 Rachel Torres
Rachel Torres Christina L. Tague
Christina L. Tague Joseph P. McFadden2
Joseph P. McFadden2
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Clim., 27 March 2024
Sec. Climate, Ecology and People
Volume 6 - 2024 | https://doi.org/10.3389/fclim.2024.1280615
This article is part of the Research TopicUrban ecohydrology & hydrometeorology: Integrating water and greenery management to improve climate resilience in citiesView all 6 articles
In Southern California cities, urban trees play a vital role in alleviating heat waves through shade provision and evaporative cooling. Trees in arid to semi-arid regions may rely on irrigation, which is often the first municipal water use to be restricted during drought, causing further drought stress. Finding a balance between efficient water use and maintaining tree health will be crucial for long-term urban forestry and water resources management, as climate change will increase drought and extreme heat events. This study aimed to quantify how urban tree water and carbon fluxes are affected by irrigation reductions, and how that relationship changes with tree species and temperature. We used an ecohydrologic model that mechanistically simulates water, carbon, and energy cycling, parameterized for 5 common tree species in a semi-arid urban area. We simulated a range of irrigation reductions based on average outdoor water use data from the city for a recent extreme drought as well as with warmer temperatures. We then analyzed the response of model outcomes of plant carbon fluxes, leaf area index (LAI), and water use. Results show that reducing irrigation up to 25%, a comparable amount as the California state mandate in 2014, has minimal effects on tree primary productivity and water use efficiency. We found that transpiration was linearly related to irrigation input, which could lead to a short-term loss of evaporative cooling with irrigation reductions during drought. However, primary productivity and LAI had a nonlinear response to irrigation, indicating shade provision could be maintained throughout drought with partial irrigation reductions. Results varied across tree species, with some species showing greater sensitivity of productivity to both irrigation reductions and potentially warmer droughts. These results have implications for water resources management before and during drought, and for urban tree climate adaptation to future drought.
Cities in Southern California face the challenge of adapting to warmer temperatures and heat waves that exacerbate urban heat islands (UHI), and one strategy to combat heat is to increase and maintain urban tree cover. Climate change will cause not only rising temperatures but increased aridity and frequency of drought (Cayan et al., 2010; Cook et al., 2015; Diffenbaugh et al., 2015; McKinnon et al., 2021). Because a large portion of urban water use goes toward outdoor use, the combination of drought and heat waves requires more efficient water use for Southern California to sustain urban water resources for landscape management (Pincetl et al., 2018; Luthy Richard et al., 2020).
Urban trees contribute to climate change adaptation by offering several ecosystem services including stormwater filtration, carbon sequestration, UHI mitigation, and psychological benefits for residents (Bolund and Hunhammar, 1999; Livesley et al., 2016; McPherson et al., 2017; Sudimac et al., 2022). However, for trees to provide cooling they require adequate water, which becomes limited during drought because of scarcity and conservation mandates on outdoor water use (Gober et al., 2012; Yang and Wang, 2017; Gao et al., 2020).
Outdoor water use is a significant fraction of total water use in residential areas, estimated to be between 18–35% in Southern California (Gleick et al., 2003), and up to 50% in some areas of Los Angeles (Mini et al., 2014). Because it takes up a substantial portion of the urban water balance, outdoor water use efficiency has been studied throughout Southern California. Outdoor water use has been quantified through estimating evapotranspiration of trees and turfgrass using portable chambers in the Los Angeles region (Pataki et al., 2011; Litvak et al., 2017b). Other tools for estimating outdoor water use include using water billing data, remote sensing, or monitoring reference evapotranspiration with adjustments for plant types (Nouri et al., 2013; Mini et al., 2014).
Vegetation outdoor water use varies by vegetation type and ecophysiologic traits. Irrigation has shown to be a main driver of lawn transpiration (Bijoor et al., 2014; Litvak and Pataki, 2016; Litvak et al., 2017a). While some urban trees, particularly those in larger urban green spaces may rely primarily on precipitation inputs, many urban trees also receive substantial irrigation water. Irrigation inputs have caused some urban trees in Los Angeles to acclimate and rely on irrigation along with natural water sources (Bijoor et al., 2012; Chen et al., 2015). Tree water use and drought tolerance depend on not only how much irrigation is applied but also species ecophysiological traits, including regulation of stomatal conductance in response to the environment (McDowell et al., 2008; Ibsen et al., 2023). Drought tolerant species or native species may require less irrigation. Phenology also affects annual water use as leaf area affects transpiration, for example deciduous trees decrease transpiration in the winter.
While it is important to quantify vegetation water use to improve urban water efficiency, fully accounting for the value of outdoor water use should include the indirect effects of ecosystem services provided by trees. In semi-arid climates, outdoor water contributes to UHI mitigation through maintaining healthy tree canopy cover for shade provision and increasing latent heat from evapotranspiration (Gillner et al., 2015; Winbourne et al., 2020). Studies have used atmospheric coupled land surface models to demonstrate the effects of irrigation on cooling the microclimate in semi-arid cities (Yang et al., 2015; Vahmani and Ban-Weiss, 2017; Broadbent et al., 2018; Reyes et al., 2018; Gao et al., 2020). For urban parks in a variety of climates, it's been shown increasing leaf area index (LAI), and therefor shade, is an effective way to reduce heat stress at the surface (Brown et al., 2015; Rahman et al., 2020). Because of the ability to reduce heat stress on people, tree maintenance should be considered along with water use restrictions, especially when heat waves coincide with drought. However, balancing both maintaining tree ecosystem services and conserving water may lead to trade-offs.
Studies on outdoor water conservation mandates during a recent California drought have focused on the effectiveness at saving water (Liang et al., 2017; Palazzo et al., 2017; Quesnel and Ajami, 2019), and others have looked at effects on social perceptions (Gonzales and Ajami, 2017; McCumber, 2017), temperature (Vahmani and Ban-Weiss, 2017; Allen et al., 2021), and disease vector abundance (Bhattachan et al., 2021). A few studies have looked at the effects on greenness (Quesnel et al., 2019; Allen et al., 2021; Miller et al., 2022) but did not consider the indirect effects on urban tree carbon and water fluxes.
This study examines the role of irrigation in maintaining urban tree productivity during the 2012–2016 California drought in Santa Barbara, California, a coastal semi-arid city. This drought was classified as “exceptional,” with drought indicators below the 2nd percentile, by the U.S. Drought Monitor (Diffenbaugh et al., 2015). This event was one of the most severe droughts in a millennium for California (Griffin and Anchukaitis, 2014). In 2014, extreme warm temperatures coincided with low precipitation, further straining water resources (AghaKouchak et al., 2014; Lund Jay et al., 2018). In 2015, California set a mandatory 25% statewide water use reduction, with urban water districts assigned to different amounts ranging from 4–36% (Exec. Order No. B-29-15, 2015). In this study we use an ecohydrologic model to quantify tradeoffs between water conservation and tree ecosystem services during drought. We use the 2012-2016 as a salient example of both drought climatology and expected water reductions. Since future droughts may coincide with more extreme warm temperatures (Cook et al., 2015; Diffenbaugh et al., 2015), we also consider how these tradeoffs might change for future warmer droughts. We ask the following two questions:
1. How do reductions in irrigation during drought affect tree ecosystem services during and following the drought?
2. How do the these relationships differ under a drought with higher temperatures?
Using an ecohydrologic model, we simulated tree response to historic drought conditions, and a drought scenario with similar precipitation but warmer temperatures, under varied irrigation amounts for select urban tree species in a coastal semi-arid city. We analyzed the effects of reduced irrigation on model outputs of transpiration, net primary productivity (NPP), plant water use efficiency (WUE), and leaf area index (LAI). These model outputs are indicators of ecosystem processes driving the subsequent provisioning, regulating, and cultural ecosystem services (Sun et al., 2017).
We used the Regional Hydro-Ecologic Simulation System (Tague and Band, 2004), RHESSys, to simulate vegetation ecosystem services with varying scenarios of water input. RHESSys and its hydrologic and carbon cycling sub-models have been validated through comparison with observed streamflow (Shields and Tague, 2012), flux towers (Zierl et al., 2007), remote sensing of vegetation growth, sap flow (Tsamir et al., 2019), tree rings (e.g., Vicente-Serrano et al., 2015) and other observations. RHESSys was previously used in semi-arid urban watersheds for analyzing hydrologic parameter uncertainty on evapotranspiration (Shields and Tague, 2012), estimating the effects of urban development on runoff (Tague and Pohl-Costello, 2008), and assessing parameter uncertainty in urban trees and their drought resilience (Torres, 2024).
RHESSys simulates carbon, water, energy, and nutrient cycling at several spatial scales. Thus RHESSys simulates both daily time series of hydrologic variables, including soil moisture, evaporation and transpiration along with plant responses, including stomatal conductance, short term carbon assimilation and its impact of year to year changes in tree biomass, height and LAI. In RHESSys, a watershed is divided into patches, which vary by topography. For our study, we use a single patch to represent a localized area with a single tree species, at 10 m2 resolution, with only vertical hydrologic flow. Vertical water inputs include precipitation which can be intercepted by the canopy, infiltrate into the subsurface, or create overland flow. Irrigation is added to any precipitation that infiltrates into the top layer of soil. Soil subsurface layers include the rooting zone defined by the vegetation rooting depth, an unsaturated layer, and a saturated layer. Water flowing out of the saturated layer goes to deeper groundwater storage. Vegetation evapotranspiration is calculated daily using Penman-Monteith (Monteith, 1965). Vapor pressure deficit for the Penman-Monteith is estimated based on minimum and maximum daily temperatures. Soil evaporation is based on energy, atmospheric conditions, and soil moisture. We emphasize that because RHESSys computes daily evapotranspiration as a function of atmospheric conditions, and available moisture, it implicitly accounts for time-varying irrigation demand.
Vegetation carbon cycling in the model is driven by daily estimates of photosynthesis through the Farquhar model, where the rate of carbon gain per unit LAI depends on available water, nutrients, and light (Farquhar and von Caemmerer, 1982). Vegetation maintenance and growth respiration depends on these environmental factors and temperature (Ryan et al., 1997), and net primary productivity (NPP) is calculated from these two fluxes as the difference between gross photosynthesis and plant respiration. Assimilated carbon gets allocated to either the leaves, stems, coarse roots, or fine roots, based on vegetation parameters. The amount allocated uses a semi-mechanistic approach that accounts for resource limitations and LAI (Landsberg and Waring, 1997; Dickinson et al., 1998).
Vegetation parameters in RHESSys play an important role for differentiating tree species carbon cycling and water use. This current study builds on a previous study that calibrated species-specific vegetation parameters using remote sensing data in Santa Barbara (Torres, 2024). In the prior study, remote sensing products, including estimated LAI (Alonzo et al., 2016) and changes to the normalized differential vegetation index (NDVI) during drought (Miller et al., 2020) at the tree species level were used to constrain and select parameters. Calibration and parameter sensitivity analysis included vegetation parameters that control ecophysiological mechanisms including carbon allocation, phenology, carbon turnover, and stomatal function. This prior study found that the dominant parameters that affect tree species responses to water availability include maximum stomatal conductance, vapor pressure deficit at stomatal closure, and leaf water potential at stomatal closure. A full list of parameters is included in the Supplementary material. In this current study, parameter uncertainty was maintained for each tree species by using a subset of 50 parameter sets that gave reasonable performance from Torres (2024). Uncertainty in parameter estimates can arise from multiple sources, including within species variation in tree responses in trees sampled in Miller et al. (2020) and low sensitivity of variables used for calibration to underlying parameters. By running all model scenarios with multiple parameter sets for each tree species we account for within species trait variation. We acknowledge that this approach cannot fully account for spatial heterogeneity in species traits given uncertainty in remote sensing data (as well as limited spatial coverage). Nor does this approach account for potential longer term adaptation of species traits with climate change (Ibsen et al., 2023). Nonetheless it allows us to account for observed species differences from the Santa Barbara setting.
RHESSys also includes soil and subsurface parameters that influence drainage rates and water storage capacity. For parameters that control subsurface drainage rates we use a previous study that calibrated to streamflow in the same watershed (Chen, 2016). For storage related parameters, we consider that different species may be preferentially associated with particular rooting zone storage capacity. To account for this we follow the same apporach used to select vegetation parameters by using (Torres, 2024) that included species specific soil storage parameters in its calibration. As with vegetation parameters, while this approach does represent differences in soil conditions associated with particular tree species in the study site, it cannot fully account for local heterogeneity that may be associated with soil hydrologic properties in urban settings. We also note that the remote sensing data used for calibration focused largely on urban trees in urban green space rather than street trees (Miller et al., 2020). Street trees may have substantially different soils conditions due to site preparation (Ghosh et al., 2016; Grabosky and Bassuk, 2016).
For each parameter set, RHESSys is used to spinup vegetation carbon and nitrogen stores along with structural characteristics, including LAI. Table 1 summarizes these initial conditions. Climate data used for spinup is described below.
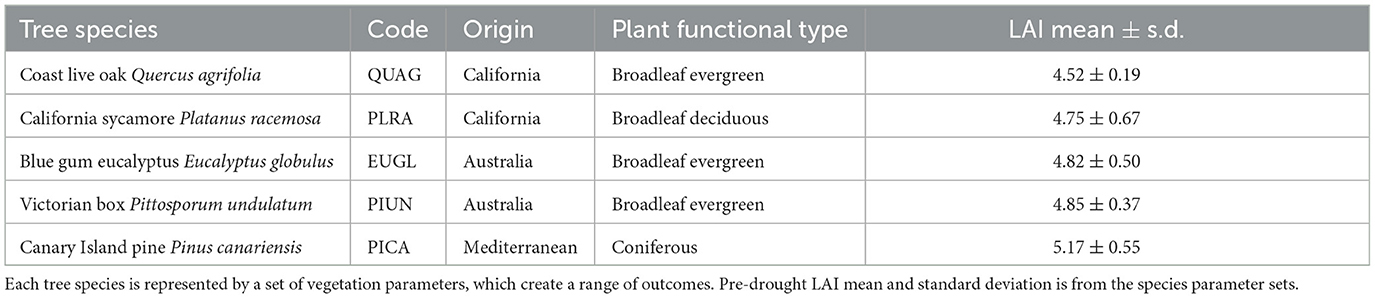
Table 1. Tree species used in model scenarios.
The tree species that were parameterized are Coast Live Oak (Quercus agrifolia), California Sycamore (Platanus racemos), Blue Gum Eucalyptus (Eucalyptus globulus), Victorian Box (Pittosporum undulatum), and Canary Island Pine (Pinus canariensis). These species are among the most common, non-palm trees, in the study area, as documented by Alonzo et al. (2014). They are representative of native, non-native, deciduous, and evergreen trees (Table 1).
These species are a part of a diverse urban forest located in Santa Barbara, California, a coastal city with a Mediterranean climate comprised of hot, dry summers and wet winters. The average rainfall in the last 50 years is 482 mm. In 2012–2016 there was a multiyear drought that became “exceptional” in the U.S. Drought Monitor scale in Santa Barbara county in 2014 (Diffenbaugh et al., 2015). The same year, the City of Santa Barbara declared a “Stage One” drought and asked residents to reduce water consumption by 20%. In 2015 the state mandated that Santa Barbara cut 16% of water consumption (Exec. Order No. B-29-15, 2015) .
The model study period includes water years 2011–2017, with 2011 and 2017 identified as non-drought years (annual average is 699 mm) and 2012–2016 as drought years (annual average is 258 mm). Several climate datasets were used as an input for the model. The first is a temperature dataset from NOAA (station USC00047902) that includes daily maximum and minimum temperatures. Daily precipitation is from a rain gauge located in downtown Santa Barbara (station ID 234), from Santa Barbara County Public Works Water Resources. The model goes through a “spin up” period to establish soil and vegetation carbon and nitrogen stores for each tree species, and this uses observed climate data from 1940-2010. Model outputs are then taken from 2011–2017. During the drought period 2012–2016, which was considered exceptionally severe due to low precipitation coinciding with high temperatures (AghaKouchak et al., 2014), observed annual average maximum temperature was 23.3° in 2014 and annual average minimum temperature was 13.1° in 2014. Long term annual average maximum and minimum temperatures are 21.2° and 11.1° respectively.
Along with the observed drought, we simulated how this drought would affect urban trees if it were to occur in a future climate with potentially warmer temperatures. The climate change scenario inputs have the same precipitation data, with both minimum and maximum temperatures increased by 1.8°. We do this to isolate the impact of a temperature increase. This increase is based on analyzing downscaled climate model projections for the city of Santa Barbara from the tool Cal-Adapt (Pierce et al., 2014). The projections show an overall trend of warming, with increased precipitation variability in the Santa Barbara region. To determine potential temperature changes during future drought years, we looked at the temperature difference in years from the RCP8.5 projections with precipitation anomalies similar to the 2012–2016 drought. The mean annual positive temperature difference was about 1.8°. Thus a 1.8° warmer drought will be at expectation for higher emission scenarios (RCP8.5). We note, however, that this temperature increase is also likely to occur for some droughts even with more moderate emissions scenarios. We also note that a 1.8° is within range of temperature changes by 2070 for this region as reported in the California's Fourth Climate Change Assessment compared to historic temperatures from 1961–1990 (Langridge, 2018).
Monthly irrigation data for the years 2010–2020 were obtained from the City of Santa Barbara Water Resources landscaping accounts, which include large landowners, private residences, and businesses (Water Resources, 2010-2020). To protect the privacy of residents and account owners, the individual accounts were aggregated to a net monthly irrigation amount, which was then normalized by the aggregate area to get the average monthly amount per unit area. The annual irrigation inputs ranged from 331–566 mm in the baseline scenario (Figure 1).

Figure 1. Water inputs to the model include rainfall, shown as gray columns, and irrigation, shown as colored lines. Years when water conservation mandates were set are marked with vertical lines, one for the city of Santa Barbara, and a second for the state of California. The yellow line displays the trend of baseline irrigation if it was not reduced with the conservation mandates. The baseline scenario is the monthly outdoor water use provided by the city.
Monthly irrigation was disaggregated to weekly values using the R package tempdisagg (Sax and Steiner, 2013), which works similar to a linear regression model to transform a low frequency time series to a higher frequency series. Weekly values were then split into two so that irrigation input occurred twice per week. Input occurred twice per week during the night, following previous modeling studies that included urban irrigation as an input (Vahmani and Hogue, 2014; Yang et al., 2015; Reyes et al., 2018). For this study, the focus is on effects due to reductions to total annual input. In the baseline scenario before the conservation mandates, the average daily input was 4.5 mm, with an average of 7mm during summer months. For reference, the average daily reference evapotranspiration was 3.1 mm and maximum was 6.7 mm for the California irrigation management information system (CIMIS) station in downtown Santa Barbara (station 107, California Deptartment of Water Resources, 2021).
With a baseline scenario from the monthly data from the city, we created 5 scenarios: baseline irrigation and with baseline irrigation reduced by 25%, 50%, 75%, and 90% for all years of the simulation. This created scenarios with average annual irrigation input during the study years of: 424 mm, 318 mm, 212 mm, 106 mm, and 42 mm. The baseline data does include a period of time with mandatory state water restrictions, beginning in 2014 with a proclamation of a state of emergency and advancing in April 2015 to a statewide 25% reduction in potable urban usage. Irrigation scenarios were input as reductions for all years, including years following the mandate (Figure 1).
It should be noted that the water use data used in this study was from a water resources account that represents large areas of private land. While it does reflect the seasonal changes to water use and the monthly amounts, it is specific to Santa Barbara, and the amounts may be above or below average for urban outdoor water use in Southern California. Despite this potential bias, the annual irrigation rates used are comparable to the higher amounts estimated for outdoor water use rates in Los Angeles, California (Mini et al., 2014).
Model simulations included the five tree species with 50 permutations of vegetation parameters to account for parameter uncertainty, two drought scenarios, and 5 irrigation scenarios (Table 2), ran over the same time period, 2011–2017.
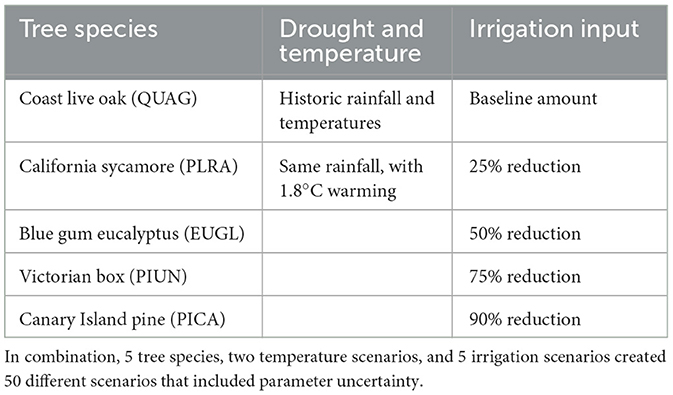
Table 2. Model simulation set up, with each column displaying a variable in the simulations.
We used RHESSys model outputs of transpiration, NPP, tree-level water use efficiency (WUE), and LAI to test for differences between irrigation scenarios over the course of the 2012–2016 drought. WUE was calculated as the ratio of annual kgC of tree NPP to annual mm of tree transpiration. RHESSys runs at a daily time step, and outputs were aggregated to the total amount for transpiration, NPP, and WUE, and the average LAI over each water year. While we did not explicitly convert output of transpiration to latent energy or changes in temperature, we consider it as an estimate of plant water use and as a proxy for potential evaporative cooling above the canopy. LAI resilience is the ratio of post-drought LAI to pre-drought LAI, following a previous study that analyzed how LAI resilience for the same tree species varied with altered climate conditions (Torres, 2024).
After aggregating RHESSys daily outputs to annual values for each irrigation reduction and temperature scenario, we statistically compared the rate of change in each outcome with irrigation reduction to compare across species and temperature. We also compared across drought years (2012–2016) and non-drought years (2011, 2017). For transpiration, we ran a linear regression to compare the rate of increase in transpiration with irrigation input. For LAI resilience and WUE, the linear regression was performed paired with a post-hoc Tukey's HSD to check for significance in difference in means of the outcome with reduced irrigation compared to the baseline irrigation. A piece-wise linear regression was used to determine changes in annual NPP with irrigation input during drought years. We used the R package “segmented” (Muggeo, 2008) to test for the existence of breakpoints and to compare rates of change in NPP before and after the breakpoints.
During the drought period with historic temperatures and baseline irrigation, average annual transpiration varied between 407–502 mm across all tree species parameter sets, compared to 462–713 mm in the non-drought years (Figure 2). Transpiration under a warmer climate had similar ranges, suggesting tree transpiration in Santa Barbara is more limited by water availability than temperature and vapor pressure deficit. Water limited transpiration is also apparent in comparing how mean annual transpiration varies with irrigation input during drought and non-drought years. Transpiration increased with more irrigation applied, but the rate of increase differed for drought and non-drought years (Figure 2). The less irrigation there is, the greater the difference in transpiration between a drought and non-drought year. For example, in comparing the means of the groups of irrigation amounts, under a non-drought year the difference between baseline irrigation and a 90% decrease was 130 mm/yr, but during a drought year the average difference between these groups was 350 mm/yr. The linear slopes for change in annual transpiration with irrigation input are shown in Table 3.

Figure 2. Annual transpiration for each irrigation scenario, with boxplots showing tree species parameter uncertainty. Drought years include 2012–2016, and non-drought are years before and after drought. Horizontal dashed lines show average annual transpiration for baseline irrigation of each tree and climate scenario. (A–E) Show the drought scenario with actual observed temperatures, and (F–J) show the same drought with warmer temperatures. Columns show each tree species: (A, F) Eucalyptus globulus; (B, G) Pinus canariensis; (C, H) Pittosporum undulatum; (D, I) Platenus racemosa; (E, J) Quercus agrifolia.
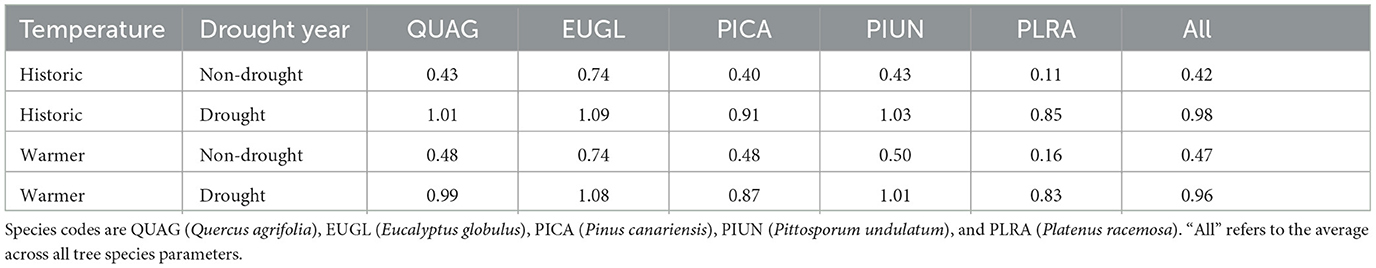
Table 3. Slopes of relationship between annual transpiration and irrigation input for each tree species and temperature scenario, also separated by drought (2012–2016) and non-drought years (2011, 2017) in our simulation.
The change in transpiration with irrigation reductions depends on not only the amount of total water input but also on the tree species. Tree species variation in transpiration depends on key parameters that control stomatal behavior and leaf lifespan. During drought years there is less variation in slope between species relative to non-drought years. The tree species with the lowest slope is the only deciduous tree (PLRA), while the highest slope is the blue gum eucalyptus tree (EUGL) (Table 3). For most of the tree species, the rate of change in transpiration with irrigation reduction doubles during a drought year, with the exceptions of PLRA, which had a greater difference, and EUGL which had less of a difference. The variation in slopes between drought years and non-drought years is more substantial than the variation in slopes between the historic temperature and warmer temperature scenarios.
Across tree parameter sets in the baseline irrigation scenario with the historic climate, average annual NPP during the drought ranged from 0.61–1.09kgC/m2, compared to 0.56–1.21kgC/m2in a non-drought year. These average values with baseline irrigation are shown in Figure 3 as dashed horizontal lines. This range is reduced to averages of 0.23–0.70 kgC/m2 during drought and 0.48–1.31 kgC/m2 in non-drought years with the maximum irrigation reductions of 90%.
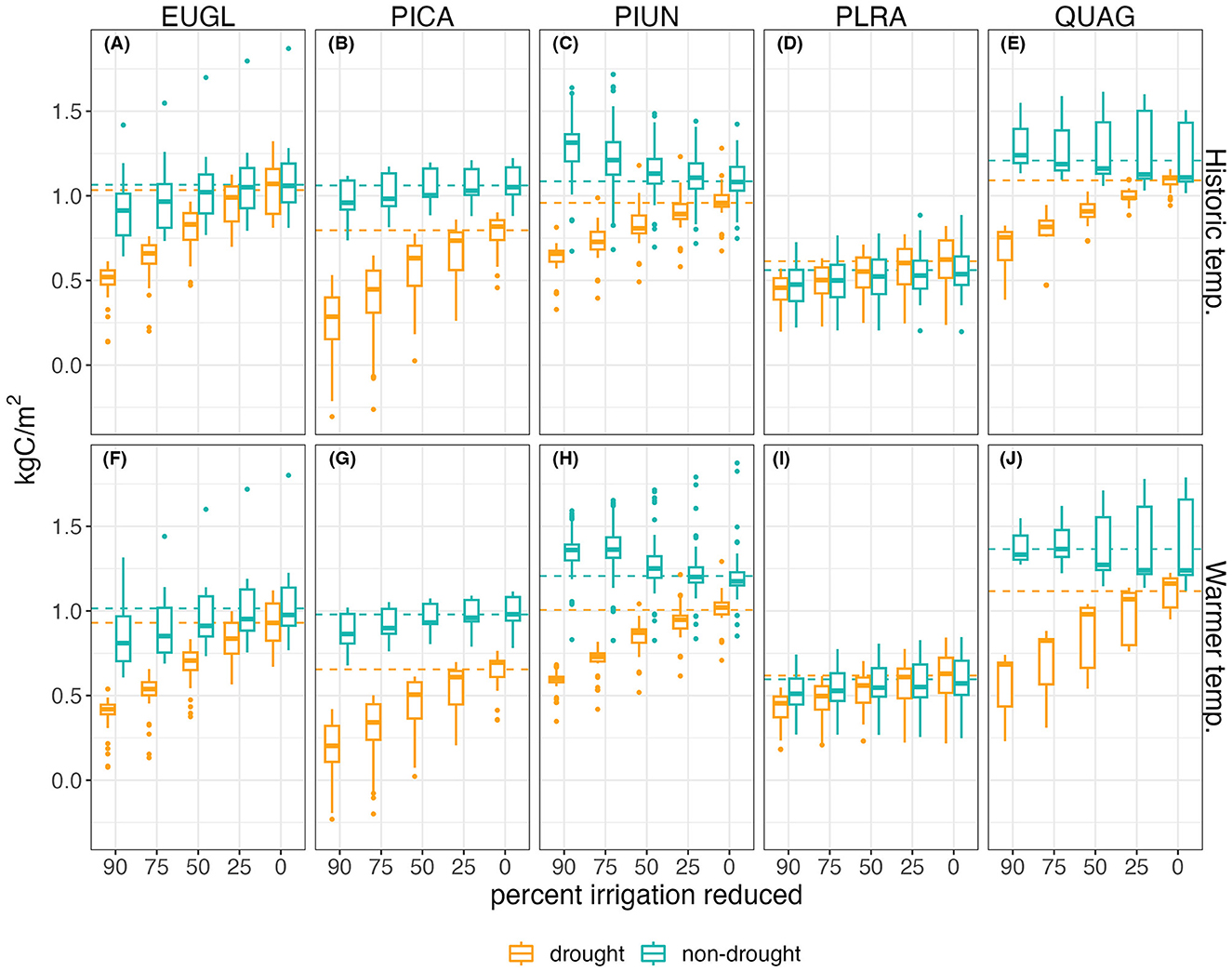
Figure 3. Annual net primary productivity (NPP) for each irrigation scenario, with boxplots showing tree species parameter uncertainty. Drought years include 2012–2016, and non-drought are years before and after drought. The horizontal dashed line shows the average annual amount for each tree species and climate with baseline irrigation. (A–E) Show the drought scenario with actual observed temperatures, and (F–J) show the same drought with warmer temperatures. Columns show each tree species: (A, F) Eucalyptus globulus; (B, G) Pinus canariensis; (C, H) Pittosporum undulatum; (D, I) Platenus racemosa; (E, J) Quercus agrifolia.
While annual transpiration declines linearly with decreased irrigation, annual NPP declines at a non-linear rate during drought years (Figure 3). When is not a drought year, there is less of a decline, and not a significant difference in mean NPP between baseline and irrigation reduction of –25%. Using a break point analysis in a piece-wise linear regression we identified the point in the relationship of average annual NPP to average irrigation input during a drought year. This point occurred between the 25% to 50% irrigation reduction scenarios. From baseline irrigation to the breakpoint reduction during a drought year, the rate of decline in average annual NPP with irrigation has a gradual slope between 0.42–1.69kgC/m2/mm depending on tree species. Beyond reductions of 25–50%, this rate can double or more to a range of 0.93–3.12 kgC/m2/mm (Figure 4). Species differences are greater for irrigation reductions above 25%, with PLRA showing relatively small declines in NPP. With warming, the slope of baseline irrigation to the breakpoint is similar to the slope for the historic temperature drought, with similar patterns across tree species and tree parameter uncertainty. However, when irrigation is further reduced, species respond differently to the warmer drought. The evergreen broadleaf trees QUAG and PIUN had higher slopes with warming, while the evergreen coniferous tree, PICA, slope decreased with warming. The deciduous tree, PLRA, had the lowest average slope for both temperature scenarios.
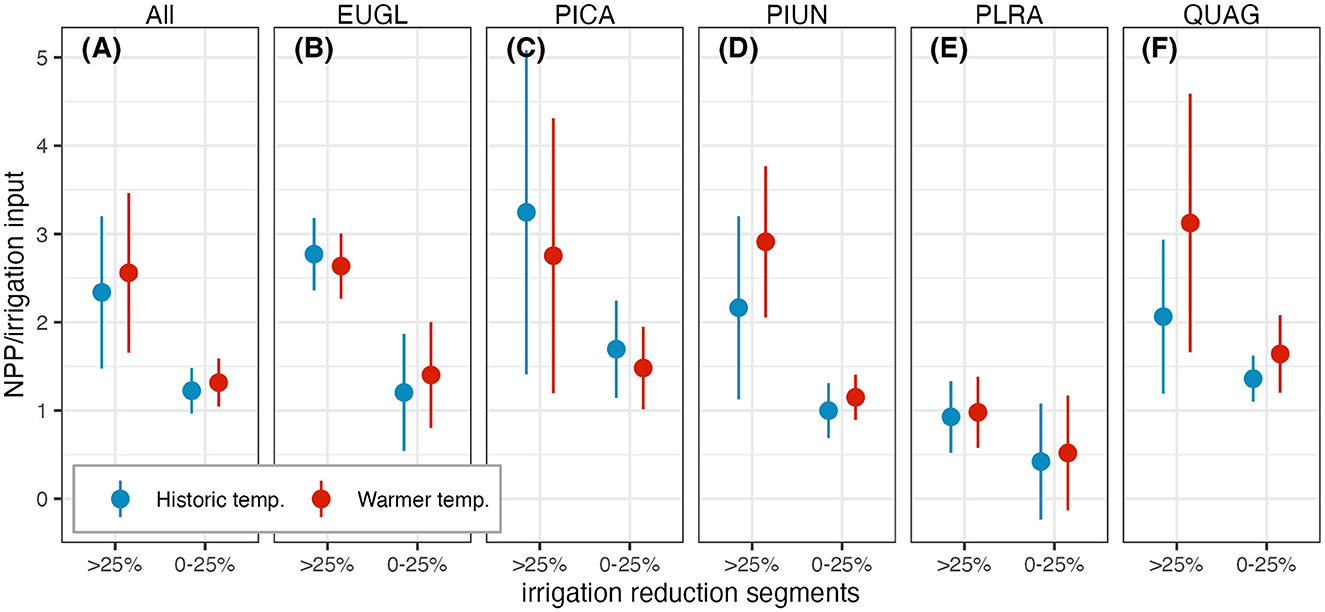
Figure 4. For a piece-wise linear relationship between annual net primary productivity (NPP) and irrigation reduction during drought years, two slopes were calculated. “Pre” shows the slope before the breakpoint, between 25–50%, and “post” is the slope after the breakpoint. The y-axis shows the slope and 95% confidence intervals for both segments. Colors represent temperature, where blue is historic and red is warming. Box (A) shows the relationship for the average NPP across all tree types. Boxes (B–F) show tree species: (B) Eucalyptus globulus; (C) Pinus canariensis; (D) Pittosporum undulatum; (E) Platenus racemosa; (F) Quercus agrifolia.
Water use efficiency (WUE), quantified here as the amount of tree NPP per amount of transpiration on average shows an increase during the drought (Figure 5). Across all tree parameter sets, average WUE ranged from 0.67 to 3.45 in the baseline drought years, and 0.73–2.85 during the non-drought years. There is steeper linear decline in transpiration (Figure 2) relative to a more gradual nonlinear decline in NPP (Figure 3), causing WUE to increase with decreasing irrigation. WUE increased on average with less irrigation, but with variation across tree species. For EUGL and PICA during drought years, average WUE increases under the scenarios of irrigation reduction from 25–50%, but then declines under higher reductions. For the other tree species parameter sets, WUE continues to increase under greater irrigation reductions. For two of the evergreen species, QUAG and PICA, irrigation reduction from 75–90% under warming has a greater average WUE during the non-drought years than the drought years. PICA however does have some large outliers in lower irrigation scenarios, due to some parameter sets resulting in negative annual NPP.
Figure 5. Water use efficiency (WUE) as tree species annual NPP divided by tree transpiration, for each tree species, temperature scenario, and drought and non-drought year. Variation for each tree species shows parameter uncertainty. Horizontal dashed lines show average value for baseline irrigation. (A–E) Show the drought scenario with actual observed temperatures, and (F–J) show the same drought with warmer temperatures. Columns show each tree species: (A, F) Eucalyptus globulus; (B, G) Pinus canariensis; (C, H) Pittosporum undulatum; (D, I) Platenus racemosa; (E, J) Quercus agrifolia.
LAI resilience is calculated as the ratio of post-drought LAI to pre-drought LAI. Similar to patterns of NPP, LAI drought resilience declines at a greater rate with irrigation reductions above 25%–50%. The average LAI resilience across tree species for the baseline scenario was 0.97 and 0.95 with warming, and this declines to 0.96 and 0.94 at 25% irrigation reduction—not statistically significant differences for both current climate and warmer temperatures. Beyond 25%, the LAI resilience declines to as low as 0.46 with 90% reduced, and 0.37 with warming (Figure 6). Species showed substantial differences in the loss of resilience with irrigation reductions, although for historic drought, all show resilience close to 1 for full irrigation, and most show only minor reductions in resilience for 25% reductions. EUGL shows greatest declines in resilience with irrigation reduction, while PIUN maintains resilience even with 90% reduction under the warmer drought. During the drought years, all tree species had a greater loss of canopy mid-drought with irrigation reductions greater than 50% compared to the baseline irrigation.

Figure 6. LAI resilience, the ratio of post-drought LAI to pre-drought LAI, for each tree species and irrigation scenario. Variation for each tree species shows parameter uncertainty. Horizontal line is shown at 1 for reference, where 1 is a return to pre-drought LAI. (A–E) Show the drought scenario with actual observed temperatures, and (F–J) show the same drought with warmer temperatures. Columns show each tree species: (A, F) Eucalyptus globulus; (B, G) Pinus canariensis; (C, H) Pittosporum undulatum; (D, I) Platenus racemosa; (E, J) Quercus agrifolia.
For the baseline irrigation scenario, model outputs of annual tree transpiration, annual NPP, WUE, and LAI are comparable with other studies. With observed historic temperatures, annual transpiration for all tree species averaged 450 mm/year, which is comparable to in-situ studies that measured urban tree transpiration in coastal Southern California using sap-flux measurements and empirical models, some of which include the same species in our study, Canary Island Pine (PICA) and California Sycamore (PLRA) (McCarthy and Pataki, 2010; Pataki et al., 2011; Litvak et al., 2017a,b). The tree species with the highest estimated annual transpiration rates were from the blue gum eucalyptus (EUGL). The lowest annual transpiration rates during drought was the coniferous tree, the Canary Island Pine (PICA), which agrees with lower daily values observed in a Los Angeles field study of tree evapotranspiration (Litvak et al., 2017a) .
Vegetation carbon fluxes and carbon storage in cities are challenging to quantify due to limited data and lack of transferability from natural vegetation models (McHale et al., 2009; Zhuang et al., 2022). The average annual NPP across all species for the baseline irrigation was higher than field studies of natural forests (Malone et al., 2016; Alexander et al., 2023), which may reflect irrigation inputs as well as differences in soil and species adaptations in urban microclimates. Across the range of model estimates of annual NPP with parameter uncertainty, the lower values were within range of comparable studies that estimated vegetation productivity in arid to semi-arid cities (Nowak et al., 2013; Shields and Tague, 2015; McHale et al., 2017), as well as for a sub-humid city (Mngadi et al., 2022). Tree species differed in NPP response to irrigation reductions, with some having a steep slope for irrigation reductions beyond the breakpoint of 25–50% (EUGL, PICA, PIUN), and others having a more gradual slope (PLRA, QUAG). Species also differed in how the slopes changed with a warmer temperature. For example, QUAG, the coast live oak, the slope increased with warming, while for PICA, the conifer, the slope decreased with warming (Figure 4).
With baseline irrigation, for all tree species the average WUE increased during drought years. This is in agreement with Malone et al. (2016), who found an increase in WUE of natural vegetation in arid to semi-arid regions of California during 2012-2016 drought. There are fewer studies of WUE in semi-arid urban settings, but in more temperate climates WUE also increased when trees were water limited (Fu et al., 2020; Grossiord et al., 2020; Horike et al., 2023). For an irrigated setting in Los Angeles, California, shrub species from arid regions had a smaller WUE compared to species from temperate regions due to the availability of excess water from irrigation (Goedhart and Pataki, 2012). Increased WUE with low irrigation rates relative to higher irrigation rates was also found in an urban arid setting (Stabler, 2008). In our study two of the tree species (QUAG, PIUN) experienced this pattern for both drought and non-drought years.
All tree types maintained average LAI throughout the drought period with baseline irrigation. LAI can be compared to the remote sensing index equivalent water thickness (EWT) due to the relationship between canopy level water content and LAI (Roberts et al., 2004). Model simulations of constant average annual LAI for the baseline temperature and irrigation scenario are in agreement with observed EWT for a similar time period in Santa Barbara done by Miller et al. (2022), who found that tree canopy varied with seasonal summer drought, but the peak values in the spring were similar across years 2013–2015.
To determine whether trade-offs with irrigation reduction exist, we compared model outputs across scenarios of irrigation input for each tree species and temperature. For all tree species, annual transpiration was linearly related to water input. In drought years, the slope nearly doubles compared to non-drought years for most of the tree species (Table 3). The steeper slope shows a greater loss of transpiration with smaller amounts of irrigation reduction, which would result in short-term loss of evaporative cooling during drought. In other words, the importance of irrigation for maintaining evaporative cooling increases during drought. This is notable given that in semi-arid climates even non-drought years have long periods during the summer with little or no precipitation inputs. With warmer temperatures, the slopes increased in non-drought years relative to the historic temperatures for all tree species except EUGL, as shown in Table 3. The increased slope represents a greater sensitivity to water input under warmer temperatures. This may reflect the higher water demand due to increased vapor pressure deficit, which is expected to increase transpiration in the future (Kirschbaum, 2000; Marchin et al., 2016; Grossiord et al., 2017).
Despite reductions in transpiration with irrigation reduction, annual NPP had a nonlinear response to the irrigation reduction scenarios. This is partially due to the effects of water limitations on the seasonal timing and amount of productivity. With 25% less irrigation than the baseline amount and observed temperatures, there was not a statistically significant difference in annual NPP or LAI resilience across all tree species, suggesting that over-watering was occurring before the drought. Maintaining adequate water supply during summer is costly even in non-drought years. Thus efforts toward general moderate (25%) reductions in irrigation during non-drought years may be worthwhile in the future (Wilson et al., 2016). While there would be some reduction in evaporative cooling (e.g., transpiration) these are small for non-drought years and changes in ecosystem services related to carbon cycling and shading would be minimal.
In comparing the rates of change between annual NPP and irrigation input during drought, the smaller slope from baseline irrigation to the breakpoint suggests a gradual loss of NPP with small amounts of irrigation reduction could occur while still maintaining productivity, which could aid in shade provision. If the goal is to maintain a resilient tree canopy post-drought, but prioritize water conservation during a drought, irrigation reductions up to 50% from the baseline will cause loss of NPP and LAI during the drought, but can maintain resilient LAI post-drought. For reductions greater than 50%, we found there would be multi-year effects in NPP and LAI— trees would have less recovery post-drought and there may be structural loss and low to negative amounts of productivity during the drought. There are notable differences across species, and these are most apparent when irrigation reductions are greater than 50%. Species such as PLRA, the California Sycamore typically found in riparian areas, may tolerate more severe watering restrictions, although LAI resilience does declines with 75% or more irrigation reduction for warmer droughts. The relative small declines in resilience and NPP for irrigation reductions < 50% for all species are useful for informing broad watering restrictions; while species differences can be used to guide longer term decisions about landscaping and tree selection for climate change (Esperon-Rodriguez et al., 2022).
The different responses in tree water use and NPP caused an increase in annual water use efficiency (WUE) during drought compared to non-drought years, across tree species and temperature scenarios (Figure 5). The increased WUE during drought happens for each irrigation scenario for all tree species except PICA, the coniferous tree. Across all tree species, compared to the baseline irrigation, average WUE also increases with 25% irrigation reduction, reflecting a greater loss of transpiration and little to no loss in NPP. WUE could be further monitored in urban systems as an indicator of efficient water use for tree planting in semi-arid to arid cities.
In this study, we used RHESSys, a coupled dynamic ecohydrologic model, to estimate urban stand productivity for a range of climate and irrigation scenarios. As with all model studies there are important limitations. Species specific vegetation parameters and soil parameters were derived from a prior study that used remote sensing of vegetation drought response for parameter selection in the Santa Barbara urban area. Uncertainty in these parameters arises from both uncertainty in remote sensing data sampling limitations and equifinality (multiple parameter sets giving similar correspondence with remote sensing patterns). Soils and vegetation in urban environments are highly heterogeneous. Soils can also be altered by landscaping and are often compacted (De Kimpe and Morel, 2000; Morgenroth and Buchan, 2009; Edmondson et al., 2011), which lowers infiltration, soil moisture, and tree water accessibility (Gregory et al., 2006; Phillips et al., 2019; Fidal and Kjeldsen, 2020). We note that trees sampled in this prior study were predominately located in urban green spaces such as parks, which may have markedly different soils from urban street trees. Finally we also ignore trees along waterways that may receive additional groundwater inputs. Future work could utilize data from local urban street trees to explore how soil management and site selection might alter the predicted relationships among irrigation, climate and productivity. Measurements of infiltration capacity of urban soils that could also inform parameterization of these future modeling studies (Yang and Zhang, 2011; Phillips et al., 2019; Schaffitel et al., 2020).
Our parameterization and modeling approach also ignore the potential for some ecophysiologic traits to adapt to local conditions and a changing climate (Ibsen et al., 2023). Few ecohydrologic models incorporate this plasticity but as our understanding of trait adaptation matures, future models may incorporate these dynamics. Our study was also restricted to the 5 species of trees focused on in the prior study. There are a diversity of urban tree species used within the Southern California region and other semi-arid urban settings (Avolio et al., 2015). Expanding on this current study to explore a broader range of tree species could be used to develop combined irrigation and species selection strategies. Similarly future work could expand on the range of climate scenarios considered, and importantly consider more complex irrigation and water management strategies such as mulching to reduce water loss (Wang et al., 2021), or storm water redirection to fulfill irrigation requirements (Thom et al., 2022).
Because outdoor water for landscaping is one of the highest sectors of urban water use in Southern California (Gleick et al., 2003; Pincetl et al., 2018), reducing outdoor water use and improving efficiency is essential to climate change adaptation for future drought and heatwaves. Decision makers might consider limiting outdoor water use during wet years when over-watering normally occurs to save water for dry years when vegetation needs it more (Chen et al., 2015). During drought, urban forestry management should be considered along with water conservation efforts, and allocating water use toward trees that provide shade and cooling. In addition, investing in rainwater catchment and recycled wastewater would be a more efficient way of allocating water toward urban trees and their ecosystem services (Livesley et al., 2021).
This study used an ecohydrologic model in a semi-arid city to show that urban tree ecosystem services for select species can be maintained during a multi-year drought with moderate irrigation reductions of up to 25%. The drought period from 2012–2016 was an “exceptional” drought with both substantial precipitation declines and higher than normal temperatures. Similar and potential warmer droughts are expected to increase in Southern California in the future (AghaKouchak et al., 2014; Cook et al., 2015). Irrigation in semi-arid cities is important for maintaining tree-related ecosystem services not only because of the direct potential for evaporative cooling, but because of its indirect importance for maintaining tree productivity which contributes to building canopy for shade provision. When irrigation is reduced during drought years, both evapotranspiration and plant productivity become water limited. The amount of irrigation reduction, however, affects these two fluxes differently. Tree transpiration declined linearly with irrigation reduction, resulting in a short-term loss of evaporative cooling. NPP declined non-linearly with reduced irrigation and for moderate (< 25%) reductions at a more gradual rate than transpiration loss. Tree resilience, as indicated by LAI, following the drought was also maintained for irrigation reductions of less than 50% , even for warmer droughts. These results suggest that it is possible to maintain tree productivity during multi-year droughts and post-drought tree cover for shade provision with partial irrigation reductions.
Our results also highlight both consistency across species and important differences. All species showed linear declines in transpiration (and consequently evaporative cooling) with irrigation reduction during droughts but much more stable transpiration for non-drought years—suggesting a general pattern of overwatering when not in drought. All species also show resilience to drought for irrigation reductions less than 50% and generally small declines in NPP, a finding that can guide general watering restrictions. Species differences however are notable, particularly for more substantial irrigation reductions. Species specific model results should be interpreted with caution, and we note that for some species, such as QUAG and PICA, uncertainty in model estimates are significant. Nonetheless, these estimates of species differences in transpiration, NPP, and LAI can motivate and guide additional species specific monitoring to guide landscape planning and tree selection.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
RT: Writing – original draft, Software, Methodology, Formal analysis, Conceptualization. CT: Writing – review & editing, Software, Conceptualization. JM: Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
We would like to acknowledge Santa Barbara Water Resources for their generosity in data sharing of outdoor water use, that made this study possible. We would also like to acknowledge the Tague Lab at the Bren School of Environmental Science and Management, especially Janet Choate, for providing support and guidance throughout the process of this study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fclim.2024.1280615/full#supplementary-material
AghaKouchak, A., Cheng, L., Mazdiyasni, O., and Farahmand, A. (2014). Global warming and changes in risk of concurrent climate extremes: insights from the 2014 california drought. Geophy. Res. Lett. 41, 8847–8852. doi: 10.1002/2014GL062308
Alexander, J. D., McCafferty, M. K., Fricker, G. A., and James, J. J. (2023). Climate seasonality and extremes influence net primary productivity across california's grasslands, shrublands, and woodlands. Environ. Res. Lett. 18:064021. doi: 10.1088/1748-9326/acd396
Allen, M. A., Roberts, D. A., and McFadden, J. P. (2021). Reduced urban green cover and daytime cooling capacity during the 2012–2016 California drought. Urban Clim. 36:100768. doi: 10.1016/j.uclim.2020.100768
Alonzo, M., Bookhagen, B., and Roberts, D. A. (2014). Urban tree species mapping using hyperspectral and lidar data fusion. Rem. Sens. Environ. 148, 70–83. doi: 10.1016/j.rse.2014.03.018
Alonzo, M., McFadden, J. P., Nowak, D. J., and Roberts, D. A. (2016). Mapping urban forest structure and function using hyperspectral imagery and lidar data. Urban Forest. Urban Green. 17, 135–147. doi: 10.1016/j.ufug.2016.04.003
Avolio, M., Pataki, D. E., Gillespie, T., Jenerette, G. D., McCarthy, H. R., Pincetl, S., et al. (2015). Tree diversity in southern California's urban forest: the interacting roles of social and environmental variables. Front. Ecol. Evol. 3:73. doi: 10.3389/fevo.2015.00073
Bhattachan, A., Skaff, N. K., Irish, A. M., Vimal, S., Remais, J. V., and Lettenmaier, D. P. (2021). Outdoor residential water use restrictions during recent drought suppressed disease vector abundance in Southern California. Environ. Sci. Technol. 55, 478–487. doi: 10.1021/acs.est.0c05857
Bijoor, N. S., McCarthy, H. R., Zhang, D., and Pataki, D. E. (2012). Water sources of urban trees in the Los Angeles metropolitan area. Urban Ecosyst. 15, 195–214. doi: 10.1007/s11252-011-0196-1
Bijoor, N. S., Pataki, D. E., Haver, D., and Famiglietti, J. S. (2014). A comparative study of the water budgets of lawns under three management scenarios. Urban Ecosyst. 17, 1095–1117. doi: 10.1007/s11252-014-0361-4
Bolund, P., and Hunhammar, S. (1999). Ecosystem services in urban areas. Ecol. Econ. 29, 293–301. doi: 10.1016/S0921-8009(99)00013-0
Broadbent, A. M., Coutts, A. M., Tapper, N. J., and Demuzere, M. (2018). The cooling effect of irrigation on urban microclimate during heatwave conditions. Urban Clim. 23, 309–329. doi: 10.1016/j.uclim.2017.05.002
Brown, R. D., Vanos, J., Kenny, N., and Lenzholzer, S. (2015). Designing urban parks that ameliorate the effects of climate change. Landsc. Urban Plan. 138, 118–131. doi: 10.1016/j.landurbplan.2015.02.006
California Deptartment of Water Resources C. (2021). California irrigation management information system final report. Department of Land, Air and Water Resources, University of California, Davis.
Cayan, D. R., Das, T., Pierce, D. W., Barnett, T. P., Tyree, M., and Gershunov, A. (2010). Future dryness in the southwest US and the hydrology of the early 21st century drought. PNAS 107, 21271–21276. doi: 10.1073/pnas.0912391107
Chen, X. (2016). Factors affecting the streamflow and in-stream nitrate concentration in semi-arid areas: sub-surface flow-generation, vertical distribution of soil nitrate and drainage properties, and the connectivity of impervious areas. Ph.D. Dissertation, University of California Santa Barbara.
Chen, Y.-J., McFadden, J. P., Clarke, K. C., and Roberts, D. A. (2015). Measuring spatio-temporal trends in residential landscape irrigation extent and rate in Los Angeles, California using SPOT-5 satellite imagery. Water Resour. Manage. 29, 5749–5763. doi: 10.1007/s11269-015-1144-2
Cook, B. I., Ault, T. R., and Smerdon, J. E. (2015). Unprecedented 21st century drought risk in the American Southwest and Central Plains. Sci. Adv. 1:e1400082. doi: 10.1126/sciadv.1400082
De Kimpe, C. R., and Morel, J.-L. (2000). Urban soil management: a growing concern. Soil Sci. 165:31. doi: 10.1097/00010694-200001000-00005
Dickinson, R. E., Shaikh, M., Bryant, R., and Graumlich, L. (1998). Interactive canopies for a climate model. J. Clim. 11, 2823–2836. doi: 10.1175/1520-0442(1998)011<2823:ICFACM>2.0.CO;2
Diffenbaugh, N. S., Swain, D. L., and Touma, D. (2015). Anthropogenic warming has increased drought risk in California. Proc. Natl. Acad. Sci. USA. 112, 3931–3936. doi: 10.1073/pnas.1422385112
Edmondson, J. L., Davies, Z. G., McCormack, S. A., Gaston, K. J., and Leake, J. R. (2011). Are soils in urban ecosystems compacted? A citywide analysis. Biol. Lett. 7, 771–774. doi: 10.1098/rsbl.2011.0260
Esperon-Rodriguez, M., Tjoelker, M. G., Lenoir, J., Baumgartner, J. B., Beaumont, L. J., Nipperess, D. A., et al. (2022). Climate change increases global risk to urban forests. Nat. Clim. Change 12, 950–955. doi: 10.1038/s41558-022-01465-8
Exec. Order No. B-29-15 (2015). Available online at: https://www.waterboards.ca.gov/waterrights/water_issues/programs/drought/docs/040115_executive_order.pdf
Farquhar, G. D., and von Caemmerer, S. (1982). “Modelling of photosynthetic response to environmental conditions”, in Physiological Plant Ecology II: Water Relations and Carbon Assimilation, Encyclopedia of Plant Physiology, eds. O. L. Lange, P. S. Nobel, C. B. Osmond, and H. Ziegler (Berlin, Heidelberg: Springer), 549–587. doi: 10.1007/978-3-642-68150-9_17
Fidal, J., and Kjeldsen, T. R. (2020). Accounting for soil moisture in rainfall-runoff modelling of urban areas. J. Hydrol. 589:125122. doi: 10.1016/j.jhydrol.2020.125122
Fu, L., Xu, Y., Xu, Z., Wu, B., and Zhao, D. (2020). Tree water-use efficiency and growth dynamics in response to climatic and environmental changes in a temperate forest in Beijing, China. Environ. Int. 134:105209. doi: 10.1016/j.envint.2019.105209
Gao, K., Santamouris, M., and Feng, J. (2020). On the cooling potential of irrigation to mitigate urban heat island. Sci. Total Environ. 740:139754. doi: 10.1016/j.scitotenv.2020.139754
Ghosh, S., Scharenbroch, B. C., Burcham, D., Ow, L. F., Shenbagavalli, S., and Mahimairaja, S. (2016). Influence of soil properties on street tree attributes in Singapore. Urban Ecosyst. 19, 949–967. doi: 10.1007/s11252-016-0530-8
Gillner, S., Vogt, J., Tharang, A., Dettmann, S., and Roloff, A. (2015). Role of street trees in mitigating effects of heat and drought at highly sealed urban sites. Landsc. Urban Plan. 143, 33–42. doi: 10.1016/j.landurbplan.2015.06.005
Gleick, P. H., Wolff, G. H., and Cushing, K. K. (2003). Waste not, want not: The potential for urban water conservation in California. Pacific Institute for Studies in Development, Environment, and Security Oakland, CA.
Gober, P., Middel, A., Brazel, A., Myint, S., Chang, H., Duh, J.-D., et al. (2012). Tradeoffs between water conservation and temperature amelioration in phoenix and portland: implications for urban sustainability. Urban Geogr. 33, 1030–1054. doi: 10.2747/0272-3638.33.7.1030
Goedhart, C. M., and Pataki, D. E. (2012). Do arid species use less water than mesic species in an irrigated common garden? Urban Ecosyst. 15, 215–232. doi: 10.1007/s11252-011-0201-8
Gonzales, P., and Ajami, N. (2017). Social and structural patterns of drought-related water conservation and rebound. Water Resour. Res. 53, 10619–10634. doi: 10.1002/2017WR021852
Grabosky, J., and Bassuk, N. (2016). Seventeen years growth of street trees in structural soil compared with a tree lawn in New York City. Urban Forest. Urban Green. 16, 103–109. doi: 10.1016/j.ufug.2016.02.002
Gregory, J. H., Dukes, M. D., Jones, P. H., and Miller, G. L. (2006). Effect of urban soil compaction on infiltration rate. J. Soil Water Conserv. 61, 117–124. Available online at: http://www.jswconline.org/content/61/3/117.abstract
Griffin, D., and Anchukaitis, K. J. (2014). How unusual is the 2012–2014 California drought? Geophy. Res. Lett. 41, 9017–9023. doi: 10.1002/2014GL062433
Grossiord, C., Buckley, T. N., Cernusak, L. A., Novick, K. A., Poulter, B., Siegwolf, R. T. W., et al. (2020). Plant responses to rising vapor pressure deficit. New Phytol. 226, 1550–1566. doi: 10.1111/nph.16485
Grossiord, C., Sevanto, S., Borrego, I., Chan, A. M., Collins, A. D., Dickman, L. T., et al. (2017). Tree water dynamics in a drying and warming world. Plant, Cell Environ. 40, 1861–1873. doi: 10.1111/pce.12991
Horike, H., Kinoshita, T., Kume, A., and Hanba, Y. T. (2023). Responses of leaf photosynthetic traits, water use efficiency, and water relations in five urban shrub tree species under drought stress and recovery. Trees 37, 53–67. doi: 10.1007/s00468-021-02083-1
Ibsen, P. C., Santiago, L. S., Shiflett, S. A., Chandler, M., and Jenerette, G. D. (2023). Irrigated urban trees exhibit greater functional trait plasticity compared to natural stands. Biol. Lett. 19:20220448. doi: 10.1098/rsbl.2022.0448
Kirschbaum, M. U. F. (2000). Forest growth and species distribution in a changing climate. Tree Physiol. 20, 309–322. doi: 10.1093/treephys/20.5-6.309
Landsberg, J. J., and Waring, R. H. (1997). A generalised model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance and partitioning. Forest Ecol. Manage. 95, 209–228. doi: 10.1016/S0378-1127(97)00026-1
Langridge, R. (2018). Central Coast Summary Report. California's Fourth Climate Change Assessment Publication number: SUM-CCCA4–2018-006. University of California, Santa Cruz.
Liang, L. L., Anderson, R. G., Shiflett, S. A., and Jenerette, G. D. (2017). Urban outdoor water use and response to drought assessed through mobile energy balance and vegetation greenness measurements. Environ. Res. Lett. 12:084007. doi: 10.1088/1748-9326/aa7b21
Litvak, E., Manago, K. F., Hogue, T. S., and Pataki, D. E. (2017a). Evapotranspiration of urban landscapes in Los Angeles, California at the municipal scale: evapotranspiration of urban landscapes. Water Resour. Res. 53, 4236–4252. doi: 10.1002/2016WR020254
Litvak, E., McCarthy, H. R., and Pataki, D. E. (2017b). A method for estimating transpiration of irrigated urban trees in California. Landsc. Urban Plan. 158, 48–61. doi: 10.1016/j.landurbplan.2016.09.021
Litvak, E., and Pataki, D. E. (2016). Evapotranspiration of urban lawns in a semi-arid environment: an in situ evaluation of microclimatic conditions and watering recommendations. J. Arid Environ. 134, 87–96. doi: 10.1016/j.jaridenv.2016.06.016
Livesley, S. J., Marchionni, V., Cheung, P. K., Daly, E., and Pataki, D. E. (2021). Water smart cities increase irrigation to provide cool refuge in a climate crisis. Earth's Fut. 9:e2020EF001806. doi: 10.1029/2020EF001806
Livesley, S. J., McPherson, G. M., and Calfapietra, C. (2016). The Urban Forest and Ecosystem Services: Impacts on Urban Water, Heat, and Pollution Cycles at the Tree, Street, and City Scale. J. Environ. Quality 45:119. doi: 10.2134/jeq2015.11.0567
Lund, J., Medellin-Azuara, J., Durand, J., and Stone, K. (2018). Lessons from California's 2012–2016 drought. J. Water Resour. Plan. Manage. 144:04018067. doi: 10.1061/(ASCE)WR.1943-5452.0000984
Luthy Richard, G., Wolfand Jordyn, M., and Bradshaw Jonathan, L. (2020). Urban water revolution: sustainable water futures for california cities. J. Environ. Eng. 146:04020065. doi: 10.1061/(ASCE)EE.1943-7870.0001715
Malone, S. L., Tulbure, M. G., Prez-Luque, A. J., Assal, T. J., Bremer, L. L., Drucker, D. P., et al. (2016). Drought resistance across california ecosystems: evaluating changes in carbon dynamics using satellite imagery. Ecosphere 7:e01561. doi: 10.1002/ecs2.1561
Marchin, R. M., Broadhead, A. A., Bostic, L. E., Dunn, R. R., and Hoffmann, W. A. (2016). Stomatal acclimation to vapour pressure deficit doubles transpiration of small tree seedlings with warming. Plant, Cell Environ. 39, 2221–2234. doi: 10.1111/pce.12790
McCarthy, H. R., and Pataki, D. E. (2010). Drivers of variability in water use of native and non-native urban trees in the greater los angeles area. Urban Ecosyst. 13, 393–414. doi: 10.1007/s11252-010-0127-6
McCumber, A. (2017). Building “natural” beauty: drought and the shifting aesthetics of nature in Santa Barbara, California. Nat. Cult. 12, 246–262. doi: 10.3167/nc.2017.120303
McDowell, N., Pockman, W. T., Allen, C. D., Breshears, D. D., Cobb, N., Kolb, T., et al. (2008). Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol. 178, 719–739. doi: 10.1111/j.1469-8137.2008.02436.x
McHale, M. R., Burke, I. C., Lefsky, M. A., Peper, P. J., and McPherson, E. G. (2009). Urban forest biomass estimates: is it important to use allometric relationships developed specifically for urban trees? Urban Ecosyst. 12, 95–113. doi: 10.1007/s11252-009-0081-3
McHale, M. R., Hall, S. J., Majumdar, A., and Grimm, N. B. (2017). Carbon lost and carbon gained: a study of vegetation and carbon trade-offs among diverse land uses in phoenix, arizona. Ecol. Applic. 27, 644–661. doi: 10.1002/eap.1472
McKinnon, K. A., Poppick, A., and Simpson, I. R. (2021). Hot extremes have become drier in the United States Southwest. Nat. Clim. Chang. 11, 598–604. doi: 10.1038/s41558-021-01076-9
McPherson, E. G., Xiao, Q., van Doorn, N. S., de Goede, J., Bjorkman, J., Hollander, A., et al. (2017). The structure, function and value of urban forests in California communities. Urban Forest. Urban Green. 28, 43–53. doi: 10.1016/j.ufug.2017.09.013
Miller, D. L., Alonzo, M., Meerdink, S. K., Allen, M. A., Tague, C. L., Roberts, D. A., et al. (2022). Seasonal and interannual drought responses of vegetation in a california urbanized area measured using complementary remote sensing indices. ISPRS J. Photogram. Rem. Sens. 183, 178–195. doi: 10.1016/j.isprsjprs.2021.11.002
Miller, D. L., Alonzo, M., Roberts, D. A., Tague, C. L., and McFadden, J. P. (2020). Drought response of urban trees and turfgrass using airborne imaging spectroscopy. Rem. Sens. Environ. 240:111646. doi: 10.1016/j.rse.2020.111646
Mini, C., Hogue, T. S., and Pincetl, S. (2014). Estimation of residential outdoor water use in Los Angeles, California. Landscape Urban Plan. 127, 124–135. doi: 10.1016/j.landurbplan.2014.04.007
Mngadi, M., Odindi, J., Mutanga, O., and Sibanda, M. (2022). Estimating aboveground net primary productivity of reforested trees in an urban landscape using biophysical variables and remotely sensed data. Sci. Total Environ. 802:149958. doi: 10.1016/j.scitotenv.2021.149958
Morgenroth, J., and Buchan, G. (2009). Soil moisture and aeration beneath pervious and impervious pavements. J. Arboric. 35, 135–141. doi: 10.48044/jauf.2009.024
Muggeo, V. M. R. (2008). Segmented: an R package to fit regression models with broken-line relationships. R News 8, 20–25.
Nouri, H., Beecham, S., Hassanli, A. M., and Kazemi, F. (2013). Water requirements of urban landscape plants: a comparison of three factor-based approaches. Ecol. Eng. 57, 276–284. doi: 10.1016/j.ecoleng.2013.04.025
Nowak, D. J., Greenfield, E. J., Hoehn, R. E., and Lapoint, E. (2013). Carbon storage and sequestration by trees in urban and community areas of the United States. Environ. Pollut. 178, 229–236. doi: 10.1016/j.envpol.2013.03.019
Palazzo, J., Liu, O. R., Stillinger, T., Song, R., Wang, Y., Hiroyasu, E. H. T., et al. (2017). Urban responses to restrictive conservation policy during drought. Water Resour. Res. 53, 4459–4475. doi: 10.1002/2016WR020136
Pataki, D. E., McCarthy, H. R., Litvak, E., and Pincetl, S. (2011). Transpiration of urban forests in the Los Angeles metropolitan area. Ecol. Applic. 21, 661–677. doi: 10.1890/09-1717.1
Phillips, T. H., Baker, M. E., Lautar, K., Yesilonis, I., and Pavao-Zuckerman, M. A. (2019). The capacity of urban forest patches to infiltrate stormwater is influenced by soil physical properties and soil moisture. J. Environ. Manage. 246, 11–18. doi: 10.1016/j.jenvman.2019.05.127
Pierce, D. W., Cayan, D. R., and Thrasher, B. L. (2014). Statistical downscaling using localized constructed analogs (LOCA). J. Hydrometeor. 15, 2558–2585. doi: 10.1175/JHM-D-14-0082.1
Pincetl, S., Porse, E., Mika, K. B., Litvak, E., Manago, K. F., Hogue, T. S., et al. (2018). Adapting urban water systems to manage scarcity in the 21st Century: the case of Los Angeles. Environ. Manag. 63, 293–308. doi: 10.1007/s00267-018-1118-2
Quesnel, K. J., Ajami, N., and Marx, A. (2019). Shifting landscapes: decoupled urban irrigation and greenness patterns during severe drought. Environ. Res. Lett. 14:064012. doi: 10.1088/1748-9326/ab20d4
Quesnel, K. J., and Ajami, N. K. (2019). Large landscape urban irrigation: a data driven approach to evaluate conservation behavior. Water Resour. Res. 55, 771–786. doi: 10.1029/2018WR023549
Rahman, M. A., Stratopoulos, L. M. F., Moser-Reischl, A., Zölch, T., Häberle, K.-H., Rötzer, T., et al. (2020). Traits of trees for cooling urban heat islands: a meta-analysis. Build. Environ. 170:106606. doi: 10.1016/j.buildenv.2019.106606
Reyes, B., Hogue, T., and Maxwell, R. (2018). Urban irrigation suppresses land surface temperature and changes the hydrologic regime in semi-arid regions. Water 10:1563. doi: 10.3390/w10111563
Roberts, D. A., Ustin, S. L., Ogunjemiyo, S., Greenberg, J., Dobrowski, S. Z., Chen, J., et al. (2004). Spectral and structural measures of northwest forest vegetation at leaf to landscape scales. Ecosystems 7, 545–562. doi: 10.1007/s10021-004-0144-5
Ryan, M. G., Lavigne, M. B., and Gower, S. T. (1997). Annual carbon cost of autotrophic respiration in boreal forest ecosystems in relation to species and climate. J. Geophys. Res. 102, 28871–28883. doi: 10.1029/97JD01236
Sax, C., and Steiner, P. (2013). Temporal disaggregation of time series. R J. 5:80. doi: 10.32614/RJ-2013-028
Schaffitel, A., Schuetz, T., and Weiler, M. (2020). A distributed soil moisture, temperature and infiltrometer dataset for permeable pavements and green spaces. Earth Syst. Sci. Data 12, 501–517. doi: 10.5194/essd-12-501-2020
Shields, C., and Tague, C. (2015). Ecohydrology in semiarid urban ecosystems: Modeling the relationship between connected impervious area and ecosystem productivity. Water Resour. Res. 51, 302–319. doi: 10.1002/2014WR016108
Shields, C. A., and Tague, C. L. (2012). Assessing the role of parameter and input uncertainty in ecohydrologic modeling: implications for a semi-arid and urbanizing coastal california catchment. Ecosystems 15, 775–791. doi: 10.1007/s10021-012-9545-z
Stabler, L. B. (2008). Management regimes affect woody plant productivity and water use efficiency in an urban desert ecosystem. Urban Ecosyst. 11, 197–211. doi: 10.1007/s11252-008-0050-2
Sudimac, S., Sale, V., and Kühn, S. (2022). How nature nurtures: amygdala activity decreases as the result of a one-hour walk in nature. Mol. Psychiat. 27, 4446–4452. doi: 10.1038/s41380-022-01720-6
Sun, G., Hallema, D., and Asbjornsen, H. (2017). Ecohydrological processes and ecosystem services in the Anthropocene: a review. Ecol. Proc. 6:35. doi: 10.1186/s13717-017-0104-6
Tague, C., and Pohl-Costello, M. (2008). The potential utility of physically based hydrologic modeling in ungauged urban streams. Ann. Assoc. Am. Geogr. 98, 818–833. doi: 10.1080/00045600802099055
Tague, C. L., and Band, L. E. (2004). RHESSys: regional hydro-ecologic simulation system an object-oriented approach to spatially distributed modeling of carbon, water, and nutrient cycling. Earth Inter. 8, 1–42. doi: 10.1175/1087-3562(2004)8<1:RRHSSO>2.0.CO;2
Thom, J. K., Fletcher, T. D., Livesley, S. J., Grey, V., and Szota, C. (2022). Supporting growth and transpiration of newly planted street trees with passive irrigation systems. Water Resour. Res. 58:e2020WR029526. doi: 10.1029/2020WR029526
Torres, R. (2024). Urban tree drought resilience and outdoor water conservation in a Mediterranean climate: insights from an ecohydrologic model. PhD thesis, University of Calfiornia, Santa Barbara.
Tsamir, M., Gottlieb, S., Preisler, Y., Rotenberg, E., Tatarinov, F., Yakir, D., et al. (2019). Stand density effects on carbon and water fluxes in a semi-arid forest, from leaf to stand-scale. Forest Ecol. Manage. 453:117573. doi: 10.1016/j.foreco.2019.117573
Vahmani, P., and Ban-Weiss, G. (2017). Climatic consequences of adopting drought-tolerant vegetation over Los Angeles as a response to California drought. Geophys. Res. Lett. 45, 8240–8249. doi: 10.1002/2016GL069658
Vahmani, P., and Hogue, T. S. (2014). Incorporating an urban irrigation module into the noah land surface model coupled with an urban canopy model. J. Hydrometeorol. 15, 1440–1456. doi: 10.1175/JHM-D-13-0121.1
Vicente-Serrano, S. M., Camarero, J. J., Zabalza, J., Sangesa-Barreda, G., López-Moreno, J. I., and Tague, C. L. (2015). Evapotranspiration deficit controls net primary production and growth of silver fir: implications for circum-mediterranean forests under forecasted warmer and drier conditions. Agric. Forest Meteorol. 206, 45–54. doi: 10.1016/j.agrformet.2015.02.017
Wang, B., Niu, J., Berndtsson, R., Zhang, L., Chen, X., Li, X., et al. (2021). Efficient organic mulch thickness for soil and water conservation in urban areas. Sci. Rep. 11:6259. doi: 10.1038/s41598-021-85343-x
Water Resources (2010-2020). [City of Santa Barbara Dedicated Irrigation Accounts Water Consumption] [Unpublished raw data]. City of Santa Barbara Public Works.
Wilson, T. S., Sleeter, B. M., and Cameron, D. R. (2016). Future land-use related water demand in california. Environ. Res. Lett. 11:054018. doi: 10.1088/1748-9326/11/5/054018
Winbourne, J. B., Jones, T. S., Garvey, S. M., Harrison, J. L., Wang, L., Li, D., et al. (2020). Tree transpiration and urban temperatures: current understanding, implications, and future research directions. BioScience 70, 576–588. doi: 10.1093/biosci/biaa055
Yang, J., and Wang, Z.-H. (2017). Planning for a sustainable desert city: the potential water buffering capacity of urban green infrastructure. Landsc. Urban Plann. 167, 339–347. doi: 10.1016/j.landurbplan.2017.07.014
Yang, J., Wang, Z.-H., Chen, F., Miao, S., Tewari, M., Voogt, J. A., et al. (2015). Enhancing hydrologic modelling in the coupled weather research and forecasting urban modelling system. Boundary-Layer Meteorol. 155, 87–109. doi: 10.1007/s10546-014-9991-6
Yang, J.-L., and Zhang, G.-L. (2011). Water infiltration in urban soils and its effects on the quantity and quality of runoff. J. Soils Sedim. 11, 751–761. doi: 10.1007/s11368-011-0356-1
Zhuang, Q., Shao, Z., Gong, J., Li, D., Huang, X., Zhang, Y., et al. (2022). Modeling carbon storage in urban vegetation: progress, challenges, and opportunities. Int. J. Appl. Earth Observ. Geoinform. 114:103058. doi: 10.1016/j.jag.2022.103058
Keywords: ecohydrology, urban water use, ecosystem services, urban forestry, water conservation, drought resilience, climate change adaptation
Citation: Torres R, Tague CL and McFadden JP (2024) Exploring potential trade-offs in outdoor water use reductions and urban tree ecosystem services during an extreme drought in Southern California. Front. Clim. 6:1280615. doi: 10.3389/fclim.2024.1280615
Received: 21 August 2023; Accepted: 12 March 2024;
Published: 27 March 2024.
Edited by:
Tobi Eniolu Morakinyo, University College Dublin, IrelandReviewed by:
Francesco Ferrini, University of Florence, ItalyCopyright © 2024 Torres, Tague and McFadden. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rachel Torres, UmFjaGVsLlRvcnJlc0BodW1ib2xkdC5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.