 Xin Zhou
Xin Zhou Lili Chen2†
Lili Chen2† Wenxin Zheng
Wenxin Zheng Feng Zhou
Feng Zhou- 1Departments of Pathology, The International Peace Maternal and Child Health Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, China
- 2Department of Oncology, Zhejiang University School of Medicine Women’s Hospital, Hangzhou, Zhejiang, China
- 3Laboratory of Pathology, National Cancer Institute, National Institutes of Health, Bethesda, MD, United States
- 4Department of Pathology, Department of Obstetrics and Gynecology, University of Texas Southwestern Medical Center, Dallas, TX, United States
- 5Harold C. Simon Comprehensive Cancer Center, University of Texas Southwestern Medical Center, Dallas, TX, United States
- 6Shanghai Key Laboratory of Embryo Original Diseases, Shanghai, China
Background: Microbial community dynamics have been involved in numerous diseases, including cancer. The diversity of intertumoral microbiota in human papillomavirus independent endocervical adenocarcinoma (HPVI ECA) is not well-characterized.
Objective: Our objective is to delineate the intratumoral microbiota profile in HPVI ECA and investigate its potential influence on oncogenesis.
Methods: We analyzed 45 HPVI ECA cases, comprising 36 gastric-type ECA (GEA) and 9 clear cell carcinomas (CCC). We compared the microbial composition within cancerous and adjacent noncancerous tissue samples using 5R-16S ribosomal DNA sequencing. Further, we investigated the correlation between specific microbes and clinical-pathological metrics as well as patient outcomes.
Results: Our findings demonstrate notable differences in the microbial spectra between cancerous and adjacent noncancerous tissues. Amongst HPVI ECA subtypes, GEAs exhibit more microbial variations compared to CCCs. Using the Random Forest algorithm, we identified two distinct microbial signatures that could act as predictive biomarkers for HPVI ECA and differentiate between GEA and CCC. Varied microbial abundances was related to clinical characteristics of HPVI ECA patients. In addition, high levels of Micrococcus and low levels of unknown genus75 from the Comamonadaceae family were associated with poorer outcomes in HPVI ECA patients. Similarly, an abundance of Microbacterium correlated with reduced overall survival (OS), and a high presence of Streptococcaceae family microbes was linked to reduced recurrence-free survival (RFS) in GEA patients. Intriguingly, a high abundance of Micrococcus was also associated with a worse OS in GEA patients.
Conclusion: The study reveals distinct microbial signatures in HPVI ECA, which have potential as biomarkers for disease prognosis. The correlation between these tumor-associated microbiota features and clinicopathological characteristics underscores the possibility of microbiome-based interventions. Our research provides a foundation for more in-depth studies into the cervical microbiome’s role in HPVI ECA.
Introduction
Endocervical adenocarcinoma (ECA) is the second most common histological subtype of cervical cancer, accounting for approximately 20-25% of all cases in Western countries (Bergström et al., 1999; Smith et al., 2000; Adegoke et al., 2012). While the majority of cervical squamous cell carcinomas (SCC) are associated with human papillomavirus (HPV) infection, a notable 10-25% of ECAs are HPV-independent (HPVI) (Kusanagi et al., 2010; Karamurzin et al., 2015; Stolnicu et al., 2018). The 2020 World Health Organization (WHO) classification of tumors of the female reproductive organs includes several subtypes of HPVI category, namely gastric-type ECA (GEA), clear cell carcinoma (CCC), mesonephric carcinoma, endometrioid carcinoma and adenocarcinoma not otherwise specified (Parra-Herran et al, 2020). Among these, GEA is the most prevalent subtype of HPVI ECA, representing 10-15% of all ECAs globally and up to 25% in Asian populations (Kusanagi et al., 2010; Karamurzin et al., 2015). CCC is another subtype of HPVI ECA, constituting 4-9% of all ECA cases (Noller et al., 1974; Saigo et al., 1986).
HPVI ECAs often present with more aggressive behavior, a propensity for extrauterine spread, and are frequently diagnosed at an advanced-stage, leading to a less favorable prognosis when compared to HPV associated ECAs (Kojima et al., 2007; Karamurzin et al., 2015; Nishio et al., 2019). The critical role of HPV in most cervical adenocarcinomas notwithstanding, it is insufficient to explain the etiology and progression of HPVI ECA alone. This gap in understanding underscores the urgent need for research into additional factors that may influence the onset and advancement of this cancer type.
Emerging evidence posits the microbiota as a pivotal regulatory agent and potential biomarker in the pathogenesis of various cancers, including that of the cervix (Meng et al., 2018; Liu et al., 2019a; Wu et al., 2021). Imbalances in microbial community homeostasis can trigger cancer initiation and growth through a variety of complex processes. These include the induction of inflammation, impairment of immune function, toxin production, DNA damage, activation of oncogenic pathways, and alteration of anticancer drug pharmacokinetics (García-Castillo et al., 2016; Byrd et al., 2023). Furthermore, the extensive microbiota, considered a secondary genome within the host, is instrumental in molding the tumor microenvironment and thus impacts both cancer progression and the efficacy of treatment (Pushalkar et al., 2018; Jain et al., 2021).
Current research has established links between cervical/vaginal microbiota and cervical lesions (Mitra et al., 2015; Audirac-Chalifour et al., 2016; Wu et al., 2021; Lin et al., 2022; Jiang et al., 2023; Teka et al., 2023; Wu et al., 2023), but the specific microbial species or communities associated with ECA incidence, clinical pathological features, and prognosis remain largely unexplored, particularly for HPVI ECA. This study, aims to delineate the differences in microbial abundance between cancerous and adjacent noncancerous tissues and further investigate the intratumoral microbiota’s role in cancer prediction and subtype differentiation. We also investigate the associations between microbiota profile and clinical-pathological variables, as well as prognostic implications for the disease.
Method and material
Sample collection
Paraffin-embedded samples from 45 HPVI ECA cases (36 GEA and 9 CCC) from March 2018 to February 2023. Patients’ clinicopathologic information, including age, clinical stage, and follow-up data were extracted from the electronic clinical information system database of Women’s Hospital. Hematoxylin and eosin and immunohistochemistry (IHC) slides were reviewed by two gynecological pathologists (X.Z. and F.Z.) in a blinded fashion and the pathologic diagnosis was confirmed. The sample collection procedure and further studies were approved by the Institutional Review Board and Ethics Committee of the Zhejiang University School of Medicine Women’s Hospital, China (IRB-20210276R).
Subsequently, 5R-16S ribosomal DNA (5R-16S rDNA) MiSeq sequencing was performed on these samples. All experiments were conducted following the guidelines and instructions approved by the manufacturer. Clinical staging of patients was categorized according to the staging system for gynecologic tumors developed by the International Federation of Gynecology and Obstetrics (FIGO) (Bhatla et al., 2018).
Human papillomavirus RNA in situ hybridization
High-risk HPV (hrHPV) testing was performed using RNA in situ hybridization (RISH) (Advanced Cell Diagnostics, Hayward, CA, USA), which recognizes 18 hrHPV genotypes (16,18, 26, 31, 33, 35, 39, 45, 51, 52, 53, 56, 58, 59, 66, 68, 73, and 82) according to the manufacturer’s instructions. The presence of intracellular punctate yellow-to-brownish reaction was defined as positive staining for HPV RNA (Wang et al., 2023).
DNA extraction and 5R-16S rDNA sequencing
DNA from various samples was extracted using the formalin-fixed and paraffin embedded (FFPE) Genome DNA Extraction Kit (Concert, Xiamen, China) following the manufacturer’s guidelines. The integrity and fragment size of the extracted DNA were assessed through 1% agarose gel electrophoresis. Additionally, the quality of the extracted DNA was measured using NanoDrop 2000 (Thermo Scientific, Waltham, MA, USA).
Microbial community analysis was performed using 5R-16S rDNA sequencing technology (Lianchuan Biotechnology, Hangzhou, China). To amplify the five regions (V2, V3, V5, V6, and V8) on the 16S rDNA, specific primers were utilized in two rounds of amplification. Primers and amplification procedures were applied as described in previous reference (Nejman et al., 2020). Following amplification, the PCR products underwent purification using AMPure XT beads (Beckman Coulter Genomics, Danvers, MA, USA) and quantification via Qubit (Invitrogen, USA). Subsequently, amplicon pools were readied for sequencing, with the size and quantity of the amplicon library evaluated on an Agilent 2100 Bioanalyzer (Agilent, USA) and using the Library Quantification Kit for Illumina (Kapa Biosciences, Woburn, MA, USA), respectively. Following sequencing of the libraries on the NovaSeq 6000 platform, a total of 18,814,031 reads were obtained after quality control and filtering, with an average of 209,045 reads per sample.
Microbiome data analysis
Paired-end reads were sorted into samples using unique barcodes and then trimmed by removing the barcode and primer sequences using cutadapt (v1.9). FLASH (v1.2.8) was employed for merging paired-end reads. Quality filtering of raw reads was performed with fqtrim (v0.94) under specific filter conditions to obtain high-quality clean reads. Chimeric sequences were filtered using Vsearch software (v2.3.4). Following dereplication of reads and discarding of singletons using DADA2, the representative sequence with single-base accuracy is obtained, resulting in the ASV (Amplicon Sequence Variants) feature table and feature sequence. The ASV feature sequence alignment for species annotation was performed using BLAST, with the SILVA and NT-16S alignment databases. Subsequently, species abundance per sample was calculated based on the ASV abundance table. Annotation confidence was set at a threshold of 0.7.
The α diversity indexes (observed species, Chao1, Shannon, Simpson) was implemented to display the diversity of bacteria among the different samples. The β diversity analysis was used to evaluate global differences between microbial community composition. Bray-Curtis for Nonmetric multidimensional scaling (NMDS) was carried out to visualize the β diversity between different groups. The α diversity and β diversity were calculated by QIIME, relative abundance was used in bacteria taxonomy. Linear discriminant analysis effect size (LEfSe) was employed to assess differences in abundance of taxonomic units between different groups (Segata et al., 2011), with those having linear discriminant analysis (LDA) values greater than 3 and p-values less than 0.05 considered to have significant differential abundance. Wilcoxon rank-sum tests were employed for statistical comparisons of taxonomic units at the phylum and genus levels between different groups. Additionally, using Phylogenetic Investigation of Communities by Reconstruction of Unobserved States2 (PICRUSt2) based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) database and Cluster of Ortholog Genes (COG) database (Douglas et al., 2020), the functional composition of bacterial communities between different groups was predicted using STAMP v2.1.3 (Parks et al., 2014). A p-value less than 0.05 was considered statistically significant.
Microbiota signature identification
In this study, we employed the Random Forest (RF) algorithm for the selection of microbiota signatures. RF is a popular supervised machine learning method used for modeling various omics data, including microbiome data, and it is highly effective for both prediction and interpretation purposes. A classification model was established using the RF algorithm from the RandomForest R package (https://cran.r-project.org/web/packages/randomForest/index.html). The receiver operating characteristic (ROC) curve was constructed using the pROC R package (https://cran.r-project.org/web/packages/pROC/index.html), and the performance of the prediction model was evaluated through the area under the ROC curve (AUC).
Statistics analysis
Data analysis was performed using GraphPad Prism and R. Statistical tests were based on two-sided comparisons, with significance set at p < 0.05. When stratifying according to clinical pathological features, the Wilcoxon test was employed to determine differential enrichment of bacterial taxa. The Pearson “cor.test () “ function in R version 4.0.2 was utilized to analyze the correlation between specific microbial relative abundance and tumor staging. Consistent with our previous studies (Sun et al., 2024), overall survival (OS) was defined as the time from surgery to death or the end of follow-up, while recurrence-free survival (RFS) was defined as the interval between total hysterectomy and first recurrence, metastasis or last observation. Following this, samples were classified into high-abundance and low-abundance groups based on the microbial composition abundance. Kaplan‐Meier survival analysis was conducted to assess microbial communities associated with RFS and OS.
Result
Clinicopathologic characteristics of patients
This study included 45 patients with HPVI ECA, with 36 patients with GEA and 9 with CCC. The median age of these patients was 49 years (49 years for the GEA and 58 years for the CCC). The median follow-up duration for the cohort was 27 months with GEA patients having a median of 26 months and CCC patients 32 months. Details on patient demographics, pathologic diagnoses, clinical stage (FIGO stage), tumor maximum diameter (TMD), deep stromal invasion, lymph node metastasis (LNM), and lymphovascular invasion (LVI), and follow-up data are summarized in Table 1 and Figure 1A.
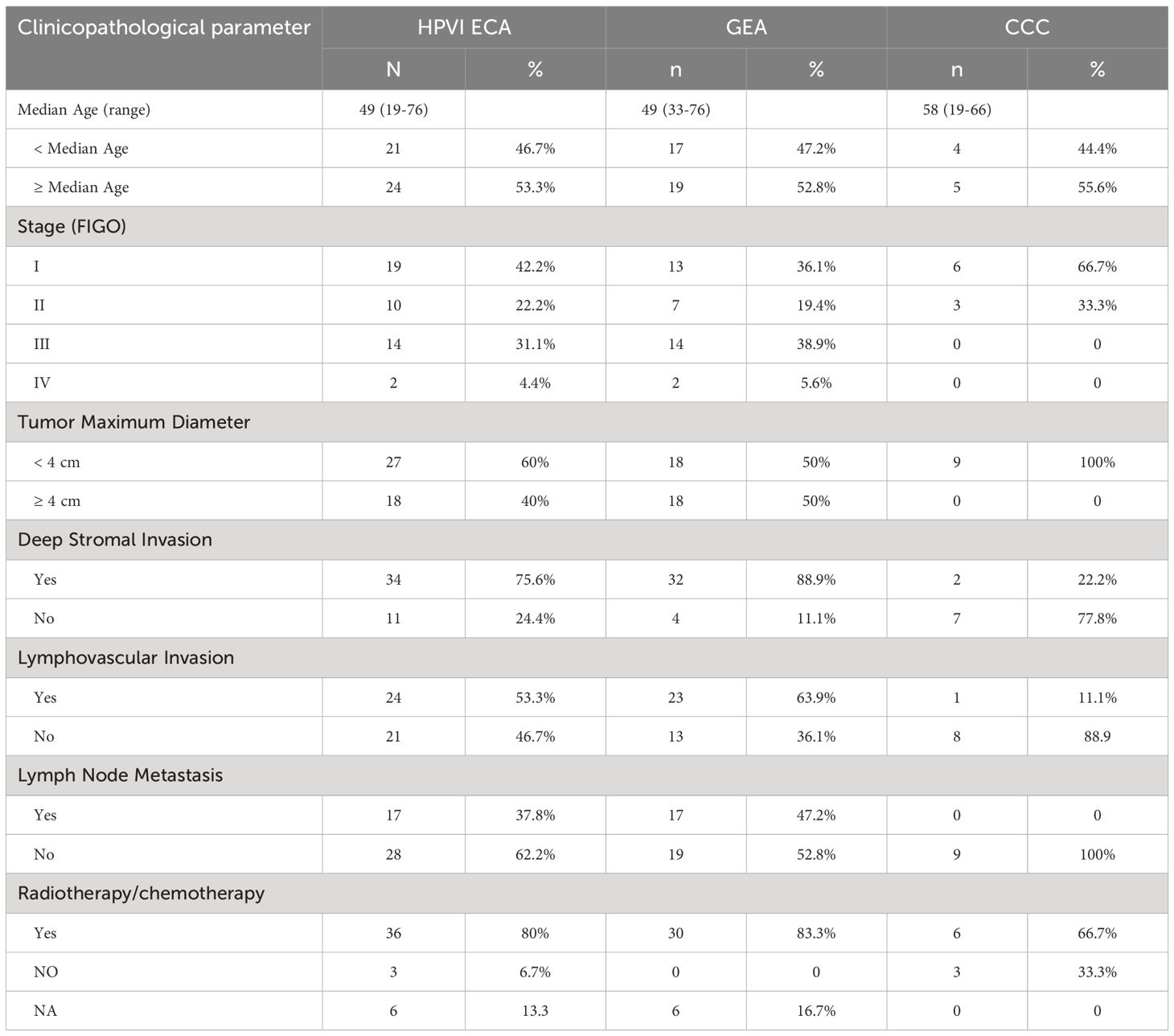
Table 1. Clinicopathological information of patients.
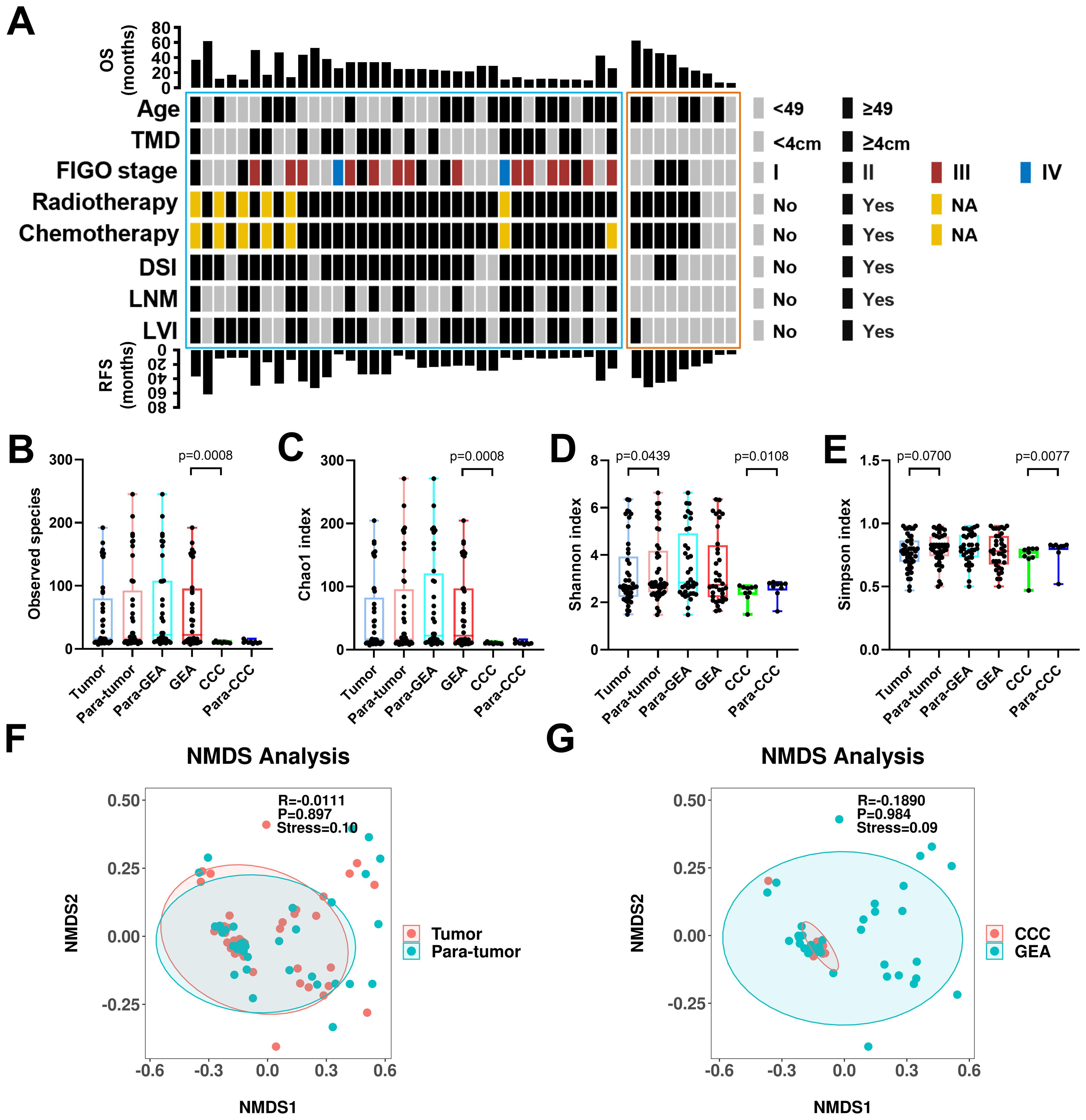
Figure 1. Clinical-pathological characteristics and intra-tumoral microbiota diversity of all patients. (A) Clinical-pathological characteristics and clinical outcomes of all 45 human papillomavirus independent endocervical adenocarcinoma (HPVI ECA) patients. Cases in the blue box belong to gastric-type endocervical adenocarcinoma (GEA) patients, and cases in the red box belong to clear cell carcinoma (CCC) patients. (B-E) The α-diversity among these groups was evaluated using the two-sided Wilcoxon sum rank test for the following indices: Box plots of observed species (B), Chao1 index (C), Shannon index (D), and Simpson index (E) in HPVI ECA samples. (F, G) β-diversity assessed by non-metric multidimensional scaling (NMDS) using Bray-Curtis. (F) HPVI ECA cancerous tissue versus adjacent non-cancerous tissue, (G) GEA versus CCC. TMD, tumor maximum diameter; DSI, deep stromal invasion; LNM, lymph node metastasis; LVI, lymphovascular invasion.
Intratumor microbial diversity in HPVI ECA
We compared the microbial communities within cancerous lesions (tumor group) to those in the adjacent non-cancerous tissues (para-tumor group) across the 45 HPVI ECA patients. We also analyzed the differences between the GEA and CCC groups. In our result, no significant differences in the Observed species were noted between the tumor and para-tumor groups. However, species richness was significantly greater in the GEA than in the CCC (Observed species, p = 0.0008) (Figure 1B). The Chao1 species richness index also confirmed this significant difference (Chao1 p = 0.0008) (Figure 1C). Marginally significant variations were seen in microbial community α-diversity between cancerous and adjacent non-cancerous tissues (Shannon p = 0.0438, Simpson p = 0.0700). However, between CCC and its adjacent tissue, the α-diversity shows a significant difference (Shannon p = 0.0108, Simpson p = 0.0077) (Figures 1D, E). Regarding β-diversity, Bray-Curtis dissimilarity assessments and NMDS plots revealed no significant difference between the tumor and para-tumor groups or between the GEA and CCC groups, though a certain explanatory meaning observed in these grouping scheme (0.05 < stress < 0.1), as shown in Figures 1F, G.
Altered composition of the intratumor microbiome in HPVI ECA
At the phylum level, Proteobacteria, Thermi, and Deinococcus-thermus were predominantly found across all groups, as shown in Figure 2A. A more detailed examination at the genus level highlighted a significant enrichment of Deinococcus, as well as Unknown genus75 of family Comamonadaceae, Cupriavidus, Meiothermus, and Porphyrobacter across all groups. Notably, Nevskia was markedly more abundant in the GEA and its adjacent non-cancerous groups, as presented in Figure 2B. The average abundance ratios of the top 10 microbial compositions at the phylum and genus levels for each group are presented in Supplementary Table S1. Microbial community compositions at other taxonomic levels are detailed in Supplementary Figure S1.
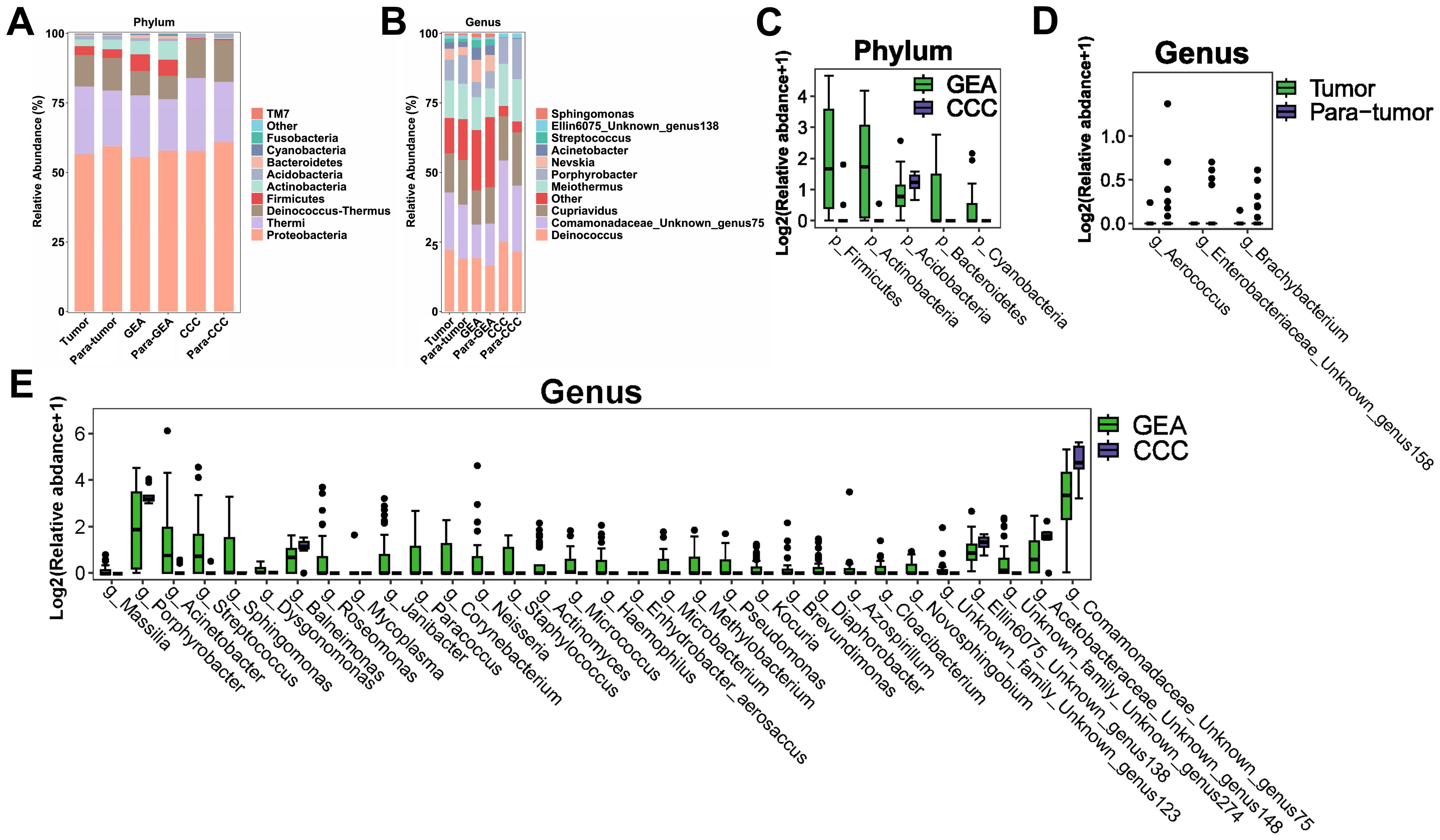
Figure 2. Microbial composition and differences between cancerous lesions and adjacent tissues. (A) Stacked bar chart showing the average relative abundance of the top 10 phyla across all groups. (B) Stacked bar chart depicting the average relative abundance of the top 10 genera across all groups. (C-E) Boxplots evaluating the statistical differences of intra-tumoral microbiota at the phylum and genus levels. (C) Genera with differential abundance between Tumor and Para-tumor groups. (D) Phyla with differential abundance between gastric-type endocervical adenocarcinoma (GEA) and clear cell carcinoma (CCC) groups. (E) Genera with differential abundance between GEA and CCC groups.
Comparative analysis using the Wilcoxon rank-sum test revealed no significant differences in microbial composition at the phylum level between the HPVI ECA and adjacent non-cancerous groups. However, when comparing GEA and CCC groups, relative abundances of Actinobacteria, Firmicutes, Betaproteobacteria, Bacteroidetes, Cyanobacteria, and Acidobacteria were significantly different (Figure 2C). At the genus level, differences in the microbial composition of Brachybacterium, Unknown genus158 of family Enterobacteriaceae, and Aerococcus were statistically significant within cancer tissues compared to paired adjacent tissues, as indicated in Figure 2D. Box plots in Figure 2E showed 32 microbial genera were significantly different between the GEA and CCC. Taxonomic differences at the phylum and genus levels and their corresponding p-values are detailed in Supplementary Table S2.
Furthermore, LEfSe was utilized to corroborate these statistical differences and to identify significant taxonomic groups within the various microbial communities. A substantial increase in the relative abundance of Teiothermus, Thermaceae, and Thermales was observed in cancerous lesions, as shown in Figure 3A. At the genus level, LEfSe results identified 25 microbial genera with significant differences between GEA and CCC (Figure 3B). Additionally, we found that among all taxonomic categories, 77 bacteria had increased relative abundance in the GEA, while 22 bacteria were more abundant in CCC, detailed in Supplementary Figure S2.
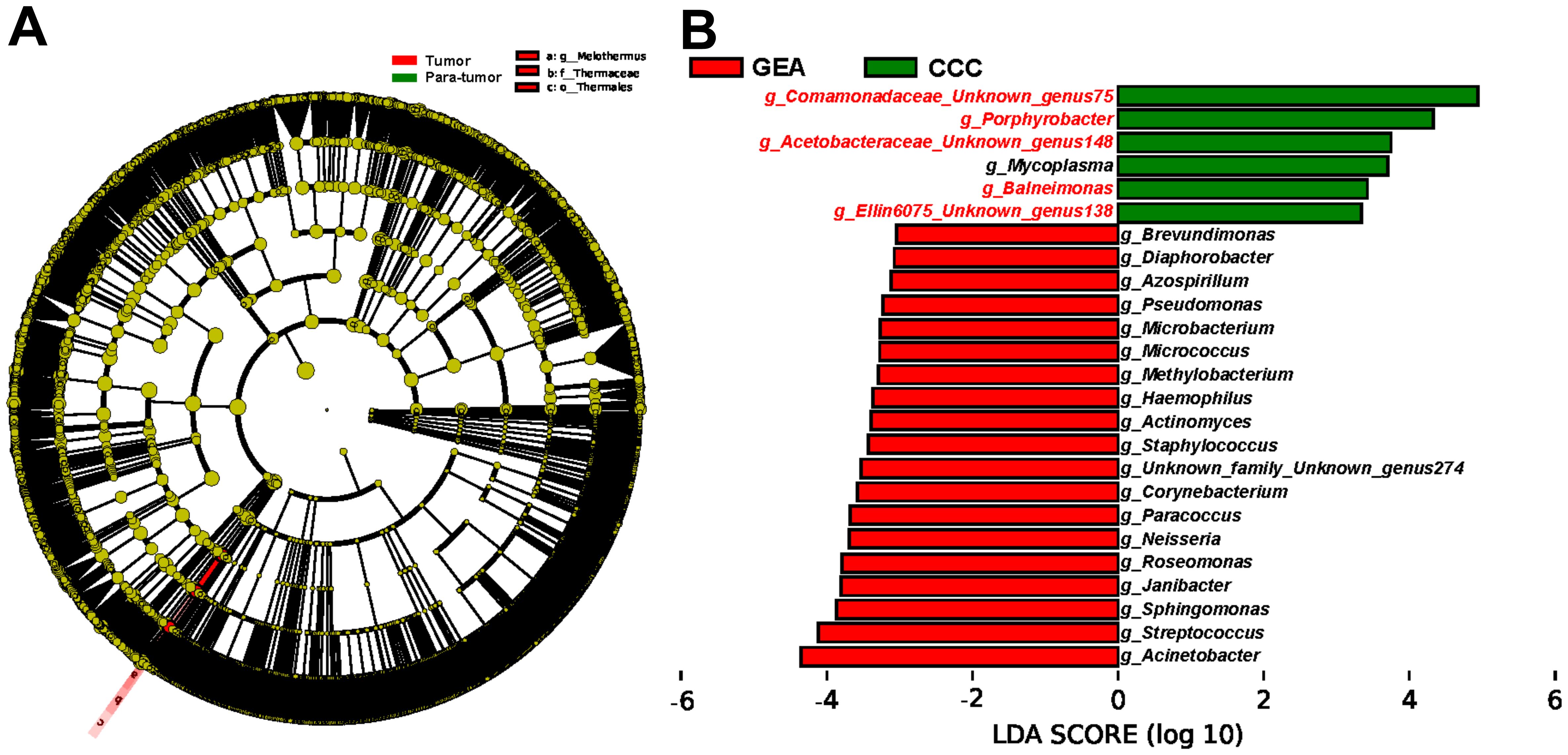
Figure 3. Identification of enriched microbial taxa between groups using Linear Discriminant Analysis Effect Size (LEfSe) method. (A) Cladogram illustrating taxonomic enrichment of microbial taxa between cancerous lesion and adjacent tissues. (B) Linear Discriminant Analysis (LDA) scores of specific microbial taxa at the genus level between gastric-type endocervical adenocarcinoma (GEA) and clear cell carcinoma (CCC) groups. Genera highlighted in red font indicate taxa overlapping with microbes in the subsequent Random Forest (RF) model distinguishing GEA from CCC, with an LDA score threshold of 3.
The role of intratumoral microbiota in HPVI ECA prediction and subtype differentiation
Exploration of microbial signatures within tumor micro environments has now made it possible to identify potential diagnostic markers for HPVI ECA. In general terms, a “microbial signature” refers to a collection of microbial taxa that are closely associated with a specific trait and have a high predictive value within a given model (Rivera-Pinto et al., 2018). The RF method, a machine learning algorithm, excels at identifying an optimal set of variables with significant discriminative power from a large pool of dependent or independent variables. It is renowned for its capability to capture complex dependency patterns between the outcome and the covariates (Hornung and Wright, 2019). A training cohort consisting of 31 pairs of HPVI ECA cancerous and adjacent tissue samples was used to develop a RF prediction model. And the top six genera were selected through a 10-fold cross-validation (Figures 4A, B). The detailed genera composition in each sample was listed in Supplementary Table S3. The model’s effectiveness was confirmed not only in the training cohort but also in a validation cohort of 14 pairs of HPVI ECA samples, achieving AUC scores of 0.764 and 0.694 for the training and validation sets, respectively (Figure 4C).

Figure 4. Utilization of Random Forest (RF) algorithm for tumor prediction and discerning tumor subtypes based on intratumoral microbiota signature. (A) Prioritization of the top 6 genera between human papillomavirus independent endocervical adenocarcinoma (HPVI ECA) tissues and adjacent tissues using 10-fold cross-validation method. (B) Cross-validation error curves between cancerous and adjacent non-cancerous tissues demonstrating the 10-fold cross-validation method employed to determine prioritization of top features for constructing simplified models. (C) The Receiver Operating Characteristic (ROC) curve illustrating predictive performance of simplified RF model constructed based on the top 6 genera. Blue and red curves represent model performance in training set (31 pairs) and validation set (14 pairs) respectively. (D) Prioritization of the top 6 genera between gastric-type endocervical adenocarcinoma (GEA) and clear cell carcinoma (CCC) using 10-fold cross-validation method. (E) Cross-validation error curves between GEA and CCC illustrating the 10-fold cross-validation method employed to determine prioritization of top features for constructing simplified models. (F) ROC curve illustrating predictive performance of simplified RF model for distinguishing between GEA and CCC based on the top 6 genera.
Additionally, LEfSe analysis highlighted significant alterations between microbial profiles of GEA and CCC groups, underscoring the importance of microbial signatures in differentiating these conditions. To ensure an adequate sample size, all samples of GEA and CCC were included in the analysis. Adhering to the principle of simplicity, the top 6 genera were then selected for the final model (Figures 4D, E), which exhibited outstanding predictive accuracy (AUC = 0.944) as shown in Figure 4F. When constructing these models, a taxon was included only if it was present in over 20% samples.
Microbial abundances and their clinical implications
Further investigation into the correlation between specific microbes and clinical characteristics of HPVI ECA patients revealed varied microbial abundances. Patients with LNM showed a notably lower presence of Unknown genus138 of family Ellin6075 (p=0.0135). Balneimonas exhibited significantly lower abundance in patients with deep stromal invasion (p=0.0333). Notably, the low-abundance of Unknown genus75 of family Comamonadaceae was also linked to LVI (p=0.0429). Additionally, patients with larger tumors (≥ 4cm) exhibited a decrease in Balneimonas (p=0.0336), Unknown genus138 of family Ellin6075 (p=0.0426), Unknown genus148 of Acetobacteraceae (p=0.0405), and an increase in Micrococcus (p=0.0134) (Figure 5A). Advanced tumor stages correlated significantly with the abundance of certain microbes: Balneimonas (p=0.018, r=-0.35), Unknown genus75 of family Comamonadaceae (p=0.033, r=-0.32) were inversely related to advanced tumor stage, while Micrococcus exhibited a positive correlation (p=0.02, r=0.35) (Figure 5B).
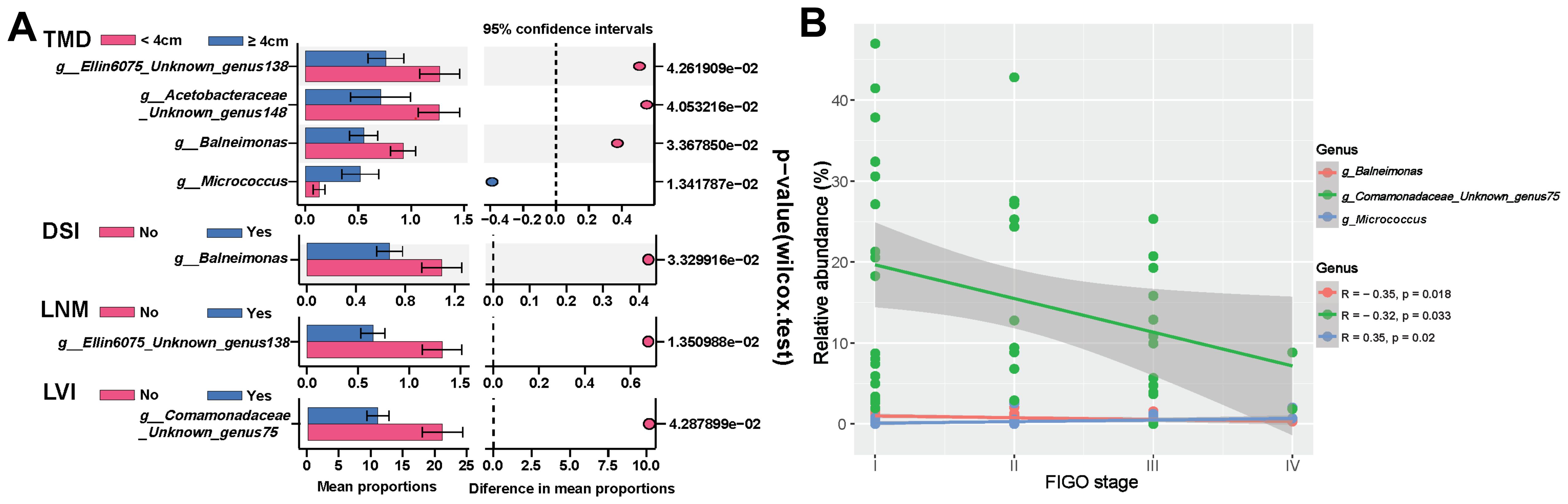
Figure 5. Correlation between intratumoral microbiota and clinical-pathological features of human papillomavirus independent endocervical adenocarcinoma (HPVI ECA). (A) Differential enrichment of specific microbiota across stratifications based on distinct clinical characteristics. (B) Relationship between specific microbiota and tumor stage. TMD, tumor maximum diameter; DSI, deep stromal invasion; LNM, lymph node metastasis; LVI, lymphovascular invasion.
Survival analysis indicated that specific microbial abundances could be indicative of recurrence and survival probabilities in patients (Figure 6). High levels of Micrococcus and low levels of Unknown genus75 of family Comamonadaceae were associated with poorer outcomes in HPVI ECA patients. Intriguingly, a high abundance of Micrococcus was also associated with worse OS in GEA patients.
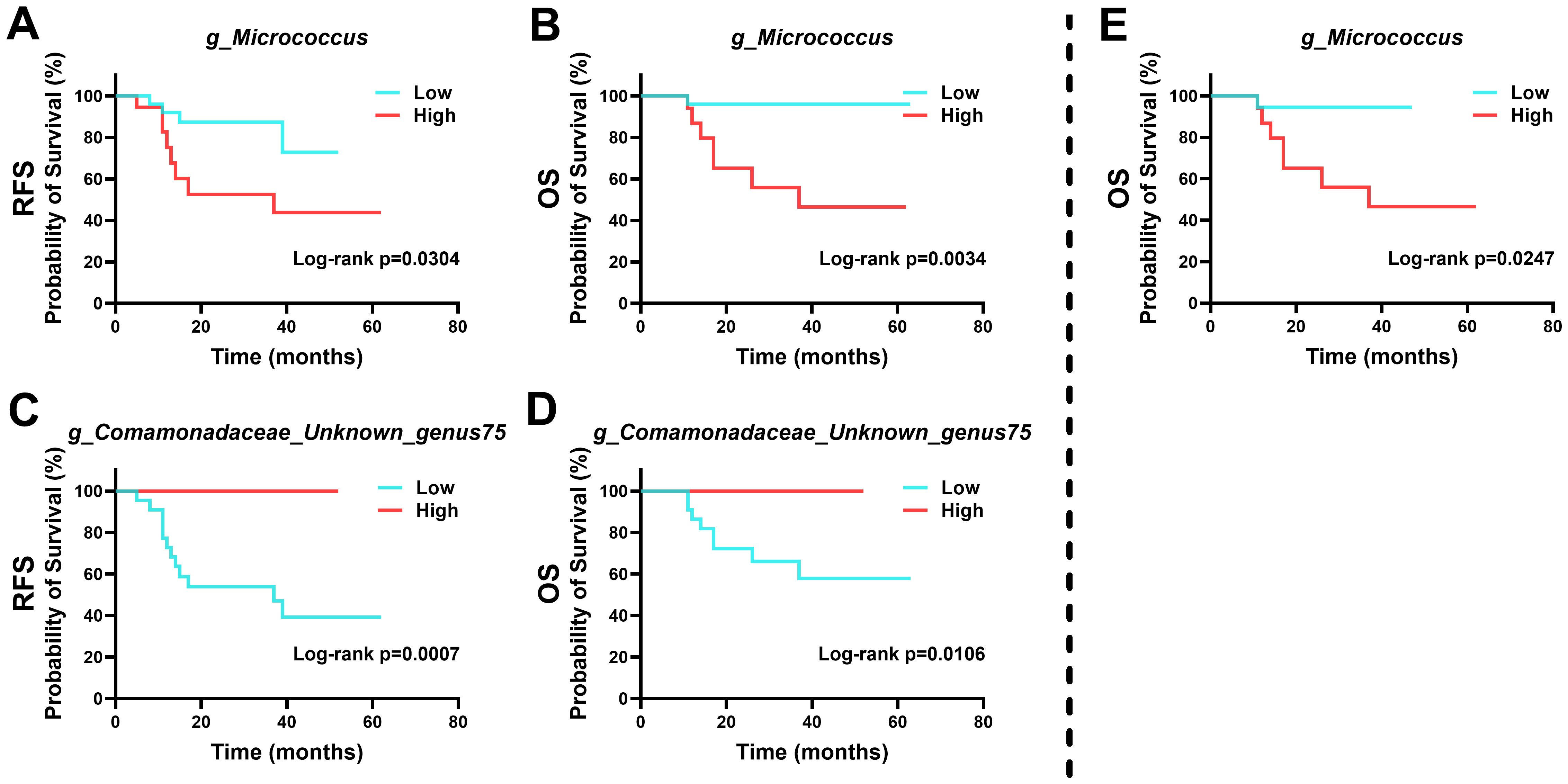
Figure 6. Kaplan-Meier survival curves based on intra-tumoral microbial abundance. (A-D) Relationship between intra-tumoral microbiota and prognosis of human papillomavirus independent endocervical adenocarcinoma (HPVI ECA) patients. (E) Relationship between intra-tumoral microbiota and prognosis of gastric-type endocervical adenocarcinoma (GEA) patients.
Predicted functional impact of the HPVI ECA microbiome
The study annotated and analyzed KEGG pathways between the GEA and CCC groups, uncovering 106 pathways with significant variances (Supplementary Table S4). The GEA group exhibited an increase in 64 pathways including those involved in proteasome function, polycyclic aromatic hydrocarbon degradation, and various metabolic processes, while the CCC group showed enrichment in 42 pathways, including those related to sphingolipid metabolism and biosynthesis of certain amino acids and bile acids (Figure 7). These data suggest a significant metabolic reprogramming within the microbial communities in the HPVI ECA subtype. However, when comparing HPVI ECA tumors to adjacent tissues, no significant changes were noted in KEGG pathways, indicating a microbial similarity between these groups. Yet, nine COG functional categories differed significantly between the two groups (Supplementary Figure S3, Supplementary Table S5).
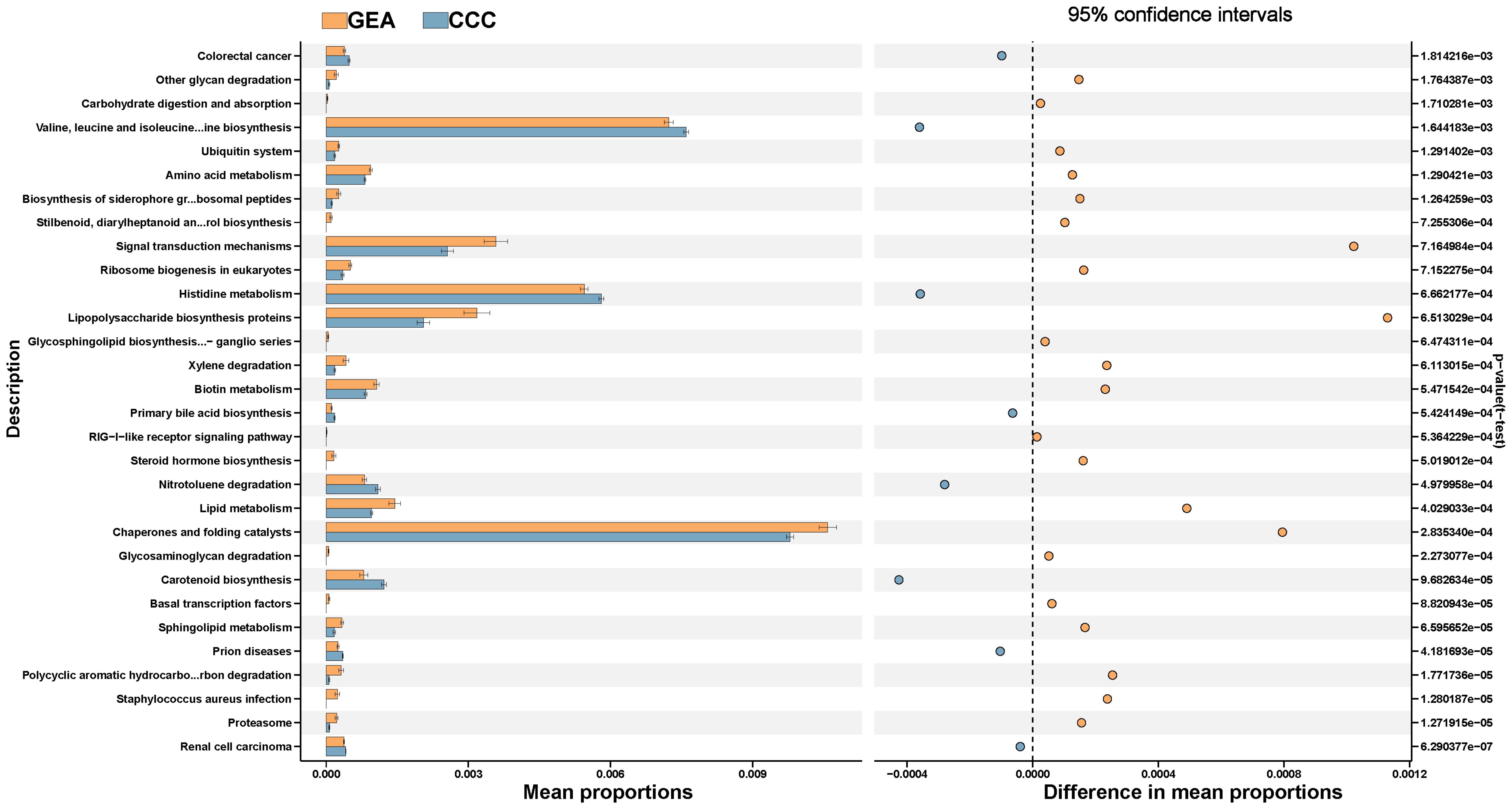
Figure 7. The top 30 Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways between gastric-type endocervical adenocarcinoma (GEA) and clear cell carcinoma (CCC) groups, along with their respective 95% confidence intervals and p-values.
Discussion
Mounting evidence increasingly implicates dysbiosis of the reproductive tract microbiota as a key risk factor for the onset of cervical cancer (Hu et al., 2018; Zhou et al., 2022). Despite a rising incidence of cervical adenocarcinoma (Smith et al., 2000), no studies have reported on microbial dysbiosis related to the development of this disease in patients. In our research, we concentrated on tissue samples from HPVI ECA to evaluate both the composition and the variation of the microbiota, alongside its prognostic implications. Our principal discoveries are as follows: 1) microbial profiles in HPVI ECA exhibit alterations, displaying slightly deviations from neighboring tissues; 2) amongst HPVI ECA subtypes, GEA reveals more pronounced microbial profiles changes compared to CCC; 3) We employed the RF algorithm to reveal distinct microbial signatures for predicting HPV-induced ECA carcinogenesis composed of seven bacterial species, as well as another microbial signature discriminating between HPVI ECA subtypes (GEA and CCC) involving six bacterial species; 4) We identified specific bacterial profiles that correlated with both the prognosis and the clinical-pathological features of HPVI ECA.
By employing 5R-16S rDNA sequencing, we delineated the microbial composition of 45 matched pairs of HPVI ECA carcinomas and their adjacent non-cancerous tissue. The disparity in microbial species abundance and richness between cancerous and adjacent non-cancerous tissues was negligible in terms of both α-diversity and β-diversity, which may be due to their physical closeness. However, in GEA, microbial abundance was significantly greater than in CCC. Literature suggests that a diminished microbial community diversity might correlate with disease progression, while a higher diversity typically signals a healthier condition, as noted in studies on gastric cancer (Liu et al., 2019b) and pancreatic cancer (Riquelme et al., 2019). Similar to these findings, our study discovered significantly lower microbial α-diversity (Shannon p = 0.0439) indices between HPV-associated endocervical adenocarcinoma (ECA) carcinogenic lesions and adjacent tissues, indicating a decrease in diversity associated with cancer. These patterns suggest that the disease’s advancement could be related to either a simplification or a diversification of microbial communities within the ecosystem of the microbiota.
Within the scope of our investigation, the predominant phylum in both the cancerous and adjacent tissues was Proteobacteria, aligning with previous findings on the microbiota in cervical cancer (Chang et al., 2023a; Mulato-Briones et al., 2024). Additionally, the relative abundance of Thermi and Deinococcus-thermus exceeded 10%, yet did not display significant variances between the groups. Notably, although microbes such as Firmicutes and Actinobacteria were reported as dominant in earlier studies (Chang et al., 2023a; Mulato-Briones et al., 2024), they did not appear as prevalent in our research, possibly due to the distinct nature of our samples or specific microbial traits linked to HPVI ECA. Importantly, we observed variations in the relative abundance of these microbes between GEA and CCC, hinting at their nuanced involvement in the evolution of HPVI ECA subtypes. At the genus level, the microbial community within HPVI ECA cancerous tissues, as opposed to adjacent non-cancerous tissues, was predominantly composed of Deinococcus, Unknown genus75 of family Comamonadaceae, Cupriavidus, Meiothermus, Porphyrobacter. Notably, Meiothermus as identified in further LEfSe analysis, was more prevalent in cancerous lesions, suggesting its potential as a biomarker for the onset and progression of HPVI ECA.
The RF algorithm has proven to be an effective method for predicting different disease states, particularly through the identification of microbial signatures (Yu et al., 2017; Nakatsu et al., 2018; Half et al., 2019; Yang et al., 2023). This allows researchers to delve into the complexities of microbial dysbiosis and identify targets for managing these imbalances. Prior research by Wu et al. (Wu et al., 2023) and Chang et al (Chang et al., 2023b). has applied the RF algorithm to identify distinctive microbial features in patients with cervical cancer, and to correlate these with tumor stages and prognostic outcomes. Additionally, Jiang et al. have constructed a RF-based risk model aimed at predicting the metastasis of cervical cancer (Jiang et al., 2023). Notably, the two RF models established in our study both demonstrate high predictive accuracy. Furthermore, there were significant differences in the abundance of these bacteria between the two groups. The highly consistent results among these analyses explain the remarkable ability of this group of bacteria to distinguish between the two HPVI ECA subtypes. Out of the ten bacterial markers identified through the two RF models, Anaerococcus (Chen et al., 2020) and Porphyrobacter (Lin et al., 2022) has been demonstrated a direct association with cervical intraepithelial neoplasia. Moreover, Micrococcus, which is implicated in inflammatory responses in endometrial cancer, has been linked to a higher incidence of cervical cancer (Lu et al., 2021), and its increased abundance is associated with cervical cancer (Tango et al., 2020).
We further explored the relationship between the abundance of certain microbe and the clinical outcomes of patients. Notably, a higher abundance of Micrococcus has been linked with worse prognosis in patients with HPVI ECA, and is positively correlated with the advanced clinical stage, indicating its potential role in the progression of cancer. The discovery of microbial markers such as Balneimonas, Unknown genus75 of family Comamonadaceae and Unknown genus138 of family Ellin6075 could provide valuable insights into the mechanisms underlying tumor metastasis. The development of agents targeting these bacteria and their associated derivatives holds tremendous promise in cancer therapy. Various bacterial derivatives have been reported to selectively target cancer cells and exert cytotoxic effects, including bacterial toxins, enzymes, peptides, and some secondary bacterial metabolites. Current research has identified several Micrococcus with anticancer activity, such as the ethyl acetate extract and yellow pigment from Micrococcus luteus, which exhibit anticancer effects against cancer cells (Mithun and Rao, 2012; Ushasri and Shalomi, 2015). Other reports indicate that the carotenoids pigment from Micrococcus roseus possess antioxidant and anticancer properties (Rostami et al., 2016). Furthermore, a recent study revealed that the yellow pigment from Micrococcus terreus can activate cysteine to induce apoptosis in cervical cancer cell lines (Shukla and Nadumane, 2021). This burgeoning field of research underscores the potential of bacterial derivatives as valuable additions to the arsenal of cancer therapeutics, warranting further exploration and clinical translation.
The research presented here offers a preliminary exploration of the microbial community within cervical adenocarcinoma tumors. However, it is not without limitations, particularly the potential for the 16S rDNA amplification and sequencing approach to overlook certain members of the microbial community. This technique may not capture as comprehensive a picture as metagenomic sequencing and leaves some microbial taxa unidentified. Additionally, the limited sample size and lack of diversity in the tissue samples may constrain the wider applicability of these findings. The study underscores the necessity for broader and more detailed mechanistic studies to validate these findings and proposes further investigation into the relationship between intra-tumoral and vaginal microbiota, which may contribute to the advancement of cervical cancer prevention and treatment strategies.
Conclusion
In summary, our investigation has charted the landscape of tumor-associated microbiota in HPVI ECA, uncovering microbial signatures with potential as predictive biomarkers. Utilizing RF algorithms, we’ve linked specific microbiota profiles to clinical outcomes and established their correlation with disease progression. Despite certain limitations, our findings offer a promising gateway to better understand the role of microbiota in HPVI ECA, which may lead to improved diagnostic and therapeutic strategies. As we seek to enhance cervical cancer management, this research lays the groundwork for further exploration into the microbiome’s impact on patient prognosis.
Data availability statement
The datasets generated for this study can be found in the SRA of NCBI: https://www.ncbi.nlm.nih.gov/sra/PRJNA1142715.
Ethics statement
The studies involving humans were approved by Institutional Review Board and Ethics Committee of the Women’s Hospital, School of Medicine, Zhejiang University, China. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author contributions
XZ: Conceptualization, Data curation, Investigation, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing. LC: Conceptualization, Investigation, Methodology, Software, Supervision, Validation, Writing – original draft, Writing – review & editing. WL: Conceptualization, Data curation, Formal analysis, Methodology, Supervision, Writing – original draft, Writing – review & editing. WZ: Conceptualization, Formal analysis, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing. HZ: Conceptualization, Data curation, Formal analysis, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing. FZ: Conceptualization, Data curation, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interest
The authors declared that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer XC declared a shared affiliation with the authors to the handling editor at the time of review.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2024.1440017/full#supplementary-material
References
Adegoke, O., Kulasingam, S., Virnig, B. (2012). Cervical cancer trends in the United States: a 35-year population-based analysis. J. Womens Health (Larchmt) 21, 1031–1037. doi: 10.1089/jwh.2011.3385
Audirac-Chalifour, A., Torres-Poveda, K., Bahena-Román, M., Téllez-Sosa, J., Martínez-Barnetche, J., Cortina-Ceballos, B., et al. (2016). Cervical microbiome and cytokine profile at various stages of cervical cancer: A pilot study. PLoS One 11, e0153274. doi: 10.1371/journal.pone.0153274
Bergström, R., Sparén, P., Adami, H. O. (1999). Trends in cancer of the cervix uteri in Sweden following cytological screening. Br. J. Cancer 81, 159–166. doi: 10.1038/sj.bjc.6690666
Bhatla, N., Aoki, D., Sharma, D. N., Sankaranarayanan, R. (2018). Cancer of the cervix uteri. Int. J. Gynaecol Obstet 143 Suppl;2, 22–36. doi: 10.1002/ijgo.12611
Byrd, D. A., Fan, W., Greathouse, K. L., Wu, M. C., Xie, H., Wang, X. (2023). The intratumor microbiome is associated with microsatellite instability. J. Natl. Cancer Inst 115, 989–993. doi: 10.1093/jnci/djad083
Chang, L., Qiu, L., Lei, N., Zhou, J., Guo, R., Gao, F., et al. (2023a). Characterization of fecal microbiota in cervical cancer patients associated with tumor stage and prognosis. Front. Cell Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1145950
Chang, L., Qiu, L., Lei, N., Zhou, J., Guo, R., Gao, F., et al. (2023b). Characterization of fecal microbiota in cervical cancer patients associated with tumor stage and prognosis. Front. Cell Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1145950
Chen, Y., Qiu, X., Wang, W., Li, D., Wu, A., Hong, Z., et al. (2020). Human papillomavirus infection and cervical intraepithelial neoplasia progression are associated with increased vaginal microbiome diversity in a Chinese cohort. BMC Infect. Dis. 20, 629. doi: 10.1186/s12879-020-05324-9
Douglas, G. M., Maffei, V. J., Zaneveld, J. R., Yurgel, S. N., Brown, J. R., Taylor, C. M., et al. (2020). PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 38, 685–688. doi: 10.1038/s41587-020-0548-6
García-Castillo, V., Sanhueza, E., McNerney, E., Onate, S. A., García, A. (2016). Microbiota dysbiosis: a new piece in the understanding of the carcinogenesis puzzle. J. Med. Microbiol. 65, 1347–1362. doi: 10.1099/jmm.0.000371
Half, E., Keren, N., Reshef, L., Dorfman, T., Lachter, I., Kluger, Y., et al. (2019). Fecal microbiome signatures of pancreatic cancer patients. Sci. Rep. 9, 16801. doi: 10.1038/s41598-019-53041-4
Hornung, R., Wright, M. N. (2019). Block Forests: random forests for blocks of clinical and omics covariate data. BMC Bioinf. 20, 358. doi: 10.1186/s12859-019-2942-y
Hu, J., Gao, Y., Zheng, Y., Shang, X. (2018). KF-finder: identification of key factors from host-microbial networks in cervical cancer. BMC Syst. Biol. 12, 54. doi: 10.1186/s12918-018-0566-x
Jain, T., Sharma, P., Are, A. C., Vickers, S. M., Dudeja, V. (2021). New insights into the cancer-microbiome-immune axis: decrypting a decade of discoveries. Front. Immunol. 12. doi: 10.3389/fimmu.2021.622064
Jiang, L., Duan, B., Jia, P., Zhang, Y., Yan, X. (2023). The role of intratumor microbiomes in cervical cancer metastasis. Cancers (Basel) 15, (2). doi: 10.3390/cancers15020509
Karamurzin, Y. S., Kiyokawa, T., Parkash, V., Jotwani, A. R., Patel, P., Pike, M. C., et al. (2015). Gastric-type endocervical adenocarcinoma: an aggressive tumor with unusual metastatic patterns and poor prognosis. Am. J. Surg. Pathol. 39, 1449–1457. doi: 10.1097/PAS.0000000000000532
Kojima, A., Mikami, Y., Sudo, T., Yamaguchi, S., Kusanagi, Y., Ito, M., et al. (2007). Gastric morphology and immunophenotype predict poor outcome in mucinous adenocarcinoma of the uterine cervix. Am. J. Surg. Pathol. 31, 664–672. doi: 10.1097/01.pas.0000213434.91868.b0
Kusanagi, Y., Kojima, A., Mikami, Y., Kiyokawa, T., Sudo, T., Yamaguchi, S., et al. (2010). Absence of high-risk human papillomavirus (HPV) detection in endocervical adenocarcinoma with gastric morphology and phenotype. Am. J. Pathol. 177, 2169–2175. doi: 10.2353/ajpath.2010.100323
Lin, S., Zhang, B., Lin, Y., Lin, Y., Zuo, X. (2022). Dysbiosis of cervical and vaginal microbiota associated with cervical intraepithelial neoplasia. Front. Cell Infect. Microbiol. 12. doi: 10.3389/fcimb.2022.767693
Liu, H., Pan, L. L., Lv, S., Yang, Q., Zhang, H., Chen, W., et al. (2019a). Alterations of gut microbiota and blood lipidome in gestational diabetes mellitus with hyperlipidemia. Front. Physiol. 10. doi: 10.3389/fphys.2019.01015
Liu, X., Shao, L., Liu, X., Ji, F., Mei, Y., Cheng, Y., et al. (2019b). Alterations of gastric mucosal microbiota across different stomach microhabitats in a cohort of 276 patients with gastric cancer. EBioMedicine 40, 336–348. doi: 10.1016/j.ebiom.2018.12.034
Lu, W., He, F., Lin, Z., Liu, S., Tang, L., Huang, Y., et al. (2021). Dysbiosis of the endometrial microbiota and its association with inflammatory cytokines in endometrial cancer. Int. J. Cancer 148, 1708–1716. doi: 10.1002/ijc.33428
Meng, C., Bai, C., Brown, T. D., Hood, L. E., Tian, Q. (2018). Human gut microbiota and gastrointestinal cancer. Genomics Proteomics Bioinf. 16, 33–49. doi: 10.1016/j.gpb.2017.06.002
Mithun, V., Rao, C. J. I. (2012). Isolation and molecular characterization of anti-cancerous compound producing marine bacteria by using 16S rRNA sequencing and GC-MS techniques. Int. J. Modern Eng. Res. 2, 4510–4515.
Mitra, A., MacIntyre, D. A., Lee, Y. S., Smith, A., Marchesi, J. R., Lehne, B., et al. (2015). Cervical intraepithelial neoplasia disease progression is associated with increased vaginal microbiome diversity. Sci. Rep. 5, 16865. doi: 10.1038/srep16865
Mulato-Briones, I. B., Rodriguez-Ildefonso, I. O., Jiménez-Tenorio, J. A., Cauich-Sánchez, P. I., Méndez-Tovar, M., Aparicio-Ozores, G., et al. (2024). Cultivable microbiome approach applied to cervical cancer exploration. Cancers (Basel) 16, 314. doi: 10.3390/cancers16020314
Nakatsu, G., Zhou, H., Wu, W. K. K., Wong, S. H., Coker, O. O., Dai, Z., et al. (2018). Alterations in enteric virome are associated with colorectal cancer and survival outcomes. Gastroenterology 155, 529–541.e5. doi: 10.1053/j.gastro.2018.04.018
Nejman, D., Livyatan, I., Fuks, G., Gavert, N., Zwang, Y., Geller, L. T., et al. (2020). The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 368, 973–980. doi: 10.1126/science.aay9189
Nishio, S., Mikami, Y., Tokunaga, H., Yaegashi, N., Satoh, T., Saito, M., et al. (2019). Analysis of gastric-type mucinous carcinoma of the uterine cervix - An aggressive tumor with a poor prognosis: A multi-institutional study. Gynecol Oncol. 153, 13–19. doi: 10.1016/j.ygyno.2019.01.022
Noller, K. L., Decker, D. G., Dockerty, M. B., Lanier, A. P., Smith, R. A., Symmonds, R. E. (1974). Mesonephric (clear cell) carcinoma of the vagina and cervix. A retrospective analysis. Obstet Gynecol 43, 640–644.
Parks, D. H., Tyson, G. W., Hugenholtz, P., Beiko, R. G. (2014). STAMP: statistical analysis of taxonomic and functional profiles. Bioinformatics 30, 3123–3124. doi: 10.1093/bioinformatics/btu494
Parra-Herran, C., Hoang, L. N., Park, K. J., Kong, C. S., Ohishi, Y., Kato, N., et al, WHO Classification of Tumours Editorial Board, Female Genital Tumours (2020). “Tumours of the uterine cervix/glandular tumours and precursors,” in WHO Classification of Tumours Series, 5th edition (International Agency for Reseach on Cancer, Lyon, France), 367–371.
Pushalkar, S., Hundeyin, M., Daley, D., Zambirinis, C. P., Kurz, E., Mishra, A., et al. (2018). The pancreatic cancer microbiome promotes oncogenesis by induction of innate and adaptive immune suppression. Cancer Discovery 8, 403–416. doi: 10.1158/2159-8290.CD-17-1134
Riquelme, E., Zhang, Y., Zhang, L., Montiel, M., Zoltan, M., Dong, W., et al. (2019). Tumor microbiome diversity and composition influence pancreatic cancer outcomes. Cell 178, 795–806.e12. doi: 10.1016/j.cell.2019.07.008
Rivera-Pinto, J., Egozcue, J. J., Pawlowsky-Glahn, V., Paredes, R., Noguera-Julian, M., Calle, M. L. (2018). Balances: a new perspective for microbiome analysis. mSystems 3, 12. doi: 10.1128/mSystems.00053-18
Rostami, H., Hamedi, H., Yolmeh, M. (2016). Some biological activities of pigments extracted from Micrococcus roseus (PTCC 1411) and Rhodotorula glutinis (PTCC 5257). Int. J. Immunopathology Pharmacol. 29, 684–695. doi: 10.1177/0394632016673846
Saigo, P. E., Cain, J. M., Kim, W. S., Gaynor, J. J., Johnson, K., Lewis, J. L., Jr (1986). Prognostic factors in adenocarcinoma of the uterine cervix. Cancer 57, 1584–1593. doi: 10.1002/1097-0142(19860415)57:8<1584::aid-cncr2820570825>3.0.co;2-8
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W. S., et al. (2011). Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60. doi: 10.1186/gb-2011-12-6-r60
Shukla, M., Nadumane, V. K. (2021). Yellow pigment from a novel bacteria, Micrococcus terreus, activates caspases and leads to apoptosis of cervical and liver cancer cell lines. J. Appl. Pharm. Sci. 11, 077–084. doi: 10.7324/JAPS.2021.110811
Smith, H. O., Tiffany, M. F., Qualls, C. R., Key, C. R. (2000). The rising incidence of adenocarcinoma relative to squamous cell carcinoma of the uterine cervix in the United States–a 24-year population-based study. Gynecol Oncol. 78, 97–105. doi: 10.1006/gyno.2000.5826
Stolnicu, S., Barsan, I., Hoang, L., Patel, P., Terinte, C., Pesci, A., et al. (2018). International endocervical adenocarcinoma criteria and classification (IECC): A new pathogenetic classification for invasive adenocarcinomas of the endocervix. Am. J. Surg. Pathol. 42, 214–226. doi: 10.1097/PAS.0000000000000986
Sun, Y., Zhou, X., Lucas, E., Chen, L., Zhang, H., Chen, H., et al. (2024). Expression of B7-H3 and TIM-3 in gastric-type endocervical adenocarcinoma: prevalence, association with PD-L1 expression, and prognostic significance. J. Pathol. Clin. Res. 10, e345. doi: 10.1002/cjp2.345
Tango, C. N., Seo, S. S., Kwon, M., Lee, D. O., Chang, H. K., Kim, M. K. (2020). Taxonomic and functional differences in cervical microbiome associated with cervical cancer development. Sci. Rep. 10, 9720. doi: 10.1038/s41598-020-66607-4
Teka, B., Yoshida-Court, K., Firdawoke, E., Chanyalew, Z., Gizaw, M., Addissie, A., et al. (2023). Cervicovaginal microbiota profiles in precancerous lesions and cervical cancer among Ethiopian women. Microorganisms 11, (4). doi: 10.3390/microorganisms11040833
Ushasri, R., Shalomi, C. G. W. (2015). A study on in vitro anti breastcancer activity of crude ethanol and acetone pigment extracts of Micrococcus luteus by MTT assay and analysis of pigment by thin layer chromatography. Int. J. Pharm. Biol. Sci. 5, 59–65.
Wang, S., Zhou, X., Niu, S., Chen, L., Zhang, H., Chen, H., et al. (2023). Assessment of HER2 in gastric-type endocervical adenocarcinoma and its prognostic significance. Mod Pathol. 36, 100148. doi: 10.1016/j.modpat.2023.100148
Wu, S., Ding, X., Kong, Y., Acharya, S., Wu, H., Huang, C., et al. (2021). The feature of cervical microbiota associated with the progression of cervical cancer among reproductive females. Gynecol Oncol. 163, 348–357. doi: 10.1016/j.ygyno.2021.08.016
Wu, M., Yu, H., Gao, Y., Li, H., Wang, C., Li, H., et al. (2023). Leveraging 16S rRNA data to uncover vaginal microbial signatures in women with cervical cancer. Front. Cell Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1024723
Yang, J., He, Q., Lu, F., Chen, K., Ni, Z., Wang, H., et al. (2023). A distinct microbiota signature precedes the clinical diagnosis of hepatocellular carcinoma. Gut Microbes 15, 2201159. doi: 10.1080/19490976.2023.2201159
Yu, G., Hu, N., Wang, L., Wang, C., Han, X. Y., Humphry, M., et al. (2017). Gastric microbiota features associated with cancer risk factors and clinical outcomes: A pilot study in gastric cardia cancer patients from Shanxi, China. Int. J. Cancer 141, 45–51. doi: 10.1002/ijc.30700
Keywords: cervical cancer, gastric-type endocervical adenocarcinoma, clear cell carcinomas, intratumoral microbiota, prognosis
Citation: Zhou X, Chen L, Lin W, Zheng W, Zhang H and Zhou F (2024) Diagnostic and prognostic potential of the intra-tumoral microbiota profile in HPV-independent endocervical adenocarcinoma. Front. Cell. Infect. Microbiol. 14:1440017. doi: 10.3389/fcimb.2024.1440017
Received: 28 May 2024; Accepted: 24 July 2024;
Published: 16 August 2024.
Edited by:
Changwu Wu, Central South University, ChinaReviewed by:
Xueqiong Zhu, Second Affiliated Hospital and Yuying Children’s Hospital of Wenzhou Medical University, ChinaXuejun Chen, Zhejiang University School of Medicine, China
Copyright © 2024 Zhou, Chen, Lin, Zheng, Zhang and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Feng Zhou, ZnVuZ2NoZXdAemp1LmVkdS5jbg==; Huijuan Zhang, emhhbmdoajgxNUBzanR1LmVkdS5jbg==
†These authors have contributed equally to this work