 Marzia Verduci
Marzia Verduci Laurie J. Goodrich
Laurie J. Goodrich Jean-Francois Therrien
Jean-Francois Therrien Ernesto Ruelas Inzunza
Ernesto Ruelas Inzunza- 1Acopian Center for Conservation Learning, Hawk Mountain Sanctuary, Orwigsburg, PA, United States
- 2Universidad Veracruzana, Instituto de Biotecnología y Ecología Aplicada, Xalapa, Mexico
Migratory raptors can fuel their journeys by feeding along the way. To maximize migration success, raptors are suspected to time their movements with the availability of their prey. Feeding rates can vary among species depending on migration distance (long vs. short distance), flight strategies (flapping vs. soaring), and within species according to individual timing (early vs. late migrants). We assessed the feeding rate of four eastern North American migratory raptors over a period of 35 years. Using visual observations collected during autumn migration in the Central Appalachians, we assessed the presence/absence of distended crops of migrants. We quantified the feeding rates of species with different migration ecologies (migration distance, flight strategies), compared within-season (timing) species, and assessed if feeding rates varied over the last 35 years. We found a higher feeding rate in short-distance than in long-distance migrants and in power-flapping vs soaring migrants. We detected a long-term significant increase in feeding rate for most species, more evident in early migratory individuals coupled with a decrease in the feeding rate of late migrants. Understanding how feeding rates vary according to migratory behavior and how the migration ecology of raptors is changing under a scenario of global environmental change is important for their conservation.
Introduction
Migration is a crucial aspect of the life history of many species of raptors. A successful journey relies critically on the interplay of various factors such as weather conditions, habitat, and food availability encountered along the way. To face the challenge of migration and the encountered conditions, raptors need to be in good body condition. Strategies that would promote successful journeys can affect the overall fitness of individuals and has consequences on the entire population dynamics (Bildstein, 2006).
Raptors can fuel their migratory movements in several ways: by building fat reserves prior to migration and, subsequently, burning fat en route, by feeding regularly during migration, and/or by soaring on orographic updrafts (Bildstein, 2006; Goodrich and Smith, 2008). Feeding en route is a common strategy used by several migratory raptor species (Rudebeck, 1950; Rudebeck, 1951; Dekker, 1980; Newton, 2006). Prey availability during migration is thus a key component of a successful migration and in turn, for species conservation (Kerlinger, 1989). Raptors that feed on migrating species are suspected to time their movements to coincide with those of their prey (Kjellén et al., 2001; Bildstein, 2006). Some migrating raptors rely on aerial insects during migration, such as dragonflies (Nicoletti, 1996). Indeed, insects can play an important role in the ecology of migratory raptors, and species that depend on them may travel long distances to find them (Newton, 1998) or feed opportunistically during migration (Nicoletti, 1996; Yosef, 1996).
Birds and especially raptors are excellent indicators of environmental health and ecosystem integrity. Recent evidence of population losses across much of the North American avifauna showed an alarming loss approaching 3 billion birds (Rosenberg et al., 2019). This might have consequences on the prey availability and on the success of a migration journey for migrating raptors (Moore and Kerlinger, 1987). Studying the strategies associated with successful migratory journeys is thus of utmost importance for species conservation.
In raptors, the effect of weather conditions and habitat availability during migration has been studied more broadly than feeding behavior and food availability (Niles et al., 1996; Sullivan et al., 2016; Therrien et al., 2017). Feeding rates of raptors during migration can vary both within and among species, with individuals migrating longer distances more likely to feed en route than those covering shorter distances (Candler and Kennedy, 1995). In contrast, some long-distance migratory species are hypothesized to feed less than short-distance migrants as they are assumed to have built up fat reserves prior to departure (Smith et al., 1986). Feeding rates may also differ between individuals migrating earlier compared to those later in the season as weather, prey availability, and other conditions can change by date. There are several examples of raptors observed feeding during migration (Shelly and Benz, 1985; Gorney and Yom-Tov, 1994; Nicoletti, 1996; Yosef, 1996), but relatively few studies have evaluated the feeding rates of migrating raptors throughout a migration season and across many years, how the proportion of feeding vs. non-feeding individuals varies between early vs. late migrants, and how these proportions vary across years. It is thus increasingly interesting to evaluate the feeding rates of migratory raptors in the current context of global climate change (Lehikoinen, 2009) and other widespread changes such as declines in songbird populations (Rosenberg et al., 2019).
We studied the feeding rates of four migrating raptor species over several periods that span 35 years. Our studied raptors include one complete, long-distance migrant, the Broad-winged Hawk (Buteo platypterus), and three partial, short-distance migrants: Sharp-shinned Hawk (Accipiter striatus), Cooper’s Hawk (Accipiter cooperii), and Red-tailed Hawk (Buteo jamaicensis). During migration, Broad-winged Hawks have been observed to feed on insects but during the nesting season, they also feed on small mammals, birds, and reptiles (McCabe et al., 2019; Goodrich et al., 2020). In contrast, the diet of Sharp-shinned and Cooper’s hawks is mainly composed of birds (Bildstein et al., 2020; Rosenfield et al., 2020) but, depending on the geographic area and time of year, the diet of the two Accipiter species could vary such that, and at least for Cooper’s Hawk, there could be greater use of mammals (Rosenfield et al., 2020; Rosenfield et al., 2022), whereas Red-tailed Hawks prey mainly on small mammals but also take birds and reptiles (Preston, 2000). Our four focal species differ in their flight strategies, with Broad-winged and Red-tailed hawks relying mostly on soaring flight, taking advantage of thermal lift to migrate, while Sharp-shinned and Cooper’s hawks more often use powered-flapping flight and gliding on orographic lift with occasional soaring flight (Bildstein, 2006; Duerr et al., 2012).
The four studied species are listed as Least Concern according to the IUCN Criteria (IUCN, 2022). Even though this is not a critical category, all these species need special attention, as they exhibited a decline according to recent results from the Raptor Population Index in North America (Oleyar et al., 2021). Most importantly, knowledge gained on relatively common species can be of great assistance when assessing limiting factors in threatened species or to prevent further loss in the common ones.
In this study, we (1) compare the feeding rate of migrating raptors using powered-flapping vs. soaring flight and the feeding rate of complete, long-distance migrants vs. partial, short-distance migrants, (2) examine differences in feeding rates between early and late migrants among species, and (3) assess if feeding rates varied over the last 35 years per species. Based on the assumption that powered-flapping flight requires more energy than soaring, we hypothesized that species using mainly powered-flapping flight would be more likely to feed during migration than the primarily soaring ones. We assumed that long-distance migrants would be less likely to feed, as they are known to build up fat stores before departure (Bildstein, 2006), compared to short-distance migrants. We predicted that migrating raptors are more likely to feed during the late portion of their migration, when conditions allowing efficient soaring flight are much reduced and thus energy demand for migration is higher, compared to early migrants (Maransky et al., 1997; Bildstein, 2006). In addition, we predicted that the proportion of individuals feeding during the early portion of their migration will increase over years, based on the assumption that avian prey species are generally advancing their migration phenology in relation to climate change (Parmesan, 2006) and this might involve changes in the timing in some key prey, such as songbirds. Lastly, we predicted that there will be no significant change in feeding rate for raptors feeding mainly on mammals (i.e., Red-tailed Hawk).
Methods
Study area and data collection
Our study took place at Hawk Mountain Sanctuary, a well-known raptor migration watch site in the Central Appalachian Mountains of Pennsylvania, United States (for a detailed description of this site and raptor migration protocols see Bednarz et al., 1990, Bildstein, 2006; Therrien et al., 2012). Observations were conducted at the North Lookout (40.641, -75.991), located 464 m above sea level atop the Kittatinny Ridge, a leading line concentrating raptors during migration.
Migrating raptors are often seen at eye level or just above eye level at the site. Observers used binoculars to count migrating raptors passing through and recorded migrating individual Broad-winged, Red-tailed, Cooper’s, and Sharp-shinned hawks exhibiting a distended crop (a proxy for recent feeding) during autumn migration, i.e., between 15 August–15 December in 1987, 2002, from 2011–2019, and in 2022.
Observers focused on birds that were close enough to determine the presence or absence of a distended crop. A crop was recorded as “full” if there was a bulge in the ventral profile of the throat, which was used as evidence of recent prey ingestion (Shelly and Benz, 1985), and “empty” if no distension was noted for the migrants assessed. We recognize that crop assessment is limited by the height of migrants; however, we assumed birds with crops are equally likely to fly higher and lower above the lookout. Crop assessment did not occur every day and not all migrants were assessed for crops.
Statistical analysis
To compare the feeding rate (proportion) of species relying on powered-flapping vs. soaring flight, we divided the four species into two groups (soaring vs. flapping) and compared them using a Chi-square test. Similarly, to compare the feeding rate of short- vs. long-distance migrants, we divided the four species into two groups (short- vs. long-distance) and compared groups using a Chi-squared test. For each year and for every studied species, we identified the median migration passage date of all migrating individuals counted at the site. The day identified as the median date was the one where half of the total of the individuals had passed. We then assigned individual observations to either early or late migrants according to whether they passed before or after the median date. To assess if migrating raptors had an increased frequency of feeding during the early vs the late portion of their migration passage window, we used a Generalized Linear Model (GLM) with a Bernoulli distribution (appropriate for binary [presence/absence] data).
To assess if the propensity of feeding during the early (or late) migration varied over time, we used a GLM with the response variable, the probability of showing a distended crop during either early or late migration, as a function of the years as the explanatory variable. We used R and RStudio (4.2.0) software with packages ddply and ggplot2, for statistical analyses R Core Team (2023). RStudio: Integrated Development Environment for R. Posit Software, PBC, Boston, MA. URL http://www.posit.co/). All analyses were run for each species separately.
Results
We recorded an annual mean of 10 Broad-winged Hawks, 17 Cooper’s Hawks, 70 Red-tailed Hawks, and 116 Sharp-shinned Hawks with distended crops (n=12 years; Appendix Table 1).
We found a higher proportion of distended crops in species using mainly powered-flapping flight: Cooper’s Hawk, 40% of the individuals recorded (209/500 hawks), Sharp-shinned hawks, 28% of the individuals recorded (1394/4896 hawks), than in species relying on soaring flight: Broad-winged Hawk: 11% of the individuals recorded (101/829 hawks) and Red-tailed Hawk: 23% of the individuals recorded (843/3657 hawks), with significant differences between these two migratory strategies (χ2 = 95.66, df =1, P < 0.01).
We found a higher proportion of distended crops in short-distance migrants: Cooper’s Hawk, Sharp-shinned Hawk, and Red-tailed Hawk (see values above), than in long-distance migrants Broad-winged Hawk (values above), with significant differences between species adopting these two migration strategies (χ2 = 86.595, df = 1, P < 0.01).
The within-season feeding rate of individuals for most of the species was higher in early than late-season migrants: (Broad-winged Hawk: slope = 469.8, Z = 3.04, P < 0.01, Figure 1; Red-tailed Hawk: slope = 71.7, Z = 4.4, P < 0.01, Figure 1; and Sharp-shinned Hawk: slope = 118.9, Z = 7.7, P < 0.01, Figure 1). Cooper’s Hawk showed a higher feeding rate during the late season (slope = -0.9, Z = -4.7, P < 0.01; Figure 1).
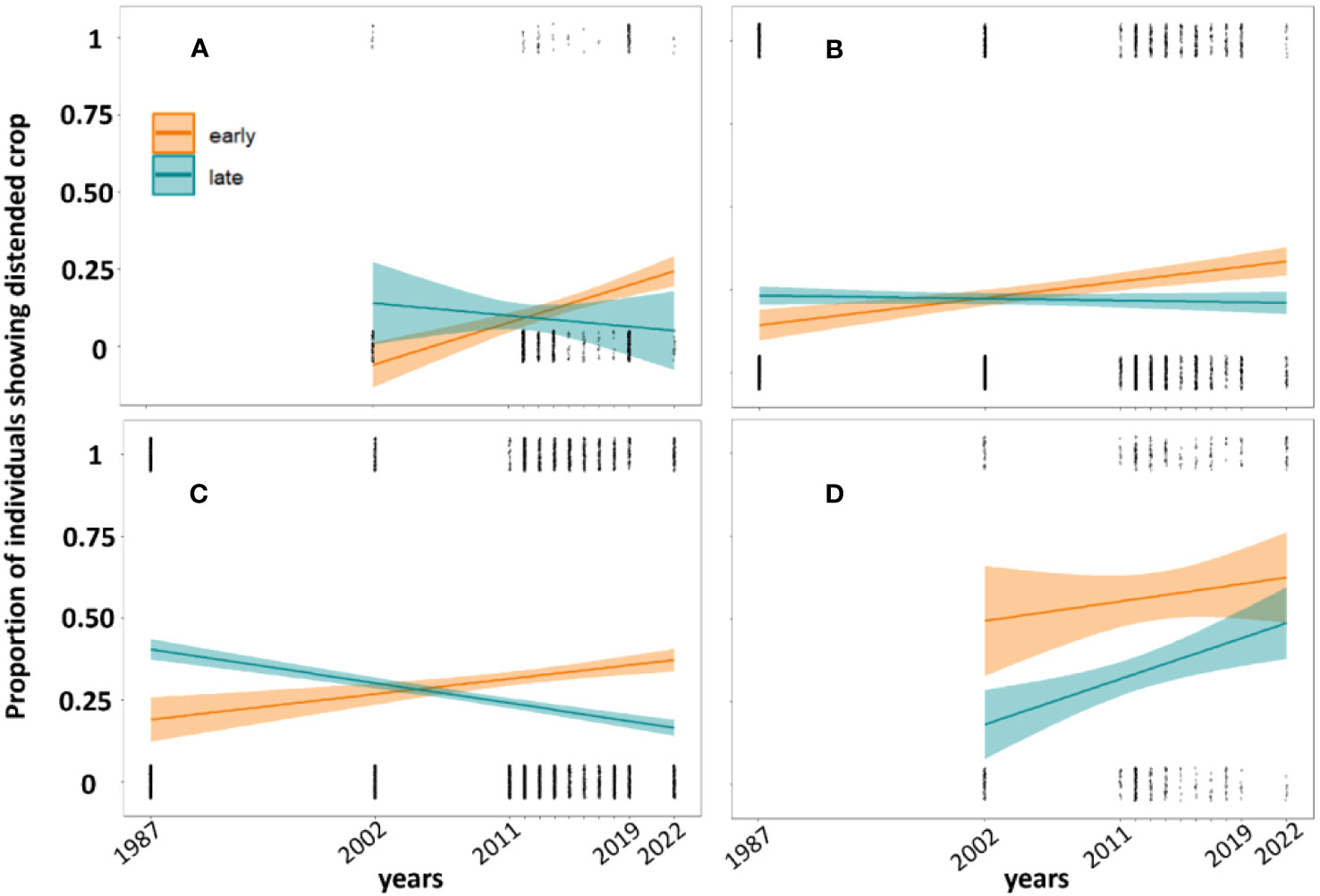
Figure 1 Proportion of individual hawks (A): Broad-winged, (B): Red-tailed, (C): Sharp-shinned, (D): Cooper’s exhibiting a distended crop during autumn migration at Hawk Mountain Sanctuary between early vs late migrants over time from 1987 to 2022.
From 1987–2022, however, we saw a significant decrease in feeding rates in late migrants for most species (Broad-winged Hawk: slope = - 0.2, Z = - 3.04, P = 0.002; Red-tailed Hawk: slope = -0.03, Z = -4.4, P < 0.01; Sharp-shinned Hawk: slope = -0.05, Z = -7.7, P < 0.01). A single species, the Cooper’s Hawk, didn’t show a significant decrease in feeding rate over time (Figure 1).
We found a significant long-term increase in feeding rate for three species from 1987–2022 (Broad-winged Hawk: slope = 0.1, Z = 4.9, P < 0.01, Figure 2; Red-tailed Hawk: slope = 0.008, Z = 2.3; P = 0.01, Figure 2; Cooper’s Hawk: slope: 0.06; Z = 3.5, P < 0.01, Figure 2). The Sharp-shinned showed a significant decrease in feeding rate over time (slope = -0.01, Z = -5.5, P < 0.01; Figure 2).
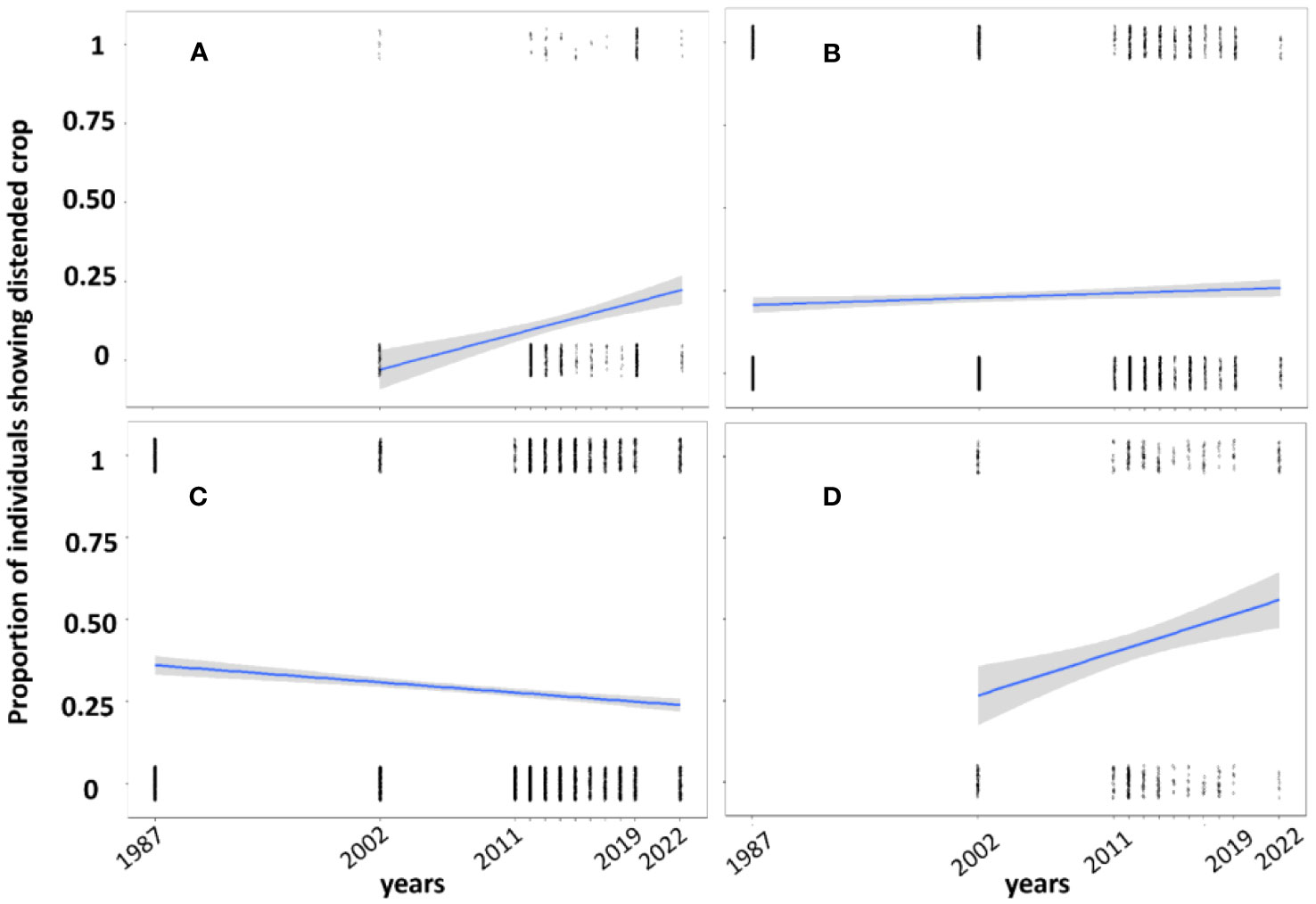
Figure 2 Proportion of individual hawks (A): Broad-winged, (B): Red-tailed, (C): Sharp-shinned, (D): Cooper’s exhibiting a distended crop overall during autumn migration at Hawk Mountain sanctuary over time (1987-2022).
Discussion
Both short and long-distance migrants appear to regularly feed during migration. This finding supports the fact that most individuals need to replenish energy levels at some point during migration and that this is done by both, soaring species (Broad-winged and Red-tailed hawks) and short-distance migrants (Sharp-shinned, Cooper’s, and Red-tailed hawks). Feeding by raptors during migration has been reported in other studies using telemetry on soaring raptors, such as Broad-winged and Swainson’s hawks. Indeed, the studies revealed that they stop and feed regularly during migration, especially during the early portion of the journey (Kochert et al., 2011; McCabe et al., 2020).
Not surprisingly, the overall proportion of individuals feeding during migration showed a higher percentage of individuals with distended crops for species mainly using powered-flapping flight compared to mostly soaring ones, most likely related to the higher energy demand of this flying strategy (Bildstein, 2006). The overall proportion of birds feeding during migration showed a higher percentage of individuals with distended crops for the species with short-distance migration strategies compared to the long-distance migrants. This observation might be related to the fact that long-distance migrants may have built fat stores before the departure or that traveling in large groups could prevent feeding opportunities. These factors could explain why they could feed less frequently during migration (Bildstein, 2006).
Three of the four studied species showed a significant decrease in feeding rate during the late portion of their migration over the last two decades. This is in line with our prediction that raptors are less likely to feed during the late portion of migration because climate change is leading raptors’ songbird prey to migrate early, but also includes the species feeding mainly on mammals (Red-tailed Hawk). With ongoing global changes, many late migrating raptors are likely facing the fact that potential prey may have already moved out or became less available within a given area (Parmesan, 2006).
It is important to consider that the method used in this study is based on visual observations of raptors, during migration, and the assessment of the evidence of distended crop. According to the raptor diet, we acknowledge that it might be difficult to assess the presence of a distended crop in Broad-winged hawks, an insect-eater, after recent ingestion of prey (i.e., a dragonfly), because this might not exhibit an obvious distended crop (Shelly and Benz, 1985). Moreover, this study was conducted at a single site. To better understand the feeding behavior of migratory raptors under a scenario of global climate change, more studies scattered across a broad geographical region and data collection with specific attention on feeding behavior are encouraged.
Changes in climate are affecting species worldwide and they could lead to negative implications for the ecosystems if the changes in the life history traits of predators, such as migration timing, do not synchronize to those of their prey (Parmesan, 2006). Assessing feeding rate in migrating raptors and how this rate is changing over time is crucial to understand how these global changes are affecting the population of the raptor species. With food availability being an aspect of primary concern, information regarding how species adjust to varying conditions can play an important role in the conservation of several raptor species.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
All authors have contributed to designing the study, collecting data, analyzing data and writing of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
We thank Hawk Mountain volunteers, trainees, and staff who collected and compiled data over the years, particularly Jason Ryan, Jane Ching, and Rebecca Laritz. Special thanks to the James A. Kushlan Award and the Hawk Migration Association of North America (HMANA) for supporting this project. Thanks to the Acopian Center for Conservation Learning, Hawk Mountain Sanctuary, for supporting researchers from all over the world. This is Hawk Mountain Sanctuary’s contribution to conservation science #395.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bednarz J. C., Klem D. Jr., Goodrich L. J., Senner S. E. (1990). Migration counts of raptors at Hawk Mountain, Pennsylvania, as indicators of population trends, 1934-1986. The Auk 107(1), 96–109.
Bildstein K. L. (2006). Migrating Raptors of the World: Their Ecology and Conservation (Ithaca, New York (United States): Ithaca, Cornell University Press).
Bildstein K. L., Meyer K. D., White C. M., Marks J. S., Kirwan G. M. (2020). “Sharp-shinned hawk (Accipiter striatus),” in version 1.0. in Birds of the World. Eds. Billerman S. M., Keeney B. K., Rodewald P. G., Schulenberg T. S. (Ithaca, NY: Cornell Lab of Ornithology). doi: 10.2173/bow.shshaw.01
Candler G., Kennedy P. (1995). Flight strategies of migrating osprey: fasting vs. foraging. J. Raptor Res. 29, 85–92.
Dekker D. (1980). Hunting success rates, foraging habits, and prey selection of peregrine falcons migrating through central Alberta. Can. Field Nat. 94, 371–382.
Duerr A. E., Miller T. A., Lanzone M., Brandes D., Cooper J., O'Malley K., et al. (2012). Testing an emerging paradigm in migration ecology shows surprising differences in efficiency between flight modes. PloS One 7, e35548. doi: 10.1371/journal.pone.0035548
Goodrich J. L., Smith J. P. (2008). “Raptor migration in North America,” in State of North America’s Birds of Prey. Eds. Bildstein K. L., Smith J. P., Inzunza E. R., Veit R. R. (Washington, DC: Nuttall Ornithological Club and American Ornithologists' Union), 37–149.
Goodrich L. J., Crocoll S. T., Senner S. E. (2020). “Broad-winged Hawk (Buteo platypterus),” in version 1.0. in Birds of the World. Ed. Poole. A. F. (Ithaca, NY: Cornell Lab of Ornithology). doi: 10.2173/bow.brwhaw.01
Gorney E., Yom-Tov Y. (1994). Fat, hydration condition, and moult of Steppe Buzzards Buteo buteo vulpinus on spring migration. Ibis 136, 185–192. doi: 10.1111/j.1474-919X.1994.tb01083.x
Kjellén N., Hake M., Alerstam T. (2001). Timing and speed of migration in male, female and juvenile ospreys Pandion haliaetus between Sweden and Africa as revealed by field observation, radar and satellite tracking. J. Avian Biol. 32, 57–67. doi: 10.1034/j.1600-048X.2001.320108.x
Kochert M. N., Fuller M. R., Schueck L. S., Bond L., Bechard M. J., Woodbridge B., et al. (2011). Migration patterns, use of stopover areas, and austral summer movements of Swainson’s Hawks. Condor. 113, 89–116. doi: 10.1525/cond.2011.090243
Lehikoinen A. (2009). Climate forcing on avian life history (Helsinki (Finland: University of Helsinki).
Maransky B., Goodrich L. J., Bildstein K. L. (1997). Seasonal shifts in the effects of weather on the visible migration of Red-tailed Hawks at Hawk Mountain, Pennsylvania 1992-1994. Wilson Bull. 109, 246–252.
McCabe R. A., Goodrich L. J., Barber D., Master T., Watson J. L., Bayne E. M., et al. (2020). Satellite tracking reveals age and origin differences in migration ecology of two populations of Broad-winged Hawks (Buteo platypterus). Wilson J. Ornith. 132, 1–14. doi: 10.1676/1559-4491-132.1.1
McCabe R. A., Goodrich L. J., Master T., Bordner Z. (2019). Broad-winged hawk nesting behavior in forested landscape of Pennsylvania. J. Raptor Res. 53, 293–308. doi: 10.3356/JRR-18-56
Moore F., Kerlinger P. (1987). Stopover and fat deposition by North American wood-warblers (Parulinae) following spring migration over the Gulf of Mexico. Oecologia 74, 47–54. doi: 10.1007/BF00377344
Newton I. (1998). “Migration patterns in west palearctic raptors,” in Holoarctic Birds of Prey. Eds. Chancellor R. D., Meyburg B.-U., Ferrero J. J. (Merida and Berlin: Proceedings of an International Conference), 603–612.
Newton I. (2006). Can conditions experienced during migration limit the population level of birds? J. Ornith. 147, 146–166. doi: 10.1007/s10336-006-0058-4
Nicoletti F. J. (1996). American Kestrel and Merlin migration correlated with Green Darner Movement at Hawk Ridge. Loon. 68, 216–221.
Niles L., Burger J., Clarck K. (1996). The influence of weather, geography, and habitat on migrating raptors on cape may peninsula. Condor. 98, 382–394. doi: 10.2307/1369156
Oleyar D., Ethier D., Goodrich L., Brandes D., Smith R., Brown J., et al. (2021) The Raptor Population Index: 2019 Analyses and Assessments. Available at: https://www.rpi-project.org/2019-analysis/.
Parmesan C. (2006). Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 37, 637–669. doi: 10.1146/annurev.ecolsys.37.091305.110100
R Core Team. (2023). _R: A Language and Environment for Statistical Computing_. (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/
Rosenberg K. V., Dokter A. M., Blancher P. J., Sauer J. R., Smith A. C., Smith P. A., et al. (2019). Decline of the North American avifauna. Science 366, 120–124. doi: 10.1126/science.aaw1313
Rosenfield R. N., Grosshuesch D. A., Stewart A. C., Stout W. E., Driscoll T. G., Seibel J. P. (2022). Commentary: untenables and clarifications about Accipiter migration, resurgence in urban habitats, and prey preferences in North America. Wilson J. Ornith. 134, 560–565. doi: 10.1676/21-00032
Rosenfield R. N., Madden K. K., Bielefeldt J., Curtis O. E. (2020). “Cooper's Hawk (Accipiter cooperii),” in version 1.0, in Birds of the World. Ed. Rodewald P. G. (Ithaca, NY: Cornell Lab of Ornithology). doi: 10.2173/bow.coohaw.01
Rudebeck G. (1950). The choice of prey and modes of hunting of predatory birds with special reference to their selective effect. Oikos. 2, 65–88. doi: 10.2307/3564663
Rudebeck G. (1951). The choice of prey and modes of hunting of predatory birds with special reference to their selective effect. Oikos. 3, 200–231. doi: 10.2307/3565185
Shelly E., Benz S. (1985). Observation of aerial hunting, food carrying and crop size of migrant raptors (Quarryville and Kempton, Pennsylvania (United States): ICBP Technical Publication No. 5), 299–301.
Smith N. G., Goldstein D. L., Bartholomew G. A. (1986). Is long-distance migration possible for soaring hawks using only stored fat? Auk 103, 607–611.
Sullivan A. R., Flaspohler D. J., Froese R. E., Ford D. (2016). Climate variability and the timing of spring raptor migration in eastern North America. J. Avian Biol. 47, 208–218. doi: 10.1111/jav.00692
Therrien J. F., Goodrich L. J., Barber D. R., Bildstein K. L. (2012). A long‐term database on raptor migration at Hawk Mountain Sanctuary, northeastern United States: Ecological Archives E093‐174. Ecology 93(8), 1979–1979.
Therrien J. F., Lecomte N., Zgirski T., Jaffré M., Beardsell A., Goodrich L. J., et al. (2017). Long-term phenological shifts in migration and breeding area residency in eastern North American raptors. Auk. 134, 871–881. doi: 10.1642/AUK-17-5.1
Yosef R. (1996). Raptors feeding on migration at Eilat, Israel: Opportunistic behavior or migratory strategy? J. Raptor Res. 30, 242–245.
Appendix

Table 1 Median passage date and number of migrating individuals for which feeding rate was assessed for each species annually at Hawk Mountain, (USA).
Keywords: eastern North America, soaring, hawks, timing, distance, global change
Citation: Verduci M, Goodrich LJ, Therrien J-F and Ruelas Inzunza E (2023) Feeding rates of raptors during autumn migration in the Central Appalachians 1987–2022. Front. Conserv. Sci. 4:1250142. doi: 10.3389/fcosc.2023.1250142
Received: 29 June 2023; Accepted: 06 November 2023;
Published: 24 November 2023.
Edited by:
Oded Berger-Tal, Ben-Gurion University of the Negev, IsraelReviewed by:
Ali T. Qashqaei, Borderless Wildlife Conservation Society, IranRobert Rosenfield, University of Wisconsin–Stevens Point, United States
Copyright © 2023 Verduci, Goodrich, Therrien and Ruelas Inzunza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marzia Verduci, bWFyemlhLnZlcmR1Y2lAaG90bWFpbC5pdA==