 Yang Qu
Yang Qu Zexi Zhou
Zexi Zhou Tae-Ho Lee
Tae-Ho Lee- 1School of Education and Social Policy, Northwestern University, Evanston, IL, United States
- 2Department of Human Development and Family Sciences, The University of Texas at Austin, Austin, TX, United States
- 3Department of Psychology, Virginia Tech, Blacksburg, VA, United States
Children and their parents are wired to connect as it provides the foundation for developing children to adapt to an increasingly complex environment. Although extensive studies demonstrate the importance of parent-child dyadic similarity at the behavioral, psychological, and physiological levels in fostering children's learning and psychological wellbeing, little is known about parent-child similarity at the neural level until recently. Drawing on our own work and the work by other scholars, this review summarizes recent advances in empirical research on parent-child neural similarity. Specifically, this review elaborates the theoretical importance of studying parent-child neural similarity and showcases how parent-child neural similarity is assessed using different neuroimaging approaches. We further synthesize empirical evidence about the contextual and individual factors that may contribute to variability in parent-child neural similarity, summarize how such neural similarity is related to different aspects of child adjustment, and highlight important directions for future research. Taken together, we hope that this integrative review can demonstrate cutting-edge research that explores neural similarity in parent-child dyads, and provide researchers with a clear roadmap to examine parent-child neural similarity in order to gain a better understanding of parental socialization process and brain development.
1. Introduction
Children and parents are wired to connect as it provides the foundation for developing children to adapt to an increasingly complex environment (Ainsworth et al., 2015). For a long time, human developmental studies have demonstrated that parent-child dyads often show similarities at the behavioral, psychological, and physiological levels (Kochanska and Aksan, 1995; Harrist and Waugh, 2002; DePasquale, 2020; Rogers et al., 2022) and such dyadic similarities play a major role in children's adjustment across development (Boyum and Parke, 1995; Feldman, 2007; Feng et al., 2007). Despite extensive studies highlighting the importance of parent-child dyadic similarity in fostering children's learning and psychological wellbeing, little is known about parent-child similarity at the neural level until recently. In the past few years, a growing number of studies begin to investigate parent-child neural similarity using a variety of neuroimaging approaches, such as functional magnetic resonance imaging (fMRI), functional near-infrared spectroscopy (fNIRS), and electroencephalography (EEG) (e.g., Lee et al., 2017a,b, 2018; Reindl et al., 2018; Nguyen et al., 2021b; Haresign et al., 2022). Therefore, our review aims to summarize recent advances in empirical research on parent-child neural similarity. First, we explain why parent-child neural similarity serves as an important lens to understand human development within the family context and provides unique information beyond prior methods (e.g., survey, observation, and interview). Second, we showcase how parent-child neural similarity is assessed and quantified using different neuroimaging approaches in recent research. Third, we synthesize empirical evidence regarding the contextual and individual factors that may contribute to variability in parent-child neural similarity and delineate how such neural similarity is related to different aspects of child adjustment. Finally, we point out important directions for future research. Taken together, this integrative review systematically demonstrates cutting-edge research that explores neural similarity in parent-child dyads, and provides researchers with a clear roadmap to further this endeavor in order to gain a better understanding of socialization process and brain development.
2. The importance of understanding neural similarity in parent-child dyads
Examining neural similarity in parent-child dyads can provide novel theoretical and empirical insights in multiple ways. First, it can offer direct evidence on how parents and children's brains are wired together from a dyadic perspective. Although previous neuroimaging studies have shed light on the role of parents in children's brain development (e.g., Belsky and De Haan, 2011; Whittle et al., 2014), the scope of these studies has largely been limited because much of these works just scan children's brain while evaluating parental profiles via traditional measures such as self-report or observation. For example, prior research has examined how parent-child relationships, parental depression, and even the mere presence of parents, modulate adolescents' brain development and psychological adjustment (Qu et al., 2015, 2016a; Telzer et al., 2015). The paucity of understanding in dyadic similarity at the neural level is partly due to the lack of studies that take parents' brain into consideration to explain children's neurobiological development beyond a single-brain perspective. Similarly, a growing body of literature on parental brain typically just assesses parents' neural responses to multiple types of stimuli related to children (e.g., infant cry, baby movies; Feldman, 2015; Kringelbach et al., 2016; Lee et al., 2019; Stark et al., 2019). For example, research found that mothers showed enhanced activity in specific brain regions (e.g., inferior frontal regions, striatum) when hearing their own infants' cries across cultures (Bornstein et al., 2017). However, without employing brain scans for both parents and their children, it remains unknown about the shared neural processes in parent-child dyads.
Second, the brain-level measures of parent-child similarity can provide insights into the mechanisms of within-family interpersonal concordance. Scholars have long been interested in understanding interpersonal similarity in parent-child dyads, including physiological (e.g., heart rate; Feldman et al., 2011), hormonal (e.g., cortisol levels; Papp et al., 2009), and emotional similarity (e.g., shared affect; Feldman et al., 1999). However, there are still open questions about why such coordinated behavior and psychophysiology occur and what might predict such concordance. Recent neuroimaging studies indicate that parent-child neural similarity is related to their behavioral similarity. For example, parent-child dyads with similar default-mode network connectivity showed greater similarity in daily sleep behavior routines (Lee et al., 2017a). Therefore, examining the similarity of neural configruation profiles between children and parents help understand the underlying mechanisms of dyadic similarity in affect, behavior, and psychophysiology.
Third, parent-child neural similarity can serve as a unique lens and provide a novel perspective to understand the well-established theories on human development. For example, examining parent-child neural similarity may enable more in-depth investigation of the attachment theory. Specifically, parent-child neural similarity can increase the understanding of the mechanisms regarding how attachment to primary caregivers may get under the skin (Pietromonaco et al., 2013; Pietromonaco and Powers, 2015) and have long-lasting effects on individuals' psychological, emotional, and social development from a neurobiological perspective (Lee et al., 2018). It allows researchers to examine many central propositions of the attachment theory such as the consistency hypothesis. Decades of research demonstrates the continuity of attachment over time and across relationships (e.g., from parent-child relationship to romantic relationship and friendship; Rubin et al., 2004; Selcuk et al., 2010; Feldman et al., 2013; Ulmer-Yaniv et al., 2022). The advances in quantifying neural similarity may offer novel empirical evidence for testing such theoretical hypotheses (e.g., how parent-child neural similarity may predict whom children become friends with later in life). Similarly, understanding parent-child neural similarity can provide insights into the theories regarding parental socialization and children's internalization. Given that parents and children may not be able to articulate their psychological processes, assessment of dyadic neural similarity can provide a powerful tool to sensitively capture parents and children's autonomic responses to interactions. Understanding this invisible process can shed light on theories on parental socialization and parent-child interactions. For example, scholars argue that elucidating dyadic neural synchrony during parent-child interactions can help distinguish competing theories regarding the interactive directionality in parent-child dyads (e.g., a uni-directional process in which children implicitly model their parents vs. bidirectional influences between children and parents) (Ratliff et al., 2021).
3. Measurements of parent-child neural similarity
It is worth to note that there are different methods to examine parent-child neural similarity and they can be broadly divided into two approaches based on which features of interpersonal process they focus on. The first approach, with relatively long tradition in dyadic studies, is to focus on the moment-to-moment inter-personal synchronization of neural signals when both parents and children are engaged in interactive activities (e.g., conversation) or exposed to the same stimuli at the same time (e.g., listen to the same music) (e.g., Reindl et al., 2018; Nguyen et al., 2020; Turk et al., 2022). This approach is interested in the contexts in which parents and children show in-synched temporal changes of neural activity, namely real-time neural synchronization, to capture their reciprocal processes with assumptions that such dyadic synchrony occurs instantaneously and may reflect their harmonized psychological and behavioral status at the given moments. Given that this approach directly compares the temporal characteristics of neural signals in parent-child dyads online, it is necessary to adopt neuroimaging techniques with higher temporal resolution such as electroencephalography (EEG), functional near-infrared spectroscopy (fNIRS), and magnetic electroencephalography (MEG), and thus the analyses are more often focused to quantify temporal coherence of inter-brain signal fluctuation (i.e., temporal synchrony). For instance, the phase synchronization level of EEG signal oscillations at a certain frequency band (e.g., gamma band, 30–80 Hz; Figure 1A) is calculated directly between electrodes attached to the brain of each individual (Babiloni and Astolfi, 2014; Hari et al., 2015; Hasson and Frith, 2016). However, the limitations of spatial resolution in the neural synchrony approach prevent the identification of the specific brain regions or networks mainly responsible for synchronization, and hinder the investigation of whole-brain activity, especially the limbic system.
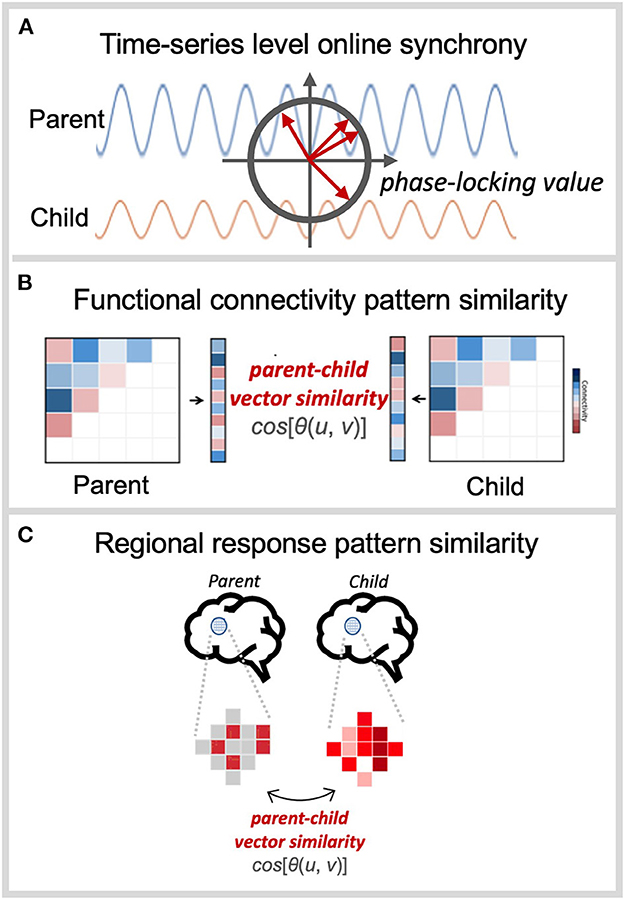
Figure 1. Illustration of approach for assessing parent-child neural similarity using (A) online time-series synchrony, (B) functional connectivity pattern, and (C) task-based pattern similarity.
The measure of parent-child neural similarity has recently been extended to fMRI approach. Different from the first approach, the implications of fMRI approach focus on neural pattern similarity through either intrinsic connectivity pattern (Lee et al., 2017a,b) or response pattern to a certain event (Lee et al., 2018) between parents and children. Even though fMRI has limitation in capturing the real-time interactive process, the approach using fMRI is possible to quantify the extent to which parent-child dyads' neural profiles remain in-tuned over time as well as how similarly they respond to the environments. The connectivity-based pattern similarity has been examined more through the resting-state fMRI data (Figure 1B), as it reflects neural foundation of individuals' psychological status, namely intrinsic neural systems (Cole et al., 2014). That is, this approach measures the similarity of the intrinsic brain system with the assumption that the interactive process over time tunes the functional configurations of the brains, resulting in similar neural profiles between parents and children. For example, recent work assessed the similarity of functional connectivity patterns across intrinsic networks to evaluate the extent to which the brain's functional architectures are similarly formed between parents and children (Lee et al., 2017b).
Another way of using the pattern similarity is to examine how similarly parents and children respond to a specific event (i.e., representational pattern similarity; Kriegeskorte et al., 2008; Diedrichsen and Kriegeskorte, 2017). This approach focuses more on the similarity of the neural response pattern to task-specific events between parents and children, rather than on the connectivity configuration similarity (Figure 1C). In fact, this approach can be implemented using various neuroimaging modalities such as EEG, fNIRS, and MEG, as the response pattern can be measured across electrodes, after stimulus onsets. However, fMRI can offer greater benefits as it allows for the examination of specific regions of interest (ROIs) considered as a core region for the task-specific process. For example, a recent study by Lee et al. (2018) aimed to evaluate the neural representational similarity between parents and children when they are exposed to stressful stimuli. The researchers first identified the brain regions particularly involved in stress processing (e.g., insula), and they calculated the neural response pattern similarity during stress-inducing task between parents and children within the ROI.
Notably, recent studies attempt to measure parent-child neural synchrony using fMRI (Ratliff et al., 2021), especially with hyper-scanning protocols (using two fMRI machines simultaneously). Although temporal resolution in fMRI is relatively less precision than EEG/fNIRS/MEG methods, it provides important information of low frequency trends in neural processing, and thus fMRI-based neural synchrony studies increase in last couple of years. However, given the simultaneous scanning of parent-child dyads during reciprocal interactions, this approach requires two fMRI scanners on site.
4. Variability and antecedents of parent-child neural similarity
It is also important to note that similar to dyadic concordance at behavioral or physiological levels, there is significant variability in dyadic concordance at the neural level (e.g., Lee et al., 2018). Therefore, scholars have begun to identify the antecedents that may modulate the level of parent-child neural similarity. Below we summarize familial and individual factors that receive relatively more attention in prior research.
4.1. Quality of parent-child interaction or relationship
Theoretical work suggests that harmonious interpersonal interactions are associated with more in-tune neural states between two individuals (Wheatley et al., 2012). Therefore, it has been thought that parent-child positive interaction or relationship quality may increase the levels of neural similarity. Indeed, scholars have examined neural similarity across different aspects of positive parent-child interactions, such as cooperation (vs. competition), positive (vs. negative) emotion during interaction, and high (vs. low) behavioral reciprocity (e.g., Reindl et al., 2018; Miller et al., 2019; Nguyen et al., 2020; Santamaria et al., 2020). This line of research has consistently demonstrated that during positive interactions, parent-child dyads show increased neural synchrony in regions involved in cognitive and social processes (e.g., prefrontal cortex and temporo-parietal junction) (Reindl et al., 2018; Miller et al., 2019; Nguyen et al., 2020, 2021a,b; Morgan et al., 2023). Interestingly, some research compared parent-child dyads with stranger-child dyads, and found that children do not show significant brain-to-brain synchrony with strangers during positive interactions (e.g., Reindl et al., 2018). This finding suggests that increased neural synchrony during positive interactions may be specific to parent-child dyads.
In addition to momentary parent-child interaction quality, the long-term relationship quality also seems to influence neural similarity of parent-child dyads. Consistent with prior evidence that positive parent-child relationship quality plays a vital role in increasing shared dyadic process in affect, behavior and physiology (Davis et al., 2018; Han et al., 2019; Birk et al., 2022), a recent study demonstrated that mother-child dyads with greater family connectedness show more similar neural response patterns to stress (Lee et al., 2018). However, other research did not find the link between parent-child attachment quality and inter-brain synchrony during cooperation (Miller et al., 2019). It is possible that parent-child relationship quality may play a larger role in dyadic neural similarity during negative or stressful situations, which calls for more future research to explore.
4.2. Characteristics of parents
Research highlights the roles of parents' beliefs, state, and parenting practices in parent-child neural similarity. With regard to parents' beliefs, research suggests that when fathers believed they should be involved with and act sensitively toward their children, there was increased father-child neural synchrony during cooperation (Nguyen et al., 2021b). Moreover, recent research has also begun to examine the complex role of parenting stress in parent-child neural similarity. Although greater parenting stress was associated with lower levels of brain-to-brain synchrony in the prefrontal cortex when mother-child dyads watched animation videos together (Azhari et al., 2019), a different pattern was shown when parent-child dyads engaged in shared play. During joint play, greater parenting stress was related to less synchrony in posterior areas, but more synchrony in frontal areas of the prefrontal cortex (Azhari et al., 2023). These seemingly contradictory findings highlight the importance of understanding the nature of the dyadic activity. It has been argued that even though parenting stress leads to less similarities in brain regions involved in emotional processing and regulation, joint interactions also require greater neural synchrony in brain regions that support attentional regulation among dyads with more stressed parents (Azhari et al., 2023).
Given the important implications of parenting practices for children's adjustment (e.g., Morris et al., 2017, 2021), scholars have also examined the role of parenting practices in parent-child neural similarity. However, emerging evidence is less clear. Some research suggests that fewer supportive parent emotion socialization was associated with cross-brain connectivity between multiple emotion-related brain regions in parent-child dyads (Ratliff et al., 2021), whereas other research indicates that higher levels of parental psychological control may lead to negative association in frontal resting EEG asymmetry among parent-child dyads (Wang et al., 2018). Therefore, more research is needed to investigate how different aspects of parenting practices (e.g., parental autonomy support and control) play a role in parent-child neural similarity in a variety of settings.
In addition to parents' characteristics, researchers also explored whether parent-child neural similarity is disrupted by exposure to stressors at the family level. For example, a burgeoning literature demonstrated that family adversity decreases parent-child behavioral and physiological synchrony (e.g., Clearfield et al., 2014; Creaven et al., 2014; Suveg et al., 2016). In line with this literature, recent work showed that demographic factors such as low family income and parents' educational attainment are associated with decreased parent-child neural synchrony in the context of experimentally-induced stress (Hoyniak et al., 2021). These findings not only identify the negative impact of stress on families facing adversity, but also highlight the importance of distinguishing between family contexts and parents' characteristics (e.g., parenting stress) when examining parent-child neural similarity.
4.3. Characteristics of children
Although only limited work has considered the role of children, empirical evidence indicates that children's characteristics may also influence parent-child neural similarity. For example, recent research found that higher child irritability was associated with less parent-child neural synchrony in the lateral prefrontal cortex, suggesting that children with higher levels of irritability may have difficulties in achieving synchrony (Quiñones-Camacho et al., 2020). Moreover, children's greater autonomy in the collaborative task (e.g., actively approach working on the task and initiate goal-directed behavior) was associated with higher neural synchrony during parent-child cooperation (Nguyen et al., 2020), highlighting the active role of children in shaping dyadic neural similarity. It is important to point out that the familial and individual factors examined in prior research may not be isolated from each other. For example, children's irritability may influence parent-child interaction quality, which further play a role in parent-child neural similarity. Therefore, it is important to examine the unique and overlapping effects of these factors on parent-child neural similarity.
5. Developmental consequences of parent-child neural similarity
Researchers not only quantify parent-child neural similarity across various contexts, but also examine the implications of such similarity for children's performance and adjustment. Decade of research demonstrates that parent-child dyadic similarities at the behavioral and psychological levels are associated with children's positive outcomes (Boyum and Parke, 1995; Feldman, 2007; Feng et al., 2007). Moreover, evidence from social neuroscience suggests that greater inter-brain synchrony in teams predicts better collective performance (e.g., Gumilar et al., 2021; Reinero et al., 2021). In line with this research, extant studies have examined how neural similarity in parent-child dyads is related to short-term outcomes during parent-child interactions. For example, increased parent-child neural synchrony in the dorsolateral prefrontal and frontopolar cortex during prior cooperation was predictive of their better cooperative performance in subsequent trials (Reindl et al., 2018), but not vice versa, suggesting the potential causal link between neural synchrony and performance. Furthermore, mother-child neural synchrony predicted the dyad's problem-solving success beyond their behavioral reciprocity (Nguyen et al., 2020), highlighting the unique role of neural synchrony in predicting task performance.
More importantly, a series of research examine how parent-child neural similarity is associated with children's adjustment. For example, children who had higher level of dyadic neural similarity with their parents showed better psychological adjustment such as higher emotional competence (Lee et al., 2017b), more optimal sleep quality (Lee et al., 2017a), lower level of stress (Lee et al., 2018), and more rapid decreases in internalizing behaviors (Quiñones-Camacho et al., 2022). Similarly, recent evidence suggests that parent-child neural synchrony in emotion-related brain regions during conflict discussion was associated with fewer adolescent-reported depressive symptoms (Ratliff et al., 2021). Together, these studies provide initial and promising evidence that parent-child neural similarity has predictive validity for children's performance and wellbeing.
6. Future directions
Extant research has provided exciting advances in understanding parent-child neural similarity, which also points to the directions for future research.
6.1. Taking a life-span perspective to examine changes of parent-child neural similarity over the course of development
Although prior research investigates parent-child neural similarity and its relation to children's adjustment, each study focuses on a limited age range. Therefore, it remains unknown how neural similarity in parent-child dyads emerge and change over the course of development. It is important for future research to examine developmental changes in parent-child neural similarity using cross-sectional designs that include parent-child dyads across stages or longitudinal designs that follow parent-child dyads over a long period of time. For example, it is interesting to examine how parent-child neural similarity changes over adolescence, a time during which children are individuating from parents and oriented toward peers (Collins and Steinberg, 2006). Also, as close social bonds such as intergenerational relationships can have greater implications for physical and psychological wellbeing along with aging (Charles and Carstensen, 2010; Birditt and Fingerman, 2013), how parent-child neural similarity may play a role in older adults and their adult children's life may also be worth investigation. For example, heightened neural similarity may confer benefits to both older adults and their adult children. Previous work indicates that parent-child relationship can be ambivalent, which involves both positive and negative feelings among adults and their aging parents (Fingerman et al., 2008). When adults and their aging parents have less ambivalent feelings about their relationship, they may be more likely to show neural similarity during interactions, which provides a basis for higher-quality interactions and more dyadic support exchanges (e.g., providing instrumental and emotional support) in daily life. Therefore, instead of treating parent-child neural similarity as a static snapshot, depicting its developmental changes and identifying its consequences across developmental stages will provide new insights into the understanding of family dynamics at the neural level.
6.2. Understanding parent-child neural similarity in diverse sociocultural contexts
It is important to examine how sociocultural contexts—both distal contexts (e.g., culture) and proximal situations (e.g., family relationships, lab-induced cooperation vs. competition)—influence parent-child neural similarity. Although extant research focuses on the role of proximal situations, the role of culture in parent-child neural similarity receives little attention. Scholars have highlighted the importance of examining parent-child neural similarity in diverse cultural contexts (Chen and Qu, 2021). Move beyond current literature focusing on brain development in Western cultures (Qu et al., 2021), comparing parent-child similarity in multiple cultures allow scholars to investigate whether there are mean-level differences in parent-child similarity across cultures, and whether the antecedents and consequences of parent-child similarity also vary across cultures. Importantly, advances in this line of research will increase the theoretical understanding of cultural socialization and transmission. Guided by their cultural values and beliefs, parents' socialization goals for their children as well as children's internalization of these goals may vary across cultures (Qu et al., 2016b; Ng et al., 2019). Therefore, parent-child neural similarity can provide neurobiological insights into how parents' culturally shaped socialization goals are transmitted to their children. For example, researchers can investigate how parent-child neural similarity facilitates the intergenerational transmission of cultural values (Arredondo, 2023).
Although prior research showed that both long-term family environment and momentary parent-child interaction quality may influence parent-child neural similarity (Lee et al., 2018; Nguyen et al., 2020; Hoyniak et al., 2021), it is also valuable to consider “when” and “in what context” parent-child neural similarity plays a positive role in child adjustment. Prior research on parent-child similarity at physiological level suggests that such similarity may not always be promotive and protective, especially in negative family contexts (Smith et al., 2016; Suveg et al., 2016; Davis et al., 2018; Creavy et al., 2020). Therefore, it is possible that heightened neural similarity in parent-child dyads does not always lead children to better psychological adjustment, especially when children live in adverse family environment (e.g., low parental warmth and high family conflict). For example, when parents show low warmth or more depressive symptoms, children who show greater dyadic neural similarity with their parents may exhibit maladaptive neural patterns to stress as their parents, leading to negative developmental outcomes over time. Thus, it is crucial to explore the potential family environment factors that may moderate the effects of parent-child neural similarity on children's outcomes.
6.3. Integrating various designs and advanced computational tools to capture the complexity of parent-child neural similarity
As we described above, researchers have employed different tools (e.g., fMRI, fNIRS, EEG, MEG), different tasks (e.g., problem solving, conflict discussion), and different designs (e.g., verbal and non-verbal stimuli, naturalistic stimuli, hyperscanning) (e.g., Lee et al., 2017a,b,c; Nguyen et al., 2020; Santamaria et al., 2020; Ratliff et al., 2021; Zivan et al., 2022). To rigorously compare the findings across different settings, it will be theoretically important to integrate and examine multiple designs or tasks in the same study (e.g., comparing parent-child neural similarity during verbal vs. non-verbal communication or during movie-watching vs. free play). This will allow researchers to test the generalizability of event-specific parent-child neural similarity. For example, do parent-child dyads show similar patterns of neural synchrony during free play and conflict discussion? Does parent-child neural synchrony in these settings play a similar role in children's developmental outcomes over time?
Moreover, innovative analytical techniques are developed to quantify parent-child neural similarity at multiple levels. For example, significant work has begun to examine the neural underpinnings of parent-child dyadic similarity by estimating multi-dimensional neural pattern similarity across voxels, regions, and networks between parents and their children (Lee et al., 2017a,b, 2018). However, few studies so far have considered parent-child similarity in brain structure. Prior research on adults has established the association from neural structure to neural function and further to behavior (Leong et al., 2016, 2018). It is possible that the brain structural similarity in parent-child dyads provide the infrastructure across regions and networks for parent-child functional similarity, which subsequently relates to children's outcomes. Therefore, researchers can employ multimodal neuroimaging assessment for both functional and structural similarity, examine the association between these two levels, and identify their unique and overlapping effects on children's outcomes.
7. Conclusion
With rapid development in the research on parent-child neural similarity, this promising line of research requires scholars to systematically integrate empirical evidence and clarify theoretical contributions. To address this issue, our review synthesizes recent advances in the study of parent-child neural similarity, delineates different types of measurements, and points out important directions for future research. The advances in this vibrant field not only provide novel insights into how parents and children's brains are wired together from a dyadic perspective, but also identify important antecedents and consequences of parent-child neural similarity. Ultimately, this endeavor will provide researchers with a unique lens to understand parental socialization process, parent-child interactions, and brain development.
Author contributions
YQ, ZZ, and T-HL wrote the first draft of the manuscript. All authors contributed to manuscript revision, read, and approved the submitted version.
Funding
This research was supported by funding from the National Science Foundation (BCS 1944644), the Society for Research in Child Development Small Grants Program for Early Career Scholars, the Center for Culture, Brain, Biology, and Learning at Northwestern University to YQ, and a research award by the Virginia Tech Institute for Society, Culture and Environment to T-HL.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ainsworth, M. D. S., Blehar, M. C., Waters, E., Wall, S. (2015). Patterns of Attachment: A Psychological Study of the Strange Situation. London: Psychology Press. doi: 10.4324/9780203758045
Arredondo, M. M. (2023). Shining a light on cultural neuroscience: recommendations on the use of fNIRS to study how sociocultural contexts shape the brain. Cultur. Divers. Ethnic. Minor. Psychol. 29, 106–117. doi: 10.1037/cdp0000469
Azhari, A., Bizzego, A., Esposito, G. (2023). Parent–child dyads with greater parenting stress exhibit less synchrony in posterior areas and more synchrony in frontal areas of the prefrontal cortex during shared play. Soc. Neurosci. 1–12. doi: 10.1080/17470919.2022.2162118
Azhari, A., Leck, W. Q., Gabrieli, G., Bizzego, A., Rigo, P., Setoh, P., et al. (2019). Parenting stress undermines mother-child brain-to-brain synchrony: a hyperscanning study. Sci. Rep. 9, 11407. doi: 10.1038/s41598-019-47810-4
Babiloni, F., Astolfi, L. (2014). Social neuroscience and hyperscanning techniques: past, present and future. Neurosci. Biobehav. Rev. 44, 76–93. doi: 10.1016/j.neubiorev.2012.07.006
Belsky, J., De Haan, M. (2011). Annual research review: parenting and children's brain development: the end of the beginning. J. Child. Psychol. Psychiatry. 52, 409–428. doi: 10.1111/j.1469-7610.2010.02281.x
Birditt, K. S., Fingerman, K. L. (2013). “Parent-child and intergenerational relationships in adulthood,” in Handbook of Family Theories: A Content-based Approach, eds M. A. Fine, and F. D. Fincham (London: Routledge), 71–86.
Birk, S. L., Stewart, L., Olino, T. M. (2022). Parent-child synchrony after early childhood: a systematic review. Clin. Child Fam. Psychol. Rev. 25, 529–551. doi: 10.1007/s10567-022-00383-7
Bornstein, M. H., Putnick, D. L., Rigo, P., Esposito, G., Swain, J. E., Suwalsky, J. T., et al. (2017). Neurobiology of culturally common maternal responses to infant cry. Proc. Nat. Acad. Sci. 114, E9465–E9473. doi: 10.1073/pnas.1712022114
Boyum, L. A., Parke, R. D. (1995). The role of family emotional expressiveness in the development of children's social competence. J. Marriage Fam. 57, 593–608. doi: 10.2307/353915
Charles, S. T., Carstensen, L. L. (2010). Social and emotional aging. Annu. Rev. Psychol. 61, 383–409. doi: 10.1146/annurev.psych.093008.100448
Chen, P.-H. A., Qu, Y. (2021). Taking a computational cultural neuroscience approach to study parent-child similarities in diverse cultural contexts. Front. Hum. Neurosci. 15, 703999. doi: 10.3389/fnhum.2021.703999
Clearfield, M. W., Carter-Rodriguez, A., Merali, A.-R., Shober, R. (2014). The effects of SES on infant and maternal diurnal salivary cortisol output. Infant Behav. Dev. 37, 298–304. doi: 10.1016/j.infbeh.2014.04.008
Cole, M. W., Bassett, D. S., Power, J. D., Braver, T. S., Petersen, S. E. (2014). Intrinsic and task-evoked network architectures of the human brain. Neuron 83, 238–251. doi: 10.1016/j.neuron.2014.05.014
Collins, W. A., Steinberg, L. (2006). “Adolescent development in interpersonal context,” in Handbook of Child Psychology: Vol. 3. Social, Emotional, and persoNality Development, ed N. Eisenberg, 6th ed. (Hoboken, NJ: JohnWiley), 1003–1067.
Creaven, A., Skowron, E. A., Hughes, B. M., Howard, S., Loken, E. (2014). Dyadic concordance in mother and preschooler resting cardiovascular function varies by risk status. Dev. Psychobiol. 56, 142–152. doi: 10.1002/dev.21098
Creavy, K. L., Gatzke-Kopp, L. M., Zhang, X., Fishbein, D., Kiser, L. J. (2020). When you go low, I go high: negative coordination of physiological synchrony among parents and children. Dev. Psychobiol. 62, 310–323. doi: 10.1002/dev.21905
Davis, M., West, K., Bilms, J., Morelen, D., Suveg, C. (2018). A systematic review of parent-child synchrony: it is more than skin deep. Dev. Psychobiol. 60, 674–691. doi: 10.1002/dev.21743
DePasquale, C. E. (2020). A systematic review of caregiver–child physiological synchrony across systems: associations with behavior and child functioning. Dev. Psychopathol. 32, 1754–1777. doi: 10.1017/S0954579420001236
Diedrichsen, J., Kriegeskorte, N. (2017). Representational models: a common framework for understanding encoding, pattern-component, and representational-similarity analysis. PLoS Comput. Biol. 13, e1005508. doi: 10.1371/journal.pcbi.1005508
Feldman, R. (2007). Parent-infant synchrony: biological foundations and developmental outcomes. Curr. Dir. Psychol. Sci. 16, 340–345. doi: 10.1111/j.1467-8721.2007.00532.x
Feldman, R. (2015). The adaptive human parental brain: implications for children's social development. Trends Neurosci. 38, 387–399. doi: 10.1016/j.tins.2015.04.004
Feldman, R., Bamberger, E., Kanat-Maymon, Y. (2013). Parent-specific reciprocity from infancy to adolescence shapes children's social competence and dialogical skills. Attach. Hum. Dev. 15, 407–423. doi: 10.1080/14616734.2013.782650
Feldman, R., Greenbaum, C. W., Yirmiya, N. (1999). Mother–infant affect synchrony as an antecedent of the emergence of self-control. Dev. Psychol. 35, 223–231. doi: 10.1037/0012-1649.35.1.223
Feldman, R., Magori-Cohen, R., Galili, G., Singer, M., Louzoun, Y. (2011). Mother and infant coordinate heart rhythms through episodes of interaction synchrony. Infant Behav. Dev. 34, 569–577. doi: 10.1016/j.infbeh.2011.06.008
Feng, X., Shaw, D. S., Skuban, E. M., Lane, T. (2007). Emotional exchange in mother-child dyads: stability, mutual influence, and associations with maternal depression and child problem behavior. J. Fam. Psychol. 21, 714–725. doi: 10.1037/0893-3200.21.4.714
Fingerman, K. L., Pitzer, L., Lefkowitz, E. S., Birditt, K. S., Mroczek, D. (2008). Ambivalent relationship qualities between adults and their parents: implications for the well-being of both parties. J. Gerontol. B. Psychol. Sci. Soc. Sci. 63, 362–371. doi: 10.1093/geronb/63.6.P362
Gumilar, I., Sareen, E., Bell, R., Stone, A., Hayati, A., Mao, J., et al. (2021). A comparative study on inter-brain synchrony in real and virtual environments using hyperscanning. Comput. Graph. 94, 62–75. doi: 10.1016/j.cag.2020.10.003
Han, Z. R., Gao, M. M., Yan, J., Hu, X., Zhou, W., Li, X., et al. (2019). Correlates of parent-child physiological synchrony and emotional parenting: differential associations in varying interactive contexts. J. Child Fam. Stud. 28, 1116–1123. doi: 10.1007/s10826-019-01337-4
Haresign, I. M., Phillips, E. A. M., Whitehorn, M., Goupil, L., Noreika, V., Leong, V., et al. (2022). Measuring the temporal dynamics of inter-personal neural entrainment in continuous child-adult EEG hyperscanning data. Dev. Cogn. Neurosci. 54, 1–11. doi: 10.1016/j.dcn.2022.101093
Hari, R., Henriksson, L., Malinen, S., Parkkonen, L. (2015). Centrality of social interaction in human brain function. Neuron 88, 181–193. doi: 10.1016/j.neuron.2015.09.022
Harrist, A. W., Waugh, R. M. (2002). Dyadic synchrony: its structure and function in children's development. Dev. Rev. 22, 555–592. doi: 10.1016/S0273-2297(02)00500-2
Hasson, U., Frith, C. D. (2016). Mirroring and beyond: coupled dynamics as a generalized framework for modelling social interactions. Philos. Trans. R. Soc. B: Biol. Sci. 371, 20150366. doi: 10.1098/rstb.2015.0366
Hoyniak, C. P., Quiñones-Camacho, L. E., Camacho, M. C., Chin, J. H., Williams, E. M., Wakschlag, L. S., et al. (2021). Adversity is linked with decreased parent-child behavioral and neural synchrony. Dev. Cogn. Neurosci. 48, 100937. doi: 10.1016/j.dcn.2021.100937
Kochanska, G., Aksan, N. (1995). Mother-child mutually positive affect, the quality of child compliance to requests and prohibitions, and maternal control as correlates of early internalization. Child Dev. 66, 236–254. doi: 10.2307/1131203
Kriegeskorte, N., Mur, M., Bandettini, P. A. (2008). Representational similarity analysis-connecting the branches of systems neuroscience. Front. Syst. Neurosci. 4, 2008. doi: 10.3389/neuro.06.004.2008
Kringelbach, M. L., Stark, E. A., Alexander, C., Bornstein, M. H., Stein, A. (2016). On cuteness: unlocking the parental brain and beyond. Trends Cogn. Sci. 20, 545–558. doi: 10.1016/j.tics.2016.05.003
Lee, T. H., Miernicki, M. E., Telzer, E. H. (2017a). Behavioral and neural concordance in parent-child dyadic sleep patterns. Dev. Cogn. Neurosci. 26, 77–83. doi: 10.1016/j.dcn.2017.06.003
Lee, T. H., Miernicki, M. E., Telzer, E. H. (2017b). Families that fire together smile together: resting state connectome similarity and daily emotional synchrony in parent-child dyads. Neuroimage 152, 31–37. doi: 10.1016/j.neuroimage.2017.02.078
Lee, T. H., Qu, Y., Telzer, E. H. (2017c). Love flows downstream: mothers' and children's neural representation similarity in perceiving distress of self and family. Soc. Cogn. Affect. Neurosci. 12, 1916–1927. doi: 10.1093/scan/nsx125
Lee, T. H., Qu, Y., Telzer, E. H. (2018). Dyadic neural similarity during stress in mother–child dyads. J. Res. Adolesc. 28, 121–133. doi: 10.1111/jora.12334
Lee, T. H., Qu, Y., Telzer, E. H. (2019). Neural representation of parental monitoring and links to adolescent risk taking. Front. Neurosci. 13, 1286. doi: 10.3389/fnins.2019.01286
Leong, J. K., MacNiven, K. H., Samanez-Larkin, G. R., Knutson, B. (2018). Distinct neural circuits support incentivized inhibition. Neuroimage 178, 435–444. doi: 10.1016/j.neuroimage.2018.05.055
Leong, J. K., Pestilli, F., Wu, C. C., Samanez-Larkin, G. R., Knutson, B. (2016). White-matter tract connecting anterior insula to nucleus accumbens correlates with reduced preference for positively skewed gambles. Neuron 89, 63–69. doi: 10.1016/j.neuron.2015.12.015
Miller, J. G., Vrtička, P., Cui, X., Shrestha, S., Hosseini, S. M. H., Baker, J. M., et al. (2019). Inter-brain synchrony in mother-child dyads during cooperation: an fNIRS hyperscanning study. Neuropsychologia 124, 117–124. doi: 10.1016/j.neuropsychologia.2018.12.021
Morgan, J. K., Santosa, H., Conner, K. K., Fridley, R. M., Forbes, E. E., Iyengar, S., et al. (2023). Mother–child neural synchronization is time linked to mother–child positive affective state matching. Soc. Cogn. Affect. Neurosci. 18, nsad001. doi: 10.1093/scan/nsad001
Morris, A. S., Criss, M. M., Silk, J. S., Houltberg, B. J. (2017). The impact of parenting on emotion regulation during childhood and adolescence. Child Dev. Perspect. 11, 233–238. doi: 10.1111/cdep.12238
Morris, A. S., Ratliff, E. L., Cosgrove, K. T., Steinberg, L. (2021). We know even more things: a decade review of parenting research. J. Res. Adolesc. 31, 870–888. doi: 10.1111/jora.12641
Ng, J., Xiong, Y., Qu, Y., Cheung, C., Ng, F. F. -Y., Wang, M., et al. (2019). Implications of Chinese and American mothers' goals for children's emotional distress. Dev. Psychol. 55, 2616–2629. doi: 10.1037/dev0000834
Nguyen, T., Schleihauf, H., Kayhan, E., Matthes, D., Vrtička, P., Hoehl, S., et al. (2020). The effects of interaction quality on neural synchrony during mother-child problem solving. Cortex 124, 235–249. doi: 10.1016/j.cortex.2019.11.020
Nguyen, T., Schleihauf, H., Kayhan, E., Matthes, D., Vrtička, P., Hoehl, S., et al. (2021a). Neural synchrony in mother–child conversation: exploring the role of conversation patterns. Soc. Cogn. Affect. Neurosci. 16, 93–102. doi: 10.1093/scan/nsaa079
Nguyen, T., Schleihauf, H., Kungl, M., Kayhan, E., Hoehl, S., Vrtička, P., et al. (2021b). Interpersonal neural synchrony during father–child problem solving: an fNIRS hyperscanning study. Child Dev. 92, e565–e580. doi: 10.1111/cdev.13510
Papp, L. M., Pendry, P., Adam, E. K. (2009). Mother-adolescent physiological synchrony in naturalistic settings: within-family cortisol associations and moderators. J. Fam. Psychol. 23, 882–894. doi: 10.1037/a0017147
Pietromonaco, P. R., DeBuse, C. J., Powers, S. I. (2013). Does attachment get under the skin? Adult romantic attachment and cortisol responses to stress. Curr. Dir. Psychol. Sci. 22, 63–68. doi: 10.1177/0963721412463229
Pietromonaco, P. R., Powers, S. I. (2015). Attachment and health-related physiological stress processes. Curr. Opin. Psychol. 1, 34–39. doi: 10.1016/j.copsyc.2014.12.001
Qu, Y., Fuligni, A. J., Galván, A., Lieberman, M. D., Telzer, E. H. (2016a). Links between parental depression and longitudinal changes in youths' neural sensitivity to rewards. Soc. Cogn. Affect. Neurosci. 11, 1262–1271. doi: 10.1093/scan/nsw035
Qu, Y., Fuligni, A. J., Galván, A., Telzer, E. H. (2015). Buffering effect of positive parent-child relationships on adolescent risk taking: a longitudinal neuroimaging investigation. Dev. Cogn. Neurosci. 15, 26–34. doi: 10.1016/j.dcn.2015.08.005
Qu, Y., Jorgensen, N. A., Telzer, E. H. (2021). A call for greater attention to culture in the study of brain and development. Perspect. Psychol. Sci. 16, 275–293. doi: 10.1177/1745691620931461
Qu, Y., Pomerantz, E. M., Deng, C. (2016b). Mothers' goals for adolescents in the United States and China: content and transmission. J. Res. Adolesc. 26, 126–141. doi: 10.1111/jora.12176
Quiñones-Camacho, L. E., Fishburn, F. A., Camacho, M. C., Hlutkowsky, C. O., Huppert, T. J., Wakschlag, L. S., Perlman, S. B. (2020). Parent-child neural synchrony: a novel approach to elucidating dyadic correlates of preschool irritability. J. Child Psychol. Psychiatry 61, 1213–1223. doi: 10.1111/jcpp.13165
Quiñones-Camacho, L. E., Hoyniak, C. P., Wakschlag, L. S., Perlman, S. B. (2022). Getting in synch: unpacking the role of parent–child synchrony in the development of internalizing and externalizing behaviors. Dev. Psychopathol. 34, 1901–1913. doi: 10.1017/S0954579421000468
Ratliff, E. L., Kerr, K. L., Misaki, M., Cosgrove, K. T., Moore, A. J., DeVille, D. C., et al. (2021). Into the unknown: examining neural representations of parent-adolescent interactions. Child Dev. 92, e1361–e1376. doi: 10.1111/cdev.13635
Reindl, V., Gerloff, C., Scharke, W., Konrad, K. (2018). Brain-to-brain synchrony in parent-child dyads and the relationship with emotion regulation revealed by fNIRS-based hyperscanning. Neuroimage 178, 493–502. doi: 10.1016/j.neuroimage.2018.05.060
Reinero, D. A., Dikker, S., Van Bavel, J. J. (2021). Inter-brain synchrony in teams predicts collective performance. Soc. Cogn. Affect. Neurosci. 16, 43–57. doi: 10.1093/scan/nsaa135
Rogers, C. R., Qu, Y., Lee, T. H., Liu, S., Kim, S. H. (2022). Similarities and discrepancies across family members at multiple levels: Insights from behavior, psychophysiology, and aeuroimaging. Front. Psychol. 12, 831048. doi: 10.3389/fpsyg.2021.831048
Rubin, K. H., Dwyer, K. M., Booth-LaForce, C., Kim, A. H., Burgess, K. B., Rose-Krasnor, L. (2004). Attachment, friendship, and psychosocial functioning in early adolescence. J. Early Adolesc. 24, 326–356. doi: 10.1177/0272431604268530
Santamaria, L., Noreika, V., Georgieva, S., Clackson, K., Wass, S., Leong, V., et al. (2020). Emotional valence modulates the topology of the parent-infant inter-brain network. Neuroimage 207, 116341. doi: 10.1016/j.neuroimage.2019.116341
Selcuk, E., Günaydin, G., Sumer, N., Harma, M., Salman, S., Hazan, C., et al. (2010). Self-reported romantic attachment style predicts everyday maternal caregiving behavior at home. J. Res. Pers. 44, 544–549. doi: 10.1016/j.jrp.2010.05.007
Smith, J. D., Woodhouse, S. S., Clark, C. A., Skowron, E. A. (2016). Attachment status and mother–preschooler parasympathetic response to the strange situation procedure. Biol. Psychol. 114, 39–48. doi: 10.1016/j.biopsycho.2015.12.008
Stark, E. A., Stein, A., Young, K. S., Parsons, C. E., Kringelbach, M. L. (2019). “Neurobiology of human parenting,” in Handbook of Parenting: Biology and Ecology of Parenting, ed M. H. Bornstein (Routledge/Taylor and Francis Group), 250–284. doi: 10.4324/9780429401459-7
Suveg, C., Shaffer, A., Davis, M. (2016). Family stress moderates relations between physiological and behavioral synchrony and child self-regulation in mother–preschooler dyads. Dev. Psychobiol. 58, 83–97. doi: 10.1002/dev.21358
Telzer, E. H., Ichien, N. T., Qu, Y. (2015). Mothers know best: redirecting adolescent reward sensitivity toward safe behavior during risk taking. Soc. Cogn. Affect. Neurosci. 10, 1383–1391. doi: 10.1093/scan/nsv026
Turk, E., Vroomen, J., Fonken, Y., Levy, J., van den Heuvel, M. I. (2022). In sync with your child: the potential of parent-child electroencephalography in developmental research. Dev. Psychobiol. 64, e22221. doi: 10.1002/dev.22221
Ulmer-Yaniv, A., Waidergoren, S., Shaked, A., Salomon, R., Feldman, R. (2022). Neural representation of the parent–child attachment from infancy to adulthood. Soc. Cogn. Affect. Neurosci. 17, 609–624. doi: 10.1093/scan/nsab132
Wang, H., Mai, X., Han, Z. R., Hu, Y., Lei, X. (2018). Linkage between parent-child frontal resting electroencephalogram (EEG) asymmetry: the moderating role of emotional parenting. J. Child Fam. Stud. 27, 2990–2998. doi: 10.1007/s10826-018-1121-5
Wheatley, T., Kang, O., Parkinson, C., Looser, C. E. (2012). From mind perception to mental connection: synchrony as a mechanism for social understanding. Soc. Personal. Psychol. Compass 6, 589–606. doi: 10.1111/j.1751-9004.2012.00450.x
Whittle, S., Simmons, J. G., Dennison, M., Vijayakumar, N., Schwartz, O., Yap, M. B., et al. (2014). Positive parenting predicts the development of adolescent brain structure: a longitudinal study. Dev. Cogn. Neurosci. 8, 7–17. doi: 10.1016/j.dcn.2013.10.006
Keywords: dyadic, family, neural similarity, neuroimaging, parent-child
Citation: Qu Y, Zhou Z and Lee T-H (2023) Parent-child neural similarity: Measurements, antecedents, and consequences. Front. Cognit. 2:1113082. doi: 10.3389/fcogn.2023.1113082
Received: 01 December 2022; Accepted: 10 March 2023;
Published: 29 March 2023.
Edited by:
George R. Mangun, University of California, Davis, United StatesReviewed by:
Joao Guassi Moreira, University of California, Los Angeles, United StatesCopyright © 2023 Qu, Zhou and Lee. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yang Qu, eWFuZ3F1QG5vcnRod2VzdGVybi5lZHU=; Tae-Ho Lee, dGFlaG9sQHZ0LmVkdQ==