a serious threat to albatrosses, and there are several fisheries in California waters that are known to have taken albatrosses (Jannot et al., 2021). There are existing regulations requiring vessels of various sizes to employ bycatch mitigation measures specifically for albatrosses, including streamer lines and night-setting (Jannot et al., 2021). As with wind energy, establishment of an albatross breeding colony in the Channel Islands would not substantially alter their abundance in this area and would be unlikely to result in any fishery management measures that are not already in place.
Discussion
Establishing breeding colonies of Laysan and Black-footed Albatrosses in the Channel Islands is feasible using existing methods of social attraction and translocation. These techniques have been applied with many seabird species worldwide and with these albatross species in particular (VanderWerf et al., 2019; VanderWerf et al., 2022; Spatz et al., 2023). Social attraction would be simpler and less expensive but probably would have a lower chance of success in the near future because few albatrosses visit the shallow waters inside the continental shelf where the Channel Islands are located. Translocation would be more labor intensive and expensive and would require more complex logistics and infrastructure but is more likely to achieve success sooner.
Any attempt to create an albatross colony in the Channel Islands would need to be considered a long-term project; establishing a colony using either method would require many years, and an incipient colony would grow slowly over multiple decades consistent with their life history. Laysan and Black-footed Albatrosses do not begin breeding until they are 7-9 years old (Arata et al., 2009; VanderWerf and Young, 2016), so it would be at least that long before any translocated birds began breeding. If translocation is attempted, enough birds would need to be moved to establish a viable colony; 100 birds has been a target in previous albatross translocations (VanderWerf et al., 2019). It is difficult to move and care for more than about 25 albatross chicks at a time, so it probably would be necessary to conduct translocations for four years. Natal philopatry in albatrosses is high, and social attraction is most likely to attract young birds that have not yet chosen where to breed. The number of albatrosses that might visit would be small at first, and albatrosses often take years to select a mate. This also would be a complex project from regulatory and logistical perspectives that could require many years to plan, engage stakeholders, conduct environmental review and compliance, and prepare the sites for release or attraction. Should this endeavor be treated by State and Federal authorities as a conservation introduction outside of the species’ indigenous range, additional time would be needed for planning in order to align with agency policy than if it were deemed a translocation to augment an existing population.
All three North Pacific albatross species are visiting California waters more often today than just a few years ago. Laysan Albatrosses are visiting California waters more often, and occasionally landing on the Channel Islands, in part because their numbers are growing in newly established colonies in Mexico (Hernández-Montoya et al., 2014; Henry et al., 2021). Black-footed Albatrosses are increasingly visiting California and the main Hawaiian Islands because of lost breeding habitat on low-lying islands in the Northwestern Hawaiian Islands. Short-tailed Albatrosses are again visiting the Channel Islands more frequently, reclaiming some of their previous foraging range, as the population recovers from near extinction caused by hunting by humans on the nesting islands in Japan (VanderWerf et al., 2022).
The increase in the numbers of albatrosses in the California Current can be expected to continue, and it is possible one or more species will naturally colonize the Channel Islands. If they do, there is a good chance they would choose an unsuitable area where they would be vulnerable to predation or where they would conflict with existing land uses. Facilitating the establishment of colonies in suitable locations would help to mitigate impacts of human activities on existing albatross colonies and would help to avoid potential conflicts. Social attraction and translocation both offer opportunities to influence where albatrosses settle in the Channel Islands.
The rate at which albatross colonies in the Northwestern Hawaiian Islands become inundated is difficult to predict, but some effects are already occurring and are virtually certain to worsen in the next several decades (Baker et al., 2006; Reynolds et al., 2015; Reynolds et al., 2017; VanderWerf et al., 2019; VanderWerf et al., 2022). The time that would elapse before any Channel Islands colony could be created means that any benefits to the species would not begin to occur until well after population declines have already occurred in the Northwest Hawaiian Island colonies. In planning conservation translocations, it is critical to take into account the trajectories of both the anticipated decline in the source population and the estimated growth rates of the desired new population, in order to ensure that extinction risks overall are reduced (McDonald-Madden et al., 2011). Given the considerable biological constraints in building a demographically and genetically robust colony, as well as the difficulties of detecting long-term trends in such long-lived species (Bakker et al., 2018), we recommend that managers take a proactive and precautionary approach to bolstering the resiliency of these species.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
EV: Conceptualization, Investigation, Methodology, Writing – original draft. NH: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Writing – review & editing. SM: Conceptualization, Funding acquisition, Investigation, Writing – review & editing. CK: Investigation, Writing – review & editing. AW: Conceptualization, Investigation, Methodology, Writing – review & editing. LY: Conceptualization, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This project was funded by The Nature Conservancy.
Acknowledgments
This report was funded by a contract to Pacific Rim Conservation from The Nature Conservancy. For information and discussion about the Channel Islands and logistical support during site visits, we thank Channel Islands National Park, especially David Mazurkiewicz, Annie Little, and Ken Convery, and the U. S. Navy, especially Bill Hoyer. We thank Paul Collins, curator emeritus of the Santa Barbara Natural History Museum, for generously sharing information about the Channel Islands, allowing us to use information about recent albatross observations from his upcoming book on Birds of the Channel Islands, and providing unpublished information about archaeological evidence of albatrosses in the Channel Islands. We also thank Lara Brenner of TNC for helping to advance the idea of managed albatross relocation in the Channel Islands and Irina Koroleva of TNC for making the map in Figure 1.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2023.1279373/full#supplementary-material
References
Anders A. D., Burger J. R., Pepi V. E. (2009). Barking Sands Pacific Missile Range Facility, proposed Laysan albatross management plan to reduce bird-aircraft strike hazard potential. NAVFAC Pacific 16.
Arata J. A., Sievert P. R., Naughton M. B. (2009). Status assessment of Laysan and black-footed albatrosses, North Pacific Ocean 1923-2005. U.S. Geological Survey Scientific Investigations Report 2009-5131 (U.S. Geological Survey Scientific). 80.
Baker J. D., Littnan C. L., Johnston D. W. (2006). Potential effects of sea level rise on the terrestrial habitats of endangered and endemic megafauna in the Northwestern Hawaiian Islands. Endangered Species Res. 2, 21–30. doi: 10.3354/esr002021
Bakker V. J., Finkelstein M. E., Doak D. F., VanderWerf E. A., Young L. C., Arata J. A., et al. (2018). The albatross of assessing and managing risk for long-lived pelagic seabirds. Biol. Conserv. 217, 83–95. doi: 10.1016/j.biocon.2017.08.022
Balazs G. H., Ralph C. J. (1979). A Steller’s Sea Eagle at Kure and Midway: First Hawaii record. ‘Elepaio 39, 117.
Beltran R. S., Kreidler N., Van Vuren D. H., Morrison S. A., Zavaleta E. S., Newton K., et al. (2014). Passive recovery of vegetation after herbivore eradication on Santa Cruz Island, California. Restor. Ecol. 22, 790–797. doi: 10.1111/rec.12144
BirdLife International. (2018). The IUCN red list of threatened species 2018. Available at: https://dx.doi.org/10.2305/IUCN.UK.2018-2.RLTS.T22698001A132618432.en.
Blight L. K., Ryder J. L., Bertram D. F. (1999). Predation on Rhinoceros Auklet eggs by a native population of Peromyscus. Condor 101, 871–876. doi: 10.2307/1370079
Buxton R. T., Jones C., Moller H., Towns D. R. (2014). Drivers of seabird population recovery on New Zealand islands after predator eradication. Conserv. Biol. 28, 333–344. doi: 10.1111/cobi.12228
Chiba H., Kawakami K., Suzuki H., Horikoshi K. (2007). The distribution of seabirds in the Bonin Islands, Southern Japan. Yamashina Institute Ornithology 39, 1–17. doi: 10.3312/jyio.39.1
Collins P. W., Guthrie D. A., Whistler E. L., Vellanoweth R. L., Erlandson J. M. (2018). Terminal Pleistocene–Holocene avifauna of San Miguel and Santa Rosa Islands: identifications of previously unidentified avian remains recovered from fossil sites and prehistoric cave deposits. Western North Am. Nat. 78, 370–403. doi: 10.3398/064.078.0311
Collins P. W., Jones H. L., Stahl J. T. (In Press). Birds of California’s Channel Islands (Santa Barbara, California: Natural History Museum of Santa Barbara).
Collins P. (unpublished). Short-tailed Albatross – Historical Literature. 115, Unpublished report of the Natural History Museum of Santa Barbara.
Coonan T. J., Schwemm C. A., Garcelon D. K. (2010). Decline and Recovery of the Island Fox: A Case Study for Population Recovery (Cambridge, England: Cambridge University Press).
Croll D. A., Ellis A. A., Adams J., Cook A. S., Garthe S., Goodale M. W., et al. (2022). Framework for assessing and mitigating the impacts of offshore wind energy development on marine birds. Biol. Conserv. 276, 109795.
Deguchi T., Jacobs J., Harada T., Perriman L., Watanabe Y., Sato. F. (2012). Translocation and hand-rearing techniques for establishing a colony of threatened albatross. Bird Conserv. Int. 22, 66–81. doi: 10.1017/S0959270911000438
Deguchi T., Sato F., Eda M., Izumi H., Suzuki H., Suryan R. M., et al. (2017). Translocation and hand-rearing result in shorttailed albatrosses returning to breed in the Ogasawara Islands 80 years after extirpation. Anim. Conserv. 20, 341e349.
Dias M. P., Martin R., Pearmain E. J., Burfield I. J., Small C., Phillips R. A., et al. (2019). Threats to seabirds: a global assessment. Biol. Conserv. 237, 525–537. doi: 10.1016/j.biocon.2019.06.033
Duhr M., Flint E. N., Hunter S. A., Taylor R. V., Flanders B., Howald G., et al. (2019). Control of house mice preying on adult albatrosses at Midway Atoll National Wildlife Refuge. Island Invasives: Scaling up to Meet the Challenge. Eds. Veitch C. R., Ciout M. N., Russell J. C., West C. J., 21–25.
Dunn P. O., Møller A. P. (2019). Effects of Climate Change on Birds, second edition (Oxford, UK: Oxford University Press).
Eda M., Kawakami K., Chiba H., Suzuki H., Horikoshi K., Koike H. (2008). Genetic characteristics of the Black-footed Albatross Diomedea nigripes on the Bonin Islands and their implications for the species' demographic history and population structure. Ornithological Sci. 7, 109–116. doi: 10.2326/1347-0558-7.2.109
Erlandson J. M., Rick T. C., Braje T. J., Casperson M., Culleton B., Fulfrost B., et al. (2011). PaleoIndian seafaring, maritime technologies, and coastal foraging on California’s Channel Islands. Science 331, 1181–1185. doi: 10.1126/science.1201477
Fisher H. I. (1969). ). Eggs and egg laying in the Laysan Albatross Diomedea immutabilis. Condor 71, 102–112. doi: 10.2307/1366071
Flint B., Kendall W., Breton A., Courtot K., Krause C., Naughton M., et al. (2011). Management of Black-footed Albatross in the face of climate change. U.S. Fish and Wildlife Service. 17, unpublished.
Freeman B. G., Freeman A. M. C. (2014). Rapid upslope shifts in New Guinean birds illustrate strong distributional responses of tropical montane species to global warming. Proc. Natl. Acad. Sci. 111, 4490–4494. doi: 10.1073/pnas.1318190111
Glassow M. A. (2010). Channel Islands National Park Archeological Overview and Assessment (Channel Islands National Park: Department of the Interior).
Gummer H. (2003). Chick translocation as a method of establishing new surface-nesting seabird colonies: a review (Wellington, New Zealand: New Zealand Department of Conservation).
Guthrie D. A., Thomas H. W., Kennedy G. L. (2002). “A new species of extinct Late Pleistocene puffin (Aves: Alcidae) from the southern California Channel Islands,” in Proceedings of the Fifth California Islands Symposium, Santa Barbara (CA): Santa Barbara Museum of Natural History, Vol. 2. 525–530.
Henry III R. W., Shaffer S. A., Antolos M., Félix-Lizárraga M., Foley D. G., Hazen E. L., et al. (2021). Successful long-distance breeding range expansion of a top marine predator. Front. Ecol. Evol. 9, 274. doi: 10.3389/fevo.2021.620103
Hernández-Montoya J. C., Luna-Mendoza L., Aguirre-Muñoz A., Méndez-Sánchez F., Félix-Lizárraga M., Barredo-Barberena J. M. (2014). Laysan Albatross on Guadalupe Island, México: current status and conservation actions. Monogr. Western North Am. Nat. 7, 543–555. doi: 10.3398/042.007.0101
Hernández Montoya J. C., Juárez-Rodríguez M., Méndez-Sánchez F., Aguirre-Muñoz A., Rojas-Mayoral E., Íñigo-Elias E., et al. (2019). Sexual dimorphism and foraging trips of the Laysan Albatross (Phoebastria immutabilis) on Guadalupe Island. Animals 9 (6), 364.
Hipfner M. J., Blight L. K., Lowe R. W., Wilhelm S. I., Robertson G. J., Barrett R. T., et al. (2012). Unintended consequences: how the recovery of sea eagle Haliaeetus spp. populations in the northern hemisphere is affecting seabirds. Mar. Ornithology 40, 39–52.
Hoegh-Guldberg O., Hughes L., McIntyre S., Lindenmayer D. B., Parmesan C., Possingham H. P., et al. (2008). Assisted colonization and rapid climate change. Science 321, 345–346. doi: 10.1126/science.1157897
Jacobs J., Deguchi T., Perriman L., Flint E., Gummer H., Uhart M. (2020). Guidelines for translocations of albatrosses and petrels. Available at: https://acap.aq/en/resources/acap-conservation-guidelines/2640-translocation-guidelines/file.
Jannot J. E., Wuest A., Good T. P., Somers K. A., Tuttle V. J., Richerson K. E., et al. (2021). Seabird Bycatch in U.S. West Coast Fisheries 2002–18 (U.S. Department of Commerce, NOAA Technical Memorandum NMFS-NWFSC-165).
Jones H. P., Kress S. W. (2012). A review of the world's active seabird restoration projects. J. Wildlife Manage. 76, 2–9. doi: 10.1002/jwmg.240
Jones M. G. W., Ryan P. G. (2010). Evidence of mouse attacks on albatross chicks on sub-Antarctic Marion Island. Antarctic Sci. 22, 39–42. doi: 10.1017/S0954102009990459
Junak S. (2008). A Flora of San Nicolas Island California (Santa Barbara, California: Santa Barbara Botanic Garden).
Karasov-Olson A., Schwartz M. W., Olden J. D., Skikne S., Hellmann J. J., Allen S., et al. (2021b). Ecological risk assessment of managed relocation as a climate change adaptation strategy. Natural Resource Report NPS/NRSS/CCRP/NRR—2021/2241 (Fort Collins, Colorado: National Park Service). doi: 10.36967/nrr-2284919
Karasov-Olson A., Schwartz M. W., Skikne S. A., Hellmann J. J., Olden J. D., Lawrence D. J., et al. (2021a). Co-development of a risk assessment strategy for managed relocation. Ecol. Solutions Evidence 2, e12092. doi: 10.1002/2688-8319.12092
Kostyack J., Lawler J. J., Goble D. D., Olden J. D., Scott J. M. (2011). Beyond reserves and corridors: policy solutions to facilitate the movement of plants and animals in a changing climate. BioScience 61, 713–719. doi: 10.1525/bio.2011.61.9.10
Lawler J. J., Olden J. D. (2011). Reframing the debate over assisted colonization. Front. Ecol. Environ. 9, 569–574. doi: 10.1890/100106
Leirness J. B., Adams J., Ballance L. T., Coyne M., Felis J. J., Joyce T., et al. (2021). Modeling at-sea density of marine birds to support renewable energy planning on the Pacific Outer Continental Shelf of the contiguous United States (Camarillo (CA: US Department of the Interior, Bureau of Ocean Energy Management), 385.
MacLean S. A., Beissinger S. R. (2017). Species’ traits as predictors of range shifts under contemporary climate change: A review and meta-analysis. Global Change Biol. 23, 4094–4105. doi: 10.1111/gcb.13736
Maier D. S., Simberloff D. (2016). Assisted migration in normative and scientific context. J. Agric. Environ. Ethics 29, 857–882. doi: 10.1007/s10806-016-9628-5
McDonald-Madden E., Runge M. C., Possingham H. P., Martin T. G. (2011). Optimal timing for managed relocation of species faced with climate change. Nat. Climate Change 1, 261–265. doi: 10.1038/nclimate1170
McEachern A. K., Atwater T., Collins P. W., Faulkner K., Richards D. (2016). “Managed island ecosystems,” in Ecosystems of California. Eds. In Mooney H. A., Zavaleta E. (Oakland, California: University of California Press), 755–778.
McLachlan J. S., Hellmann J. J., Schwartz M. W. (2007). A framework for debate of assisted migration in an era of climate change. Conserv. Biol. 21, 297–302. doi: 10.1111/j.1523-1739.2007.00676.x
Millus S. A., Stapp P., Martin P. (2007). Experimental control of a native predator may improve breeding success of a threatened seabird in the California Channel Islands. Biol. Conserv. 138, 484–492. doi: 10.1016/j.biocon.2007.05.017
Morrison S. A. (2014). A bird in our hand: weighing uncertainty about the past against uncertainty about the future in Channel Islands National Park. George Wright Forum 31, 77–93.
Morrison S. A., Sillett T. S., Funk C. W., Ghalambor C. K., Rick T. C. (2018). California Island Rediscovery: building an archive to improve conservation today and equip the historical ecologist of tomorrow. Western North Am. Nat. 78, 837–851. doi: 10.3398/064.078.0423
Murray K. G., Winnett-Murray K., Eppley Z. A., Hunt G. L., Schwartz D. B. (1983). Breeding biology of the Xantus’ Murrelet. Condor 85, 12–21. doi: 10.2307/1367880
National Park Service (2015). Channel Islands National Park final general management plan/wildeness study/Environmental Impact Statement (Ventura, California: National Park Service, U.S. Department of the Interior).
Newsome S. D., Collins P. W., Rick T. C., Guthrie D. A., Erlandson J. M., Fogel M. L. (2010). Pleistocene to historic shifts in bald eagle diets on the Channel Islands, California. Proc. Natl. Acad. Sci. 107, 9246–9251. doi: 10.1073/pnas.0913011107
Newsome S. D., Collins P. W., Sharpe P. (2015). Foraging ecology of a reintroduced population of breeding Bald Eagles on the Channel Islands, California, USA, inferred from prey remains and stable isotope analysis. Condor: Ornithological Appl. 117, 396–413. doi: 10.1650/CONDOR-14-213.1
Porcasi J. F. (1999). Prehistoric exploitation of albatross on the southern California Channel Islands. J. California Great Basin Anthropology 21 (1), 94–112.
Pyle R. L., Pyle P. (2017). “The Birds of the Hawaiian Islands: Occurrence, History, Distribution, and Status,” (Honolulu, HI, U.S.A: B.P. Bishop Museum). Available at: http://hbs.bishopmuseum.org/birds/rlp-monograph.
Reynolds M. H., Berkowitz P., Klavitter J. L., Courtot K. N. (2017). Lessons from the Tōhoku tsunami: A model for island avifauna conservation prioritization. Ecol. Evol. 7, 5873–5890. doi: 10.1002/ece3.3092
Reynolds M. H., Courtot K. N., Berkowitz P., Storlazzi C. D., Moore J., Flint E. (2015). Will the effects of sea-level rise create ecological traps for Pacific island seabirds? PloS One. doi: 10.1371/journal.pone.0136773
Ricciardi A., Simberloff D. (2009). Assisted colonization is not a viable conservation strategy. Trends Ecol. Evol. 24, 248–253. doi: 10.1016/j.tree.2008.12.006
Richardson D. M., Hellmann J. J., McLachlan J. S., Sax D. F., Schwartz M. W., Gonzalez P., et al. (2009). Multidimensional evaluation of managed relocation. Proc. Natl. Acad. Sci. 106, 9721–9724. doi: 10.1073/pnas.0902327106
Rick T. C., Sillett T. S., Ghalambor C. K., Hofman C. A., Ralls K., Anderson R. S., et al. (2014). Ecological change on California's channel islands from the pleistocene to the anthropocene. BioScience 64, 680–692. doi: 10.1093/biosci/biu094
Schoenherr A. A., Feldmeth C. R., Emerson M. J. (2003). Natural History of the Islands of California (Berkeley, CA: University of California Press).
Seddon P. J. (2010). From reintroduction to assisted colonization: moving along the conservation translocation spectrum. Restor. Ecol. 18, 796–802. doi: 10.1111/j.1526-100X.2010.00724.x
Spatz D. R., Newton K. M., Heinz R., Tershy B., Holmes N. D., Butchart S. H., et al. (2014). The biogeography of globally threatened seabirds and island conservation opportunities. Conserv. Biol. 28 (5), 1282–1290.
Spatz D. R., Young L. C., Holmes N. D., Jones H. P., VanderWerf E. A., Lyons D. E., et al. (2023). Tracking the global application of conservation translocation and social attraction to reverse seabird declines. Proc. Natl. Acad. Sci. 120, e2214574120. doi: 10.1073/pnas.2214574120
Thomas C. D., Cameron A., Green R. E., Bakkenes M., Beaumont L. J., Collingham Y. C., et al. (2004). Extinction risk from climate change. Nature 427, 145–148. doi: 10.1038/nature02121
USFWS [U.S. Fish and Wildlife Service]. (2005). Regional Seabird Conservation Plan, Pacific Region. U.S. Fish and Wildlife Service, Migratory Birds and Habitat Programs, Pacific Region (Portland, Oregon: U.S. Fish and Wildlife Service), 264.
VanderWerf E. A., Young L. C. (2017). A summary and gap analysis of seabird monitoring in the U.S. Tropical Pacific. Rep. prepared U.S. Fish Wildlife Service Region 1 Portland OR (Honolulu, HI) 59, 1.
VanderWerf E. A., Kress S., Bedolla-Guzmán Y., Spatz D., Taylor G., Gummer H. (2022). “Chapter 19 in conservation of marine birds,” in Restoration: Social attraction and translocation. Eds. Young L., VanderWerf E. (London, United Kingdom: Elsevier).
VanderWerf E. A., Young L. C. (2011). Estimating survival and life stage transitions in the Laysan Albatross using multi-state mark-recapture models. Auk 128, 726–736. doi: 10.1525/auk.2011.10285
VanderWerf E. A., Young L. C. (2016). Juvenile survival, recruitment, population size, and effects of avian poxvirus in Laysan Albatross (Phoebastria immutabilis) on Oahu, Hawaii. Condor 118, 804–814. doi: 10.1650/CONDOR-16-49.1
VanderWerf E. A., Young L. C., Kohley C. R., Dalton M. E., Fisher R., Fowlke L., et al. (2019). Establishing Laysan and black-footed albatross breeding colonies using translocation and social attraction. Global Ecol. Conserv. 19, e00667. doi: 10.1016/j.gecco.2019
Vokhshoori N. L., McCarthy M. D., Collins P. W., Etnier M. A., Rick T., Eda M., et al. (2019). Broader foraging range of ancient short-tailed albatross populations into California coastal waters based on bulk tissue and amino acid isotope analysis. Mar. Ecol. Prog. Ser. 610, 1–13. doi: 10.3354/meps12839
Wallingford P. D., Morelli T. L., Allen J. M., Beaury E. M., Blumenthal D. M., Bradley B. A., et al. (2020). Adjusting the lens of invasion biology to focus on the impacts of climate-driven range shifts. Nat. Climate Change 10, 398–405. doi: 10.1038/s41558-020-0768-2
Wanless R. M., Aguirre-Muñoz A., Angel A., Jacobsen J. K., Keitt B. S., McCann J. (2009). Birds of Clarion island, Revillagigedo Archipelago, Mexico. Wilson J. Ornithology 121, 745–751. doi: 10.1676/08-140.1
Young L. C., Behnke J. H., Vanderwerf E. A., Raine A. F., Mitchell C., Kohley C. R., et al. (2018). The Nihoku Ecosystem Restoration Project: A case study in predator exclusion fencing, ecosystem restoration, and seabird translocation. Pacific Cooperative Studies Unit Technical Report 198 (Honolulu, HI: University of Hawai’i at Mānoa, Department of Botany), 83.
Young L., Suryan R. M., Duffy D., Sydeman W. J. (2012). Climate change and seabirds of the California Current and Pacific Islands ecosystems: observed and potential impacts and management implication. Final Report to the U.S. Fish and Wildlife Service (Portland, Oregon: U.S. Fish and Wildlife Service).
Young L. C., VanderWerf E. A. (2016). The beginning of Black-footed Albatross colonization on O’ahu, Hawai`i. Elepaio 76, 1–4.
Young L. C., VanderWerf E. A., Lohr M. T., Miller C. J., Titmus A. J., Peters D., et al. (2013). Multi-species predator eradication within a pest-proof fence at Ka’ena Point, Hawai’i. Biol. Invasions 15, 2627–2638. doi: 10.1007/s10530-013-0479-y
Keywords: albatross, assisted colonization, California Channel Islands, climate change, managed relocation, social attraction, translocation
Citation: VanderWerf EA, Holmes ND, Morrison SA, Kohley CR, Wegmann A and Young LC (2024) Assisted colonization of albatrosses in the California Channel Islands: conservation basis and suitability assessment. Front. Conserv. Sci. 4:1279373. doi: 10.3389/fcosc.2023.1279373
Received: 17 August 2023; Accepted: 21 December 2023;
Published: 06 February 2024.
Edited by:
Doug P. Armstrong, Massey University, New ZealandReviewed by:
Nicola J. Van Wilgen, South African National Parks, South AfricaMarc Romano, United States Department of the Interior, United States
Melissa Price, University of Hawaii at Manoa, United States
Copyright © 2024 VanderWerf, Holmes, Morrison, Kohley, Wegmann and Young. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eric A. VanderWerf, ZXJpY0BwYWNpZmljcmltY29uc2VydmF0aW9uLm9yZw==